
Multiparametric protocol for the determination of thiol redox state in the living matter
14
Methods in Free Radical Biology and Medicine Multiparametric protocol for the determination of thiol redox state in living matter Konstantinos Grintzalis, Ioannis Papapostolou, Dimitris Zisimopoulos, Irene Stamatiou, Christos D. Georgiou n Genetics, Cell and Developmental Biology Section, Department of Biology, University of Patras, Patras, Greece article info Article history: Received 15 April 2014 Received in revised form 13 June 2014 Accepted 20 June 2014 Available online 1 July 2014 Keywords: Thiol redox state Glutathione Cysteine Nonprotein thiols Protein thiols abstract Thiol redox state (TRS) evaluation is mostly restricted to the estimation of GSH and GSSG. However, these TRS parameters can estimate the GSSG/GSH potential, which might be useful for indicating abnormalities in redox metabolism. Nonetheless, evaluation of the multiparameric nature of TRS is required for a more accurate assessment of its physiological role. The present protocol extends the partial assessment of TRS by current methodologies. It measures 15 key parameters of TRS by two modular subprotocols: one for the glutathione (GSH)- and cysteine (CSH)-based nonprotein (NP) thiols/ mixed disulfides (i.e., GSH, GSSG, GSSNP, CSH, CSSNP, NPSH, NPSSNP, NP x SH NPSSNP , NP x SH NPSH ), and the other for their protein (P) thiols/mixed disulfides (i.e., PSH, PSSG, PSSC, PSSNP, PSSP, NP x SH PSSNP ). The protocol eliminates autoxidation of GSH and CSH (and thus overestimation of GSSG and CSSNP). Its modularity allows the determination GSH and GSSG also by other published specific assays. The protocol uses three assays; two are based on the photometric reagents 4,4 0 -dithiopyridine (DTP) and ninhydrin (NHD), and the third on the fluorometric reagent o-phthaldialdehyde (OPT). The initial assays employing these reagents have been extensively modified and redesigned for increased specificity, sensitivity, and simplicity. TRS parameter values and their standard errors are estimated automatically by sets of Excel- adapted algebraic equations. Protocolsensitivity for NPSH, PSH, NPSSNP, PSSP, PSSNP, CSH, CSSNP, PSSC, NP x SH NPSSNP , and NP x SH NPSH is 1 nmol –SH/CSH, for GSSNP 0.2 nmol, for GSH and GSSG 0.4 nmol, and for PSSG 0.6 nmol. The protocol was applied on human plasma, a sample of high clinical value, and can be also applied in any organism. & 2014 Elsevier Inc. All rights reserved. Introduction Evaluating cellular thiol redox state (TRS) is very important in a wide variety of biological processes [1,2]. TRS comprises many protein (P) and nonprotein (NP) thiol parameters such as glu- tathione (GSH), cysteine (CSH), and reduced NP and P thiols (NPSH and PSH, respectively), as well as their symmetric and mixed disulfides. The commonly used indicators for the assessment of TRS are restricted to GSH and its symmetric oxidized disulfide (GSSG). The role of GSH in intracellular redox state signaling, enzymic antioxidant defense, and in response to environmental factors is well established [3,4]. Additionally, GSH regulation is related to many physiological processes, in which CSH and PSH and their mixed and symmetric protein disulfides (PSSNP and PSSP, respectively) are also involved [5]. Glutathione disulfide (GSSG) can activate many enzymes (e.g., glucose-6-phosphatase and acid phosphatase) and inhibit others (e.g., glycogen synthe- tase, pyruvate kinase, adenylate cyclase, phosphorylase/phospha- tase, and ribonucleotide reductase) [4]. Aerobic organisms need a system to restore key sulfhydryl groups to their reduced state after exposure to oxidant stress. Without a process to reduce protein disulfides, vulnerable cystei- nyl residues of essential enzymes might remain oxidized, leading to changes in catalytic activity. This function is fulfilled by the thiol-disulfide exchange catalyzed by thiol transferases in the Contents lists available at ScienceDirect journal homepage: www.elsevier.com/locate/freeradbiomed Free Radical Biology and Medicine http://dx.doi.org/10.1016/j.freeradbiomed.2014.06.024 0891-5849/& 2014 Elsevier Inc. All rights reserved. Abbreviations: TRS, thiol redox state; GSH, glutathione; GSSG, oxidized glu- tathione; GSSNP, mixed disulfides of glutathione with nonprotein (NP) thiols; CSH, cysteine; CSSNP, mixed disulfides of cysteine with nonprotein (NP) thiols; NPSH, reduced nonprotein (NP) thiols; NPSSNP, mixed disulfides of nonprotein thiols (NPS); NP x SH NPSSNP , the NP thiol component of NPSSNP, excluding the NPSH components GSH and CSH; NP x SH NPSH , the NP thiol component of NPSH, excluding the NPSH components GSH and CSH; PSH, reduced protein (P) thiols; PSSG, mixed disulfides of glutathione (GS) with protein thiols (PS); PSSC, mixed disulfides of cysteine (CS) with protein (PS) thiols; PSSP, mixed disulfides of protein thiols (PS); PSSNP, mixed disulfides of nonprotein (NP) with protein (PS) thiols; NP x SH PSSNP , the NP thiol component of PSSNP, excluding the NPSH components GSH and CSH; Note: abbreviations of the reagents used in the present study are listed in the section of Materials under “Reagents.” n Corresponding author. Fax: þ30 2610 997840. E-mail addresses: [email protected] (K. Grintzalis), [email protected] (I. Papapostolou), [email protected] (D. Zisimopoulos), [email protected] (I. Stamatiou), [email protected] (C.D. Georgiou). Free Radical Biology and Medicine 74 (2014) 85–98
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Multiparametric protocol for the determination of thiol redox state in the living matter

Methods in Free Radical Biology and Medicine
Multiparametric protocol for the determination of thiol redox statein living matter
Konstantinos Grintzalis, Ioannis Papapostolou, Dimitris Zisimopoulos,Irene Stamatiou, Christos D. Georgiou n
Genetics, Cell and Developmental Biology Section, Department of Biology, University of Patras, Patras, Greece
a r t i c l e i n f o
Article history:Received 15 April 2014Received in revised form13 June 2014Accepted 20 June 2014Available online 1 July 2014
Keywords:Thiol redox stateGlutathioneCysteineNonprotein thiolsProtein thiols
a b s t r a c t
Thiol redox state (TRS) evaluation is mostly restricted to the estimation of GSH and GSSG. However,these TRS parameters can estimate the GSSG/GSH potential, which might be useful for indicatingabnormalities in redox metabolism. Nonetheless, evaluation of the multiparameric nature of TRS isrequired for a more accurate assessment of its physiological role. The present protocol extends thepartial assessment of TRS by current methodologies. It measures 15 key parameters of TRS by twomodular subprotocols: one for the glutathione (GSH)- and cysteine (CSH)-based nonprotein (NP) thiols/mixed disulfides (i.e., GSH, GSSG, GSSNP, CSH, CSSNP, NPSH, NPSSNP, NPxSHNPSSNP, NPxSHNPSH), and theother for their protein (P) thiols/mixed disulfides (i.e., PSH, PSSG, PSSC, PSSNP, PSSP, NPxSHPSSNP).The protocol eliminates autoxidation of GSH and CSH (and thus overestimation of GSSG and CSSNP). Itsmodularity allows the determination GSH and GSSG also by other published specific assays. The protocoluses three assays; two are based on the photometric reagents 4,40-dithiopyridine (DTP) and ninhydrin(NHD), and the third on the fluorometric reagent o-phthaldialdehyde (OPT). The initial assays employingthese reagents have been extensively modified and redesigned for increased specificity, sensitivity, andsimplicity. TRS parameter values and their standard errors are estimated automatically by sets of Excel-adapted algebraic equations. Protocol sensitivity for NPSH, PSH, NPSSNP, PSSP, PSSNP, CSH, CSSNP, PSSC,NPxSHNPSSNP, and NPxSHNPSH is 1 nmol –SH/CSH, for GSSNP 0.2 nmol, for GSH and GSSG 0.4 nmol, and forPSSG 0.6 nmol. The protocol was applied on human plasma, a sample of high clinical value, and can bealso applied in any organism.
& 2014 Elsevier Inc. All rights reserved.
Introduction
Evaluating cellular thiol redox state (TRS) is very important in awide variety of biological processes [1,2]. TRS comprises many
protein (P) and nonprotein (NP) thiol parameters such as glu-tathione (GSH), cysteine (CSH), and reduced NP and P thiols (NPSHand PSH, respectively), as well as their symmetric and mixeddisulfides. The commonly used indicators for the assessment ofTRS are restricted to GSH and its symmetric oxidized disulfide(GSSG). The role of GSH in intracellular redox state signaling,enzymic antioxidant defense, and in response to environmentalfactors is well established [3,4]. Additionally, GSH regulation isrelated to many physiological processes, in which CSH and PSHand their mixed and symmetric protein disulfides (PSSNP andPSSP, respectively) are also involved [5]. Glutathione disulfide(GSSG) can activate many enzymes (e.g., glucose-6-phosphataseand acid phosphatase) and inhibit others (e.g., glycogen synthe-tase, pyruvate kinase, adenylate cyclase, phosphorylase/phospha-tase, and ribonucleotide reductase) [4].
Aerobic organisms need a system to restore key sulfhydrylgroups to their reduced state after exposure to oxidant stress.Without a process to reduce protein disulfides, vulnerable cystei-nyl residues of essential enzymes might remain oxidized, leadingto changes in catalytic activity. This function is fulfilled by thethiol-disulfide exchange catalyzed by thiol transferases in the
Contents lists available at ScienceDirect
journal homepage: www.elsevier.com/locate/freeradbiomed
Free Radical Biology and Medicine
http://dx.doi.org/10.1016/j.freeradbiomed.2014.06.0240891-5849/& 2014 Elsevier Inc. All rights reserved.
Abbreviations: TRS, thiol redox state; GSH, glutathione; GSSG, oxidized glu-tathione; GSSNP, mixed disulfides of glutathione with nonprotein (NP) thiols;CSH, cysteine; CSSNP, mixed disulfides of cysteine with nonprotein (NP) thiols;NPSH, reduced nonprotein (NP) thiols; NPSSNP, mixed disulfides of nonproteinthiols (NPS); NPxSHNPSSNP, the NP thiol component of NPSSNP, excluding the NPSHcomponents GSH and CSH; NPxSHNPSH, the NP thiol component of NPSH, excludingthe NPSH components GSH and CSH; PSH, reduced protein (P) thiols; PSSG, mixeddisulfides of glutathione (GS) with protein thiols (PS); PSSC, mixed disulfides ofcysteine (CS) with protein (PS) thiols; PSSP, mixed disulfides of protein thiols (PS);PSSNP, mixed disulfides of nonprotein (NP) with protein (PS) thiols; NPxSHPSSNP,the NP thiol component of PSSNP, excluding the NPSH components GSH and CSH;Note: abbreviations of the reagents used in the present study are listed in thesection of Materials under “Reagents.”
n Corresponding author. Fax: þ30 2610 997840.E-mail addresses: [email protected] (K. Grintzalis),
[email protected] (I. Papapostolou),[email protected] (D. Zisimopoulos), [email protected] (I. Stamatiou),[email protected] (C.D. Georgiou).
Free Radical Biology and Medicine 74 (2014) 85–98

presence of GSH. Many proteins are activated or inhibited in vitroby the disulfide exchange between the protein and the GSH orother small-molecule disulfides (R0SHþRSSR2R0SSRþRSH).Whereas many proteins are active when the key sulfhydryls arein the thiol form, others require them to be in the oxidized,disulfide form. Because the thiol-transferase reaction is bidirec-tional, the equilibrium is determined by the redox state of the cell.Moreover, cells avoid CSH autooxidation to CSSC, producingpotentially toxic oxygen radicals by storing most of the nonproteinCSH as GSH [4].
The role of NP thiols other than GSH and GSSG in mixeddisulfides (NPSSNP), such as coenzyme A (CoASH), in the TRS ofcells is usually underestimated, although they have significantcontribution to the physiological consequences of oxidative stress[5,6]. Such examples are the increase of CoASSG mixed disulfidelevels during hydroperoxide metabolism [7], its inhibitory effecton enzymes such as GSSG reductase (GR), phosphofructokinaseand fatty acid synthase, and its activating action on fructose16-bisphosphatase [4].
PSH, PSSP, and PSSNP are also important indicators of oxidativestress. Although PSSNP are often designated in the literature asPSSG [8] due to GSH contribution (glutathionylation), this designa-tion is limiting since they may also contain CSH (designated asPSSC; cysteinylation)—a particular form of protein S-thiolation iscysteyl-glycylation [9]—and to a lesser degree other minor NP thiols(NPxSHPSSNP, e.g., CoASH). Interdisulfide bridges between differentprotein thiols as well as intradisulfide bridges within proteinsundergoing oxidative tertiary structure modification can producesymmetric disulfides (PSSP) [10]. These can be significant thiolredox markers since they are involved in inactivation of enzymes,transporters, and transcriptional factors during oxidative stressconditions [10], and also in pathological processes associated withoxidative stress in animal and plant cells under stress [10–13].
