
Early-phase GVHD gene expression profile in target versus non-target tissues: kidney, a possible...
10
ORIGINAL ARTICLE Early-phase GVHD gene expression profile in target versus non-target tissues: kidney, a possible target? B Sadeghi 1 , H Al-Chaqmaqchi 1 , S Al-Hashmi 1 , D Brodin 2 , Z Hassan 1,3 , M Abedi-Valugerdi 4 , A Moshfegh 5 and M Hassan 1,3 GVHD is a major complication after allo-SCT. In GVHD, some tissues like liver, intestine and skin are infiltrated by donor T cells while others like muscle are not. The mechanism underlying targeted tropism of donor T cells is not fully understood. In the present study, we aim to explore differences in gene expression profile among target versus non-target tissues in a mouse model of GVHD based on chemotherapy conditioning. Expression levels of JAK–signal transducers and activators of transcription (STAT), CXCL1, ICAM1 and STAT3 were increased in the liver and remained unchanged (or decreased) in the muscle and kidney after conditioning. At the start of GVHD the expression levels of CXCL9, ITGb2, SAA3, MARCO, TLR and VCAM1 were significantly higher in the liver or kidney compared with the muscle of GVHD animals. Moreover, biological processes of inflammatory reactions, leukocyte migration, response to bacterium and chemotaxis followed the same pattern. Our data show that both chemotherapy and allogenicity exclusively induce expression of inflammatory genes in target tissues. Moreover, gene expression profile and histopathological findings in the kidney are similar to those observed in the liver of GVHD mice. Bone Marrow Transplantation advance online publication, 23 July 2012; doi:10.1038/bmt.2012.120 Keywords: gene expression; GVHD; kidney; target tissues; mouse model INTRODUCTION Allo-SCT is the treatment of choice for a variety of malignant and non-malignant diseases, especially in the field of hematology. 1 However, its application is limited owing to several complications, especially GVHD. 2 It is assumed that conditioning induces tissue injury and subsequent lipopolysaccharides (LPS) extravasation promotes a cytokine storm, leading to activation of donor T cells by host/ donor APCs. 3,4 Finally, allo-reactive donor T cells migrate and infiltrate peripheral tissues. Hypothetically all organs should be possible targets for donor T cells; nevertheless, the liver, gastrointestinal tract, skin and lungs 3,5,6 are the principal target organs. 7 Although damage to other organs (for example, kidney) has been described, 8,9 most clinicians do not consider the kidney a target organ for acute GVHD (aGVHD). Several hypotheses have been discussed, but clear alterations of target versus non-target tissues have not been explored yet. Inflammatory cytokine secretion and chemokine expression have an important role in lymphocyte activation and migration to inflammation sites. 10–13 Mapara et al. 14 , Ma et al. 15 and others have shown higher expression of recruiting chemokines on the target tissues of GVHD mice. Overexpression or aberrant expression of MHC molecules 16 has been suggested as a possible cause for the targeted invasion of donor T cells. 3 These reports shed light on the principal molecular changes in the process of GVHD; however, owing to elevated levels of cytokines in the serum and systemic inflammation following GVHD, these findings cannot be considered exclusive changes in target organs—especially when compared with syngeneically transplanted animals. Moreover, none of these reports describe any changes in gene expression profile of non-target tissues like muscle. We hypothesized that comprehensive gene expression analysis of target versus non-target tissues after conditioning regimen and in the early phases of GVHD might help us to understand the underlying mechanisms of T-cell trafficking. Hence, in the present report, we classified the liver as affected (target) and muscle as less-affected (non-target) organs, and explored the role of the kidney in aGVHD. Then we compared gene expression profiles before and after conditioning, as well as in the early phase of aGVHD within and between groups (allo- and syngeneic SCT). MATERIALS AND METHODS Animals Female BALB/c and male C57BL/6 mice, 10–14 weeks old, were purchased from Scanbur (Sollentuna, Sweden). They were maintained under specific pathogen-free conditions in an animal facility with individually ventilated cages. Animals were fed autoclaved mouse chow and tap water ad libitum and were allowed to acclimatize to their surroundings for 1 week. All experiments described here were approved by the southern Stockholm ethics committee for animal research in accordance with Swedish Animal Welfare law. Induction of aGVHD GVHD induction was performed as previously described. 17 Briefly, recipient mice received BU (80 mg/kg) followed by CY (200 mg/kg). At day 0, recipient mice were injected via the lateral tail vein with 2 10 7 and 3 10 7 viable cells of bone marrow cell (BMC) and splenocyte (SP) from C57BL/6 (allogeneic) or BALB/c (syngeneic) as donor, respectively. 1 Experimental Cancer Medicine, Institution for Laboratory Medicine, Karolinska Institute, Stockholm, Sweden; 2 Bioinformatics and Expression Analysis core facility, Karolinska Institute, Stockholm, Sweden; 3 Clinical Research Centre (KFC, Novum), Karolinska University Hospital-Huddinge, Stockholm, Sweden; 4 Department of Biochemistry and Biophysics, Arrhenius Laboratories for the Natural Sciences, Stockholm University, Stockholm, Sweden and 5 Department of Oncology and Pathology, Cancer Center Karolinska (CCK), Karolinska Hospital and Institute, Stockholm, Sweden. Correspondence: Dr B Sadeghi, Division of Clinical Immunology, F79, Karolinska Institutet, Karolinska University Hospital-Huddinge, SE 14186 Stockholm, Sweden. E-mail: [email protected] Received 27 February 2012; accepted 20 May 2012 Bone Marrow Transplantation (2012), 1–10 & 2012 Macmillan Publishers Limited All rights reserved 0268-3369/12 www.nature.com/bmt
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Early-phase GVHD gene expression profile in target versus non-target tissues: kidney, a possible...

ORIGINAL ARTICLE
Early-phase GVHD gene expression profile in target versus
non-target tissues: kidney, a possible target?B Sadeghi1, H Al-Chaqmaqchi1, S Al-Hashmi1, D Brodin2, Z Hassan1,3, M Abedi-Valugerdi4, A Moshfegh5 and M Hassan1,3
GVHD is a major complication after allo-SCT. In GVHD, some tissues like liver, intestine and skin are infiltrated by donor T cells while
others like muscle are not. The mechanism underlying targeted tropism of donor T cells is not fully understood. In the present
study, we aim to explore differences in gene expression profile among target versus non-target tissues in a mouse model of GVHD
based on chemotherapy conditioning. Expression levels of JAK–signal transducers and activators of transcription (STAT), CXCL1,
ICAM1 and STAT3 were increased in the liver and remained unchanged (or decreased) in the muscle and kidney after conditioning.
At the start of GVHD the expression levels of CXCL9, ITGb2, SAA3, MARCO, TLR and VCAM1 were significantly higher in the liver or
kidney compared with the muscle of GVHD animals. Moreover, biological processes of inflammatory reactions, leukocyte migration,
response to bacterium and chemotaxis followed the same pattern. Our data show that both chemotherapy and allogenicity
exclusively induce expression of inflammatory genes in target tissues. Moreover, gene expression profile and histopathological
findings in the kidney are similar to those observed in the liver of GVHD mice.
Bone Marrow Transplantation advance online publication, 23 July 2012; doi:10.1038/bmt.2012.120
Keywords: gene expression; GVHD; kidney; target tissues; mouse model
INTRODUCTION
Allo-SCT is the treatment of choice for a variety of malignant andnon-malignant diseases, especially in the field of hematology.1
However, its application is limited owing to several complications,especially GVHD.2
It is assumed that conditioning induces tissue injury andsubsequent lipopolysaccharides (LPS) extravasation promotes acytokine storm, leading to activation of donor T cells by host/donor APCs.3,4 Finally, allo-reactive donor T cells migrate andinfiltrate peripheral tissues. Hypothetically all organs should bepossible targets for donor T cells; nevertheless, the liver,gastrointestinal tract, skin and lungs3,5,6 are the principal targetorgans.7 Although damage to other organs (for example, kidney)has been described,8,9 most clinicians do not consider the kidney atarget organ for acute GVHD (aGVHD). Several hypotheses havebeen discussed, but clear alterations of target versus non-targettissues have not been explored yet. Inflammatory cytokinesecretion and chemokine expression have an important role inlymphocyte activation and migration to inflammation sites.10–13
Mapara et al.14, Ma et al.15 and others have shown higherexpression of recruiting chemokines on the target tissues of GVHDmice. Overexpression or aberrant expression of MHC molecules16
has been suggested as a possible cause for the targeted invasionof donor T cells.3 These reports shed light on the principalmolecular changes in the process of GVHD; however, owingto elevated levels of cytokines in the serum and systemicinflammation following GVHD, these findings cannot beconsidered exclusive changes in target organs—especially whencompared with syngeneically transplanted animals. Moreover,
none of these reports describe any changes in gene expressionprofile of non-target tissues like muscle.We hypothesized that comprehensive gene expression analysis
of target versus non-target tissues after conditioning regimen andin the early phases of GVHD might help us to understand theunderlying mechanisms of T-cell trafficking. Hence, in the presentreport, we classified the liver as affected (target) and muscle asless-affected (non-target) organs, and explored the role of thekidney in aGVHD. Then we compared gene expression profilesbefore and after conditioning, as well as in the early phase ofaGVHD within and between groups (allo- and syngeneic SCT).
