
Protein disulfide isomerase, but not binding protein, overexpression enhances secretion of a...
11
Protein Disulfide Isomerase, But Not Binding Protein, Overexpression Enhances Secretion of a Non-Disulfide-Bonded Protein in Yeast Jason D. Smith, Benjamin C. Tang, Anne Skaja Robinson Department of Chemical Engineering, University of Delaware, 259 Colburn Laboratory, Newark, Delaware 19716; telephone: 302-831-0557; fax: 302- 831-6262; e-mail: robinson @che.udel.edu Received 5 May 2003; accepted 21 August 2003 Published online 31 December 2003 in Wiley InterScience (www.interscience.wiley.com). DOI: 10.1002/bit.10853 Abstract: In eukaryotes, secretory proteins are folded and assembled in the endoplasmic reticulum (ER). Many hete- rologous proteins are retained in the ER due to suboptimal folding conditions. We previously reported that heterolo- gous secretion of Pyrococcus furiosus h-glucosidase in Saccharomyces cerevisiae resulted in the accumulation of a large fraction of inactive h-glucosidase in the ER. In this work, we determine the effect of introducing additional genes of ER-resident yeast proteins, Kar2p (binding protein [BiP]) and protein disulfide isomerase (PDI), on relieving this bottleneck. Single-copy expression of BiP and PDI worked synergistically to improve secretion by f 60%. In an effort to optimize BiP and PDI interactions, we created a library of h-glucosidase expression strains that incorporated four combinations of constitutively or inducibly-expressed BiP and PDI genes integrated to random gene copynumbers in the yeast chromosome. Approximately 15% of the trans- formants screened had secretion level improvements higher than that seen with single BiP/PDI gene overexpression, and the highest secreting strain had threefold higher h-glucosi- dase levels than the control. Nineteen of the improved strains were re-examined for h-glucosidase secretion as well as BiP and PDI levels. Within the improved trans- formants BiP and PDI levels ranged sevenfold and tenfold over the control, respectively. Interestingly, increasing BiP levels decreased h-glucosidase secretion, whereas increas- ing PDI levels increased h-glucosidase secretion. The action of PDI was unexpected because h-glucosidase is not a disulfide-bonded protein. We suggest that PDI may be acting in a chaperone-like capacity or possibly creating mixed disulfides with the h-glucosidase’s lone cysteine residue during the folding and assembly process. B 2004 Wiley Periodicals, Inc. Keywords: Archaea; binding protein (BiP); protein disulfide isomerase (PDI); Pyrococcus furiosus; Saccharomyces cerevisiae; optimization; secretory pathway; endoplasmic reticulum; h-glucosidase; chaperone; Hsp70 INTRODUCTION Overexpression of a foreign protein in any recombinant host can lead to misfolding of the foreign protein due to non-native, suboptimal conditions and/or overloading of the host’s machinery. Gene copy optimization is a typical strategy used to optimize heterologous protein expression. However, in the case of secreted proteins, this strategy can lead to saturation or overloading of the secretory pathway, typically in the endoplasmic reticulum (ER) (Parekh et al., 1995; Parekh and Wittrup, 1997; Robinson et al., 1994; Shusta et al., 1998). By varying gene copynumber, Parekh and Wittrup (1997) reached 180 mg/L of secreted BPTI in Saccharomyces cerevisiae. They found that BPTI accumulated in the ER in an unfolded form, which they believed was due to dilution of the necessary folding machinery. Robinson and Wittrup (1995) showed that overexpression of heterologous proteins in S. cerevisiae led to decreased soluble levels of ER- resident immunoglobulin heavy-chain binding protein (BiP) and protein disulfide isomerase (PDI). Consequently, it has been shown that reduction of BiP levels resulted in decreased heterologous protein secretion (Robinson et al., 1996). BiP and PDI, among others, participate directly in the active folding of proteins in the ER and only properly folded proteins are allowed to exit the ER. Therefore, manipulation of the ER folding environment may produce further im- provements in S. cerevisiae’s secretory productivity. BiP and PDI are commonly studied, abundant ER-resident proteins. BiP, also known as Kar2p in yeast, is an Hsp70- class heat shock protein (Haas and Wabl, 1983) that functions in protein folding as well as being an important part of the cell’s translocation machinery (Matlack et al., 1999). BiP aids in protein folding through the binding of hydrophobic amino acid stretches in the immature protein. Binding of these exposed stretches stabilizes the protein and prevents it from aggregating with other unfolded/misfolded proteins. Through ATP hydrolysis, BiP releases the bound polypeptide allowing for continuous cycles of binding and releasing thus giving the polypeptide a chance to bury its B 2004 Wiley Periodicals, Inc. Correspondence to: A. Robinson Contract grant sponsors: DuPont (Young Investigator Award); ACS Petroleum Research Foundation Contract grant number: 35916-G4
Transcript of Protein disulfide isomerase, but not binding protein, overexpression enhances secretion of a...

Protein Disulfide Isomerase, But Not BindingProtein, Overexpression Enhances Secretionof a Non-Disulfide-Bonded Protein in Yeast
Jason D. Smith, Benjamin C. Tang, Anne Skaja Robinson
Department of Chemical Engineering, University of Delaware, 259 ColburnLaboratory, Newark, Delaware 19716; telephone: 302-831-0557; fax: 302-831-6262; e-mail: [email protected]
Received 5 May 2003; accepted 21 August 2003
Published online 31 December 2003 in Wiley InterScience (www.interscience.wiley.com). DOI: 10.1002/bit.10853
Abstract: In eukaryotes, secretory proteins are folded andassembled in the endoplasmic reticulum (ER). Many hete-rologous proteins are retained in the ER due to suboptimalfolding conditions. We previously reported that heterolo-gous secretion of Pyrococcus furiosus h-glucosidase inSaccharomyces cerevisiae resulted in the accumulation of alarge fraction of inactive h-glucosidase in the ER. In thiswork, we determine the effect of introducing additionalgenes of ER-resident yeast proteins, Kar2p (binding protein[BiP]) and protein disulfide isomerase (PDI), on relieving thisbottleneck. Single-copy expression of BiP and PDI workedsynergistically to improve secretion by f60%. In an effortto optimize BiP and PDI interactions, we created a library ofh-glucosidase expression strains that incorporated fourcombinations of constitutively or inducibly-expressed BiPand PDI genes integrated to random gene copynumbers inthe yeast chromosome. Approximately 15% of the trans-formants screened had secretion level improvements higherthan that seenwith single BiP/PDI gene overexpression, andthe highest secreting strain had threefold higher h-glucosi-dase levels than the control. Nineteen of the improvedstrains were re-examined for h-glucosidase secretion aswell as BiP and PDI levels. Within the improved trans-formants BiP and PDI levels ranged sevenfold and tenfoldover the control, respectively. Interestingly, increasing BiPlevels decreased h-glucosidase secretion, whereas increas-ing PDI levels increased h-glucosidase secretion. The actionof PDI was unexpected because h-glucosidase is not adisulfide-bonded protein. We suggest that PDI may beacting in a chaperone-like capacity or possibly creatingmixed disulfides with the h-glucosidase’s lone cysteineresidue during the folding and assembly process. B 2004Wiley Periodicals, Inc.
Keywords: Archaea; binding protein (BiP); protein disulfideisomerase (PDI); Pyrococcus furiosus; Saccharomycescerevisiae; optimization; secretory pathway; endoplasmicreticulum; h-glucosidase; chaperone; Hsp70
INTRODUCTION
Overexpression of a foreign protein in any recombinant
host can lead to misfolding of the foreign protein due to
non-native, suboptimal conditions and/or overloading of
the host’s machinery. Gene copy optimization is a typical
strategy used to optimize heterologous protein expression.
However, in the case of secreted proteins, this strategy can
lead to saturation or overloading of the secretory pathway,
typically in the endoplasmic reticulum (ER) (Parekh et al.,
1995; Parekh and Wittrup, 1997; Robinson et al., 1994;
Shusta et al., 1998).
