
Progesterone Receptor-Induced Bending of its Target DNA
15
Progesterone Receptor-Induced Bending of its Target DNA: Distinct Effects of the A and B Receptor Forms Paul Prendergast, Zhaoxing Pan, and Dean P. Edwards Department of Pathology (P.P., D.P.E.) and Molecular Biology Program (D.P.E.) Department of Preventive Medicine and Biometrics (Z.P.) University of Colorado Health Sciences Center Denver, Colorado 80262 We have used circular permutation and phasing electrophoretic mobility shift assays to determine the ability of the A and B forms of human proges- terone receptor (PR) to bend target DNA. Studies were done with baculovirus-expressed full-length receptors purified to apparent homogeneity. By circular permutation analysis, both forms of PR induced substantial distortions in the structure of target DNA with calculated distortion angles ((up) of 57” for PR-A and 64” for PR-B. The apparent bend centers for both forms of PR were similarly located a few base pairs (-4 to -2 bp) from the middle of the progesterone response element. No differ- ences were detected in the magnitude of distortion or apparent bend centers when PR was bound to hormone agonist (R5020) or the antagonist RU466. Phasing analysis, which can determine the orien- tation of a DNA bend, revealed that both forms of PR mediated directional bends toward the major groove of the DNA helix. Calculated directed bend angles (cud were 40” for PR-B and 31” for PR-A. The chromatin high mobility group protein HMG-1, which acts as an accessory factor to enhance the binding affinity of purified PR for progesterone re- sponse elements, had minimal influence on PR- mediated DNA bending. This result, taken together with the fact that HMG-1 can form a ternary com- plex with PR and DNA, is consistent with the con- clusion that HMG-1 facilitates PR binding by sta- bilizing a receptor-induced DNA conformation that is required for assembly of a high affinity PR-DNA complex. The results of this study also suggest that DNA bending may be coupled to transcriptional regulation since PR-B is generally a stronger tran- scriptional activator than PR-A and also mediates a larger bend in target DNA than PR-A. (Molecular Endocrinology 10: 393-407, 1996) 0888-8809/96/$3.00/O Molecular Endocrinology Copyright 0 1996 by The Endocrine Society INTRODUCTION The progesterone receptor (PR) is a member of the steroid receptor gene family of ligand-dependent tran- scriptional regulators that mediate their effects on gene transcription by binding to specific hormone re- sponse elements of steroid-responsive genes (1, 2). The mechanism by which receptor-DNA interaction promotes changes in gene transcription is not well understood. This likely involves protein-protein inter- actions either with basal transcription factors (3-5) or with intermediary coactivators (6-9). Many prokaryotic and eukaryotic sequence-specific transcriptional activators have been shown to be ca- pable of bending DNA when bound to their recognition element (10-20). Because the DNA helix is relatively inflexible over short distances, interaction between proteins bound to neighboring sites may require de- formation of the intervening DNA structure (11,21,22). Therefore, DNA bending may be coupled to transcrip- tion as a mechanism with which to facilitate contact of upstream activators with other factors in the transcrip- tional apparatus. Several studies with prokaryotic transcriptional activators that bend DNA have shown that an intrinsic DNA- bending sequence, or an unre- lated DNA-bending protein and insertion of its binding sequence, can functionally substitute for the transcrip- tion factor (20, 23, 24). This indicates that protein- induced changes in DNA topology may be sufficient for enhancement of transcription. In eukaryotes, DNA bending also appears to play a role in transcriptional regulation. There is a class of eukaryotic transcrip- tional activators that bind to recognition sequences located on a variety of cellular and viral promoters but have little intrinsic activation potential. These factors act by sharply bending DNA to facilitate assembly of higher order nucleoprotein complexes, thus creating the structural framework for other transcription factors to interact and function. Examples of such context- dependent architectural transcription factors (25) are the lymphocyte enhancer factor LEF-1 (26) the chro- 393 Downloaded from https://academic.oup.com/mend/article/10/4/393/2713356 by guest on 09 July 2022
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of Progesterone Receptor-Induced Bending of its Target DNA

Progesterone Receptor-Induced Bending of its Target DNA: Distinct Effects of the A and B Receptor Forms
Paul Prendergast, Zhaoxing Pan, and Dean P. Edwards
Department of Pathology (P.P., D.P.E.) and Molecular Biology Program (D.P.E.) Department of Preventive Medicine and Biometrics (Z.P.) University of Colorado Health Sciences Center Denver, Colorado 80262
We have used circular permutation and phasing electrophoretic mobility shift assays to determine the ability of the A and B forms of human proges- terone receptor (PR) to bend target DNA. Studies were done with baculovirus-expressed full-length receptors purified to apparent homogeneity. By circular permutation analysis, both forms of PR induced substantial distortions in the structure of target DNA with calculated distortion angles ((up) of 57” for PR-A and 64” for PR-B. The apparent bend centers for both forms of PR were similarly located a few base pairs (-4 to -2 bp) from the middle of the progesterone response element. No differ- ences were detected in the magnitude of distortion or apparent bend centers when PR was bound to hormone agonist (R5020) or the antagonist RU466. Phasing analysis, which can determine the orien- tation of a DNA bend, revealed that both forms of PR mediated directional bends toward the major groove of the DNA helix. Calculated directed bend angles (cud were 40” for PR-B and 31” for PR-A. The chromatin high mobility group protein HMG-1, which acts as an accessory factor to enhance the binding affinity of purified PR for progesterone re- sponse elements, had minimal influence on PR- mediated DNA bending. This result, taken together with the fact that HMG-1 can form a ternary com- plex with PR and DNA, is consistent with the con- clusion that HMG-1 facilitates PR binding by sta- bilizing a receptor-induced DNA conformation that is required for assembly of a high affinity PR-DNA complex. The results of this study also suggest that DNA bending may be coupled to transcriptional regulation since PR-B is generally a stronger tran- scriptional activator than PR-A and also mediates a larger bend in target DNA than PR-A. (Molecular Endocrinology 10: 393-407, 1996)
0888-8809/96/$3.00/O Molecular Endocrinology Copyright 0 1996 by The Endocrine Society
INTRODUCTION
The progesterone receptor (PR) is a member of the steroid receptor gene family of ligand-dependent tran- scriptional regulators that mediate their effects on gene transcription by binding to specific hormone re- sponse elements of steroid-responsive genes (1, 2). The mechanism by which receptor-DNA interaction promotes changes in gene transcription is not well understood. This likely involves protein-protein inter- actions either with basal transcription factors (3-5) or with intermediary coactivators (6-9).
Many prokaryotic and eukaryotic sequence-specific transcriptional activators have been shown to be ca- pable of bending DNA when bound to their recognition element (10-20). Because the DNA helix is relatively inflexible over short distances, interaction between proteins bound to neighboring sites may require de- formation of the intervening DNA structure (11,21,22). Therefore, DNA bending may be coupled to transcrip- tion as a mechanism with which to facilitate contact of upstream activators with other factors in the transcrip- tional apparatus. Several studies with prokaryotic transcriptional activators that bend DNA have shown that an intrinsic DNA- bending sequence, or an unre- lated DNA-bending protein and insertion of its binding sequence, can functionally substitute for the transcrip- tion factor (20, 23, 24). This indicates that protein- induced changes in DNA topology may be sufficient for enhancement of transcription. In eukaryotes, DNA bending also appears to play a role in transcriptional regulation. There is a class of eukaryotic transcrip- tional activators that bind to recognition sequences located on a variety of cellular and viral promoters but have little intrinsic activation potential. These factors act by sharply bending DNA to facilitate assembly of higher order nucleoprotein complexes, thus creating the structural framework for other transcription factors to interact and function. Examples of such context- dependent architectural transcription factors (25) are the lymphocyte enhancer factor LEF-1 (26) the chro-
393
Dow
nloaded from https://academ
ic.oup.com/m
end/article/10/4/393/2713356 by guest on 09 July 2022

MOL END0 .1996 VollONo.4 394
matin high mobility group protein HMG I (Y) (27), mam- malian sex-determining factor SW (28) and the zinc finger protein WI that binds to several sites in the c-fos promoter (29).
The functional significance of DNA bending by eukaryotic sequence-specific DNA-binding proteins that possess intrinsic activation function is less clear. Studies with eukaryotic promoters have shown that intrinsic DNA-bending sequences can only partially substitute for specific protein-DNA interactions. This indicates that DNA bending may be involved but is not sufficient as a transcriptional regulatory mechanism as has been shown with prokaryotic genes (30,31). Sub- stantial differences in the magnitude and orientation of bending of the same DNA target element have been reported for various hetero- and homodimers of the bZlP family of transcriptional activators (11, 32) as well as hetero- and homodimeric forms of Myc and Max (14). This differential DNA bending has been sug- gested to contribute to the functional diversity of these related proteins that bind to the same, or similar, se- quences by creating spatial orientations that promote distinct protein-protein interactions. DNA bending may also be involved in transcriptional regulation by providing an optimal stereochemical configuration for protein DNA contacts. For certain sequence-specific DNA-binding proteins, correlations have been ob- served between the strength of DNA binding and the extent of DNA bending (33, 34). Additionally, intrinsic DNA bends have been shown to dramatically increase the binding affinity of certain transcription factors (34-36).
Steroid receptor-mediated bending of DNA has been demonstrated by circular permutation assay for thyroid hormone receptor (TR) (37, 38), estrogen re- ceptor (ER) (39-41), and retinoic acid X receptor (RXR), the latter when complexed to different dimer partners (42). Additionally, ER and various RXR heterodimers have been shown by phasing analysis to cause a directional bend toward the major groove of the DNA helix (31, 42). The influence of PR on target DNA conformation has not been explored and is of partic- ular interest for two reasons. First, human PR is ex- pressed as two distinct molecular forms, full length PR-B and N-terminally truncated PR-A (43), and each receptor form has been shown to exhibit different functional properties dependent on the cellular and target promoter context (44, 45). Second, human PR requires a nuclear accessory factor for high affinity binding to progesterone response elements (PREs) in vitro (46) which has been identified as the chromatin high mobility group protein 1 (HMG-1) (47). Interest- ingly, HMG-1 has no known DNA sequence specificity but prefers to bind to distorted DNA structures and, when present in high concentrations, can bend DNA (47-49). In the present study we have used circular permutation and phasing analyses to determine the ability of purified A and B forms of human PR to bend target DNA. Additionally, we have compared the ef- fects of a progestin agonist (R5020) and antagonist
(RU486) and the influence of HMG-1 on PR-mediated DNA bending.