MATERIALS AND METHODS
Animals
Female BALB/c and male C57BL/6 mice, 10–14weeks old, were purchasedfrom Scanbur (Sollentuna, Sweden). They were maintained under specificpathogen-free conditions in an animal facility with individually ventilatedcages. Animals were fed autoclaved mouse chow and tap water ad libitumand were allowed to acclimatize to their surroundings for 1week. Allexperiments described here were approved by the southern Stockholmethics committee for animal research in accordance with Swedish AnimalWelfare law.
Induction of aGVHD
GVHD induction was performed as previously described.17 Briefly, recipientmice received BU (80mg/kg) followed by CY (200mg/kg). At day 0,recipient mice were injected via the lateral tail vein with 2� 107 and3� 107 viable cells of bone marrow cell (BMC) and splenocyte (SP) fromC57BL/6 (allogeneic) or BALB/c (syngeneic) as donor, respectively.
1Experimental Cancer Medicine, Institution for Laboratory Medicine, Karolinska Institute, Stockholm, Sweden; 2Bioinformatics and Expression Analysis core facility, Karolinska
Institute, Stockholm, Sweden; 3Clinical Research Centre (KFC, Novum), Karolinska University Hospital-Huddinge, Stockholm, Sweden; 4Department of Biochemistry and Biophysics,
Arrhenius Laboratories for the Natural Sciences, Stockholm University, Stockholm, Sweden and 5Department of Oncology and Pathology, Cancer Center Karolinska (CCK),
Karolinska Hospital and Institute, Stockholm, Sweden. Correspondence: Dr B Sadeghi, Division of Clinical Immunology, F79, Karolinska Institutet, Karolinska University
Hospital-Huddinge, SE 14186 Stockholm, Sweden.
E-mail: [email protected]
Received 27 February 2012; accepted 20 May 2012
Bone Marrow Transplantation (2012), 1–10
& 2012 Macmillan Publishers Limited All rights reserved 0268-3369/12
www.nature.com/bmt
Assessment of GVHD
Recipient mice were examined daily from the start of conditioning until theindicated sampling day. Animals were evaluated for five clinical symptomsof GVHD: weight loss, posture, activity, fur texture and skin integrity asdescribed elsewhere.17
Histopathology
To confirm GVHD, histopathological evaluation was performed asdescribed previously.17 Tissue samples were fixed in neutral bufferedformalin for 24 h, transferred to 70% ethanol, dehydrated and embeddedin paraffin according to standard procedure. Sections of 4 mm wereprepared and stained by hematoxylin and eosin for histopathologicalevaluation.
Tissue preparation
At day � 7 (control), 0 (after conditioning) and þ 7 (start of GVHD)between three to six mice per group were killed. To exclude the effects ofcirculatory cells in tissues, the entire mouse body was perfused throughthe heart with 20–30-mL-ice-cold PBS containing 2% FBS and 10mM EDTA.Then portions of the liver, kidney and muscle were isolated, snap frozen inliquid nitrogen and transferred to � 150 1C until analysis.
Microarray analysis
RNA extraction was done according to Qiagen RNeasy kit instructionsmanual (WVR, Stockholm, Sweden). The integrity of extracted RNA wasconfirmed using the Agilent 2100Bioanalyzer (Agilent Technologies, PaloAlto, CA, USA).cRNA sysnthesis and microarray analysis were done according to
Affymetrix manual (Affymetrix Inc., Santa Clara, CA, USA). Scanned imageswere inspected and analyzed using established quality control criteria.
Data analysis
Data analysis was performed with Affymetrix GeneChip Operating Software(GCOS, v1.1) using the MAS5 method (TGT¼ 100). A quantitative signaland a qualitative detection call was generated for each sample andtranscript. Within each tissue, pair-wise comparisons of treated samplesversus the untreated sample were performed, generating a correspondingquantitative signal log ratio and a qualitative change call.
Enrichment analysis
To investigate induced treatment effects in the different tissues in afunctional context, enrichment analysis was performed using WebGestalt(http://bioinfo.vanderbilt.edu/webgestalt/). Differently expressed geneswithin each tissue were selected and tested for enrichment of the geneontology subcategory (http://www.geneontology.org) and KEGG pathways(http://www.genome.ad.jp/kegg/). Hypergeometric was considered forstatistical method, significance level was 0.01, minimum number of genesfor a category considered 4 and P-value correction by Benjamini &Hochberg, statistical method (BH).Principal component analysis plot sample similarities in expression
patterns were visualized by three-dimensional principal componentanalysis plots. A set of gene ontology terms were selected for visualizationand genes associated with the respective term and present in at least onesample were subjected to principal component analysis using QlucoreOmics Explorer 2.0 (www.qlucore.com/).
RESULTS
Establishment of GVHD
In this model (C57BL/6 to BALB/c), preliminary signs of clinical andimmunological manifestations of aGVHD emerge 1 week after allo-
Skin allo Colon allo
Skin syn
Colon syn
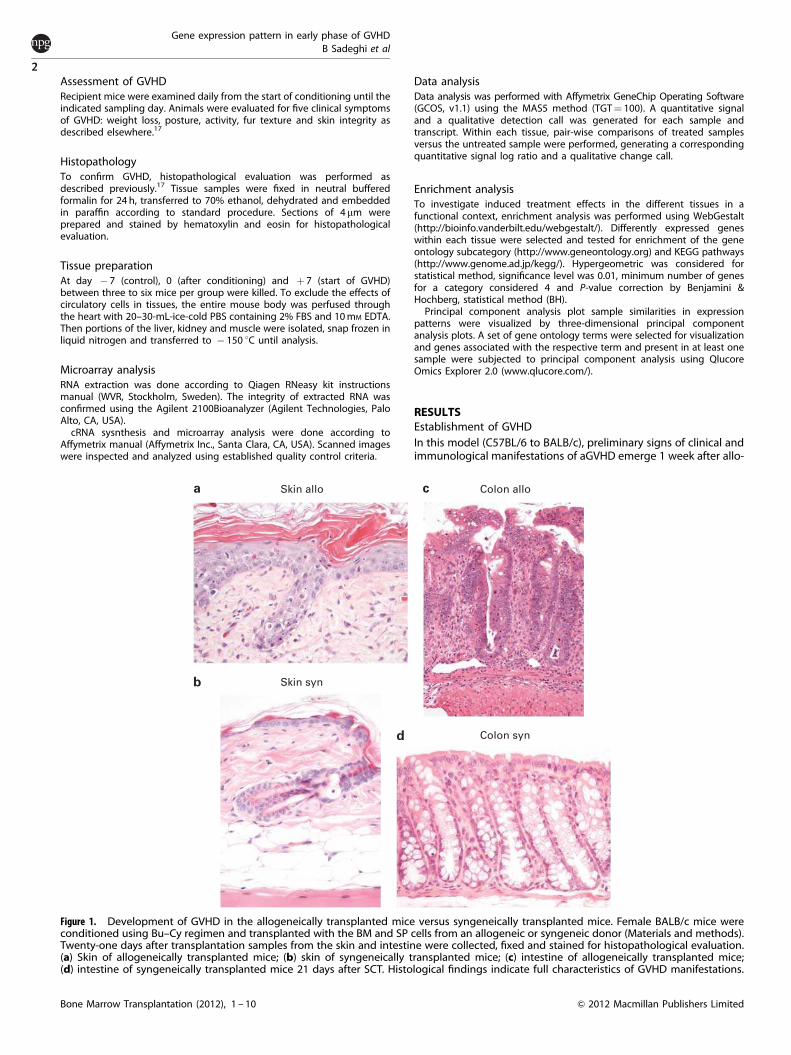
Figure 1. Development of GVHD in the allogeneically transplanted mice versus syngeneically transplanted mice. Female BALB/c mice wereconditioned using Bu–Cy regimen and transplanted with the BM and SP cells from an allogeneic or syngeneic donor (Materials and methods).Twenty-one days after transplantation samples from the skin and intestine were collected, fixed and stained for histopathological evaluation.(a) Skin of allogeneically transplanted mice; (b) skin of syngeneically transplanted mice; (c) intestine of allogeneically transplanted mice;(d) intestine of syngeneically transplanted mice 21 days after SCT. Histological findings indicate full characteristics of GVHD manifestations.
Gene expression pattern in early phase of GVHD
B Sadeghi et al
2
Bone Marrow Transplantation (2012), 1 – 10 & 2012 Macmillan Publishers Limited
SCT.17,18 Twenty-one days after transplantation histologicalfindings in the skin (inter-follicular epidermal hyperplasia,apoptotic figures and disappearance of hypodermal fat layer)and intestine (severe disturbance in crypts, hyperplasia,hyperchromasi and apoptotic features) of the allogeneic groupwas compatible with GVHD manifestations and confirmed theoccurrence of GVHD (Figure 1). Weight changes and other clinicalindices of GVHD were also observed in allogeneically transplantedanimals, whereas the syngeneic group recovered to normalconditions (data not shown).
