
Production and Characterization of Monoclonal Antibodies to Purified Deglycosylated Cystic Fibrosis...
10
Tor&at Vol . 29, No. 10, pp . 1195-1204, 1991 . Printed i° Great Britain . PRODUCTION AND CHARACTERIZATION OF MONOCLONAL ANTIBODIES AGAINST NAJA NAJA ATRA COBROTOXIN BRADLEY G. STILES, 1 BURTON C . LII~GERDING,2 FRANCIS W . SEXTON I and $EIAWN B . GuFST2 'Department of Toxinology, Pathophysiology Division; and Virology Division ; U.S . Army Modical Research Institute of Infectious Diseases, Frederick, MD 21702-5011, U.S .A . (Received 7 April 1991 ; accepted 30 May 1991) INTRODUCTION n Ptm pk B . G . STILES, B . C . LIDGERDING, F . W. SE7CTON and S. B . GUEST . Production and characterization of monoclonal antibodies against Naja raja atra cobrotoxin . Toxicon 29, 1195-1204, 1991 .-Twelve monoclonal antibodies against cobrotoxin from Naja raja atra venom were tested for cross-reactivity with eight different snake toxins, binding to linear epitopes, prevention of cobrotoxin binding to acetylcholine receptor (AchR) in vitro, and protection in mice concomitantly given a lethal dose of cobrotoxin . The antibodies were highly specific, as evidenced by little reactivity with other snake toxins . None of the monoclonal antibodies bound to reduced cobrotoxin or synthesized 8-mer regions spanning the whole molecule, thus suggesting the recognition of conformational epitopes . The in vitro binding of toxin to AchR was competitively inhibited (23-79%) with a 1 .66 :1 mole ratio of antibody : AchR . Preincubation of monoclonal antibody with toxin before adding AchR (3 : 1 mole ratio of AchR : antibody) inhibited the in vitro binding of toxin to AchR by 20-80% . Monoclonal antibodies added after the preincubation of toxin with AchR did not dissociate the toxin-AchR complex . An antibody : toxin mole ratio of 2 .5 : 1, with 6 rug of cobrotoxin, delayed the time to death of mice 3 .7-23 .8-fold over control mice . The monoclonal antibodies that most effectively prevented in vitro binding of toxin to AchR also provided the longest delay in time to death in mice . COBROTOXTN, isolated from the venom of Naja raja atra, is a short-chain postsynaptic neurotoxin composed of 62 amino acids and is dependent on four disulfide bonds for biological activity (YANG, 1965, 1974 ; YANG et al., 1969, 1970) . The long-chain post- synaptic neurotoxins, such as a-bungarotoxin, differ in structure by having 70-73 amino acids and five disulfide bonds (YANG, 1974) . Snake venom postsynaptic neurotoxins have homologous amino acid regions and consequently have similar conformations and binding affinity constants for acetylcholine receptor (AchR) (WESER and CHANGEUX, 1974 ; YANG, 1974; WALKINSHAW et al., 1980; DUFTON and HIRER, 1983 ; Low and CORFIELD, 1986) . Various investigators have studied the binding of snake venom postsynaptic neurotoxin regions to AchR . JUILLERAT et al. (1982) showed that a peptide loop region (amino acids 1648) of a Naja raja philippinensis short-chain neurotoxin binds AchR with more affinity 1195
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Production and Characterization of Monoclonal Antibodies to Purified Deglycosylated Cystic Fibrosis...

Tor&at Vol . 29, No. 10, pp . 1195-1204, 1991 .Printed i° Great Britain .
PRODUCTION AND CHARACTERIZATION OF MONOCLONALANTIBODIES AGAINST NAJA NAJA ATRA COBROTOXIN
BRADLEY G. STILES, 1 BURTON C. LII~GERDING,2 FRANCIS W. SEXTON I and $EIAWN B .GuFST2
'Department of Toxinology, Pathophysiology Division; and Virology Division ; U.S . Army Modical ResearchInstitute of Infectious Diseases, Frederick, MD 21702-5011, U.S .A .
(Received 7 April 1991 ; accepted 30 May 1991)
INTRODUCTION
n Ptm pk
B. G. STILES, B . C . LIDGERDING, F. W. SE7CTON and S. B. GUEST . Productionand characterization of monoclonal antibodies against Naja raja atracobrotoxin . Toxicon 29, 1195-1204, 1991 .-Twelve monoclonal antibodiesagainst cobrotoxin from Naja raja atra venom were tested for cross-reactivitywith eight different snake toxins, binding to linear epitopes, prevention ofcobrotoxin binding to acetylcholine receptor (AchR) in vitro, and protection inmice concomitantly given a lethal dose of cobrotoxin . The antibodies werehighly specific, as evidenced by little reactivity with other snake toxins . Noneof the monoclonal antibodies bound to reduced cobrotoxin or synthesized8-mer regions spanning the whole molecule, thus suggesting the recognition ofconformational epitopes . The in vitro binding of toxin to AchR wascompetitively inhibited (23-79%) with a 1 .66 :1 mole ratio ofantibody : AchR . Preincubation of monoclonal antibody with toxin beforeadding AchR (3 : 1 mole ratio of AchR : antibody) inhibited the in vitrobinding of toxin to AchR by 20-80% . Monoclonal antibodies added after thepreincubation of toxin with AchR did not dissociate the toxin-AchR complex.An antibody : toxin mole ratio of 2.5 : 1, with 6 rug of cobrotoxin, delayed thetime to death of mice 3.7-23.8-fold over control mice . The monoclonalantibodies that most effectively prevented in vitro binding of toxin to AchRalso provided the longest delay in time to death in mice .