RESULTS
Facilitated Binding of Purified PR to a PRE Contained in a Large DNA Fragment
Our earlier DNA-binding studies with purified human PR and the accessory factor HMG-1 were done with short PRE-containing oligonucleotides (47). Longer DNA fragments are required for bending studies be- cause the DNA molecule must be substantially larger than the segment that interacts with the protein (50). Therefore, it was necessary to establish conditions for binding of purified PR to longer PRE-containing DNA fragments and to determine whether HMG-1 would also enhance binding under these conditions. Be- cause HMG-1 binds DNA without any known se- quence specificity, we did not know whether it would facilitate PR binding to PREs in the presence of excess nonspecific sites. Both the A and B forms of PR, expressed as full-length proteins in a baculovirus in- sect cell system (51), were purified to apparent homo- geneity by methods previously described (47). As shown in Fig. lA, purified PR-B (lane 2) exhibited minimal binding to a 142-bp DNA fragment containing a single PRE (the DNA fragment was one of several circularly permutated probes used below). Addition of HMG-1 dramatically enhanced the formation of PR- DNA complexes (lane 4), which did not occur by the addition of ovalbumin (lane 3) or other nonspecific carrier proteins (not shown). The presence of both PR and HMG-1 in the enhanced complex was shown by supershift with a PR-specific monoclonal antibody (MAb) B-30 (lane 6) and with a MAb to HMG-1 (lane 7). This contrasts with the lack of supershift by an unre- lated MAb (lane 5). The specificity of the enhanced complex was demonstrated by competition with ex- cess unlabeled homologous DNA (lanes 8 and 9) as opposed to an unrelated DNA fragment that failed to reduce binding to the labeled probe (lanes IO and 11). HMG-1 alone at the same concentration used to en- hance PR-B binding did not bind to the DNA probe (lane 12). Similar results to those in Fig. 1A were ob- tained with purified PR-A (not shown).
In previous studies using a short PRE-containing oligonucleotide as the DNA probe, we showed by saturation binding analysis that purified PR alone does bind to PREs, but only at high concentrations. Addition of HMG-1 increased the apparent affinity allowing maximal binding at a rl O-fold lower concentration of PR (47). To establish conditions for maximal binding of purified PR and purified PR in the presence of HMG-1 to a PRE contained in the longer (142-bp) DNA frag- ment, we performed the saturation DNA-binding anal- ysis shown in Fig. 1B. Purified PR-B alone bound to the 142-bp DNA fragment in a dose-dependent man- ner but only at concentrations greater than 25 nM.
Dow
nloaded from https://academ
ic.oup.com/m
end/article/10/4/393/2713356 by guest on 09 July 2022

Progesterone Receptor-Induced DNA Bending 395
A
PR-DNA --c
Free DNA --) (142 w
B ioo-
k! EO-
n u 60- B
z :: 40-
3
8 20-
PR-E + HUG1
1 2 3 4 5 6 7 8 9 10 11 12
r PR-6 + HMG-1
PR-B Alone
%
P&B + oval.
”
0 20 40 60 80
nM PR-B
Fig. 1. The High Mobility Group Protein HMG-1 Enhances PR-B Binding to a 142- bp DNA Fragment Containing a Single PRE
A, PR-B expressed in Sf9 cells and bound to hormone (R5020) in viva was affinity purified to apparent homogeneity and incubated for 1 h at 4 C with 0.3 ng of an end-labeled 142-bp DNA fragment containing a centrally positioned PRE (Xhol fragment excised from pBend2-PRE). Lane 1 is free DNA. Lane 2 is purified PR-B alone (20 nM); the other lanes contain purified PR-B (20 nM) plus the following: lane 3, 250 ng ovalbumin; lane 4, 150 ng HMG-I; lanes 5-7, 150 ng HMG-1 plus 1 pg of an unrelated control MAb (cuER); the B-30 MAb to human PR-B @PR); and a MAb to HMG-1 (a HMG-I). Lanes 8 and 9 contain a 20-fold and 150-fold excess of unlabeled homologous DNA (sp); lanes IO and II, a 20-fold and 150-fold excess of unrelated DNA (n.s.). Lane 12 con- tains 150 ng HMG-1 alone. B, Affinity-purified PR-B was incubated with the same 3’P-labeled 142- bp PRE containing DNA fragment as above and submitted to nondenaturing gel electrophoresis. PR concentrations were varied as indicated against a constant amount of (0.3 ng) r’P]DNA, either in the absence of added protein or with the addition of 250 ng ovalbumin or 150 ng HMG-1. [32P]DNA complexes at each receptor concentration were quantified by Phosphorlmager analysis and plotted as percent upshifted PRE.
Also, the binding was not saturable over the range of PR concentrations used. Addition of HMG-1 produced a leftward shift of the curve, resulting in binding at much lower concentrations (2 nM) where PR exhibited little or no binding on its own (Fig. 1 B), and saturation was reached at = 20 nM. Unrelated proteins such as ovalbumin had no effect on the saturation binding curve (Fig. 1 B). Similar results were obtained with pu- rified PR-A (not shown). It should be noted that purified PR alone in Fig. IA was used at 20 nM where minimal binding is detected in the absence of HMG-1.
In subsequent DNA-bending experiments, purified PR alone was used at the highest concentration on the saturation binding curve (70 nM), and purified PR in the presence of HMG-1 was used at lower con- centrations (IO-20 nlvl). It should also be noted that human PR binds poorly, if at all, to PREs in the absence of ligand. Sequence-specific DNA binding is hormone-dependent (46). The nuclear accessory protein does not substitute for ligand; thus, efficient binding in vitro requires both ligand and HMG-1 (46, 47). Therefore, in all studies in this paper PR was bound to either the progestin agonist R5020 or the progestin antagonist RU486 in Sf9 insect cells be- fore extraction and purification. HMG-1 in Fig. 1 and in all subsequent experiments was purified to ap- parent homogeneity from calf-thymus as previously described (47). Thus we have used all purified com- ponents for reconstitution of a high affinity PR-DNA complex.
PR Mediates Distortion of Target DNA: PR-B Induces a larger Distortion than PR-A
To initially investigate the ability of PR to mediate a bend in target DNA, we performed circular permuta- tion electrophoretic mobility shift assays. This is a commonly used method for detecting protein-induced DNA bending that is based on the principle that a DNA fragment with a bend in the center will exhibit a slower electrophoretic mobility relative to the same DNA frag- ment with a bend positioned at either end (52). A set of DNA probes of identical length (142 bp) and sequence were constructed that contained a single PRE at dif- ferent positions relative to either end. These are shown schematically in Fig. 2A. Each 32P-labeled probe was incubated with a low concentration of purified PR in the presence of HMG-1 and submitted to gel-mobility shift assay. Representative autoradiographs of the gels are shown in Fig. 2B, and results from multiple independent experiments are summarized graphically in Fig. 2C. With both the A and B forms of PR, DNA complexes exhibited a slightly faster electrophoretic mobility when the PRE was positioned at either end of the DNA fragment (Fig. 2B, BarnHI and Bglll lanes), and the mobility became progressively slower as the PRE was moved toward the middle of the DNA frag- ment (Fig. 2B, Pvull lane). In contrast, the free r’P]DNA fragments showed little, if any, mobility vari- ation. Thus, the PRE position-dependent mobility vari-
Dow
nloaded from https://academ
ic.oup.com/m
end/article/10/4/393/2713356 by guest on 09 July 2022

MOL END0 1996 396
B
PR-A --)
Free DNA *
C 0.70
0.75 i
A
PR-A + HMG-1
PR-A + HMG-1
0.80
0.85
0.90
0.95
PR-A
1.05 Lb
-60-60-40-20 0 7.0 40 60 80
P&B + HMG-1
PR-B -
Free DNA -
PR-B + HMG-1 i AB Dimers + HMG-1
PR-B
1./ e
-80-60-40-20 0 20 40 60 80
Position of PRE
1 AB Dimers
-\;/li -80-60-40-20 0 20 40 60 80
VollONo.4
Fig. 2. PR-Mediated Distortion of Target DNA as Detected by Circular Permutation Assay A, Schematic of the circular permutated DNA fragments. Seven DNA fragments (142 bp) containing a single PRE oligonucle-
otide 5’-TCGACGGTACAAACTGTTCCC-3’ (stippled box) were prepared by restriction digestion of pBend2-PRE using Erg/II,
Dow
nloaded from https://academ
ic.oup.com/m
end/article/10/4/393/2713356 by guest on 09 July 2022
Progesterone Receptor-Induced DNA Bending 397
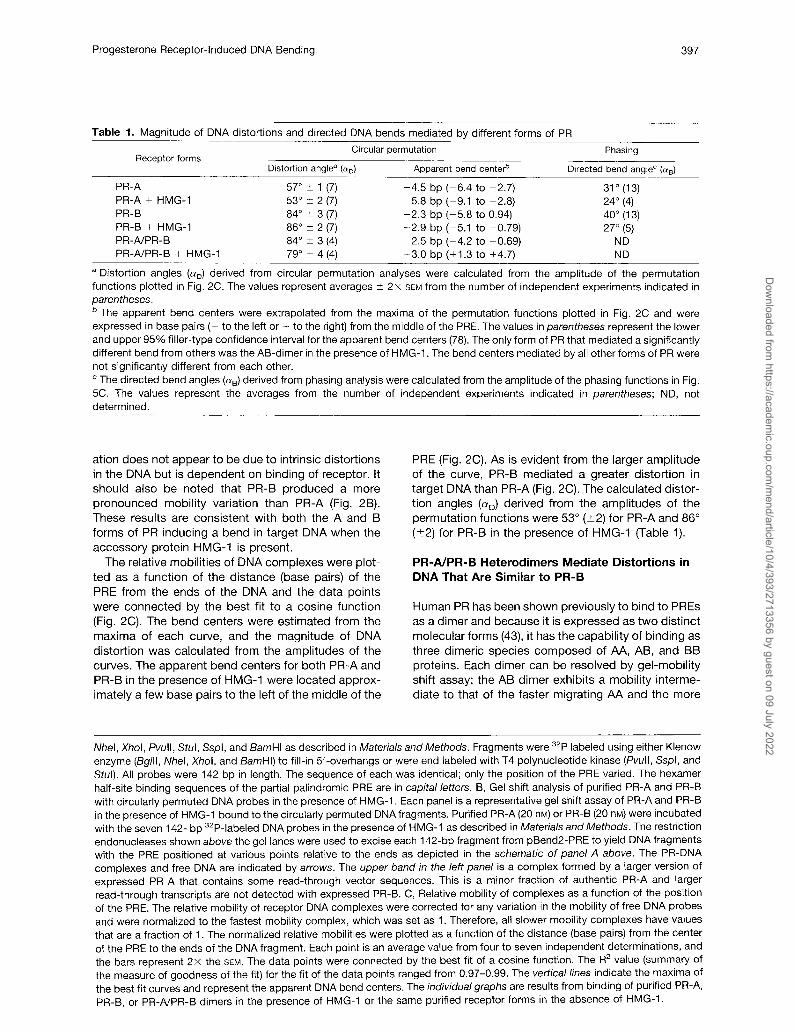
Table 1. Magnitude of DNA distortions and directed DNA bends mediated by different forms of PR
Receptor forms Circular permutation Phasing
Distortion angle* (Q) Apparent bend center” Directed bend angleC (+)
PR-A 57” i 1 (7) -4.5 bp (-6.4 to -2.7) 31” (13) PR-A + HMG-1 53” i 2 (7) -5.8 bp (-9.1 to -2.8) 24” (4) PR-B 84” 2 3 (7) -2.3 bp (-5.8 to 0.94) 40” (13) PR-B + HMG-1 86” 2 2 (7) -2.9 bp (-5.1 to -0.79) 27” (5) PR-A/PR-B 84” -+ 3 (4) -2.5 bp (-4.2 to -0.69) ND PR-A/PR-B + HMG-1 79” -c 4 (4) +3.0 bp (+1.3 to +4.7) ND
a Distortion angles (01~) derived from circular permutation analyses were calculated from the amplitude of the permutation functions plotted in Fig. 2C. The values represent averages 2 2x SEM from the number of independent experiments indicated in parentheses. ’ The apparent bend centers were extrapolated from the maxima of the permutation functions plotted in Fig. 2C and were expressed in base pairs (- to the left or + to the right) from the middle of the PRE. The values in parentheses represent the lower and upper 95% filler-type confidence interval for the apparent bend centers (78). The only form of PR that mediated a significantly different bend from others was the AB-dimer in the presence of HMG-1. The bend centers mediated by all other forms of PR were not significantly different from each other. ‘The directed bend angles (+) derived from phasing analysis were calculated from the amplitude of the phasing functions in Fig. 5C. The values represent the averages from the number of independent experiments indicated in parentheses; ND, not determined.