Protein thiols/disulfides can be extracellular and part of mem-brane and subcellular structures, contributing to protein stability[14] and the regulation of redox homeostasis [1]. Of specialmetabolic interest is the involvement of the ROS-generatedprotein cysteine sulfenic acid in the formation of mixed GSHdisulfides via thiol-disulfide exchange reactions [15–18]. Redox-dependent signaling events involving the posttranslational oxida-tive modification of proteins have now been accepted as animportant regulatory process, controlled by a number of redox-dependent modifications of some protein cysteinyl thiols, includ-ing an interchange between the reduced thiol and several differentoxidized disulfide states [19]. It has been proposed that catalyti-cally important sulfhydryl groups in PSH are protected fromoxidative stress by reacting reversibly with GSH to form PSSG[20]. PSSG can inactivate enzymes (e.g., fructose-1,6-biphosphatealdolase) and make proteins more or less susceptible to proteolysis[21]. Also, specific oxidation/reduction of particular protein thiolsmay represent an important event in cellular and oxidativesignaling cascades [22,23]. In addition, protein thiols could reactwith nitric oxide radical to form S-nitrosothiols, which areinvolved in signal transduction and posttranslational proteinmodification [24]. All these render the simultaneous quantitativeassessment of thiols in organisms very important, since cellularTRS is associated with normal as well as abnormal metabolicprocesses.
TRS evaluation: Methodologies and limitations
TRS is normally evaluated by the total cellular thiol content [25]or by GSH and GSSG [8,26,27]. GSH is estimated by photometricassays based on Ellman’s reagent (5,50-dithiobis-2-nitrobenzoicacid, or DTNB) [28], which are not GSH specific since they do not
discriminate it from CSH [29]. GSSG is usually determined byenzymatic assays [8,30,31], while GSH has been also evaluatedmore specifically by HPLC after derivatization with N-ethylmalei-mide (NEM) [30]. GSH and GSSG are also evaluated by theo-phthaldialdehyde (OPT)-based fluorometric assay [32]. However,if proteins in the sample are not effectively removed (e.g., byprecipitation) they could cause interference because OPT, as withGSH, can bind covalently with closely spaced (�3 Å) sulfhydryland ε-amino groups of cysteine and lysine residues [33,34].Adopting this assay in the present protocol (see next section),we modified it to eliminate such protein interference and increaseits specificity and sensitivity. Similarly, PSSG is underestimated byphotometric assays because of oxidation of the measured GSHduring the procedure [35,36]. There is also lack of assays for thesimultaneous determination of CSH, nonprotein oxidized disul-fides of CSH (cystine, CSSC, CSH-GSH disulfide, CSSG), and PSSC.The available methodologies, mainly HPLC based, measure the CSHcomponent of the P/NP-CSH disulfides (via derivatized thioldetection by LC-MS after tri-n-butylphosphine, TBP, reduction[37], or by LC fluorescence following TBP [38] and tris(2-carbox-yethyl)phosphine, TCEP [39], disulfide reduction).
Concerning protein thiols, there are many methods availablefor measuring changes in the reduced protein thiol status of cellsand tissues [19]. In general, these methods make use of chemicalsthat react with thiol groups with a variety of tags or labels, whichallow the extent of incorporation to be measured. There arechemical methods involving thiol derivatization by thiol-labelingagents based on maleimide, iodoacetamide, iodoacetate, andthiosulfates. These agents may take the form of an affinity labelsuch as biotin, a fluorophore, a radionucleotide, or a label thatchanges the molecular weight of the protein. Other protein thiol-labeling agents include alkyl halides, arylating agents, thiosulfates,and disulfide compounds (such as DTNB). Antibodies to GSH havebeen used to investigate the thiol oxidation state of samples inenzyme-linked immunosorbent assays (ELISA). Moreover, cells canbe metabolically labeled by incubation with [35S]cysteine, and canthen undergo a variety of treatments, and S-thiolation (normallyinterpreted as S-glutathiolation) can be measured [19].
Normally, the PSH component of PSSNP, PSSG, and PSSC hasbeen determined by Ellman’s reagent-based assays [40,41], whichdo not discriminate the NP thiol component of PSSG, PSSC, andPSSNP [41]. Other methodologies quantify protein thiols withoutcorrelating the PS thiol component of PSSG, PSSC, PSSNP, and PSSPto its corresponding NP thiol component, which they do notmeasure [42]. However, simultaneous determination of incompletesets of P- and NP-TRS parameters based on GSH and CSH has beenperformed by GC-MS [37], laser-induced fluorescence capillaryelectrophoresis [43], and HPLC methodologies [38,39,42,44–46], aswell as by fluorometric and photometric assays [47–52]. However,in a case where plasma total thiols were determined after reductionwith TCEP, the involved HPLC methodology [39] does not discrimi-nate P from NP thiols (e.g., CSH and GSH resulting from thereduction of PSSC and PSSG, respectively).
Advantages of a multiparametric TRS protocol
Restricting TRS evaluation to the estimation of GSH and GSSGcan offer a calculation of the GSSG/GSH potential, which might beuseful at best as an analytical tool to disclose disturbances in redoxmetabolism [53]. Even so, a new technique of noninvasivelymeasuring thiol redox potentials has shown that the GSH/GSSGratio is 50,000 rather than about 100 in the cytosol [54], and thatthe cytosolic GSSG concentration is much more tightly regulatedthan expected (GSSG is rapidly transported into the vacuole by theABC-C transporter Ycf1 in yeast), providing also a mechanistic
K. Grintzalis et al. / Free Radical Biology and Medicine 74 (2014) 85–9886

explanation for the discrepancy with conventional measurements[54]. Given the partial assessment of TRS by current methodolo-gies, the evaluation of its multiparameric nature is required for theaccurate assessment of its physiological role.
This study presents a multiparametric protocol that estimates15 of the most important P and NP thiol/disulfide TRS parameters.The protocol is modular to adapt to any experimental needs asthey relate to TRS. The protocol represents a major extension andredesigning of two TRS protocols previously developed by ourlab [50,51]. Although limited in TRS parameters these protocolsmay have been, the scientific attention they have received is areflection of the usefulness in biological research of TRS evalua-tion. In general, the protocol differs in offering a more extensiveset of TRS parameters that are identified with increased specificity,sensitivity, and simplicity, while their estimation (and theirstatistical analysis) is automated by Excel-adapted algebraic setsof equations (see Supplementary Material). More specifically, themajor differences and innovations of the present protocol are thefollowing:
� It eliminates the artificial oxidation of GSH during sampleprocessing (and subsequent overestimation of GSSG), by usinga modification of the NEM-derivatization procedure for GSHreported in the (GSH-GSSG-restricted) Giustarini et al. 2013protocol [30]. However, the present protocol does not adopt theprocedures for GSH and GSSG (via HPLC, and the GR recyclingassay, respectively) of the aforementioned protocol, becausethey are somewhat complex and time consuming for an alsotime consuming multiparametric TRS protocol. Nonetheless,the modularity of the present protocol allows their selection (aswell as that of the GR recycling assay in Rahman et al. 2006protocol [31]) as optional. Given the applicability of the GSH-NEM derivatization step on other experimental models besidesblood [30,55], its adoption by the present protocol extents itsuse to any organism as well.
� It eliminates autoxidation of CSH (and thus overestimationof CSSNP) by inclusion of catalase (CAT) in the sample
homogenization buffers. CSH autoxidation to cystine can becaused by H2O2 in the presence of transition metals [56,57].
� It measures higher numbers of TRS parameters and of quitedifferent thiol types, and with much higher specificity andsimplicity than the Patsoukis and Georgiou 2004 and 2005protocols [50,51]. For example, the protocol also measures CSHand its P/NP disulfides, excluding the less TRS importantN-acetylcysteine that is measured by the Patsoukis and Geor-giou 2004 protocol [50].
� It measures GSH and GSSG with high specificity (equaling thatof the Giustarini et al. 2013 protocol [30]; data not shown) byusing a simple chemical assay that is based on the fluorescentprobe OPT. The Patsoukis and Georgiou 2005 and 2005 proto-cols, instead, measure GSH and GSSG nonspecifically by thephotometric DTNB and the fluorescent monobromobimaneprobes, respectively.
� It measures general TRS thiols with the photometric probe 4,40-dithiopyridine (DTP), a highly specific reagent for -SH groups,and not with the prone to interference DTNB probe (which hasbeen used in other assays and also by the Patsoukis andGeorgiou 2004 protocol).
� It uses TBP or TCEP to reduce thiol disulfides by a nearly 100%efficiency instead of the less effective disulfide reductantssodium borohydride (NaBH4) and NaBH4/dithithiothreitol(DTT) used in the Patsoukis and Georgiou 2004 and 2005protocols, respectively.
� It replaces the ineffective trichloroacetic acid (TCA) proteinprecipitation step with a certain combination of deoxycholate(DOC) and TCA, which precipitates proteins as low as 2–5 mg byZ90% effectiveness [58,59]. Protein precipitation with TCA aloneis used in the Patsoukis and Georgiou 2004 and 2005 protocols,and is normally used in most thiol protocols including theGiustarini et al. 2013 protocol [30].
The present protocol consists of two subprotocols: one for thequantification of the NP-TRS and the other for the P-TRS parametersafter appropriate reduction of involved disulfide bonds (Fig. 1).
Fig. 1. Schematic representation of the quantification of nonprotein (NP) and protein (P) thiol parameters of TRS by the present protocol. The main NP and P TRS parameters(estimated after reduction of the P/NP symmetric/mixed disulfides with TBP or TCEP) are highlighted in green and orange bold, respectively. Abbreviations in parenthesisshow the reagents OPT, NHD, and DTP that are involved in the quantification of the indicated TRS parameters; optional methods for GSH (as GS-NEM) and GSSGquantification by HPLC and the GR recycling assay, respectively, are also stated (in parentheses). TRS parameters shown inside parentheses without the equality symbol arethose derived by corresponding algebraic equations (see section “Estimation of the TRS parameters”), which involve certain fractions (A to L) quantified by the protocol. (Forinterpretation of the references to color in this figure legend, the reader is referred to the web version of this article.)
K. Grintzalis et al. / Free Radical Biology and Medicine 74 (2014) 85–98 87

In addition to GSH and GSSG, the NP-TRS protocol quantifies theNP-thiols/disulfides GSSNP (¼ GSHGSSNP), CSH, CSSNP (¼ CSHCSSNP),NPSH (¼ GSHþCSHþNPxSHNPSH), NPxSHNPSH (the unidentified NPthiol component/s of NPSH, excluding GSH and CSH), NPSSNP (¼GSSGþGSSNPþCSSNPþNPxSHNPSSNP), and NPxSHNPSSNP (the uni-dentified NP thiol component/s of NPSSNP, excluding GSH andCSH); GSH, GSSNP, NPSSNP, CSSNP, NPxSHNPSH, and NPxSHNPSSNP
are derived algebraically. The P-TRS subprotocol measures theP-thiols/disulfides PSH (containing reduced protein cysteines), PSSG,PSSC, PSSNP (¼ PSSGþPSSCþNPxSHPSSNP), NPxSHPSSNP (the uniden-tified NP thiol component/s of PSSNP, excluding GSH and CSH), andPSSP (intra- plus interprotein disulfides); PSSP and NPxSHPSSNP arederived algebraically.
The two subprotocols are based on three different thiol probes.These are the photometric reagents DTP and ninhydrin (NHD), andthe fluorometric reagent OPT (Fig. 2). Although the mechanism ofOPT reaction with GSH is already known (i.e., via formation ofcovalent bonds, one with the sulfhydryl and the other with the ε-amino groups of cysteine and lysine, respectively [33]), the factthat OPT also reacts specifically with GSSG at pH Z12 [32,33,60]has not been explained. We propose a possible mechanisminvolving GSSG reaction with OPT at alkaline pH Z12, where itis hydrolyzed to GS̄ (Fig. 2) as similarly are hydrolyzed proteindisulfides and low molecular weight aliphatic and aromatic dis-ulfides (2RSSRþ4OH̄-3RS̄þRSO2̄þ2 H2O [61]). Then, GS̄ wouldreact with OPT to form a GS-OPT fluorophore [33]. It should benoted that GS-OPT remains stable at the pH range 8-12, asindicated by the stability of its fluorescence [60]. Thus, it is stableat pHs 8 and 12 used for GSH and GSSG determination,
respectively [32]. Below pH 8, GS-OPT is unstable as shown bythe decrease of its fluorescence intensity [60].
The initial DTP-, NHD-, and OPT-based assays have beenmodified in the present protocol for increased specificity, stability,and ease of use. The DTP reagent quantifies the NP thiols NPSH,NPSSNP, PSH, PSSNP, PSSP, NPxSHNPSH, NPxSHNPSSNP, andNPxSHPSSNP. For the determination of the -SH groups in the Pthiols PSH and NPSH, in the PS- part of PSSP and PSSNP (after theirreduction), and in the NPS- part of PSSNP and NPSSNP (after theirreduction), we chose DTP instead of DTNB for the followingreasons: DTP reacts with thiols in a thiol-disulfide exchangereaction accompanied by the stoichiometric release of the chro-mogenic compound 4-thiopyridone. Because the electrostaticmilieu of cysteine thiols in proteins may have substantial effectson the rates of thiol-disulfide exchange reactions, DTP has a greatadvantage over DTNB due to its hydrophobic nature and some-what small size. Protonation of the pyridyl nitrogens increases thereactivity of DTP dramatically. This almost completely compen-sates for the decrease in reaction rate at low pH due to protonationof the nucleophilic thiolate anion. Therefore, the reagent can beused at low pH where thiol-disulfide exchange reactions normallydo not occur. DTP gives off only two 4-PS moles when reactingwith NPSH or mixed (P/NP) disulfides (after their reduction), andis not involved in SH/SS exchange reactions, making it the best -SHreagent [10,11]. On the other hand, DTNB-based assays have thefollowing disadvantages: 5-thio-2-nitrobenzoate, the product ofDTNB reaction with -SH groups, can be also formed by thehydrolysis of DTNB, making DTNB not a safe quantitative indicatorof thiols. DTNB instability due to hydrolysis has been estimated asan artificial 1.2 μM h–1 increase in thiol concentration at pH 8.0[40]. Another limitation in using DTNB is that unwanted oxidationof thiols occurs at pH values above 7.0 (DTNB assays are performedusually at pH 7.5), because SH/SS exchange reactions of the finalproduct are generally occurring at this pH range [45].
