
Biochemical and molecular characterization of purified chicken pancreatic phospholipase A 2
14
Biochemical and molecular characterization of galectins from zebrafish (Danio rerio): notochord-specific expression of a prototype galectin during early embryogenesis Hafiz Ahmed, Shao-J. Du, Nuala O’Leary, and Gerardo R. Vasta 1 Center of Marine Biotechnology, University of Maryland Biotechnology Institute, 701 East Pratt Street, Baltimore, MD 21202 Received on July 21, 2003; revised on September 17, 2003; accepted on October 7, 2003 Galectins are a family of b-galactoside-binding lectins that on synthesis are either translocated into the nucleus or released to the extracellular space. Their developmentally regulated expression, extracellular location, and affinity for extracel- lular components (such as laminin and fibronectin) suggest a role in embryonic development, but so far this has not been unequivocally established. Zebrafish constitute an ideal model for developmental studies because of their external fertiliza- tion, transparent embryos, rapid growth, and availability of a large collection of mutants. As a first step in addressing the biological roles in zebrafish embryogenesis, we identified and characterized members of the three galectin types: three proto- galectins (Drgal1-L1, Drgal1-L2, Drgal1-L3), one chimera galectin (Drgal3), and one tandem-repeat galectin (Drgal9- L1). Like mammalian prototype galectin-1, Drgal1-L2 pre- ferentially binds to N-acetyllactosamine. Genomic structure of Drgal1-L2 revealed four exons, with the exon–intron boundaries conserved with the mammalian galectin-1. Interestingly, this gene also encodes an alternatively spliced form of Drgal1-L2 that lacks eight amino acids near the carbohydrate-binding domain. Zebrafish galectins exhibited distinct patterns of temporal expression during embryo devel- opment. Drgal1-L2 is expressed postbud stage, and its expression is strikingly specific to the notochord. In contrast, Drgal1-L1 is expressed maternally in the oocytes. Drgal1-L3, Drgal3, and Drgal9-L1 are expressed both maternally and zygotically, ubiquitously in the adult tissues. The distinct temporal and spatial patterns of expression of members of the zebrafish galectin repertoire suggest that each may play distinct biological roles during early embryogenesis. Key words: Danio rerio/developmental expression/ notochord/specificity/zebrafish galectins Introduction There is growing evidence that complex carbohydrate struc- tures encode information that modulate interactions between cells or between cells and the extracellular matrix by specifically binding to cell surface–associated or soluble carbohydrate-binding receptors (Bakkers et al., 1997; Hathaway and Shur, 1997; Laine, 1997). The expression patterns of such carbohydrate-binding receptors are of particular interest in the context of development because the structures of potential ligand molecules, such as glyco- proteins, glycolipids, and other glycans, are subject to change by glycosidases and glycosyltransferases (Shur, 1993; Wassarman, 1995). The rapidity and complexity of the changes in exposed carbohydrate residues during early development suggest that they may play important roles during embryogenesis. Galectins, a family of b-galactoside-binding development- ally regulated proteins formerly known as S-type lectins, have been proposed to participate in a variety of biologi- cal functions, including cell–cell and cell–extracellular matrix interactions that mediate developmental processes (Barondes et al., 1994; Hirabayashi, 1997). However, the detailed mechanisms of their biological role(s) remain unclear. Unlike other lectin families, such as C-type lectins, galectins are a relatively homogeneous, evolutionarily con- served and ubiquitous group, with representatives identified in most animal taxa examined so far, including the parazoa and both protostome and deuterostome lineages (Cooper, 2002; Hirabayashi, 1997; Vasta et al., 1999). Furthermore, from the biochemical, structural, and genetic standpoints, galectins constitute one of the best-characterized lectin families (Ahmed et al., 1996a; Cho and Cummings, 1995; Liao et al., 1994; Ohyama and Kasai, 1988; Schwarz et al., 1998; Sparrow et al., 1987). Based on structural features, galectins have been classi- fied in three types: proto, chimera, and tandem-repeat (Hirabayashi and Kasai, 1993). Prototype galectins contain one carbohydrate-recognition domain (CRD) per subunit and are usually homodimers of noncovalently linked subunits. In contrast, chimera-type galectins are monomeric with a C-terminal CRD similar to the proto type, joined to an N-terminal peptide of yet unknown functional proper- ties. Tandem-repeat galectins, in which two CRDs are joined by a linker peptide, are also monomeric. Proto- and tandem-repeat types make up several distinct galectin subtypes. Galectin subtypes have been numbered following the order of their discovery (Barondes et al., 1994), and so far, 14 have been described in mammals (Dunphy et al., 2002). Substantial efforts have been invested toward the elucida- tion of the role(s) of galectins in embryogenesis and devel- opment using mammalian models (Colnot et al., 1997), but because of the diversity of the galectin repertoire and the technical difficulties in working ex utero with mammalian embryos, results have been inconclusive (Colnot et al., 1998). Lower vertebrates and invertebrates appear to have 1 To whom correspondence should be addressed; e-mail: [email protected] Glycobiology vol. 14 no. 3 # Oxford University Press 2004; all rights reserved. 219 Glycobiology vol. 14 no. 3 pp. 219–232, 2004 DOI: 10.1093/glycob/cwh032 Advance Access publication December 23, 2003 by guest on June 3, 2013 http://glycob.oxfordjournals.org/ Downloaded from
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Biochemical and molecular characterization of purified chicken pancreatic phospholipase A 2

Biochemical and molecular characterization of galectins from zebrafish (Danio rerio):notochord-specific expression of a prototype galectin during early embryogenesis
Hafiz Ahmed, Shao-J. Du, Nuala O'Leary, and GerardoR. Vasta1
Center of Marine Biotechnology, University of Maryland BiotechnologyInstitute, 701 East Pratt Street, Baltimore, MD 21202
Received on July 21, 2003; revised on September 17, 2003; accepted onOctober 7, 2003
Galectins are a family of b-galactoside-binding lectins that onsynthesis are either translocated into the nucleus or releasedto the extracellular space. Their developmentally regulatedexpression, extracellular location, and affinity for extracel-lular components (such as laminin and fibronectin) suggest arole in embryonic development, but so far this has not beenunequivocally established. Zebrafish constitute an ideal modelfor developmental studies because of their external fertiliza-tion, transparent embryos, rapid growth, and availability of alarge collection of mutants. As a first step in addressing thebiological roles in zebrafish embryogenesis, we identified andcharacterized members of the three galectin types: three proto-galectins (Drgal1-L1, Drgal1-L2, Drgal1-L3), one chimeragalectin (Drgal3), and one tandem-repeat galectin (Drgal9-L1). Like mammalian prototype galectin-1, Drgal1-L2 pre-ferentially binds to N-acetyllactosamine. Genomic structureof Drgal1-L2 revealed four exons, with the exon±intronboundaries conserved with the mammalian galectin-1.Interestingly, this gene also encodes an alternatively splicedform of Drgal1-L2 that lacks eight amino acids near thecarbohydrate-binding domain. Zebrafish galectins exhibiteddistinct patterns of temporal expression during embryo devel-opment. Drgal1-L2 is expressed postbud stage, and itsexpression is strikingly specific to the notochord. In contrast,Drgal1-L1 is expressed maternally in the oocytes. Drgal1-L3,Drgal3, and Drgal9-L1 are expressed both maternally andzygotically, ubiquitously in the adult tissues. The distincttemporal and spatial patterns of expression of members ofthe zebrafish galectin repertoire suggest that each may playdistinct biological roles during early embryogenesis.
Key words: Danio rerio/developmental expression/notochord/specificity/zebrafish galectins
Introduction
There is growing evidence that complex carbohydrate struc-tures encode information that modulate interactionsbetween cells or between cells and the extracellular matrix
by specifically binding to cell surface±associated or solublecarbohydrate-binding receptors (Bakkers et al., 1997;Hathaway and Shur, 1997; Laine, 1997). The expressionpatterns of such carbohydrate-binding receptors are ofparticular interest in the context of development becausethe structures of potential ligand molecules, such as glyco-proteins, glycolipids, and other glycans, are subject tochange by glycosidases and glycosyltransferases (Shur,1993; Wassarman, 1995). The rapidity and complexity ofthe changes in exposed carbohydrate residues during earlydevelopment suggest that they may play important rolesduring embryogenesis.
Galectins, a family of b-galactoside-binding development-ally regulated proteins formerly known as S-type lectins,have been proposed to participate in a variety of biologi-cal functions, including cell±cell and cell±extracellularmatrix interactions that mediate developmental processes(Barondes et al., 1994; Hirabayashi, 1997). However, thedetailed mechanisms of their biological role(s) remainunclear. Unlike other lectin families, such as C-type lectins,galectins are a relatively homogeneous, evolutionarily con-served and ubiquitous group, with representatives identifiedin most animal taxa examined so far, including the parazoaand both protostome and deuterostome lineages (Cooper,2002; Hirabayashi, 1997; Vasta et al., 1999). Furthermore,from the biochemical, structural, and genetic standpoints,galectins constitute one of the best-characterized lectinfamilies (Ahmed et al., 1996a; Cho and Cummings, 1995;Liao et al., 1994; Ohyama and Kasai, 1988; Schwarz et al.,1998; Sparrow et al., 1987).
Based on structural features, galectins have been classi-fied in three types: proto, chimera, and tandem-repeat(Hirabayashi and Kasai, 1993). Prototype galectins containone carbohydrate-recognition domain (CRD) per subunitand are usually homodimers of noncovalently linkedsubunits. In contrast, chimera-type galectins are monomericwith a C-terminal CRD similar to the proto type, joined toan N-terminal peptide of yet unknown functional proper-ties. Tandem-repeat galectins, in which two CRDs arejoined by a linker peptide, are also monomeric. Proto- andtandem-repeat types make up several distinct galectinsubtypes. Galectin subtypes have been numbered followingthe order of their discovery (Barondes et al., 1994), and so far,14 have been described in mammals (Dunphy et al., 2002).