ation does not appear to be due to intrinsic distortions in the DNA but is dependent on binding of receptor. It should also be noted that PR-B produced a more pronounced mobility variation than PR-A (Fig. 28). These results are consistent with both the A and B forms of PR inducing a bend in target DNA when the accessory protein HMG-1 is present.
The relative mobilities of DNA complexes were plot- ted as a function of the distance (base pairs) of the PRE from the ends of the DNA and the data points were connected by the best fit to a cosine function (Fig. 2C). The bend centers were estimated from the maxima of each curve, and the magnitude of DNA distortion was calculated from the amplitudes of the curves. The apparent bend centers for both PR-A and PR-B in the presence of HMG-1 were located approx- imately a few base pairs to the left of the middle of the
PRE (Fig. 2C). As is evident from the larger amplitude of the curve, PR-B mediated a greater distortion in target DNA than PR-A (Fig. 2C). The calculated distor- tion angles (Q) derived from the amplitudes of the permutation functions were 53” (k2) for PR-A and 86” (52) for PR-B in the presence of HMG-1 (Table 1).
PR-A/PR-B Heterodimers Mediate Distortions in DNA That Are Similar to PR-B
Human PR has been shown previously to bind to PREs as a dimer and because it is expressed as two distinct molecular forms (43), it has the capability of binding as three dimeric species composed of AA, AB, and BB proteins. Each dimer can be resolved by gel-mobility shift assay; the AB dimer exhibits a mobility interme- diate to that of the faster migrating AA and the more
Nhel, Xhol, Pvull, Stul, Sspl, and BamHl as described in Materials and Methods. Fragments were 32P labeled using either Klenow enzyme @g/II, Nhel, Xhol, and BarnHI) to fill-in 5’-overhangs or were end labeled with T4 polynucleotide kinase (Pvull, Sspl, and Stul). All probes were 142 bp in length. The sequence of each was identical; only the position of the PRE varied. The hexamer half-site binding sequences of the partial palindromic PRE are in capita/ letters. B, Gel shift analysis of purified PR-A and PR-B with circularly permuted DNA probes in the presence of HMG-1. Each panel is a representative gel shift assay of PR-A and PR-B in the presence of HMG-1 bound to the circularly permuted DNAfragments. Purified PR-A (20 nM) or PR-B (20 nM) were incubated with the seven 142- bp 3’P-labeled DNA probes in the presence of HMG-1 as described in Materials and Methods. The restriction endonucleases shown above the gel lanes were used to excise each 142-bp fragment from pBend2-PRE to yield DNA fragments with the PRE positioned at various points relative to the ends as depicted in the schematic of pane/ A above. The PR-DNA complexes and free DNA are indicated by arrows. The upper band in the left pane/ is a complex formed by a larger version of expressed PR-A that contains some read-through vector sequences. This is a minor fraction of authentic PR-A and larger read-through transcripts are not detected with expressed PR-B. C, Relative mobility of complexes as a function of the position of the PRE. The relative mobility of receptor DNA complexes were corrected for any variation in the mobility of free DNA probes and were normalized to the fastest mobility complex, which was set as 1. Therefore, all slower mobility complexes have values that are a fraction of 1. The normalized relative mobilities were plotted as a function of the distance (base pairs) from the center of the PRE to the ends of the DNA fragment. Each point is an average value from four to seven independent determinations, and the bars represent 2x the SEM. The data points were connected by the best fit of a cosine function. The R” value (summary of the measure of goodness of the fit) for the fit of the data points ranged from 0.97-0.99. The vertical lines indicate the maxima of the best fit curves and represent the apparent DNA bend centers. The individual graphs are results from binding of purified PR-A, PR-B, or PR-A/PR-B dimers in the presence of HMG-1 or the same purified receptor forms in the absence of HMG-1.
Dow
nloaded from https://academ
ic.oup.com/m
end/article/10/4/393/2713356 by guest on 09 July 2022
MOL END0 1996 398
VoliONo.4
slowly migrating BB dimers (53). Because the distor- tion angle mediated by PR-B was greater than that of PR-A, we asked whether the AB dimer would alter DNA structure in a distinct manner, or assume the property of one or the other form of PR. Circular per- mutation assays were performed in the same manner as Fig. 2 except that separately purified PR-A and PR-B, at low concentrations in the presence of the accessory factor HMG-1, were mixed before the gel- shift assay. Three distinct DNA complexes were de- tected that correspond to previously identified AA, AB, and BB dimers (Fig. 3). Additionally, a PRE position- dependent mobility variation was observed with all three complexes indicating that AB dimers also medi- ate distortion of target DNA.
The relative mobilities of the AB dimers complexed to the different DNA probes were plotted as a function of the position of the PRE, and the data points were connected by the best fit to a cosine function. From the amplitude of the curves, it is evident that the de- gree of DNA distortion mediated by AB dimers was more similar to that mediated by PR-B than PR-A (Fig. 2C). The calculated distortion angle mediated by AB- dimers in the presence of HMG-1 was 79” (+4) (Table 1). The apparent bend center, however, was different from that mediated by either PR-A or PR-B. As extrap-
PM -t PR-A/PR-B +
PR-A +
Free DNA -
PR-A/PR-B + HMG-1
Fig. 3. Gel Shift Analysis of AB Receptor Heterodimers Bound to Circularly Permuted DNA Probes
PR-A and PR-B were purified separately and mixed before addition of the DNA- binding reaction. The mixed receptors, at a final concentration of 30 nM, were incubated with the seven circularly permutated DNA probes as in Fig. 2 in the presence of 150 ng HMG-1. The three different dimeric forms of PR bound to DNA are indicated by the arrows (PR-B, PR-A/PR-B, PR-A).
olated from the curve maximum (Fig. 2C), the bend was located slightly to the right of the middle of the PRE (+3 bp). In contrast, the apparent bend centers for all other forms of PR were located between -2 bp to -4 bp to the left of the middle of the PRE. Thus the magnitude of DNA distortion mediated by AB dimers was closer to that of PR-B than PR-A whereas the apparent bend center was unique.
PR Has an Intrinsic Ability to Distort the Structure of Target DNA
All the studies described to this point were done with purified PR in the presence of HMG-1. To determine whether HMG-1 cooperates with PR to distort DNA or whether PR has an intrinsic ability to alter DNA con- formation, circular permutation assays were per- formed with purified PR in the absence of HMG-1. In these experiments, receptor was used at a higher con- centration (70 nM) where some binding occurs in the absence of the accessory factor (see Fig. 1B). The results of the circular permutation analysis are sum- marized graphically in Fig. 2C and quantitatively in Table 1. As determined from the amplitudes of the permutation functions (Fig. 2C), all forms of PR medi- ated a similar magnitude of DNA distortion in the ab- sence and presence of accessory factor. Calculated distortion angles in the absence of HMG-1 were 57” (+ 1) for PR-A, 84” (?3) for PR-B, and 84” (? 3) for AB-dimers. Thus, purified PR alone, when present at a high concentration, has an intrinsic ability to distort the structure of target DNA with approximately the same magnitude as low concentrations of PR in the pres- ence of the accessory protein HMG-1. The apparent bend centers extrapolated from the curve maxima (Fig. 2C) for PR-A (-4.5 bp) and PR-B (-2.3 bp) in the absence of HMG-1 were also indistinguishable from the values obtained in the presence of HMG-1 (Table 1). However, HMG-1 did appear to alter the center of DNA bending mediated by AB-dimers, shifting it from -2.5 bp to +3.0 bp from the middle of the PRE.