Effect of chemotherapy conditioning on gene expression profile
We evaluated the gene expression profile in the liver, kidney andmuscle after chemotherapy conditioning (day 0). Expressions of164, 114 and 660 genes were changed in the liver, kidney andmuscle, respectively. Table 1 lists the 20 genes most highlyaffected by Bu–Cy regimen in three different tissues. Adistinguishable effect of conditioning was that genes involved inhemoglobin synthesis exclusively decreased in all tissues (Table 1).Reciprocally, expression levels of only two genes, FK506-bindingprotein 5 and Vanin 1, increased in all studied tissues afterconditioning.Inflammation-related genes-like CXCL1, IFNg-induced GTPase
(Igtp), signal transducers and activators of transcription (STAT3)and intercellular adhesion molecule 1 (ICAM-1) were upregulatedby 13-, 2.8-, 2.3- and 2.1-fold, respectively, in the liver, while notchanging or decreasing in the kidney or muscle.
Effect of conditioning on expression of different biologicalprocesses
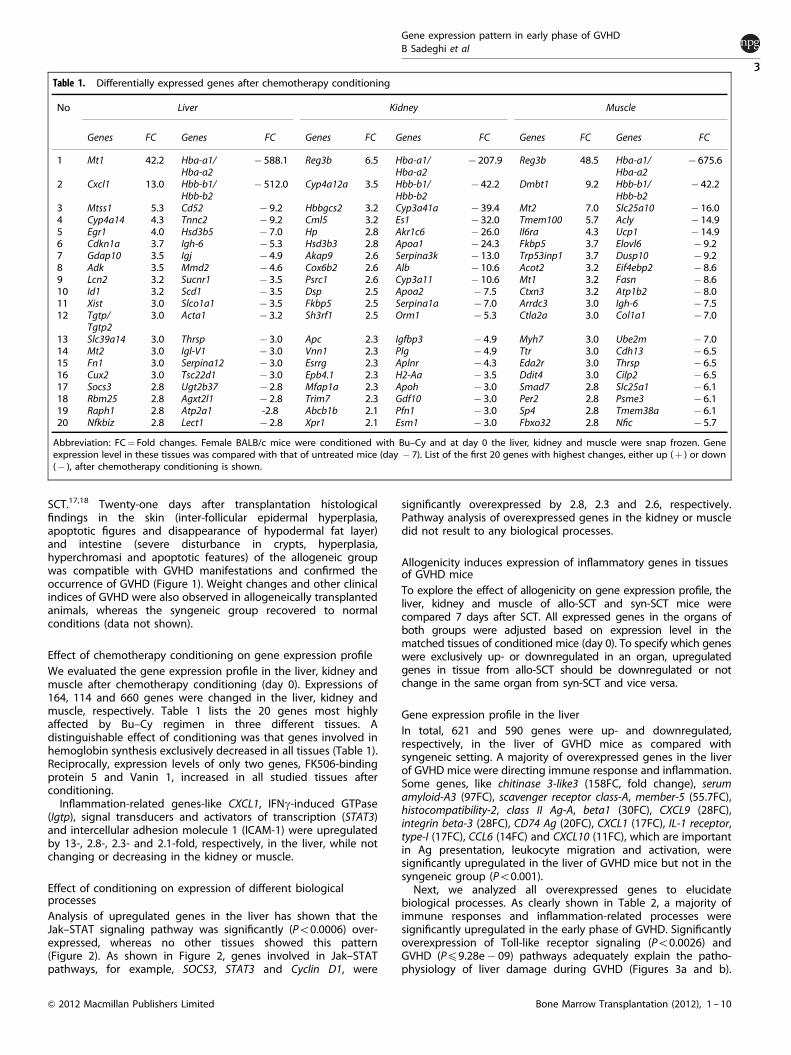
Analysis of upregulated genes in the liver has shown that theJak–STAT signaling pathway was significantly (Po0.0006) over-expressed, whereas no other tissues showed this pattern(Figure 2). As shown in Figure 2, genes involved in Jak–STATpathways, for example, SOCS3, STAT3 and Cyclin D1, were
significantly overexpressed by 2.8, 2.3 and 2.6, respectively.Pathway analysis of overexpressed genes in the kidney or muscledid not result to any biological processes.
Allogenicity induces expression of inflammatory genes in tissuesof GVHD mice
To explore the effect of allogenicity on gene expression profile, theliver, kidney and muscle of allo-SCT and syn-SCT mice werecompared 7 days after SCT. All expressed genes in the organs ofboth groups were adjusted based on expression level in thematched tissues of conditioned mice (day 0). To specify which geneswere exclusively up- or downregulated in an organ, upregulatedgenes in tissue from allo-SCT should be downregulated or notchange in the same organ from syn-SCT and vice versa.
Gene expression profile in the liver
In total, 621 and 590 genes were up- and downregulated,respectively, in the liver of GVHD mice as compared withsyngeneic setting. A majority of overexpressed genes in the liverof GVHD mice were directing immune response and inflammation.Some genes, like chitinase 3-like3 (158FC, fold change), serumamyloid-A3 (97FC), scavenger receptor class-A, member-5 (55.7FC),histocompatibility-2, class II Ag-A, beta1 (30FC), CXCL9 (28FC),integrin beta-3 (28FC), CD74 Ag (20FC), CXCL1 (17FC), IL-1 receptor,type-I (17FC), CCL6 (14FC) and CXCL10 (11FC), which are importantin Ag presentation, leukocyte migration and activation, weresignificantly upregulated in the liver of GVHD mice but not in thesyngeneic group (Po0.001).Next, we analyzed all overexpressed genes to elucidate
biological processes. As clearly shown in Table 2, a majority ofimmune responses and inflammation-related processes weresignificantly upregulated in the early phase of GVHD. Significantlyoverexpression of Toll-like receptor signaling (Po0.0026) andGVHD (Pp9.28e� 09) pathways adequately explain the patho-physiology of liver damage during GVHD (Figures 3a and b).
Table 1. Differentially expressed genes after chemotherapy conditioning
No Liver Kidney Muscle
Genes FC Genes FC Genes FC Genes FC Genes FC Genes FC
1 Mt1 42.2 Hba-a1/Hba-a2
� 588.1 Reg3b 6.5 Hba-a1/Hba-a2
� 207.9 Reg3b 48.5 Hba-a1/Hba-a2
� 675.6
2 Cxcl1 13.0 Hbb-b1/Hbb-b2
� 512.0 Cyp4a12a 3.5 Hbb-b1/Hbb-b2
� 42.2 Dmbt1 9.2 Hbb-b1/Hbb-b2
� 42.2
3 Mtss1 5.3 Cd52 � 9.2 Hbbgcs2 3.2 Cyp3a41a � 39.4 Mt2 7.0 Slc25a10 � 16.04 Cyp4a14 4.3 Tnnc2 � 9.2 Cml5 3.2 Es1 � 32.0 Tmem100 5.7 Acly � 14.95 Egr1 4.0 Hsd3b5 � 7.0 Hp 2.8 Akr1c6 � 26.0 Il6ra 4.3 Ucp1 � 14.96 Cdkn1a 3.7 Igh-6 � 5.3 Hsd3b3 2.8 Apoa1 � 24.3 Fkbp5 3.7 Elovl6 � 9.27 Gdap10 3.5 Igj � 4.9 Akap9 2.6 Serpina3k � 13.0 Trp53inp1 3.7 Dusp10 � 9.28 Adk 3.5 Mmd2 � 4.6 Cox6b2 2.6 Alb � 10.6 Acot2 3.2 Eif4ebp2 � 8.69 Lcn2 3.2 Sucnr1 � 3.5 Psrc1 2.6 Cyp3a11 � 10.6 Mt1 3.2 Fasn � 8.610 Id1 3.2 Scd1 � 3.5 Dsp 2.5 Apoa2 � 7.5 Ctxn3 3.2 Atp1b2 � 8.011 Xist 3.0 Slco1a1 � 3.5 Fkbp5 2.5 Serpina1a � 7.0 Arrdc3 3.0 Igh-6 � 7.512 Tgtp/
Tgtp23.0 Acta1 � 3.2 Sh3rf1 2.5 Orm1 � 5.3 Ctla2a 3.0 Col1a1 � 7.0
13 Slc39a14 3.0 Thrsp � 3.0 Apc 2.3 Igfbp3 � 4.9 Myh7 3.0 Ube2m � 7.014 Mt2 3.0 Igl-V1 � 3.0 Vnn1 2.3 Plg � 4.9 Ttr 3.0 Cdh13 � 6.515 Fn1 3.0 Serpina12 � 3.0 Esrrg 2.3 Aplnr � 4.3 Eda2r 3.0 Thrsp � 6.516 Cux2 3.0 Tsc22d1 � 3.0 Epb4.1 2.3 H2-Aa � 3.5 Ddit4 3.0 Cilp2 � 6.517 Socs3 2.8 Ugt2b37 � 2.8 Mfap1a 2.3 Apoh � 3.0 Smad7 2.8 Slc25a1 � 6.118 Rbm25 2.8 Agxt2l1 � 2.8 Trim7 2.3 Gdf10 � 3.0 Per2 2.8 Psme3 � 6.119 Raph1 2.8 Atp2a1 -2.8 Abcb1b 2.1 Pfn1 � 3.0 Sp4 2.8 Tmem38a � 6.120 Nfkbiz 2.8 Lect1 � 2.8 Xpr1 2.1 Esm1 � 3.0 Fbxo32 2.8 Nfic � 5.7
Abbreviation: FC¼ Fold changes. Female BALB/c mice were conditioned with Bu–Cy and at day 0 the liver, kidney and muscle were snap frozen. Gene
expression level in these tissues was compared with that of untreated mice (day � 7). List of the first 20 genes with highest changes, either up (þ ) or down
(� ), after chemotherapy conditioning is shown.