By varying gene copynumber, Parekh and Wittrup (1997)
reached 180 mg/L of secreted BPTI in Saccharomyces
cerevisiae. They found that BPTI accumulated in the ER in
an unfolded form, which they believed was due to dilution of
the necessary folding machinery. Robinson and Wittrup
(1995) showed that overexpression of heterologous proteins
in S. cerevisiae led to decreased soluble levels of ER-
resident immunoglobulin heavy-chain binding protein (BiP)
and protein disulfide isomerase (PDI). Consequently, it has
been shown that reduction of BiP levels resulted in decreased
heterologous protein secretion (Robinson et al., 1996). BiP
and PDI, among others, participate directly in the active
folding of proteins in the ER and only properly folded
proteins are allowed to exit the ER. Therefore, manipulation
of the ER folding environment may produce further im-
provements in S. cerevisiae’s secretory productivity.
BiP and PDI are commonly studied, abundant ER-resident
proteins. BiP, also known as Kar2p in yeast, is an Hsp70-
class heat shock protein (Haas and Wabl, 1983) that
functions in protein folding as well as being an important
part of the cell’s translocation machinery (Matlack et al.,
1999). BiP aids in protein folding through the binding of
hydrophobic amino acid stretches in the immature protein.
Binding of these exposed stretches stabilizes the protein and
prevents it from aggregating with other unfolded/misfolded
proteins. Through ATP hydrolysis, BiP releases the bound
polypeptide allowing for continuous cycles of binding and
releasing thus giving the polypeptide a chance to bury its
B 2004 Wiley Periodicals, Inc.
Correspondence to: A. Robinson
Contract grant sponsors: DuPont (Young Investigator Award); ACS
Petroleum Research Foundation
Contract grant number: 35916-G4

exposed hydrophobic residues in the process of proper
folding (Rowling and Freedman, 1993). BiP also plays a role
in proofreading, through ER-associated degradation (Brod-
sky et al., 1999).
PDI, an abundant ER foldase (Rowling and Freedman,
1993), is an essential yeast protein that catalyzes the oxida-
tion, reduction, and isomerization of disulfide bonds (Laboi-
ssire et al., 1995). PDI also has a well-demonstrated, albeit
poorly understood, chaperone ability (Dai and Wang, 1997;
Klappa et al., 1998; LaMantia and Lennarz, 1993); that is,
there is no known specific binding site as there is with BiP.
By manipulating chaperone and foldase levels, improve-
ments in yeast/eukaryotic secretion have been found for a
number of proteins. Robinson et al. (1994) reported four- to
tenfold higher secretion yields of two heterologous proteins
in the yeast S. cerevisiae by overexpressing a single inte-
grated copy of PDI. Similarly, Shusta et al. (1998) reported
approximately twofold increases in secretion of a single-
chain antibody fragment by overexpression of either BiP or
PDI in yeast. When both PDI and BiP were co-overex-
pressed, secretion levels increased eightfold. In addition,
secreted levels of the highly disulfide-bonded thaumatin
were increased approximately fivefold upon overexpression
of protein disulfide isomerase in the filamentous fungus
Aspergillus awamori (Moralejo et al., 2001). In an
Escherichia coli cell-free translation system for single-chain
antibody fragments, the addition of PDI increased functional
antibody levels (Ryabova et al., 1997).
We had previously demonstrated heterologous secretion
of the hyperthermophilic, tetrameric Pyrococcus furiosus h-
glucosidase in Saccharomyces cerevisiae to approximately
12 mg/L (Smith and Robinson, 2002). Characterization of
h-glucosidase expression revealed an ER bottleneck, with
experimental evidence indicating misfolding of the mono-
mer. In this study we optimize the folding conditions of the
ER by manipulation of the ER-resident proteins, BiP and
PDI, to improve h-glucosidase secretion levels.
MATERIALS AND METHODS
Cell Strains
The yeast strain BJ5464 (a ura3-52 trp1 leu2D1 hisD200
pep4::HIS3 prb1D1.6R can1 GAL) was obtained from the
Yeast Genetic Stock Center (Berkeley, CA) and served as
the basis for all yeast constructs. Strain JSIY017 was ob-
tained by transformation of BJ5464 with the pITy4-hintegrating plasmid containing the secretion-engineered
Pyrococcus furiosus h-glucosidase gene (Smith and
Robinson, 2002).
Media and Growth Conditions
Yeast cells transformed with integrating plasmids were
typically grown in the rich media YPD and YPG. YPD (2%
peptone; Difco, Carlsbad, CA), with 1% yeast extract
(Difco) and 2% glucose (Fisher, Pittsburgh, PA), was used
as rich growth media and YPG (2% peptone; Difco), with
1% yeast extract (Difco) and 2% galactose (Fisher), was
used as expression media.
Minimal media was used for yeast that had been
transformed with plasmids containing auxotrophic selec-
tion markers. Growth (SD-2� SCAA) or expression (SG-
2� SCAA) minimal media contained 0.67% yeast nitrogen
base (Difco), 2% glucose or galactose, 1.47% citric acid
monohydrate (Sigma), 0.42% sodium citrate (Sigma), and
synthetic amino acid supplement (2� SCAA) (Wittrup and
Benig, 1994). Leucine, tryptophan, and uracil were added
as needed to concentrations of 400 mg/L, 40 mg/L, and
40 mg/L, respectively.
Transformants were typically grown for 48 h in glucose-
based media at 30jC, 275 rpm. Five optical density (OD)-
milliliter equivalents were isolated by centrifugation (2000� g, 5 min) and the supernatant removed. Cell pellets were
resuspended in 5 mL of galactose-based media and ex-
pressed for 24 h for YPG or 48 h for SG-2� SCAA at 30jC,
275 rpm. Supernatant and cell fractions were isolated for
further analysis.
Plasmids
A CEN-ARS vector with a URA3 metabolic marker, pCT37,
was used for galactose-induced expression of yeast PDI
(Tachibana and Stevens, 1992). Similarly, pGalKar2-LEU
is a CEN-ARS vector containing a galactose-inducible yeast
KAR2 gene and a LEU2 metabolic marker (Robinson and
Wittrup, 1995).
Integrating plasmids were constructed using the pITy
integrating plasmid (Parekh et al., 1996). The plasmid
pITy4-h (Smith and Robinson, 2002) was previously used to
achieve optimal expression of h-glucosidase. A 64-bp
polylinker containing the restriction sites (from 5V to 3V)KpnI, AatII, EcoRI, EagI, BamHI, XbaI, and SacI was
purchased from Operon and was ligated into pITy4-h, which
had a KpnI–SacI expression cassette that had been removed
through restriction digestion, resulting in plasmid pITypL.
Sequential EagI and AatII digests were performed on
pGalKar2-LEU to remove the GAL1 promoter-KAR2
expression cassette (including KAR2’s transcription termi-
nator). The expression cassette was ligated into pITypL
digested with AatII and EagI, thus creating plasmid pITy-
GAL-KAR2. In a similar manner, pGAPDH-KAR2 (Rob-
inson et al., 1996) was digested sequentially with EagI and
AatII to remove the GAPDH promoter-KAR2 expression
cassette. Subsequent ligation with pITypL digested with
AatII and EagI created pITyGAPDH-KAR2.
The PDI gene was amplified from pCT37 using
polymerase chain reaction (PCR). The primer 5V-GCTCGAGATGAAGTTTTCTGCTGGTGC-3V was used
to create an XhoI site 5V of the start codon, and the primer
5V-CGGTCTAGATTTTCTCTTTCCCTAAGGCG-3V was
used to create an XbaI site 170 bp downstream of the
PDI stop codon. Both pITyGAL-KAR2 and pITyGAPDH-
SMITH ET AL.: PDI OVEREXPRESSION ENHANCES SECRETION OF YEAST PROTEIN 341

KAR2 were digested with SalI and XbaI (partial) to remove
the KAR2 gene. Subsequent ligation with the PCR-ampli-
fied gene product digested with XhoI and XbaI resulted in
integrating vectors with PDI under control of the GAL1
inducible promoter (pITyGAL-PDI) and the GAPDH
constitutive promoter (pITyGAPDH-PDI).