The Progestin Antagonist RU466 Does Not Alter the Ability of PR to Distort DNA Structure
In previous studies we and others have shown that specific PR-PRE complexes in standard gel-mobility shift assays migrate discernibly faster when receptor is bound to the antagonist RU486 as compared with progestin agonists (53-55). This difference in mobility was interpreted to be due to ligand stabilization of distinct conformations of the receptor protein as op- posed to an alteration in the conformation of the target DNA. These earlier studies were done with PR in crude cell extracts so it was also not possible to determine whether the altered mobility was due to the presence or absence of another protein. To resolve these ques- tions, we have purified PR bound to RU486 to appar- ent homogeneity and have compared electrophoretic mobilities of DNA complexes on standard gel-shift assays with that of purified PR bound to R5020. Both
Dow
nloaded from https://academ
ic.oup.com/m
end/article/10/4/393/2713356 by guest on 09 July 2022

Progesterone Receptor-induced DNA Bending 399
A
PR-DNA -)-
Complex
Free DNA --t
PR-B
PR-B l&-l HH
B 0.70. PR-A RU486
0.75
0.80
0.85
0.90
h 0.95
2
"0
1.00 w
1.05
E 1.10 a2
.z 2
0.70
z 0.75
d 0.80
0.85
0.90
0.95
1 .oo
1.05
1.101 .‘ -80-60-40-20 0 20 40 60 80
Position of PRE Fig. 4. The Progestin Antagonist RU486 Does Not Alter PR- Mediated Distortion of Target DNA
A, Purified PR-B bound to RU486 exhibits altered electro- phontic mobility when complexed to target DNA in standard gel mobility shift assays. PR-B expressed in Sf9 cells was bound in viva to either the progestin R5020 or the progestin antagonist RU486 and was affinity purified to apparent ho- mogeneity. Each receptor was incubated at high concentra- tion (70 nM) in the absence of HMG-1 or at a low concentra
forms of purified PR were used at high concentrations (70 nM) where accessory factor is not required for DNA binding, and the DNA was a 142-bp circular permuta- tion probe with a single PRE positioned near the center of the DNA fragment. DNA complexes formed by pu- rified PR bound to RU486 displayed a faster mobility than DNA complexes formed by purified PR bound to R5020 (Fig. 4A, left panel). Similar differences in elec- trophoretic mobility were observed with low concen- trations of purified PR in the presence of the accessory factor HMG-1 (Fig. 4A, right panel).
Purified PR-A and PR-B bound to RU486 were also analyzed by circular permutation assay using the same set of DNA probes that were used with purified PR bound to R5020. Relative mobilities of the DNA complexes were plotted as a function of the position of the PRE, and the data points were best fitted to a cosine function (Fig. 4B). As evident from the ampli- tudes and maxima of the curves, both forms of PR (PR-A and PR-B), when bound to RU486, induced distortions in DNA with magnitudes and apparent bend centers that were similar to that mediated by the same receptors bound to R5020 (compare Figs. 2C and Fig. 4B). Distortion angles and apparent bend centers calculated from the permutation functions in Fig. 4B are given in Table 2. These were not measur- ably different from the values calculated for the differ- ent forms of PR bound to agonist (compare Tables 1 and 2). These results indicate that the differential elec- trophoretic mobility of PR bound to RU486 and R5020 on standard gel-mobility shift assays is not due to the presence of another protein, or to differences in bend- ing of target DNA, but rather reflects distinct confor- mations of the receptor. Thus, differential PR-medi- ated DNA bending does not appear to contribute to the antagonist action of RU486.
PR-A and PR-B Mediate Directional Bends Toward the Major Groove of the DNA Helix as Determined by Phasing Analysis
Although the circular permutation assay is widely used to detect protein-mediated distortions in DNA, it has
tion (20 nM) in the presence of HMG-1 (150 ng) with a 32P- labeled 142-bp DNA fragment containing a single PRE (Xhol fragment of pBend2-PRE). PR-DNA complexes were electro- phoresed on 6% polyacrylamide gels, dried, and autoradio- graphed. The additional band at the top of the gel in the right most lane is an HMG-1 protein aggregate that does not enter the gel. This is occasionally seen with enhanced PR-DNA complexes and is not related to RU486. B, Results of circular permutation analysis with purified PR-A and PR-B bound to RU486. The relative mobilities of PR-DNA complexes were plotted as a function of the position of the PRE, and the data points were connected by the best fit of a cosine function. Each point is an average value from three independent de- terminations, and the bars represent 2X SEM. The R2 value for the fit of the data points ranged from 0.97-0.99. The vertical line indicates the maxima of the best fit curves and repre- sents the apparent bend centers.
Dow
nloaded from https://academ
ic.oup.com/m
end/article/10/4/393/2713356 by guest on 09 July 2022

MOL END0 . 1996 VollONo.4 400
Table 2. DNA distortion mediated by PR bound to RU486
Circular permutation Receptor form Distortion angle”
(4 Apparent bend centet”
PR-A 55” i 2 (3) -5.2 bp (-7.2 to -4.6) PI?-A + HMG-1 55” i 3 (3) -5.9 bp (-7.7 to -2.8) PR-B 90” i 4 (3) -1.4 bp (-2.2 to -0.51) PR-B + HMG-1 90” 2 4 (3) -1.7 bp (-4.0 to 0.52)
a Distortion angles (01~) derived from circular permutation analysis were calculated from the amplitude of the circular permutation functions plotted in Fig. 4B (for PR-A and PR-B) and for PR-A and PR-B plus HMG-1 (not shown). The values represent averages from three independent experiments -t 2X SEM.
b The apparent bend centers were extrapolated from the maxima of the circular permutation functions in Fig. 4B and were expressed in base pairs from the middle of the PRE. The values in parentheses represent the lower and upper 95% filler-type confidence interval (78) for the apparent bend center.
limitations in that mobility variations dependent upon the position of the protein-binding site may not always reflect a directed bend (change in direction of the DNA helix). Also, the method does not detect the orientation (toward the minor or major groove of the DNA helix) of the bend (11,56). Therefore, we have performed phas- ing analysis to determine whether PR induces a di- rected bend and to determine the orientation. A set of DNA probes of similar length and sequence were pre- pared that contain an intrinsic sequence-directed bend (produced by three phased dA:T, tracts), which was separated from a single PRE by variable length spacers. The distances separating the intrinsic DNA bend and the PRE were varied incrementally over ap- proximately one and a half turns of the DNA helix. This resulted in variably placing the receptor-binding site and the dA:T, tracts on the same and opposite faces of the DNA helix. I f PR were to introduce a directional bend, the electrophoretic mobility of the DNA com- plexes would be expected to vary dependent on the spacing between the dA:T, tracts and the PRE. Be- cause phased dA:T, tracts bend DNA toward the mi- nor groove, the pattern of the mobility variation is diagnostic of the orientation of the induced bend (56).
Seven DNA fragments were prepared that contained a PRE separated from the intrinsic bend by variable length spacers ranging from 23-35 bp (Fig. 5A, top schematic). Each of the probes was end labeled and incubated with purified A or B receptors, and free and bound DNA complexes were separated by nondena- turing gel electrophoresis. A representative phasing gel with purified PR-A shows that DNA complexes exhibited a mobility variation dependent on the spac- ing between the PRE and the intrinsic bend (Fig. 5A, bottom panel). It should be noted that the mobility of the free DNA probes also varied as a function of the positioning of the intrinsic bend. However, the mobility variation of the PR-DNA complexes followed a differ-
ent pattern than that of the free DNA probes (Fig. 5A). A similar pattern of mobility variation as a function of the spacing between the dA:T, tracts and the PRE was also observed with PR-B (not shown). After normaliza- tion to variations in the mobility of free probes, the relative mobilities of PR-DNA complexes were plotted as a function of the spacer length, and the data points from multiple independent determinations were plot- ted as a best fit to a cosine function (Fig. 5B). For both A and B receptors, the slowest mobility occurred when the centers of the dA:T, tracts and PRE were sepa- rated by 27 bp or = 2 l/2 turns of the DNA helix. The fastest mobility complexes were observed when the dA:T, tracts and PRE were separated by 32 bp, or m 3 full turns of the DNA helix. This pattern of mobility variation is consistent with PR-A and PR-B mediating bends toward the major groove of the DNA helix (56). Directed bend angles ((Y& calculated from the ampli- tude of the plotted phasing functions from Fig. 5B were 31” for PR-A and 40” for PR-B (Table 1).
In the presence of HMG-1, a similar pattern of mo- bility variation as a function of spacer length was ob- served for both the A and B receptors (Fig. 5B). Thus, HMG-1 neither altered the orientation of PR-mediated DNA bending nor did it have a major effect on the magnitude of the directed bend (Le. amplitude of the curves in Fig. 5B). However, in the presence of HMG-1, there was a modest but measurable reduction in the calculated directed angles mediated by PR-A (reduction from 31” to 24”) and PR-B (reduction from 40” to 27”) (Table 1).
HMG-1 binds to DNA with no known sequence specificity but prefers to interact with bends or kinks in DNA. Furthermore, at high concentrations HMG-1 has been shown to bend DNA (47-49). To ascertain whether the small decrease in the amplitude of the phasing functions in the presence of HMG-1 (Fig. 5B) was due to HMG-1 binding to the intrinsic dA:T, tract and itself affecting DNA structure, we have determined whether HMG-1 alone can bind to the phasing probes. As shown in Fig. 5C, purified HMG-1 did bind to a small extent, producing two upshifted complexes (in- dicated by arrows with slightly reduced mobility as compared with the free DNA probe). In contrast, HMG-1 alone did not form a stable complex with the circular permutation probes (Fig. 1) nor in our previous studies with PRE oligonucleotides (47). This differen- tial ability to bind to phasing, as opposed to other DNA probes, suggests that HMG-1 can form a stable com- plex with the intrinsic DNA bend site. It should also be noted that HMG-1 in the presence of PR does not form a separate binary HMG-l/DNA complex with the phasing probes (Fig. 5A). With both sets of probes (permutation and phasing) HMG-1 formed a ternary complex with PR and DNA as determined by super- shift with antibodies (Fig. IA for permutation probes and not shown for phasing probes). These results suggest that HMG-1 has a minimal effect on the di- rected bend angle mediated by PR and that the re-
duced amplitude of the phasing function was most
Dow
nloaded from https://academ
ic.oup.com/m
end/article/10/4/393/2713356 by guest on 09 July 2022

Progesterone Receptor-Induced DNA Bending 401
A Phasing Analysis
PR - B + HMG-1
PR - A + HMG-1
likely due to an independent effect of HMG-1 on the structure of the phasing probes.