Substantial efforts have been invested toward the elucida-tion of the role(s) of galectins in embryogenesis and devel-opment using mammalian models (Colnot et al., 1997), butbecause of the diversity of the galectin repertoire and thetechnical difficulties in working ex utero with mammalianembryos, results have been inconclusive (Colnot et al.,1998). Lower vertebrates and invertebrates appear to have
1To whom correspondence should be addressed; e-mail:[email protected]
Glycobiology vol. 14 no. 3 # Oxford University Press 2004; all rights reserved. 219
Glycobiology vol. 14 no. 3 pp. 219±232, 2004DOI: 10.1093/glycob/cwh032Advance Access publication December 23, 2003
by guest on June 3, 2013http://glycob.oxfordjournals.org/
Dow
nloaded from

smaller galectin repertoires (Cooper, 2002). Thus speciescurrently used as model organisms for genetic and develop-mental studies, such as the zebrafish (Danio rerio), fruitfly(Drosophila melanogaster), and nematode (Caenorhabditiselegans) may be more suitable alternatives for studies on thefunctional roles of galectins. Galectins have been identifiedand characterized in these species by biochemical andmolecular approaches (Cooper, 2002; Hirabayashi et al.,1997; Pace et al., 2002), as well as current genomeprojects (C. elegans Sequencing Consortium, 1998; BerkleyDrosophila Genome Project, NIH Zebrafish GenomeProject).
Zebrafish should constitute a particularly useful animalmodel for investigating biological roles of galectins in earlyembryo development of vertebrates because this speciesoffers a number of advantages over mammalian systems.First, fertilization is external; the embryos develop rapidlyin vitro and are transparent, making it possible to visualizethe expression of putative genes mediating cell adhesionand/or migration. Second, the early expression of thosegene(s) can be easily manipulated in zebrafish embryos,enabling visualization of the phenotypic consequences oftheir misexpression. Third, a growing collection of interest-ing mutations that affect early embryonic development arebeing generated and mapped, providing an excellentresource for genetic studies of the function and mechanismsof action of galectin(s) genes. Fourth, zebrafish cell lineshave been established that might be very useful for studyinggene expression and protein export at a cellular level.Finally, because zebrafish are evolutionarily closer to mam-mals than the above-mentioned invertebrate models, struc-tural and functional information obtained in this organismmay be more applicable to the interpretation of similarobservations in mammalian systems.
In this study we report for the first time the identificationand characterization of members of the three galectin typesin zebrafish and their differential expression during earlydevelopment.
Results
Purification and characterization of galectins fromzebrafish
Lactose-binding lectins were purified from whole adultzebrafish extracts by affinity chromatography on lactosyl-Sepharose as described under Materials and methods. Onsodium dodecyl sulfate±polyacrylamide gel electrophoresis(SDS±PAGE), the lactose eluate resolved in four bandscorresponding to 14.5, 15.5, 22, and 24 kDa, of which the14.5 kDa (designated as Drgal1-L2) was separated byanion-exchange chromatography on DEAE-Sepharose,based on its acidic properties (Figure 1Ab). On westernblot, only the 14.5-kDa component of the lactose eluatecross-reacted with an anti-[striped bass proto galectin] anti-body (Figure 1B). Drgal1-L2 behaved as a dimer in gelpermeation chromatography (Figure 1C). The yield ofthe purified Drgal1-L2 was approximately 2 mg/g adultfish tissue and had a specific activity of 1±2� 104 mg/mlwith protease-treated rabbit erythrocytes. To determinethe amino acid sequence of Drgal1-L2, the protein was
trypsin-digested, and selected peptides sequenced. Fromthe 38 peaks separated by high-performance liquidchromatography (HPLC) from the tryptic digest, 9 werepredominant (data not shown). Four of these fractionswere further purified, and the peptides were sequenced(a total of 56 amino acids) by using a combination oftandem mass spectrometry and Edman degradation. Theselected peptides yielded the following amino acid sequences:peptide 16 (mass 1168.4): YMHFEGEVR; peptide 23 (mass1238.4): DNNFPFIQDK; peptide 32 (mass not confirmed):VGQTLTITGVPKP(D); and peptide 38 (mass 2677.8):ITFTNEEFLVTLPDGSEIHFPNR. Based on these pep-tide sequences, molecular probes were synthesized forcloning the Drgal1-L2 cDNA.
Carbohydrate specificity of Drgal1-L2
The carbohydrate-binding specificity of Drgal1-L2 wasdetermined by analyzing the binding of the Drgal1-L2-HRP conjugate to asialofetuin (ASF) in the presence ofseveral saccharides in a solid phase assay under the optimalconditions established (Ahmed et al., 1996a, 2002). Foreach test saccharide, a complete inhibition curve was estab-lished, and the molar concentrations that inhibited thebinding of the lectin conjugate to ASF by 50% (I50) werecalculated and normalized with respect to lactose, whichwas included in each plate as a standard (Table I). Thebinding of the Drgal1-L2 to Galb1,4GlcNAc and thiodi-galactoside were about 8±12-fold higher than that tolactose. The binding of Drgal1-L2 to Galb1,3GalNAc wasnegligible. The overall binding-inhibition pattern ofDrgal1-L2 suggests a specificity very similar to thatobserved in mammalian 14 kDa galectins exhibiting
A B C
67 29 13.7kDa
a b
kDa24.022.0
15.514.5
Fig. 1. (A) Purification of galectin-1 like protein from zebrafish(D. rerio): the extract from adult zebrafish was absorbed on lactosyl-Sepharose, and the lactose eluate was separated on DEAE-Sepharosecolumn. Samples of (a) flow-through and (b), bound fractions weresubjected to SDS±PAGE (15%) followed by silver staining.(B) Serological cross-reactivity of D. rerio galectin with the 14.5-kDagalectin from striped bass (Morone saxatalis): the lactose eluate waselectrophoresed, transferred onto a PVDF membrane, and incubatedwith anti-[14.5-kDa striped bass galectin] antibody followed by goatanti-rabbit IgG-peroxidase conjugate. Color for peroxidase activitywas developed with chloronaphthol according to the manufacturer'sinstructions. (C) Gel permeation chromatography of the purified14.5-kDa galectin from D. rerio. The sample (1 mg) was loaded ontoSuperose 6 column as described in Materials and methods.
H. Ahmed et al.
220
by guest on June 3, 2013http://glycob.oxfordjournals.org/
Dow
nloaded from

conserved (Type I) CRDs (Ahmed and Vasta, 1994). Bind-ing of Drgal1-L2 to plastic-coated laminin was linear up toapproximately 0.6 mg/well of laminin (Figure 2).
cDNA cloning and sequence analysis of Drgal1-L2
The full-length cDNA for Drgal1-L2 (GenBank accessionnumber AY421704) was obtained by PCR followed by50- and 30-rapid amplification of cDNA ends (RACE)(Figure 3). The cDNA was approximately 850 bp with aconserved polyadenylation signal sequence at nucleotidepositions 816±821. The cDNA encodes a polypeptide of134 amino acids with a calculated molecular weight of15,254 and theoretical pI of 5.25. Comparison of the proteinsequence deduced from the cDNA with the sequences ob-tained for the tryptic peptides revealed full sequence iden-tity. Comparison of the Drgal1-L2 amino acid sequence
with sequences of selected galectins (Figure 4) confirmedthat the D. rerio lectin is a member of the galectin family.When considering the sequence of the CRD and itsmolecular size (subunit or native), it clearly belongs tothe prototype galectins and is more closely related to thegalectin-1 group than to any other galectin family members.When compared to galectins from ectothermic vertebrates,the highest percent identity was observed with electric eel(64%; Paroutaud et al., 1987), followed by conger eel (32%;Muramoto and Kamiya, 1992), and toad Bufo arenarum(32%; Ahmed et al., 1996a). The percent identities withgalectin-1 from homeotherm vertebrates, such as human(Gitt and Barondes, 1986) and bovine (Ahmed et al.,1996b), ranged from 30% to 35%, whereas it was 28%identical to the 16-kDa proto type galectin from C. elegans(Hirabayashi et al., 1996).
Recently, several galectin sequences from the zebrafishgenome project became available in the GenBank database(Cooper, 2002). Analyses of these sequences revealed twoadditional prototype galectin-1 like lectins (Drgal1-L1 andDrgal1-L3), a chimera galectin (Drgal3), and two tandem-repeat galectin9-like lectins (Drgal9-L1 and Drgal9-L2).Drgal1-L1 (calculated molecular weight of 15,556 and pIof 4.93) is 79% identical to Drgal1-L2 (Figure 4). Drgal1-L3(calculated molecular weight of 14,804 and pI of 6.64) isonly 49% identical to Drgal1-L2 and 45% to Drgal1-L1.Because the cDNA sequences of Drgal3 and Drgal9-L1 inthe database are incomplete, we used 50- and 30-RACE todetermine their full-length sequences. The cDNA forDrgal3 (GenBank accession number AY421705) had anopen reading frame (ORF) (45±728) and a potential initia-tion codon (Figure 5). The ORF encoded 228 amino acidswith a predicted molecular weight of 25,017 and calculatedpI of 6.29. It is noteworthy that the N-terminal portion(90 amino acids) of Drgal3 was significantly shorter relativeto the known chimera galectins from higher vertebrates,such as human (114 aa; Robertson et al., 1990) and rat(128 aa; Oda et al., 1993). The full-length cDNA ofDrgal9-L1 (GenBank accession number AY421706) wasalso cloned by 50- and 30-RACE (Figure 6). The cDNAcontained an ORF (53±1012) that encoded 320 amino
Fig. 2. Binding of 14.5 kDa galectin to laminin. Binding of the galectin(10 mg/100 ml/well) to variable amount of laminin (0.02±0.3 mg/well) withand without lactose. After washing the plate, the bound galectin wasdetected with rabbit anti-galectin antibody, followed by goat anti-rabbitIgG peroxidase and development with ABTS substrate.