Integration into the genome was facilitated through
linearization of pITyGAL-KAR2 and pITyGAPDH-KAR2
with BsaBI and pITyGAL-PDI and pITyGAPDH-PDI with
XhoI before transformation into BJ5464. Transformant
colonies were selected on the basis of resistance to G418
(Sigma Co., St. Louis, MO) as previously described (Parekh
et al., 1996). Multiple pools of transformants (f20 to
30 transformant colonies each) from independent trans-
formations were transformed with pRS314-h, a CEN-ARS
pRS314 vector (ATCC, Manassas, VA) containing a TRP1
metabolic marker and a galactose-inducible P. furiosus
h-glucosidase modified for yeast secretion (Smith and
Robinson, 2002). Transformants were selected on the basis
of growth on minimal media lacking tryptophan.
Immunodetection
Cell extracts were typically prepared as described by
Robinson et al. (1996). For Western blots, cell extracts and
culture supernatants were first separated by sodium
dodecylsulfate–polyacrylamide gel electrophoresis (SDS-
PAGE). Protein transfer to nitrocellulose and Western blot
protocol were performed as previously described (Robinson
et al., 1996). Anti-BiP polyclonal primary antibody (a gift
from Jeff Brodsky, University of Pittsburgh) and anti-PDI
polyclonal primary antibody (Stressgen, San Diego, CA)
were used at 1:10,000 and 1:1000 dilutions, respectively.
Goat anti-rabbit secondary antibody conjugated to horse-
radish peroxidase (Amersham Biosciences, Piscataway, NJ)
was used for BiP and PDI detection at a 1:2000 dilution.
Antibody–antigen complexes were visualized with ECL
Plus detection reagent (Amersham) and Western blots were
imaged using the fluorimager (Model 595, Amersham).
Images were quantitated by densitometric analysis using
NIH Image 1.44 software (NIH, Bethesda, MD).
B-Glucosidase Activity Assay
The colorimetric assay used to determine h-glucosidase
activity was modified from a previously described protocol
(Wood and Bhat, 1998). Kinetic assays were performed at
90jC on a spectrophotometer (Model DU600, Beckman,
Fullerton, CA) at an analytical wavelength of 405 nm.
Quartz cuvettes (Starna Cells, Atascadero, CA) containing
0.1 M sodium acetate, pH 5.0 (capped tightly with Teflon
stoppers), were equilibrated to 90jC, after which 30 AL of
20 mM p-nitrophenol-h-D-glucopyranoside (pNPG; Sigma)
and 10 AL of isolated culture supernatant were added (1 mL
total volume). Runs were immediately initiated and
measurements were recorded every 30 s and compared
with a buffer blank. Autocleavage rates of pNPG were
undetectable during the time course of the experiments.
Activities are reported as the maximum rate of pNPG
disappearance normalized by the culture optical density.
Activities measured from cultures derived from air
shakers were typically two- to threefold lower than those
derived from water-bath shaker cultures; however, final
culture optical densities were not significantly affected.
RESULTS
Single BiP/PDI Gene Dose Results
Previously, a Pyroccocus furiosus h-glucosidase trans-
formant of the wild-type BJ5464 Saccharomyces cerevisiae
was isolated (Smith and Robinson, 2002). This trans-
formant, JSIY017, retained high levels of intracellular
h-glucosidase in the ER. The first approach to improving
h-glucosidase folding and secretion through ER manipu-
lation involved the addition of a single copy of BiP or PDI,
alone or in combination. To do this, JSIY017 was
transformed individually and in combination with pCT37
and pGalKar2-LEU, CEN-ARS single-copy plasmids con-
taining galactose-inducible PDI and BiP, respectively.
Empty CEN-ARS plasmids pRS315 and pRS316 (ATCC,
Manassas, VA) containing LEU2 and URA3 metabolic
markers, respectively, were transformed into JSIY017 as
needed to maintain identical metabolic requirements (leu�,
ura�, trp+). Cells were grown and expressed as described in
Materials and Methods. Cell fractions isolated from both
growth and expression media were assayed for BiP and PDI
levels using Western blots (see Materials and Methods). h-
Glucosidase levels in culture supernatant fractions were
determined by activity assays (Materials and Methods) and
are listed in Table I.
Individual overexpression of PDI and BiP resulted in
modest h-glucosidase secretion improvements of approx-
imately 25% and 20%, respectively, over the control
(JSIY017 transformed with pRS315 and pRS316). Co-
overexpression of both BiP and PDI gave a synergistic
effect, resulting in approximately 60% increased extra-
cellular h-glucosidase. Although secretion levels were
increased, intracellular h-glucosidase levels remained es-
sentially unchanged as measured by Western blot analysis
(data not shown), indicating that co-overexpression of ER
chaperones and foldases failed to completely remove the
ER bottleneck. All transformants had approximately iden-
tical growth characteristics in both growth and expression
media; therefore, the metabolic requirements of the cells
were likely equivalent and thus did not impact our results.
In expressing cells, BiP levels were practically unchanged
from the control when BiP, PDI, or both were overexpressed.
In contrast, BiP levels increased twofold when BiP was
overexpressed in BJ5464 parental cells (Table I). PDI
overexpression led to an approximately twofold increase in
PDI levels over the control case when either PDI or both PDI
and BiP were overexpressed.
342 BIOTECHNOLOGY AND BIOENGINEERING, VOL. 85, NO. 3, FEBRUARY 5, 2004
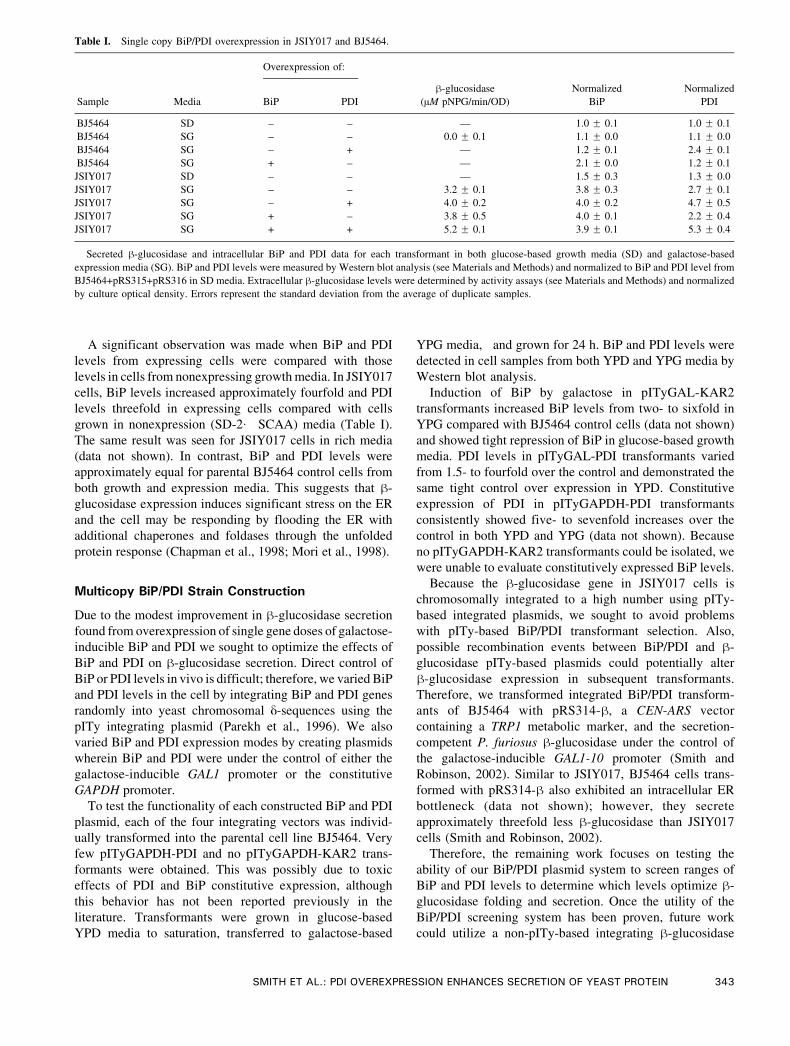
A significant observation was made when BiP and PDI
levels from expressing cells were compared with those
levels in cells from nonexpressing growth media. In JSIY017
cells, BiP levels increased approximately fourfold and PDI
levels threefold in expressing cells compared with cells
grown in nonexpression (SD-2� SCAA) media (Table I).