Gene expression pattern in early phase of GVHD
B Sadeghi et al
3
& 2012 Macmillan Publishers Limited Bone Marrow Transplantation (2012), 1 – 10

Interestingly CD14 (22.7FC), LBP (5.3FC), TLR1 (5.3FC) and TLR2(7FC) genes were significantly upregulated.
Gene expression profile in the kidney
Gene expression analysis shows that numerous genes related toinflammation and the immune response were upregulated in thekidney of GVHD mice (day þ 7), while remaining unchanged oreven decreasing in the syngeneic setting. The genes Chitinase 3-like3(955-fold), CXCL9 (72-fold), IFN-inducible GTPase-1 (72-fold), IFN g-inducible protein-47 (49-fold), Histocompatibility-2, Q region (49-fold),Histocompatibility-2, class II, locus Mb-2 (49-fold), Macrophageactivation-2 like (45-fold), IFN-inducible GTPase-1 (45-fold), Gran-zyme-B (39-fold), Granzyme-K (34-fold), CD3 Ag, g-polypeptide (34-fold) and CXCL11 (32-fold), which directly or indirectly are relevant toimmune responses and/or inflammation, were significantly upregu-lated in the kidney of GVHD mice but not in the syngeneic group.As shown in Figure 3c, overexpression of MHC (both classes) in
combination with intermediate genes, for example, CLIP (11.3FC),TAPBP (4.6FC), TAP-1 (26FC) and TAP-2 (15FC), genes that areinvolved in Ag processing and presentation pathways, wassignificantly higher in the kidney of GVHD mice.
Gene expression profile in the muscle
In our analysis, we have found that a total of 3918 genes werechanged in the muscle of GVHD mice as compared with the
syngeneic group. Among them several inflammatory genes thatmainly relate to systemic inflammation and cytokine effects wereoverexpressed in the GVHD group. Although a majority ofobserved pathways (470%) belonged to cell biological andmetabolic clusters,o30% of them were directed to Ag processingand presentation as was expected. As opposed to the kidney and/or liver, important genes for chemotaxis-like IL-1-b, chemokinesand adhesive molecules, for example, CXCL9, CXCL1, CCL1, Marco,Saa-1, Saa-3, STAT1 and VCAM, either were not expressed or weresignificantly less expressed in the muscle. In summary, despite themuscle of GVHD mice presenting more inflammatory genes thanthe muscle of syn-SCT animals, they still expressed much lessinflammatory, chemotactic genes compared with the liver andkidney of GVHD mice.
Target organs have different gene expression pattern in the earlystages of GVHD
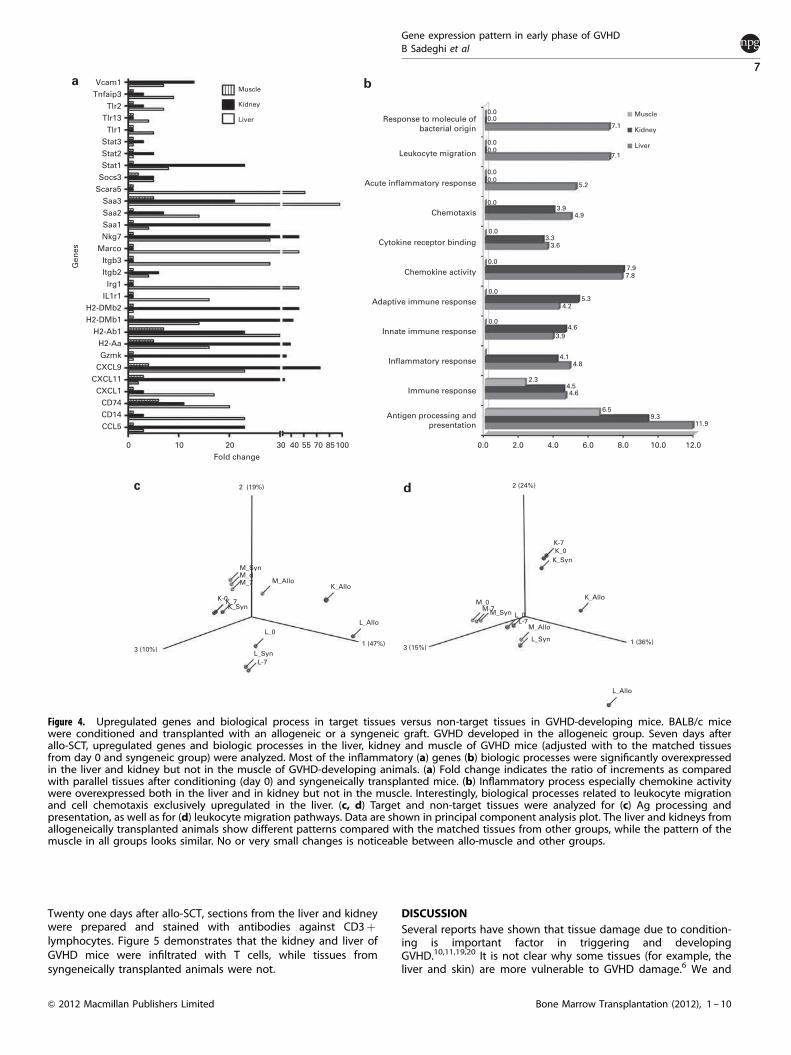
In total 345 genes were exclusively upregulated in the liver ofGVHD mice. Figure 4a compares the expression level of some ofthese genes in three different tissues of the GVHD group. Serumamyloid A-3, scavenger receptor class-A, member-5, macrophagereceptor with collagenous structure, immunoresponsive gene-1,CD14, CXCL9, TLR2 and TLR1 were significantly increased by 97-,56-, 49-, 49-, 23-, 23-, 7- and 5-fold, respectively, in the liver,whereas remaining unchanged or decreasing in the muscle of thesame mice or liver of syn-SCT animals (Figure 4a).
JAK-STAT signaling pathway
Endocytosis Lysosomal
proteolysis
Cbl
JAK
PIAS
SHP1
–p
–p
+p
+u
+u
STAT5
Homo/hetero
STAT dimers
Ras/MAPKMAPK signaling
pathway
Antiapoptosis
IFNα
CBP
Pim-1
CIS
Growth, proliferation,
fate determination,
development, immunity
c-Myc
Cell cycle
Antiapoptosis
Apoptosis
04630 3/31/09
(c) Kanehisa laboratories
BclXL
Sprouty
Spred
Apoptosis
+p
+p +p
SHP2 GRB SOS
PI3K AKT
STAM
STAT
P48
SOCS
CycD
CytokineR
Ubiquitin
mediated
proteolysis
EPO
IFN/IL10
IL2/3
IL6
UPD
Cytokine
Cytokine-cytokine
receptor
interaction
Figure 2. Pathway analysis of the liver after chemotherapy conditioning. Female BALB/c mice were conditioned with Bu–Cy regimen and atday 0, the liver was snap frozen. All upregulated genes (compared with untreated mice) were analyzed. Chemotherapy conditioningsignificantly (Po0.0006) induced upregulation of the JAK–STAT pathway in the liver, whereas the kidney and muscle did not show the samepattern. Arrows indicate overexpressed genes.
Gene expression pattern in early phase of GVHD
B Sadeghi et al
4
Bone Marrow Transplantation (2012), 1 – 10 & 2012 Macmillan Publishers Limited

Figure 4b illustrates and compares the expression level ofbiological processes or molecular functions in the liver, kidney andmuscle 7 days after allo-SCT. As shown in Figure 4b, a majority ofimmune-related processes are upregulated in the liver or kidneybut not in the muscle.
