
Recognition of HLA-A2-restricted mammaglobin-A-derived epitopes
13
Report Recognition of HLA-A2-restricted mammaglobin-A-derived epitopes by CD8+ cytotoxic T lymphocytes from breast cancer patients Andre´ s Jaramillo 1 , Kishore Narayanan 1 , Lacey G. Campbell 1 , Nicholas D. Benshoff 1 , Lonnie Lybarger 2 , Ted H. Hansen 2,3 , Timothy P. Fleming 1 , Jill R. Dietz 1 , and T. Mohanakumar 1,3 1 Department of Surgery; 2 Department of Genetics; 3 Department of Pathology and Immunology, Washington University School of Medicine, St. Louis, MO, USA Key words: CD8+ T lymphocytes, epitope, HLA-A2, mammaglobin-A, tumor-specific antigen Summary A breast cancer-associated antigen, mammaglobin-A, is specifically expressed in 80% of primary breast tumors. The definition of immune responses against this highly expressed breast cancer-specific antigen should be of great value in the development of new therapeutic strategies for breast cancer. Thus, the purpose of this study was to identify HLA-A2-restricted mammaglobin-A-derived epitopes recognized by CD8+ cytotoxic T lymphocytes (CTL). We identified seven mammaglobin-A-derived candidate epitopes that bind the HLA-A2 molecule (Mam-A2.1-7) by means of a HLA class I-peptide binding computer algorithm from the Bioinformatics & Molecular Analysis Section of the National Institutes of Health. Subsequently, we determined that CD8+ CTLs from breast cancer patients reacted to the Mam-A2.1 (83– 92, LIYDSSLCDL), Mam-A2.2 (2–10, KLLMVLMLA), Mam-A2.3 (4–12, LMVLMLAAL), Mam-A2.4 (66–74, FLNQTDETL), and Mam-A2.7 (32–40, TINPQVSKT) epitopes using an IFN-c ELISPOT assay. Interestingly, healthy individuals also showed high reactivity to the Mam-A2.2 epitope. Two CD8+ CTL lines generated in vitro against TAP-deficient T2 cells loaded with the candidate epitopes showed significant cytotoxic activity against the Mam-A2.1-4 epitopes. These CD8+ CTL lines recognized a HLA-A2+ breast cancer cell line expressing the Mam-A2.1 epitope. In addition, DNA vaccination of HLA-A2+/human CD8+ double-transgenic mice with a DNA construct encoding the Mam-A2.1 epitope and the HLA-A2 molecule induced a significant expansion of epitope-specific CD8+ CTLs that recognize the same HLA- A2+/Mam-A2.1+ breast cancer cell line. In conclusion, these results demonstrate the immunotherapeutic potential of mammaglobin-A for the treatment and prevention of breast cancer. Introduction Tumor-specific CD8+ cytotoxic T lymphocytes (CTL) have been well documented in melanoma, renal cell carcinoma, and ovarian cancers [1–3]. However, it has been difficult to consistently iden- tify a similar tumor-specificity in the CD8+ CTL response to other types of tumors including breast cancer [1–3]. Despite this problem, it has been shown that tumor-infiltrating cells obtained from metastatic effusions of breast cancer patients con- tained CD8+ CTLs that recognized autologous tumor cells in a HLA class I-restricted tumor-spe- cific manner [4]. With the development of our understanding of epitope recognition by CD8+ CTLs, it has been possible to identify breast cancer- associated antigens expressed in a wide variety of tumors such as MUC1 and Her2/neu [5–11]. It has been shown that vaccination with dendritic cells pulsed with HER2/neu- or MUC1-derived peptides Breast Cancer Research and Treatment 88: 29–41, 2004. Ó 2004 Kluwer Academic Publishers. Printed in the Netherlands.