Table I. Inhibition of Drgal1-L2 binding to asialofetuin by a seriesof saccharidesa
Compound I50 (mM)Relative inhibitoryactivityb
Galb1,4Glc 4.4±6.5c 1.0
Galb1,4Glcb-OPhNH2 (p) 3.2 1.4
Galb1,4GlcNAc 0.68 7.9
Galb1,3GlcNAc 3.2 1.4
Galb1,3GlcNAcb-OPhNO2 (p) 2.0 2.3
Galb1S1bGal 0.44 12.2
Galb1,3Ara 3.1 2.1
Galb1,4Man 2.9 1.5
Galb1,4Fruf 6.5 1.0
Galb1,6GlcNAc 140 0.03
Galb1,3GalNAc 950 0.007
MeO-2Galb1,4Glc 1.5 2.9
Fuca1,2Galb1,4Glc 12.5 0.35
Fuca1,2[GalNAca1,3]Galb1,4Glc 4500 (37%)d 50.01
NeuAca2,3Galb1,4Glc 160 0.027
Glcb1,4Glc 425,000 (16%) 50.0002
GlcNAcb1,4Galb1,4Glc 4400 (38%) 50.011
NeuAca2,6Galb1,4Glc 4200 (5%) 50.023
Fuca1,3[Galb1,4] Glc 4200 (10%) 50.023
Gal 1400 0.004
Gala-OMe 1000 0.006
Gala-OPhNO2 (p) 40 0.15
Galb-OMe 1400 0.004
Galb-OPhNO2 (p) 380 0.016
Gala1,6Glc 1400 0.004
aEach saccharide was tested in duplicate and average value was taken.
bLactose was included in each plate as a standard and I50 for each
saccharide was normalized with respect to lactose.cLac I50 varied from 4.4±6.5 mM in 10 experiments.
d% Inhibition of binding was indicated in parenthesis because I50 was
not achieved at the highest concentration tested.
Molecular characterization and developmental expression of zebrafish galectins
221
by guest on June 3, 2013http://glycob.oxfordjournals.org/
Dow
nloaded from

acids with a predicted molecular weight of 36,097 and cal-culated pI of 8.67. The sequence revealed two CRDs (38%identical): N-terminal (147 aa) and C-terminal (140 aa)connected by a 33-aa-long linker peptide.
Gene organization and linkage analysis of Drgal1-L2
A genomic clone for Drgal1-L2 was isolated by polymerasechain reaction (PCR) from a Genome Walker genomiclibrary. The Drgal1-L2 gene contains four exons (see Figure 3for positions of the introns), with highly conserved exon/intron boundaries as in mammalian galectin-1 (Hirabayashiand Kasai, 1993). Linkage analysis using the LN54 hybridpanel, placed the Drgal1-L2 gene in the linkage group 3, at5.66 centiRay (1 cR is ~148 kb) from the marker fc23e05.
Identification of a Drgal1-L2 isoform (Drgal1-L2d)
During sequencing of Drgal1-L2, we identified a cDNAclone encodes a galectin isoform (Drgal1-L2d) that wasidentical to the Drgal1-L2 except that it lacks eight aminoacids (FAINIGHS) corresponding to part of the exon III(see Figure 3). Examination of the genomic sequence
around the deleted region revealed a cryptic splicingsequence at the deleted site, suggesting that the isoformDrgal1-L2d probably arises from alternate splicing ofDrgal1-L2 gene (Figure 3). To rule out the presence of asecond gene encoding for the Drgal1-L2d isoform, PCRamplification of zebrafish genomic DNA was carried outwith primers 5 and 6, which would amplify both genes. ThePCR product was subcloned, and multiple clones weresequenced. All sequences were identical and represent asingle gene corresponding to Drgal1-L2. No clones corre-sponding to Drgal1-L2d were identified.
Galectin expression in developing and adult zebrafish
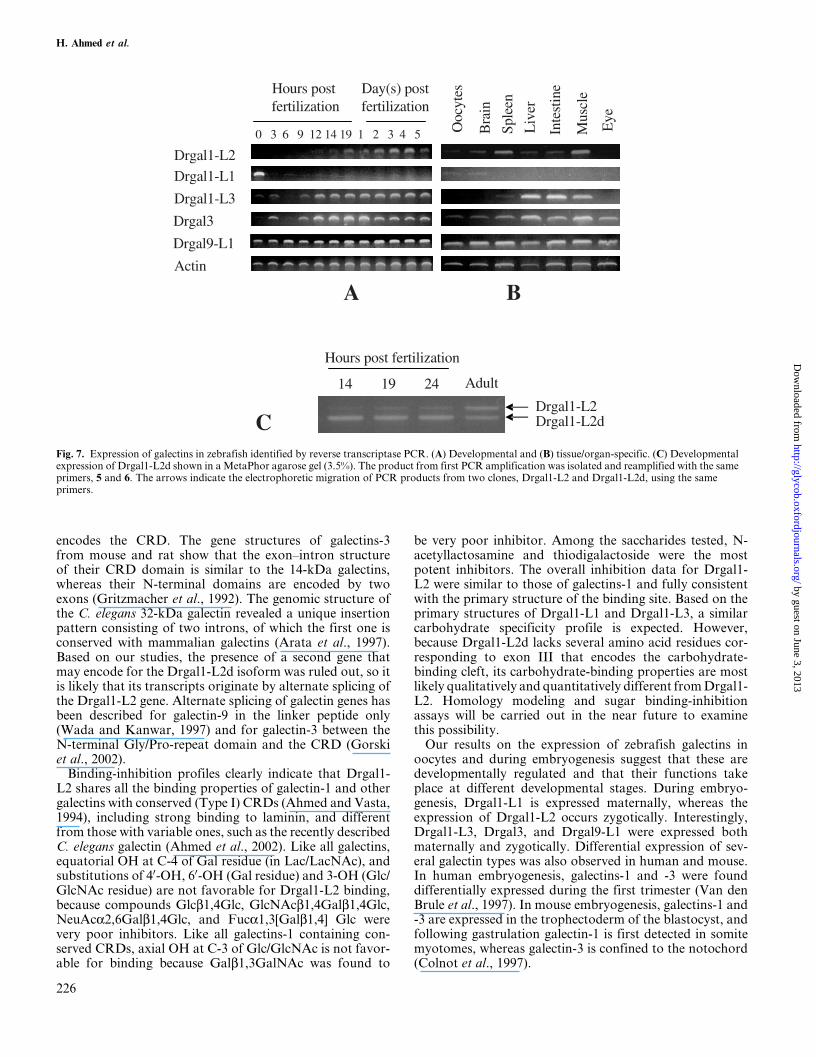
The temporal expression of each galectin was analyzed inzebrafish oocytes and embryos by reverse transcriptionPCR. Drgal1-L1 was expressed in 0 h postfertilization, butdeclined in blastula and post-blastula stage embryos. Thesedata suggest that Drgal1-L1 is maternally expressed. In con-trast, little or no Drgal1-L2 expression could be detected innewly fertilized eggs and embryos at 3±9 h postfertilization,but become evident at 12 h postfertilization, and remained
Fig. 3. Nucleotide and deduced amino acid sequence from cDNA for the Drgal1-L2. Sequences for the peptides are underscored. Sequence for the clone(350 bp) generated from the degenerate primers starts at nucleotide 92 and terminates at 433. The stop codon is marked by an asterisk. The putativepolyadenylation signal sequence in the 30-untranslated region of the transcript is in italics. The position of introns are indicated by vertical lines. Thecryptic site for alternate splicing of Drgal1-L2 gene is shown in the beginning of exon III.
H. Ahmed et al.
222
by guest on June 3, 2013http://glycob.oxfordjournals.org/
Dow
nloaded from

high in later stages (14±19 h postfertilization; 1±5 day(s) post-fertilization (Figure 7A). Interestingly, Drgal1-L3 wasexpressed both maternally and zygotically with a sharp declineat 6 h postfertilization (Figure 7A). The expression of Drgal3was detected at 0 h postfertilization and increased at 3 hpostfertilization. Its expression declined at 6 h postfertilizationbut increased at 9 h postfertilization and remained high at laterstages (Figure 7A). Unlike all galectins, the expression ofDrgal9-L1 was strong in all stages tested (Figure 7). Figure 7Bshows organ expression in adult zebrafish. Drgal1-L2 isexpressed in brain, spleen, liver, intestine, and muscle as ana-lyzed by reverse transcription PCR. For Drgal1-L1, weakexpression was observed only in oocytes and brain. Drgal1-L3 was strongly expressed in liver, intestine, and muscle andweakly in oocytes and spleen. But Drgal3 and Drgal9-L1 wereexpressed in all organs tested (oocytes, brain, spleen, liver,intestine, muscle, and eye). The isoform Drgal1-L2d wasdevelopmentally expressed (Figure 7C).
The spatial expression of Drgal1-L2 was examinedby whole mount in situ hybridization. Results on 16- and24-h postfertilization embryos confirmed that Drgal1-L2is expressed in early embryonic stages, particularly inthe notochord (Figure 8A, C, E, F). The embryos probedwith sense RNA did not show any staining (Figure 8B, D).