The kidney could be a target tissue after allo-SCT
Our analysis showed that 311 genes were exclusively upregulatedin the kidney of allo-SCT mice. These genes were unchanged oreven decreased in the kidney of syn-SCT animals and also themuscle of GVHD mice. Most of the overexpressed genes in thekidney of GVHD mice were related to Ag presenting and immuneresponse (Figure 4a). Although stimulated biological processes inthe kidney are comparable to those observed in the liver, genes(Figure 4a) and biological processes (Figure 4b) showed differentexpression levels and patterns in these two organs. Notably, bothare significantly different from those observed in the muscle.
Figure 4b shows that some biological processes, mainly thoserelated to chemokine function, cell taxis (Po9.75e� 05) andleukocyte migration/trafficking (Po1.71e� 05) are exclusivelyoverexpressed in the liver.To evaluate the whole picture of gene expression and pathways
across different tissues and conditions, we have analyzed allrelevant data regarding Ag processing and presentation, andleukocyte migration, and have shown in principal componentanalysis graph (Figures 4c and d). As shown in Figures 4c and d,the liver and kidney of GVHD animals have different patterns ascompared with similar tissues in controls (� 7) after conditioning(0) and in the syngeneic group (Syn), whereas the pattern in themuscle of GVHD animals does not show a similar pattern.
T-cell infiltration in the kidney of GVHD mice
We next explored the infiltration of CD3þ T lymphocytes intarget tissues as a result of GVHD progress and establishment.
Table 2. List of biological processes that were significantly over expressed in the liver of GVHD mice
No Biologic process Go code C O E R Raw P-value Adj P-value
1 Response to stimulus 0050896 1160 138 56.74 2.43 2.74e� 27 1.51e� 242 Defense response 0006952 259 60 12.67 4.74 1.30e� 25 3.59e� 233 Immune system process 0002376 479 81 23.43 3.46 8.99e� 25 1.65e� 224 Immune response 0006955 278 58 13.60 4.27 2.74e� 22 3.78e� 205 Response to stress 0006950 632 85 30.91 2.75 3.76e� 19 4.15e� 176 Inflammatory response 0006954 155 39 7.58 5.14 3.75e� 18 3.45e� 167 Ag processing and presentation 0019882 39 21 1.91 11.01 4.53e� 18 3.57e� 168 Response to external stimulus 0009605 365 58 17.85 3.25 2.51e� 16 1.73e� 149 Response to wounding 0009611 223 44 10.91 4.03 4.73e� 16 2.90e� 1410 Ag processing and presentation of peptide ag 0048002 25 16 1.22 13.08 1.03e� 15 5.69e� 1411 Ag processing and presentation of exogenous peptide ag 0002478 17 11 0.83 13.23 3.10e� 11 1.43e� 0912 Ag processing and presentation of exogenous ag 0019884 21 12 1.03 11.68 3.05e� 11 1.43e� 0913 Response to chemical stimulus 0042221 376 48 18.39 2.61 4.35e� 10 1.85e� 0814 Acute inflammatory response 0002526 51 16 2.49 6.41 1.13e� 09 4.46e� 0815 Positive regulation of immune system process 0002684 112 23 5.48 4.20 2.92e� 09 1.07e� 0716 Multi-organism process 0051704 172 29 8.41 3.45 3.17e� 09 1.09e� 0717 Chemotaxis 0006935 81 19 3.96 4.80 7.12e� 09 2.18e� 0718 Taxis 0042330 81 19 3.96 4.80 7.12e� 09 2.18e� 0719 Regulation of immune system process 0002682 168 28 8.22 3.41 7.85e� 09 2.28e� 0720 Response to other organism 0051707 114 22 5.58 3.95 2.18e� 08 6.02e� 0721 Ag processing and presentation of peptide ag via MHC Class II 0002495 13 8 0.64 12.58 3.13e� 08 7.85e� 0722 Ag processing and presentation of exogenous peptide ag
via MHC Class II0019886 13 8 0.64 12.58 3.13e� 08 7.85e� 07
23 Ag processing and presentation of peptide or polysaccharideag via MHC Class II
0002504 14 8 0.68 11.68 7.00e� 08 1.68e� 06
24 Response to biotic stimulus 0009607 144 24 7.04 3.41 9.62e� 08 2.21e� 0625 Leukocyte migration 0050900 31 11 1.52 7.25 1.14e� 07 2.52e� 0626 Ag processing and presentation of peptide ag via MHC Class I 0002474 11 7 0.54 13.01 1.75e� 07 3.58e� 0627 Response to bacterium 0009617 79 17 3.86 4.40 1.74e� 07 3.58e� 0628 Leukocyte mediated immunity 0002443 73 16 3.57 4.48 3.09e� 07 6.09e� 0629 Immune effector process 0002252 93 18 4.55 3.96 4.12e� 07 7.84e� 0630 Regulation of response to stimulus 0048583 169 25 8.27 3.02 5.55e� 07 1.02e� 0531 Adaptive immune response 0002250 68 15 3.33 4.51 6.68e� 07 1.15e� 0532 Adaptive immune response based on somatic recombination
of immune receptors built from ig super family domains0002460 68 15 3.33 4.51 6.68e� 07 1.15e� 05
33 Positive regulation of response to stimulus 0048584 88 17 4.30 3.95 8.94e� 07 1.50e� 0534 Positive regulation of immune response 0050778 71 15 3.47 4.32 1.20e� 06 1.95e� 0535 Innate immune response 0045087 74 15 3.62 4.14 2.10e� 06 3.31e� 0536 Positive regulation of T Cell differentiation 0045582 20 8 0.98 8.18 2.26e� 06 3.47e� 0537 Positive regulation of lymphocyte differentiation 0045621 21 8 1.03 7.79 3.50e� 06 5.22e� 0538 Acute-phase response 0006953 22 8 1.08 7.43 5.27e� 06 7.66e� 0539 Lymphocyte mediated immunity 0002449 64 13 3.13 4.15 9.90e� 06 0.000140 Response to organic substance 0010033 195 25 9.54 2.62 8.00e� 06 0.0001
Abbreviations: C¼number of reference genes in the category, O¼number of genes in the gene set and also in the category, E¼ expected number in the
category, R¼ ratio of enrichment; adj P¼ adjusted P value. BALB/c mice were conditioned with Bu–Cy regimen and underwent allogeneic or syngeneic SCT
(Materials and methods). Seven days after transplantation, genes that were exclusively upregulated in the liver of GVHD (allogeneic) compare with the liver of
syngeneic setting were analyzed to elucidate induced biological processes. Majority of induced biological processes were inflammatory and immune
responses.
Gene expression pattern in early phase of GVHD
B Sadeghi et al
5
& 2012 Macmillan Publishers Limited Bone Marrow Transplantation (2012), 1 – 10
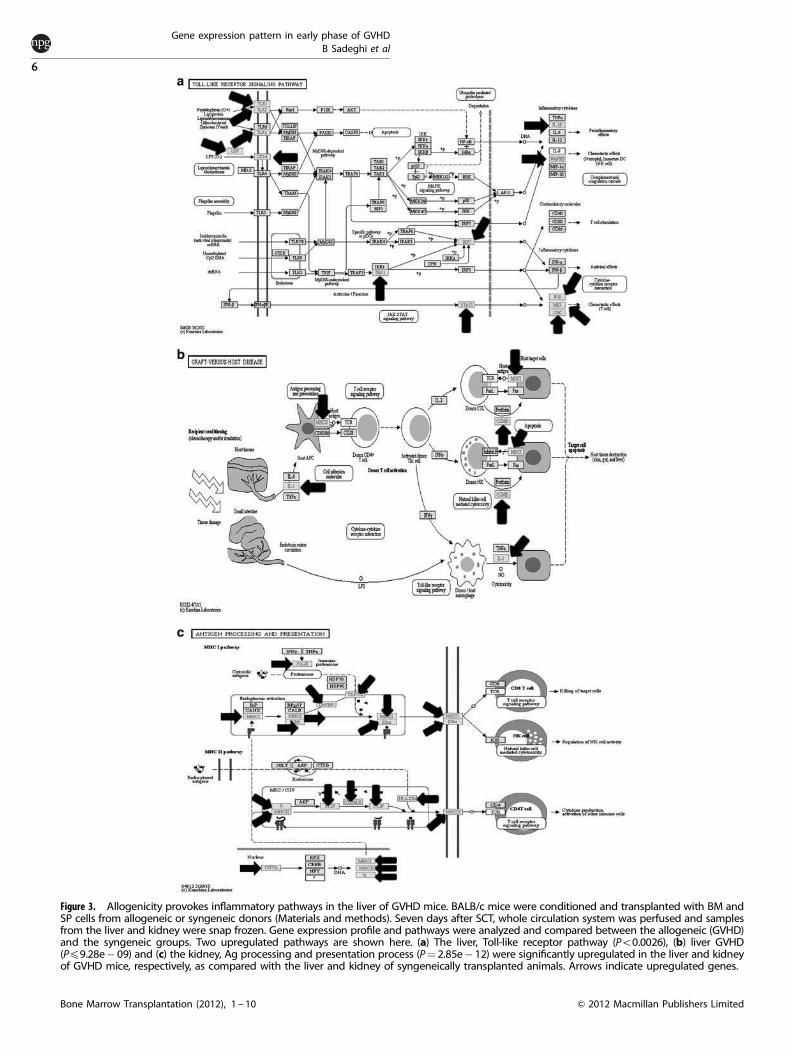
Figure 3. Allogenicity provokes inflammatory pathways in the liver of GVHD mice. BALB/c mice were conditioned and transplanted with BM andSP cells from allogeneic or syngeneic donors (Materials and methods). Seven days after SCT, whole circulation system was perfused and samplesfrom the liver and kidney were snap frozen. Gene expression profile and pathways were analyzed and compared between the allogeneic (GVHD)and the syngeneic groups. Two upregulated pathways are shown here. (a) The liver, Toll-like receptor pathway (Po0.0026), (b) liver GVHD(Pp9.28e� 09) and (c) the kidney, Ag processing and presentation process (P¼ 2.85e� 12) were significantly upregulated in the liver and kidneyof GVHD mice, respectively, as compared with the liver and kidney of syngeneically transplanted animals. Arrows indicate upregulated genes.