COBROTOXTN, isolated from the venom of Naja raja atra, is a short-chain postsynapticneurotoxin composed of 62 amino acids and is dependent on four disulfide bonds forbiological activity (YANG, 1965, 1974 ; YANG et al., 1969, 1970). The long-chain post-synaptic neurotoxins, such as a-bungarotoxin, differ in structure by having 70-73 aminoacids and five disulfide bonds (YANG, 1974). Snake venom postsynaptic neurotoxins havehomologous amino acid regions and consequently have similar conformations andbinding affinity constants for acetylcholine receptor (AchR) (WESER and CHANGEUX, 1974 ;YANG, 1974; WALKINSHAW et al., 1980; DUFTON and HIRER, 1983 ; Low and CORFIELD,1986).Various investigators have studied the binding of snake venom postsynaptic neurotoxin
regions to AchR . JUILLERAT et al. (1982) showed that a peptide loop region (amino acids1648) of a Naja raja philippinensis short-chain neurotoxin binds AchR with more affinity
1195

1196
B. G. STILES et at .
than acetylcholine, but less than native toxin. A peptide loop region from a-bungarotoùn(amino acids 26-41) also recognizes AchR with lesser affinity than the native toxin(McDArI>EL et al., 1987). Other binding studies with a-bungarotoùn have used AchRpeptides from the a subunit (MuLAC-JERICEVIC and ATASSI, 1987; Mur..AC-JERICEVIC et al .,1988), as well as three different loop regions from a-bungarotoùn which include aminoacids 3-16, 261, and 45-59 (RUAN et al., 1990). The highest affinity of an a-bungaro-toxin peptide (amino acids 26-41) for AchR peptides was approximately 10-fold less thannative toxin for the same receptor peptides .
Various immunological studies with snake venom postsynaptic neurotoains havedemonstrated neutralizing polyclonal antibodies that cross-react with other snake neuro-toxins (MENEZ, 1985). One molecule of cobrotoxin or Naja nigricollis toùn a accommo-dates three or four antibody molecules, respectively (CHANG and YANG, 1969; YANG et al.,1978 ; MENEZ et al., 1979). Antibody against Laticauda semifasciata venom or purifiederabutoùn b cross-reacts with many different short-chain postsynaptic neurotoùns(BoQuET et al ., 1973 ; ABE and TAMnrA, 1979), except cobrotoxin, which shares 58% aminoacid homology with erabutoxin b (YANG et al., 1969 ; ABE and TAMIxA, 1979).More recent studies have been done with monoclonal antibodies against snake venom
postsynaptic neurotoùns (BOULAIN et al., 1982; DANSE et al., 1986 ; ia:iUANG et al., 1989 ;KASE et al ., 1989 ; PACHNER and RICALTüN, 1989). Three different investigations haveresulted in monoclonal antibodies that neutralize a-bungarotoùn in vivo (DANSE et al .,1986; CwuANC et al., 1989 ; KASE et al ., 1989). These antibodies were developed againstformylated or native a-bungarotoùn (DANSE et al ., 1986; CY~uANC et al., 1989 ; KASE et al.,1989). A neutralizing epitope on a-bungarotoùn was identified within amino acids 34 1(CFILJANG et al., 1989) and 34-38 (KASE et al., 1989). This is also the same region of thetoxin that binds AchR peptides with the highest affinity (RUAN et al., 1990). Themonoclonal antibody which recognized amino acids 34-38 (SSRGK) of a-bungarotoùnalso neutralized other long-chain newotoxins containing an identical sequence, but notclosely related neurotoùns differing by only one amino acid (S-35 to I-35) (KASE et al.,1989).The few monoclonal antibody studies done with snake venom short-chain postsynaptic
neurotoxins include those of BOULAIN et al. (1982) and TREMFAU et al. (1986) . Bothinvestigations resulted in different monoclonal antibodies against N. nigricollis toùn awhich prevented the in vitro binding of toxin to AchR and neutralized the lethal effects oftoxin a in mice . The antibodies differed in epitope recognition and specificity, with oneantibody binding to seven different short-chain, but no long-chain, neurotoùns (TREMF.AUet al., 1986). Although there are various reports on the neutralizing capabilities ofpolyclonal antisera against N. n. atra cobrotoxin (GIANG and YANG, 1969, 1973; YANGet al., 1977), there is only one report of a monoclonal antibody against cobrotoxin (YANG,1985) but no characterization work was done regarding binding specificity and toxinneutralization .
This study evaluates 12 monoclonal antibodies against N. n. atra cobrotoxin and theirability to cross-react with other snake venom toùns, prevent in vitro binding of cobratoxin to AchR, neutralize cobrotoxin in vivo, and finally react with linear epitopes oncobrotoxin .
MATERIALS AND METHODS
Cett linesThe non-immunoglobulin-secreting marine plasmacyloma, S1?2/0-Agl4 cell line (Sttut.~wN er d., 1978) and all

Cobrotoxin Monoclonal Antibodies
1197
developed clones were grown in Optimem medium (Gibco/BRL, Grand Island, NY, U.S.A .) with 7.5% fetalbovine serum, 2mM glutamine, 50 kg/ml gentamycin (Whiltaker Bioproducts, Walkersvilk, MD, U.S.A .),2 pg/ml Fungizone (Giboo/BRL), and 15% conditioned medium . Conditioned medium was prepared by filteringEagle's minimum easeatial medium from 7-14-day-old MRC-5 cells through a 0.22-fan filter (Millipore .Bedford, MA, U.S.A.). MRC-5 cells were obtained from the Salk Institute (Swiftwater, PA, U.S .A .), andmaintained is antibiotio-free medium .
Monoclonal antibody productionMale BALB/c mice (25 g) were each injected i.p . with 1 Kg of N . n. afro cobrotoxin mixed in 200 pl of RIBi
adjuvant (Hamilton, MT, U.S.A .). Mice were injected three times, once every week, and antibody filets weretested by an enzyme-linked immunosorbent assay (ELISA) as described below. Spleens were taken from micekilled three days after a final boost and fused with SP2/0-Ag14 cells at a 1 : 2 ratio using 50% (v/v) polyethYleneglycol 1500 MW (Boehringer-Mannheim, Indianapolis, IN, U.S.A.) as the fusogen (F.~x~nr and Osr©eurrc,1985) . Fused cells were plated in 9lrwell plates and grown in Optimem medium containing hypoxanthine,atainopterin, and thymidine (Boehringer-Mannheim) for 14 days to select for antibody-producing cells .