DISCUSSION
We have shown by circular permutation and phasing analysis that human PR can mediate a directed bend upon binding to a PRE that is oriented toward the major groove of the DNA helix. Several other steroid receptors have been observed to mediate bends in target DNA, including the thyroid hormone receptor (TR) (37, 38) ER (31, 39-41), and RXR either as a homodimer or heterodimerized to various protein part- ners (42). These reports, taken together with the re- sults of the present study with PR, suggest that DNA bending may be a general property of the steroid receptor family. Other DNA-bending studies were per- formed with relatively crude preparations of receptors taken as cell extracts or as synthesized protein in rabbit reticulocyte lysates. The present study with full-
Fig. 5. PR-mediated Directional DNA Bending Detected by Phasing Analysis
A, Schematic of the DNA probes used for phasing analysis and results with purified PR-A. Each DNAfragment contained a single PRE separated by a variable length spacer (23-35 bp) from an intrinsic DNA bend directed by three in-phased dA:T, tracts. The centers of the intrinsic bend and PRE were sep- arated by 23, 25, 27, 29, 31, 33, and 35 bp. The seven phasing probes were end-labeled with 32P by Klenow fill-in and incubated with a high concentration (70 nM) of purified PR-A and electrophoresed as in Fig. 2 with the minor mod- ifications described in Materials and Methods. Free DNA and PR-A-DNA complexes are indicated by arrows. The numbers above each lane indicate the distance in base pairs from the center of the dA:T, tract to the center of the PRE. It should be noted that the free DNA probes exhibit mobility variations dependent on the position of the intrinsic bend. This variation is dissimilar to the mobility variation of the PR-DNA com- plexes. B, Summary of phasing analyses. The relative mobil- ities of PR-DNA complexes corrected for variation in mobil- ities of the free DNA were normalized to the average mobility of all complexes as described in Materials and Methods. The normalized relative mobilities were then plotted as a function of the spacer length (base pairs) between the center of the intrinsic bend and the PRE. The data points were connected by the best fit of a cosine function. The values represent an average of four to 13 independent experiments. The individ- ual graphs are results from binding of purified PR-B alone (70 nM), purified PR-B (20 nM) plus HMG-1, purified PR-A alone (70 nM), and purified PR-A (20 nM) plus HMG-1. The R’ values for the fit of the data to a cosine function were 0.752 for PR-B, 0.640 for PR-B + HMG-1,0.704 for PR-A, and 0.757 for PR-A + HMG-1. C, Binding of HMG-1 to phasing probes. Purified HMG-1 at the same concentration used to enhance PR bind- ing was incubated alone with the seven 32P-labeled phasing DNA probes and submitted to electrophoretic gel mobility assays under the same conditions as those used for PR-DNA binding. The arrows indicate positions of free DNA and two HMG-l/DNA complexes which likely represent single and multiple molecules of HMG-1 bound per DNA fragment.
Dow
nloaded from https://academ
ic.oup.com/m
end/article/10/4/393/2713356 by guest on 09 July 2022

MOL END0 . 1996 VollONo.4 402
length human PR purified to apparent homogeneity demonstrates that DNA bending is an intrinsic prop- erty of steroid receptors that does not require other proteins.
The orientation and location of the DNA bends me- diated by the two molecular forms of PR were found to be indistinguishable. As detected by phasing analysis, both PR-A and PR-B induced bends toward the major groove of the DNA helix. As derived from circular per- mutation analysis, the bend was located within the PRE but was not symmetrically positioned with re- spect to the hexamer core half-sites. The bend centers were located several base pairs (-4 bp for PR-A and -2 bp for PR-B) from the midpoint between the two PRE half-sites. This small difference in the location of the bend centers calculated for PR-A and PR-B was not within the limit of resolution of the assay. In con- trast, the magnitudes of the bends mediated by PR-A and PR-B were substantially different. This differential was detected both by circular permutation and phas- ing analysis, and in both cases PR-B induced a larger bend angle than PR-A. We do not believe that the differential magnitude of DNA bending mediated by A and B receptors is due simply to the different sizes of the proteins. No relationship has been reported be- tween the size of a protein and its ability to bend DNA. In fact, small molecular weight proteins have been observed to produce some of the sharpest bends in DNA. For example, no relationship was observed be- tween molecular mass and the degree of DNA bending mediated by numerous members of the bZlP family of proteins (32). Additionally, studies with DNA-binding domain fragments and truncation mutants have indi- cated that sequence regions outside of the DNA- binding domains contribute to DNA bending (11, 32). Thus, the larger degree of DNA bending mediated by PR-B is likely due to unique structural properties con-
7 ferred by the extended N-terminal sequences. Kerppola and Curran (32) proposed an independent
DNA-bending model for protein heterodimers where the overall bending property is the sum of the inde- pendent bends induced by each dimer subunit. This was best illustrated by certain bZlP heterodimers (32) and Myc/Max heterodimers (14) that fail to generate a significant directed DNA bend when the heterodimer is formed by subunits that induce off-setting DNA bends with opposite orientations. It is not clear from our analysis of AB dimers whether the independent DNA- bending model applies to PR. By circular permutation analysis the magnitude of DNA bending mediated by AB dimers and the apparent bend center were indis- tinguishable from BB dimers and were not intermedi- ate between AA and BB dimers. However, we did not determine by phasing analysis the orientation or mag- nitude of the directed bend angle induced by AB dimers. The orientation is not likely to be different from that induced by either PR-A or PR-B since A and B homodimers both bend DNA toward the major groove.
The distortion angles determined by circular permu- tation assay for PR-B ((up = 84”) and PR-A ((ho = 57”)
were larger than the directed bend angles for PR-B ((Ye = 40”) and PR-A ((Ye = 31”) determined by phasing assay. Similar quantitative discrepancies have been reported for several other sequence- specific tran- scription factors, as well as RXR-TR complexes (10, 32, 42, 57, 58). Similar to results with human PR re- ported here, RXR-TR complexes mediated distortion angles derived from circular permutation analysis that, dependent upon the sequence of the binding site, ranged from 58” to 84”. In contrast directed bend angles measured by phasing assay ranged from 22” to 31” (42). The reason for the apparent discrepancies between the results of circular permutation and phas- ing analysis is not clear. It has been suggested that the observed differences are due to the fact that the cir- cular permutation assay is sensitive to localized flexi- bility in the target DNA (10, 32) or to unusual structure of a protein as has been reported for GCN4 (57). Indeed, points of increased flexibility have been dem- onstrated to influence the electrophoretic probility of DNA whereas localized flexibility, as compared with a fixed bend, is not phase-sensitive (59). Thus we be- lieve that the apparent bend angles determined by circular permutation analysis are likely to be an over- estimate and that the true magnitudes of the directed bends are closer to the values obtained by phasing analysis. It is important to note that the differential bending mediated by A and B receptors was detected by both assay methods.
One reason for studying PR-induced DNA bending was prompted by our previous results showing that the HMG-1 protein, which itself can bend DNA, dra- matically enhanced the binding affinity of PR to PREs (47). We questioned whether a stable PR-DNA com- plex might require structural alterations in target DNA and that receptor and HMG-1 cooperated to induce the change. However, as determined by circular per- mutation and phasing analyses, we observed little dif- ference in the magnitude, orientation, or apparent bend centers mediated by PR in the presence and absence of HMG-1. The only significant influence of HMG-1 on PR-mediated DNA bending was an alter- ation in the apparent bend center mediated by AB dimers; shifting it from -2 bp from the middle of the PRE in its absence to +3 bp in its presence. This suggests that HMG-1 may alter the spatial configura- tion of AB-dimer/DNA complexes. A nuclear acces- sory factor that enhances sequence- specific binding of TR was also observed to influence the center of DNA bending mediated by TR (38). However, the TR nuclear accessory factor differs mechanistically from HMG-1 in that it heterodimnerizes with TR, and with certain target DNA sequences it had a much greater effect on the bend center, shifting its location as much as 17 bp (38).
Although the mechanism by which HMG-1 en- hances sequence-specific DNA binding of PR re- mains unresolved, the known properties of HMG-1 taken together with the results of this paper are consistent with an induced-fit model presented
Dow
nloaded from https://academ
ic.oup.com/m
end/article/10/4/393/2713356 by guest on 09 July 2022

Progesterone Receptor-Induced DNA Bending 403
schematically in Fig. 6A. HMG-1 is thought to func- tion as a general DNA chaperone by manipulating the structure of DNA during transactions that require transient flexibility of DNA such as replication, re- combination, repair, and transcription (62). We pro- pose that a stable receptor-DNA complex requires a combination of specific nucleotide recognition se- quence and altered DNA structure and that PR in- duces the change in DNA conformation. HMG-1 is envisioned to enhance PR binding by stabilizing the receptor-mediated alteration in DNA conformation which is energetically unfavorable but required for assembly of a stable PR-DNA complex. This model is further supported by studies with ER where intro- duction of tertiary structure in a single-stranded es- trogen response element by denaturation/renatur-
ation, dramatically increased the binding affinity of ER (63, 64).
The functional role for DNA bending by eukaryotic transcriptional activators is not well defined. It has been speculated that DNA bending may be coupled to transcriptional regulation by facilitating protein-protein interactions between upstream sequence-specific
A) Induced Fit
@@
Stabilization of
DNA Bending
B) Transcriptional Regulation
HMG-1 HMG-1 HMG-1 HMG-I
Fig. 6. Proposed Functional Roles of P&Mediated DNA Bending
A, Induced fit model for assembly of a stable PR-DNA complex. We propose that high affinity receptor binding to DNA requires both specific nucleotide recognition sequence and an altered conformation of DNA that is induced by the receptor. HMG-1 enhances the binding of PR to PREs by stabilizing receptor-mediated bending at the PRE which is an energetically unfavored conformation. B, Transcriptional reg- ulation. DNA bending may be coupled to transcriptional reg- ulation as a means to facilitate PR interaction with coactiva- tors or other factors in the basal transcriptional apparatus. This is supported by the finding that PR-B, which is a gen- erally stronger transcriptional activator than PR-A, mediates a larger bend in target DNA than PR-A.
DNA-binding proteins and other factors in the basal transcriptional apparatus. There is no direct functional evidence for this as yet. Data suggesting such a mech- anism are correlative. For example, the numerous het- erodimers that form between members of the bZlP family of proteins are capable of inducing distinct con- formations in the same DNA element (32). This differ- ential DNA bending has been suggested to contribute to specific transcriptional responses mediated by re- lated proteins through the same DNA element by cre- ating different spatial orientations. Studies with TR have observed a correlation between the degree of distortion of different mutant TRE elements and func- tional responses (37, 38), and the degree of bending mediated by TR and various RXR complexes was found to be dependent on the sequence of the recep- tor-binding site (42). Additionally, the magnitude of DNA bending was found to correlate with the strength of transcriptional activation when the DNA-binding do- main fragment of ER was compared with full-length ER (39, 40) and the A and B forms of human PR were analyzed (this paper). However, in all studies reported to date, ligand was found to have no influence on steroid receptor-mediated DNA bending (37-42), and no difference was detected either for ER (31) or PR (this paper) bound to agonist or antagonist. This sug- gests that DNA bending is involved in receptor-medi- ated transcription but is not sufficient. It is well docu- mented that ligand-induced conformational changes in the receptor are important for activation (65). Thus, the combined effects of DNA bending and conforma- tional changes in protein structure are likely required for the correct spatial orientation between receptors and other factors to allow a productive interaction with the transcriptional apparatus. More direct studies will be needed to define the physiological role for steroid receptor-induced DNA bending.