Gene expression pattern in early phase of GVHD
B Sadeghi et al
6
Bone Marrow Transplantation (2012), 1 – 10 & 2012 Macmillan Publishers Limited
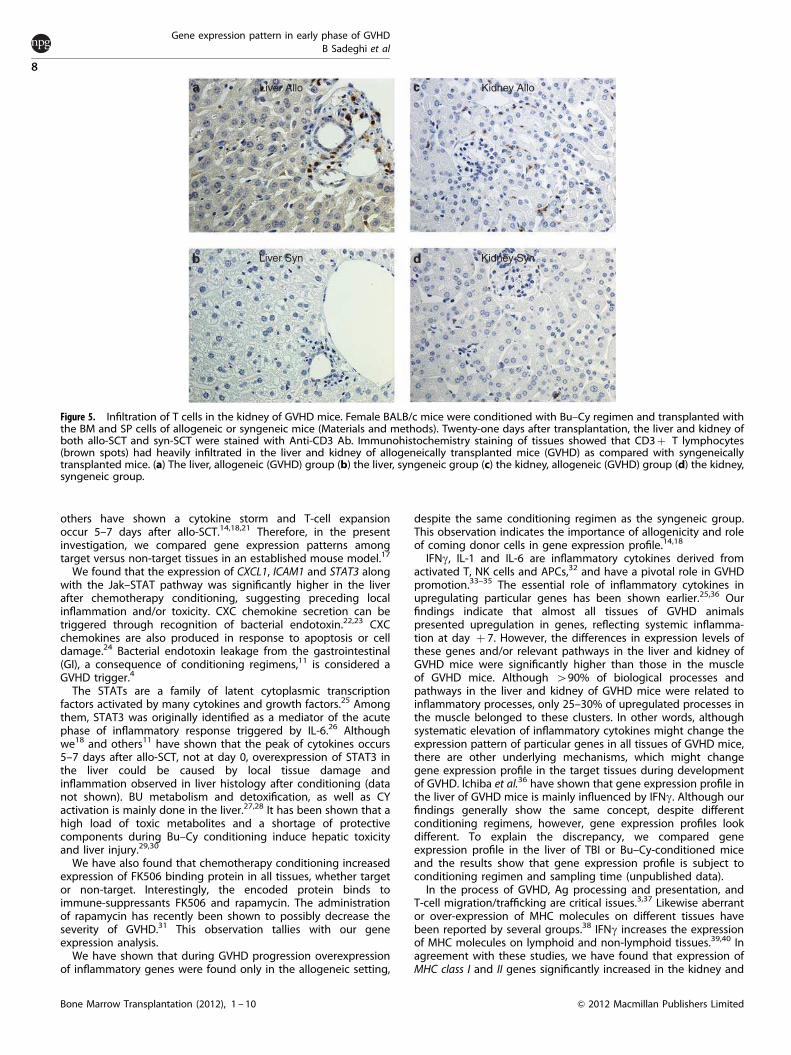
Twenty one days after allo-SCT, sections from the liver and kidneywere prepared and stained with antibodies against CD3þ
lymphocytes. Figure 5 demonstrates that the kidney and liver of
GVHD mice were infiltrated with T cells, while tissues from
syngeneically transplanted animals were not.
DISCUSSION
Several reports have shown that tissue damage due to condition-ing is important factor in triggering and developingGVHD.10,11,19,20 It is not clear why some tissues (for example, theliver and skin) are more vulnerable to GVHD damage.6 We and
Muscle
Kidney
Liver0.00.0
0.00.0
0.00.0
0.0
0.0
0.0
0.0
0.0
11.99.3
6.5
4.64.5
4.8
2.3
4.1
3.9
4.6
4.25.3
7.87.9
3.63.3
4.93.9
5.2
7.1
Muscle
Kidney
Liver
7.1Response to molecule of
bacterial origin
Leukocyte migration
Acute inflammatory response
Chemotaxis
Cytokine receptor binding
Chemokine activity
Adaptive immune response
Innate immune response
Inflammatory response
Immune response
Antigen processing and
presentation
0.0 2.0 4.0 6.0 8.0 10.0 12.0
Gen
es
Vcam1
Tnfaip3
Tlr2
Tlr13
Tlr1
Stat3
Stat2
Stat1
Socs3
Scara5
Saa3
Saa2
Saa1
Nkg7
Marco
Itgb3
Itgb2
Irg1
IL1r1
H2-DMb2
H2-DMb1
H2-Ab1
H2-Aa
Gzmk
CXCL9
CXCL11
CXCL1
CD74
CD14
CCL5
2 (19%)
M_SynM_dM_7
K-0
3 (10%)
L_0
M_AlloK_Allo
L_Allo
1 (47%)3 (15%)
2 (24%)
K-7
K_0
K_Syn
K_Allo
M_Allo
L_Syn
L_0L-7
M_0M-7
M_Syn
1 (36%)
L_Allo
L_Syn
L-7
K_7K_Syn
0 10 20 30 40 55 70 85100
Fold change
Figure 4. Upregulated genes and biological process in target tissues versus non-target tissues in GVHD-developing mice. BALB/c micewere conditioned and transplanted with an allogeneic or a syngeneic graft. GVHD developed in the allogeneic group. Seven days afterallo-SCT, upregulated genes and biologic processes in the liver, kidney and muscle of GVHD mice (adjusted with to the matched tissuesfrom day 0 and syngeneic group) were analyzed. Most of the inflammatory (a) genes (b) biologic processes were significantly overexpressedin the liver and kidney but not in the muscle of GVHD-developing animals. (a) Fold change indicates the ratio of increments as comparedwith parallel tissues after conditioning (day 0) and syngeneically transplanted mice. (b) Inflammatory process especially chemokine activitywere overexpressed both in the liver and in kidney but not in the muscle. Interestingly, biological processes related to leukocyte migrationand cell chemotaxis exclusively upregulated in the liver. (c, d) Target and non-target tissues were analyzed for (c) Ag processing andpresentation, as well as for (d) leukocyte migration pathways. Data are shown in principal component analysis plot. The liver and kidneys fromallogeneically transplanted animals show different patterns compared with the matched tissues from other groups, while the pattern of themuscle in all groups looks similar. No or very small changes is noticeable between allo-muscle and other groups.