Production of antibody against tobrotoxin by hybridomas was detected with an ELISA as described below.Antibody-producing cells were cloned twice by the limiting dilution method of RF.rvea et al. (1985) without afeeder layer. The sub-isotype of the monoclonal antibodies was determined on cell culture fluids by ELISA withmonospectifit antisera (Hyclone, Logen, UT, U.S.A .) . Selected cloned hybridomas were grown in BALB/c strainCJ mice to produce oscine fluid . Monoclonal antibodies were purified from ascàtes fluid by affinity chromato-graphy with protein G-agarose (Pharmacie, Piacataway, NJ, U.S.A.), dialysed against phosphate buffeted saline(PBS; pH 7.4) and stored at -70°C.
Frtzymt-linked imnwnosorbcnt assays (ELISA)Antibody titers against cobrotoxin in mice were monitored with an ELISA by coating each well on an
Immulon II miaotiter plate (Dynatech, Chantilly, VA, U.S.A .) with 100 pl of cobrotoxin (2 pg/ml carbonatebuffer, pH 9.6) overnight at 4°C. Unoccupied sites in each well were then blocked with 300 pl ofPBS containing1% gelatin (PBSG) for 30 min at 37°C . Mouse sorom was diluted 1 : 100 in PBS containing 0.1 % Tween 20 and0.1% gelatin (PHSTG), and added (100ît1/well) for lhr at 37°C . Wells were washed four times with PBScontaining 0.1% Tween 20 (PBST), 100 Ill of PHSTG containing anti-mouse antibody conjugated with alkalinephosphatage (Sigma, St Louis, MO, U.S.A .) was added, and incubated for 1 hr at 37°C . Wells were then washedfive times with PHST, 100It1 of pare-nitrophenyl phosphate substrate (Kirkegaard and Perry Laboratories,Gaithersburg, MD, U.S.A .) was added, and the absorbance was read at 405 nm after 30 min at roomtemperature .
Detection of hybridomas producing antibody against cobrotoxin was accomplished by initially coating eachImmulon II nticrotiter well with 100 pl of a 1 l+g oobrotoxinJml carbonate buffer solution (pH 9.6) overnight at4°C. After blocking unoccupied sites for 30 thin at 37°C with PHSG, 100It1 of undiluted growth mediumooataining antibodies was added per well and incubated 1 hr at 37°C . Wells were then washed twice andanti-mouse antibody conjugated with alkaline phosphatage in PHSTG was added for 1 hr at 37°C . Wells werefinally washed four times, substrate added for 30 min, and the absorbance read at 405 tun.The cross-reactivity of cobrotoxin monoclonal antibodies with purified snake venom toxins (toxins were
purchased from Natural Product Sciences, Salt Lake City, UT, U.S.A.) wan tested with crotamine and crotoxinfrom Crotalus drtrisrus terriftcus, cardiotoxin and cobrotoxin from Naja raja afro, ß-bungarotoxin fromBungarvs multicinctus, and erabutoxin b from Laticauda semijasciata . Purified N. n. oxiana neurotoxins I and II,and B. tratlticinctus a-bungarotoxin (Sigma) were also tested. All toxins were adsorbed onto Immuloa IImicrotiter wells at 4°C overnight with a 10 pg toxin/ml carbonate buffer solution (pH 9.6 ; 100 Ill/well) .Monoclonal antibodies (100 Ill/well of a 10 pg/nil PHSTG solution) were then added and the cross-reactivity wasdetermined by an ELISA as previously described for mouse serum.
Acrtylchollne receptor assaysAcetyk holine receptor (AchR) assays were done essentially as previously described (Snt.>:s, 1991). Monoclonal
antibodies were diluted in PHSTG and 100 p1 was added to wells containing adsorbed cobroloxin (10014 of al0ltg toxin/ml carbonate buffer solution; pH 9.6) I hr before, during, or after the addition of AchR (50ItglmlPHSTG) to the plate . The receptor bound to toxin was detected with a 1 : l00 dilution ofguinea-pig anti-AchRincubated for 1 hr at room temperature. After washing twice with PBST, 100It1 of and-guinea-pig antibodyalkaline phosphatage conjugate (Sigma) was added per well for an additional 1 hr at room temperature. Theplate was finally washed five rimes, substrate added, and absorbance read at 405 nm.

1198
H. G. STILES et al.
TABLE l . REACTIVITY OF MONOCLONAL ANTIBODR~3 (I pg/well) WITHCOBROTOXIN (I lig~WCII)
Control wells contained blocking solution (1% gelatin),monoclonal antibody, conjugate and substrate . Mouse hyperimmuneand preimmune sera (1 :100 of each) control readings for nativeCobrotoxin were >2.800 and 0.05610.006, respectively . Mousehyperimmune and preimmune sera (1 : 100 of each) control readingsfor reduced Cobrotoxin were 0.30610.040 and 0.00710.005,respectively . All antibodies were tested in triplicate and the averageand S.D. determined .
Neutralization assaySwiss Webster mice (23 g1I .8 g) were each injected i.p . with 300 pl of wbrotoxin diluted in PBS (6 ~g/mouse)
and monoclonal antibody (200pg/ml or 1000 pgJml PBS final dilution) . Toxin and antibody were preincubatodtogether for 30 min at room temperature before the injections.
TABLE 2. INHIBTTION OF TOXIN~ACIIR COMPLEXING (PREINCUBATION OF TOXIN WTTHANTIBODY BEFORE AchR)
" Poncent competition (% Comp) was computed by comparing test wellscontaining 1 Rg wbrotoxin, 5 pg AchR, and monoclonal antibody with negativecontrol wells containing 1 pg Cobrotoxin, 5 kg AchR, andmouse preimmune serum(1 : 100). Negative control values were OD,~ = 2.08910.035 . Positive Control wellscontained 1 Kg Cobrotoxin, 5 Ng AchR, plus mouse hyperimmune serum (1 : (00) andresulted in an OD,~ value of 0.52810.026. All antibodies were tested in triplicateand the average and S.D. calculated.
