
Regulated Revenues and Hospital Behavior - Boston University
Upload
michiganstateCategory
view
1download
0
Conservation of thiol-oxidative stress responses regulated bySigR orthologues in actinomycetesmmi_8115 1..19
Min-Sik Kim,1§ Yann S. Dufour,2,3†§ Ji Sun Yoo,1
Yoo-Bok Cho,1 Joo-Hong Park,1 Gi-Baeg Nam,1
Hae Min Kim,1 Kang-Lok Lee,1 Timothy J. Donohue2
and Jung-Hye Roe1*1Laboratory of Molecular Microbiology, School ofBiological Sciences, and Institute of Microbiology, SeoulNational University, Seoul 151-742, Korea.2Department of Bacteriology and 3BACTER Institute,University of Wisconsin, Madison, WI 53706, USA.
Summary
Numerous thiol-reactive compounds cause oxidativestress where cells counteract by activation of survivalstrategies regulated by thiol-based sensors. In Strep-tomyces coelicolor, a model actinomycete, a sigma/antisigma pair SigR/RsrA controls the response tothiol-oxidative stress. To unravel its full physiologicalfunctions, chromatin immuno-precipitation combinedwith sequence and transcript analyses were employedto identify 108 SigR target genes in S. coelicolor and topredict orthologous regulons across actinomycetes.In addition to reported genes for thiol homeostasis,protein degradation and ribosome modulation, 64additional operons were identified suggesting newfunctions of this global regulator. We demonstrate thatSigR maintains the level and activity of the housekeep-ing sigma factor HrdB during thiol-oxidative stress, anovel strategy for stress responses. We also foundthat SigR defends cells against UV and thiol-reactivedamages, in which repair UvrA takes a part. Using arefined SigR-binding sequence model, SigR ortho-logues and their targets were predicted in 42 actino-mycetes. This revealed a conserved core set of SigRtargets to function for thiol homeostasis, proteinquality control, possible modulation of transcriptionand translation, flavin-mediated redox reactions, andFe-S delivery. The composition of the SigR regulonreveals a robust conserved physiological mechanismto deal with thiol-oxidative stress from bacteria tohuman.
Introduction
Cells are exposed to various forms of oxidative stressorssuch as reactive oxygen species (ROS), natural or xeno-biotic redox-active compounds, or some antibiotics, whichelicit the formation of ROS inside the cell. The responsemechanism involves various enzymes to remove suchoxidants, systems to repair and recycle damaged cellcomponents, and to maintain optimal cell physiology(Imlay, 2008; Zuber, 2009). One form of oxidative damagethat frequently occurs in proteins and small molecules isthe oxidation of cysteine thiols by ROS (Kiley and Storz,2004; Jacob et al., 2006). Cysteine thiols can also bemodified by reactive nitrogen species (RNS) and thiol-reactive electrophiles (Hess et al., 2005; Rudolph andFreeman, 2009)
The presence of ROS, RNS and other thiol-reactivecompounds can be sensed directly through thiol-basedsensor-regulators, which modulate the expression ofgenes encoding functions that constitute the biologicalstress response (Antelmann and Helmann, 2011). Thebest studied examples of thiol-based sensor-regulatorsthat respond to ROS in bacteria include H2O2-sensingOxyR in Escherichia coli, organic peroxide-sensing OhrRin Bacillus subtilis, and the anti-sigma factor RsrA thatsenses thiol oxidation and modulates regulator SigR activ-ity in Streptomyces coelicolor (Paget and Buttner, 2003;D’Autreaux and Toledano, 2007; Antelmann and Helmann,2011). In eukaryotes, thiol-based redox switches havebeen well exemplified in Yap1 of Saccharomyces cerevi-siae and the mammalian Nrf2/Keap1 system (D’Autreauxand Toledano, 2007; Kensler et al., 2007; Brandes et al.,2009). Representative target genes of these thiol-basedregulators are those that encode thiol homeostasis systemsuch as thioredoxin (Trx), glutaredoxin (Grx) and smallmolecular thiol systems (regulated by OxyR, SigR/RsrA,Yap1, Nrf2/Keap1), catalases and peroxidases (by OxyRand Yap1), organic hydroperoxidase (by OhrR), detoxifica-tion of electrophiles (by SigR and Nrf2), and some pro-teolytic system (by OxyR, SigR, Yap1, Nrf2) (Kensler et al.,2007; den Hengst and Buttner, 2008; Imlay, 2008; Antel-mann and Helmann, 2011).
A zinc-containing anti-sigma (ZAS) factor RsrA thatbinds an ECF (group 4) sigma factor SigR in S. coelicolorresponds to diamide-induced thiol oxidation by formingdisulphide bonds, releasing SigR to transcribe its target
Accepted 23 May, 2012. *For correspondence. E-mail [email protected]; Tel. (+82) 2 880 6706; Fax (+82) 2 882 6706. †Present address:Department of Molecular, Cellular and Developmental Biology, YaleUniversity, New Haven, CT 06520, USA. §These two authors contrib-uted equally.
Molecular Microbiology (2012) � doi:10.1111/j.1365-2958.2012.08115.x
© 2012 Blackwell Publishing Ltd

genes (Kang et al., 1999; Li et al., 2003b; Bae et al., 2004).In addition to thiol-oxidants, the presence of non-oxidativethiol-reactive compounds also induces expression of theSigR regulon, suggesting that RsrA may respond to thesecompounds either directly through thiolation of reactivecysteines or indirectly through changes in reduced thiolpools such as mycothiol, the functional equivalent in acti-nomycetes of glutathione (Newton et al., 2008; Park andRoe, 2008). To date, around 44 SigR target genes havebeen identified based on promoter sequence similarity, S1mapping and microarray expression profiling (Paget et al.,1998; 2001; Park and Roe, 2008; Kallifidas et al., 2010).The products of known SigR target genes include thiol-redox proteins such as thioredoxin systems (TrxBA, TrxC),the first enzyme in mycothiol synthesis (MshA), and aglutaredoxin-like protein called mycoredoxin (Mrx). Theyalso include proteolytic components of protein qualitycontrol (PepN, SsrA, ClpP1P2, ClpX, ClpC, Lon), cysteineproduction (CysM), methionine reduction (MsrA, MsrB),guanine synthesis (GuaB), ribosome-associated function(RpmE, RelA), and electrophile detoxification (Mca) (Pagetet al., 2001; Park and Roe, 2008; Kallifidas et al., 2010).Therefore, perturbation of intracellular thiols, as sensedthrough RsrA, induces expression of gene products thatcontributes to protein quality control, detoxification of thiol-conjugative xenobiotics, and modulation of transcriptionand translation.
Because SigR/RsrA activity controls the cellularresponse to a variety of stressors, we expect the SigRregulon to be large and encompass additional stressresponse functions in S. coelicolor. Therefore, we per-formed chromatin immuno-precipitation on a chip (ChIP-chip) assays and discovered > 100 direct SigR targetpromoters transcribing > 160 genes. The comprehensivecharacterization of the S. coelicolor SigR regulon helpedus understand in more details the global effects of andcellular response to thiol-oxidative stress. Furthermore, toidentify functions that are most critical to survive thiol-oxidative stress response, we used the SigR-binding con-sensus sequence to predict orthologous thiol-oxidativestress regulons in other actinomycetes. The analysisrevealed a core conserved regulon as well as lineage-specific adaptations.
Results and discussion
Identification of SigR target genes by chromatinimmuno-precipitation on a chip (ChIP-chip) andsequence analysis
To identify SigR target sites in the S. coelicolor genome weperformed a ChIP-chip experiment using anti-SigR serumwith wild-type cell cultures treated with diamide to activateSigR activity. We also performed the same ChIP-chip
experiment with a DsigR mutant to subtract non-specificchromatin enrichment because S. coelicolor possessesmany additional sigma factors that may cross-react withthe anti-SigR serum. After subtracting non-specific signal,we identified 122 regions significantly enriched by theimmuno-precipitation of SigR (P-value � 0.05). Next, toidentify more precisely putative SigR promoters, wesearched within the enriched genomic regions for putativeSigR promoters by detecting sequences that differed fromthe previously proposed SigR promoter (GGAAT-N18-GTT;(Paget et al., 2001) by at most 2 nucleotides with either of2 possible spacer lengths (18 and 19 nucleotides) on eitherDNA strand. Only 5 of the 122 enriched regions did notcontain at least one sequence matching our criteria,increasing the confidence that the regions enriched by theChIP-chip assay are indeed SigR binding sites. Overall,176 putative SigR promoter sequences were detected, and108 of them are oriented towards annotated genes within500 nt from the start codon.
In Table 1, we summarized genes predicted to be tran-scribed from these 108 promoters. Detailed informationabout the position of SigR-bound regions (enriched ChIPpeaks), position and sequence of the predicted SigR pro-moters is presented in Table S1. Considering operon struc-tures (http://scocyc.streptomyces.org.uk), more than 163genes are predicted to be under the direct control of SigR.Among these, 44 promoters have been previously reportedto produce SigR-dependent transcripts by S1 mappingexperiments (Paget et al., 2001; Park and Roe, 2008;Kallifidas et al., 2010) or microarray studies (Kallifidaset al., 2010). Re-examination of the transcriptionalmicroarray data (Kallifidas et al., 2010), using slightly dif-ferent criteria (see Experimental procedures), predictsSigR-dependent transcripts from a total of 62 promoterswithin 57 of the regions bound by SigR in vivo. We markedthese promoters in Table S1. Therefore, our ChIP-chipanalysis revealed between 46 and 64 new SigR targetpromoters, undetectable from transcriptome profilingalone. The lack of detection in microarray analysis alonecould be due partly to low level of gene expressionand/or additional transcription from one or more SigR-independent promoter.
When 108 SigR target operons were classified on thebasis of known and predicted functions, many of them fallin the functional groups related with maintenance of thiolredox homeostasis and sulphur-containing amino acids(11 operons; groups 1 and 2 in Table 1), protein folding anddegradation (7 operons; group 5), and oxidoreductases(15 operons; group 8) (Table 1), fortifying previous propos-als that the SigR regulon functions to achieve redoxhomeostasis and protein quality control under conditionsof thiol-oxidative stress (Paget et al., 2001; Park andRoe, 2008; Kallifidas et al., 2010). Within these groups,we noticed newly added genes encoding a putative
2 M.-S. Kim et al. �
© 2012 Blackwell Publishing Ltd, Molecular Microbiology