The same result was seen for JSIY017 cells in rich media
(data not shown). In contrast, BiP and PDI levels were
approximately equal for parental BJ5464 control cells from
both growth and expression media. This suggests that h-
glucosidase expression induces significant stress on the ER
and the cell may be responding by flooding the ER with
additional chaperones and foldases through the unfolded
protein response (Chapman et al., 1998; Mori et al., 1998).
Multicopy BiP/PDI Strain Construction
Due to the modest improvement in h-glucosidase secretion
found from overexpression of single gene doses of galactose-
inducible BiP and PDI we sought to optimize the effects of
BiP and PDI on h-glucosidase secretion. Direct control of
BiP or PDI levels in vivo is difficult; therefore, we varied BiP
and PDI levels in the cell by integrating BiP and PDI genes
randomly into yeast chromosomal y-sequences using the
pITy integrating plasmid (Parekh et al., 1996). We also
varied BiP and PDI expression modes by creating plasmids
wherein BiP and PDI were under the control of either the
galactose-inducible GAL1 promoter or the constitutive
GAPDH promoter.
To test the functionality of each constructed BiP and PDI
plasmid, each of the four integrating vectors was individ-
ually transformed into the parental cell line BJ5464. Very
few pITyGAPDH-PDI and no pITyGAPDH-KAR2 trans-
formants were obtained. This was possibly due to toxic
effects of PDI and BiP constitutive expression, although
this behavior has not been reported previously in the
literature. Transformants were grown in glucose-based
YPD media to saturation, transferred to galactose-based
YPG media, and grown for 24 h. BiP and PDI levels were
detected in cell samples from both YPD and YPG media by
Western blot analysis.
Induction of BiP by galactose in pITyGAL-KAR2
transformants increased BiP levels from two- to sixfold in
YPG compared with BJ5464 control cells (data not shown)
and showed tight repression of BiP in glucose-based growth
media. PDI levels in pITyGAL-PDI transformants varied
from 1.5- to fourfold over the control and demonstrated the
same tight control over expression in YPD. Constitutive
expression of PDI in pITyGAPDH-PDI transformants
consistently showed five- to sevenfold increases over the
control in both YPD and YPG (data not shown). Because
no pITyGAPDH-KAR2 transformants could be isolated, we
were unable to evaluate constitutively expressed BiP levels.
Because the h-glucosidase gene in JSIY017 cells is
chromosomally integrated to a high number using pITy-
based integrated plasmids, we sought to avoid problems
with pITy-based BiP/PDI transformant selection. Also,
possible recombination events between BiP/PDI and h-
glucosidase pITy-based plasmids could potentially alter
h-glucosidase expression in subsequent transformants.
Therefore, we transformed integrated BiP/PDI transform-
ants of BJ5464 with pRS314-h, a CEN-ARS vector
containing a TRP1 metabolic marker, and the secretion-
competent P. furiosus h-glucosidase under the control of
the galactose-inducible GAL1-10 promoter (Smith and
Robinson, 2002). Similar to JSIY017, BJ5464 cells trans-
formed with pRS314-h also exhibited an intracellular ER
bottleneck (data not shown); however, they secrete
approximately threefold less h-glucosidase than JSIY017
cells (Smith and Robinson, 2002).
Therefore, the remaining work focuses on testing the
ability of our BiP/PDI plasmid system to screen ranges of
BiP and PDI levels to determine which levels optimize h-
glucosidase folding and secretion. Once the utility of the
BiP/PDI screening system has been proven, future work
could utilize a non-pITy-based integrating h-glucosidase
Table I. Single copy BiP/PDI overexpression in JSIY017 and BJ5464.
Overexpression of:
Sample Media BiP PDI
h-glucosidase
(AM pNPG/min/OD)
Normalized
BiP
Normalized
PDI
BJ5464 SD – – — 1.0 F 0.1 1.0 F 0.1
BJ5464 SG – – 0.0 F 0.1 1.1 F 0.0 1.1 F 0.0
BJ5464 SG – + — 1.2 F 0.1 2.4 F 0.1
BJ5464 SG + – — 2.1 F 0.0 1.2 F 0.1
JSIY017 SD – – — 1.5 F 0.3 1.3 F 0.0
JSIY017 SG – – 3.2 F 0.1 3.8 F 0.3 2.7 F 0.1
JSIY017 SG – + 4.0 F 0.2 4.0 F 0.2 4.7 F 0.5
JSIY017 SG + – 3.8 F 0.5 4.0 F 0.1 2.2 F 0.4
JSIY017 SG + + 5.2 F 0.1 3.9 F 0.1 5.3 F 0.4
Secreted h-glucosidase and intracellular BiP and PDI data for each transformant in both glucose-based growth media (SD) and galactose-based
expression media (SG). BiP and PDI levels were measured by Western blot analysis (see Materials and Methods) and normalized to BiP and PDI level from
BJ5464+pRS315+pRS316 in SD media. Extracellular h-glucosidase levels were determined by activity assays (see Materials and Methods) and normalized
by culture optical density. Errors represent the standard deviation from the average of duplicate samples.
SMITH ET AL.: PDI OVEREXPRESSION ENHANCES SECRETION OF YEAST PROTEIN 343

plasmid such that a high h-glucosidase gene copy number
and BiP/PDI engineering could be employed concurrently.
BJ5464 cells were transformed with various combina-
tions of BiP and PDI plasmids (galactose-inducible and
constitutive promoters) and selected for resistance to G418.
Approximately 20 to 30 colonies from independent trans-
formations were pooled together and then transformed with
pRS314-h. Transformants were then selected on minimal
media lacking tryptophan and lacking G418, because inte-
grated pITy plasmids are typically stable. This sequential,
single-selection process decreased overall population
variation due to colony pooling, but enabled transformation
with all three expression plasmids.
By transforming BJ5464 with the various combinations
of BiP (Kar2p in yeast) and PDI expression plasmids we
created a fully inducible system, two mixed inducible–
constitutive systems, and a fully constitutive system, all
containing a single gene dose of galactose-inducible h-
glucosidase. The systems were denoted as follows: (1)
GAL-KAR2+GAL-; (2) GAL-KAR2+GAPDH-PDI;
(3) GAPDH-KAR2+GAL-PDI; and (4) GAPDH-KAR2+-
GAPDH-PDI. BJ5464-C (BJ5464 transformed with
pRS314) served as the control for BiP and PDI levels and
BJ5464-h (BJ5464 transformed with pRS314-h) served as
the control for h-glucosidase levels.
Similar to the case for individual integrants of BiP and
PDI into BJ5464, low efficiencies were obtained for
combinations including constitutively expressed BiP vec-
tors, again presumably due to the possible toxic effects of
constitutive BiP overexpression. Therefore, in total, 100
transformants of each GAL-KAR2+GAL-PDI+h and
GAL-KAR2+GAPDH-PDI+h, 20 transformants of
GAPDH-KAR2+GAL-PDI+h, and 19 transformants of
GAPDH-KAR2+GAPDH-PDI+h, where h denotes the
addition of a h-glucosidase gene, were isolated and screened
for improved h-glucosidase secretion.
B-Glucosidase Screen
For all four systems, duplicate samples were grown to
saturation in 5-mL SD-2� SCAA(�trp) (abbreviated SD)
minimal media cultures, then transferred to 5-mL SG-2�SCAA(�trp) (abbreviated SG) minimal media cultures, and
then allowed to express h-glucosidase for 48 h, as described
in Materials and Methods. Culture supernatants were
isolated from SG media and assayed for h-glucosidase
activity (Fig. 1). Activities were normalized by cell optical
density and values reported are relative to BJ5464-h control.