Antibody 10 pg/ml (% Comp)" 100 ng/ml (% Comp) 1 ng/ml (% Comp)
1B93 1 .67110.066 (20) 2.087±0 .188 (0) 2.3I810.129 (0)1SE1 1 .65210.048 (21) 1.92710 .054 (4) 2.17510.153 (0)2A7 1 .63210.060 (22) 1.87010.030 (10) 2.068 t0.050 (1)6A10 1 .47410.056 (29) 2.00810.I13 (4) 2.19210.092 (0)13H7 1 .39510.013 (33) 1.91110.139 (9) 2.20410.049 (0)13H8 1 .37710.112 (34) 2.00810.106 (4) 2.21410.069 (0)ISF2 1 .25110.050 (40) I.92110.072 (8) 2.04710.123 (2)1OD12 1 .096t0.163 (47) I.83110.134 (12) 2.10510.096 (0)9B5 1 .07410.271 (48) 1.98810.045 (4) 2.15210.069 (0)IB92 0.81810.152 (61) 1.58810.095 (9) 2.10310.071 (0)1D8 0.65210.124 (71) l.79810.066 (14) 2.05010 .1 13 (1)BH5 0.45710.041 (78) 1.73210.167 (17) I .95710.177 (6)lE8 0.41510.096 (80) 1.76510.014 (15) 2.042 f0.133 (2)
Antibody Cobrotoxin
OD,~
Reduced Cobrotoxin Control'
2A7 0.811 ±0.022 0.01710.010 0.00310.001lE8 0.905±0.013 0.08010.010 0.00410.002IOD12 1 .0I210.01 l 0.00010.000 0.004±0.0011B93 1.05710.040 0.042 ±0.012 0.01010.0036A10 1 .06610.116 0.01610.012 0.02310.0101D8 1 .10510.038 0.07610.003 0.03310.0058H5 I .11010.009 0.07210.001 0.01610.00213H7 1 .29810.027 0.01610.007 0.00910.00413H8 I .40010.079 0.0161O.O14 0.02810.01115E1 1 .707±0.032 0.02410.015 0.025±0.0039B5 1 .79210.053 0.085f0.028 0.03910.013ISF2 I .90210.020 0.08810.004 0.00810.002
Cobrotoxin Monoclonal Antibodies
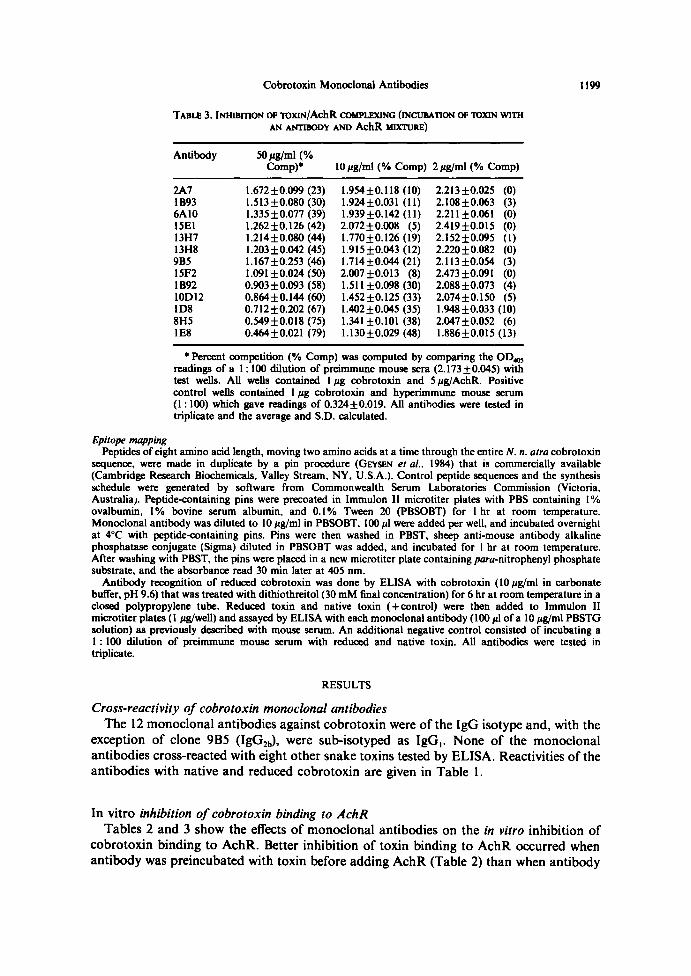
TABI~ 3. hvlliBmoN oe Toxnv/AchR coun't..exxtNC (tNCUBAnoN oF ToxIN wlrllAN ANTIBODY AND AC11R è®tTUAE)
RESULTS
' Percent competition (% Comp) was computed by comparing the OD,~readings of a 1 : 100 dilution of preimmune mouse sera (2.17310.045) withtest wells . All wells contained I ug cobrotoxin and 5 pg/AchR. Positivecontrol wells contained 1 Pg cobrotoxin and hyperimmune mouse serum(1 : 100) which gave readings of 0.32410.019. All antibodies were tested intriplicate and the average and S.D . calculated .
1199
Epilope mappingPeptides ofeight amino acid length, moving two amino acids at a time through the entire N. n. atra cobrotoxin
sequence, were made in duplicate by a pin procedure (GetrseN et ai., 1984) that is commercially available(Cambridge Research Biochemicals, Valley Stream, NY, U.S.A .) . Control peptide sequences and the synthesisschedule were generated by software from Commonwealth Serum Laboratories Commission (Victoria,Australia . Peptide-containing pins were precoated in Immulon II microtiter plates with PBS containing Iovalbumin, 1 % bovine serum albumin, and 0.1 % Tween 20 (PBSOBT) for I hr at room temperature .Monoclonal antibody was diluted to 10 ug/ml in PBSOBT . 100 pl were added per well, and incubated overnightat 4°C with peptide~ontaining pins. Pins were then washed in PBST, sheep anti-mouse antibody alkalinephosphatase conjugate (Sigma) diluted in PBSOBT was added, and incubated for I hr at room temperature .After washing with PBST, the pins were placed in a new microtiter plate containing para-nitrophenyl phosphatesubstrate, and the absorbance read 30 min later at 405 nm .