The NHD reagent quantifies specifically free CSH and the CSpart of CSSNP and PSSC after their reduction. NHD allows thediscrimination of CSH in mixtures of NP thiols that are determinedby the nonspecific DTP and DTNB assays. The initial NHD-basedassay [51,62] has been modified in the present protocol in order tostabilize the absorbance of the chromophore and increase thesensitivity of the method.
The OPT reagent quantifies specifically GSH and GSSG, and theGS part of GSSNP and PSSG after their reduction. The initial assay[32] has been modified for use in the present protocol in order tobecome more specific by removing interfering proteins [33,34]. Forthis, we replaced the metaphosphoric acid step in the initial assayby use of a more effective precipitation step; we used a certainDOC-TCA combination which precipitates as slow as 5 mg ml�1
proteins with Z95% efficiency [58,59] (thus also allowing themeasurement of very low protein samples). This step is followedby a protein pellet and aqueous fraction washing step with diethylether, in order to remove any TCA traces (besides DOC), whichinterfere with the OPT assay. These modifications achieve a�4000-fold increase in sensitivity for GSH and GSSG over theinitial assay, due to the high sample dilutions it requires in order toeliminate metaphosphoric acid interference. Moreover, the proteinthiol TRS parameters can be evaluated in samples with low proteinconcentration, as it is estimated by an ultrasensitive (100–200 ng)Coomassie brilliant blue (CBB) G-250 assay [63].
The determination of NP and P thiol disulfides requires priorreduction of -SS- bonds, which is commonly done by dithiothrei-tol. However, the reduction reaction is rather time-consuming andmost importantly there is an absolute requirement for the com-plete removal of dithiothreitol prior P/NP thiol quantification [40].An alternative reductant, borohydride, has been also employed,taking advantage of the easy removal of its excess by acid
Fig. 2. Reaction mechanisms of thiol/mixed disulfide-based -SH groups, cysteines,and GSSG/GSH with the respective DTP [40], NHD [71], and OPT reagents used inthe multiparametric TRS protocol. OPT use in the GSSG determination involvesalkaline hydrolysis of GSSG (at pH 12), where the resulting GS̄ reacts with OPT toform the GS-OPT fluorophore (as also GSH does) [33] (a detailed explanation of theOPT-based GSSG assay reaction mechanism is presented in the Introduction).
K. Grintzalis et al. / Free Radical Biology and Medicine 74 (2014) 85–9888

neutralization. However, its reductive effectiveness is low at thealkaline pH required for certain parameters of TRS [50,51]. Wechose trialkylphosphines because they are very strong -SS- groupreductants (more powerful than dithiothreitol [48]), reduce dis-ulfides stoichiometrically and irreversibly, and are nonreactivetoward many other functional groups [49]; TCEP [39] and TBP[37,38] have been already used in disulfide bond reduction.Hydrophobic TBP was selected for the DTP-based assays in theprotocol because it can be easily removed by extraction in butanol-cyclohexane (5:1 v/v). On the other hand, the water-soluble TCEPwas preferred for the NHD- and OPT-based assays because it doesnot interfere with them.
The present protocol was applied in human plasma, a sample ofestablished clinical value, for two main reasons: (1) As anyextracellular fluid, blood plasma is considered a “difficult” samplebecause of its low concentration levels on GSH and GSSG (bothpresent at �1 to 2 μM) [30], and (2) because the evaluation of theother 13 parametres of TRS could be also of clinical usefulness.
Materials
Instrumentation
Balance (Kern, 770/65/6 J)Bench-top centrifuge (Hermle, Model Z206A)Glass Pasteur pipettes (internal diameter 0.5 cm, 22 cm long, by
Hirschmann Laborgeräte GmbH & Co, Germany)Microcentrifuge clear tubes, 1.5 and 2 ml (VWR, Cat. No. 89000-
028)Micropipettes (adjustable volume) 2.5 μl, 10 μl, 20 μl, 100 μl,
200 μl, 1 ml, and tips (Eppendorf Research)Microcuvette for absorbance measurements (12.5�12.5�45 mm
external dimensions, 4 mm internal window, and 9 mm bottom,1.16 ml, quartz; Starna 9/B/9/Q/10)
Microcuvette for fluorescence measurements (45�4 mm,0.5 ml, quartz; Starna SOG/Q), with its FCA4 adapter
pH meter (Metrohm, 827 pHlab)Spectrofluorometer (Shimadzu, Model RF-1501)Spectrophotometer (Hitachi, Model UV-VIS U-1800)
Reagents
Acetic acid (glacial) 100% (Merck, Cat. No. 1.00063.1011)!CAUTION Corrosive.
Acetone (AC; Merck, Cat. No. 01-6300117)! CAUTION Highlyflammable and harmful.
Acivicin (Sigma-Aldrich, Cat. No. SML0312).Aldrithiol (4,40-dithiopyridine or DTP; Sigma-Aldrich, Cat. No.
143057)! CAUTION Irritant.Catalase from bovine liver (CAT; Sigma, Cat. No. C-9322).Chloroform (CHCl3; Merck, Cat. No. 1.02445)! CAUTION Highly
flammable and harmful.Cumene hydroperoxide, CumOOH (Sigma, Cat. No. C-0524)!
CAUTION Oxidizing, Corrosive.Cyclohexane (Sigma-Aldrich, Cat. No. 179191)! CAUTION Highly
flammable and harmful.L-Cysteine (CSH; Sigma-Aldrich, Cat. No. C-7352).Deoxycholic acid, sodium salt (DOC; Sigma-Aldrich, Cat. No.
D-6750).Diethyl ether (DE; Merck, Cat. No. 1.00921)! CAUTION Highly
flammable and harmful.Dimethyl sulfoxide (DMSO; Sigma-Aldrich, Cat. No. 276855)!
CAUTION Harmful.Ethylenediaminetetraacetic acid (Na2EDTA; Merck, Cat. No.
324503)! CAUTION Irritant, dangerous for the environment.
Formamide (100% w/w or 25 M, Merck, Cat. No. 1.09684.1000)!CAUTION Very toxic.
Glutathione (GSH; Sigma-Aldrich, Cat. No.G-4251).L-Glutathione oxidized disodium salt (GSSG; Sigma-Aldrich,
Cat. No. G-4626).Glutathione peroxidase from bovine erythrocytes (GPx; Sigma-
Aldrich, Cat. No. G-6137).HCl conc. (Z37% w/w; Fluka, Cat. No. 84415)! CAUTION
Corrosive.Isobutanol (Merck, Cat. No. 1.00984.1000)! CAUTION Irritant,
dangerous for the environment.Methanol (MetOH; 100%) for HPLC (Sigma-Aldrich, Cat. No.
34860)! CAUTION Highly flammable and harmful.N-Ethylmaleimide (NEM; Sigma-Aldrich, Cat. No. E-3876)!
CAUTION Highly flammable and corrosive.Ninhydrin (NHD; Serva, Cat. No. 30410).N,N-Dimethylformamide (DMF, Merck, Cat. No. 1.03053.1000)!
CAUTION Very toxic.Sodium phosphate (Νa2HPO4; Merck, Cat. No. 30412).o-Phthaldialdehyde (OPT; Sigma-Aldrich, Cat. No. P-1378)!
CAUTION Irritant.NaOH (Merck, Cat. No. 567530)! CAUTION Corrosive.Tri-n-butylphosphine 97% (TBP; Sigma-Aldrich, Cat. No.
247049)! CAUTION Highly flammable and corrosive.Tris(2-carboxyethyl)phosphine hydrochloride, powder, Z98%
(TCEP; Sigma-Aldrich, Cat. No. C4706)! CAUTION Highly flam-mable and corrosive.
Trichloroacetic acid (TCA; Merck, Cat. No. 1.00807.0250)! CAU-TION Corrosive, dangerous for the environment.
Urea (Sigma-Aldrich, Cat. No. U1250).Water (ddH2O; purified by a Milli-Q system, Millipore Corp).
Reagent setup
� 100% AC: Keep at -20 1C.� 500 mM phosphate buffer, 1 mM acivicin, 30 mM EDTA, pH 7.5
(500PAE buffer, pH 7.5): Prepare fresh by dissolving 0.891 gNa2 HPO4.2H2O, 111 mg Na2EDTA, 1.8 mg in 8 ml ddH2O. Adjustthe pH to 7.5 and final volume of the solution to 10 ml.
� 2000 Units CAT ml-1: Prepare fresh by dissolving 1 mg CAT in1 ml ddH2O. Keep on ice.
� 500 mM phosphate buffer, 1 mM acivicin, 30 mM EDTA,300 mM NEM, pH 7.5 (500PAEN buffer, pH 7.5): Prepare freshby dissolving 37.5 mg NEM in 1 ml 500PAE buffer, pH 7.5. Keepon ice.
� 300 Units GPx ml-1: Prepare fresh by dissolving 1 mg GPx in1 ml ddH2O. Keep on ice.
� 10 mM CumOOH: Dilute CumOOH stock (5.52 M) to 50 mMwith 100% MetOH. Dilute further this stock to 10 mM withddH2O.� 50, 100% TCA: Prepare the 100% TCA fresh by dissolving 10 gTCA in ddH2O to a final 10 ml. Prepare the 50% TCA by 2Xdilution of 100% TCA with ddH2O.� 1% DOC: Prepare fresh by dissolving 0.01 g DOC in ddH2O tofinal 1 ml.
� 10% TCA, 0.02% DOC, 50PAE buffer, pH 7.5, washed with DE(10TCA-DOC-50PAE-DE): Prepare fresh by mixing 2.5 ml 100%TCA with 19.5 ml ddH2O, 2.5 ml 500PAE buffer, pH 7.5, and0.5 ml 1% DOC, followed by centrifugation at 16,000g for 2 min.Then, wash 3� with 25 ml DE by vortexing, followed bycentrifugation at 16,000g for 2 min and discard the upperDE phase.
� 10% TCA, 0.02% DOC, 50PAEN buffer, pH 7.5, washed with DE(10TCA-DOC-50PAEN-DE): Prepare fresh by mixing 0.5 ml 100%TCAwith 3.9 ml ddH2O, 0.5 ml 500PAEN buffer, pH 7.5, and 0.1 ml1% DOC, followed by centrifugation at 16,000g for 2 min.
K. Grintzalis et al. / Free Radical Biology and Medicine 74 (2014) 85–98 89

Then, wash 3� with 5 ml DE by vortexing, followed by centri-fugation at 16,000g for 2 min and discard the upper DE phase.
� 4, 0.1 M Acetic acid, pH 4.5: For preparing the 4 M solution,dilute the concentrated stock (16.6 M) 4.165X with ddH2O andadjust to pH 4.5 with 10 M NaOH. For preparing the 0.1 Msolution dilute the 4 M stock solution 40X with ddH2O, andadjust to pH 4.5 with 1 M NaOH.
� 12 Μ Formamide: Prepare fresh by mixing 0.25 ml 4 M aceticacid, pH 4.5, with 25.75 ml ddH2O and 24 ml 100% formamide,and adjust pH to 4.5 with 1 M NaOH.
� 0.5, 1 M phosphate buffer, pH 8.0: Prepare the 1 M solution bydissolving 7.12 g Na2 HPO4.2H2O in ddH2O to final 40 ml andadjust the pH to 8.0 (with 10 M HCl). The 0.5 M solution ismade by 2X dilution of the 1 M stock.
� 50 mM TBP: Prepare fresh by diluting 80X with DMF thecommercial TBP stock (4 M).
� 2.5 Μ Urea: Prepare fresh by dissolving 7.5 g urea in a finalvolume 50 ml ddH2O.� 50 mM TCEP: Prepare fresh by dissolving 14 mg TCEP in 1 mlddH2O. The reagent must be prepared fresh before use.
� 10% TCA, 0.02% DOC, 7 M formamide, 0.25 M phosphate buffer(pH 8.0), 2 mM TBP (TCA-DOC-F-P-T): Prepare fresh (20.4 ml)by mixing 12 ml 12 Μ formamide with 5.2 ml 1 M phosphatebuffer, pH 8.0, 800 μl 50 mM TBP, 400 ml 1% DOC, and 2 ml 50%TCA, followed by centrifugation at 16,000g for 2 min.
� 10% TCA, 0.02% DOC, 2 M urea, 8 mM phosphate buffer (pH 8.0),2 mM TCEP (TCA-DOC-U-P-T): Prepare fresh (12 ml) by mixing10 ml 2.5 M urea with 200 μl 0.5 M phosphate buffer, pH 8.0,500 μl 50 mM TCEP, 240 ml 1% DOC (final 0.02%), 1200 μl 100%TCA (final 10%), followed by centrifugation at 16,000g for 2 min.Then, wash 3� with equal volume DE by vortexing, followedby centrifugation at 16,000g for 2 min and discard the upperDE phase.