Growth characteristics for all cultures, including the control,
were roughly equivalent (data not shown).
Figure 1. h-Glucosidase screen of the system transformants. Transformants of GAL-KAR2+GAL-PDI+h (A), GAL-KAR2+GAPDH-PDI+h (B),
GAPDH-KAR2+GAL-PDI+h (C), and GAPDH-KAR2+GAPDH-PDI+h (D) were grown in glucose-based growth media to saturation, transferred to
galactose-based media, and expressed for 48 h at 30jC. Culture supernatant samples were isolated and assayed for h-glucosidase activity (see Materials
and Methods). Activity values were normalized to BJ5464-h control cells and reported as relative h-glucosidase levels.
344 BIOTECHNOLOGY AND BIOENGINEERING, VOL. 85, NO. 3, FEBRUARY 5, 2004

A significant number of transformants were found with
h-glucosidase secretion levels higher than the original BiP/
PDI single gene dose experiments. Approximately 10% of
GAL-KAR2+GAL-PDI+h, 15% of GAL-KAR2+GAPDH-
PDI+h and GAPDH-KAR2+GAPDH-PDI+h, and 65%
of GAPDH-KAR2+GAL-PDI+h transformants had im-
proved secretion levels, higher than the 60% improvement
seen for the single BiP/PDI gene overexpression case.
Strains with constitutive BiP expression showed h-glucosi-
dase improvements that were, on average, much higher than
those for inducible BiP and constitutive PDI strains. Strains
with constitutive BiP and inducible PDI expression not only
had a higher percentage of improved transformants, but h-
glucosidase levels were considerably higher than those for
the other combinations. Overall, the screening data suggest
that BiP and PDI expression characteristics (constitutive vs.
Table II. BiP, PDI, and h-glucosidase levels from improved system transformants.
Relative
increase
System name Strain # SD/SG
h-glucosidase
(AM pNPG/min�OD) BiP (AU) PDI (AU)
Approx.
h-glucosidase BiP PDI
GAL-KAR2 + GAL-PDI 16 SD 30.3 F 4.5 26.9 F 4.0 1.3 0.9
SG 2.28 F 0.30 67.3 F 4.8 61.8 F 4.6 2.4 2.8 2.1
62 SD 36.2 F 16.9 41.1 F 13.8 1.5 1.4
SG 1.52 F 0.14 149 F 0 73.1 F 10.6 1.6 6.2 2.5
71 SD 22.3 F 1.2 41.5 F 2.6 0.9 1.4
SG 0.92 F 0.16 164 F 12 56.8 F 13.3 1.0 6.8 2.0
84 SD 27.9 F 3.4 27.8 F 8.4 1.2 1.0
SG 2.11 F 0.11 77.0 F 4.1 28.4 F 1.1 2.2 3.2 1.0
GAL-KAR2 + GAPDH-PDI 40 SD 20.7 F 4.7 94.8 F 5.2 0.9 3.3
SG 2.16 F 0.00 86.6 F 2.2 95.8 F 3.8 2.3 3.6 3.4
67 SD 25.5 F 2.1 26.4 F 0.3 1.1 0.9
SG 1.48 F 0.13 120 F 1 35.6 F 1.3 1.5 5.0 1.2
72 SD 27.1 F 1.9 30.1 F 7.8 1.1 1.1
SG 1.19 F 0.12 44.4 F 5.3 31.4 F 1.2 1.2 1.8 1.1
76 SD 20.4 F 1.2 35.2 F 3.9 0.8 1.2
SG 1.68 F 0.24 63.3 F 2.9 31.2 F 1.1 1.8 2.6 1.1
80 SD 15.2 F 0.7 21.0 F 1.5 0.6 0.7
SG 1.17 F 0.29 123 F 11 41.2 F 1.1 1.2 5.1 1.4
GAPDH-KAR2 + GAL-PDI 1 SD 83.5 F 4.5 25.7 F 2.9 3.5 0.9
SG 2.18 F 0.26 79.3 F 2.1 56.9 F 3.1 2.3 3.3 2.0
4 SD 36.4 F 1.3 37.7 F 1.2 1.5 1.3
SG 2.54 F 0.12 37.1 F 3.9 114 F 4 2.7 1.5 4.0
9 SD 16.9 F 4.4 31.7 F 4.5 0.7 1.1
SG 2.17 F 0.20 21.6 F 0.5 239 F 10 2.3 0.9 8.4
11 SD 32.2 F 1.1 41.6 F 7.9 1.3 1.5
SG 2.45 F 0.18 31.9 F 3.0 224 F 8 2.6 1.3 7.9
12 SD 58.9 F 7.2 24.3 F 0.8 2.4 0.8
SG 2.65 F 0.35 64.8 F 3.2 114 F 6 2.8 2.7 4.0
15 SD 22.2 F 2.3 14.8 F 6.6 0.9 0.5
SG 2.06 F 0.51 47.0 F 18.6 146 F 58 2.2 2.0 5.1
16 SD 73.6 F 1.0 27.9 F 4.3 3.1 1.0
SG 2.95 F 0.18 69.1 F 0.7 284 F 15 3.1 2.9 9.9
GAPDH-KAR2 + GAPDH-PDI 6 SD 63.3 F 2.0 120 F 11 2.6 4.2
SG 1.93 F 0.24 63.4 F 0.6 153 F 32 2.0 2.6 5.3
7 SD 24.1 F 0.7 235 F 5 1.0 8.2
SG 2.15 F 0.10 32.5 F 0.8 239 F 21 2.2 1.3 8.4
11 SD 29.4 F 6.8 213 F 10 1.2 7.5
SG 2.24 F 0.17 38.2 F 2.6 202 F 9 2.3 1.6 7.1
BJ5464-h SD 21.9 F 2.3 25.8 F 0.9 0.9 0.9
SG 0.96 F 0.04 33.9 F 4.4 23.7 F 3.1 1 1.4 0.8
BJ5464-C SD 24.1 F 0.3 28.9 F 2.3 1.0 1.0
SG 0.03 F 0.03 17.1 F 0.9 9.0 F 3.3 0.7 0.3
A subset of improved strains from Figure 1 were grown and expressed in duplicate as in Table I. Cell samples were taken in SD and SG media and
analyzed for BiP and PDI levels by Western blot analysis. SG culture supernatants were assayed for h-glucosidase activity as before (reported in AM pNPG/
min/OD). BiP and PDI levels were measured by densitometric analysis of Western blots and reported in arbitrary units. Transformant h-glucosidase levels
were normalized relative to the control (BJ5464+pRS314-h), and BiP and PDI levels were normalized to the overall control (BJ5464+pRS314). Boldface
entries highlight those samples that were used for expression vs. time studies (Fig. 3). Reported errors represent the standard deviation between duplicate
expression experiments. Errors in "approximate relative increases" were typically F10%.
SMITH ET AL.: PDI OVEREXPRESSION ENHANCES SECRETION OF YEAST PROTEIN 345

inducible) play an important role in improved h-glucosi-
dase secretion.
Investigation of Improved Strains
To examine these improvements in more detail, a subset of
improved transformants from each system was re-expressed
in 5-mL cultures. In all, four GAL-KAR2+GAL-PDI+h,
five GAL-KAR2+GAPDH-PDI+h, seven GAPDH-KAR2+
GAL-PDI+h, and three GAPDH-KAR2+GAPDH-PDI+htransformants were investigated further. Transformants
GAL-KAR2+GAPDH-PDI+h #80 and GAL-KAR2+GAL-
PDI+h #71 had secretion levels close to that of the BJ5464-hcontrol, and were used as a type of ‘‘negative control.’’
The 19 transformants and 2 controls (BJ5464-C and
BJ5464-h) were grown and expressed as before. Cell
samples were taken in both SD and SG minimal media and
intracellular BiP and PDI levels were determined using
Western blot analysis. Cell culture supernatants were
isolated from SG media cultures and assayed for h-
glucosidase activity. BiP, PDI, and h-glucosidase values
for all samples in both SD and SG media are summarized
in Table II.