The in situ hybridization results were further corroboratedby whole mount antibody staining using anti-Drgal1-L2 anti-bodies on 24-h postfertilization embryos (Figure 8H).The antibody was validated as specific for Drgal1-L2 onwestern blot: only a single band corresponding to 14.5 kDa
(Drgal1-L2) was detected in both the crude zebrafish extractand the lactose eluted galectin mix (Figure 8G). Immunostain-ing results revealed that the Drgal1-L2 protein is localized inthe notochord (Figure 8H). The preimmune serum controlsdid not show comparable staining in notochord (not shown).
Discussion
In this study we characterize the diversity and develop-mental expression of the zebrafish galectin repertoire. Wepurified, biochemically characterized, and cloned Drgal1-L2 and its isoform Drgal1-L2d and cloned the full-lengthcDNAs of the zebrafish chimera and tandem-repeat galec-tins Drgal3 and Drgal9-L1. Furthermore we identified fromthe database two additional closely related yet distinct pro-togalectins Drgal1-L1 and Drgal1-L3. All zebrafish galec-tins characterized in this study showed remarkablestructural similarities with mammalian galectins, and thisenabled their unambiguous classification within the threewell-established galectin groups. Galectin repertoires ofprotostome invertebrates and lower vertebrates appear tobe limited to one or two galectin types (Cooper, 2002). Forexample, C. elegans and Drosophila have proto- andtandem-repeat type galectins (Cooper, 2002; Hirabayashiet al., 1997; Pace et al., 2002). In this aspect, it is noteworthythat at the evolutionary level of teleosts all three galectingroups are already represented. Nevertheless, based on ourstudies and the information currently available from the
Fig. 4. Amino acid sequence comparison of the Drgal1-L2 with other galectins from vertebrates and invertebrates. Amino acid sequence of electric eel(EE), bovine spleen (Bvsp), B. arenarum (Bufo), conger eel (CE), and C. elegans 16 kDa (N16) galectins are aligned by Pileup program (GCG) sequenceanalysis package. The amino acids (shown in bold) are known to interact with N-acetyllactosamine as determined from 3D structure of the bovinegalectin-1 (Liao et al., 1994).
Molecular characterization and developmental expression of zebrafish galectins
223
by guest on June 3, 2013http://glycob.oxfordjournals.org/
Dow
nloaded from

zebrafish genome project, it seems quite clear that the zebra-fish galectin repertoire is smaller than that of mammals.Therefore, this species should be a more suitable modelorganism for the study of biological roles of galectins thanthe mammalian model systems, in which 14 distinct galec-tins have been already identified (Dunphy et al., 2002) andadditional galectin members are evident from GenBankdatabases (Cooper, 2002).
Like mammalian protogalectins, Drgal1-L2 is a homo-dimer with a single CRD per subunit. The primary structureof the zebrafish Drgal9-L1 revealed the typical organizationof the mammalian tandem-repeat galectins, where the twoCRDs are connected by a linker peptide. Similarly, despite ashorter N-terminal peptide region, the zebrafish Drgal3exhibited the typical organization of the chimera typemammalian galectins. Alignment of amino acid sequencesfor Drgal1-L2, Drgal1-L1, and Drgal1-L3 with the
sequences of selected galectins; the presence of conservedcritical residues in their CRDs; and their subunit and nativemolecular sizes clearly indicate that they belong to the pro-totype galectins and are more closely related to galectins-1than to any other subgroup. All three prototype galectinsare acidic and showed galectin-1-like sequences that containconserved (Type I) CRDs (Ahmed and Vasta, 1994). To ourknowledge, this is the first observation of the fact that threeclosely related yet distinct prototype galectins are present ina given organism (Cooper, 2002). Two prototype galectins(galectin-1-like) have been described in chicken, but theyare substantially divergent: they are only 39% identical, andone is acidic, whereas the other is almost neutral. When theamino acid sequence of Drgal1-L2 sequence was comparedwith galectins from vertebrates and invertebrates, the high-est identity was observed with electric eel (64%), whereaslower percent identities were observed with mammalian
Fig. 5. Nucleotide and deduced amino acid sequence from cDNA for the Drgal3. The stop codon is marked by an asterisk. The putative polyadenylationsignal sequence in the 30-untranslated region of the transcript is in italics. The N-terminal peptide of Drgal3 is underscored. The critical amino acidresidues that are known to interact with N-acetyllactosamine (Liao et al., 1994) are shown in bold.
H. Ahmed et al.
224
by guest on June 3, 2013http://glycob.oxfordjournals.org/
Dow
nloaded from

galectins-1 from human, bovine, mouse, and amphibians,and an invertebrate homolog (C. elegans) ranged from 28%to 35%. The observed cross-reactivity with an anti-(proto)-galectin from M. saxatilis, another teleost fish, furtherconfirmed the structural similarities observed.
Despite some difference in intron sizes, the gene encodingfor Drgal1-L2 showed an organization similar to the mam-malian galectin-1 genes. The gene organization of galectins
from human, mouse, and chicken is remarkably similar.Two galectins-1 from human and mouse (Chiariotti et al.,1991; Gitt and Barondes, 1991), galectin-2 (Gitt et al.,1992), and chicken 14-kDa galectin (Ohyama and Kasai,1988) have identical numbers of exons, and their sizes aresimilar. Each gene (about 3.2 kb) contains four exons (fromthe 50 end to the 30 end: exon I, 6±9 bp; II, 80±83; III,160±172 bp; IV, 144±150 bp), of which the largest (exon III)
Fig. 6. Nucleotide and deduced amino acid sequence from cDNA for the Drgal9-L1. The two carbohydrate-binding domains are connected by a linkpeptide (underscored). The stop codon is marked by an asterisk. The putative polyadenylation signal sequence in the 30-untranslated region for thetranscript is in italics. The critical amino acid residues that are known to interact with N-acetyllactosamine (Liao et al., 1994) are shown in bold.
Molecular characterization and developmental expression of zebrafish galectins
225
by guest on June 3, 2013http://glycob.oxfordjournals.org/
Dow
nloaded from
encodes the CRD. The gene structures of galectins-3from mouse and rat show that the exon±intron structureof their CRD domain is similar to the 14-kDa galectins,whereas their N-terminal domains are encoded by twoexons (Gritzmacher et al., 1992). The genomic structure ofthe C. elegans 32-kDa galectin revealed a unique insertionpattern consisting of two introns, of which the first one isconserved with mammalian galectins (Arata et al., 1997).Based on our studies, the presence of a second gene thatmay encode for the Drgal1-L2d isoform was ruled out, so itis likely that its transcripts originate by alternate splicing ofthe Drgal1-L2 gene. Alternate splicing of galectin genes hasbeen described for galectin-9 in the linker peptide only(Wada and Kanwar, 1997) and for galectin-3 between theN-terminal Gly/Pro-repeat domain and the CRD (Gorskiet al., 2002).
Binding-inhibition profiles clearly indicate that Drgal1-L2 shares all the binding properties of galectin-1 and othergalectins with conserved (Type I) CRDs (Ahmed and Vasta,1994), including strong binding to laminin, and differentfrom those with variable ones, such as the recently describedC. elegans galectin (Ahmed et al., 2002). Like all galectins,equatorial OH at C-4 of Gal residue (in Lac/LacNAc), andsubstitutions of 40-OH, 60-OH (Gal residue) and 3-OH (Glc/GlcNAc residue) are not favorable for Drgal1-L2 binding,because compounds Glcb1,4Glc, GlcNAcb1,4Galb1,4Glc,NeuAca2,6Galb1,4Glc, and Fuca1,3[Galb1,4] Glc werevery poor inhibitors. Like all galectins-1 containing con-served CRDs, axial OH at C-3 of Glc/GlcNAc is not favor-able for binding because Galb1,3GalNAc was found to
be very poor inhibitor. Among the saccharides tested, N-acetyllactosamine and thiodigalactoside were the mostpotent inhibitors. The overall inhibition data for Drgal1-L2 were similar to those of galectins-1 and fully consistentwith the primary structure of the binding site. Based on theprimary structures of Drgal1-L1 and Drgal1-L3, a similarcarbohydrate specificity profile is expected. However,because Drgal1-L2d lacks several amino acid residues cor-responding to exon III that encodes the carbohydrate-binding cleft, its carbohydrate-binding properties are mostlikely qualitatively and quantitatively different from Drgal1-L2. Homology modeling and sugar binding-inhibitionassays will be carried out in the near future to examinethis possibility.
Our results on the expression of zebrafish galectins inoocytes and during embryogenesis suggest that these aredevelopmentally regulated and that their functions takeplace at different developmental stages. During embryo-genesis, Drgal1-L1 is expressed maternally, whereas theexpression of Drgal1-L2 occurs zygotically. Interestingly,Drgal1-L3, Drgal3, and Drgal9-L1 were expressed bothmaternally and zygotically. Differential expression of sev-eral galectin types was also observed in human and mouse.In human embryogenesis, galectins-1 and -3 were founddifferentially expressed during the first trimester (Van denBrule et al., 1997). In mouse embryogenesis, galectins-1 and-3 are expressed in the trophectoderm of the blastocyst, andfollowing gastrulation galectin-1 is first detected in somitemyotomes, whereas galectin-3 is confined to the notochord(Colnot et al., 1997).
Fig. 7. Expression of galectins in zebrafish identified by reverse transcriptase PCR. (A) Developmental and (B) tissue/organ-specific. (C) Developmentalexpression of Drgal1-L2d shown in a MetaPhor agarose gel (3.5%). The product from first PCR amplification was isolated and reamplified with the sameprimers, 5 and 6. The arrows indicate the electrophoretic migration of PCR products from two clones, Drgal1-L2 and Drgal1-L2d, using the sameprimers.