Table 1. The SigR regulon genes.
Ga Numberb Name Description Confirmc
3 SCO0569 rpmJ 50S ribosomal protein L363 SCO0570* rpmG3 50S ribosomal protein L33 (C-type) (S1)8 SCO0882* Hypothetical protein, possible dithiol-disulphide isomerase (S1)8 SCO0884* Probable oxidoreductase, probable pyridine nucleotide-disulphide oxidoreductases1 SCO0885* trxC Thioredoxin (S1, A)8 SCO0917* Possible oxygenase7 SCO0973* Probable integral membrane protein1 SCO1084* trxA4 Putative thioredoxin S1
12 SCO1085* Probable phospholipid/glycerol acyltransferase, divergent from trxA4 (1084)8 SCO1142* Possible oxidoreductase, probable FAD-dependent pyridine nucleotide-disulphide
oxidoreductase5 SCO1238* Putative ATP-dependent Clp protease subunit6 SCO1425* Possible AsnC-family transcriptional regulatory protein
14 SCO1426* Hypothetical protein3 SCO1513* relA GTP pyrophosphokinase for ppGpp synthesis (S1)3 SCO1598 rplT 50S ribosomal protein L203 SCO1599 rpmI 50S ribosomal protein L353 SCO1600* infC Translation initiation factor IF-3 S16 SCO1618 Hypothetical protein, similar to cobalamin binding protein BtuF6 SCO1619* Possible transcriptional regulatory protein of HTH-XRE family5 SCO1643 prcA 20S proteasome alpha-subunit5 SCO1644 prcB 20S proteasome beta-subunit5 SCO1645 Hypothetical protein5 SCO1646 pup Prokaryotic ubiquitin-like protein5 SCO1647 pafD Proteasome accessory factor5 SCO1648* mpa Probable AAA ATPase similar to mycobacterium proteasome-associated ATPase S13 SCO1758* engA GTP-binding protein with multiple functions S18 SCO1869* Conserved hypothetical, probable protein dithiol-disulphide isomerase (S1, A)
11 SCO1919 Probable metal-sulphur cluster biosynthetic enzyme (A)11 SCO1920* sufU Similar to NifU in SUF-type Fe-S assembly gene cluster (1926-1919) (A), S19 SCO1936* tal2 Probable transaldolase, a component of non-oxidative pentose phosphate pathway S19 SCO1937 zwf2 Glucose 6-phosphate 1-dehydrogenase9 SCO1938 Hypothetical protein9 SCO1939 pgl 6-phosphogluconolactonase
10 SCO1958* uvrA ABC excision nuclease subunit A (A), S114 SCO1995* Unknown (S1, A)11 SCO1996 coaE Dephospho-CoA kinase (A)11 SCO1997* Unknown (S1, A)7 SCO2124* Possible hydrophobic membrane spanning regions7 SCO2154* Integral membrane protein
11 SCO2161* sufA HesB/YadR/Yfh family protein, probable SufA (S1), S111 SCO2162* nadA Quinolinate synthetase A subunit for nicotinamide synthesis, divergent from sufA
(2161). 4Fe-4S enzyme11 SCO2194* lipA Putative lipoyl synthase S17 SCO2254* Possible transmembrane efflux protein of MFS(major facilitator superfamily) S17 SCO2310* Possible transmembrane efflux protein similar to TcmA6 SCO2331* Possible MarR family transcriptional regulator6 SCO2481* Hypothetical protein, putative transcriptional regulator6 SCO2537* Possible DNA-binding protein (A), S16 SCO2538 Hypothetical protein6 SCO2539 era GTP-binding protein era3 SCO2595* obgE GTPase, Obg subfamily, ObgE S15 SCO2617 clpX ATP-dependent Clp protease ATP binding subunit5 SCO2618 clpP2 ATP-dependent Clp protease proteolytic subunit 2 (A)5 SCO2619* clpP1 ATP dependent Clp protease proteolytic subunit 1 (S1, A)8 SCO2634* Hypothetical protein, contains DsbA-like thioredoxin domain (S1, A)5 SCO2635* Probable aminopeptidase
14 SCO2642* Unknown5 SCO2643* pepN Aminopeptidase N (S1, A)7 SCO2763* ABC transporter protein, ATP-binding component S18 SCO2816* Conserved hypothetical protein, flavin utilizing monooxygenase superfamily8 SCO2849* Conserved hypothetical protein, flavin utilizing monooxygenase superfamily (S1, A)2 SCO2910 cysM Cysteine synthase (A)2 SCO2911* moaD Putative MoaD-family protein with ubiquitin fold (A)3 SCOs02* ssrA Transfer messenger RNA (tmRNA) (S1)7 SCO3083* Possible integral membrane protein (S1, A)
Conserved thiol-oxidative stress in actinomycetes 3
© 2012 Blackwell Publishing Ltd, Molecular Microbiology

Table 1. cont.
Ga Numberb Name Description Confirmc
12 SCO3091* cfa Cyclopropane-fatty-acyl-phospholipid synthase, SAM-dependent methylase (A), S113 SCO3162* Possible esterase, b-lactamase superfamily (S1, A)14 SCO3187* Hypothetical protein (S1)7 SCO3206* Probable transmembrane efflux protein (A)6 SCO3207* TetR family transcriptional regulator8 SCO3295 Possible oxidoreductase8 SCO3296* Possible oxidoreductase, similar to F420-dependent alcoholdehydrogenase S15 SCO3373* clpC Probable Clp-family ATP-binding protease similar to MecB/ClpC (S1), S1
11 SCO3403* folE GTP cyclohydrolase I for tetrahydrofolate biosynthesis (S1)1 SCO3442* mrxB Glutaredoxin-like protein, probable mycoredoxin S16 SCO3449 rsrA2-1 Putative ZAS (zinc-containing anti-sigma) factor6 SCO3450* sigR2 ECF (group 4) sigma factor close to SigR6 SCO3451* rsrA2-1 Putative ZAS (zinc-containing anti-sigma) factor
14 SCO3509* Hypothetical protein7 SCO3764 Integral membrane protein (A)7 SCO3765* Possible integral membrane protein with CBS domain
14 SCO3766* Hypothetical protein14 SCO3767* Hypothetical protein, similar to tellurite resistance protein TerB1 SCO3889 trxA Thioredoxin (A)1 SCO3890* trxB Thioredoxin reductase (S1, A)
14 SCO4039* Unknown (A)14 SCO4040* Unknown8 SCO4109* Possible oxidoreductase, probable aldo/keto reductases
14 SCO4203* Hypothetical protein1 SCO4204* mshA Glycosyltransferase for mycothiol synthesis (S1, A)1 SCO4205 Conserved hypothetical protein8 SCO4297* Possible oxidoreductase with FMN binding site, old yellow enzyme family (A)8 SCO4298 Carboxylesterase8 SCO4299 Integral membrane protein
11 SCO4418 pdxH2 Pyridoxamine 5′-phosphate oxidase oxidase11 SCO4419* pdxH3 Conserved hypothetical, probable pyridoxamine 5′-phosphate oxidase for pyridoxal
phosphate synthesis14 SCO4420* Hypothetical protein13 SCO4561* NLP/P60 family protein4 SCO4770* guaB Inosine 5′ monophosphate dehydrogenase (S1, A)4 SCO4771 guaB1 Inosine 5′ monophosphate dehydrogenase
10 SCO4797* uvrD Probable ATP-dependent DNA helicase S18 SCO4833 Phosphorylmutase8 SCO4834 Hypothetical protein8 SCO4835* Conserved hypothetical protein with alkylhydroperoxidase-like AhpD/CMD domain2 SCO4956* msrA Probable peptide methionine S-sulphoxide reductase (S1, A), S1
14 SCO4966* Possible membrane protein1 SCO4967* mca Mycothiol-S-conjugate amidase (S1, A)1 SCO4968 Hypothetical protein (A)9 SCO5042* fumC Fumarate hydratase, class II family (iron-independent) (A)6 SCO5065* Possible transcriptional regulator, LuxR family
14 SCO5163* Unknown (S1)11 SCO5178* moeB Possible sulphurylase related to molybdopterin synthesis (S1, A)1 SCO5187* mrxA Glutaredoxin-like protein, probable myc917oredoxin (S1, A)
10 SCO5188* uvrD2 ATP-dependent DNA helicase (UvrD/REP type), divergent from mrxA (5187)6 SCO5216* sigR ECF (group 4) sigma factor sR (S1, A)6 SCO5217 rsrA ZAS (zinc-containing antisigma) factor for sR (A)
14 SCO5284* Hypothetical protein5 SCO5285* lon ATP-dependent protease (S1)6 SCO5357* rho Transcription termination factor, an RNA–DNA helicase3 SCO5359* rpmE1 Ribosomal proteins L31 (S1, A)3 SCO5360 prfA Peptide release factor A3 SCO5361 hemK Probable release factor-specific methyltransferase8 SCO5465* Conserved hypothetical, probable F420-dependent NADP oxidoreductase (S1, A)
14 SCO5490* Hypothetical protein14 SCO5545* hypothetical protein (A)6 SCO5552* ndgR IclR family transcription regulator3 SCO5705* Hypothetical protein, first gene in translation-related gene cluster S13 SCO5706 infB Translation initiation factor IF-23 SCO5707 Hypothetical protein3 SCO5708 rbfA Ribosome binding factor
4 M.-S. Kim et al. �
© 2012 Blackwell Publishing Ltd, Molecular Microbiology

mycoredoxin (SCO3442), and a eukaryotic-type protea-some system that degrades proteins ligated with prokary-otic ubiquitin-like protein Pup (Mpa, PafD, Pup, PrcAB) firstreported in M. tuberculosis (Pearce et al., 2008; Burns andDarwin, 2010). This gene cluster encodes a putative AAAATPase (Mpa), an accessory factor (PafD), prokaryoticubiquitin-like protein (Pup), 20S proteasome alpha andbeta subunits (PrcAB) similar to those encoded by Myco-bacterium tuberculosis (Pearce et al., 2008; Darwin,2009). S1 mapping analysis demonstrates that mpa(SCO1648) has a SigR-dependent promoter, which isdiamide-induced, in addition to ones that are independentof SigR (Fig. 1). Since the thiol-oxidant diamide can causeprotein misfolding and aggregation (Kallifidas et al., 2010),induction of proteolytic systems by diamide-triggered SigRmatches biological necessity.
We also found a large number of SigR target genes thatmay function in modulating ribosome-associated pro-cesses (9 operons; group 3). Newly identified genes in this
group encode probable translation initiation factors (IF-2,IF-3), 50S ribosomal proteins (L20, L35), ribosome-associated GTP-binding proteins (EngA, ObgE, Era), aribosome binding factor RbfA, and a tRNA pseudouridinesynthase (TruB). Observation of many ribosome-associated genes in the SigR regulon leads us to postulatethat thiol-oxidative stress may somehow hinder translationprocesses, against which cells induce a SigR-mediatedresponse. Induction of tmRNA (ssrA) by SigR under thiol-oxidative stress suggests an increase in truncated mRNAin the A-site of ribosome in the cell. It has been shown inE. coli that ribosome stalling induces A-site bound RelE orRelE-like RNA interferases to cut mRNA (Neubauer et al.,2009; Christensen-Dalsgaard et al., 2010), whose contin-ued translation requires tmRNA that adds the SsrA tag tothe truncated protein for degradation. In this respect, arecent discovery of the role of tmRNA in relieving polyribo-somal stalling in dnaK and other stress-related mRNA tofacilitate their translation in streptomycetes highlights a
Table 1. cont.
Ga Numberb Name Description Confirmc
3 SCO5709 truB Probable tRNA pseudouridine synthase10 SCO5754* cinA Competence-DNA damage-induced protein, neighbouring clgR (5755) (S1, A)3 SCO5796* hflX Conserved ribosome binding GTPase (S1, A)6 SCO5820* hrdB Major vegetative sigma factor S1
14 SCO5864* Hypothetical protein14 SCO5865 Hypothetical protein2 SCO6061* msrB Probable peptide methionine R-sulphoxide reductase (S1, A), S1
14 SCO6126* Hypothetical protein13 SCO6127* Probable carboxylesterase, probable phenylcarbamate hydrolase11 SCO6423* Putative lipoate-protein ligase (A)8 SCO6551* Probable oxidoreductase, aldo/keto reductase family (S1, A)
12 SCO6759* phyA/hopE Probable phytoene (hopanoid) synthase (S1, A), S112 SCO6760 phyB/hopD Squalene/phytoene synthase12 SCO6761 Small (52 aa) hypothetical protein of unknown function12 SCO6762 phyC/hopC Squalene/phytoene dehydrogenase12 SCO6763 phyD/hopB Polyprenyl diphosphate synthase12 SCO6764 phyE/hopA Squalene–hopene cyclase12 SCO6765 Lipoprotein12 SCO6766 Hypothetical protein12 SCO6767 ispG 4-hydroxy-3-methyl-2-en-1-yl diphosphate synthase12 SCO6768 1-deoxy-D-xylulose-5-phospate synthase12 SCO6769 Aminotransferase12 SCO6770 DNA-binding protein12 SCO6771 Small hydrophobic protein6 SCO6775* Probable transcriptional regulator, TetR family
14 SCO6776* Unknown6 SCO7140* Probable DNA-binding protein
14 SCO7631* Possible secreted protein1 SCO7632* Probable amidase similar to mshB and mca (A), S18 SCO7784* Possible oxidoreductase, probable nitro/flavin reductase S18 SCO7785 Transcriptional regulator
a. Genes were grouped according to predicted functions such as thiol homeostasis (1), sulphur metabolism (2), modulation of ribosome and/ortranslation (3), guanine nucleotide metabolism (4), protein degradation (5), transcriptional regulators or DNA binding proteins (6), transporter orintegral membrane proteins (7), oxidoreductases (8), energy metabolism (9), DNA damage repair and/or recombination (10), cofactor metabolism(11), lipid metabolism (12), others (13), or function unpredictable (14).b. Genes with SigR-binding promoters were marked with asterisks (*).c. The genes whose SigR-dependent expression was previously confirmed by S1 or microarray (A) were marked with S1 and A in parenthesis(Paget et al., 2001; Park and Roe, 2008; Kallifidas et al., 2010). Those confirmed in this study were noted with S1.
Conserved thiol-oxidative stress in actinomycetes 5
© 2012 Blackwell Publishing Ltd, Molecular Microbiology
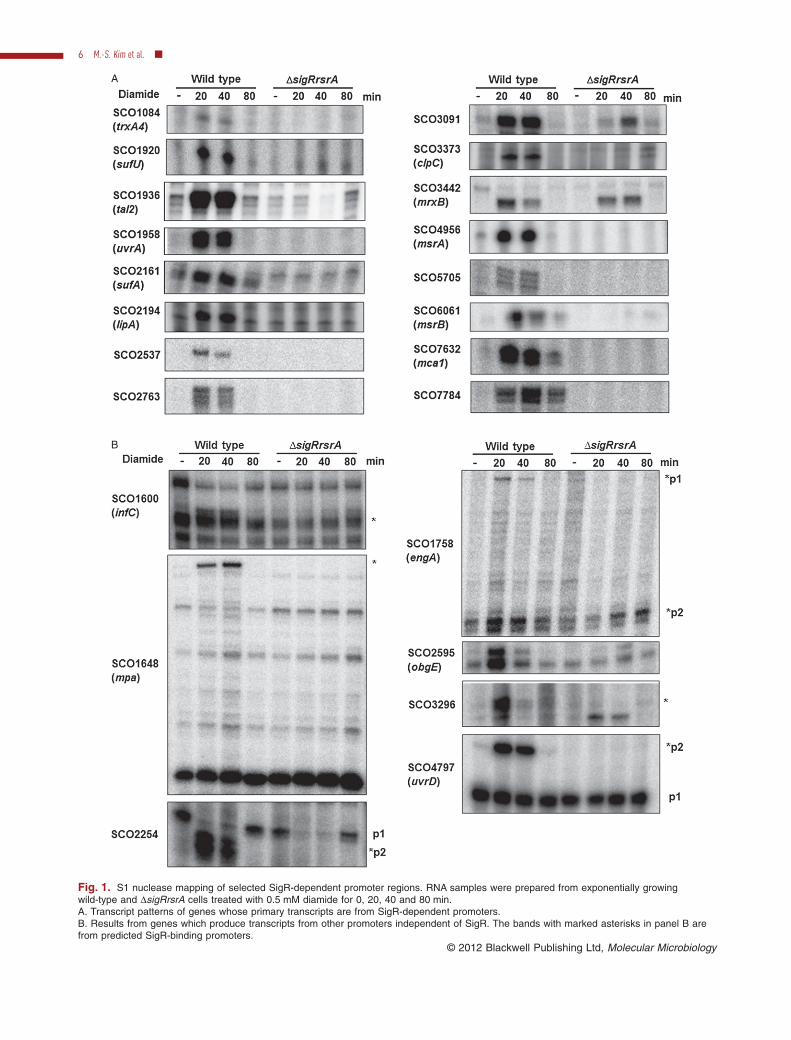
A
B
Fig. 1. S1 nuclease mapping of selected SigR-dependent promoter regions. RNA samples were prepared from exponentially growingwild-type and DsigRrsrA cells treated with 0.5 mM diamide for 0, 20, 40 and 80 min.A. Transcript patterns of genes whose primary transcripts are from SigR-dependent promoters.B. Results from genes which produce transcripts from other promoters independent of SigR. The bands with marked asterisks in panel B arefrom predicted SigR-binding promoters.
6 M.-S. Kim et al. �
© 2012 Blackwell Publishing Ltd, Molecular Microbiology