Unlike the case of multicopy h-glucosidase expression
(JSIY017 cells in Table I), BJ5464-h BiP and PDI levels
appeared similar to the BJ5464-C control. BiP and PDI
levels did not change significantly with the single gene
copy of h-glucosidase (BJ5464-h), whereas both proteins
increased approximately fourfold in JSIY017-expressing
cells. Therefore, it appears that, although both cells have
difficulty in properly exporting h-glucosidase (Smith and
Robinson, 2002), the added h-glucosidase gene copies and
increased unfolded protein in the ER of JSIY017 appeared
to cause a considerably higher stress on these cells.
A plot of the relative BiP and PDI levels derived from
SG cultures (Table II) shows that a wide range of both PDI
and BiP levels enabled secretion improvements (Fig. 2A).
However, transformants tended to have high BiP and low
PDI levels, high PDI and low BiP levels, or moderate levels
of each. In no case was a transformant isolated with high
levels of both BiP and PDI. This may indicate a true trade-
off in expression, or may be an artifact of the limited
sampling. Also, contrary to the observation from the overall
screen (Fig. 1), inducible vs. constitutive expression did not
appear to play as large a role as simply the total PDI or BiP
present in the cell.
Significant trends were found by plotting the relative
increase in h-glucosidase levels vs. that of either PDI or
BiP. Increasing intracellular BiP levels decreased improve-
ments in h-glucosidase secretion (Fig. 2C). This was
especially evident for the ‘‘negative controls’’ GAL-
KAR2+GAPDH-PDI+h #80 and GAL-KAR2+GAL-PDI +h#71, which had high levels of BiP, low levels of PDI, and
approximately the same h-glucosidase secretion levels as
BJ5464-h. PDI levels showed an opposite correlation,
where increased PDI levels increased h-glucosidase levels
(Fig. 2B).
The complication in examining the response of h-
glucosidase to PDI and BiP is that BiP and PDI work
synergistically, as observed from single BiP/PDI experi-
ments (Table I). Because BiP and PDI are not independent
of one another, the effects of BiP and PDI seen in Figure 2B
and C cannot be completely decoupled. From Figure 2A we
see that low BiP levels were typically accompanied by high
PDI and vice versa. This may explain why the data in
Figure 2B and C were more scattered at low BiP/PDI con-
centrations and then became less varied at high levels. The
dependence of h-glucosidase at low levels of BiP or PDI
actually includes a significant contribution by the other pro-
tein. For example, in Figure 2B, samples with low PDI and
h-glucosidase levels below the trend line tended to have
high levels of BiP.
Transient Detection of BiP, PDI, and B-Glucosidase
The trends seen in Figure 2 resulted from static timepoint
experiments. To more completely characterize the effects of
BiP and PDI on h-glucosidase secretion to determine whe-
ther we could clarify the role of these proteins, we followed
BiP, PDI, and intracellular h-glucosidase levels over time.
Cultures of GAPDH-KAR2+GAL-PDI+h #16, GAPDH-
KAR2+GAPDH-PDI+h #6, GAL-KAR2+GAPDH-PDI+h#40, and BJ5464-h were sampled at multiple timepoints
Figure 2. Graphical presentation of Table II data. Relative levels of BiP,
PDI, and h-glucosidase derived from SG media cultures were plotted
against each other for system GAL-KAR2+GAL-PDI+h (.), GAL-
KAR2+GAPDH-PDI+h (o), GAPDH-KAR2+GAL-PDI+h (n), and
GAPDH-KAR2+GAPDH-PDI+h (5). Plots show the relative levels of
PDI vs. BiP (A), h-glucosidase vs. PDI (B), and h-glucosidase vs. BiP (C)
over the controls (BJ5464-h for h-glucosidase levels and BJ5464-C for
BiP and PDI levels). Lines shown for (B) and (C) indicate the linear least-
squares fit of the data. R2 values for both plots were approximately 0.4. (D)
BiP and PDI Western blot images for represented transformants in both SD
and SG media. Errors in relative BiP and PDI levels, as determined by the
standard deviation of duplicate samples quantified by Western blot
analysis, are shown for (A).
346 BIOTECHNOLOGY AND BIOENGINEERING, VOL. 85, NO. 3, FEBRUARY 5, 2004

postinduction (SG media), and analyzed by Western blots for
BiP, PDI, and h-glucosidase levels (Fig. 3). These four
transformants were chosen for their range of BiP, PDI, and
secreted h-glucosidase levels as well as their variation in
expression modes. The 0- and 48-h timepoints were in good
agreement with the data from Table II. Stationary phase for
all four cultures was reached at approximately 40 to 48 h.
As presented in Figure 3, the onset of h-glucosidase
secretion and the rate of accumulation did not appear to be
affected by the levels or expression modes (constitutive or
inducible) of BiP and PDI. h-Glucosidase levels quickly
rose to approximately maximal levels within 16 to 20 h
postinduction. BiP and PDI levels also reached their highest
levels within this time frame. Overall, the behavior of BiP
and PDI was as expected for promoter regulation.
However, an interesting observation was made from
examination of BJ5464-h. In BJ5464-h control cells
(Fig.3D), BiP and h-glucosidase levels rose throughout the
course of the experiment. PDI levels for this strain showed a
small initial induction (f60%) followed by a return to
approximately initial conditions. The induction and repres-
sion of PDI seen in BJ5464-h may indicate PDI induction in
an unfolded protein response of the cell. A smaller induction
(f20%) was observed in GAL-KAR2+GAPDH-PDI+h #40.
This effect may not have been seen with the other two cell
strains because PDI levels were already elevated to high
levels (9.9- and 5.3-fold vs. 3.4-fold relative to BJ5464-C for
GAL-KAR2+GAPDH-PDI+h #40; Table II).
A change in the ER bottleneck might be observed by
comparing the levels of intracellular h-glucosidase. A
comparison of the intracellular h-glucosidase in all four
transformants is shown in Figure 4. The best secretion
transformant, GAPDH-KAR2+GAL-PDI+h #16, showed an
earlier decrease in intracellular h-glucosidase than the other
three after approximately 8 h, and reached the lowest final
levels of the four transformants. Intracellular h-glucosidase
levels for GAPDH-KAR2+GAPDH-PDI+h #6 and GAL-
KAR2+GAPDH-PDI+h #40 were approximately equal to
that of BJ5464-h until after approximately 16 h, and
reached final levels much lower than that of BJ5464-h.
Intracellular levels were inversely related to the secreted
levels of h-glucosidase for these transformants (Table II).
Because BiP levels were relatively equal among the trans-
formants, the data suggest that PDI acts to lower intra-
cellular h-glucosidase levels that, in turn, leads to increased
secreted h-glucosidase.
DISCUSSION
Single BiP/PDI Gene Expression
We previously reported the existence of an intracellular ER
bottleneck in our high h-glucosidase gene copy S. cerevisiae
transformant, JSIY017 (Smith and Robinson, 2002). There
are numerous instances in the literature wherein over-
expression of ER chaperones and foldases has been used to
increase secretion yields. Therefore, we investigated the
effect of single gene doses of the ER-resident proteins BiP
and PDI on h-glucosidase secretion in JSIY017.
Overexpression of both BiP and PDI synergistically
improved h-glucosidase secretion. Interestingly, this type
of result has been seen with recombinant single-chain
antibody secretion in S. cerevisiae, wherein overexpression
of BiP and PDI increased scFv secretion levels higher than
the additive effects of BiP and PDI alone (Shusta et al.,
Figure 3. Intracellular BiP, PDI, and h-glucosidase levels over time.
GAPDH-KAR2+GAL-PDI+h #16 (A), GAPDH-KAR2+GAPDH-PDI+h #6
(B), GAL-KAR2+GAPDH-PDI+h #40 (C), and BJ5464-h (D) were grown
to saturation in SD media. Also, 50 OD-mL equivalents of cells were
isolated and resuspended in 50 mL SG media (t = 0). Cell samples were
taken at various timepoints postinduction and analyzed by Western blot for
h-glucosidase (x), BiP (5), and PDI (o). All species were normalized by
the highest signal for that species.