� 3 mM DTP: Prepare fresh by dissolving 7 mg DTP in 1 ml 100%DMSO and dilute 10X with ddH2O.� 10% DMSO: Dilute 10X with ddH2O the 100% DMSOcommercial stock.
� Acetic acid-HCl solution: Prepare fresh mixing 45 ml 100%acetic acid with 30 ml conc. HCl.
� 100 mM NHD: Prepare fresh by dissolving 0.79 g NHD in 40 mlacetic acid-HCl solution.
� 1 mg ml-1 OPT: Prepare fresh by dissolving 1 mg OPT in 1 ml100% MetOH. Keep on ice and light-protected.
� 0.2 M NaOH: Prepare by dissolving 0.16 g NaOH in ddH2O tofinal 20 ml.
� 5 mM GSSG: Prepare fresh by dissolving 3.3 mg GSSG in 1 ml10TCA-DOC-50PAE-DE. Keep on ice.
� 5 mM GSH: Prepare fresh by dissolving 3 mg GSH in 2 ml 10TCA-DOC-50PAE-DE. Keep on ice (stable for at least 2 weeks at -20 1C).
� 5 mM CSH: Prepare fresh by dissolving 3 mg CSH in 0.5 ml0.5 M HCl, and then add 4.5 ml 10TCA-DOC-50PAE-DE. Keep onice (stable for at least two weeks at -20 1C).
Construction of standard curves
Standard curve for -SH groups in fractions A, B, H, I, and J (fordefinitions of fractions A, B, H, I, and J see Method procedure):From the 5 mM GSH solution, prepare a series of GSH standards (1to 17 μM, in 10TCA-DOC-50PAE-DE) and mix 900 μl of each with50 μl 4 M acetic acid, pH 4.5, and 50 μl 3 mM DTP. As reagentblank, 900 μl 10TCA-DOC-50PAE-DE in place of the GSH solution isused. After 10 min incubation at room temperature (RT), theabsorbance of the mixtures is measured at 325 nm against thereagent blank using a photometric quartz microcuvette. Sensitiv-ity: 1 nmol -SH.
Standard curve for fractions C, and D (for definitions of fractionsC and D see Method procedure): From the 5 mM CSH solution,prepare a series of CSH standards (1 to 12 μM, in 10TCA-DOC-50PAE-DE) and mix 2 ml of each with 2 ml 100% acetic acid and2 ml 100 mM NHD. As reagent blank, 2 ml 10TCA-DOC-50PAE-DEin place of the CSH solution is used. After 10 min incubation at100 1C, let the tubes cool, add to each 1 ml chloroform, vortexvigorously, centrifuge at 16,000g for 5 min, transfer the chloroformbottom phase into a glass microcuvette, and measure absorbanceat 560 nm against the reagent blank. Sensitivity: 1 nmol CSH.
CRITICAL: If the chloroform phase that results after the cen-trifugation step is not clear, you should not measure itsabsorbance because you will get erroneously high readings.The slight turbidity is due to the presence of minute waterdroplets entrapped in the chloroform phase during vortexingbecause the NHD reaction mixtures were not cooled well (toRT) before adding chloroform. To remove turbidity eitherrecentrifuge the chloroform phase after recooling in ice waterbath, or warm it shortly at 30 1C.
Standard curve for fraction K (¼ PSSC) (for definition of fractionK see Method procedure): From the 5 mM CSH solution, prepareCSH standards (1 to 12 μM in TCA-DOC-F-P-T) and mix 2 ml ofeach with 2 ml 100% acetic acid and 2 ml 100 mM NHD. As reagentblank, 2 ml TCA-DOC-F-P-T in place of the CSH solution is used.After 10 min incubation at 100 1C, let the tubes cool, add to each1 ml chloroform, vortex vigorously, centrifuge at 16,000g for 5 minat RT. Isolate the chloroform phase and wash 1 ml of it with 4 ml4 M acetic acid by vortexing, followed by centrifugation at 16,000gfor 5 min at RT, then isolate the acetic acid-washed chloroform(cleared from the color interference due to formamide), transfer itinto a glass microcuvette, and measure its absorbance at 560 nmagainst the reagent blank. Sensitivity: 1 nmol CSH.
CRITICAL: Same as in Standard curve for fraction C, D.CAUTION: Avoid plastic photometric cuvettes as they might beeroded by the chloroform.
Standard curve for fractions E (¼ GSSG), and F (for definitions offractions E and F see Method procedure): From the 5 mM GSSGsolution, prepare a series of GSSG standards (2 to 20 μM, in 10TCA-DOC-50PAE-DE) and mix 230 μl of each with 110 μl 0.2 M NaOHand 20 μl 1 mg ml-1 OPT. As reagent blank, 230 μl 10TCA-DOC-50PAE-DE in place of the GSSG solution is used. After 20 minincubation at RT in the dark, the fluorescence at ex/em 340/420 nm is measured against the reagent blank using a fluorometricquartz microcuvette (with the spectrofluorometer in use set at lowsensitivity and slit width 10 nm). Sensitivity: 0.4 nmol GSSG.
Standard curve for fraction G (for definition of fraction G seeMethod procedure): From the 5 mM GSH solution, prepare a seriesof GSH standards (1 to 15 μM, in 10TCA-DOC-50PAE-DE) and mix190 μl of each with 20 μl 50 mM TCEP, 130 μl 0.5 M phosphatebuffer, pH 8.0, and 20 μl 1 mg ml-1 OPT. As reagent blank, 190 μl10TCA-DOC-50PAE-DE in place of the GSH solution is used. After20 min incubation at RT in the dark, the fluorescence at ex/em340/420 nm is measured against the reagent blank using afluorometric quartz microcuvette (with the spectrofluorometerin use set at low sensitivity and slit width 10 nm). Sensitivity:0.2 nmol GSH.
Standard curve for fraction L (¼PSSG) (for definition of fraction Lsee Method procedure): From the 5 mM GSH solution, prepare aseries of GSH standards (3 to 35 μM, in TCA-DOC-U-P-T) and mix200 μl with 130 μl 0.5 M phosphate buffer, pH 8.0, and 20 μl 1 mgml-1 OPT is added. As reagent blank, 200 μl TCA-DOC-U-P-T inplace of the GSH solution is used. After 20 min incubation at RT inthe dark, the fluorescence at ex/em 340/420 nm is measured
K. Grintzalis et al. / Free Radical Biology and Medicine 74 (2014) 85–9890

against the reagent blank using a fluorometric quartz microcuvette(with the spectrofluorometer in use set at low sensitivity and slitwidth 10 nm). Sensitivity: 0.6 nmol GSH.
Method procedure
The protocol procedure (outlined in Fig. 3) is as follows:
Sample treatment
1. Split sample (internal organs, solid tissues, cells, etc; for bloodsee as follows) in 1/5 and 4/5 portions (expressed in weight orvolume for tissue/cell or liquid samples, respectively) andhomogenize as follows:(a) The 1/5-sample is initially mixed with an appropriate
volume of cold ddH2O (as to keep the final homogenizationvolume as low as possible), and to the resulting mixturevolume 1/10 vol 500PAEN buffer, pH 7.5, is added, followedby homogenization (with a homogenization method appro-priate for the sample type), and incubation for 5 min atroom temperature (RT). The blood plasma 1/5-sample iscollected in microcentrifuge tubes prefilled with 1/10 vol500PAEN buffer, pH 7.5, and mixed by tilting four to fivetimes for 5 min at RT. The 1/5-sample homogenate iscentrifuged at 16,000g for 10 min at 4 1C, the resultingclear NEM-treated homogenate supernatant (designated1/5-NEM-hs) is collected, and its protein content is mea-sured as stated elsewhere [63] in order to express its TRSparameters per protein amount.
PAUSE POINT: Samples can be stored at �80 1C becausecompared with results from fresh samples they did notaffect these results even when stored for 3 months.
NOTE: Alternatively, the 1/5-NEM-hs can be used toquantify both GSSG and GSH by the more complicated(although �5 fold more sensitive) methods described inthe Giustarini et al. 2013 protocol [30]; GSSG is deter-mined by the nonendpoint GR recycling assay as alsodescribed elsewhere [31], and GSH as GS-NEM by HPLC,starting from substeps A or B in step 2 of the “Analyticalprocedures” section [30].
(b) The 4/5-sample is also mixed with an appropriate (foreffective homogenization) volume of cold ddH2O, and tothe resulting mixture 1/10 vol 500PAE buffer, pH 7.5, and1/10 vol 2000 Units CAT ml�1 are added, followed byhomogenization; the blood plasma 4/5-sample is collectedin microcentrifuge tubes prefilled with 1/10 vol 500PAEbuffer, pH 7.5, and 1/10 vol 2000 Units CAT ml�1, and mixedby tilting four to five times. The 4/5-sample homogenate iscentrifuged at 16,000g for 10 min at 4 1C, the clearuntreated homogenate supernatant is collected (designated4/5-untr-hs), and its protein content is measured as statedin step 1a.
NOTE: Acivicin inhibits γ-GT [64], thus preventing thisenzyme from utilizing GSH as substrate, while NEM inhibitsGR (and limits artifactual ex vivo reduction of GSSG) andprevents GSH autoxidation by forming a GS-NEM adduct,thus allowing the accurate determination of GSSG [30,65].We did not use serine/borate to inhibit γ-GT in order tominimize possible interference by the homogenization buf-fers in the assays of the protocol. Instead, we counter-balanced the slow inactivation potential of acivicin byusing 4.5-fold higher concentration (100 mM) than the oneused in the Giustarini et al. 2013 protocol [30].NOTE: CAT was added to eliminate CSH autoxidation causedby H2O2 in the presence of transition metals [56,57].
Fig. 3. General outline of the multiparametric TRS protocol. TRS parameters are enclosed in white boxes, and are determined by the indicated steps (described in detail in thetext). Text in italics designates indicative starting sample fractions processed during the protocol procedure, as they are described in detail the protocol procedure.
K. Grintzalis et al. / Free Radical Biology and Medicine 74 (2014) 85–98 91

We verified this by spiking the sample homogenate withknown concentrations of CSH, and testing for recovery(Method Procedure, step 9, fraction C treatment; data notshown).
NOTE: The 1/5 and 4/5 sample portion splitting is based onthe need of a small fraction (�1/5) for the determination ofGSSG. It is a good starting approximation for most samples,and should be set experimentally considering also sampleGSSG concentration.
CRITICAL: When cell lysis homogenization buffers are used,they should not contain -SH groups (e.g., mercaptoethanol,dithiothreitol). Moreover, if they contain organic solventsand detergents these should be tested for interference withthe DTP, NHD and OPT assays of the protocol.
2. The 4/5-untr-hs (from step 1b) is further split into two 1/4 and3/4 vol portions. The 3/4 vol portion is not treated (designated3/4-untr-hs), while to the 1/4 vol portion 1/10 vol 10 mMCumOOH and 1/10 vol 300 units GPx ml�1 are added andincubated at RT for 15 min (designated 1/4-GPx-hs).NOTE: In this step, GPx will oxidize GSH specifically to GSSG.However, to ensure complete oxidation of GSH in the 1/4 volfraction, at least three dilutions of it (with 10X diluted 500PAEbuffer, pH 7.5) should be CumOOH/GPx-treated and give thesame total GSSG (¼ GSSGþGSSGGSH) concentration.
3. 1/5-NEM-hs, 1/4-GPx-hs, and 3/4-untr-hs are subjected to pro-tein precipitation by DOC-TCA (DT) [58,59]. Specifically, per1 ml 1/5-NEM-hs, 1/4-GPx-hs, and 3/4-untr-hs 20 μl 1% DOC(final 0.02%) are added and incubated for 10 min at RT, followedby addition of 115 μl 100% TCA (final 10%), incubation for 15 minin an ice-water bath, protein precipitation by centrifugation at16,000g for 5 min at 4 1C, and collection of the correspondingprotein free supernatants (pfsup) 1/5-DT-NEM-pfsup, 1/4-DT-GPx-pfsup, and 3/4-DT-untr-pfsup. The resulting correspondingprotein pellets (pp), designated 1/4-DT-GPx-pp and 3/4-DT-untr-pp, are also collected (to be used in step 4).
NOTE: The minimum protein concentration with Z90%efficiency by DOC-TCA precipitation is �2 mg ml-1 (for BSA[58,59]).Then, the 1/5-DT-NEM-pfsup, 1/4-DT-GPx-pfsup, and 3/4-DT-untr-pfsup are washed 3� with equal volume diethyl ether(DE) by vortexing, followed by centrifugation at 16,000g for5 min at 4 1C (and discarding of the upper DE phase). Then,the three washed bottom aqueous phases (aqph), desig-nated 1/5-DT-NEM-DE-aqph, 1/4-DT-GPx-DE-aqph, and 3/4-DT-untr-DE-aqph, are collected (to be used in subsequentsteps) and their exact volume is measured for normalizingthe quantification of their nonprotein thiols per supernatantprotein (before the DT-DE treatments).
NOTE: DE-washing of 1/5-DT-NEM-pfsup, 1/4-DT-GPx-pfsup,and 3/4-DT-untr-pfsup removes DOC and TCA (thus facilitat-ing their pH adjustment) and also any remains of MetOHand CumOOH from the 1/4-DT-GPx-pfsup, all of which mayinterfere in the assays applied on these aqueous phases insubsequent steps.
PAUSE POINT: Although sample immediately is recom-mended, DE-washed, protein free supernatants can bestored at -80 1C for analyzing them after maximum 2 days.