Antibody recognition of reduced cobrotoxin was done by ELISA with cobrotoxin (10 Pg/ml in carbonatebuffer, pH 9.6) that was treated with dithiothreitol (30 mM final concentration) for 6 hr at room temperature in aclosed polypropylene tube . Reduced toxin and native toxin (+control) were then added to Immulon IImicrotiter plates ( I pgJwell) and assayed by ELISA with each monoclonal antibody (100 pl ofa 10 pg/ml PBSTGsolution) as previously described with mouse serum. An additional negative control consisted of incubating aI : 100 dilution of preimmune mouse serum with reduced and native toxin. All antibodies were tested intriplicate.
Cross-reactivity of cobrotoxin monoclonal antibodiesThe 12 monoclonal antibodies against cobrotoxin were of the IgG isotype and, with the
exception of clone 9B5 (IgGZ~, were sub-isotyped as IgG, . None of the monoclonalantibodies cross-reacted with eight other snake toxins tested by ELISA. Reactivities of theantibodies with native and reduced cobrotoxin are given in Table 1 .
In vitro inhibition of cobrotoxin binding to AchRTables 2 and 3 show the effects of monoclonal antibodies on the in vitro inhibition of
cobrotoxin binding to AchR. Better inhibition of toxin binding to AchR occurred whenantibody was preincubated with toxin before adding AchR (Table 2) than when antibody
Antibody 50 Ng/ml (%Comp)' l0 pg/ml (% Comp) 2ieg/ml (% Comp)
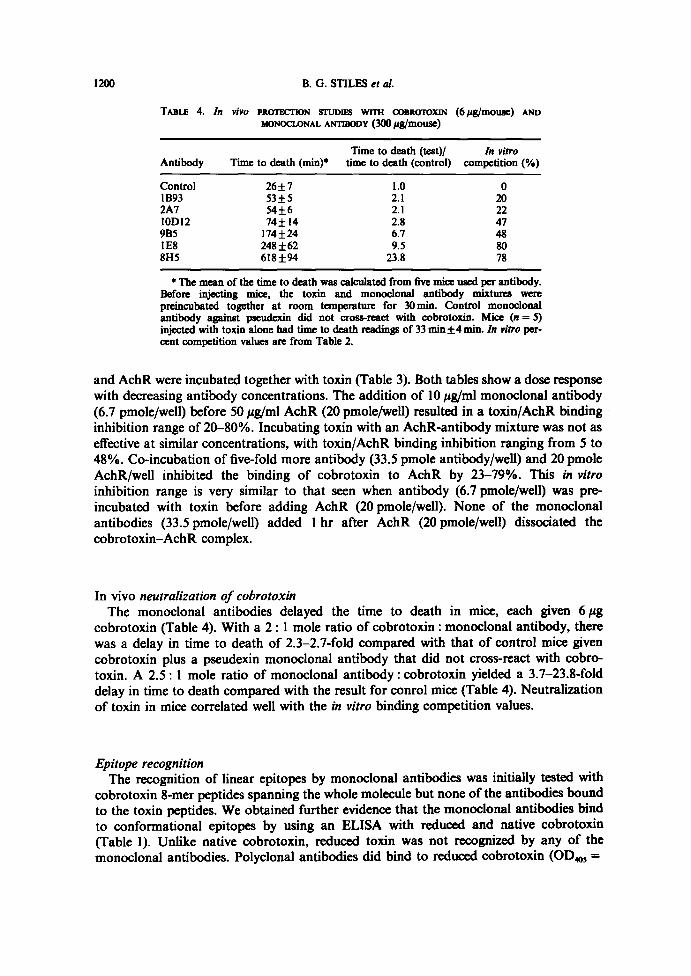
2A7 1.67210.099 (23) 1.95410.118 (10) 2.21310 .025 (0)1B93 1.513±0.080 (30) 1.92410.031 (11) 2.10810 .063 (3)6A10 1.33510.077 (39) 1.93910.142 (11) 2.21110 .061 (0)15E1 1.26210.126 (42) 2.07210.008 (5) 2.41910 .015 (0)13H7 1.21410.080 (44) 1.77010.126 (19) 2.15210 .095 ( l )13Hß L20310.042 (45) 1.91510.043 (l2) 2.22010 .082 (0)9B5 . 1.16710.253 (46) 1 .71410.044 (21) 2.11310 .054 (3)15F2 1.09I 10.024 (50) 2.00710.013 (8) 2.47310 .091 (0)1892 0.90310.093 (58) 1.51110.098 (30) 2.08810 .073 (4)1OD12 0.86410.144 (60) 1.45210.125 (33) 2.07410 .150 (5)ID8 0.71210.202 (67) 1.40210.045 (35) 1 .94810 .033 (10)ßH5 0.54910.018 (75) 1.34110.101 (38) 2.04710 .052 (6)IE8 0.46410.021 (79) 1.13010.029 (48) 1 .88610 .015 (l3)
1200
B. G. STILES et ai.
" The mean of the time to death was calculated from five mice used per antibody.Before injecting mice, the toxin and monoclonal antibody mixtures werepreincubated together at room temperature for 30min . Control monoclonalantibody against pseudexin did not cross-react with cobrotoxin . Mice (n = ~injected with toxin alone had time to death readings of 33 minf4min . In vitro per-cent competition values are from Table 2 .
and AchR were incubated together with toxin (Table 3). Both tables show a dose responsewith decreasing antibody concentrations . The addition of 10 pg/ml monoclonal antibody(6.7 pmole/well) before 50 ~g/ml AchR (20 pmole/well) resulted in a toxin/AchR bindinginhibition range of 20-80% .Incubating toxin with an AchR-antibody mixture was not aseffective at similar concentrations, with toxin/AchR binding inhibition ranging from 5 to48% . Co-incubation of five-fold more antibody (33.5 pmole antibody/well) and 20 pmoleAchR/well inhibited the binding of cobrotoxin to AchR by 23-79% . This in vitroinhibition range is very similar to that seen when antibody (6.7 pmole/well) was pre-incubated with toxin before adding AchR (20 pmole/well). None of the monoclonalantibodies (33.5 pmole/well) added 1 hr after AchR (20 pmole/well) dissociated thecobrotoxin-AchR complex.