potential new link between ribosomal modulation andstress response (Barends et al., 2010). Increased demandfor guanine nucleotides due to elevation in activity of RelAand GTP-binding proteins during stress could be met byincreased GMP synthesis by guaB and guaB1 gene prod-ucts. Increased demand for guanine nucleotide can also beexplained by the fact that guanine is most vulnerable tooxidation among four DNAbases due to its lower oxidation-reduction potential (Kawanishi et al., 2001).
Many transcription factors (15 operons; group 6) arepredicted to be SigR targets, implying that SigR is a globalregulator that can change the transcription profile throughcascades of affected transcriptional regulators. Amongnewly identified SigR targets are those encoding themajor housekeeping sigma factor HrdB (SCO5820) and aSigR-like ECF sigma factor SigR2 (SCO3450) and itsputative partner RsrA2-2 (SCO3451). The gene for anIclR-like transcription factor (SCO5552) called ‘NdgR’ thatregulates amino-acid-dependent growth and antibioticproduction (Yang et al., 2009) also contains a SigR-binding promoter sequence.
Synthesis of a number of redox-sensitive cofactors (suchas Fe-S, CoA, NAD, lipoate, folic acid, pyridoxal phosphateand molybdopterin) appears to be regulated by SigR (9operons; group 11). In addition to the previously identifiedsufA, coaE, folE and moeB genes [encoding a putativeFe-S assembly factor, coenzyme A kinase, GTP cyclohy-drolase I for folate biosynthesis, and sulphurylase possiblyinvolved in molybdopterin synthesis (Paget et al., 2001)respectively], we found that sufU, nadA, lipA and pdxH2/3[encoding putative NifU-type Fe-S assembly factor, quino-linate synthetase A subunit for nicotinamide synthesis,lipoic acid synthase, and phospho-pyridoxamine syn-thetase for pyridoxal phosphate (vitamin B6)] were alldirect targets of SigR. It is interesting to note that some ofthese proteins are either putative Fe-S assembly compo-nents (SufA, SufU) or Fe-S containing enzymes (NadA,LipA). In addition, the cofactors they synthesize containsulphur atoms (Fe-S, folate, CoA, lipoic acid), or involvesulphur-mediated reactions (LipA, MoeB). Exposed Fe-Scofactors are labile to oxidation, so we propose thatincreased SigR activity contributes to replenishingFe-S cofactors, Fe-S-containing proteins, and/or otheroxidation-labile S-containing factors upon oxidative stress.
S1 nuclease mapping confirms predictedSigR promoters
To provide additional experimental evidence for some ofthe newly identified SigR targets, we examined 25 selectedtranscripts by S1 nuclease mapping at different times afterexposing S. coelicolor cells to the thiol oxidant diamide. Weobserved SigR-specific transcripts from 24 promoterswhose 5′ ends match with the predicted location of the
promoters (Table S1). This supports the hypothesis thatnearly all the SigR regulon genes presented in Table 1produce SigR-dependent transcripts. We detected newSigR-dependent transcripts from 15 promoters whoseexpression has not been previously reported (SCO1084,1600, 1648, 1758, 1936, 2194, 2254, 2595, 2763, 3296,3442, 4797, 5705, 5820, 7784), confirming that ChIP-chipanalysis detects SigR target genes with high sensitivity.Figure 1 shows S1 mapping analysis of transcripts fromthese promoters. Some genes produce transcripts initiatedprimarily from SigR-dependent promoters (Fig. 1A; 1084,1920, 1936, 1958, 2161, 2194, 2537, 2763, 3091, 3373,4956, 5705, 6061, 7632, 7784). Others utilize additionalSigR-independent promoters (Fig. 1B; 1600, 1758, 2254,2595, 3296, 4797).
Examination of genes with multiple promoters revealsdiverse patterns of SigR-dependent modulation. Forexample, SCO1758 (engA) encoding a putative GTPaseproduces p1 transcript as predicted (250 nt upstream fromthe start codon; Table S1) and also the downstreamunpredicted p2 transcripts, whose promoter sequence(GGAT-N16-GTT; ggatcacccggtaaaggggtgtt) resemblesthe SigR consensus but with a shorter spacing of 16nucleotides. This implies a flexibility of spacing between-35 and -10 elements of SigR-dependent promoters. Insome genes such as SCO2254 the dominant transcriptsproduced under non-stressed condition disappear tran-siently following stress independent of SigR, during whichperiod the SigR-dependent transcripts are induced.
In promoters such as p2 of SCO1758 (engA; Fig. 1B)and SCO3442 (mrxB; Fig. 1A) (encoding a paralogue ofmycoredoxin and a glutaredoxin-like protein) the SigR-induced transcripts still appear in the DsigR-rsrA mutantbut with delayed response. A similar phenomenon wasalso observed for SCO3091 (cfa) encoding a putativecyclopropane-fatty acyl phospholipid synthase (Fig. 1A).The delayed induction kinetics in DsigR-rsrA mutant sug-gests that these SigR-dependent promoters may also berecognized by SigR paralogues that recognize similar pro-moter sequences as SigR. Similar induction pattern hasbeen reported for transcripts encoding Lon protease,which are induced rapidly by SigR but with slower kineticsin the absence of SigR (Kallifidas et al., 2010). SCO3450and SCO3451, which are SigR regulated genes encodinghomologues of SigR (SigR2) and RsrA (RsrA2-1), respec-tively, show a similar pattern of induction (data notshown). A likely candidate for a sigma factor that sharessimilar promoter recognition sequence with SigR isSCO5147 (SigR1), which is a close homologue of SigEfrom M. tuberculosis (MtbSigE) as exemplified in the phy-logenic comparison of ZAS-linked ECF sigma factors fromS. coelicolor and M. tuberculosis (Fig. S2). The promoterfor sigR1 has GGAAC-N18-GTT sequence that is notbound by SigR in our ChIP analysis. It is induced by
Conserved thiol-oxidative stress in actinomycetes 7
© 2012 Blackwell Publishing Ltd, Molecular Microbiology
diamide with slower kinetics (data not shown), and wepropose that it is recognized by SigR1 itself. These obser-vations indicate that a subset of SigR regulon genes canbe recognized by at least one closely related SigR para-logue in addition to or in the absence of SigR.
SigR maintains the level and activity of thehousekeeping sigma factor HrdB duringthiol-oxidative stress
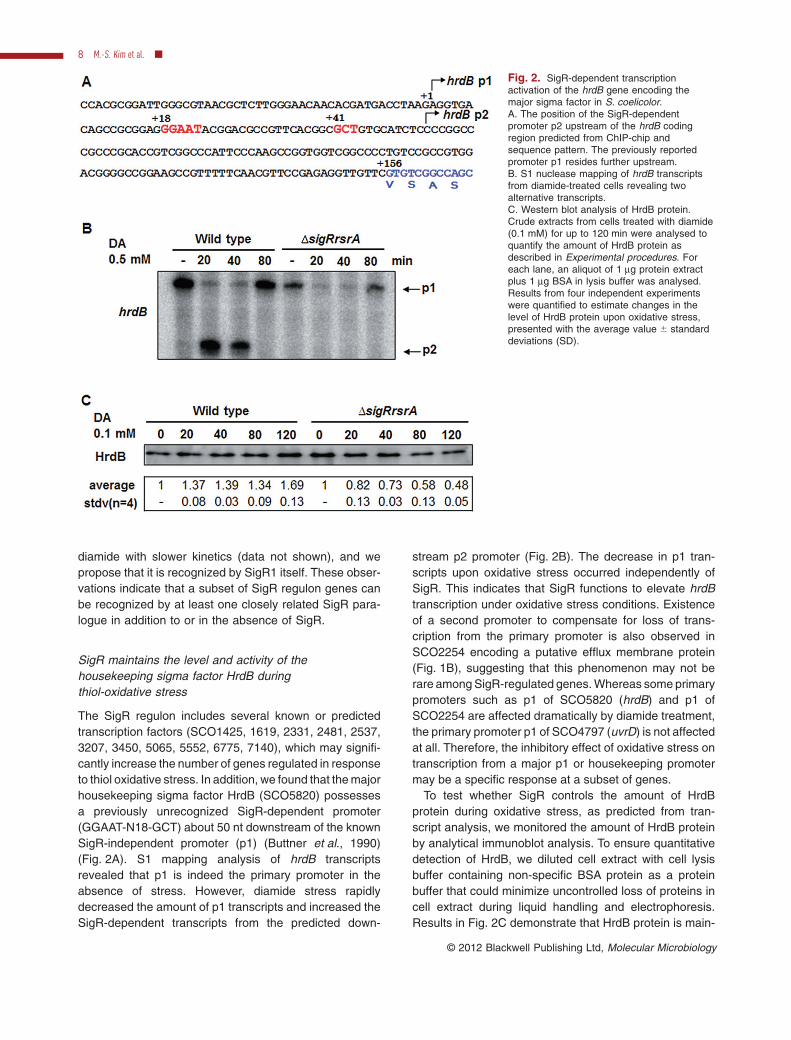
The SigR regulon includes several known or predictedtranscription factors (SCO1425, 1619, 2331, 2481, 2537,3207, 3450, 5065, 5552, 6775, 7140), which may signifi-cantly increase the number of genes regulated in responseto thiol oxidative stress. In addition, we found that the majorhousekeeping sigma factor HrdB (SCO5820) possessesa previously unrecognized SigR-dependent promoter(GGAAT-N18-GCT) about 50 nt downstream of the knownSigR-independent promoter (p1) (Buttner et al., 1990)(Fig. 2A). S1 mapping analysis of hrdB transcriptsrevealed that p1 is indeed the primary promoter in theabsence of stress. However, diamide stress rapidlydecreased the amount of p1 transcripts and increased theSigR-dependent transcripts from the predicted down-
stream p2 promoter (Fig. 2B). The decrease in p1 tran-scripts upon oxidative stress occurred independently ofSigR. This indicates that SigR functions to elevate hrdBtranscription under oxidative stress conditions. Existenceof a second promoter to compensate for loss of trans-cription from the primary promoter is also observed inSCO2254 encoding a putative efflux membrane protein(Fig. 1B), suggesting that this phenomenon may not berare among SigR-regulated genes. Whereas some primarypromoters such as p1 of SCO5820 (hrdB) and p1 ofSCO2254 are affected dramatically by diamide treatment,the primary promoter p1 of SCO4797 (uvrD) is not affectedat all. Therefore, the inhibitory effect of oxidative stress ontranscription from a major p1 or housekeeping promotermay be a specific response at a subset of genes.
To test whether SigR controls the amount of HrdBprotein during oxidative stress, as predicted from tran-script analysis, we monitored the amount of HrdB proteinby analytical immunoblot analysis. To ensure quantitativedetection of HrdB, we diluted cell extract with cell lysisbuffer containing non-specific BSA protein as a proteinbuffer that could minimize uncontrolled loss of proteins incell extract during liquid handling and electrophoresis.Results in Fig. 2C demonstrate that HrdB protein is main-
Fig. 2. SigR-dependent transcriptionactivation of the hrdB gene encoding themajor sigma factor in S. coelicolor.A. The position of the SigR-dependentpromoter p2 upstream of the hrdB codingregion predicted from ChIP-chip andsequence pattern. The previously reportedpromoter p1 resides further upstream.B. S1 nuclease mapping of hrdB transcriptsfrom diamide-treated cells revealing twoalternative transcripts.C. Western blot analysis of HrdB protein.Crude extracts from cells treated with diamide(0.1 mM) for up to 120 min were analysed toquantify the amount of HrdB protein asdescribed in Experimental procedures. Foreach lane, an aliquot of 1 mg protein extractplus 1 mg BSA in lysis buffer was analysed.Results from four independent experimentswere quantified to estimate changes in thelevel of HrdB protein upon oxidative stress,presented with the average value � standarddeviations (SD).
8 M.-S. Kim et al. �
© 2012 Blackwell Publishing Ltd, Molecular Microbiology