4. The 1/4-DT-GPx-pp and 3/4-DT-untr-pp (from step 3) arewashed once with 0.5 ml cold 100% acetone (AC) by vortexing,followed by centrifugation at 16,000g for 5 min at 4 1C, and air-drying of pellet remnant AC for 10 min at RT.
PAUSE POINT: Although it is generally recommended toanalyze samples immediately, AC-washed protein pellets
can be stored for maximum 1 week at -80 1C.The resulting AC-washed protein pellets, designated 1/4-DT-GPx-AC-pp and 3/4-DT-untr-AC-pp, are treated as follows:
(a) The 3/4-DT-untr-AC-pp is solubilized in minimum 6 ml 12 Mformamide, centrifuged at 16,000g for 5 min to collect theclear solubilizate (sol), designated 3/4-DT-untr-AC-ppsol (tobe used in step 19), and its protein concentration ismeasured in order to express its TRS parameters perprotein.
(b) The 1/4-DOC-TCA-GPx-AC-pp is solubilized in 0.2 ml 6 Murea, and then adjusted to 2.5 M urea by appropriatedilution with ddH2O and centrifuged at 16,000g for 5 min.The resulting clear solubilizate (sol), designated 1/4-DT-GPx-AC-ppsol, is collected (to be used in step 29) and itsprotein content is measured in order to express its TRSparameters per protein amount.
Subprotocol for nonprotein thiols
Fraction A [¼ NPSH (¼ GSHþCSHþNPxSHNPSH)]
5. Mix two 900-μl portions (for sample blank, SB, and sample, S)3/4-DT-untr-DE-aqph (obtained from step 3; further diluted ifneeded with 10TCA-DOC-50PAE-DE) with the reagents shownin Table 1:
6. Incubate reagent mixtures for 10 min at RT and measure theabsorbance of the mixtures at 325 nm, using a photometricquartz microcuvette. Subtract from the S value the values ofthe SB and RB, and the resulting net absorbance is convertedto -SH group (GSH equivalent) concentration from the Stan-dard curve for -SH groups in fractions A, B, H, I, J (see “Reagentsetup” section).Fraction B {¼ NPSH (¼ Fraction A)þNPSHNPSSNP [¼ GSHGSSG
(¼ 2GSSG)þGSHGSSNP (¼ GSSNP)þNPxSHGSSNPþCSHCSSNP (¼CSSNP)þNPxSHCSSNPþNPxSHNPxSSNPx]}, for indirect NPSSNPdetermination
7. Mix 2700 μl 3/4-DT-untr-DE-aqph (obtained from step 3; furtherdiluted if needed with 10TCA-DOC-50PAE-DE) with 150 μl 1 Mphosphate buffer, pH 8.0, and 150 μl 50 mM TBP incubate for30 min at 50 1C. As reagent blank, 2700 μl 10TCA-DOC-50PAE-DE in place of the sample is used. To both mixtures add 3 mlbutanol-cyclohexane (BC) (5:1 v/v), vortex, and centrifuge at16,000g for 5 min to isolate the corresponding bottom aqueousphases, designated sample-BC-aqph and RB-CB-aqph. Mix two900-μl portions (for SB and S) sample-BC-aqph and 900 ml RB-BC-aqph with the reagents shown in Table 2:
8. Incubate the reagent mixtures for 10 min at RT and measurethe absorbance of the mixtures at 325 nm. Subtract from the Svalue the values of the SB and RB, and the resulting netabsorbance is converted to GSH concentration from theStandard curve for -SH groups in fractions A, B, H, I, J (see“Reagent setup” section).Fraction C¼CSH
9. Mix two 2000-ml portions (for SB and S) 3/4-DT-untr-DE-aqph(obtained from step 3; further diluted, if needed, with 10TCA-DOC-50PAE-DE) with the reagents shown in Table 3:
Table 1
Reagents (μl) Reagent blank (RB) Sample blank (SB) Sample (S)
3/4-DT-untr-DE-aqph – 900 90010TCA-DOC-50PAE-DE 900 – –
4.5 M acetic acid, pH 4.5 50 50 503 mM DTP 50 – 5010% DMSO – 50 –
K. Grintzalis et al. / Free Radical Biology and Medicine 74 (2014) 85–9892
10. Incubate the reagent mixtures for 10 min at 100 1C, let thetubes cool in an ice-water bath, and add 1 ml chloroform.Vortex the tubes and centrifuge them to separate the chloro-form (bottom) phase. Measure the absorbance of the chloro-form phase at 560. Subtract from the S value the values of theSB and RB, and the resulting net absorbance is converted toCSH concentration from the Standard curve for fractions C, D(see “Reagent setup” section).
CRITICAL: Same as in Standard curve for fractions C, and D.Fraction D [¼ CSH (¼ Fraction C)þCSHCSSNP (¼ CSSNP)], forindirect CSSNP determination
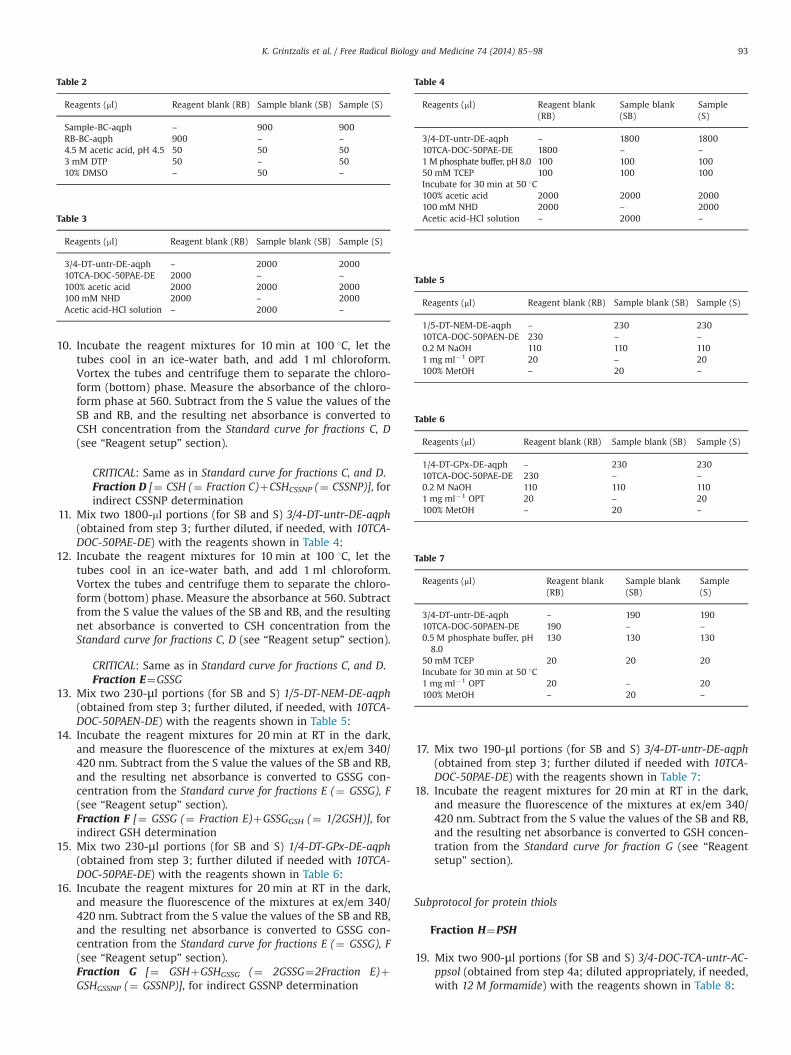
11. Mix two 1800-ml portions (for SB and S) 3/4-DT-untr-DE-aqph(obtained from step 3; further diluted, if needed, with 10TCA-DOC-50PAE-DE) with the reagents shown in Table 4:
12. Incubate the reagent mixtures for 10 min at 100 1C, let thetubes cool in an ice-water bath, and add 1 ml chloroform.Vortex the tubes and centrifuge them to separate the chloro-form (bottom) phase. Measure the absorbance at 560. Subtractfrom the S value the values of the SB and RB, and the resultingnet absorbance is converted to CSH concentration from theStandard curve for fractions C, D (see “Reagent setup” section).
CRITICAL: Same as in Standard curve for fractions C, and D.Fraction E¼GSSG
13. Mix two 230-μl portions (for SB and S) 1/5-DT-NEM-DE-aqph(obtained from step 3; further diluted, if needed, with 10TCA-DOC-50PAEN-DE) with the reagents shown in Table 5:
14. Incubate the reagent mixtures for 20 min at RT in the dark,and measure the fluorescence of the mixtures at ex/em 340/420 nm. Subtract from the S value the values of the SB and RB,and the resulting net absorbance is converted to GSSG con-centration from the Standard curve for fractions E (¼ GSSG), F(see “Reagent setup” section).Fraction F [¼ GSSG (¼ Fraction E)þGSSGGSH (¼ 1/2GSH)], forindirect GSH determination
15. Mix two 230-μl portions (for SB and S) 1/4-DT-GPx-DE-aqph(obtained from step 3; further diluted if needed with 10TCA-DOC-50PAE-DE) with the reagents shown in Table 6:
16. Incubate the reagent mixtures for 20 min at RT in the dark,and measure the fluorescence of the mixtures at ex/em 340/420 nm. Subtract from the S value the values of the SB and RB,and the resulting net absorbance is converted to GSSG con-centration from the Standard curve for fractions E (¼ GSSG), F(see “Reagent setup” section).Fraction G [¼ GSHþGSHGSSG (¼ 2GSSG¼2Fraction E)þGSHGSSNP (¼ GSSNP)], for indirect GSSNP determination
17. Mix two 190-μl portions (for SB and S) 3/4-DT-untr-DE-aqph(obtained from step 3; further diluted if needed with 10TCA-DOC-50PAE-DE) with the reagents shown in Table 7:
18. Incubate the reagent mixtures for 20 min at RT in the dark,and measure the fluorescence of the mixtures at ex/em 340/420 nm. Subtract from the S value the values of the SB and RB,and the resulting net absorbance is converted to GSH concen-tration from the Standard curve for fraction G (see “Reagentsetup” section).
Subprotocol for protein thiols
Fraction H¼PSH
19. Mix two 900-μl portions (for SB and S) 3/4-DOC-TCA-untr-AC-ppsol (obtained from step 4a; diluted appropriately, if needed,with 12 M formamide) with the reagents shown in Table 8:
Table 2
Reagents (μl) Reagent blank (RB) Sample blank (SB) Sample (S)
Sample-BC-aqph – 900 900RB-BC-aqph 900 – –
4.5 M acetic acid, pH 4.5 50 50 503 mM DTP 50 – 5010% DMSO – 50 –
Table 3
Reagents (ml) Reagent blank (RB) Sample blank (SB) Sample (S)
3/4-DT-untr-DE-aqph – 2000 200010TCA-DOC-50PAE-DE 2000 – –
100% acetic acid 2000 2000 2000100 mM NHD 2000 – 2000Acetic acid-HCl solution – 2000 –
Table 4
Reagents (ml) Reagent blank(RB)
Sample blank(SB)
Sample(S)
3/4-DT-untr-DE-aqph – 1800 180010TCA-DOC-50PAE-DE 1800 – –
1M phosphate buffer, pH 8.0 100 100 10050 mM TCEP 100 100 100Incubate for 30 min at 50 1C100% acetic acid 2000 2000 2000100 mM NHD 2000 – 2000Acetic acid-HCl solution – 2000 –
Table 5
Reagents (μl) Reagent blank (RB) Sample blank (SB) Sample (S)
1/5-DT-NEM-DE-aqph – 230 23010TCA-DOC-50PAEN-DE 230 – –
0.2 M NaOH 110 110 1101 mg ml�1 OPT 20 – 20100% MetOH – 20 –
Table 6
Reagents (μl) Reagent blank (RB) Sample blank (SB) Sample (S)
1/4-DT-GPx-DE-aqph – 230 23010TCA-DOC-50PAE-DE 230 – –
0.2 M NaOH 110 110 1101 mg ml�1 OPT 20 – 20100% MetOH – 20 –
Table 7
Reagents (μl) Reagent blank(RB)
Sample blank(SB)
Sample(S)
3/4-DT-untr-DE-aqph – 190 19010TCA-DOC-50PAEN-DE 190 – –
0.5 M phosphate buffer, pH8.0
130 130 130
50 mM TCEP 20 20 20Incubate for 30 min at 50 1C1 mg ml�1 OPT 20 – 20100% MetOH – 20 –
K. Grintzalis et al. / Free Radical Biology and Medicine 74 (2014) 85–98 93

20. Incubate the reagent mixtures for 10 min at RT and measurethe absorbance of the mixtures at 325 nm. Subtract from the Svalue the values of the SB and RB, and the resulting netabsorbance is converted to GSH concentration from theStandard curve for -SH groups in fractions A, B, H, I, and J (see“Reagent setup” section).
Reduction of protein -SS- groups (in PSSP, PSSNP, PSSC, PSSG)
21. Mix 4.5 ml 3/4-DT-untr-AC-ppsol (obtained from step 4a;diluted appropriately using 12 M formamide) with 1.95 ml1 M phosphate buffer, pH 8.0, and 0.3 ml 50 mM TBP andincubate for 30 min at 50 1C (designated 3/4-DT-untr-AC-ppsol-TBP). For the two reagent blanks (RB; one used in thesubsequent step 22a and the other in the step 22b) use forboth 4.5 ml 12 M formamide (in place of the 3/4-DOC-TCA-untr-AC-ppsol) mixed with 1.95 ml 1 M phosphate buffer, pH 8.0, and0.3 ml 50 mM TBP. The final volume of the reagent blanksolution for both reagent blanks is 6.75 ml, and is designatedRB-formamide-TBP.