An interesting property of human PR is the ex- pression of two molecular forms from the same gene that exhibit distinct functional properties. Full- length PR-B is generally found to be a stronger transcriptional activator than PR-A (44, 45). Addi- tionally, in cell and promoter contexts where PR-A is a weak or nonfunctional transcriptional activator, it can repress the activity of several other steroid re- ceptors (44, 45, 66). A transcriptional activation do- main located in the N-terminal segment unique to PR-B has been defined that appears to synergize with other transcriptional activation domains and has been suggested to be responsible for the unique functional properties of PR-B (67). The results of the present study raise the possibility that differential DNA bending may also contribute to functional re- sponses unique to the two receptor forms. PR-B is generally a stronger transcriptional activator than PR-A and was found in this study to mediate a greater degree of bending at the same PRE than PR-A. How DNA bending may be involved is shown schematically in Fig. 6B. We propose that the same PRE bound to PR-A or PR-B assumes different spa-
Dow
nloaded from https://academ
ic.oup.com/m
end/article/10/4/393/2713356 by guest on 09 July 2022

MOL END0 1996 VollONo.4 404
tial conformations resulting in receptor contacting
other proteins more efficiently (or contacting differ-
ent proteins) to direct the assembly of a transcrip-
tional unit that mediates distinct functional re-
sponses. HMG-1 may also be involved in
transcriptional regulation by stabilizing the confor-
mation of the transcription unit (Fig. 6B). Indeed
there is growing evidence that HMG-1 and the highly
related HMG-2 protein are involved in mediating the
transcriptional activity of other sequence-specific
DNA-binding proteins. HMG-l/2 have been shown
to enhance the DNA-binding and transcription ac-
tivity of a factor required for transcription of the adenovirus major late promoter (60) and the octamer
transcription factors OCT-1 and OCT-2 (61). Addi-
tionally, HMG-l/2 have been shown to act as re-
pressors of basal transcription and to be required for
activated transcription in cell-free systems com-
posed of RNA polymerase II and defined purified
basal transcription factors (68-70).
MATERIALS AND METHODS
Baculovirus Expression of Recombinant Human PR and Purification
Construction of recombinant baculovirus expressing either full-length A (94 kDa) or B (120 kDa) forms of human PR and their functional characteristics have been previously de- scribed (51). Briefly, Spodoptera frugiperda (Sf9) insect cells were plated in spinner vessels at a density of 1 Oe cells/ml in Graces insect cell medium supplemented with 10% FBS. Recombinant baculovirus was added at a multiplicity of in- fection of 2.0, and cells were allowed to incubate for 48 h at 27 C. Receptors were bound to hormone in viva by incubat- ing infected Sf9 cells at 27 C for the last 4-6 h of infection with 100 nM of the synthetic progestin R5020. Recombinant re- ceptors were prepared by lysing infected Sf9 cells in TEDG buffer [lo mM Tris base (pH 7.4), 1 mM EDTA, 1 mM dithio- threitol, 10% glycerol] containing a cocktail of protease in- hibitors (71) and 0.5 M NaCI. Samples were then centrifuged at 105,000 x g for 30 min to yield a soluble whole-cell extract (WCE). WCEs were dialyzed at 4 C overnight against TEDG to reduce salt concentrations before DNA-binding assays. The receptor concentration in WCEs was estimated by steroid binding and immunodot blot assays, which have been pre- viously described (47).
Separately expressed PR-A and PR-B were purified from WCEs by Mab affinity chromatography as previously de- scribed (47). To purify PR-A we used the AB-52 MAb that recognizes both the A and B isoform of human PR (71). TO purify PR-B, we used the B-30 MAb that recognizes only PR-B (71). Affinity resins were constructed as previously de- scribed (47). PR bound to affinity resins was eluted in alkaline pH buffer, followed by immediate neutralization and addition of excess zinc chloride (50 pm). This single-step procedure results in purification of PR to apparent homogeneity as judged by silver-stained SDS gels and Western blotting to confirm the identity of the protein band (47). Additionally, this procedure yields biologically active PR product, which re- tains the majority of hormone-binding and DNA-binding ac- tivity, albeit the latter requires the addition of HMG-1. Purified PR was frozen at -70 C and retained its biological activity for up to 3 months if not repeatedly freeze-thawed.
Purification of HMG-1 and Production of Monoclonal Antibodies
Purification of HMG-1 has been previously described (47) and is a modification of the method of Adachi et al. (72). Briefly, calf thymus was homogenized, and the acid-soluble chro- mosomal proteins were extracted with 5% perchloric acid and precipitated with cold 25% trichloroacetic acid. The pre- cipitate was pelleted by centrifugation at 10,000 rpm for 20 min, and the pellet was dissolved in cold water. The sample was then dialyzed against 10 mM Tris-HCI (pH 7.8) overnight at 4 C. The dialysate was applied directly to a PBE94 poly- buffer-exchange resin (Pharmacia-LKB, Piscataway, NJ) used under anion-exchange conditions. The column was washed with 10 mtv Tris-HCI (pH 7.8), and bound proteins were eluted with a O-l 5 M NaCl gradient. Purified calf thymus HMG-1 was used to immunize Balb/c mice for the production of monoclonals antibodies by methods previously described (71). Two hybridomas (clones 854/ElO and 990/Dll) were isolated that produce mouse IgM antibodies that recognize HMG-1 by Western blot. These MAbs were used to supershift HMG-l/PR/DNA ternary complexes detected by electro- phoretic mobility shift assay.
Plasmids and Preparation of DNA Probes
For generation of 142-bp DNA fragments containing a per- muted PRE, a 21-bp double-stranded synthetic oligonucle- otide 5’-TCGACGGTACAAACTGTTCCC-3’ was cloned into the unique SalI site of pBend2 plasmid, yielding pBend2- PRE. The parent pBend2 vector was provided by Sankar Adhya (73).‘Oligonucleotide cloning was accomplished by the linker tailing method described by Lathe et a/. (74). The oli- gonucleotide contained a 15-bp partial palindromic se- quence determined by Leiberman et al. (75) to be an optimal progesterone response element (PRE). DNA fragments of the same length (142 bp) with the PRE located at different posi- tions relative to the DNA ends were produced by digestion of oBend2-PRE with BarnHI. Ssol, Stul, Pvull, Xhol, Nhel, and bg/ll. The resulting DNA fragments were purified from 2% agarose gels by Mermaid Kit (Bio 101) as recommended by the supplier. DNA fragments that generated blunt ends were end-labeled with -#*P]ATP and T4 polynucleotide kinase (Pvull, Stul, Sspl), and those that generated 5’-overhangs were end-labeled by Klenow DNA polymerase fill-in with a[32P]dATP and @*P]dCTP. Average probe-specific radio- activity ranged from 75,000300,000 cpm/ng, and the total amount of probe used per assay ranged from 0.3-l .O ng.
Phasing plasmids pTK401-26 and pTK401-28, as de- scribed by Kerppola and Curran (lo), contain unique Xbal and SalI endonuclease restriction sites bordering an AP-l- binding sequence positioned either 26 or 28 bp from the center of an intrinsic DNA bend sequence (3 in phase dA:T, tracts). The AP-l-containing fragment was excised and re- placed with a series of oligonucleotides containing a single PRE that were engineered to have compatible XballSall sites at either end for cloning into pTK401-26 or pTK401-28. The double-stranded oligonucleotide 5’-TCGACCTGAGAACA- AACTGTTCTTAAAACGAG-3’ was cloned into pTK401-26 to yield the phasing probe with a 31-bp spacer between the center of the PRE and the dA:T, tract; 5’-TCGAAGAA- CAAACTGTTCTTAAAACGAT-3’ was cloned into pTK401-26 or pTK401-28 to yield phasing probes, respectively, with 27- and 29-bp spacers, and 5’-TCGACCTITGAGAACAAACTGT- TCTTAAAAC-GAG-S’ was cloned into pTK401-26 or pTK401-28 to yield phasing probes, respectively, with 33- and 35-bp spacers. To generate phasing probes with 23- and 25-bp spacers, plasmids pTK401-26 and pTK401-28 were digested with Sall, treated with SI nuclease, and digested with Xbal. This generated a blunt end/Xhal site for cloning the oligonucleotide 5’-AGAACAAACTGTTCTTAAAACGAT- 3’. This overall cloning strategy generated a set of plasmids
Dow
nloaded from https://academ
ic.oup.com/m
end/article/10/4/393/2713356 by guest on 09 July 2022

Progesterone Receptor-Induced DNA Bending 405
in which the center of the intrinsic bend sequence was sep- arated by 23, 25, 27, 29, 31, 33, and 35 bp from the center of the PRE. Insertion of the oligonucleotide and spacing was confirmed by DNA sequencing using Sequenase 2.0 (United States Biochemical Corp, Cleveland, OH). This resulted in phasing of the dA:T, tracts and PRE over nearly one and a half turns of the DNA helix. DNA probes for gel-shift assay were prepared by digesting each plasmid with EcoR I and Hindlll, which excises the DNA insert. The DNA fragments ranged in length from 296-308 bp and were gel purified. Each DNA probe was end-labeled by Klenow DNA polymerase fill-in with @P]dATP and a[3’P]dCTP.
To obtain a k value we measured the relative mobilities of a set of DNA standards with intrinsic DNA bends according to the relationship originally defined by Thompson and Landy (77) and modified by Kerppola and Curran (10, 32): k,+,,I~e = cos(ka/2) where p*M is the relative mobility of the standard with the intrinsic bend positioned in the middle of the DNA fragment, pe is the relative mobility of the standard with the intrinsic bend positioned at the end of the DNA fragment, and 01 is the known bend angle. The DNA bend standards, as described bv Thompson and Landv (77). contained from two to seven phased dA:T, tracts po&ioned in the middle or at end of a DNA fragment of similar size and sequence. Phased dA:T, tracts bend DNA toward the minor groove where a single dA:T, tract bends DNA by 18” & 10% (77). A k value of 0.995 was obtained under our electrophoresis conditions.