In vivo neutralization of cobrotoxinThe monoclonal antibodies delayed the time to death in mice, each given 6hg
cobrotoxin (Table 4) . With a 2 : 1 mole ratio of cobrotoxin : monoclonal antibody, therewas a delay in time to death of 2.3-2.7-fold compared with that of control mice givencobrotoxin plus a pseudexin monoclonal antibody that did not cross-react with cobro-toxin. A 2.5 : 1 mole ratio of monoclonal antibody : cobrotoxin yielded a 3.7-23.8-folddelay in time to death compared with the result for conrol mice (Table 4) . Neutralizationof toxin in mice correlated well with the in vitro binding competition values .
Epitope recognitionThe recognition of linear epitopes by monoclonal antibodies was initially tested with
cobrotoxin 8-mer peptides spanning the whole molecule but none of the antibodies boundto the toxin peptides . We obtained further evidence that the monoclonal antibodies bindto conformational epitopes by using an ELISA with reduced and native cobrotoxin(Table 1) . Unlike native cobrotoxin, reduced toxin was not recognized by any of themonoclonal antibodies . Polyclonal antibodies did bind to reduced cobrotoxin (OD,°s =
TABLE 4 .
Antibody
In vivo P1tOTEC1'ION SCUD>QS1lONOCLONAL
Time to death (min)'
WITH COBROTOXINANTiHODY (300 /!g/mOUSt)
Time to death (test)/time to death (control)
(6pg/mouse) Arm
In vitrocompetition (%)
Control 26 t 7 l .0 0IB93 53 t 5 2 .1 202A7 54t 6 2 .1 22lOD12 74t 14 2 .8 479B5 174f 24 6 .7 481E8 248 t 62 9 .5 808H5 618 t 94 23 .8 78

Cobrotoxin Monoclonal Antibodies
12(11
0.306 f0.040), but to a much lesser extent than native toxin (OD,°, > 2.800). These data
suggest that epitopes for the cobrotoxin monoclonal antibodies are conformational, and
not linear.
DISCUSSION
This investigation characterizes 12 highly specific monoclonal antibodies against N. n .
atra cobrotoxin that prevented toxin binding to AchR in vitro, and also delayed the timeto death in mice lethally challenged with cobrotoxin . There have been several monoclonalantibody studies with snake venom postsynaptic neurotoùns, such as a-bungarotoxin andtoxin a (BOULAIN et al ., 1982 ; DANSE et al., 1986; Z~tEINEAU et al., 1986; C~-tuzNC et al.,
1989; KASE et al., 1989; PACtuvElt and RICAI,~rox, 1989). There has been only one otherreport of a monoclonal antibody against cobrotoxin (YANG, 1985), but little informationwas given and no subsequent work has been published.We found a 20-80% inhibition of cobrotoùn binding to AchR in vitro when the
monoclonal antibodies were individually preincubated with toxin before the addition ofAchR . Although a cobrotoxin (6 fig) : antibody mole ratio of 2 : 1 delayed the time todeath in mice (2.3-2 .7 increase in time to death compared with that ofcontrol mice), therewas little variation between antibodies which gave distinctive results in in vitro competi-tion experiments. Mice injected with cobrotoùn (6 pg = 8.6 x 10- '° mole/mouse) andantibody (2.0 x 10-9 mole/mouse), a 1 : 2.5 mole ratio of toxin : antibody, showed a moremarked effect with a 3 .7-23.8-fold delay in the time to death compared with that ofcontrol mice . The in vitro and in vivo (1 : 2.5 mole ratio of toxin : antibody) protectionstudies correlated well . The antibodies that best prevented the binding of toxin to AchRin vitro also afforded the longest time to death in mice. Antibodies with little effect onin vitro binding of toxin to AchR concomitantly yielded shorter times to death in mice.
There was some variation noted between the in vitro and in vivo assays. Antibodies 8H5and 1E8, which had nearly identical in vitro inhibition values (78 and 80%, respectively)for toxin binding to AchR, differed by 2.5-fold in in vivo neutralizing capabilities. Anotherexample of monoclonal antibodies with similar in vitro competition values is 1OD12 and9B5 (47 and48%, respectively) . The in vivo results for 1OD12 and 9B5 differed by 2.4-fold.Antibodies with low in vitro inhibition values, such as 2A7 and 1 B93 (22 and 20%,respectively), provided identical in vivo results. Variations between assays with antibodiesthat share similar in vitro competition values may be attributed to the affinity of anantibody for toxin in solution vs toxin adsorbed to an ELISA plate.
There was little correlation between antibody reactivity with cobrotoxin and inhibitionof toùn binding to AchR in vitro (Tables 1 and 2) . The most remarkable differences werethe relatively low reactivity of antibody lE8 with cobrotoxin (OD,°S = 0.905±0.013) buthigh toùn/AchR binding inhibition in vitro (80%). On the other extreme was antibody15E1 and its higher reactivity with cobrotoxin (OD,°S = 1 .707±0.032) yet lower toxin/AchR binding inhibition in vitro (21%).A previous in vivo neutralization study with N. nigricollis toxin a (BOULAIN et al., 1982)
and a monoclonal antibody showed that a 15.5 : 1 mole ratio of antibody :toxin (3 hgtoùn/mouse) protected all mice (n = 10) for 24 hr. Since the study was not done for alonger time period, it remains uncertain if the mice were protected over an extended timeperiod. The monoclonal antibody recognized a site reportedly not involved in AchRbinding but protection was probably due to steric hindrance of toùn binding to AchR

120 2
B. G. STILES et a/.
and/or the toùn was conformationally changed such that the affinity of the toxin forAchR was decreased. Dissociation of the toxin a : AchR complex was achieved with a 900molar excess of monoclonal antibody vs toxin in a radiolabeled toxin assay (BOULAINet al., 1985). Our in vitro studies with a 13-fold molar excess of any of the cobrotoxinmonoclonal antibodies, compared with plate-adsorbed cobrotoxin able to bind AchR(STtLFS, 1991), did not dissociate the toxin/AchR complex.Another monoclonal antibody/toxin a report by Ttt>a~nu et al. (1986) describes a
different monoclonal antibody which recognized and neutralized various short~hainneurotoxins in vitro, but only delayed mouse lethality, even at a 50 : 1 mole ratio ofantibody : toxin a (MEtvEZ, 1985). The neutralizing epitope was evidently conformationaland incorporated at least the invariant amino acids Lys-27, Trp-29, and Lys-47 ('IytE~nuet al., 1986). We did not try in vivo protection experiments with such a high mole ratio ofantibody :toxin .