Gene expression pattern in early phase of GVHD
B Sadeghi et al
7
& 2012 Macmillan Publishers Limited Bone Marrow Transplantation (2012), 1 – 10
others have shown a cytokine storm and T-cell expansionoccur 5–7 days after allo-SCT.14,18,21 Therefore, in the presentinvestigation, we compared gene expression patterns amongtarget versus non-target tissues in an established mouse model.17
We found that the expression of CXCL1, ICAM1 and STAT3 alongwith the Jak–STAT pathway was significantly higher in the liverafter chemotherapy conditioning, suggesting preceding localinflammation and/or toxicity. CXC chemokine secretion can betriggered through recognition of bacterial endotoxin.22,23 CXCchemokines are also produced in response to apoptosis or celldamage.24 Bacterial endotoxin leakage from the gastrointestinal(GI), a consequence of conditioning regimens,11 is considered aGVHD trigger.4
The STATs are a family of latent cytoplasmic transcriptionfactors activated by many cytokines and growth factors.25 Amongthem, STAT3 was originally identified as a mediator of the acutephase of inflammatory response triggered by IL-6.26 Althoughwe18 and others11 have shown that the peak of cytokines occurs5–7 days after allo-SCT, not at day 0, overexpression of STAT3 inthe liver could be caused by local tissue damage andinflammation observed in liver histology after conditioning (datanot shown). BU metabolism and detoxification, as well as CYactivation is mainly done in the liver.27,28 It has been shown that ahigh load of toxic metabolites and a shortage of protectivecomponents during Bu–Cy conditioning induce hepatic toxicityand liver injury.29,30
We have also found that chemotherapy conditioning increasedexpression of FK506 binding protein in all tissues, whether targetor non-target. Interestingly, the encoded protein binds toimmune-suppressants FK506 and rapamycin. The administrationof rapamycin has recently been shown to possibly decrease theseverity of GVHD.31 This observation tallies with our geneexpression analysis.We have shown that during GVHD progression overexpression
of inflammatory genes were found only in the allogeneic setting,
despite the same conditioning regimen as the syngeneic group.This observation indicates the importance of allogenicity and roleof coming donor cells in gene expression profile.14,18
IFNg, IL-1 and IL-6 are inflammatory cytokines derived fromactivated T, NK cells and APCs,32 and have a pivotal role in GVHDpromotion.33–35 The essential role of inflammatory cytokines inupregulating particular genes has been shown earlier.25,36 Ourfindings indicate that almost all tissues of GVHD animalspresented upregulation in genes, reflecting systemic inflamma-tion at day þ 7. However, the differences in expression levels ofthese genes and/or relevant pathways in the liver and kidney ofGVHD mice were significantly higher than those in the muscleof GVHD mice. Although 490% of biological processes andpathways in the liver and kidney of GVHD mice were related toinflammatory processes, only 25–30% of upregulated processes inthe muscle belonged to these clusters. In other words, althoughsystematic elevation of inflammatory cytokines might change theexpression pattern of particular genes in all tissues of GVHD mice,there are other underlying mechanisms, which might changegene expression profile in the target tissues during developmentof GVHD. Ichiba et al.36 have shown that gene expression profile inthe liver of GVHD mice is mainly influenced by IFNg. Although ourfindings generally show the same concept, despite differentconditioning regimens, however, gene expression profiles lookdifferent. To explain the discrepancy, we compared geneexpression profile in the liver of TBI or Bu–Cy-conditioned miceand the results show that gene expression profile is subject toconditioning regimen and sampling time (unpublished data).In the process of GVHD, Ag processing and presentation, and
T-cell migration/trafficking are critical issues.3,37 Likewise aberrantor over-expression of MHC molecules on different tissues havebeen reported by several groups.38 IFNg increases the expressionof MHC molecules on lymphoid and non-lymphoid tissues.39,40 Inagreement with these studies, we have found that expression ofMHC class I and II genes significantly increased in the kidney and
Liver Allo
Liver Syn Kidney Syn
Kidney Allo
Figure 5. Infiltration of T cells in the kidney of GVHD mice. Female BALB/c mice were conditioned with Bu–Cy regimen and transplanted withthe BM and SP cells of allogeneic or syngeneic mice (Materials and methods). Twenty-one days after transplantation, the liver and kidney ofboth allo-SCT and syn-SCT were stained with Anti-CD3 Ab. Immunohistochemistry staining of tissues showed that CD3þ T lymphocytes(brown spots) had heavily infiltrated in the liver and kidney of allogeneically transplanted mice (GVHD) as compared with syngeneicallytransplanted mice. (a) The liver, allogeneic (GVHD) group (b) the liver, syngeneic group (c) the kidney, allogeneic (GVHD) group (d) the kidney,syngeneic group.
Gene expression pattern in early phase of GVHD
B Sadeghi et al
8
Bone Marrow Transplantation (2012), 1 – 10 & 2012 Macmillan Publishers Limited

liver of GVHD mice, but remained unchanged or showed limitedincrease in the muscle of the same animal. Along with MHC classII, CLIP(CD74) is an important protein for Ag presentation. CLIP hasa key role in presenting antigens by DCs through MHC-IItrafficking, protein loading and DC migration.41 The expressionof CLIP is induced by IFNg.42 We have found that the expression ofCLIP is increased in the kidney and liver of GVHD mice. It seemsthat increased ability of Ag presenting by inhabitant APCs in thekidney and liver is an important process in the development ofGVHD in these organs.43
Leukocyte trafficking to various tissues is mediated by expres-sion of adhesion molecules ICAM-1 and VCAM-1 in endothelialcells.44,45 They have an important role in recruiting donor T cellsand initiating organ damage in GVHD pathogenesis.46,47 Ourfindings showed that the expression level of ICAM-1 was increasedin the liver and kidney 7 days after allo-SCT. Remarkably, VCAM-1expression was higher in the kidney than in the liver, whereas itdid not increase in the muscle of GVHD mice or in any tissues ofsyn-SCT animals. Several groups have explored and identified therole of various chemokines in T-cell migration and trafficking totarget tissues in the GVHD process.14,36,48 In line with theseobservations, we have found that expression levels of CXCL1,CXCL9, CXCL11 and CCL5 were increased after allo-SCT.Interestingly, whereas the expression of CXCL1 and CXCL13 washigher in the liver, CXCL9, CXCL11 and CCL5 were more highlyexpressed in the kidney. Different chemokine expression patternsin these tissues could be due to the tissue nature/physiology,intensity of previous damage, expression kinetics and/or level ofinhabitant immune cells. Interestingly, biological analysis hasshown that certain processes for example, leukocyte migrationand leukocyte chemotaxis, which are vital for early T-cell invasionand trafficking, were higher in the liver as compared with thekidney. This might explain the priority of the liver as the first placefor tissue damage in the GVHD. Further kinetic studies arewarranted.Last, histopathologic staining of the kidney of GVHD mice
confirmed the presence of T cells (similar to that observed in theliver) indicating T-cell invasion to this organ after developingGVHD. In summary, our data indicate that the gene expressionprofile of the kidney in allo-SCT is comparable to that of the liver.However, expression patterns of inflammatory processes andimmune response pathways in the kidney are different from thosein the liver (Figures 4a–d). It seems that these two organsexperience different pathways inciting inflammation and immuneresponse after allo-SCT and initiating the GVHD process. A conceptthat needs to be explored more in the future!
CONFLICT OF INTEREST
The authors declare no conflict of interest.
ACKNOWLEDGEMENTS
We would like to express their gratitude to Jennifer Usterud for reading the paper
and writing assistance. The study was supported by grants from Swedish Cancer
Foundation (Cancerfonden) and The Swedish Childhood Cancer Foundation
(Barncancerfonden).
REFERENCES
1 Copelan EA. Hematopoietic stem-cell transplantation. N Engl J Med 2006; 354:
1813–1826.
2 Mielcarek M, Martin PJ, Leisenring W, Flowers ME, Maloney DG, Sandmaier BM
et al. Graft-versus-host disease after nonmyeloablative versus conventional
hematopoietic stem cell transplantation. Blood 2003; 102: 756–762.
3 Shlomchik WD. Graft-versus-host disease. Nat Rev Immunol 2007; 7: 340–352.
4 Hill GR, Ferrara JL. The primacy of the gastrointestinal tract as a target organ of
acute graft-versus-host disease: rationale for the use of cytokine shields in allo-
geneic bone marrow transplantation. Blood 2000; 95: 2754–2759.
5 Glucksberg H, Storb R, Fefer A, Buckner CD, Neiman PE, Clift RA et al. Clinical
manifestations of graft-versus-host disease in human recipients of marrow from
HL-A-matched sibling donors. Transplantation 1974; 18: 295–304.
6 Teshima T, Ferrara JL. Understanding the alloresponse: new approaches to
graft-versus-host disease prevention. Semin Hematol 2002; 39: 15–22.
7 Kataoka Y, Iwasaki T, Kuroiwa T, Seto Y, Iwata N, Hashimoto N et al. The role of
donor T cells for target organ injuries in acute and chronic graft-versus-host
disease. Immunology 2001; 103: 310–318.
8 Kimura S, Horie A, Hiki Y, Yamamoto C, Suzuki S, Kuroda J et al. Nephrotic syn-
drome with crescent formation and massive IgA deposition following allogeneic
bone marrow transplantation for natural killer cell leukemia/lymphoma. Blood
2003; 101: 4219–4221.
9 Homma CI, Kami M, Masuo S, Sakiyama M, Kojima R, Hori A et al. Graft-versus-host
disease of the kidney after rapid tapering of cyclosporin following reduced
intensity hematopoietic stem cell transplantation. Bone Marrow Transplant 2005;
35: 929–930.
10 Holler E, Kolb HJ, Mittermuller J, Kaul M, Ledderose G, Duell T et al. Modulation of
acute graft-versus-host-disease after allogeneic bone marrow transplantation by
tumor necrosis factor alpha (TNF alpha) release in the course of pretransplant
conditioning: role of conditioning regimens and prophylactic application of a
monoclonal antibody neutralizing human TNF alpha (MAK 195F). Blood 1995; 86:
890–899.
11 Hill GR, Crawford JM, Cooke KR, Brinson YS, Pan L, Ferrara JL. Total body
irradiation and acute graft-versus-host disease: the role of gastrointestinal
damage and inflammatory cytokines. Blood 1997; 90: 3204–3213.
12 Ebert LM, Schaerli P, Moser B. Chemokine-mediated control of T cell traffic in
lymphoid and peripheral tissues. Mol Immunol 2005; 42: 799–809.