H. Ahmed et al.
226
by guest on June 3, 2013http://glycob.oxfordjournals.org/
Dow
nloaded from

The strong expression of Drgal1-L2 in the notochord mayindicate its role in early embryo development. Several sce-narios are possible. First, Drgal1-L2 may be involved innotochord development by regulating adhesion or migra-tion of notochord cells during embryogenesis. This studydemonstrated that Drgal1-L2 strongly binds to laminin,and the presence and distribution of this glycan in thenotochord has been characterized in detail (Parsons et al.,2002). The presence of both the galectin and its ligand in the
notochord, a critical structure for further patterning of theembryo, suggests a relevant role for Drgal1-L2 in the zebra-fish embryogenesis. It is known that Hedgehog signalsecreted from the notochord plays an important role inslow muscle formation (Blagden et al., 1997; Currie andIngham, 1996; Du et al., 1997). Like Hedghog, Drgal1-L2may also be involved in muscle cell differentiation. Hed-ghog from the floor plate is also involved in slow muscleformation (Coutelle et al., 2001; Du et al., 1997). Drgal1-L2
Fig. 8. Whole mount in situ hybridization and immunostaining. (A±F) In situ hybridization of whole zebrafish embryo showing Drgal1-L2 expression innotochord (A, C, E, F). (A) Lateral view of 24-h postfertilization embryo; (B) 24-h postfertilization embryo with sense probe (ÿ) control; (C) 24-hpostfertilization embryo showing trunk; (D) 24-h postfertilization embryo with sense probe showing trunk; (E) lateral view of 16-h postfertilizationembryo; (F) cross-section of trunk after whole mount in situ hybridization; (G) specificity of anti-Drgal1-L2 antibodies. (I) Crude zebrafish extract and(II) the affinity-purified galectin mix were tested with the antibodies on western blot. (H) Whole mount immunostaining with anti-Drgal1-L2 antibodies.Lateral view of trunk of zebrafish embryo (24-h postfertilization) showing staining in notochord.
Molecular characterization and developmental expression of zebrafish galectins
227
by guest on June 3, 2013http://glycob.oxfordjournals.org/
Dow
nloaded from

may influence notochord formation and consequently theexpression of Hedghog signals, and thus Drgal1-L2 mayindirectly be involved in muscle cell differentiation.
Because the notochord is critical for the neural tubepatterning, Drgal1-L2 may be involved in neural tube for-mation and expression of Wnt and bone morphogeneticproteins (BMPs). It has been demonstrated that Wnt andBMPs expressed in the neural tube are critical for muscledevelopment (Stern et al., 1995; Tajbakhsh et al., 1998).Thus Drgal1-L2 may indirectly be involved in muscle devel-opment. During myogenesis, glycoproteins or proteoglycansare directly involved in transducing Wnt and Hedgehog andepidermal growth factor signaling (Lin and Perrimon, 1999;Perrimon and Bernfield, 2000), and it is possible thatDrgal1-L2 plays a role in modulating the activity of theseglycans. Moreover, during development of skeletal musclea population of myoblasts migrate to their destinationsthrough the extracellular matrix and then differentiate, soit has been suggested that lectins might control the inter-action between myoblasts and the extracellular matrix byselective modulating the interaction of integrin with fibro-nectin and laminin and thus regulates the migration of myo-blasts (Gu et al., 1994). In mammalian models it has beenshown that in skeletal muscle galectin-1 is secreted duringdifferentiation (Goldring et al., 2002; Gu et al., 1994) andaccumulates with laminin in the basement membranesurrounding each myofiber (Cooper and Barondes, 1990).
Materials and methods
Reagents
The protein assay reagent was from BioRad (Hercules,CA). Size exclusion chromatography molecular weightstandards were purchased from Amersham Pharmacia(Piscataway, NJ). The peroxidase substrate diammonium2,20-azinobis (3-ethylbenzothiazoline-6-sulfonate) (ABTS)was from Kirkegaard and Perry Laboratories (Gaithersburg,MD). Sequencing-grade reagents and solvents for proteinsequencing, amino acid analysis, and HPLC were fromApplied Biosystems (Foster City, CA). Sequencing-gradetrypsin was obtained from Roche Molecular Biochemicals(Indianapolis, IN). Horseradish peroxidase (HRP), andcarbohydrates used in binding-inhibition assays were fromSigma (St. Louis, MO). All other reagents were of the high-est grade commercially available.
Purification of D. rerio 14.5-kDa galectin
The galectin was purified following the procedure describedelsewhere (Ahmed et al., 1996a), with some modifications.Briefly, approximately 400 adult zebrafish (D. rerio; 200 gtotal weight) were homogenized with a PRO 400 tissuehomogenizer using 59 mm� 150 mm generator (PROScientific, Monroe, CT) in cold (4�C) phosphate bufferedsaline (PBS)/0.01 M 2-mercaptoethanol (ME) containing0.1 M lactose and 0.1 mM phenylmethylsulfonyl fluoride,at 2 ml buffer/g wet tissue. The homogenate was centrifugedat 27,000 � g for 1 h at 4�C, and the clear supernatantwas dialyzed against PBS/ME and absorbed on a lactosyl-Sepharose column preequilibrated with PBS/ME/0.002 MEDTA/0.5 M NaCl. The column was thoroughly washed
with equilibrating buffer followed by five bed volumes ofPBS (1:10)/ME, and the bound protein was eluted with0.1M lactose in PBS (1:10)/ME. To isolate the 14.5-kDagalectin (D. rerio protogalectin; Drgal1-L2) from othergalectin species, the eluted fractions containing proteinwere pooled and absorbed on a DEAE-Sepharose column(1 ml bed volume) preequilibrated with PBS (1:10)/ME.After washing the column with PBS (1:10)/ME, the boundprotein was eluted with PBS (1:10)/ME/0.5 M NaCl.
Analytical procedures
Agglutination tests with protease-treated rabbit erythro-cytes were carried out in bovine serum albumin(BSA)-coated 96-well Terasaki plates (Robbins Scientific,Mountain View, CA) as reported earlier (Vasta et al., 1986).Protein concentrations were determined on 96-well flat-bottom plates with the BioRad Protein Assay following amodification of the manufacturer's protocols, using BSA asstandard (Ahmed et al., 1996a). Analytical PAGE in thepresence of SDS (2%) was carried out on 15% (w/v) acryl-amide gels under reducing conditions as reported elsewhere(Laemmli and Favre, 1973). The molecular weight of thenative protein was estimated by gel permeation chromato-graphy carried out on a Pharmacia Superose 6 column (1 �30 cm) as described elsewhere (Vasta et al., 1986).
Preparation of rabbit anti-Drgal1-L2 antiserum
The anti-Drgal1-L2 antiserum was prepared in New Zealandwhite rabbits at Duncroft (Lovetsville, FL) by multiplesubcutaneous and intramuscular injections of affinity-purified Drgal1-L2 (100 mg/injection), and the antibodytiter was determined by enzyme-linked immunosorbentassay (ELISA) as previously described (Vasta et al., 1986).The specificity of the antiserum was assessed by westernblot. For this purpose, the affinity-purified galectins orzebrafish crude extract were electrotransferred onto poly-vinyl difluoride membranes, and blocked with PBS/0.05%Tween 20/3% fish gelatin (Sigma) for 1 h at room tempera-ture (RT) and incubated with anti-Drgal1-L2 antibody(1:1000 dilution) for 1 h. After three washes with PBS/0.05 % Tween 20, membranes were incubated with goatanti-rabbit IgG-HRP conjugate (1:2000 dilution) in PBS(azide free)/0.05% Tween 20/3% gelatin for 1 h, washedthree times with PBS (azide free)/Tween 20, and developedwith chloronaphthol for peroxidase activity.
Characterization of the carbohydrate specificity ofDrgal1-L2
Preparation of the Drgal1-L2-HRP conjugate. For conju-gation, the DEAE-Sepharose column containing Drgal1-L2was washed with azide-free PBS (1:10) to remove lactoseand ME, and the bound protein was eluted with 1.5 ml PBS(azide-free)/0.5 M NaCl/0.1 M lactose. The galectin wasconjugated to activated HRP at 4�C overnight with stirringfollowing the procedure previously described (Ahmed et al.,2002). Briefly, activation of HRP (4 mg in 0.5 ml PBS,pH 7.2) was performed with 1 mg sulfonated succinimidyl4-(N-maleimidomethyl)cyclohexane-1-carboxylate followedby conjugation of the activated HRP with the Drgal1-L2(100 mg). After overnight incubation at 4�C, the conjugation
H. Ahmed et al.
228
by guest on June 3, 2013http://glycob.oxfordjournals.org/
Dow
nloaded from

mixture was diluted 50-fold with cold water and adsorbedonto DEAE-Sepharose (0.5 ml) preequilibrated with azide-free PBS (1:10). The column was washed to remove lactose,and the conjugate was eluted with 4 ml PBS (azide-free)/1 MNaCl, followed by purification by affinity chromatographyon lactosyl-Sepharose as indicated. The purified galectin-HRP conjugate was dialyzed with azide-free PBS and storedin 1% BSA/50% glycerol at ÿ20�C.