In vivo neutralization studies with monoclonal antibody and the long-chain neurotoxin,a-bungarotoxin, also show only a delay in the time to death in mice (DnxsE et al., 1986;KnsE et al., 1989). The inability of monoclonal antibody to prevent lethality may be dueto the higher affinity of toxin for AchR than antibody and/or the processing and excretionof antibody may be faster than toxin (DnNSE et al., 1986). DnxsE et al . (1986) found thatmole ratios of 10 : 1 (antibody : a-bungarotoxin), using 10 pg toxin/mouse with each ofthe 26 monoclonal antibodies, increased the time to death 6-100-fold compared with theresults for control mice given toxin alone. Another study with mole ratios of 1 : 1 and 5 : 1of monoclonal antibody : a-bungarotoxin (10 hg toxin/mouse) increased the time to death4 and 13 .5-fold compared with that of control mice given toxin alone, respectively (KnsEet al., 1989). KnsE et al. (1989) found that the same monoclonal antibody againsta-bungarotoxin also recognized other long-chain neurotoxins. Unlike studies thatinvolved the short-chain neurotoxin, toxin a, linear neutralizing epitopes on a-bungaro-toxin (amino acids 34-41 and 34-38) were recognized by two different monoclonalantibodies (CxunxG et al., 1989 ; Kris et al., 1989).Our investigation suggests that the monoclonal antibodies against cobrotoxin recog-
nized conformational epitopes as none of the monoclonal antibodies reacted with reducedcobrotoxin or synthesized peptides spanning the whole toxin molecule . Since polyclonalantibody bound to reduced cobrotoxin, albeit much less effectively than native toùn, therewas some antibody recognition of linear sequences or all the toxin molecules were notthoroughly reduced .
Besides the recognition of conformational epitopes by monoclonal antibodies againstN. nigricollis toxin a, polyclonal antibodies against a cardiotoxin from N. nivea venomalso bind exclusively to conformational epitopes (05THOFF, 1989). This is of interestbecause some cardiotoxins and short-chain postsynaptic neurotoxins are strikingly similarin isoe~ectric point, amino acid length, conformation, and number of disulfide bridges(VissEx and Louw, 1978). As there was little reactivity of cobrotoxin monoclonalantibodies with other snake toxins, including N. n. atra cardiotoxin, the epitopes must be~~~.iique to cobrotoxin . Lack of reactivity by the monoclonal antibodies with the testedtoxins further suggests that linear epitopes were not recognized because many snakevenom postsynaptic neurotoxins are very homologous (Dur~-rox and Hrosx, 1983).Experiments are planned to determine the epitopes bound by the cobrotoxin monoclonalantibodies .
All monoclonal antibody studies to date with snake venom postsynaptic neurotoxinsshow that a large molar excess of antibody is required for an in vivo protective effect,

Cobrotoxin Monoclonal Antibodies
1203
although transient in many cases. Our study also demonstrates a transient, yet notabsolute, protective effect with monoclonal antibodies against cobrotoxin . The non-radioactive receptor assay provided some clues on the effectiveness of different antibodiesin preventing the binding of toxin to AchR in vivo. Furthermore, development of a trulyefficacious immunotherapy/vaccine against various short- and long-chain postsynapticneurotoxins found in snake venoms will most likely involve more epitope mapping studieswith monoclonal antibodies .
REFERENCES
AaE, T. and TAMIYA,N. (1979) Immunological studies on erabutoxin b, a sea snake toxin. Attempts to locate theamino acid residues determining antigenicity . Toxicon 17, 571-582.
BoQuET, P., PoILLEUx, G., DUMAREY, C., IZARD, Y. and RoNSSPxAY, A.-M. (1973) An attempt to classify thetoxic proteins of Elapidae and Hydrophidae venoms . Toxicon 11, 333-340.
BOULAIN, J.-C., MENEZ, A., COUDERC, J., FAURE, G., LIACOeouLOS, P. and FROMAGEOT, P. (1982) Neutralizingmonoclonal antibody specific for Naja nigricollis toxin a: Preparation, characterization, and localization of theantigenic binding site. Biochemistry 21, 2910-2915.
BOULAIN, l.-C., FROMAGEOT, P. and MENEZ, A. (1985) Further evidence showing that neurotoxin-acetykholinereceptor dissociation is accelerated by monoclonal neurotoxin-specific immunoglobulin . Molec. Immun. 22,553-556.
C~IANG, C. C. and YANG, C. C. (1969) Immunochemical studies on cobrotoxin. J. Immun. 102, 1437-1444.CwANG, C. C. and YANG, C. C. (1973) Immunochemical studies on the tryptophan-modified cobrotoxin .Biochim. biophys. Acta 295, 59504.
CSiUANG, L. Y., LIN, S. R., CHANG, S. F. and GANG, C. C. (1989) Preparation and characterization ofmonoclonal antibody specific for a-bungarotoxin and localization of the epitope. Toxicon 27, 211-219.
DArtsE, J. M., ToussAlrrr, J. L. and KEMPF, J. (1986) Neutralization of a-bungarotoxin by monoclonalantibodies . Toxicon 24, 141-151.
DuFroN, M. J. and HIRER, R. C. (1983) Conformational properties of the neurotoxins and cytotoxins isolatedfrom elapid snake venoms. CRC Crit . Rev. Biochem. 14, 113-171 .