22. The 3/4-DT-untr-AC-ppsol-TBP (prepared in step 21) is split in2.7 and 3.6 ml portions, which are treated as follows:(a) To 2.7 ml 3/4-DT-untr-AC-ppsol-TBP portion and to 2.7 ml
RB-formamide-TBP (both from step 21) add an equalvolume butanol-cyclohexane (BC) (5:1 v/v), vortex andcentrifuge at 16,000g for 5 min to isolate the bottomaqueous phase (free of TBP). To precipitate proteins inthe resulting 2.7 ml BC-washed bottom aqueous phase,60 ml 1% DOC (final 0.02%) is added and incubated for10 min at RT, and then 300 μl 100% TCA (final 10%) is addedand incubated for 15 min in an ice-water bath. Finally,proteins are precipitated by centrifugation at 16,000g for5 min at 4 1C, and the protein free supernatant (pfsup) iscollected (it contains NPSHPSSNP, which is the sumCSHPSSNPþGSHPSSNPþNPxSHPSSNP), its volume is mea-sured, and it is designated 3/4-2DT-untr-AC-ppsol-TBP-BC-pfsup. Similarly, the BC-washed RB-formamide-TBP isbrought to 0.02% DOC and 10% TCA by addition of 60 ml1% DOC and 300 μl 100% TCA, respectively, followed bycentrifugation at 16,000g for 5 min at 4 1C, and the super-natant is designated RB-formamide-TBP-BC-DT. Two 900-mlportions 3/4-2DT-untr-AC-ppsol-TBP-BC-pfsup, one for sam-ple blank (SB) and one for sample (S), and 900 ml from RB-formamide-TBP-BC-DT (for RB; the remaining can be usedfor making two more RB repeats) will be used in step 23.
(b) The 3.6 ml 3/4-DT-untr-AC-ppsol-TBP portion (from step21) is mixed with 80 ml 1% DOC (final 0.02%), incubated for10 min at RT, then mixed with 400 μl 100% TCA (final 10%),and incubated for 15 min in an ice-water bath. Then,proteins are precipitated by centrifugation at 16,000g for5 min at 4 1C, and the protein pellet (containing all -SS-groups reduced) is collected (for use in step 25) anddesignated 3/4-2DT-untr-AC-ppsol-TBP-pp. The resultingprotein free supernatant (pfsup), designated 3/4-2DT-
untr-AC-ppsol-TBP-pfsup, is also collected (it containsNPSHPSSNP, which is the sum CSHPSSNPþGSHPSSNPþNPxSHPSSNP) and its volume is measured. Similarly, a3.6 ml portion of RB-formamide-TBP (from step 21) isbrought to 0.02% DOC and 10% TCA by addition of 80 ml1% DOC and 400 μl 100% TCA, respectively, followed bycentrifugation at 16,000g for 5 min at 4 1C, and the super-natant is designated RB-formamide-TBP-DT. Two 2000-mlportions from 3/4-2DT-untr-AC-ppsol-TBP-pfsup, one forsample blank (SB) and one for sample (S), and 2000 mlfrom RB-formamide-TBP-DT (for RB; the remaining can beused for making one more RB repeat) will be used instep 27.
Fraction I [¼ NPSHPSSNP (¼ CSHPSSNPþGSHPSSNPþNPxSHPSSNP)],for PSSNP determination
23. Mix the 900 ml SB, S and RB fractions from step 22a with thereagents shown in Table 9:
24. Incubate the reagent mixtures for 10 min at RT and measurethe absorbance of the mixtures at 325 nm. Subtract from the Svalue the values of the SB and RB, and the resulting netabsorbance is converted to GSH concentration from theStandard curve for -SH groups in fractions A, B, H, I, and J (see“Reagent setup” section).Fraction J [¼ totPSH¼PSH (¼ Fraction H)þ2PSHPSSPþPSHPSSNP
(¼ Fraction I)], for PSSP determination25. Solubilize the 3/4-2DT-untr-AC-ppsol-TBP-pp (obtained in step
22b) in a minimum volume (up to 1.8 ml) 12 M formamide,centrifuge at 16,000g for 5 min to collect the clear proteinsolubilizate (designated 3/4-2DT-untr-AC-TBP-2ppsol) andmeasure its protein concentration in order to express theresult of the PSH sum per protein amount. Mix two 900-μlportions (for SB and S) 3/4-2DT-untr-AC-TBP-2ppsol (diluted ifneeded with 12 M formamide) with the reagents shown inTable 10:
26. Incubate the reagent mixtures for 10 min at RT and measurethe absorbance of the mixtures at 325 nm. Subtract from the Svalue the values of the SB and RB, and the resulting netabsorbance is converted to GSH concentration from theStandard curve for -SH groups in fractions A, B, H, I, and J (see“Reagent setup” section).Fraction K [¼ CSHPSSNP (¼ PSSC)]
27. Mix the 2000 ml SB, S and RB fractions from step 22b with thereagents shown in Table 11:
Table 8
Reagents (μl) Reagent blank(RB)
Sample blank(SB)
Sample(S)
3/4-DOC-TCA-untr-AC-ppsol
– 900 900
12 M formamide 900 – –
0.1 M acetic acid, pH 4.5 50 50 503 mM DTP 50 – 5010% DMSO – 50 –
Table 9
Reagents (μl) Reagent blank(RB)
Sample blank(SB)
Sample(S)
3/4-2DT-untr-AC-ppsol-TBP-BC-pfsup
– 900 900
RB-formamide-TBP-BC-DT 900 – –
0.1 M acetic acid, pH 4.5 50 50 503 mM DTP 50 – 5010% DMSO – 50 –
Table 10
Reagents (μl) Reagent blank(RB)
Sample blank(SB)
Sample(S)
3/4-2DT-untr-AC-TBP-2ppsol
– 900 900
12 M formamide 900 – –
0.1 M acetic acid, pH 4.5 50 50 503 mM DTP 50 – 5010% DMSO – 50 –
K. Grintzalis et al. / Free Radical Biology and Medicine 74 (2014) 85–9894

28. Incubate the reagent mixtures for 10 min at 100 1C, let thetubes cool in an ice-water bath and add 1 ml chloroform.Vortex the tubes and centrifuge them to separate the chloro-form phase (bottom). Remove the interference from formamideby isolating the chloroform phase after vortexing with 4 mlacetic acid, followed by centrifugation at 16,000g for 5 min, andmeasure absorbance at 560 nm. Subtract from the S value thevalues of the SB and RB, and the resulting net absorbance isconverted to CSH concentration from the Standard curve forfraction K (¼ PSSC) (see “Reagent setup” section).
CRITICAL: Same as in Standard curve for fractions C, and D.Fraction L [¼ GSHPSSNP (¼ PSSG)]
29. One ml 1/4-DT-GPx-AC-ppsol (obtained from step 4b; dilutedappropriately with 2.5 M urea) is mixed with 20 μl 0.5 Mphosphate buffer, pH 8.0, and reduced with 50 μl 50 mM TCEPby incubation for 30 min at 50 1C. As reagent blank (RB), 1 ml2.5 M urea (in place of 1/4-DT-GPx-AC-ppsol) is mixed with20 μl 0.5 M phosphate buffer, pH 8.0, and 50 μl 50 mM TCEP,and designated RB-urea-TCEP.
30. In the above 1.07 ml of the TCEP-reduced 1/4-DT-GPx-AC-ppsolproteins is precipitated by addition of 24 ml 1% DOC (final0.02%) and incubation for 10 min at RT, then addition of 120 μl100% TCA (final 10%) and incubation for 15 min in an ice-waterbath, and finally centrifugation at 16,000g for 5 min at 4 1C.The protein free supernatant (pfsup) is washed 3� with equalvolume DE (by vortexing and centrifugation at 16,000g for5 min at 4 1C, and discarding the upper DE phase). Theresulting DE-washed supernatant is collected (and designated1/4-2DT-GPx-AC-ppsol-TCEP-DE-pfsup) and its volume is mea-sured (it contains NPSHPSSNP, which is the sumCSHPSSNPþGSHPSSNPþNPxSHPSSNP). Similarly, the 1.07 ml RB-urea-TCEP portion (for RB from step 29) is brought to 0.02%DOC and 10% TCA by addition of 24 μl 1% DOC and 120 μl 100%TCA, respectively, followed by centrifugation at 16,000g for5 min at 4 1C. The resulting clear supernatant is washed 3�with equal volume DE (by vortexing and centrifugation at16,000g for 5 min at 4 1C, and discarding the upper DE phase),and the resulting DE-washed supernatant is designated RB-urea-TCEP-DT-DE. Two 200-ml portions 1/4-2DT-GPx-AC-ppsol-TCEP-DE-pfsup, one for sample blank (SB) and one for sample(S), and 200 ml RB-urea-TCEP-DT-DE (for RB), are mixed withthe reagents as shown in Table 12:
31. Incubate the reagent mixtures for 20 min at RT in the dark,and measure the fluorescence of the mixtures at ex/em 340/420 nm using a quartz microcuvette (with the spectrofluo-rometer in use set at low sensitivity). Subtract from thesample value the values of the sample and reagent blanksand the resulting net absorbance is converted to GSH con-centration from the Standard curve for fraction L (¼PSSG) (see“Reagent setup” section).
Estimation of the TRS parameters
The following equations were derived from the experimentalvalues for Fractions A to L as determined in the protocol procedure(for their Excel-adapted automated estimation see SupplementaryMaterial).
Nonprotein parameters
1. NPSH¼A (¼ GSHþCSHþNPxSHNPSH)2. NPSSNP¼½(B - A), deduced from B¼AþNPSHNPSSNP [¼
GSHGSSG (¼ 2GSSG)þGSHGSSNP (¼ GSSNP)þNPxSHGSSNP
þCSHCSSNP (¼ CSSNP)þNPxSHCSSNPþNPxSHNPxSSNPx]¼Aþ2NPSSNP
3. CSH¼C4. CSSNP¼D - C, deduced from D¼CSH (¼ C)þCSHCSSNP
(¼ CSSNP)5. GSSG¼E6. GSH¼2(F - E), deduced from F¼GSSG (¼ E)þGSSGGSH (¼ 1/
2GSH)7. GSSNP¼G-2F, deduced from G¼GSH [¼ 2(F - E)]þGSHGSSG (¼
2GSSG¼2E)þGSHGSSNP (¼ GSSNP)8. NPxSHNPSH¼A - C - 2(F - E), deduced from equalities (1), (3), (6)9. NPxSHNPSSNP¼B þ C - A - D - G - 2(EþF) (defined as the sum
NPxSHGSSNPþNPxSHCSSNPþNPxSHNPxSSNPx). It is deduced fromB¼AþNPSHNPSSNP [¼ GSHGSSG (¼ 2GSSG)þGSHGSSNP (¼GSSNP)þNPxSHGSSNPþCSHCSSNP (¼ CSSNP)þNPxSHCSSNPþNPxSHNPxSSNPx], and from equalities (1), (4),(5), (7)
Protein parameters
10. PSH¼H11. PSSNP¼I (¼ NPSHPSSNP¼CSHPSSNPþGSHPSSNPþNPxSHPSSNP)12. PSSP¼½(J - I - H), deduced from J¼totPSH¼PSH (¼ H)þ
2PSHPSSPþPSHPSSNP (¼ I)13. PSSC¼K (¼ CSHPSSNP)14. PSSG¼L (¼ GSHPSSNP)15. NPxSHPSSNP¼I - K - L, deduced from equalities (11), (13), (14)
Algebraic preconditions
For the validity of the above experimental fraction values of theTRS components, and of those derived as algebraic functions ofexperimentally determined fraction values, the following not strictinequalities should hold true:
NPSHZGSHþCSHNPSSNPZGSSG2NPSSNPZCSSNPþGSSNPþGSSGPSSNPZPSSGþPSSC
Statistical treatment of raw data
TRS parameters were calculated in 20 plasma samples (10/10males/females, covering a 50-60 year age span) as mean
Table 11
Reagents (ml) Reagent blank(RB)
Sample blank(SB)
Sample(S)
3/4-2DT-untr-AC-ppsol-TBP-pfsup
– 2000 2000
RB-formamide-TBP-DT 2000 – –
100% Acetic acid 2000 2000 2000100 mM NHD 2000 – 2000Acetic acid-HCl solution – 2000 –
Table 12
Reagents (μl) Reagent blank(RB)
Sample blank(SB)
Sample(S)
1/4-2DT-GPx-AC-ppsol-TCEP-DE-pfsup
– 200 200
RB-urea-TCEP-DT-DE 200 – –
0.5 M phosphate buffer, pH 8.0 130 130 1301 mg ml�1 OPT 20 – 20100% MetOH – 20 –
K. Grintzalis et al. / Free Radical Biology and Medicine 74 (2014) 85–98 95

values7SE of the mean. Parameters GSH, NPSSNP, CSSNP, GSSNP,PSSP, NPxSHNPSSNP, NPxSHNPSH, and NPxSHPSSNP are derived asmean values determined by the algebraic functions of the meanvalues of the involved fractions (see section “Estimation of the TRSparameters”). These functions were used in conjunction with theindividual SE of the involved fractions to estimate the SE of each ofthese parameters by the following general SE function adaptedfrom elsewhere [50]:
SETRS¼7 √{[(∂TRS/∂A)SEA]2þ[(∂TRS/∂B)SEB]2þ[(∂TRS/∂C)SEC]2
þ[(∂TRS/∂D)SED]2þ…}The SE equations of these TRS parameters are the following (for
their Excel-adapted automated estimation see SupplementaryMaterial):
SENPSSNP¼7 √(SEA2þSEB2)SECSSNP¼7 √(SED2þSEC2)SEGSSNP¼7 √(SEG2þSEF2)SEGSH¼7 √(SEF2þSEE2)SENPxSHNPSH
¼7 √(SEA2þSEC2þSEF2þSEE2
SENPxSHNPSSNP¼7 √(SEB2þSEC2þSEA2þSED2þ SEG2þSEE2þSEF2
SEPSSP¼7 √(SEJ2þSEI2þSEH2)SENPxSHPSSNP
¼7 √(SEI2þSEK2þSEL2
For determining the minimum statistical variation of the TRSparameters quantified by the present protocol, human plasmasamples were analyzed the same day of blood collection. The TRSparameters of each plasma sample were derived from the analysisof at least three successive sample dilutions, and their mean valueand SE were calculated. The TRS parameters were also statisticallyanalyzed for precision, assessed during a single analytical run(within-run, within-day precision or repeatability), and with time(between-run, between day repeatability, also named intermedi-ate precision). For calculating the minimum statistical variation ofTRS parameters quantified by the present protocol, at least threesuccessive dilutions of human plasma samples were analyzed thesame day of blood collection, and their mean value was calculated.The within-day % coefficient variation is calculated as StandardDeviation (SD)x100/mean, and the variance of intermediate pre-cision (σ2
total) is defined as the sum of between day variance(σ2
between) that is associated with the day-to-day variation, andthe variance of repeatability (σ2
within). The within-day coefficientvariation of the TRS protocol for all its parameters is 44% and thebetween day repeatability is 45.5%.