tained during oxidative stress condition at a level slightlyhigher than that under non-stressed condition in the wildtype. On the contrary, the level of HrdB decreased signifi-cantly in DsigRrsrA mutant upon oxidative stresscondition. Even though it is not clear why the level of HrdBprotein increases in the wild type during the stress, it isobvious that SigR does contribute to maintain the level ofHrdB protein, as predicted from transcript analysis. Wefurther monitored the activity of HrdB by examining tran-scripts from HrdB target promoters (rrnDp1, p3 and p4) inthe ribosomal RNA operon rrnD, in the wild type andmutant. All four promoters of the rrnD operon share theconsensus sequence for HrdB recognition, especially inthe -10 region, to varying extent (Baylis and Bibb, 1988;Hahn and Roe, 2007). In vitro transcription from rrnDpromoters with reconstituted holoenzyme containingHrdB (EsHrdB) has demonstrated that all four promoterscan be transcribed by EsHrdB, even though transcriptionfrom p3 and p4 promoters requires additional cellularfactors (Hahn and Roe, 2007). We found that the amountsof the abundant p3 and p4 transcripts were maintainedthroughout the stress period and even increased up to50% and 30%, respectively, during 40–80 min diamidetreatment. On the other hand, they both decreased con-tinuously in the DsigRrsrA mutant to about 40% and 60%level, respectively, during the 2 h of stress (Fig. S3). Tran-scripts from the p1 promoter decreased to 20% levelduring the stress in the mutant, whereas the level wasmaintained in the wild type (Fig. S3). These results clearlydemonstrate that SigR is necessary to maintain the leveland activity of the major housekeeping sigma factorduring thiol-oxidative stresses.
The UvrA protein is necessary to cope with UV andthiol-oxidative stresses
Among the newly discovered SigR targets are SCO1958,SCO4797 and SCO5188, which encode putative UvrA,UvrD and a paralogue of UvrD (UvrD2), respectively, pre-dicted to function in nucleotide excision repair. TheUvrABC endonuclease complex with UvrD (DNA helicaseII) has been shown to repair UV-induced DNA damages,the most prominent of which are pyrimidine dimers (Sinhaand Hader, 2002). However, whether the UvrABC systemis needed under thiol-oxidative stress condition andwhether thiol-oxidative stress causes DNA damage havenot been known. Consensus SigR promoters (GGCAT-N18-GTT for uvrA, GGCAT-N18-GTT for uvrD, GGAAA-N19-GTC for uvrD2) are located 103, 31 and 7 nt upstreamfrom the start codons of respective genes (Table S1). TheuvrA gene is transcribed at a very low level underunstressed conditions. However, upon diamide stress, it isdrastically induced from a SigR-dependent promoter(Fig. 1A). On the contrary, the uvrD gene is transcribed at
a relatively high level from a SigR-independent promoterp1 during unstressed condition. Upon induction, theupstream SigR-dependent promoter p2 is induced(Fig. 1B). The uvrD2 transcripts were not detectable by S1analysis, probably due to low amount.
In order to examine whether UvrA indeed confers UVresistance, and whether SigR and UvrA have relatedfunctions in conferring resistance to UV and thiol-reactivecompounds in S. coelicolor, we made a DuvrA strain andanalysed its sensitivity to UV and thiol-reactive com-pounds along with DsigR-rsrA mutant. Figure 3 demon-strates that DuvrA has increased sensitivity to UVirradiation, and DsigR-rsrA is also sensitive to UV, thoughnot as dramatically as DuvrA. Therefore, SigR is neces-sary to achieve optimal UV resistance possibly throughinducing UvrA system. The SigR regulon genes were notinduced by UV irradiation (data not shown), suggestingthat UV itself is not capable of activating SigR function.SigR can be activated by thiol-reactive oxidants/electrophiles such as diamide and methyl hydroquinone(MHQ), as predicted from the induction of thiol-specificoxidative stress response by these compounds in B. sub-tilis (Antelmann et al., 2008; Nguyen et al., 2009). Sincesome electrophiles can react not only with thiols in pro-teins and small molecules but also with DNA, we exam-ined whether UvrA is needed to cope with chemicalstresses caused by thiol-reactive electrophiles such asdiamide, methyl hydroquinone (MHQ) and methyl glyoxal(MG). It has been reported that MG induces not onlythiol-specific oxidative stress response but also SOSresponse, suggesting some DNA damage in B. subtilis.(Nguyen et al., 2009). MG is known to cause DNAdamage by reacting with DNA bases to form N 2-(1-carboxylethyl)-2′-deoxyguanosine (CEdG) (Synold et al.,2008). Whether this kind of damage is repaired by theUvr system has not been reported. Whether thiol-reactivediamide and MHQ could elicit DNA damage has not beenreported either. The result in Fig. 3 shows that UvrA isneeded to defend cells against these electrophiles. Asexpected, DsigR-rsrA mutant became very sensitive tothiol-reactive electrophiles. Dramatic resistance of DsigR-rsrA toward MG is unexpected. Inability to induce manythiol-homeostatic components may somehow conferbetter survival relative to the wild type in the presence ofMG in plate culture. When the sensitivity of DsigR-rsrAtoward MG was monitored in liquid culture during earlyexponential phase (OD ~ 0.3), the mutant was more sen-sitive to MG than the wild type, suggesting that the resis-tance phenotype on plate culture could depend ontreatment method (data not shown). Overall, the findingthat SigR induces DNA repair proteins expands ourunderstanding of the function of SigR to accommodateresponse towards UV- and some electrophile-incurreddamages.
Conserved thiol-oxidative stress in actinomycetes 9
© 2012 Blackwell Publishing Ltd, Molecular Microbiology

The phylogenetic analysis of SigR orthologues and theirpredicted respective regulons reveals a core conservedthiol oxidative stress regulon in Actinomycetes
The characterization of this larger SigR regulon inS. coelicolor furthers our understanding of the organ-ism’s response to thiol oxidative stress. However, it hasbeen previously observed that the composition of agiven regulon in a given organism may result from spe-cific adaptations to its ecological niche (Dufour et al.,2008; 2010a; Perez and Groisman, 2009). Therefore,the SigR regulon in S. coelicolor may comprise manyfunctions that are specific only to this organism, and notshared by other organisms. Therefore, comparison ofpredicted SigR regulons in other organisms may indicatefunctions that are commonly required to cope with thioloxidative stress in general apart from those specificallyrequired for this organism. To test this hypothesis andpredict which genes found in the S. coelicolor SigRregulon are most likely to be directly relevant to thioloxidative stress, we performed a phylogenetic analysisof SigR-RsrA orthologues and their regulons across agroup of Actinomycetes using a computational methodpreviously described (Dufour et al., 2008; 2010a). Sincethis phylogenetic analysis relies on the accurate compu-tational detection of putative SigR promoter sequences,we constructed a refined SigR promoter model by align-ing the sequences of predicted promoters correspondingto 57 enriched regions in the ChIP-chip assay thatmatches with genes transcribed in a SigR-dependentmanner (Table S1). Figure 4A demonstrates a new SigR
consensus logo, with a representative sequence ofGGAAT-N18-19-GTT.
We first detected and aligned the protein sequences ofall SigR orthologues found in 42 Actinomycetes to deter-mine the conservation of their DNA binding domains (onlyMycobacterium leprae TN did not possess SigR and RsrAorthologues). The multiple sequence alignment showedthat the protein domains determining promoter sequencespecificity (regions s2.4 and s4.2) of SigR orthologuesare well conserved across all species (Fig. S1). There-fore, we assumed that all SigR orthologues recognizevery similar if not identical promoter sequences. Thus, weidentified putative SigR targets across all the selectedgenomes by searching in each genome for sequencesmatching S. coelicolor SigR promoter model (Fig. 4A) inthe upstream of annotated genes. With the promoterscore cut-off chosen for this analysis, we captured 72 ofthe 108 SigR promoters defined by our ChIP-chip analysisin S. coelicolor. Finally, putative SigR target genes acrossthe 42 genomes were grouped by orthology and clusteredaccording to the occurrence of predicted SigR-dependentpromoters.
The emerging phylogenetic patterns predicted that thetop 25 sets of orthologous genes appear to be regulated bySigR orthologues across most of the 42 Actinomycetes,thus, forming a core conserved regulon for the response tothiol oxidative stress (Fig. 4B, Table 2). The predictionsresulting from this comparative genomics analysis are wellsupported by the experimental data presented in this reportbecause 20 of the S. coelicolor genes corresponding to thetop 25 conserved SigR targets are associated with SigR
Fig. 3. Sensitivity of DuvrA and DsigRrsrA mutants to UV and thiol-reactive oxidants and electrophiles. Serially diluted spores of wild-type(WT), DsigRrsrA, DuvrA strains were spotted on NA plates with or without added chemicals such as diamide (0.6 mM), methyl hydroquinone(MHQ, 1 mM) and methyl glyoxal (MG, 1 mM). For UV irradiation, the spotted plates were exposed to UV light from UV cross-linker for 30seconds. Plates were incubated at 30°C for 1 (untreated), 2 (UV, diamide), 5 (MHQ) or 6 days (MG).
10 M.-S. Kim et al. �
© 2012 Blackwell Publishing Ltd, Molecular Microbiology
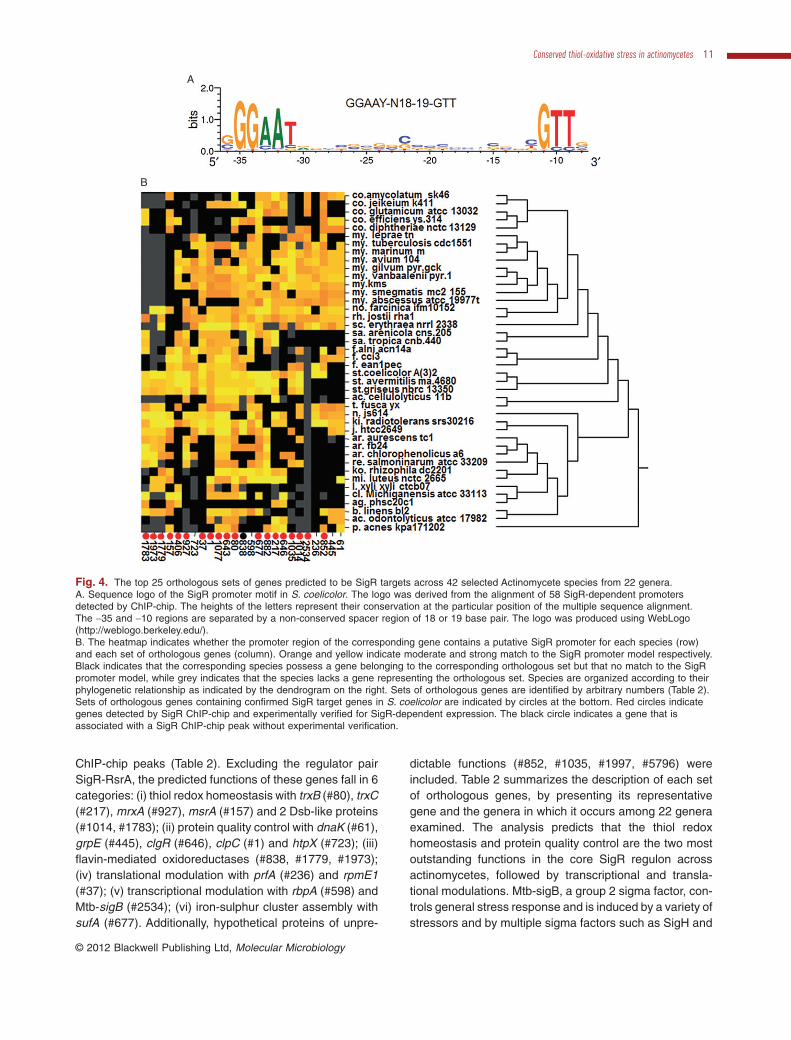
ChIP-chip peaks (Table 2). Excluding the regulator pairSigR-RsrA, the predicted functions of these genes fall in 6categories: (i) thiol redox homeostasis with trxB (#80), trxC(#217), mrxA (#927), msrA (#157) and 2 Dsb-like proteins(#1014, #1783); (ii) protein quality control with dnaK (#61),grpE (#445), clgR (#646), clpC (#1) and htpX (#723); (iii)flavin-mediated oxidoreductases (#838, #1779, #1973);(iv) translational modulation with prfA (#236) and rpmE1(#37); (v) transcriptional modulation with rbpA (#598) andMtb-sigB (#2534); (vi) iron-sulphur cluster assembly withsufA (#677). Additionally, hypothetical proteins of unpre-
dictable functions (#852, #1035, #1997, #5796) wereincluded. Table 2 summarizes the description of each setof orthologous genes, by presenting its representativegene and the genera in which it occurs among 22 generaexamined. The analysis predicts that the thiol redoxhomeostasis and protein quality control are the two mostoutstanding functions in the core SigR regulon acrossactinomycetes, followed by transcriptional and transla-tional modulations. Mtb-sigB, a group 2 sigma factor, con-trols general stress response and is induced by a variety ofstressors and by multiple sigma factors such as SigH and
A
B
Fig. 4. The top 25 orthologous sets of genes predicted to be SigR targets across 42 selected Actinomycete species from 22 genera.A. Sequence logo of the SigR promoter motif in S. coelicolor. The logo was derived from the alignment of 58 SigR-dependent promotersdetected by ChIP-chip. The heights of the letters represent their conservation at the particular position of the multiple sequence alignment.The -35 and -10 regions are separated by a non-conserved spacer region of 18 or 19 base pair. The logo was produced using WebLogo(http://weblogo.berkeley.edu/).B. The heatmap indicates whether the promoter region of the corresponding gene contains a putative SigR promoter for each species (row)and each set of orthologous genes (column). Orange and yellow indicate moderate and strong match to the SigR promoter model respectively.Black indicates that the corresponding species possess a gene belonging to the corresponding orthologous set but that no match to the SigRpromoter model, while grey indicates that the species lacks a gene representing the orthologous set. Species are organized according to theirphylogenetic relationship as indicated by the dendrogram on the right. Sets of orthologous genes are identified by arbitrary numbers (Table 2).Sets of orthologous genes containing confirmed SigR target genes in S. coelicolor are indicated by circles at the bottom. Red circles indicategenes detected by SigR ChIP-chip and experimentally verified for SigR-dependent expression. The black circle indicates a gene that isassociated with a SigR ChIP-chip peak without experimental verification.
Conserved thiol-oxidative stress in actinomycetes 11
© 2012 Blackwell Publishing Ltd, Molecular Microbiology