Electrophoretic Gel Mobility Shift Assays
Our previous gel mobility shift assays used a short 32-bp oligonucleotide containing a single PRE (47). In the present studies, minor modifications were made for binding to a PRE contained in longer DNA fragments (142 bp for circular per- mutation analysis and 296 to 308 bp for phasing assays). Purified receptor (receptor concentration is indicated in molar amounts in figure legends) was incubated for 1 h at 4 C with [32P]labeled DNA fragments (range 0.3-l .O ng) in a total re- action volume of 25 iI. Also included was 75ng of poly(dA- dTVpolv(dA-dT) and 75 no sheared fscherichia co/i DNA as nonspecific competitor DNA. The DNA-binding buffer con- tained 10 mM Tris base, pH 7.4, 50 mM NaCI, 5 mM dithio- threitol, 2 mM MgCI,, 10% glycerol, and 50 rig/ml of a carrier protein (ovalbumin) to prevent highly purified PR from binding to nonspecific sites. With the 142-bp circular permutation DNA fragments, samples were electrophoresed on 6% poly- acrylamide gels prepared at a 5O:l (wt/wt) acrylamide-bis- acrylamide ratio using 20 mM Tris-acetate, 0.5 mM EDTA in the gels and as the electrode buffer. For phasing analysis with the 296- to 308-bp DNA fragments, conditions were the same except the polyacrylamide was 5% at a 75:l (wt/wt) ratio of acrylamide-bis. To maintain constant temperature during electrophoresis, 4 C water was recirculated through the gel apparatus.
Analysis and Quantification of DNA bending By Circular Permutation Assay
Analysis and Quantification of Directed DNA Bend Angles Detected by Phasing Assays
The relative mobilities of PR-DNA complexes obtained with the seven phasing DNA probes were corrected for mobility variations of the free DNA and normalized to the average mobility of all complexes. Normalized relative mobilities were then plotted as a function of the spacer length (base pairs) between the center of the intrinsic DNA bend and the PRE. The data points were connected by the best fit of a cosine function and is referred to as the phasing function. To deter- mine the orientation of the DNA bend, we have assumed an average of 10.5 bp per turn of the DNA helix between the intrinsic bend and PRE, and that the lowest mobility complex will have a helix periodicity that cooperates with the intrinsic bend, and the fastest mobility complex will have a helix periodicity that counteracts the intrinsic bend. Since dA:T, tracts bend DNA toward the minor groove, phasing analysis also allows a determination of the orientation of the protein- induced bend.
Directed DNA bending angles ((me) were calculated from the equation of Kerppola and Curran (10):
tan (Kc@) = tanzc,2)
where ApU is the amplitude of the phasing function (difference
The relative mobilities of PR-DNA complexes (142-bp PRE permuted DNA fragments) were calculated by measuring the distance in millimeters of the complex from the top of the gel and correctina for anv variation in mobilitv of free DNA probes (by calculating the ratio of the distance-in millimeters of the complex to that of free DNA). Complexes were then normal- ized to the fastest mobility complex (fastest = 1 and slower complexes are cl). Relative mobilities were plotted as a function of the distance (base pairs) of the center of the PRE from either end of the DNA probe. (-, left; +, right). Data points, taken as the average values from multiple determina- tions, were connected by least squares (best) fit of a cosine function (76). The apparent bend center was extrapolated from the maximum of the best fit curve.
,. between probes with fastest and slowest mobility), (~c is the anale of the intrinsic DNA bend (54”). and k is a coefficient (k = 0.995) that reflects factors other than’bending that can influence the relative mobility of DNA during gel electrophoresis.
Acknowledgments
The authors wish to thank Drs. Steven K. Nordeen (University of Colorado Health Sciences Center, Denver, CO) and Ann Nardulli (University of Illinois, Urbana, IL) for helpful discus- sions, Dr. Tom Kerppola (University of Michigan, Ann Arbor, MI) for orovidina ohasina olasmids pTK401-26 and pTK401- 28: Drs: John Tcdmpsonand Arthur Landy (Brown University, Providence, RI) for plasmids pJT170-2, 4, 5, and 7, Dr. Sankar Adhya for pBend-2 plasmid, and Dr. Philip Archer and James Murphy of the Biostatistics Core of the University of Colorado Cancer Center.
The bend angle derived from the circular permutation anal- ysis was quantitated by the method of Kerppola and Curran (10, 32). We have used the term “distortion angle” as previ- ously designated (10, 32) since mobility variations detected by circular permutation analysis can reflect DNA distortions other than a directed bend or unusual protein structure (10, 32,57,59). The magnitude of DNA distortion was taken as the amplitude of the best fit curve (or circular permutation func- tion) of the relative mobilities of the PR-DNA complexes plotted as a function of the distance of the PRE from the end of the DNA probe according to the equation, A,, = 1 -cos(kao/2), where A,, is the amplitude of the circular per- mutation function, k is a coefficient that adjusts for factors other than DNA distortion that can influence electrophoretic mobility, and o~o is the distortion angle (10, 32).
Received September 15, 1995. Re-revision received and accepted February 1, 1996.
Address requests for reprints to: Dean P. Edwards, Ph.D., Department of Pathology (B216), University of Colorado Health Sciences Center, 4200 East Ninth Avenue, Denver, Colorado 80262. This work was supported in part by NIH Grant CA-46938, Universitv of Colorado Cancer Center Tissue Culture Core (P30 CA46934), and Lucille A. Markey Charitable Trust.
Dow
nloaded from https://academ
ic.oup.com/m
end/article/10/4/393/2713356 by guest on 09 July 2022

VollONo.4 MOL END0 1996 406
REFERENCES
1.
2.
3.
4.
5.
6.
7.
8.
9.
10.
11.
12.
13.
14.
15.
16.
17.
18.
19.
Tsai M-J, O’Malley BW 1994 Molecular mchanism of action of steroid/thyroid receptor superfamily members. Annu Rev Biochem 63:451-486 Truss M, Beato M 1993 Steroid hormone receptors and interaction with deoxyribonucleic acid and transcription factors. Endocr Rev 14:459-479 Ing NH, Beekman JM, Tsai SY, Tsai M-J, O’Malley BW 1992 Members of the steroid hormone receptor super- family interact with TFIIB (S300-II). J Biol Chem 267: 17617-l 7623 Tong G-X, Tanen MR, Bagchi MK 1995 Ligand modu- lates the interaction of thyroid hormone receptor p with the basal transcription machinery. J Biol Chem 270: 10601-l 0611 Jacq X, Brou C, Lutz Y, Davidson I, Chambon P, Tora L 1994 Human TAF,,SO is present in a distinct TFIID com- plex and is required for transcriptional activation by the bestrogen receptor. Cell 79:107-l 17 Halachmi S, Marden E, Martin G, MacKav H, Abbondanza’ C, Brown M. 1994 Oestrogen receptor- associated proteins: possible mediators of hormone- induced transcription. Science 264:1455-1458 Cavill& V, Dauvois S, L’Horset F, Lopez G, Hoare S, Kushner PJ, Parker MG 1995 Nuclear factor Rip 140 modulates transcriptional activation by the estrogen re- ceptor. EMBO J 14:27413751 Le Douarin B, Zechel C, Garnier J-M, Lutz Y, Tora L, Pierrat B, Heety D, Gronemeyer H, Chambon P, Losson R 1995 The N-terminal part of TIFl , a putative mediator of the ligand-dependent activation function (AF-2) of nu- clear receptors, is fused to B-raf in the oncogenic protein T18. EMtiO J 14:2020-2033 OAate SA. Tsai SY. Tsai M-J. O’Mallev BW 1995 Se- quence and characterization 0; a coactiiator for the ste- roid hormone receptor superfamily. Science 270:1354- 1356 Kerppola TK, Curran T 1991 DNA Bending by Fos and Jun: The Flexible Hinge Model. Science 254:1210-1214 Kerppola TK, Curran T 1991 Fos-jun heterodimers and jun homodimers bend DNA in opposite orientations: im- plications for transcription factor cooperativity. Cell 66: 317-326 Kim Y, Geiger JH, Hahn S, Sigler PB 1993 Crystal struc- ture of a yeast TBP/TATA-box complex. Nature 365:512- 520 Kim JL, Nikolov DB, Burley SK 1993 Co-crystal structure of TBP recognizing the minor groove of a TATA element. Nature 365:520-527 Wechsler DS, Dang CV 1992 Opposite orientations of DNA bending by c-myc and max. Proc Natl Acad Sci USA 89:7635-7639 Saikumar P, Gabriel JL, Reddy P 1994 The Myb onco- gene product induces DNA-bending. Oncogene 9:1279- 1287 Schreck R, Zorbas H, Winnacker E-L, Baeuerle P 1990 The NF-kB transcription factor induces DNA bending which is modulated by its 65-kD subunit. Nucleic Acids Res 18:6497-6502 Schroth GP, Gottesfeld JM, Bradbuty EM 1991 TFIIIA induced DNA bending: effect of low ionic strength elec- trophoresis buffer conditions. Nucleic Acids Res 19:51 l- 516 Verrijzer CP, van Oosterhout JAWM, van Weperen WW, van der Vliet PC 1991 POU proteins bend DNA via the POU-specific domain. EMBO J 10:3007-3014 Erie DA, Yang G, Schultz HC, Bustamante C 1994 DNA bending by cro protein in specific and nonspecific com- plexes: implications for protein site recognition and specificity. Science 266:1562-l 566
20. Pbrez-Martin J, Rojo F, Delorenzo V 1994 Promoters responsive to DNA bending: a common theme in pro-
21. 22.
23.
24.
25.
26.
27.
28.
29.
30.
31.
32.
33.
34.
35.
36.
37.
38.
39.
40.
41.