Figure 4. Intracellular h-glucosidase levels over time. Non-normalized
h-glucosidase levels for GAPDH-KAR2+GAL-PDI+h #16 (n), GAPDH-
KAR2+GAPDH-PDI+h #6 (5), GAL-KAR2+GAPDH-PDI+h #40 (o), and
BJ5464-h (E) are plotted for direct comparison. Intracellular h-
glucosidase levels were measured by Western blot analysis and reported
in arbitrary units.
SMITH ET AL.: PDI OVEREXPRESSION ENHANCES SECRETION OF YEAST PROTEIN 347

1998). This effect was also seen in the in vitro refolding of
reduced Fab fragments in the presence of BiP and PDI
(Mayer et al., 2000). Buchner and colleagues suggested that
BiP held open the folding intermediate, which gave PDI
access to the disulfide bonds. However, because P. furiosus
h-glucosidase has no disulfide bonds and only one cysteine
per monomer, PDI’s role as a foldase would not explain its
effect here.
Overexpression of BiP from a single-copy plasmid
pGalKar2-LEU in JSIY017 did not change BiP protein
levels. Perhaps the cell autoregulates BiP levels though
transcriptional or translational control in this case. Interest-
ingly, PDI levels do not respond in a similar manner.
Overexpression of PDI leads to a twofold increase over
JSIY017 control cells from SG media when PDI is
overexpressed by itself or in concert with BiP. Both PDI
and BiP are a part of the unfolded protein response (UPR)
(Chapman et al., 1998; Mori et al., 1998); however, our data
may suggest that BiP and PDI protein levels are controlled in
different ways.
Multicopy BiP/PDI Optimization
Due to the possible integration and selection difficulties of
using pITy-based h-glucosidase and BiP/PDI plasmids, we
employed the pITy-based BiP/PDI plasmids to improve
secretion of h-glucosidase from wild-type BJ5464 trans-
formed with the CEN-ARS zpRS314-h plasmid (Smith and
Robinson, 2002). Therefore, the utility of the varying BiP
and PDI levels was determined so that in the future this
knowledge culd be applied to high h-glucosidase expression
systems or expression systems with other hetero-
logous proteins.
In this study we optimized h-glucosidase secretion in
BJ5464 cells transformed with pRS314-h by varying BiP
and PDI levels through random chromosomal BiP/PDI gene
integration and then screening for higher h-glucosidase
secretion levels (Fig. 1). A subset of improved trans-
formants was then examined for intracellular h-glucosidase
as well as BiP and PDI levels (Fig. 2). Two general trends
emerged: increasing BiP levels decreased h-glucosidase
secretion, whereas increasing PDI levels increased h-
glucosidase secretion.
Increasing BiP levels may result in decreased h-
glucosidase secretion due to prolonged interaction with
immature h-glucosidase. Secretory proteins tend to have a
unique, individual average residence time in the ER, due in
part to the folding-mediated effects of ER-resident chaper-
ones and foldases (Pelham, 1989; Rowling and Freedman,
1993). Typically, the shorter the residence time in the ER,
the faster the proteins are secreted. On the other hand, if
chaperone levels are increased, ER residence times may
increase due to prolonged interaction with ER-resident
proteins and lower overall secretion yields. As an example,
elevated levels of Grp94p, an ER chaperone, were found to
increase the residence times for thyroglobulin in Chinese
hamster ovary cells (Muresan and Arvan, 1997).
However, from single BiP/PDI gene overexpression
results, we found that BiP and PDI together improved h-
glucosidase secretion. Therefore, although a positive role
for BiP at increased levels was not seen (Fig. 2), data for
samples GAPDH-KAR2+GAL-PDI+h #12 and #16 may
indicate that increased levels of BiP were necessary, albeit
at moderate levels.
In contrast to BIP effects, increased PDI levels increased
h-glucosidase levels (Fig. 2B). Figure 4 suggests that
increased PDI functions to lower intracellular h-glucosi-
dase, which leads to elevated secreted h-glucosidase yields.
However, the role of PDI in h-glucosidase folding and
secretion is less clear, because h-glucosidase lacks
disulfide bonds.
We postulate two possible modes of action for PDI.
Although the mechanism of action is not well understood, at
stoichiometric levels PDI can act as a chaperone (Dai and
Wang, 1997; Klappa et al., 1998; LaMantia and Lennarz,
1993). It is possible PDI may act as a chaperone for h-
glucosidase in a way that BiP cannot, a role that does not
become saturating, even at high PDI levels. Alternatively,
PDI may be forming mixed, transient disulfide bonds with
the h-glucosidase, helping to segregate its folding environ-
ment or limiting its conformational explorations in the
process of proper folding.
Thaumatin secretion was improved by increasing intra-
cellular PDI levels two- to threefold; however, according to
Moralejo et al. (2001), increasing PDI above this level
results in decreased overall yields. They reasoned that the
high levels of PDI may unfavorably alter the folding
equilibrium, leading to nonsecretable forms of thaumatin.
At high levels, PDI has been found to act as an anti-
chaperone and aggregate immature proteins regardless of
their disulfide character (Quan et al., 1995). Therefore, one
could imagine that an optimum level of PDI might be found
if it were acting as a chaperone. However, for h-glucosidase,
only a solely beneficial role for PDI was observed.
In preliminary efforts to examine the possible foldase role
of PDI in vivo, DTT was added to the culture media in an
attempt to inhibit PDI’s foldase ability. We grew four
selected transformants in SG media containing 1 mM and
5 mM DTT. Cell yields were significantly hampered in the
presence of DTT, reaching approximately 60% and 10% of
final ODs, respectively, for the 1 mM and 5 mM DTT cases,
compared with expression in the absence of DTT (data not
shown). On a per-cell basis, h-glucosidase secretion was
approximately equal for all samples in the absence or
presence of DTT (data not shown). Therefore, these results
suggest that PDI does not form mixed disulfides, but rather
acts primarily as a chaperone. However, DTT is used widely
to invoke the cellular unfolded protein response (Gasch et al.,
2000), which causes a shift in the expression of many genes,
including the ER-resident proteins BiP and PDI. As a result,
direct effects of DTT are not completely conclusive.
Additional studies are being conducted to test the effect
of site-directed mutagenesis of the sole cysteine in h-
glucosidase on its secretion and the role of PDI. We also
348 BIOTECHNOLOGY AND BIOENGINEERING, VOL. 85, NO. 3, FEBRUARY 5, 2004

plan to study PDI interactions with h-glucosidase in in vitro
refolding studies. By refolding h-glucosidase in the
presence of PDI mutants, which possess only a chaperone
(Wang, 2002) or a foldase ability (Dai and Wang, 1997),
the role of PDI could possibly be deduced.
Impact on Heterologous Protein Secretion
Often, random screens of different cell strains are
conducted to find an optimal strain for production. Here
we attempted a more rational cellular engineering approach
based on the determination of an ER bottleneck. Overall,
we have successfully created a system allowing for
screening of a range of BiP and PDI levels to improve
heterologous protein secretion. Relative to the wild-type
strain (containing one h-glucosidase gene), secretion was
improved approximately threefold in the best strain, and for
that strain PDI and BiP levels were approximately ten- and
threefold, respectively, higher than with the wild-type cells.
The scope of this approach is significantly broader than
that of previous studies of chaperone overexpression effects.
It allows for not only the selection of optimal PDI and BiP
levels, but also the identification of whether these proteins
aid or inhibit heterologous protein secretion. BiP and PDI
screening could easily be supplemented to include other
chaperones. This approach could be particularly useful in the
biotechnology industry, wherein many of the therapeutic
proteins are complex and require the assistance of chaper-
ones and foldases.
The trends seen here with h-glucosidase are likely not
directly translatable to other proteins, as different proteins
have demonstrated different behaviors in response to mole-
cular chaperones. In addition, the chaperone/foldase needs
of a single protein may change depending on environmental
changes. For example, h-glucosidase requirements at 37jC
expression are distinct from those at 30jC (unpublished
data); high PDI levels appear to lower h-glucosidase se-
cretion at 37jC.