Tab
le2.
Top
25ge
nes
that
are
high
lyco
nser
ved
asS
igR
regu
lon
mem
bers
amon
gac
tinom
ycet
es.
Ort
holo
gues
No.
Gen
eN
o.a
Gen
ena
me
Fun
ctio
nG
enus
(fre
quen
cy)b
61S
CO
3671
dnaK
Hsp
70fa
mily
chap
eron
e,no
dire
ctS
igR
targ
etin
Sco
,F
irst
gene
indn
aK-g
rpE
-dna
J-hs
pRcl
uste
r(3
671-
3668
),a
Sig
Eta
rget
inM
tbC
o,M
y,N
o,R
h,S
c,F,
T,N
,K
i,J,
Mi,
B,A
c,P
(14/
22)
445
SC
O36
70gr
pEC
o-ch
aper
one
Co,
My,
No,
Rh,
Sc,
F,T,
N,
Ki,
J,,
B,A
c(1
2/22
)85
2S
CO
3083
*P
ossi
ble
inte
gral
mem
bran
epr
otei
nC
o,M
y,N
o,R
h,S
c,F,
St,
T,N
,K
i,B
,P
(12/
22)
236
SC
O53
60*
prfA
Pep
tide
chai
nre
leas
efa
ctor
1(R
F-1
),rp
mE
1-pr
fAop
eron
My,
Rh,
Sc,
F,S
t,T,
N,
Ki(
8/22
)25
34M
T27
83M
tb-s
igB
Sig
ma
fact
orS
igB
inM
.tub
ercu
losi
sre
gula
ted
byS
igE
Co,
My,
No,
Rh,
Sc
(5/2
2)10
14S
CO
2634
*H
ypot
hetic
alpr
otei
n,co
ntai
nsD
sbA
-like
thio
redo
xin
dom
ain
Co,
My,
No,
Rh,
Sc,
F,S
t,T,
N,A
r,R
e(1
1/22
)10
35S
CO
3187
*H
ypot
hetic
alpr
otei
nM
y,N
o,R
h,F,
St,
J,A
r,R
e(8
/22)
646
SC
O57
55*
clgR
Act
ivat
orof
clpP
and
lon
prot
ease
synt
hetic
gene
s.N
eigh
bour
ing
aS
igR
-tar
get
gene
(575
4;ci
nA)
Co,
My,
No,
Rh,
Sc,
St,
N,
Ki,
Ar,
Ko,
Mi,
Cl,
Ag,
Ac,
P(1
5/22
)
217
SC
O08
85*
trxC
Thi
ored
oxin
Co,
My,
No,
Rh,
Sa,
St,
Ac,
N,
Ki,
Ko,
Mi,
Ag,
Ac,
P(1
4/22
)88
2S
CO
1997
*U
nkno
wn
Co,
My,
No,
Rh,
Sa,
F,S
t,J,
Ar,
Re,
Ko,
Mi,
P(1
3/22
)67
7S
CO
2161
*su
fAH
esB
dom
ain,
prob
able
Iron
-sul
phur
clus
ter
asse
mbl
ypr
otei
nC
o,M
y,N
o,R
h,S
c,S
a,S
t,T,
N,
Ki,
J,A
r,R
e,K
o,M
i,C
l,B
(17/
22)
598
SC
O14
21rp
bAR
NA
poly
mer
ase
bind
ing
prot
ein
Co,
My,
No,
Rh,
Sc,
Sa,
F,S
t,A
c,T,
N,
Ki,
J,A
r,R
e,K
o,M
i,B
(18/
22)
838
SC
O28
16#
Con
serv
edhy
poth
etic
alpr
otei
n,F
lavi
nut
ilizi
ngm
onoo
xyge
nase
supe
rfam
ilyC
o,M
y,N
o,R
h,S
c,F,
St,
Ki,
J,A
r,K
o,M
i,C
l,B
(14/
22)
80S
CO
3890
*tr
xBT
hior
edox
inre
duct
ase
Co,
My,
No,
Rh,
Sc,
Sa,
F,S
t,A
c,T,
Ki,
J,A
r,R
e,K
o,M
i,L,
Cl,
B,A
c,P
(21/
22)
643
SC
O52
16*
sigR
Sig
ma
fact
orR
Co,
My,
No,
Rh,
Sc,
Sa,
F,S
t,A
c,T,
N,
Ki,
J,A
r,R
e,K
o,M
i,C
l,B
,Ac,
P(2
1/22
)10
77S
CO
5217
*rs
rAA
nti-S
igR
fact
orC
o,M
y,N
o,R
h,S
c,S
a,F,
St,
Ac,
T,N
,K
i,J,
Ar,
Re,
Ko,
Mi,
Cl,
Bl,
P(2
0/22
)1
SC
O33
73*
clpC
Clp
-fam
ilyA
TP
-bin
ding
prot
ease
sim
ilar
toM
ecB
/Clp
CC
o,M
y,N
o,R
h,S
c,S
a,F,
St,
Ac,
T,N
,K
i,J,
Mi,
L,C
l,B
,Ac
(18/
22)
37S
CO
5359
*rp
mE
1R
ibos
omal
prot
eins
L31
My,
Rh,
Sc,
Sa,
F,S
t,T,
N,
Ki,
J(1
0/22
)72
3S
CO
4609
htpX
Pos
sibl
epe
ptid
ase,
M.t
uber
culo
sis
Htp
Xho
mol
ogue
,no
Sig
Rta
rget
,a
Sig
Eta
rget
inM
tbM
y,N
o,R
h,S
c,F,
St,
N,
Ki,
J,A
r(1
0/22
)
927
SC
O51
87*
mrx
AM
ycor
edox
in,
glut
ared
oxin
-like
prot
ein
My,
No,
Rh,
Sa,
F,S
t,A
c,N
,K
i,A
r,R
e(1
1/22
)40
6S
CO
5796
*U
nkno
wn
Co,
My,
Sa,
F,S
t,T,
Ki,
L,C
l,A
g(1
0/22
)15
7S
CO
4956
*m
srA
Met
hion
ine
S-s
ulph
oxid
ere
duct
ase
Co,
No,
F,S
t,N
,K
i,J,
Ar,
Ko,
Mi,
Cl,
Ag,
B,
P(1
4/22
)17
79S
CO
3296
*P
ossi
ble
oxid
ored
ucta
sesi
mila
rto
F42
0-de
pend
ent
alco
hold
ehyd
roge
nase
No,
Rh,
Sc,
St,
T,N
,K
i,J,
Ar,
Ko,
Mi,
L,B
(13/
22)
1973
SC
O32
95*
Pos
sibl
eox
idor
educ
tase
cont
aini
ngfla
vin
No,
St,
T,N
,K
i,J,
Ar,
Re,
Mi,
L,B
(11/
22)
1783
SC
O18
69*
Con
serv
edhy
poth
etic
alpr
otei
n,pr
obab
leD
sbA
-like
dith
iol-d
isul
phid
eis
omer
ase
(Frn
Esu
bfam
ily)
Co,
Rh,
Sa,
F,S
t,T,
N,
Ki,
J,A
r,R
e,C
l,A
g,B
(14/
22)
a.T
here
pres
enta
tive
gene
from
S.c
oelic
olor
(SC
O)
orM
.tub
ercu
losi
s(M
T)
inea
chor
thol
ogue
clus
ter
was
pres
ente
d.T
heS
CO
gene
sth
atw
ere
dete
rmin
edto
beS
igR
-dep
ende
ntby
ChI
P-c
hip
and
supp
orte
dby
addi
tiona
lS1
and
mic
roar
ray
data
are
mar
ked
with
aste
risks
(*).
The
gene
sde
tect
edby
ChI
P-c
hip
but
not
byad
ditio
nale
xpre
ssio
nda
taar
em
arke
dw
ith‘#
’.b
.T
hege
nera
that
poss
ess
the
corr
espo
ndin
gS
igR
targ
etge
nein
each
orth
olog
ous
clus
ter
are
pres
ente
dw
ithab
brev
iatio
ns;
Co
(Cor
yneb
acte
rium
),M
y(M
ycob
acte
rium
),N
o(N
orca
dia)
,R
h(R
hodo
cocc
us),
Sc
(Sac
char
opol
yspo
ra),
Sa
(Sal
inis
pora
),F
(Fra
nkia
),S
t(S
trep
tom
yces
),T
(The
rmob
ifida
),N
(Noc
ardi
oide
s),
Ki(
Kin
eoco
ccus
),J
(Jan
ibac
ter)
,Ar
(Art
hrob
acte
r),
Re
(Ren
ibac
-te
rium
),K
o(K
ocur
ia),
Mi(
Mic
roco
ccus
),L
(Lei
fson
ia),
Cl(
Cla
viba
cter
),A
g(A
grei
a),B
(Bre
viba
cter
ium
),A
c(A
ctin
omyc
es),
and
P(P
ropi
onib
acte
rium
).T
hefr
eque
ncy
inpa
rent
hesi
sre
pres
ents
the
num
ber
ofge
nera
that
poss
esth
eco
rres
pond
ing
Sig
Rta
rget
gene
amon
g22
gene
ra,
who
se42
geno
mes
wer
eex
amin
edin
this
stud
y.
12 M.-S. Kim et al. �
© 2012 Blackwell Publishing Ltd, Molecular Microbiology