EARLEY, E. M. and OSfF.RLING, M. C. (1985) Fusion of mouse-mouse cells to produce hybridomas secretingmonoclonal antibody . J. Tiss . Cult. Meth . 9, 141-146.
GEYSEx, H. M., MELOEN, R. H. and BARTELIxG, S. J. (1984) Use ofpeptide synthesis to probe viral antigens forepitopes to a resolution of a single amino acid . Proc . natn. Acad. Sci. U.S .A . 81, 3998-4002.
JUILLERAT, M. A., SCHWENDIMANN, B., HAYERT, J., FULPIUS, B. W. and BARGEfZI, J. P. (1982) Specific bindingto isolated acetykholine receptor ofa synthetic peptide duplicating the sequence of the presumed active centerof a lethal toxin from snake venom. J. biol. Chem . 257, 2901-2907.
KASE, R., KITAGAWA, H., HAYASHI, K., TANGUE, K. and INAGAKI, F. (1989) Neutralizing monoclonal antibodyspecific for a-bungarotoxin : Preparation and characterization ofthe antibody, and localization of the antigenicregion of a-bungarotoxin . FEBS Lett . 254, 106-110.
Low, B. W. and CoRF1Ein, P. W. (1986) Erabutoxin b: Structure/function relationships following initial proteinrefinement at 0.140 -nm resolution. Eur. J. Biochem. 161, 579-587.
McDANIEL, C. S., MANSHOURI, T. and ATAS31, M. Z. (1987) A novel peptide mimicking the interaction of aneurotoxins with acetykholine receptor . J. Prot. Chem . 6, 455-461 .
MENEZ, A. (1985) Molecular immunology of snake toxins . Pharmac. Thcr. 30, 91-113 .MENEZ, A., BOULAIN, J. C. and FROMAGEOT, P. (1979) Attempts to define the antigenic structure of Naja
nigricollts toxin a. Toxicon 17, 123.Music-JERICEVIC, B. and ATAS31, M. Z. (1987) Profile of the a-bungarotoxin binding regions on the extracellular
part of the a chain of Torpedo calijornica acetykholine receptor . Bicehem. J. 248, 847-852.Music-JEwcevlc, B., MANSxouRl, T., Yoxtx, T. and ATA39I, M. Z. (1988) The regions of a neurotoxin binding
on the extracellular part of the a subunit of human acetykholine receptor. J. Prot. Chem. 7, 173-177.OSTHOFF, G. (1989) Far-u.v. CD-spectroscopy and immunological properties of synthetic sequential peptidesderived from cardiotoxin V"1 of Naja nivea venom: An amphipathic a-helix formed by sequence IS-25 of aß-protein. Int. J. Biochem. 21, 1365-1368.
PACHNER, A. R. and RICALTON, N. (1989) In vitro neutralization by monoclonal antibodies of a-bungarotoxinbinding to acetykholine receptor. Toxicon 27, 1263-1268.
RENER, J. C., BROWN, B. B. and NARDONE, R. M. (1985) Cloning hybridoma cells by limiting dilution . J. Tics.Cult . Meth . 9, 175-177.
RUAN, K:H., $PURLING, J., Quloclto, F. A. and ATA33I, M. Z. (1990) Acetylcholine receptor-a-bungarotoxininteractions : Determination of the region-to-rogion contacts by peptide-peptide interactions and molecularmodeling of the receptor cavity. Proc. natn . Acad. Sci. U.S.A . 87, 6156-6160.

1204
B. G. STILF.S ct d.
Sxutatwx, M., Wtt.n>~, C. D. and Ko~, G. (1978) A better cell line for making hybridonas secreting spaificantibodies. NoteQe 276, 269-270.
STIIL'4, B. G. (1991) A non-radioactive receptor assay for snake venom postsynaptic neurotoxins. Toxicon 29,503-510.
Tte~au,O., Houunv, J.-C., Couneac, J., FROSUC®or, P. andMENBZ,A. (1986) Amonoclonal antibody whichreoogetized the functional site ofsnake neurotoxina and which neutralizes all short-chain variants. FEBS Lctt.208, 23lr240.
Vts~x, L. and Louw, A. I. (1978) The conformation of cardiotoxins and neurotoxins from snake venoms.Biochim . biophys . Acta 533, 80-89.
WALICINSHAW, M. D., Seeeveea, W. and MAELIC[CE, A. (1980) Three-dimensional structure of the "long"neurotoxin from cobra venom. Proc . natn. Acad. Sci . U.S .A . 77, 2400-2404.
Wept, M. and C~texaeux, J:P. (1974) Binding of Naja nigricollù (~H]a-toxin to membrane fragments fromElectrophorus and Torpedo electric organs . I . Binding of the tritiated a-neurotoxin in the absence of effector.Molec. Pharmac. 10, 1-14.
YANG, C. C. (1965) Crystallvation and properties of cobrotoxin from Formosan cobra venom. J. bio(. Chore .210, 1616-1618.
Yt tvc, C. C. (1974) Chemistry and evolution of toxins in snake venoms . Toxicon 12, 1~3.YANG, C. C. (1985) Immunochemical studies on cobrotoxin . Toxicon 23, 629.Yf,NG, C. C., Yexc, H. J. and Hu~rtc, J. S. (1969) The amino acid sequence of cobrotoxin . Biochim . biophys .Acts 188, 65-77.
YANG, C. C., Y~rtc, H. J. and G~tu, R. H. (1970) The position of disulûde bonds in cobrotoxin . Biochim .biophys. Acts 214, 355-363.
Yexc, C. C., Lrx, M. F. and C~rc, C. C. (1977) Purification of anti-cobrotoxin antibody by amitychromatography. Toxicon 15, 51-62.
Y~rrc, C. C., CFUtve, C. C., Lnv, M. F., LEA, H. Y. and Cttunxc, L. Y. (1978) Purification and properties ofnon-precipitating antibodies to cobrotoxin . Int . J. Peptide Protein Res. 12, 303-310.




