Protocol timing and planning
Timing: The timing listed in the following subheadings of theprotocol procedure represents the minimum time required forprocessing 10 samples as follows: Sample treatment (steps 1–4):total 1 h. Subprotocol for nonprotein thiols (steps 5–18): treatmentof fractions A to G, total 4 h; A, 30 min; B, 40 min; C, 30 min; D,50 min; E, 30 min; F, 30 min; G, 70 min. Subprotocol for proteinthiols (steps 19–31): treatment of fractions H to L, total 5 h; H,30 min; I, 45 min; J, 45 min; K, 90 min; L, 120 min.
Planning: Because of the multiparametric analysis offered bythe protocol, handling of many samples (e.g., 10) in the same daymay be difficult. In such case, the protocol can be executed with allrequired reagents prepared for treating all samples up to step 4, bymaking the pose points suggested in protocol steps 3 and 4.However, it is recommended to extend the sample treatment upto step 12, for the determination of CSH and CSSNP. Nonetheless,P-thiols in the AC-washed protein pellets from step 4 are normallystable up to 1 week. Then, the Subprotocol for protein thiols can beperformed next. Here the experimenter could start with the urea-solubilized protein pellet from step 4b, e.g., for PSSG determina-tion; P-thiols in the formamide-solubilized pellet from step 4a canbe analyzed next.
Calculations and expected results
This protocol assesses quantitatively 15 parameters of TRS inorganisms by three different assays applied on sample fractionsA to L. The first assay uses the DTP reagent to quantify NPSH,NPxSHNPSH, and PSH, as well as their disulfides (NPSSNP,NPxSHNPSSNP, PSSNP, NPxSHPSSNP, PSSP); the second assay specifi-cally measures CSH, CSSNP, and PSSC by the NHD reagent; thethird assay determines GSH, GSSG, GSSNP, and PSSG by the OPTreagent. The sensitivity of the protocol for NPSH, PSH, NPSSNP,PSSP, PSSNP, CSH, CSSNP, PSSC, NPxSHNPSSNP, and NPxSHNPSH is1 nmol -SH/CSH, for GSSNP 0.2 nmol, for GSH and GSSG 0.4 nmol,and for PSSG 0.6 nmol. When spiking the 1/5- and 4/5-bloodplasma fractions, in steps 1(a) and 1(b), respectively (or in thesame fractions of any nonfluid sample before homogenization),with known amounts of GSH, GSSG, and CSH, the achievedrecoveries were near 90% (data not shown).
The protocol is designed to be modular and adjustable to theexperimenter’s research priorities on some or all TRS parameters.It may be performed separately for nonprotein and protein thiols(fractions A to G and fractions H to L, respectively), or it can befocused on individual TRS parameters (Table 13).
Given the existence of lipoic acid and other lipophilic thiols invarious experimental samples, these lipophilic thiols are notcounted by the protocol for the following reasons: The protocolseparates the sample homogenate in an aqueous and a proteinfraction (for determining the nonprotein and protein thiols,respectively), by a protein precipitation step that includes homo-genate pretreatment with deoxycholate (DOC) followed by TCAprecipitation. Then, there is a diethyl ether-wash step (3) for theprotein-free supernatant, and a cold acetone wash step (4) for theprotein pellet. Thus, lipoic acid or any other lipophilic thiols areremoved from the supernatant by diethyl ether (for lipoic acidsolubility in diethyl ether see [66]), while any remnants oflipophilic residues as well the TCA-precipitated DOC and anyTCA remnants in the pellet are also removed by the acetone wash.
Another factor to consider is the applicability of the protocol inorganisms such as plants, fungi, and yeast, which can synthesizephytochelatins for heavy metal detoxification [67]. These areglutathione (GSH) related peptides, which, because of their rela-tively small molecular weight (MW), could be measured by theprotocol as GSH if present in the aqueous fraction of the sample.Nonetheless, phytochelatins are also isolated by protein precipita-tion in 6:1 vol cold ethanol (e.g., from the basidiomycete fungusBoletus edulis) [68]. However, the present protocol employs a far
Table 13Modularity of TRS protocol.
TRS parameter Fractions required
Nonprotein parametersGSH E, FGSSG EGSSNP G, FCSH CCSSNP C, DNPSH ANPSSNP A, BNPxSHNPSH A, C, F, ENPxSHNPSSNP A, B, C, D, E, F, GProtein parametersPSSG LPSSC KPSH HPSSNP IPSSP H, I, JNPxSHPSSNP I, J, K
K. Grintzalis et al. / Free Radical Biology and Medicine 74 (2014) 85–9896

more effective protein precipitation method (a mixture of deox-ycholate and TCA), which achieves Z90% recovery of smallpeptides such as ribonuclease A, even in the presence of 1 Mguanidine-HCl [58]. Moreover, the protocol uses a diethyl etherstep to wash the aqueous fraction of the sample, which renderspeptides even more insoluble. Although we cannot exclude thepossibility of very small MW peptides to escape precipitation, weexpect the majority of phytochelatin peptides to end up in theprotein pellet fraction, where they will be quantified as PSH orPSSP by the DTP reagent of the protocol.
The protocol was applied to human plasma, results werecompared with data from alternative methodologies reported inthe literature, and a good correspondence was attained (Table 14).However, no comparison data were available for the TRS para-meters GSSNP, CSSNP, NPSH, NPSSNP, PSSP, PSSNP, NPxSHNPSSNP,NPxSHPSSNP, and NPxSHNPSH. This is indicative of the partialevaluation of TRS by the current methodologies. We limited ourmeasurements to normal samples to display the applicability ofthe present assay and also set approximate normal levels mainlyfor the new TRS parameters introduced by the protocol. Beforetesting pathological samples, extensive studies are needed tooptimize normal levels by taking also into account sample varia-bility factors such as degree of hemolysis. However, care was takenin this study not to use hemolyzed samples, which are defined ashaving free hemoglobin in serum or plasma below 50 mg/dl [69].
Caveats
A near zero net absorbance or fluorescence value of a certainthiol parameter suggests that its concentration in the sample isbelow the assay sensitivity limit. Increasing the initial amount ofsample, and/or keeping its initial homogenate at a minimumvolume could address this. If the absorbance or fluorescence valueof a thiol parameter falls out of the upper limit of the linear rangeof the corresponding thiol parameter standard curve, this meansthat its concentration in the sample is very high. In this case, thesample must be diluted appropriately. High statistical variation ina thiol parameter value among replicate samples suggests highvariation in its concentration in the experimental organism.Increasing the number of different sample replicates can solvethis problem. If the algebraic preconditions defined for thisprotocol (in the subsection “Estimation of the TRS parameters”)are not met, this is likely due to experimental errors in itsexecution. The suggested solution to this problem is repetition ofthe TRS parameter measurements with new and fresh samples. Forexample, NPSH must be greater or equal to the sum of GSH andCSH because, if detected, their values are included in the value ofNPSH. Similarly, NPSH should be greater or equal to GSH or CSHalone, as long as CSH or GSH are near zero, respectively.
Conflict of Interest
The authors declare that they have no patents or financialinterests in the method or instruments.
Acknowledgments
This work was financially supported by the Greek Ministry ofEducation. We thank Prof. Dr. Helmut Sies (Institute for Biochem-istry and Molecular Biology I, Heinrich-Heine-University Düssel-dorf) for his very important and thoughtful comments.
Appendix A. Supplementary Information
Supplementary data associated with this article can be found inthe online version at http://dx.doi.org/10.1016/j.freeradbiomed.2014.06.024.
References
[1] Sies, H. Glutathione and its role in cellular functions. Free Radic. Biol. Med.27:916–921; 1999.
[2] Halliwell, B.; Gutteridge, C. M. J. Free radicals in biology and medicine. Oxford:Oxford University Press; 1999.
[3] Dickinson, D. A.; Forman, H. J. Cellular glutathione and thiols metabolism.Biochem. Pharmacol. 64:1019–1026; 2002.
[4] Wang, W.; Ballatori, N. Endogenous glutathione conjugates: occurrence andbiological functions. Pharmacol. Rev. 50:335–356; 1998.
[5] Giles, I. G.; Tasker, M. K.; Jacob, C. Hypothesis: the role of reactive sulfurspecies in oxidative stress. Free Radic. Biol. Med. 31:1279–1283; 2001.
[6] Li, J.; Huang, L. F.; Huang, K. -P. Glutathiolation of proteins by glutathionedisulfide S-oxide derived from S-nitrosoglutathione. J. Biol. Chem. 276:3098–-3105; 2001.
[7] Crane, D.; Häussinger, D.; Sies, H. Rise of coenzyme A-glutathione mixeddisulfide during hydroperoxide metabolism in perfused rat liver. Eur. J.Biochem. 127:575–578; 1982.
[8] Akerboom, M. P. T.; Sies, H. Assay of glutathione, glutathione disulfide andglutathione mixed disulfides in biological samples. In: Colowick, S. P., Kaplan,O. N., editors. Methods in enzymology. New York: Academic Press; 1981.p. 373–382.
[9] Corti, A.; Paolicchi, A.; Franzini, M.; Dominici, S.; Casini, A. F.; Pompella, A.The S-thiolating activity of membrane gamma-glutamyltransferase: formationof cysteinyl-glycine mixed disulfides with cellular proteins and in the cellmicroenvironment. Antioxid. Redox Signal. 7:911–918; 2005.
Table 14TRS parameters of human plasma.
Parameters (nmol mg-1) (μΜ) From indicativebibliography (mM)
Nonprotein parametersGSH 0.02370.004 3.170.26 3.570.3 [43]
2–5 [70]1.4–6 [44]1.570.5 [38]2–3 [46,55]3.73 [39]2 (3% RSD) [37]
GSSG 0.01370.002 1.870.21 1.2370.44 [46]2.2 (5.1% RSD) [37]
GSSNP 0.04170.006 4.370.62 Not available (NA)CSH 0.0870.01 11.170.9 11.770.95 [43]
8.3–10.7 [70]5.9–12.6 [44]9.9671.29 [46]
CSSNP 1.5570.12 215714 NANPSH 0.1970.018 25.171.5 NANPSSNP 1.9170.15 255734 NANPxSHNPSH 0.0970.02 10.971.8 NANPxSHNPSSNP 2.4370.35 316748 NAProtein parametersPSH 5.170.6 680775 400–600 [70]PSSP 98711 1280071500 NAPSSG 0.01970.002 2.670.28 2.570.76 [43]
0.7–1.9 [70]5.3–11 [44]2.3171.18 [46]
PSSC 1.1570.22 150719 142.178.4 [43]145–176 [70]178–227 [44]156.82728.44 [46]
PSSNP 5.9370.65 795788 NANPxSHPSSNP 4.7670.69 642790 NA
Note: TRS parameters were calculated in plasma samples from 10 males and 10females. All values are mean7SE after checking for equality of error variancesbetween values of males and females (Levene's test, SPSS Inc.) with two-wayANOVA analysis of variance (to identify significant differences between values), andwith the parametric post hoc multiple comparison test (Bonferroni test, P o0.05).There were no statistical differences in the TRS parameters between genders.
K. Grintzalis et al. / Free Radical Biology and Medicine 74 (2014) 85–98 97

[10] Gilbert, F. H. Thiol/disulfide exchange equilibria and disulfide bond stability.In: Packer, L., editor. Methods in enzymology. New York: Academic Press; 1995.p. 8–28.
[11] Ratbbun, B. W. Glutathione metabolism in the mammalian ocular lense. In:Vina, J., editor. Glutathione: Metabolism and physiological functions. Boca Raton:CRC Press; 1990. p. 193–206.