SigE in Mycobacterium tuberculosis (Sachdeva et al.,2010). Therefore, in many actinomycetes, it is likely thatSigR orthologues control the expression of anothergeneral stress factor of non-ECF family, suggesting thatthe stress signal initiates a transcriptional cascade, whichincludes multiple DNA binding proteins. The function ofRbpA is not well understood except that it associates withRNA polymerase and confers resistance to rifampicin inactinomycetes (Newell et al., 2006; Dey et al., 2010). PrfAterminates translation as a peptide release factor andrecycles ribosome whereas RpmE1 is the ribosomalprotein L31. How the induction of genes for PrfA andRpmE1 is beneficial under thiol-oxidative conditions is notclear. The rpmE1 gene encoding an isoform of L31 withcysteine residues for zinc-binding is induced by SigR,whereas the cysteine-negative paralogue rpmE2 isinduced by Zur under zinc-depleted condition in S. coeli-color (Owen et al., 2007; Shin et al., 2007). Whether thioloxidations in ribosomal components or translationalcomplex causes increased demand for PrfA and RpmE1and whether zinc-containing RpmE1 is sensitive to oxida-tive stress deserve future studies.
When we extended the analysis to include less con-served genes up to 250 gene sets, we found that somegene sets are still conserved significantly across actino-mycetes, whereas others are more restricted to certaingenera. Figure 5 displays the clustered pattern of thesegene sets to exhibit a relatively common group (E) andgenus-specific groups (A, B, C, D, F, G), distributed to aspecific genus such as Streptomyces (group A&E), Salin-ispora (group B&E), Frankia (group C&E), Corynebacte-rium (group D), Mycobacterium (group E&F) andArthrobacter (group G). The so-called extended regulonof S. coelicolor (group A&E) contains 79 genes, andamong genes with predicted functions the main functionsrepresented are: electron transport (9 genes), biosynthe-sis of cofactors (9 genes), transcriptional regulators (6
genes), and protein homeostasis (5 genes). However,only 29 of these 79 putative targets are associated with aChIP-chip peak from our analysis, suggesting that thecomputational detection of SigR targets that are notbroadly conserved across the phylogeny may beimpaired by a high rate of false-positive in this analysis(Fig. 5, Table S2).
SigR and its paralogues with shared promoter selectivity– Overlap of SigR regulon with other regulons
Among the top 25 conserved SigR regulon members, dnaK(#61) and grpE(#445) are not directly dependent on SigR inS. coelicolor. The consensus SigR logo is not foundupstream of the dnaK-grpE-dnaJ-hspR operon in S. coeli-color as well as in all the other Streptomyces genomesexamined in this study. In M. tuberculosis, transcription ofthe dnaK operon is activated in response to heat shock andoxidative stress by SigE, a paralogue of SigH (SigR ortho-logue) (Fig. S2), with similar recognition sequence (Songet al., 2008). Mtb-sigB gene (#2534) is also recognized bySigE in M. tuberculosis. Since Mtb-SigH induces theexpression of the sigE gene (Raman et al., 2001; Manga-nelli et al., 2002), there exist two tiers of overlap betweenregulons of SigH and SigE in M. tuberculosis, arising fromsimilar recognition sequence and from hierarchicallysequential induction. Likewise, it is conceivable that thednaK operon in S. coelicolor is regulated by a SigR paral-ogue that resembles Mtb-SigE. If this were true, SigR1(SCO5147) is the most likely candidate for activating thednaK operon in S. coelicolor (Fig. S2). SCO5147 couldplay a similar role in other species since it is found amongthe top 50 conserved sets of SigR-regulon members inactinomycetes (Table S2, group E). In M. tuberculosis, theSigE-binding consensus (GGAACY-N17-18-GTT) over-laps significantly with SigH (SigR orthologue)-binding con-sensus (GGAAYA-N17-18-GTT) (Sachdeva et al., 2010). It
Fig. 5. The top 250 orthologous sets of genes predicted to be SigR targets across 42 selected Actinomycetes. The heatmap colour coding isidentical to Fig. 4. Species are organized in the same order as in Fig. 4. Red marks indicate genes that are associated with a SigR ChIP-chippeak and differentially expressed under diamide stress, while black marks indicate genes that are only associated with a SigR ChIP-chip peak.The broader characterization of the conservation of SigR targets predicts in addition to the core regulon the existence of extended SigRregulons that are specific to lineages of related species. Lineage specific SigR regulons are indicated by letters on top, and the genecomposition of each group is summarized in Table S2.
Conserved thiol-oxidative stress in actinomycetes 13
© 2012 Blackwell Publishing Ltd, Molecular Microbiology

has been reported that a single position in the -35 elementprimarily distinguish the recognition specificity of SigE fromthat of SigH (Song et al., 2008). Considering this, it is likelythat the list of bioinformatically predicted SigR regulongenes includes not only those directly recognized by SigRorthologues, but also some that are recognized by SigE-type paralogues. In addition, since sigE transcriptionrequires SigH in M. tuberculosis, the predicted SigR/SigHregulon members may include indirectly regulated onesdue to regulator cascade. The extent of overlap betweenSigR orthologues and their close paralogues in sequencespecificity as well as hierarchical regulation may differ indifferent organisms. Experimental verification may requirenot only the comparison of transcripts between the wild-type and regulator mutants, but also regulator bindingstudies in vivo and in vitro.
Involvement of ClgR in the core SigR regulon is intriguing(Table 2). The clgR gene (SCO5755) is co-transcribed withSCO5754, a SigR target gene which encodes a homo-logue of CinA known to be induced by DNA damage andcompetence in Streptococcus pneumoniae (Martin et al.,1995). In S. coelicolor ClgR positively regulates genes forproteases (clpP1/P2, lon, clpC) that are direct targets ofSigR (Bellier and Mazodier, 2004). In C. glutamicum, ClgRcontrols transcription of genes involved in proteolysis andDNA repair such as ptrB encoding endopeptidase, hflXencoding a GTP-binding protein, recR for recombinationalrepair of UV-induced DNA damage (Engels et al., 2004).Inclusion of known ClgR targets in the SigR regulon ofS. coelicolor demonstrates an interesting regulatory circuitthat involves double positive regulators. The contribution ofSigR appears critical to allow promoter recognition as wellas provision of ClgR in transcribing ClgR target genes.
Functional overlap between prokaryotic SigR regulonand eukaryotic Nrf2 regulon suggests a broadconservation of the thiol oxidative stress response
Examination of SigR regulon genes in S. coelicolor revealsa striking functional similarity with an analogous systemin mammals as summarized in Fig. 6. The Nrf2-Keap1system in higher eukaryotes constitutes a defence mecha-nism against stresses imposed by electrophiles and oxi-dants. Nrf2 is associated with its inhibitor Keap1, whichfacilitates ubiquitination and proteosomal degradation ofNrf2. Attacked by electrophiles or oxidants, Nrf2-Keap1interaction is disrupted, and the released Nrf2 induces itsregulon genes through binding to antioxidant responseelements (ARE) (Kensler et al., 2007). Nrf2 protein itselfdoes not have any sequence homology with SigR.However, the mechanism by which it is activated involvesthiol-disulphide isomerization of its inhibitor Keap1 asoccurs in the activity modulation of SigR by RsrA (Dinkova-Kostova et al., 2002). Similarly to RsrA, Keap1 has many
cysteine residues (27 cysteines) and responds to oxidativeand electrophilic stressors. Functions of the downstreamtarget genes also overlap significantly between the SigR/RsrA and Nrf2-Keap1 systems. When activated, Nrf2induces genes for various thiol homeostatic proteins, suchas glutathione peroxidase, g-glutamyl-cystein synthetase,thioredoxin reductase and peroxiredoxin 1, which are func-tionally related with thiol redox proteins, such as thioredox-ins and their reductase, peroxiredoxin, mycothiol syntheticenzymes, and mycoredoxins regulated by SigR (Thimmu-lappa et al., 2002; Rangasamy et al., 2004). Just as Nrf2increases synthesis of proteasome subunits andg-glutamyl peptidase, SigR activates expression of lon,pepN, clp proteases and mpa (Kwak et al., 2003). Theamino acid sequence of pepN is homologous to g-glutamylpeptidase. Both Nrf2 and SigR regulate pentose phos-phate pathway enzymes such as glucose 6-phosphatedehydrogenase and transaldolase for generating reducingpotential NADPH. The SCO2763 gene product, a putativeATP-binding cassette (ABC) transporter, is a closehomologue of multidrug resistance-associated protein 1(Mrp1/Abcc1) whose expression depends on Nrf2 andis responsible for cellular extrusion of GSH-conjugates(Hayashi et al., 2003). Whether SCO2763 has a function inexporting mycothiol (MSH)-S-conjugates of electrophilesdeserves experimental study.
Fig. 6. Induction and function of SigR regulon in comparison withmammalian anti-oxidative Nrf2-Keap1 system. SigR regulon isinduced in response to thiol-reactive oxidants and electrophiliccompounds, through inactivation of antisigma RsrA by formingintramolecular disulphide bond. Nrf2 regulon is induced by similarstressors, through inactivation of inhibitory factor Keap1 thatfacilitates degradation of Nrf2, by forming multiple intra- andinter-molecular disulphide bonds, and S-alkylation. Functions ofSigR target genes overlap with those of Nrf2 targets to a largeextent, which include thiol redox homeostasis, small thiol-mediateddetoxification, protein quality control, and pentose phosphatepathway for generating NADPH. The scheme for Nrf2 activationmechanism is simplified to highlight analogy with SigR-RsrA(Kensler et al., 2007; Antelmann and Helmann, 2011).
14 M.-S. Kim et al. �
© 2012 Blackwell Publishing Ltd, Molecular Microbiology