CONCLUSIONS
We previously reported a high h-glucosidase gene copy
Saccharomyces cerevisiae transformant that could secretef12 mg/L, but also retained a large intracellular bottleneck
in the ER. In an effort to relieve this bottleneck, an
additional BiP and PDI gene was introduced into these cells,
either alone or in combination. A synergistic effect was seen
between PDI and BiP, resulting in f60% improved
secretion levels. Due to this modest increase, we attempted
to optimize the effects of BiP and PDI on h-glucosidase
secretion by randomly varying integrated BiP/PDI gene
copynumbers. Transformants were screened for improvedh-
glucosidase secretion and a subset of improved strains were
re-examined for h-glucosidase secretion and intracellular
BiP and PDI levels. Two general trends emerged: increasing
BiP levels decreased h-glucosidase secretion, whereas
increasing PDI increased h-glucosidase secretion. However,
these relationships are not independent of each other and
deviations from the overall trend are especially evident at
low BiP and PDI levels. Increased PDI levels decreased the
levels of intracellularly retained h-glucosidase. Although
preliminary results indicate that PDI acts as a unique
chaperone for h-glucosidase, PDI may possibly be forming
mixed disulfides with h-glucosidase during the folding
process. Overall, we have successfully created a screening
system allowing for qualitative understanding of multiple
chaperone/foldase effects on a heterologous protein.
The authors thank Jeff Brodsky (University of Pittsburgh), K. Dane
Wittrup (MIT), and Bob Kelly (North Carolina State University) for
their generous gifts of the BiP antibody, pITy plasmid, and h-
glucosidase gene, respectively.
References
Brodsky JL, Werner ED, Dubas ME, Goeckeler JL, Kruse KB, McCracken
AA. 1999. The requirement for molecular chaperones during the
endoplasmic reticulum-associated protein degradation demonstrates
that protein export and import are mechanistically distinct. J Biol
Chem 274:3453– 3460.
Chapman R, Sidrauski C, Walter P. 1998. Intracellular signaling from the
endoplasmic reticulum to the nucleus. Annu Rev Cell Dev Biol 14:
459–485.
Dai Y, Wang C. 1997. A mutant truncated protein disulfide isomerase with
no chaperone activity. J Biol Chem 272:27572– 27576.
Gasch AP, Spellman PT, Kao CM, Carmel-Harel O, Eisen MB, Storz G,
Botstein D, Brown PO. 2000. Genomic expression programs in the
response of yeast cells to environmental changes. Mol Biol Cell
11:4241– 4257.
Haas I, Wabl M. 1983. Immunoglobulin heavy chain binding protein.
Nature 306:387– 389.
Klappa P, Ruddock LW, Darby NJ, Freedman RB. 1998. The bV domain
provides the principal peptide-binding site of protein disulfide
isomerase but all domains contribute to binding of misfolded proteins.
EMBO J 17:927–935.
Laboissiere MCA, Sturley SL, Raines RT. 1995. The essential fuction of
protein-disulfide isomerase is to unscramble non-native disulfide
bonds. J Biol Chem 270:28006–28009.
LaMantia M, Lennarz WJ. 1993. The essential function of yeast protein
disulfide isomerase does not reside in its isomerase activity. Cell 74:
899–908.
Matlack KES, Misselwitz B, Plath K, Rapoport TA. 1999. BiP acts as a
molecular ratchet during posttranslational transport of prepro-a factor
across the ER membrane. Cell 97:553– 564.
Mayer M, Kies U, Kammermeier R, Buchner J. 2000. BiP and PDI
cooperate in the oxidative folding of antibodies in vitro. J Biol Chem
275:29421–29425.
Moralejo FJ, Watson AJ, Jeenes DJ, Archer DB, Martin JF. 2001. A
defined level of protein disulfide isomerase expression is required for
optimal secretion of thaumatin by Aspergillus awamori. Mol Genet
Genom 266:246–253.
Mori K, Ogawa N, Kawahara T, Yanagi H, Yura T. 1998. Palindrome with
spacer of one nucleotide is characteristic of the cis-acting unfolded
protein response element in Saccharomyces cerevisiae. J Biol Chem
273:9912–9920.
Muresan Z, Arvan P. 1997. Thyroglobulin transport along the secretory
pathway. J Biol Chem 272:26095– 26102.
Quan H, Fan GB, Wang CC. 1995. Independence of the chaperone activity
of protein disulfide isomerase from its thioredoxin-like active site. J
Biol Chem 270:17077–17079.
Parekh RN, Forrester K, Wittrup KD. 1995. Multicopy overexpression of
bovine pancreatic trypsin inhibitor saturates the protein folding and
SMITH ET AL.: PDI OVEREXPRESSION ENHANCES SECRETION OF YEAST PROTEIN 349

secretory capacity of Saccharomyces cerevisiae. Prot Expr Purif
6:537– 545.
Parekh RN, Shaw MR, Wittrup KD. 1996. An integrating vector for
tunable, high copy, stable integration into the dispersed Ty y sites of
Saccharomyces cerevisiae. Biotechnol Progr 12:16–21.
Parekh RN, Wittrup KD. 1997. Expression level tuning for optimal
heterologous protein secretion in Saccharomyces cerevisiae. Bio-
technol Progr 13:117– 122.
Pelham HRB. 1989. Control of protein exit from the endoplasmic
reticulum. Ann Rev Cell Biol 5:1 –23.
Robinson AS, Bockhaus JA, Voegler AC, Wittrup KD. 1996. Reduction of
BiP levels decrease heterologous protein secretion in Saccharomyces
cerevisiae. J Biol Chem 271:10017– 10022.
Robinson AS, Hines V, Wittrup KD. 1994. Protein disulfide isomerase
overexpression increases secretion of foreign proteins in Saccharo-
myces cerevisiae. Bio/Technology 12:381– 384.
Robinson AS, Wittrup KD. 1995. Constitutive overexpression of secreted
heterologous protein decreases extractable bip and protein disulfide
isomerase levels in Saccharomyces cerevisiae. Biotechnol Progr
11:171–177.
Rowling PJE, Freedman RB. 1993. Folding, assembly, and posttransla-
tional modification of proteins within the lumen of the endoplasmic
reticulum. In: Borgese N, Harris JR, editors. Subcellular biochemistry.
New York: Plenum Press. p 41–81.
Ryabova LA, Desplancq D, Spirin AS, Pluckthun A. 1997. Functional
antibody production using cell-free translation: Effects of protein
disulfide isomerase and chaperones. Nat Biotechnol 15:79–83.
Shusta EV, Raines RT, Pluckthun A, Wittrup KD. 1998. Increasing the
secretory capacity of Saccharomyces cerevisiae for production of
single-chain antibody fragments. Nature Biotechnology 16:773– 777.
Smith JD, Robinson AS. 2002. Overexpression of an archaeal protein in
yeast: secretion bottleneck at the ER. Biotechnol Bioeng 79:713– 723.
Tachibana C, Stevens TH. 1992. The yeast EUG1 gene encodes an
endoplasmic reticulum protein that is functionally related to protein
disulfide isomerase. Mol Cell Biol 12:4601 – 4611.
Wang C-C. 2002. Protein disulfide isomerase as an enzyme and a
chaperone in protein folding. In: Sies H, Packer L, editors. Methods in
enzymology. Vol. 348. San Diego, CA: Academic Press. p 66– 75.
Wood T, Bhat KM. 1988. Biomass part A cellulose and hemicellulose. In:
Wood WA, Kellogg ST, editors. Methods in enzymology. Vol. 160.
San Diego, CA: Academic Press. p 87– 112.
Wittrup KD, Benig V. 1994. Optimization of amino-acid supplements for
heterologous protein secretion in Saccharomyces cerevisiae. Bio-
technol Techn 8:161–166.
350 BIOTECHNOLOGY AND BIOENGINEERING, VOL. 85, NO. 3, FEBRUARY 5, 2004




