[12] Miquel, J.; Weber, H. Aging and increased oxidation of the sulfur pool. In: Vina,J., editor. Glutathione: Metabolism and physiological functions. Boca Raton: CRCPress; 1990. p. 187–192.
[13] Levitt, J. Responses of plants to environmental stresses. New York: AcademicPress; 1972.
[14] Trivedi, M. V.; Laurence, J. S.; Siahaan, T. J. The role of thiols and disulfides onprotein stability. Curr. Protein Pept. Sci. 10:614–625; 2009.
[15] Gupta, V.; Carroll, K. S. Sulfenic acid chemistry, detection and cellular lifetime.Biochim. Biophys. Acta 1840:847–875; 2014.
[16] Alvarez, B.; Carballal, S.; Turell, L.; Radi, R. Formation and reactions of sulfenicacid in human serum albumin. Methods Enzymol. 473:117–136; 2010.
[17] Requejo, R.; Hurd, T. R.; Costa, N. J.; Murphy, M. P. Cysteine residues exposedon protein surfaces are the dominant intramitochondrial thiol and may protectagainst oxidative damage. FEBS J. 277:1465–1480; 2010.
[18] Carballal, S.; Alvarez, B.; Turell, L.; Botti, H.; A., F. B.; Radi, R. Sulfenic acid inhuman serum albumin. Amino Acids 32:543–551; 2007.
[19] Eaton, P. Protein thiol oxidation in health and disease: techniques formeasuring disulfides and related modifications in complex protein mixtures.Free Radic. Biol. Med. 40:1889–1899; 2006.
[20] van der Vliet, A.; Cross, E. C.; Halliwell, B.; O'Neill, A. C. Plasma protein sulfhydryloxidation: effect of low molecular weight thiols. In: Packer, L., editor. Methods inenzymology. New York: Academic Press; 1995. p. 448–455.
[21] Cotgreave, A. I.; Weis, M.; Atzori, L.; Moldéus, P. Glutathione and proteinfunction. In: Vina, J., editor. Glutathione: metabolism and physiological func-tions. Boca Raton: CRC Press; 1990. p. 155–175.
[22] Brigelius-Flohé, R. Tissue-specific functions of individual glutathione perox-idases. Free Radic. Biol. Med. 27:951–965; 1999.
[23] Finkel, T. Signal transduction by reactive oxygen species. J. Cell Biol. 194:7–15;2011.
[24] Ewing, F. J.; Janero, R. D. Specific S-nitrosothiol (thionitrite) quantification assolution nitrite after vanadium(III) reduction and ozone-chemiluminescentdetection. Free Radic. Biol. Med. 25:621–628; 1998.
[25] Täger, M.; Piecyk, A.; Köhnlein, T.; Thiel, U.; Ansorge, S.; Welte, T. Evidence of adefective thiol status of alveolar macrophages from COPD patients andsmokers. Free Radic. Biol. Med. 29:1160–1165; 2000.
[26] David, L. L.; Shearer, R. T. State of sulfhydryl in selenite cataract. Toxicol. Appl.Pharmacol. 74:109–115; 1984.
[27] Tietze, F. Enzymic method for quantitative determination of nanogramamounts of total and oxidized glutathione: applications to mammalian bloodand other tissues. Anal. Biochem. 27:502–522; 1969.
[28] Ellman, L. G. Tissue sulfhydryl groups. Arch. Biochem. Biophys. 82:70–77; 1959.[29] Redegeld, F. A. M.; Koster, A. S.; van Bennekom, W. P. Determination of tissue
glutathione. In: Vina, J., editor. Glutathione: metabolism and physiologicalfunctions. Boca Raton: CRC Press; 1990. p. 11–20.
[30] Giustarini, D.; Dalle-Donne, I.; Milzani, A.; Fanti, P.; Rossi, R. Analysis of GSHand GSSG after derivatization with N-ethylmaleimide. Nat. Protoc. 8:1660–-1669; 2013.
[31] Rahman, I.; Kode, A.; Biswas, S. K. Assay for quantitative determination ofglutathione and glutathione disulfide levels using enzymatic recyclingmethod. Nat. Protoc. 1:3159–3165; 2006.
[32] Hissin, P. J.; Hilf, R. A fluorometric method for determination of oxidized andreduced glutathione in tissues. Anal. Biochem. 74:214–226; 1976.
[33] Puri, R. N.; Roskoski Jr R. Reaction of low molecular weight aminothiols witho-phthalaldehyde. Anal. Biochem. 173:26–32; 1988.
[34] Lenton, K. J.; Therriault, H.; Wagner, J. R. Analysis of glutathione andglutathione disulfide in whole cells and mitochondria by postcolumn deriva-tization high-performance liquid chromatography with ortho-phthalaldehyde.Anal. Biochem. 274:125–130; 1999.
[35] Harrap, R. K.; Jackson, C. R.; Riches, G. P.; Smith, A. C.; Hill, T. B. The occurrenceof protein-bound mixed disulfides in rat tissues. Biochim. Biophys. Acta (Prot.Struct.) 310:104–110; 1973.
[36] Ball, R. C. Estimation and identification of thiols in rat spleen after cysteine orglutathione treatment: relevance to protection against nitrogen mustards.Biochem. Pharmacol. 15:809–816; 1966.
[37] Salazar, J. F.; Schorr, H.; Herrmann, W.; Herbeth, B.; Siest, G.; Leroy, P.Measurement of thiols in human plasma using liquid chromatography withprecolumn derivatization and fluorescence detection. J. Chromatogr. Sci.37:469–476; 1999.
[38] Ferin, R.; Pavao, M. L.; Baptista, J. Methodology for a rapid and simultaneousdetermination of total cysteine, homocysteine, cysteinylglycine and glu-tathione in plasma by isocratic RP-HPLC. J. Chromatogr. B Analyt. Technol.Biomed. Life Sci. 911:15–20; 2012.
[39] Kusmierek, K.; Bald, E. Reversed-phase liquid chromatography method for thedetermination of total plasma thiols after derivatization with 1-benzyl-2-chloropyridinium bromide. Biomed. Chromatogr. 23:770–775; 2009.
[40] Riener, C. K.; Kada, G.; Gruber, H. J. Quick measurement of protein sulfhydrylswith Ellman's reagent and with 4,40-dithiodipyridine. Anal. Bioanal. Chem.373:266–276; 2002.
[41] Giustarini, D.; Dalle-Donne, I.; Lorenzini, S.; Selvi, E.; Colombo, G.; Milzani, A.;Fanti, P.; Rossi, R. Protein thiolation index (PTI) as a biomarker of oxidativestress. Free Radic. Biol. Med. 53:907–915; 2012.
[42] Hoff, S.; Larsen, F. H.; Andersen, M. L.; Lund, M. N. Quantification of proteinthiols using ThioGlo 1 fluorescent derivatives and HPLC separation. Analyst138:2096–2103; 2013.
[43] Carru, C.; Deiana, L.; Sotgia, S.; Pes, G. M.; Zinellu, A. Plasma thiols redox statusby laser-induced fluorescence capillary electrophoresis. Electrophoresis25:882–889; 2004.
[44] Bald, E.; Chwatko, G.; Glowacki, R.; Kusmierek, K. Analysis of plasma thiols byhigh-performance liquid chromatography with ultraviolet detection. J. Chro-matogr. A 1032:109–115; 2004.
[45] Hansen, R. E.; Ostergaard, H.; Norgaard, P.; Winther, J. R. Quantification ofprotein thiols and dithiols in the picomolar range using sodium borohydrideand 4,40-dithiodipyridine. Anal. Biochem. 363:77–82; 2007.
[46] Rossi, R.; Giustarini, D.; Milzani, A.; Dalle-Donne, I. Cysteinylation andhomocysteinylation of plasma protein thiols during ageing of healthy humanbeings. J. Cell. Mol. Med. 13:3131–3140; 2009.
[47] Chin, C. C. Q.; Wold, F. The use of tributylphosphine and 4-(aminosulfonyl)-7-fluoro-2,1,3-benzoxadiazole in the study of protein sulfhydryls and disulfides.Anal. Biochem. 214:128–134; 1993.
[48] Kirley, T. L. Reduction and fluorescent labelling of cyst(e)ine-containing proteins forsubsequent structural analyses. Anal. Biochem. 180:231–236; 1989.
[49] Krijt, J.; Vacková, M.; Kozich, V. Measurement of homocysteine and otheraminothiols in plasma: advantages of using tris(2-carboxyethyl)phosphine asreductant compared with tri-n-butylphosphine. Clin. Chem. 47:1821–1828; 2001.
[50] Patsoukis, N.; Georgiou, C. D. Determination of the thiol redox state oforganisms: new oxidative stress indicators. Anal. Bioanal. Chem. 378:1783–-1792; 2004.
[51] Patsoukis, N.; Georgiou, C. D. Fluorometric determination of thiol redox state.Anal. Bioanal. Chem. 383:923–929; 2005.
[52] Owusu-Apenten, R. Colorimetric analysis of protein sulfhydyl groups in milk:applications and processing effects. Crit. Rev. Food Sci. Nutr. 45:1–23; 2005.
[53] Flohé, L. The fairytale of the GSSG/GSH redox potential. Biochim. Biophys. Acta1830:3139–3142; 2013.
[54] Morgan, B.; Ezerina, D.; Amoako, T. N.; Riemer, J.; Seedorf, M.; Dick, T. P.Multiple glutathione disulfide removal pathways mediate cytosolic redoxhomeostasis. Nat. Chem. Biol. 9:119–125; 2012.
[55] Rossi, R.; Milzani, A.; Dalle-Donne, I.; Giustarini, D.; Lusini, L.; Colombo, R.;Di Simplicio, P. Blood glutathione disulfide: in vivo factor or in vitro artifact?Clin. Chem. 48:742–753; 2002.
[56] Saez, G.; Thornalley, J. P.; Hill, O. A. H.; Hems, R.; Bannister, V. J. Theproduction of free radicals during the autoxidation of cysteine and their effecton isolated rat hepatocytes. Biochim. Biophys. Acta 719:24–31; 1982.
[57] Munday, R.; Munday, C. M.; Winterbourn, C. C. Inhibition of copper-catalyzedcysteine oxidation by nanomolar concentrations of iron salts. Free Radic. Biol.Med. 36:757–764; 2004.
[58] Arnold, U.; Ulbrich-Hofmann, R. Quantitative protein precipitation fromguanidine hydrochloride-containing solutions by sodium deoxycholate/tri-chloroacetic acid. Anal. Biochem. 271:197–199; 1999.
[59] Bensadoun, A.; Weinstein, D. Assay of proteins in the presence of interferingmaterials. Anal. Biochem. 70:241–250; 1976.
[60] Cohn, V. H.; Lyle, J. A fluorometric assay for glutathione. Anal. Biochem.14:434–440; 1966.
[61] Donovan, J. W.; White, T. M. Alkaline hydrolysis of the disulfide bonds ofovomucoid and of low molecular weight aliphatic and aromatic disulfides.Biochemistry 10:32–38; 1971.
[62] Ogwu, V.; Cohen, G. A simple colorimetric method for the simultaneousdetermination of N-acetylcysteine and cysteine. Free Radic. Biol. Med. 25:362–-364; 1998.
[63] Georgiou, C. D.; Grintzalis, K.; Zervoudakis, G.; Papapostolou, I. Mechanism ofCoomassie brilliant blue G-250 binding to proteins: a hydrophobic assay fornanogram quantities of proteins. Anal. Bioanal. Chem. 391:391–403; 2008.
[64] Obrador, E.; Carretero, J.; Ortega, A.; Medina, I.; Rodilla, V.; Pellicer, J. A.; Estrela, J.M. Gamma-glutamyl transpeptidase overexpression increases metastatic growth ofB16 melanoma cells in the mouse liver. Hepatology 35:74–81; 2002.
[65] Dubler, R. E.; Anderson, B. M. Simultaneous inactivation of the catalyticactivities of yeast glutathione reductase by N-alkylmaleimides. Biochim.Biophys. Acta 14:70–85; 1981.
[66] Witt, W.; Rüstow, B. Determination of lipoic acid by precolumn derivatizationwith monobromobimane and reversed-phase high-performance liquid chro-matography. J. Chromatogr. B Biomed. Sci. Appl 705:127–131; 1998.
[67] Cobbett, C.; Goldsbrough, P. Phytochelatins and metallothioneins: roles inheavy metal detoxification and homeostasis. Annu. Rev. Plant. Biol. 53:159–-182; 2002.
[68] Collin-Hansen, C.; Andersen, R. A.; Steinnes, E. Isolation and N-terminalsequencing of a novel cadmium-binding protein from Boletus edulis. J. Phys.IV 107:311–314; 2003.
[69] Zhao, J.; Kan, Q.; Wen, J.; Li, Y.; Sheng, Y.; Yang, L.; Wu, J.; Zhang, S. Hemolysisof blood samples has no significant impact on the results of pharmacokineticdata. J. Bioequiv. Availab 4:82–85; 2012.
[70] Turell, L.; Radi, R.; Alvarez, B. The thiol pool in human plasma: the centralcontribution of albumin to redox processes. Free Radic. Biol. Med. 7:244–253; 2013.
[71] Friedman, M. Applications of the ninhydrin reaction for analysis of aminoacids, peptides, and proteins to agricultural and biomedical sciences. J. Agric.Food Chem. 52:385–406; 2004.
K. Grintzalis et al. / Free Radical Biology and Medicine 74 (2014) 85–9898




