Conclusions
Genome-wide identification of direct SigR binding sitesunder thiol-oxidative stress conditions, combined with tran-script and sequence analyses, revealed many new genesof SigR regulon in S. coelicolor. The wide spectrum oftarget genes that include cascades of regulators indicatesthat SigR is a global regulator. As expected, SigR inducesmany genes related with homeostasis of thiols in proteinsand small molecules (mycothiols), and proper proteinquality control, which are most prominently affectedby thiol-reactive/oxidative stressors. Additionally, SigRinduces many genes for ribosome-associated compo-nents, transcriptional regulators including sigma factors,numerous oxidoreductases (many of which are predictedto contain flavin), DNAdamage repair, cofactor metabolismthat include several Fe-S containing enzymes/proteins,and lipid synthesis. The full list of SigR target genesenabled prediction and experimental demonstration of newphysiological functions of the SigR regulon under thiol-oxidative stress condition. We found that SigR directlycontributes to maintain the level and activity of the majorhousekeeping sigma factor HrdB through increasing itstranscription during oxidative stress. SigR also confersresistance to UV and thiol-reactive compounds, likely byinducing enzymes of DNA damage repair. Generation ofa refined SigR promoter model enabled comparativegenomics analysis of the SigR regulon, which revealed theexistence of a core SigR regulon conserved across 42selected Actinomycetes. The most prominent functions ofthe conserved SigR-orthologue regulons are related withthiol redox homeostasis, including the maintenance ofsulphur-containing amino acids, and protein quality controlinvolving chaperones and proteolytic systems. In addition,genes for translational modulation, transcriptional regula-tory cascades, oxidoreductases and Fe-S assembly func-tions are also included. The bioinformatic analysis alsopredicts the existence of variable extended regulons spe-cific for each genus. These results complement the experi-mental characterization of the SigR regulon in S. coelicolorto identify genes and functions that are most likely to becritical for the biological response to thiol oxidative stress.Comparison of SigR-regulated gene functions with thoseof thiol-oxidative stress response system (Nrf2-Keap1) inmammals reveals many shared gene functions, suggest-ing a robust physiological mechanism to deal with thiol-reactive stresses across distantly related life forms.
Experimental procedures
Strains and media
Spores of S. coelicolor A3(2) strain M145 were inoculated inYEME liquid medium containing 5 mM MgCl2•6H2O and 10%sucrose, and were grown at 30°C (Kieser et al., 2000). NA
plates (0.8% nutrient broth, 2% agar powder) were used forspotting test. E. coli was grown in LB broth (1% tryptone,0.5% yeast extract and 1% NaCl). E. coli ET12567, a non-methylating strain containing pUZ8002 for donor functionsand BW25113 harbouring pIJ790 for linear DNA recombina-tion were used for disruption as recommended (Gust et al.,2002).
Chromatin immunoprecipitation on a chip(ChIP-chip) analysis
The probes used to fabricate the custom-made 385 KNimbleGen microarray (Roche NimbleGen, Madison WI) forthe ChIP-chip experiment were designed using the ‘chipD’software (Dufour et al., 2010b). The parameters were set toobtain complete tilling of the S. coelicolor A3(2) genome withoverlapping isothermal probes that ranged from 35 to 75bases with an average spacing of 24 bases. In addition,probes alternate between the forward and reverse orienta-tion to capture both DNA strands. The probe sequencesand the ChIP-chip data are available at the NCBI’s GeneExpression Omnibus database with the accession numberGPL14825 (http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GPL14825).
Streptomyces coelicolor cells (wild-type M145 and Dsi-gRrsrA mutant strains) were grown to exponential phase (OD~ 0.3) and were treated with 0.5 mM diamide for 20 min toprepare samples for the ChIP-chip experiment. The sampleswere prepared as previously described (Dufour et al., 2008).Briefly, the cell cultures were fixed with 1% formaldehyde for4 min at 30°C followed by quenching with 0.125 M glycine for30 min in ice, washed with phosphate-buffered saline, andfrozen. Cells were suspended in 100 mMTris (pH 8.0),300 mM NaCl, 2% Triton X-100, and 1 mM phenylmethylsul-phonyl fluoride, and sonicated to lyse cells and fragment thegenomic DNA. Micrococcal nuclease (100 U ml-1) and RNaseA(1 mg ml-1) were added to the preparation to further fragmentthe genomic DNA and degrade RNA. Cell debris was removedby centrifugation and an aliquot was removed to analyse DNAfragmentation by agarose gel eletrophoresis (desired size of~200–1000 bp with enrichment for ~500 bp molecules). Anti-SigR rabbit polyclonal serum (7.5 ml) was used to immuno-precipitate cross-linked DNA fragments for each sampleprepared from about 50 ml culture. A mock immuno-precipitation was performed on the DsigRrsrA cells using thesame serum to detect non-specific interactions. The enrichedDNA was amplified via ligation-mediated PCR amplificationusing oligonucleotide linkers (oJW102: GCGGTGACCCGG-GAGATCTGAATTC; oJW103: GAATTCAGATC). The DNAenriched using anti-SigR serum was labelled with Cy5 dyewhile DNA from the mock precipitation was labelled with Cy3.Labelled DNA from three independent immuno-precipitationexperiments were pooled together in equal proportion toobtain the 4 mg of DNA that was hybridized on a microarray.Sample hybridization and microarray scanning were per-formed by Roche NimbleGen (Madison, WI) according to theirstandard protocol.
For each array, the Cy3 and Cy5 signals were normalizedusing the vsn method (Huber et al., 2002) in the R statisticalsoftware environment before calculating the log2 ratio of theexperimental signal (Cy5) over the control signal (Cy3). A plot
Conserved thiol-oxidative stress in actinomycetes 15
© 2012 Blackwell Publishing Ltd, Molecular Microbiology

of the intensity ratios versus intensity averages revealed nodye bias. Therefore, no further correction was applied to thedata. To remove the signal generated by non-specific DNAimmuno-precipitation, the signal from the DsigRrsrA cells wassubtracted from the signal obtained from the wild-type cells.Only one array was hybridized for each S. coelicolor strain.Regions of the genome enriched for occupancy by SigR weredetected using the TAMALPAIS method (Bieda et al., 2006)at P-value � 0.05 using a threshold set at the 99th percentileof the log2 ratio. The relative peak intensities were calculatedby averaging the 10 highest consecutive probe signals foreach selected peak.
Re-examination of expression profiling experimentsunder diamide stress
The microarray data of Kallifidas et al. (2010) werere-examined. To increase the detection sensitivity of SigR-dependent transcripts, the change in transcript abundancesin the wild-type strain (M600) was divided by the change intranscript abundances in the DsigR mutant strain (J2139)measured 20 min after the addition of diamide. Transcriptsthat were at least 1.5-fold more abundant in the wild-typestrain were selected to identify putative members of the SigRregulon. Taking the ratio of the changes in abundance mayhelp identify SigR-dependent transcripts that did not shownet increase in abundances in the wild-type strain becausehigher activity of the SigR-dependent promoter may beaccompanied by lower activity of an alternative promoter.
S1 nuclease mapping
Cells were treated with 0.1 ~ 0.5 mM diamide at the expo-nential growth phase (OD ~ 0.3) for 20–120 min. Afterharvest, they were disrupted by sonication in Kirby mix. RNAisolation and S1 nuclease mapping was performed accordingto a standard procedure (Kieser et al., 2000). Promoterregions (from -500 to +60 nt relative to each start codon ofSigR regulon gene) were amplified by PCR and used asprobes. Probes were labelled with [g-32P]-ATP and hybridizedwith 5–50 mg RNA at 50°C. After S1 nuclease treatment,protected DNAs were separated on 5% polyacrylamide gelcontaining 7 M urea. Signals were detected and quantified byBAS-2500 (Fuji film).
Western blot analysis
Cells were grown and treated with diamide as described forsample preparation for S1 mapping. Harvested cells werere-suspended in lysis buffer [20 mM Tris-HCl (pH 7.9), 10%(v/v) glycerol, 5 mM EDTA, 0.1 mM DTT, 10 mM MgCl2, 1 mMPMSF, 0.15 M NaCl]. The suspension was sonicated withultrasonicator (Sonics and Materials), and cleared bycentrifugation. Protein concentration in crude cell extract wasdetermined by Bradford reagent solution (Bio-Rad) usingBSA as a standard. For each sample, ~ 10 ml cell extractcontaining 50 mg protein was further diluted to the final con-centration of 0.125 mg ml-1 with lysis buffer (up to 400 ml) thatadditionally contain 50 mg BSA to serve as a protein buffer.
Aliquots of 8 ml containing 1 mg crude protein extract and thesame amount of BSA were resolved on 10% SDS-PAGE.Immuno-detection by polyclonal mouse antibody againstHrdB protein and the anti-mouse IgG secondary antibody at1:5000 dilution ratio, followed by ECL detection system(Amersham Life Science), was done as described previously(Kim et al., 2009).
Sensitivity test
The disruption mutant of SCO1958 (uvrA) was obtained byusing PCR-targeting mutagenesis protocol (Gust et al., 2002)as described previously (Kim et al., 2009). The wild-type andmutant spores (105, 104, 103) were spotted on NA media. ForUV sensitivity assay, the spotted spores were exposed to UVin the CL-1000 UV cross-linker at 50 or 100 J m-2 beforeincubating. For chemical sensitivity, spores were spotted onNA media containing 0.6 mM diamide, 1 mM methylhydro-quinone or 1 mM methylglyoxal, and incubated from 2 to 6days at 30°C.
Determination of orthologous genes and phylogenetictree reconstruction
All the genome sequences were obtained from the IntegratedMicrobial genome database on September 2008 (http://img.jgi.doe.gov) (Markowitz et al., 2006). Groups of orthologousgenes were determined using the OrthoMCL 1.4 software(http://www.orthomcl.org/) using the default parameters (Liet al., 2003a). To determine the species phylogenetic tree, wefirst selected about 20 proteins from groups of orthologuesthat have only one member in each genome and aligned theamino acid sequences with the MUSCLE 3.7 software (http://www.drive5.com/muscle/) (Edgar, 2004). The alignmentswere then concatenated and poorly aligned positions wereremoved using the GBlocks 0.91b software (http://molevol.cmima.csic.es/castresana/Gblocks.html) (Talavera and Cas-tresana, 2007). Finally, the phylogenetic tree was computedfrom the multiple alignment using the Fasttree 1.0.0 software(http://www.microbesonline.org/fasttree/) set to defaultparameters (Price et al., 2009).
Comparative genomics reconstruction of theSigR regulon
The phylogenetic analyses used to predict the conservationof SigR target genes across related species was conductedas previously described (Dufour et al., 2010a). The SigR pro-moter sequence model used to predict target genes acrossall the genomes sequences was constructed by aligning58 SigR promoter sequences from S. coelicolor using theHMMER 1.8.5 software (http://hmmer.janelia.org/). To detectputative SigR-dependent promoters we scored the 300 basepair sequence upstream of every annotated gene in eachgenome. The distribution of scores of each genome wasnormalized to a normal distribution to eliminate the influenceof varying base composition in the background sequencesamong species. In addition, because bacterial genomes areorganized in polycistronic operons, the promoter scores were
16 M.-S. Kim et al. �
© 2012 Blackwell Publishing Ltd, Molecular Microbiology

propagated to the remaining genes of the operon accordingto the probability that consecutive genes are co-transcribed.Operon predictions were obtained from the VIMSS database(http://www.microbesonline.org/operons/) (Price et al., 2005).Finally, genes with a z-score of � 2.0 were selected as puta-tive SigR targets.
Acknowledgements
This work was supported by grants for NRL of MolecularMicrobiology to J.H. Roe (NRF-2009–0079278) and for theIntelligent Synthetic Biology Center of Global Frontier Projectfrom MEST (2011–0031960). Ji-Sun Yoo received doctoralfellowship from the second-stage BK21 Program for LifeSciences at SNU.
References
Antelmann, H., and Helmann, J.D. (2011) Thiol-based redoxswitches and gene regulation. Antioxid Redox Signal 14:1049–1063.
Antelmann, H., Hecker, M., and Zuber, P. (2008) Proteomicsignatures uncover thiol-specific electrophile resistancemechanisms in Bacillus subtilis. Expert Rev Proteomics 5:77–90.
Bae, J.B., Park, J.H., Hahn, M.Y., Kim, M.S., and Roe, J.H.(2004) Redox-dependent changes in RsrA, an anti-sigmafactor in Streptomyces coelicolor: zinc release and disul-fide bond formation. J Mol Biol 335: 425–435.
Barends, S., Zehl, M., Bialek, S., de Waal, E., Traag, B.A.,Willemse, J., et al. (2010) Transfer-messenger RNA con-trols the translation of cell-cycle and stress proteins inStreptomyces. EMBO Rep 11: 119–125.
Baylis, H.A., and Bibb, M.J. (1988) Transcriptional analysis ofthe 16S rRNA gene of the rrnD gene set of Streptomycescoelicolor A3(2). Mol Microbiol 2: 569–579.
Bellier, A., and Mazodier, P. (2004) ClgR, a novel regulator ofclp and lon expression in Streptomyces. J Bacteriol 186:3238–3248.
Bieda, M., Xu, X., Singer, M.A., Green, R., and Farnham, P.J.(2006) Unbiased location analysis of E2F1-binding sitessuggests a widespread role for E2F1 in the humangenome. Genome Res 16: 595–605.
Brandes, N., Schmitt, S., and Jakob, U. (2009) Thiol-basedredox switches in eukaryotic proteins. Antioxid RedoxSignal 11: 997–1014.
Burns, K.E., and Darwin, K.H. (2010) Pupylation versus ubiq-uitylation: tagging for proteasome-dependent degradation.Cell Microbiol 12: 424–431.
Buttner, M.J., Chater, K.F., and Bibb, M.J. (1990) Cloning,disruption, and transcriptional analysis of three RNA poly-merase sigma factor genes of Streptomyces coelicolorA3(2). J Bacteriol 172: 3367–3378.
Christensen-Dalsgaard, M., Jorgensen, M.G., and Gerdes, K.(2010) Three new RelE-homologous mRNA interferases ofEscherichia coli differentially induced by environmentalstresses. Mol Microbiol 75: 333–348.
D’Autreaux, B., and Toledano, M.B. (2007) ROS as signallingmolecules: mechanisms that generate specificity in ROShomeostasis. Nat Rev Mol Cell Biol 8: 813–824.
Darwin, K.H. (2009) Prokaryotic ubiquitin-like protein (Pup),
proteasomes and pathogenesis. Nat Rev Microbiol 7: 485–491.
Dey, A., Verma, A.K., and Chatterji, D. (2010) Role of an RNApolymerase interacting protein, MsRbpA, from Mycobacte-rium smegmatis in phenotypic tolerance to rifampicin.Microbiology 156: 873–883.
Dinkova-Kostova, A.T., Holtzclaw, W.D., Cole, R.N., Itoh, K.,Wakabayashi, N., Katoh, Y., et al. (2002) Direct evidencethat sulfhydryl groups of Keap1 are the sensors regulatinginduction of phase 2 enzymes that protect against carcino-gens and oxidants. Proc Natl Acad Sci USA 99: 11908–11913.
Dufour, Y.S., Landick, R., and Donohue, T.J. (2008) Organi-zation and evolution of the biological response to singletoxygen stress. J Mol Biol 383: 713–730.
Dufour, Y.S., Kiley, P.J., and Donohue, T.J. (2010a) Recon-struction of the core and extended regulons of global tran-scription factors. PLoS Genet 6: e1001027.
Dufour, Y.S., Wesenberg, G.E., Tritt, A.J., Glasner, J.D.,Perna, N.T., Mitchell, J.C., and Donohue, T.J. (2010b)chipD: a web tool to design oligonucleotide probes forhigh-density tiling arrays. Nucleic Acids Res 38: W321–W325.
Edgar, R.C. (2004) MUSCLE: multiple sequence alignmentwith high accuracy and high throughput. Nucleic Acids Res32: 1792–1797.
Engels, S., Schweitzer, J.E., Ludwig, C., Bott, M., and Schaf-fer, S. (2004) clpC and clpP1P2 gene expression inCorynebacterium glutamicum is controlled by a regulatorynetwork involving the transcriptional regulators ClgR andHspR as well as the ECF sigma factor sigmaH. Mol Micro-biol 52: 285–302.
Gust, B., Kieser, T., and Chater, K. (2002) Redirect Technol-ogy: PCR-targeting System in Streptomyces coelicolor.Norwich: John Innes Centre.
Hahn, M.Y., and Roe, J.H. (2007) Partial purification of factorsfor differential transcription of the rrnD promoters for riboso-mal RNA synthesis in Streptomyces coelicolor. J Microbiol45: 534–540.
Hayashi, A., Suzuki, H., Itoh, K., Yamamoto, M., and Sug-iyama, Y. (2003) Transcription factor Nrf2 is required forthe constitutive and inducible expression of multidrugresistance-associated protein 1 in mouse embryo fibro-blasts. Biochem Biophys Res Commun 310: 824–829.
den Hengst, C.D., and Buttner, M.J. (2008) Redox control inactinobacteria. Biochim Biophys Acta 1780: 1201–1216.
Hess, D.T., Matsumoto, A., Kim, S.O., Marshall, H.E., andStamler, J.S. (2005) Protein S-nitrosylation: purview andparameters. Nat Rev Mol Cell Biol 6: 150–166.
Huber, W., von Heydebreck, A., Sultmann, H., Poustka, A.,and Vingron, M. (2002) Variance stabilization applied tomicroarray data calibration and to the quantification of dif-ferential expression. Bioinformatics 18 (Suppl. 1): S96–104.
Imlay, J.A. (2008) Cellular defenses against superoxide andhydrogen peroxide. Annu Rev Biochem 77: 755–776.
Jacob, C., Knight, I., and Winyard, P.G. (2006) Aspects of thebiological redox chemistry of cysteine: from simple redoxresponses to sophisticated signalling pathways. Biol Chem387: 1385–1397.
Kallifidas, D., Thomas, D., Doughty, P., and Paget, M.S.
Conserved thiol-oxidative stress in actinomycetes 17
© 2012 Blackwell Publishing Ltd, Molecular Microbiology