Solid phase binding-inhibition assay. Binding of theDrgal1-L2 to ASF, and its inhibition by sugars wereassessed and optimized as reported elsewhere (Ahmedet al., 1996a, 2002). Briefly, ASF (0.5 mg/100 ml/well) in0.1 M Na2CO3/0.02% NaN3 (pH 9.6) was adsorbed ontothe wells of microtiter plates (Immulon; Dynatech Labora-tories, Chantilly, VA) at 37�C for 3 h, and the boundglycoprotein was fixed with 2% formaldehyde in PBS at37�C for 30 min. The plates were washed three times withPBS (azide-free)/0.05% Tween 20, and incubated with thegalectin-HRP conjugate (10 ng/100 ml/well for bindingassays) or with preincubated mixture of equal volume ofconjugate and varying concentrations of test ligands (forbinding-inhibition assays). After incubation for 1 h at 4�C,the plates were washed with ice-cold azide-free PBS-Tween20 buffer, and the bound peroxidase activity was assayedwith ABTS. Binding of Drgal1-L2 to laminin and its inhibi-tion by lactose were examined by ELISA, as previouslydescribed (Ahmed et al., 1990).
Peptide sequencing of Drgal1-L2
The purified Drgal1-L2 was subjected to SDS±PAGE asdescribed before and after staining with Coommassie bluethe band was cut out and in-gel digested with trypsin in0.05 M Tris±HCl (pH 8.5)/1 M guanidine hydrochloride(E/S� 1:50, w/w, at 30�C, 10±20 h). The tryptic digestswere acidified to pH ~ 2 using 10% trifluoracetic acid, andthe fragments were purified by HPLC, analyzed by matrix-assisted laser desorption/ionization mass spectrometry,and sequenced using Edman degradation as previouslydescribed (Ahmed et al., 1996a).
Galectins primers
The oligonucleotides (degenerated and gene-specific) weredesigned and synthesized at BioAnalytical Services, Centerof Marine Biotechnology, University of Maryland Biotech-nology Institute (Baltimore, MD). The galectin primersused are defined by numbers as follows.
Drgal1-L2 primers:
1 50-GGICARACIYTIACIATHACIGGIGTICC-30 (forward)2 50-CKIACYTCICCYTCRAARTGC-30 (reverse)3 50-CKRTTNGGRAARTGDATYTC-30 (reverse)4 50-CTGTGCTCCTCACACCAGCTGCCGC-30 (reverse)5 50-CTATAGTGCACTGGTCCCCATGGCC-30 (reverse)6 50-GGGGTGCCAAAGCCTGATTCCAC-30 (forward)7 50-CCCAAGCTTCGATGGCCGGTGTGC-30 (forward)8 50-GGGAATTCCCCTATTTAATTTCAACCCCTTG-
GATCCTGAC-30 (reverse)9 50-CACAGCCCTGAAGACATCGCTCTCC-30 (forward)
10 50-GAGCCTTGACGGTTCGGGA-30 (reverse)11 50-CTGGTGACTCTTCCTGATGGTTC-30 (forward)12 50-GTGGAATCAGGCTTTGGA-30 (reverse)
Drgal1-L1 primers based on the EST sequence (GenBankaccession number AW174841):
13 50-CGCGGAATGTTCGTGATG-30 (forward)14 50-CCCTTGGATCCTAGCTTGGC-30 (reverse)
Drgal1-L3 primers based on the available sequence(Unigene Dr. 15088):
15 50-GCAGCTCCACCAACAACTCAG-30 (forward)16 50-CGTGTGTGAAGGCATCGTCT-30 (reverse)
Drgal3) primers based on the expressed sequence tagsequence (GenBank accession number AI942624):
17 50-GTGCCATCAGCTACACCACG-30 (forward)18 50-GATGCTGAGACCTGTGATCTGG-30 (reverse)19 50-CTCACGCTCTTCTCTGCCC-30 (reverse)
Drgal9-L1 primers based on the expressed sequence etagsequence (GenBank accession number AW419961):
20 50-CAGTGTGGTTCTCATTCAGAGGC-30 (forward)21 50-GCAGTGGAGGGAATGGTGG-30 (forward)22 50-CCTGAGCCACAAGGATCTGC-30 (reverse)
cDNA cloning and sequence analysis of galectins
cDNA cloning of galectins was carried out using a PCR-based protocol. For this purpose, total RNA was isolatedfrom zebrafish embryos, adult whole body, or selectedtissues using a standard guanidine thiocyanate, phenol/chloroform extraction (Sambrook et al., 1989). Occasion-ally isolation of total RNA was performed using RneasyTotal RNA System (Qiagen, Valencia, CA). Poly (A)�RNA was isolated from the total RNA on poly (dT)-Dynabeads using mRNA purification kit (Dynal, Oslo,Norway). Reverse transcription was done with Maloneymurine leukemia virus reverse transcriptase (Life Tech-nologies). All PCR amplifications were carried out withTaq DNA polymerase (Promega, Madison, WI) in thebuffer and Mg2� solution provided by the manufacturer.Annealing temperatures varied from 50�C to 65�C. PCRproducts were cloned into the pGEM-T vector (Promega).Plasmids for DNA sequencing were prepared using theQIAprep Miniprep Kit (Qiagen). DNA sequences weredetermined by the dye termination cycle sequencing methodusing an ABI PRISM BigDye Terminator Cycle Sequencingready reaction Kits with AmpliTaq DNA Polymerase(Applied Biosystems, Foster City, CA). All other manipula-tions of nucleic acids, such as ligation, transformation, gelelectrophoresis, gel elution, and preparation of buffers,were carried out following standard protocols (Sambrooket al., 1989).
For the cloning and sequencing of Drgal1-L2, adultzebrafish cDNA was amplified by PCR with degeneratedprimer sets 1 and 2. The authenticity of the PCR productwas assessed with a nested primer 3 in combination withthe primer 1. Validated PCR products were cloned intopGEM-T vector and sequenced. The cDNA sequencewas completed by 50- and 30-RACE. The 50-end of the
Molecular characterization and developmental expression of zebrafish galectins
229
by guest on June 3, 2013http://glycob.oxfordjournals.org/
Dow
nloaded from

full-length Drgal1-L2 was obtained from the 50 RNAligase-mediated RACE kit (Ambion, Austin, TX) withouter adapter primer and a gene-specific reverse primer 4according to the manufacturer's protocol. The PCRproduct generated after reamplification with the inneradapter primer and the nested gene-specific reverse primer5 was authenticated with the nested primer 6 in combina-tion with the primer 5. For 30-RACE PCR, the first strandcDNA synthesis was performed using MCSdT16VN lock-docking primer, 50-CCGCATGCGGCCGCAGATCTA-GATATCGAT16VN-30, synthesized commercially (Sigma-Genosys) based on Borson et al. (1992). The 30-RACEPCR product, generated with primer 6 and adapter primerMCSRACE, 50-CCGCAGATCTAGATATCGA-30, wassubcloned and sequenced.
The full-length cDNA for a Drgal1-L2 isoform (Drgal1-L2d) was obtained by PCR amplification using primers 7and 8. The full-length cDNAs for two other protogalectins(Drgal1-L1 and Drgal1-L3) were obtained based on avail-able sequences in the database using the primer sets 13±14and 15±16, respectively. The 50 and 30 end of the full-lengthchimera and tandem-repeat galectins from D. rerio (Drgal3and Drgal9-L1, respectively) were also performed asdescribed. The 50-RACE PCR product for Drgal3 wasobtained with outer adapter primer and primer 18 andwas authenticated with two nested primers 17 and 19. The30-RACE PCR product for Drgal3 was obtained with pri-mer 17 and MCSRACE and authenticated with primer 16in combination with the primer 17. The 30-RACE PCR forDrgal9-L1 was performed with primer 20 and MCSRACE,and the product was confirmed with two nested primers, 21and 22. The PCR products of expected size were subclonedinto pGEM-T vector and sequenced.
Genomic structure of Drgal1-L2
Genomic DNA was prepared from adult zebrafish testisfollowing a standard protocol (Sambrook et al., 1989).The purified genomic DNA was digested individually withseven restriction enzymes (EcoR V, Sca I, Dra I, Pvu II, Ssp I,Hpa I, Sma I). Seven libraries were constructed followingUniversal GenomeWalker kit (Clontech, Palo Alto, CA).For amplification of the exon I±exon III region, adapterprimer AP1 and primer 4 were used to generate a PCRproduct, which was reamplified with nested adapter primerAP2 and 5. The sequence of the product was determinedwith an internal primer, 12. For amplification of exon III±exon IV region, primers 9 and 10 were used, and the productwas authenticated with two nested reverse primers, 4 and 5,in combination with forward primer 9. Amplification ofexon IV (in part) and the 30 noncoding region was carriedout with primers 11 and AP2. The PCR products ofexpected size were subcloned into pGEM-T vector andsequenced.
Chromosomal mapping of Drgal1-L2
The Drgal1-L2 gene was mapped by radiation hybrid map-ping using LN54 panel developed by Hukriede et al. (1999).Briefly, 93 radiation hybrid and 3 control cell lines (giftfrom Dr. Marc Ekker, Loeb Health Research Institute atthe Ottawa Hospital, Ottawa, ON, Canada) were tested on
PCR assay in duplicate using gene-specific primers 4 and 6.The results obtained through PCR assay were scoredaccording to Hudson et al. (1995): 1, 0, and 2 indicate apositive, negative, and ambiguous hybrid data, respectively.The mapping of Drgal1-L2 against the LN54 panel wasobtained by accessing a Web tool through ZFIN (http://zfish.uoregon.edu/zfin)
Expression of galectins in selected zebrafish developmentalstages and adult organs by reverse transcriptase PCR
Total RNA was purified from several embryonic stages(1±19 h postfertilization and 1±5 days postfertilization;~200 embryos from each stage) from adult organs of zebra-fish. Poly (A)� RNA was isolated from the total RNA onpoly (dT)-Dynabeads as described. For reverse transcrip-tase PCR analysis, first strand cDNAs were generated fromthe purified mRNA using the cDNA synthesis kit (LifeTechnologies) and used as template for PCR amplification.The presence of the Drgal1-L2 isoform (Drgal1-L2d) wasdetermined from PCR products amplified by primers 5and 6, following separation on a 3.5% MetaPhor agarose(FMC BioProducts, Rockland, ME).