42
43.
caryotic gene expression. Microbial Rev 58:268-290 Travers AA 1990 Why bend DNA? Cell 60:177-180 ton Hippel PH 1994 Protein-DNA recognition: new per- spectives and underlying themes. Science 263:769-770 Bracco L, Kotlarz D, Kolb A, Diekmann S, But H 1989 Synthetic curved DNA sequences can act as transcrip- tional activators in Escherichia co/i. EMBO J 8:4289- 4296 PBrez-Martin J, Espinosa M 1993 Protein-induced bend- lng as a transcriptional switch. Science 260:805-807 Wolffe AP 1994 Architectural transcription factors. Science 264:1100-l 101 Giese K, Cox J, Grosschedl R 1992 The HMG domain of lymphoid enhancer factor 1 bends DNA and facilitates assembly of functional nucleoprotein structures. Cell 69: 185-195 Thanos D, Maniatis T 1992 The high mobility group pro- tein HMG I (Y) is required for NF-GB-dependent virus induction of the human IFN-P gene. Cell 71:777-789 Pontiggia A, Rimini R, Harley VR, Goodfellow PN, Lovell- Badge R, Bianchi ME 1994 Sex-reversing mutations ef- fect the architecture of SRY-DNA complexes. EMBO J 13:6115-6124 Natesan S, Gilman MZ 1993 DNA bending and orienta- tion-dependent function of Wl in the c-fos promoter. Genes Dev 7:2497-2509 Kim J, Klooster S, Shapiro DJ 1995 Intrinsically bent DNA in a eukaryotic transcription factor recognition sequence potentiates transcription activation. J Biol Chem 270: 1282-l 288 Nardulli AM, Grobner C, Cotter D 1995 Estrogen receptor induced DNA bending: orientation or the bend and re- placement of an estrogen response element with an in- trinsic DNA binding sequence. Mol Endocrinol 9:1064- 1076 Kerppola TK, Curran T 1993 Selective DNA bending by a variety of bZlP proteins. Mol Cell Biol 13:5479-5489 Gartenberg MR, Crothers DM 1988 DNA sequence de- terminants of CAP-induced bending and protein binding affinity. Nature 333:824-829 Spana C, Corces VG 1990 DNA bending is a determinant of binding specificity for a Drosophila zinc finger protein. Genes Dev 4:1505-l 515 Ryder K, Silver S, DeLuca AL, Fanning E, Tegtmeyer P 1986 An altered DNA conformation in origin region I is a determinant of the binding of SV-40 large T-antigen. Cell 44:719-725 Patvin JD, McCormick RJ, Sharp PA, Fisher DE 1995 Pre-bending of a promoter sequence enhances affinity for the TATA-binding factor. Nature 373:724-727 Leidig F, Shepard AR, Zhang W, Stelter A, Cattini PA, Baxter JD. Eberhardt NL 1992 Thvroid hormone respon- siveness in human growth hormone-related genes. j Biol Chem 267:913-921 King IN, de Soyza T, Catanzaro DF, Lavin TN 1993 Thy- roid hormone receptor-induced bending of specific DNA sequences is modified by an accessory factor. J Biol Chem 268:495-501 Nardulli AM, Shapiro DJ 1992 Binding of the estrogen receptor DNA-binding domain to the estrogen response element induces DNA bending. Mol Cell Biol 12:2037- 2042 Nardulli AM, Greene GL, Shapiro DJ 1993 Human estro- gen receptor bound to an estrogen response element bends DNA. Mol Endocrinol 7:331340 Sabbah M, Le Ricousse S, Redeuilh G, Baulieu E-E 1992 Estrogen receptor-induced bending of the xenopus vitel- logenin A2 gene hormone response element. Biochem Biophys Res Commun 185944-952 Lu XP, Eberhardt NL, Pfahl M 1993 DNA bending by retinoid x receptor-containing retinoid and thyroid hor- mone receptor complexes. Mol Cell Biol 13:6509-&l 9 Kastner P, Krust A, Turcotte B, Stropp U, Tora L,
Dow
nloaded from https://academ
ic.oup.com/m
end/article/10/4/393/2713356 by guest on 09 July 2022

Gronemeyer H, Chambon P 1990 Two distinct estrogen- regulated promoters generate transcripts encoding the two functionally different human progesterone receptor forms A and B. EMBO J 9:1603-1614
44. Vegeto E, Shahbaz MM, Wen DX, Goldman ME, O’Malley BW, McDonnell DP 1993 Human progesterone receptor A form is a cell- and promoter-specific repressor of hu- man progesterone receptor B function. Mol Endocrinol 7:1244-l 255
45. Wen DX, Xu Y-F, Mais De, Goldman ME, McDonnell DP 1994 The A and B isoforms of the human progesterone receptor operate through distinct signaling pathways within target cells. Mol Cell Biol 14:8356-8364
46. Prendergast P, Otiate SA, Christensen K, Edwards DP 1994 Nuclear accessory factors enhance the binding of progesterone receptor to specific target DNA. J Steroid Biochem Mol Biol 48:1-13
47. Otiate SA, Prendergast P, Wagner JP, Nissen M, Reeves R. Pettiiohn DE. Edwards DP 1994 The DNA-bendina protein HMG-1 enhances progesterone receptor binding to its target DNA sequences. Mol Cell Biol 14:3376-3391
48. Pil PM, Chow CJ, Lippard SL 1993 High mobility-group 1 protein mediates DNA bending as determined by ring closures. Proc Natl Acad Sci USA 90:9465-9469
49. Bianchi ME 1994 Prokaryotic HU and eukatyotic HMGl: a kinked relationship. Mol Microbial 14:1-5
50. Crothers DM, Gartenberg MR, Shrader TE 1991 DNA bending in protein-DNA complexes. Methods Enzymol 208:118-l 47
51. Christensen K, Estes PA, ORate SA, Beck CA, DeMarzo A, Altmann M, Lieberman BA, St John J, Nordeen SK, Edwards DP 1991 Characterization and functional prop- et-ties of the A and B forms of human progesterone receptors synthesized in a baculovirus system. Mol Endocrinol 5:1755-l 770
52. Wu H-M, Crothers DM 1984 The locus of sequence- directed and protein-induced DNA bending. Nature 308: 509-513
53. DeMarzo AM, OAate SA, Nordeen SK, Edwards DP 1992 Effects of the steroid antagonist RU486 on dimerization of the human progesterone receptor. Biochemistry 31: 10491-10501
54. El-Ashry D, Onate SA, Nordeen SK, Edwards DP 1989 Human progesterone receptor complexed with the an- tagonist RU486 binds to hormone response elements in a structurally altered form. Mol Endocrinol 3:1545-l 558
55. Meyer M-E, Pornon A, Ji J, Bocquel M-T, Chambon P, Gronemeyer H 1990 Agonist and antagonist activities of RU486 on the functions of human progesterone recep- tor. EMBO J 9:3923-3932
56. Zinkel SS, Crothers DM 1987 DNA bend direction by phase sensitive direction. Nature 328:178-l 81
57. Gartenberg MR, Ampe C, Steitz TA, Crothers DM 1990 Molecular characterization of the GCN4-DNA complex. Proc Natl Acad Sci USA 87:6034-6038
58. Avitahl N, Calame K 1994 The C/EBP family of proteins distorts DNA upon binding but does not introduce a large directed bend. J Biol Chem 269:23553-23562
59 Mills JB, Cooper JP, Hagerman PJ 1994 Electrophoretic evidence that single-stranded regions of one or more nucleotides dramatically increase the flexibility of DNA. Biochemistry 33:1797-l 803
60 Watt F, Molly PL 1988 High mobility group proteins 1 and 2 stimulate binding of a specific transcription factor to the adenovirus late promoter. Nucleic Acids Res 16: 1471-1486
Progesterone Receptor-Induced DNA Bending
61. Zwilling S, Konig H, Wirth T 1995 High mobility group
protein 2 functionally interacts with the POU domains of octamer transcription factors. EMBO J 14:1198-l 208
62. Travers AA, Ner SS, Churchill MEA 1994 DNA chaper- ones: a solution to a persistence problem. Cell 77:167- 169
63. Lannigan DA, Notides AC 1989 Estrogen receptor selec- tively binds to the “coding strand” of an estrogen respon- sive element. Proc Nat1 Acad Sci USA 86:863-867
64. Mukherjee R 1993 Selective binding of the estrogen re- ceptor to one strand of the estrogen responsive element. Nucleic Acid Res 21:2655-2661
65. Allan GF. Lena X. Tsai SY. Weiael NL. Edwards DP. Tsai M-J, O’Malle; SW 1992 Horm&e and antihormone in- duce distinct conformational changes which are central to steroid receptor activation. J Biol Chem 267:19513- 19520
66.
67.
68.
69.
70.
71.
72.
73.
74.
75.
76.
77.
78.
Tung L, Mohamed MK, Hoeffler JP, Takimoto GS, Horwitz KB 1993 Antagonist-occupied human proges- terone B-receptors activate transcription without binding to progesterone response elements and are dominantly inh.ibited by A receptors. Mol Endocrinol 7:1244-1255 Sat-torius CA, Melville MY, Hovland RA. Tuna L. Takimoto GS, Horwitz’ KB 1994 A third transactiv&on domain function (AF3) of human progesterone receptor located in the unique N-terminal segment of the B isoform. Mol Endocrinol 8:1347-l 360 Shykind BM, Kim J, Sharp PA 1995 Activation of the TFIID-TFIIA complex with HMG-2. Genes Dev 9:1354- 1365 Ge H, Roeder RG 1994 The high mobility group protein HMG-1 can reversibly inhibit class II gene transcription by interaction with the TATA-binding protein. J Biol Chem 269:17136-l 7140 Stelzer G, Goppelt A, Lottspeich F, Meisterernst M 1994 Repression of basal transcription by HMG2 is counter- acted by TFIIH-associated factors in an ATP-dependent process. Mol Cell Biol 14:4712-4721 Estes PA, Suba EJ, Lawler-Heavner D, Elashry-Stowers D, Wei LL, Toft DO, Sullivan WP, Horwitz KB, Edwards DP 1987 Immunologic analysis of human breast cancer progesterone receptors. I. lmmunoaffinity purification of transformed receptors and production of monoclonal an- tibodies. Biochemistry 26:6250-6262 Adachi Y. Mizuno S. Yoshida M 1990 Efficient larae scale purification of non-histone chromosomal proteins HMG-1 and HMG-2 by using polybuffer exchanger PBE94. J Chromatogr 530:39-46 Kim J, Zwieb C, Wu C, Adhya S 1989 Bending of DNA by gene-regulatory proteins: construction and use of a DNA bending vector. Gene 85:15-23 Lathe R, Kieny MP, Skory S, Lecocq JP 1984 Linker tailing: Unphosphorylated linker oligonucleotides for join- ing DNA termini. DNA 3:173-l 82 Lieberman BA, Bona BJ, Edwards DP, Nordeen SK 1993 The constitution of a progesterone response element. Mol Endocrinol 7515-527 Ware JH, Bowden RE 1977 Circadian rhythm analysis when output is collected at intervals. Biometrics 33:566- 572 Thompson JF, Landy A 1988 Empirical estimation of protein-induced DNA bending angles: applications to 8 site-specific recombination complexes. Nucleic Acids Res 16:9687-9705 Zerbe GO, Laska E, Meisner M, Kushner HB 1982 On multivariate confidence regions and simultaneous confi- dence limits for ratios. Communications in Statistics- Theory and Methods 11:2401-2425
Dow
nloaded from https://academ
ic.oup.com/m
end/article/10/4/393/2713356 by guest on 09 July 2022




