(2010) The sigmaR regulon of Streptomyces coelicolor A32reveals a key role in protein quality control during disul-phide stress. Microbiology 156: 1661–1672.
Kang, J.G., Paget, M.S., Seok, Y.J., Hahn, M.Y., Bae, J.B.,Hahn, J.S., et al. (1999) RsrA, an anti-sigma factor regu-lated by redox change. EMBO J 18: 4292–4298.
Kawanishi, S., Hiraku, Y., and Oikawa, S. (2001) Mechanismof guanine-specific DNA damage by oxidative stress andits role in carcinogenesis and aging. Mutat Res 488:65–76.
Kensler, T.W., Wakabayashi, N., and Biswal, S. (2007) Cellsurvival responses to environmental stresses via theKeap1-Nrf2-ARE pathway. Annu Rev Pharmacol Toxicol47: 89–116.
Kieser, T., Bibb, M., Buttner, M., Chater, K., and Hopwood, D.(2000) Practical Streptomyces Genetics. Norwich: TheJohn Innes Foundation.
Kiley, P.J., and Storz, G. (2004) Exploiting thiol modifications.PLoS Biol 2: e400.
Kim, M.S., Hahn, M.Y., Cho, Y., Cho, S.N., and Roe, J.H.(2009) Positive and negative feedback regulatory loops ofthiol-oxidative stress response mediated by an unstableisoform of sigmaR in actinomycetes. Mol Microbiol 73:815–825.
Kwak, M.K., Wakabayashi, N., Greenlaw, J.L., Yamamoto,M., and Kensler, T.W. (2003) Antioxidants enhance mam-malian proteasome expression through the Keap1-Nrf2signaling pathway. Mol Cell Biol 23: 8786–8794.
Li, L., Stoeckert, C.J., and Roos, D.S. (2003a) OrthoMCL:identification of ortholog groups for eukaryotic genomes.Genome Res 13: 2178–2189.
Li, W., Bottrill, A.R., Bibb, M.J., Buttner, M.J., Paget, M.S.,and Kleanthous, C. (2003b) The role of zinc in the disul-phide stress-regulated anti-sigma factor RsrA from Strep-tomyces coelicolor. J Mol Biol 333: 461–472.
Manganelli, R., Voskuil, M.I., Schoolnik, G.K., Dubnau, E.,Gomez, M., and Smith, I. (2002) Role of theextracytoplasmic-function sigma factor sigma(H) in Myco-bacterium tuberculosis global gene expression. Mol Micro-biol 45: 365–374.
Markowitz, V.M., Korzeniewski, F., Palaniappan, K., Szeto,E., Werner, G., Padki, A., et al. (2006) The integratedmicrobial genomes (IMG) system. Nucleic Acids Res 34:D344–D348.
Martin, B., Garcia, P., Castanie, M.P., and Claverys, J.P.(1995) The recA gene of Streptococcus pneumoniae is partof a competence-induced operon and controls lysogenicinduction. Mol Microbiol 15: 367–379.
Neubauer, C., Gao, Y.G., Andersen, K.R., Dunham, C.M.,Kelley, A.C., Hentschel, J., et al. (2009) The structuralbasis for mRNA recognition and cleavage by the ribosome-dependent endonuclease RelE. Cell 139: 1084–1095.
Newell, K.V., Thomas, D.P., Brekasis, D., and Paget, M.S.(2006) The RNA polymerase-binding protein RbpA confersbasal levels of rifampicin resistance on Streptomycescoelicolor. Mol Microbiol 60: 687–696.
Newton, G.L., Buchmeier, N., and Fahey, R.C. (2008) Bio-synthesis and functions of mycothiol, the unique protectivethiol of Actinobacteria. Microbiol Mol Biol Rev 72: 471–494.
Nguyen, T.T., Eiamphungporn, W., Mader, U., Liebeke, M.,Lalk, M., Hecker, M., et al. (2009) Genome-wide responses
to carbonyl electrophiles in Bacillus subtilis: control of thethiol-dependent formaldehyde dehydrogenase AdhA andcysteine proteinase YraA by the MerR-family regulatorYraB (AdhR). Mol Microbiol 71: 876–894.
Owen, G.A., Pascoe, B., Kallifidas, D., and Paget, M.S.(2007) Zinc-responsive regulation of alternative ribosomalprotein genes in Streptomyces coelicolor involves zur andsigmaR. J Bacteriol 189: 4078–4086.
Paget, M.S., and Buttner, M.J. (2003) Thiol-based regulatoryswitches. Annu Rev Genet 37: 91–121.
Paget, M.S., Kang, J.G., Roe, J.H., and Buttner, M.J. (1998)sigmaR, an RNA polymerase sigma factor that modulatesexpression of the thioredoxin system in response to oxida-tive stress in Streptomyces coelicolor A3(2). EMBO J 17:5776–5782.
Paget, M.S., Molle, V., Cohen, G., Aharonowitz, Y., andButtner, M.J. (2001) Defining the disulphide stressresponse in Streptomyces coelicolor A3(2): identification ofthe sigmaR regulon. Mol Microbiol 42: 1007–1020.
Park, J.H., and Roe, J.H. (2008) Mycothiol regulates and isregulated by a thiol-specific antisigma factor RsrA andsigma(R) in Streptomyces coelicolor. Mol Microbiol 68:861–870.
Pearce, M.J., Mintseris, J., Ferreyra, J., Gygi, S., and Darwin,K.H. (2008) Ubiquitin-like protein involved in the protea-some pathway of Mycobacterium tuberculosis. Science322: 1104–1107.
Perez, J.C., and Groisman, E.A. (2009) Evolution of transcrip-tional regulatory circuits in bacteria. Cell 138: 233–244.
Price, M.N., Huang, K.H., Alm, E.J., and Arkin, A. (2005) Anovel method for accurate operon predictions in allsequenced prokaryotes. Nucleic Acids Res 33: 880–892.
Price, M.N., Dehal, P.S., and Arkin, A. (2009) FastTree: com-puting large minimum evolution trees with profiles insteadof a distance matrix. Mol Biol Evol 26: 1641–1650.
Raman, S., Song, T., Puyang, X., Bardarov, S., Jacobs, W.R.,and Husson, R.N. (2001) The alternative sigma factor SigHregulates major components of oxidative and heat stressresponses in Mycobacterium tuberculosis. J Bacteriol 183:6119–6125.
Rangasamy, T., Cho, C.Y., Thimmulappa, R.K., Zhen, L.,Srisuma, S.S., Kensler, T.W., et al. (2004) Genetic ablationof Nrf2 enhances susceptibility to cigarette smoke-inducedemphysema in mice. J Clin Invest 114: 1248–1259.
Rudolph, T.K., and Freeman, B.A. (2009) Transduction ofredox signaling by electrophile-protein reactions. SciSignal 2: re7.
Sachdeva, P., Misra, R., Tyagi, A.K., and Singh, Y. (2010) Thesigma factors of Mycobacterium tuberculosis: regulation ofthe regulators. FEBS J 277: 605–626.
Shin, J.H., Oh, S.Y., Kim, S.J., and Roe, J.H. (2007) Thezinc-responsive regulator Zur controls a zinc uptakesystem and some ribosomal proteins in Streptomycescoelicolor A3(2). J Bacteriol 189: 4070–4077.
Sinha, R.P., and Hader, D. (2002) UV-induced DNA damageand repair: a review. Photochem Photobiol Sci 1: 225–236.
Song, T., Song, S.E., Raman, S., Anaya, M., and Husson,R.N. (2008) Critical role of a single position in the -35element for promoter recognition by Mycobacterium tuber-culosis SigE and SigH. J Bacteriol 190: 2227–2230.
Synold, T., Xi, B., Wuenschell, G.E., Tamae, D., Figarola, J.L.,
18 M.-S. Kim et al. �
© 2012 Blackwell Publishing Ltd, Molecular Microbiology

Rahbar, S., and Termini, J. (2008) Advanced glycation endproducts of DNA: quantification of N2-(1-Carboxyethyl)-2′-deoxyguanosine in biological samples by liquid chromatog-raphy electrospray ionization tandem mass spectrometry.Chem Res Toxicol 21: 2148–2155.
Talavera, G., and Castresana, J. (2007) Improvement of phy-logenies after removing divergent and ambiguously alignedblocks from protein sequence alignments. Syst Biol 56:564–577.
Thimmulappa, R.K., Mai, K.H., Srisuma, S., Kensler, T.W.,Yamamoto, M., and Biswal, S. (2002) Identification of Nrf2-regulated genes induced by the chemopreventive agentsulforaphane by oligonucleotide microarray. Cancer Res62: 5196–5203.
Yang, Y.H., Song, E., Kim, E.J., Lee, K., Kim, W.S., Park,S.S., et al. (2009) NdgR, an IclR-like regulator involved in
amino-acid-dependent growth, quorum sensing, and anti-biotic production in Streptomyces coelicolor. Appl MicrobiolBiotechnol 82: 501–511.
Zuber, P. (2009) Management of oxidative stress in Bacillus.Annu Rev Microbiol 63: 575–597.
Supporting information
Additional supporting information may be found in the onlineversion of this article.
Please note: Wiley-Blackwell are not responsible for thecontent or functionality of any supporting materials suppliedby the authors. Any queries (other than missing material)should be directed to the corresponding author for thearticle.
Conserved thiol-oxidative stress in actinomycetes 19
© 2012 Blackwell Publishing Ltd, Molecular Microbiology
Copyright © 2022 FDOKUMEN




