Whole mount in situ hybridization
In situ hybridization was carried out as previously described(Du and Dienhart, 2001). Briefly, whole zebrafish embryoswere fixed in 4% paraformaldehyde in azide-free PBS(pH 7.2) overnight at 4�C. After washing with PBS/0.1%Tween 20, embryos were dechorionated, soaked with acet-one atÿ20�C for 10 min, and washed five times (5 min each)with PBS-Tween. The embryos were prehybridized with amixture of 50% formamide, 5� sodium chloride (0.15 M)/sodium citrate (0.015 M) (SSC), 0.5 mg/ml tRNA, 50 mg/mlheparin, and 0.1% Tween 20 for at least 4 h at 70�C andhybridized with digoxigenin-labeled RNA (antisense orsense) probe for 4±12 h at the same temperature. Theembryos were washed with a 1:1 mixture of prehybridiza-tion buffer (without tRNA and heparin) and 2� SSC for15 min at 70�C followed by two washes (30 min each at70�C) with 0.2� SSC/0.1% 3-[(3-cholamidopropyl)dimethyl-ammonia]-1-propane sulfonate (CHAPS). Subsequently,the embryos were washed with maleic acid buffer (MAB)(0.1 M maleic acid/0.15 M NaCl, pH 7.5) for 5 min at RT,blocked with 10% goat serum/2% blocking reagent (Roche)in MAB for at least 4 h at RT, and incubated overnightat 4�C with a 1:2000 dilution of antidigoxigenin antibodyconjugated with alkaline phosphatase (Roche). This wasfollowed by six washes in PBS-Tween for 15 min each atRT and two washes with alkaline phosphatase buffer (0.1 MTris/0.05 M MgCl2/0.1 M NaCl/0.1% Tween 20, pH 9.5)for 10 min each. Color was developed with a mixture of4.5 ml nitro blue tetrazolium (75 mg/ml) and 3.5 ml 5-bromo-4-chloro-3-indolyl phosphate (BCIP) (50 mg/ml). For thepreparation of probes, plasmid vector pGEM (Promega)containing zebrafish galectin cDNA was linearized byrestriction digest with Spe I and Sac II. Antisense andsense RNA probes were synthesized by incorporatingdigoxigenin-UTP (Roche) using T7 and SP6 RNA polymer-ase (Promega) according to manufacturer's protocol.
H. Ahmed et al.
230
by guest on June 3, 2013http://glycob.oxfordjournals.org/
Dow
nloaded from

Examination of Drgal1-L2 protein expression bywhole mount antibody staining
To study Drgal1-L2 expression in zebrafish embryos, wholemount antibody staining was performed using high-titeranti-Drgal1-L2 specific antibodies (active up to 100,000dilution) following the protocol of Du et al. (1997). Forantibody labeling, embryos were fixed with 4% paraformal-dehyde for 1 h at RT, washed twice with PBS-Tween for5 min each, and soaked in cold acetone for 10 min atÿ20�C.Subsequently, embryos were washed twice with PBS-Tween5 min each, washed once with 0.1% BSA/1% dimethyl sulf-oxide/PBS (BDP) for 5 min, and incubated with avidin(Vector Laboratories, Burlingame, CA) (4 drops/ml) inblocking buffer (10% of goat serum in BDP) for 30 min atRT. The embryos were then washed twice with BDP for5 min each and incubated with 1:10,000 dilution of anti-Drgal1-L2 antibody in blocking buffer containing biotin(Vector) (4 drops/ml) overnight at 4�C. The embryos werethen washed three times for 30 min with BDP, followed byincubation with diluted (1:1000) biotin-labeled secondaryantibody (goat anti-rabbit IgG) (Vector) in BDP for 1 h atRT. Embryos were then washed three times for 30 min withBDP and incubated with 1:1 diluted avidin-biotin complexsolution (Vector) for 30 min at RT. Finally, embryos werewashed three times for 30 min in BDP and color-developedwith DAB substrate (Vector) according to the manufac-turer's protocol.
Acknowledgments
Supported by grant MCB-00-77928 from the NSF toG.R.V. and grant RO1 GM58537-01 from the NIH toS.J.D.
Abbreviations
ABTS, diammonium 2,20-azinobis(3-ethylbenzothiazoline-6-sulfonate); ASF, asialofetuin; BSA, bovine serum albu-min; CRD, carbohydrate recognition domain; ELISA,enzyme-linked immunosorbent assay; HRP, horseradishperoxidase; MAB, maleic acid buffer; ME, 2-mercapto-ethanol; ORF, open reading frame; PAGE, polyacrylamidegel electrophoresis; PCR, polymerase chain reaction; PBS,phosphate buffered saline; RACE, rapid amplification ofcDNA ends; RT, room temperature; SDS, sodium dodecylsulfate; SSC, sodium chloride (0.15 M)/sodium citrate(0.015 M).
References
Ahmed, H., and Vasta, G.R. (1994) Galectins: conservation of function-ally and structurally relevant amino acid residues defines two types ofcarbohydrate recognition domains. Glycobiology, 4, 545±549.
Ahmed, H., Allen, H.J., Sharma, A., and Matta, K.L. (1990) Humansplenic galaptin: carbohydrate-binding specificity and characterizationof the combining site Biochemistry, 29, 5315±5319.
Ahmed, H., Pohl, J., Fink, N.E., Strobel, F., and Vasta, G.R. (1996a) Theprimary structure and carbohydrate specificity of a b-galactosyl-binding lectin from toad (Bufo arenarum Hensel) ovary reveal closer
similarities to the mammalian galectin-1 than to the galectin from theclawed frog Xenopus laevis. J. Biol. Chem., 271, 33083±33094.
Ahmed, H., Fink, N.E., Pohl, J., and Vasta, G.R. (1996b) Galectin-1 frombovine spleen: biochemical characterization, carbohydrate specificityand tissue-specific isoform profiles J. Biochem. (Tokyo), 120, 1007±1019.
Ahmed, H., Bianchet, M.A., Amzel, L.M., Hirabayashi, J., Kasai, K.,Giga-Hama, Y., Tohda, H., and Vasta, G.R. (2002) Novel carbohy-drate specificity of the 16 kDa galectin from Caenorhabditis elegans:binding to blood group precursor oligosaccharides (type 1, type 2, Ta,and Tb) and gangliosides. Glycobiology, 12, 451±461.
Arata, Y., Hirabayashi, J., and Kasai, K. (1997) Structure of the 32-kDagalectin gene of the nematode Caenorhabditis elegans. J. Biol. Chem.,272, 26669±26677.
Bakkers, J., Semino, C.E., Stroband, H., Kijne, J.W., and Robbins, P.W.(1997) A important developmental role for oligosaccharides duringearly embryogenesis of cyprinid fish. Proc. Natl Acad. Sci. USA, 94,7982±7986.
Barondes, S.H., Cooper, D.N.W., Gitt, M.A., and Leffler, H. (1994)Galectins. Structure and function of a large family of animal lectins.J. Biol. Chem., 269, 20807±20810.
Blagden, C.S., Currie, P.D., Ingham, P.W., and Hughes, S. M (1997)Notochord induction of zebrafish slow muscle mediated by Sonichedgehog. Genes Dev., 11, 2163±2175.
Borson, N.D., Salo, W.L., and Drewes, L.R. (1992) A lock-dockingoligo(dT) primer for 50 and 30 RACE PCR. PCR Methods Appl., 2,144±148.
C. elegans Sequencing Consortium. (1998) Genome sequence of thenematode C. elegans: a platform for investigating biology. Science,282, 2012±2018.
Chiariotti, L., Wells, V., Bruni, C.B., and Mallucci, L. (1991) Structureand expression of the negative growth factor mouse beta-galactosidebinding protein gene. Biochim. Biophys. Acta, 1089, 54±60.
Cho, M. and Cummings, R.D. (1995) Galectin-1, a b-galactoside-bindinglectin in Chinese hamster ovary cells. I. Physical and chemicalcharacterization. J. Biol. Chem., 270, 5198±5206.
Colnot, C., Ripoche, M., Fowlis, D., Cannon, V., Scaerou, F.,Cooper D.N.W., and Poirier, F. (1997) The role of galectins in mousedevelopment. Trends Glycosci. Glycotechnol., 9, 31±40.
Colnot, C., Fowlis, D., Ripoche, M., Bouchaert, I., and Poirier, F. (1998)Embryonic implantation in galectin 1/galectin 3 double mutant mice.Develop. Dynamics, 211, 306±313.
Cooper, D.N.W. (2002) Galectinomics: a lesson in complexity. Biochim.Biophys. Acta, 1572, 209±23115.
Cooper, D.N.W. and Barondes, S.H. (1990) Evidence for export of amuscle lectin from cytosol to extracellular matrix and for a novelsecretory mechanism. J. Cell Biol., 110, 1681±1691.
Coutelle, O., Blagden, C.S., Hampson, R., Halai, C., Rigby, P.W., andHughes, S.M. (2001) Hedgehog signalling is required for maintenanceof myf5 and myoD expression and timely terminal differentiation inzebrafish adaxial myogenesis. Dev. Biol., 236, 136±150.