13 New JY, Li B, Koh WP, Ng HK, Tan SY, Yap EH et al. T cell infiltration and chemokine
expression: relevance to the disease localization in murine graft-versus-host dis-
ease. Bone Marrow Transplant 2002; 29: 979–986.
14 Mapara MY, Leng C, Kim YM, Bronson R, Lokshin A, Luster A et al. Expression
of chemokines in GVHD target organs is influenced by conditioning and
genetic factors and amplified by GVHR. Biol Blood Marrow Transplant 2006; 12:
623–634.
15 Ma HH, Ziegler J, Li C, Sepulveda A, Bedeir A, Grandis J et al. Sequential activation
of inflammatory signaling pathways during graft-versus-host disease (GVHD):
early role for STAT1 and STAT3. Cell Immunol 2011; 268: 37–46.
16 Halloran PF, Autenried P, Ramassar V, Urmson J, Cockfield S. Local T cell responses
induce widespread MHC expression. Evidence that IFN-gamma induces its own
expression in remote sites. J Immunol 1992; 148: 3837–3846.
17 Sadeghi B, Aghdami N, Hassan Z, Forouzanfar M, Rozell B, Abedi-Valugerdi M et al.
GVHD after chemotherapy conditioning in allogeneic transplanted mice. Bone
Marrow Transplant 2008; 42: 807–818.
18 Sadeghi B, Al-Hashmi S, Hassan Z, Rozell B, Concha H, Lundmark C et al. Expansion
and activation kinetics of immune cells during early phase of GVHD in
mouse model based on chemotherapy conditioning. Clin Dev Immunol 2010;
2010: 142943.
19 Perez-Simon JA, Diez-Campelo M, Martino R, Brunet S, Urbano A, Caballero MD
et al. Influence of the intensity of the conditioning regimen on the characteristics
of acute and chronic graft-versus-host disease after allogeneic transplantation.
Br J Haematol 2005; 130: 394–403.
20 Ferrara JL, Cooke KR, Teshima T. The pathophysiology of acute graft-versus-host
disease. Int J Hematol 2003; 78: 181–187.
21 Beilhack A, Schulz S, Baker J, Beilhack GF, Wieland CB, Herman EI et al. In vivo
analyses of early events in acute graft-versus-host disease reveal sequential
infiltration of T-cell subsets. Blood 2005; 106: 1113–1122.
22 Moore TA, Newstead MW, Strieter RM, Mehrad B, Beaman BL, Standiford TJ.
Bacterial clearance and survival are dependent on CXC chemokine receptor-2
ligands in a murine model of pulmonary Nocardia asteroides infection. J Immunol
2000; 164: 908–915.
23 Eisele NA, Lee-Lewis H, Besch-Williford C, Brown CR, Anderson DM. Chemokine
receptor CXCR2 mediates bacterial clearance rather than neutrophil recruitment
in a murine model of pneumonic plague. Am J Pathol 2011; 178: 1190–1200.
24 Hotchkiss RS, Nicholson DW. Apoptosis and caspases regulate death and
inflammation in sepsis. Nat Rev Immunol 2006; 6: 813–822.
25 Levy DE, Darnell Jr JE. Stats: transcriptional control and biological impact. Nat Rev
Mol Cell Biol 2002; 3: 651–662.
26 Ihle JN. The Stat family in cytokine signaling. Curr Opin Cell Biol 2001; 13: 211–217.
27 Hassan M, Ehrsson H. Metabolism of 14C-busulfan in isolated perfused rat liver.
Eur J Drug Metab Pharmacokinet 1987; 12: 71–76.
28 de Jonge ME, Huitema AD, Rodenhuis S, Beijnen JH. Clinical pharmacokinetics of
cyclophosphamide. Clin Pharmacokinet 2005; 44: 1135–1164.
29 Sadeghi B, Jansson M, Hassan Z, Mints M, Hagglund H, Abedi-Valugerdi M et al.
The effect of administration order of BU and CY on engraftment and toxicity in
HSCT mouse model. Bone Marrow Transplant 2008; 41: 895–904.
Gene expression pattern in early phase of GVHD
B Sadeghi et al
9
& 2012 Macmillan Publishers Limited Bone Marrow Transplantation (2012), 1 – 10

30 DeLeve LD, Wang X. Role of oxidative stress and glutathione in busulfan toxicity
in cultured murine hepatocytes. Pharmacology 2000; 60: 143–154.
31 Rosenbeck LL, Kiel PJ, Kalsekar I, Vargo C, Baute J, Sullivan CK et al. Prophylaxis
with sirolimus and tacrolimus þ /- antithymocyte globulin reduces the risk of
acute graft-versus-host disease without an overall survival benefit following
allogeneic stem cell transplantation. Biol Blood Marrow Transplant 2011; 17:
916–922.
32 Yang YG, Wang H, Asavaroengchai W, Dey BR. Role of Interferon-gamma in GVHD
and GVL. Cell Mol Immunol 2005; 2: 323–329.
33 Deeg HJ. Cytokines in graft-versus-host disease and the graft-versus-leukemia
reaction. Int J Hematol 2001; 74: 26–32.
34 Symington FW, Symington BE, Liu PY, Viguet H, Santhanam U, Sehgal PB. The
relationship of serum IL-6 levels to acute graft-versus-host disease and hepator-
enal disease after human bone marrow transplantation. Transplantation 1992; 54:
457–462.
35 Hill GR, Cooke KR, Brinson YS, Bungard D, Ferrara JL. Pretransplant chemotherapy
reduces inflammatory cytokine production and acute graft-versus-host
disease after allogeneic bone marrow transplantation. Transplantation 1999; 67:
1478–1480.
36 Ichiba T, Teshima T, Kuick R, Misek DE, Liu C, Takada Y et al. Early changes in gene
expression profiles of hepatic GVHD uncovered by oligonucleotide microarrays.
Blood 2003; 102: 763–771.
37 Ferrara JL, Reddy P. Pathophysiology of graft-versus-host disease. Semin Hematol
2006; 43: 3–10.
38 Mason DW, Dallman M, Barclay AN. Graft-versus-host disease induces expression
of Ia antigen in rat epidermal cells and gut epithelium. Nature 1981; 293: 150–151.
39 Schiltz PM, Gomez GG, Read SB, Kulprathipanja NV, Kruse CA. Effects of
IFN-gamma and interleukin-1beta on major histocompatibility complex antigen
and intercellular adhesion molecule-1 expression by 9L gliosarcoma: relevance to
its cytolysis by alloreactive cytotoxic T lymphocytes. J Interferon Cytokine Res 2002;
22: 1209–1216.
40 Collins T, Korman AJ, Wake CT, Boss JM, Kappes DJ, Fiers W et al. Immune
interferon activates multiple class II major histocompatibility complex genes and
the associated invariant chain gene in human endothelial cells and dermal
fibroblasts. Proc Natl Acad Sci USA 1984; 81: 4917–4921.
41 Faure-Andre G, Vargas P, Yuseff MI, Heuze M, Diaz J, Lankar D et al. Regulation of
dendritic cell migration by CD74, the MHC class II-associated invariant chain.
Science 2008; 322: 1705–1710.
42 Rahmsdorf HJ, Harth N, Eades AM, Litfin M, Steinmetz M, Forni L et al. Interferon-
gamma, mitomycin C, and cycloheximide as regulatory agents of MHC class
II-associated invariant chain expression. J Immunol 1986; 136: 2293–2299.
43 Shlomchik WD, Couzens MS, Tang CB, McNiff J, Robert ME, Liu J et al. Prevention
of graft versus host disease by inactivation of host antigen-presenting cells.
Science 1999; 285: 412–415.
44 Orito H, Fujimoto M, Ishiura N, Yanaba K, Matsushita T, Hasegawa M et al.
Intercellular adhesion molecule-1 and vascular cell adhesion molecule-1 coop-
eratively contribute to the cutaneous Arthus reaction. J Leukoc Biol 2007; 81:
1197–1204.
45 Ebnet K, Kaldjian EP, Anderson AO, Shaw S. Orchestrated information transfer
underlying leukocyte endothelial interactions. Annu Rev Immunol 1996; 14: 155–177.
46 Norton J, Sloane JP, al-Saffar N, Haskard DO. Expression of adhesion molecules in
human intestinal graft-versus-host disease. Clin Exp Immunol 1992; 87: 231–236.
47 Itoh S, Matsuzaki Y, Kimura T, Unno R, Ikegami T, Shoda J et al. Suppression of
hepatic lesions in a murine graft-versus-host reaction by antibodies against
adhesion molecules. J Hepatol 2000; 32: 587–595.
48 Bouazzaoui A, Spacenko E, Mueller G, Miklos S, Huber E, Holler E et al. Chemokine
and chemokine receptor expression analysis in target organs of acute graft-
versus-host disease. Genes Immun 2009; 10: 687–701.
Gene expression pattern in early phase of GVHD
B Sadeghi et al
10
Bone Marrow Transplantation (2012), 1 – 10 & 2012 Macmillan Publishers Limited




