Currie, P.D. and Ingham, P.W (1996) Induction of a specific muscle celltype by a Hedgehog-like protein in zebrafish. Nature, 382, 452±455.
Du, S.J. and Dienhart, M. (2001) Gli2 mediation of hedgehog signals inslow muscle induction in zebrafish. Differentiation, 67, 84±91.
Du, S. J., Devoto, S., Westerfield, M., and Moon, R.T. (1997) Positive andnegative regulation of muscle cell identity by members of the hedgehogand TGF gene families. J. Cell Biol., 139, 145±156.
Dunphy, J.L., Barcham, G.J., Bischof, R.J., Young, A.R., Nash, A., andMeeusen, E.N. (2002) Isolation and characterization of a noveleosinophil-specific galectin released into the lungs in response toallergen challenge. J. Biol. Chem., 277, 14916±14924.
Gitt, M.A. and Barondes, S.H. (1986) Evidence that a human solublebeta-galactoside-binding lectin is encoded by a family of genes. Proc.Natl Acad. Sci. USA, 83, 7603±7607.
Gitt, M.A., and Barondes, S.H. (1991) Genomic sequence and organiza-tion of two members of a human lectin gene family. Biochemistry, 30,82±89.
Gitt, M.A., Massa, S.M., Leffler, H., and Barondes, S.H. (1992) Isolationand expression of a gene encoding L-14-II, a new human solublelactose-binding lectin. J. Biol. Chem., 267, 10601±10606.
Molecular characterization and developmental expression of zebrafish galectins
231
by guest on June 3, 2013http://glycob.oxfordjournals.org/
Dow
nloaded from

Goldring, K., Jones, G.E., Thiagarajah, R., and Watt, D.J. (2002) Theeffect of galectin-1 on the differentiation of fibroblasts and myoblastsin vitro. J. Cell. Sci., 115, 355±366.
Gorski, J.P., Liu, F.T., Artigues, A., Castagna, L.F., and Osdoby, P.(2002) New alternatively spliced form of galectin-3, a member of thebeta-galactoside-binding animal lectin family, contains a predictedtransmembrane-spanning domain and a leucine zipper motif. J. Biol.Chem., 277, 18840±18848.
Gritzmacher, C.A., Mehl, V.S., and Liu, F.T. (1992) Genomic cloningof the gene for an IgE-binding lectin reveals unusual utilization of50 untranslated regions. Biochemistry, 31, 9533±9538.
Gu, M., Wang, W., Song, W.K., Cooper, D.N.W., and Kaufman, S.J.(1994) Selective modulation of the interaction of a7b1 integrin withfibronectin and laminin by L-14 lectin during skeletal muscledifferentiation. J. Cell Sci., 107, 175±181.
Hathaway, H.J. and Shur, B.D. (1997) Transgenic approachesto glycobiology. In Gabius, H.-J. and Gabius, S. (Eds.),Glycosciences: status and perspectives. Chapman & Hill, Weinheim,pp. 507±517.
Hirabayashi, J. (ed.) (1997) Recent topics on galectins. Trends Glycosci.Glycotechnol., 9, 1±180.
Hirabayashi, J. and Kasai, K. (1993) The family of metazoan metal-independent b-galactoside-binding lectins:structure, function andmolecular evolution. Glycobiology, 3, 297±304.
Hirabayashi, J., Ubukata, T., and Kasai, K. (1996) Purification andmolecular characterization of novel 16-kDa galectin from thenematode Caenorhabditis elegans. J. Biol. Chem., 271, 2497±2505.
Hirabayashi, J., Arata, Y., and Kasai, K. (1997) Galectins from thenematode Caenorhabditis elegans and the Genome Project. TrendsGlycosci. Glycotechnol., 9, 113±122.
Hudson, T.J., Stein, L.D., Gerety, S.S., Ma, J., Castle, A.B., Silva, J.,Slomin, D.K., Baptista, R., Kruglyak, L., Xu, S.H., and others.(1995) An STS-based map of the human genome. Science, 270,1945±1954.
Hukriede, N.A., Joly, L., Tsang, M., Miles, J., Tellis, P., Epstein, J.A.,Barbazuk, W.B., Li, F.N., Paw, B., Postlethwait, J.H., and others.(1999) Radiation hybrid mapping of the zebrafish genome. Proc. NatlAcad. Sci. USA, 96, 9745±9750.
Laemmli, U.K. and Favre, M. (1973) Maturation of the headof bacteriophage T4. I. DNA packaging events. J. Mol. Biol., 80,575±599.
Laine, R.A. (1997) The information-storing potential of the sugar code. InGabius, H.-J. and Gabius, S. (Eds.), Glycosciences: status andperspectives. Chapman & Hill, Weinheim, pp. 5±14.
Liao, D.I., Kapadia, G., Ahmed, H., Vasta, G.R., and Herzberg, O.(1994) Structure of S-lectin, a developmentally regulated vertebrateb-galactoside-binding protein. Proc. Natl Acad. Sci. USA, 91, 1428±1432.
Lin, X. and Perrimon, N. (1999) Dally cooperates with DrosophilaFrizzled 2 to transduce Wingless signalling. Nature, 400, 281±284.
Muramoto, K. and Kamiya, H. (1992) The amino-acid sequence of alectin from conger eel, Conger myriaster, skin mucus. Biochim.Biophys. Acta, 1116, 129±136.
Oda, Y., Herrmann, J., Gitt, M., Turck, C.W., Burlingame, A.L.,Barondes, S.H., and Leffler, H. (1993) Soluble lactose-binding lectinfrom rat intestine with two different carbohydrate-binding domains inthe same peptide chain. J. Biol. Chem., 268, 5929±5939.
Ohyama, Y. and Kasai, K. (1988) Isolation and characterization of the chick14K beta-galactoside-binding lectin gene. J. Biochem., 104, 173±177.
Pace, K.E., Lebestky, T., Hummel, T., Arnoux, P., Kwan, K., and Baum, L.G.(2002) Characterization of a novel Drosophila melanogaster galectin.Expression in developing immune, neural, and muscle tissues. J. Biol.Chem., 277, 13091±13098.
Paroutaud, P., Levi, G., Teichberg, V.I., and Strosberg, A.D. (1987)Extensive amino acid sequence homologies between animal lectins.Proc. Natl Acad. Sci. USA, 84, 6345±6348.
Parsons, M.J., Pollard, S.M., Saude, L., Feldman, B., Coutinho, P.,Hirst, E.M., and Stemple, D.L. (2002) Zebrafish mutants identify anessential role for laminins in notochord formation. Development, 129,3137±3146.
Perrimon, N. and Bernfield, M, (2000) Specificities of heparan sulphateproteoglycans in developmental processes. Nature, 404, 725±728.
Robertson, M.W., Albrandt, K., Keller, D., and Liu, F.T. (1990) HumanIgE-binding protein: a soluble lectin exhibiting a highly conservedinterspecies sequence and differential recognition of IgE glycoforms.Biochemistry, 29, 8093±8100.
Sambrook, J., Fritsch, E.F., and Maniatis, T. (1989) Molecular cloning. Alaboratory manual, 2nd ed. Cold Spring Harbor Laboratory Press,Cold Spring Harbor, New York.
Schwarz, F., Ahmed, H., Bianchet, M.A., Amzel, L.M., and Vasta, G.R.(1998) Thermodynamics of bovine spleen galectin-1 binding todisaccharides: correlation with structure and its effect on oligomeriza-tion at the denaturation temperature. Biochemistry, 37, 5867±5877.
Shur, B.D. (1993) Glycosyltransferases as cell adhesion molecules. Curr.Opin. Cell. Biol., 5, 854±863.
Sparrow, C.P., Leffler, H., and Barondes, S.H. (1987) Multiple solubleb-galactoside-binding lectins from human lung. J. Biol. Chem., 262,7383±7390.
Stern, H.M., Brown, A.M., and Hauschka, S.D. (1995) Myogenesis inparaxial mesoderm: preferential induction by dorsal neural tube andby cells expressing Wnt-1. Development, 121, 3675±3686.
Tajbakhsh, S., Borello, U., Vivarelli, E., Kelly, R., Papkoff, J., Duprez, D.,Buckingham, M., and Cossu, G. (1998) Differential activation ofMyf5 and MyoD by different Wnts in explants of mouse paraxialmesoderm and the later activation of myogenesis in the absence ofMyf5. Development, 125, 4155±4162.
Van den Brule, F.A., Fernandez, P.L., Buicu, C., Liu, F.T., Jackers, P.,Lambotte, R., and Castronovo, V. (1997) Differential expression ofgalectin-1 and galectin-3 during first trimester human embryogenesis.Dev. Dyn., 209, 399±405.
Vasta, G.R., Hunt, J.C., Marchalonis, J.J., and Fish, W.W. (1986)Galactosyl-binding lectins from the tunicate Didemnum candidum.Purification and physicochemical characterization. J. Biol. Chem., 261,9174±9181.
Vasta, G.R., Quesenberry, M., Ahmed, H., and O'Leary, N. (1999) C-typelectins and galectins mediate innate and adaptive immune functions:their roles in the complement activation pathway. Dev. Comp.Immunol., 23, 401±420.
Wada, J. and Kanwar, Y.S. (1997) Identification and characterization ofgalectin-9, a novel beta-galactoside-binding mammalian lectin. J. Biol.Chem., 272, 6078±6086.
Wassarman, P.M. (1995) Towards molecular mechanisms for gameteadhesion and fusion during mammalian fertilization. Curr. Opin. CellBiol., 7, 658±664.
H. Ahmed et al.
232
by guest on June 3, 2013http://glycob.oxfordjournals.org/
Dow
nloaded from




