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Recognition of HLA-A2-restricted mammaglobin-A-derived epitopes

Report
Recognition of HLA-A2-restricted mammaglobin-A-derived epitopes
by CD8+ cytotoxic T lymphocytes from breast cancer patients
Andres Jaramillo1, Kishore Narayanan1, Lacey G. Campbell1, Nicholas D. Benshoff1,Lonnie Lybarger2, Ted H. Hansen2,3, Timothy P. Fleming1, Jill R. Dietz1, andT. Mohanakumar1,31Department of Surgery; 2Department of Genetics; 3Department of Pathology and Immunology, WashingtonUniversity School of Medicine, St. Louis, MO, USA
Key words: CD8+ T lymphocytes, epitope, HLA-A2, mammaglobin-A, tumor-specific antigen
Summary
A breast cancer-associated antigen, mammaglobin-A, is specifically expressed in 80% of primary breasttumors. The definition of immune responses against this highly expressed breast cancer-specific antigenshould be of great value in the development of new therapeutic strategies for breast cancer. Thus, thepurpose of this study was to identify HLA-A2-restricted mammaglobin-A-derived epitopes recognized byCD8+ cytotoxic T lymphocytes (CTL). We identified seven mammaglobin-A-derived candidate epitopesthat bind the HLA-A2 molecule (Mam-A2.1-7) by means of a HLA class I-peptide binding computeralgorithm from the Bioinformatics & Molecular Analysis Section of the National Institutes of Health.Subsequently, we determined that CD8+ CTLs from breast cancer patients reacted to the Mam-A2.1 (83–92, LIYDSSLCDL), Mam-A2.2 (2–10, KLLMVLMLA), Mam-A2.3 (4–12, LMVLMLAAL), Mam-A2.4(66–74, FLNQTDETL), and Mam-A2.7 (32–40, TINPQVSKT) epitopes using an IFN-c ELISPOT assay.Interestingly, healthy individuals also showed high reactivity to the Mam-A2.2 epitope. Two CD8+ CTLlines generated in vitro against TAP-deficient T2 cells loaded with the candidate epitopes showed significantcytotoxic activity against the Mam-A2.1-4 epitopes. These CD8+ CTL lines recognized a HLA-A2+ breastcancer cell line expressing the Mam-A2.1 epitope. In addition, DNA vaccination of HLA-A2+/humanCD8+ double-transgenic mice with a DNA construct encoding the Mam-A2.1 epitope and the HLA-A2molecule induced a significant expansion of epitope-specific CD8+ CTLs that recognize the same HLA-A2+/Mam-A2.1+ breast cancer cell line. In conclusion, these results demonstrate the immunotherapeuticpotential of mammaglobin-A for the treatment and prevention of breast cancer.
Introduction
Tumor-specific CD8+ cytotoxic T lymphocytes(CTL) have been well documented in melanoma,renal cell carcinoma, and ovarian cancers [1–3].However, it has been difficult to consistently iden-tify a similar tumor-specificity in the CD8+ CTLresponse to other types of tumors including breastcancer [1–3]. Despite this problem, it has beenshown that tumor-infiltrating cells obtained from
metastatic effusions of breast cancer patients con-tained CD8+ CTLs that recognized autologoustumor cells in a HLA class I-restricted tumor-spe-cific manner [4]. With the development of ourunderstanding of epitope recognition by CD8+CTLs, it has been possible to identify breast cancer-associated antigens expressed in a wide variety oftumors such as MUC1 and Her2/neu [5–11]. It hasbeen shown that vaccination with dendritic cellspulsed with HER2/neu- orMUC1-derived peptides
Breast Cancer Research and Treatment 88: 29–41, 2004.� 2004 Kluwer Academic Publishers. Printed in the Netherlands.

induces expansion of peptide-specific CD8+ CTLsin breast cancer patients [12]. However, the fre-quency of expression of Her2/neu is 20–30% onbreast tumors [13, 14]. Therefore, the incidence ofpresentation of CD8+ CTL epitopes derived fromthis antigen is too low to be considered for clinicallyeffective immune therapies for breast cancer. Incontrast, MUC1 is expressed on more than 80% ofbreast tumors and is therefore a suitable candidatefor broadly applicable immune therapies [15–17].
A new breast cancer-associated antigen, mam-maglobin-A, has been identified using a differentialscreening approach [18–20]. Mammaglobin-A hasseveral properties which makes it a clinically rele-vant breast cancer-associated marker [21–24]. LikeMUC1, mammaglobin-A is expressed on morethan 80% of breast tumors [19, 20, 24]. Interest-ingly, the levels of mammaglobin-A expression arecomparable among well-differentiated, moderatelydifferentiated, and poorly differentiated breast tu-mors [24]. Due to the exclusive expression of thisantigen by breast cancer cells, mammaglobin-A hasbeen shown to be a superior marker for detectionof breast cancer cells in peripheral blood, bonemarrow, and lymph nodes as compared to othermarkers such as HER2/neu and cytokeratin-19[21–23].
The definition of immune responses againstbroadly expressed breast cancer-specific antigenssuch as mammaglobin-A should be of great help inthe development of new therapeutic strategies forthe treatment and prevention of breast cancer. Inthis regard, we have previously identified fourHLA-A3-restricted mammaglobin-A-derived epi-topes recognized by breast cancer patients in vivoand showed that a CD8+ CTL line developed invitro against these epitopes recognizes one of them(23–31, PLLENVISK) on breast cancer cells [25].Studies by other investigators have also shownthat CD8+ CTL lines developed in vitro againstHLA-A2-restricted mammaglobin-A-derived can-didate epitopes recognize one of them (80–89,FMQLIYDSSL) on breast cancer cells [26]. In thepresent study, we identified four additional HLA-A2-restricted mammaglobin-A-derived epitopesrecognized by breast cancer patients in vivo. Inaddition, we show that CD8+ CTLs generated byin vitro against these epitopes and in vivo by meansof DNA vaccination have the ability to recognizebreast cancer cells in a HLA-A2-restricted, epi-tope-specific fashion.
Materials and methods
Study subjects
Eight HLA-A2-positive patients with histologicaldiagnosis of breast carcinoma and five HLA-A2-positive healthy female individuals were includedin this study. HLA typing of the study subjects wasperformed using sequence-specific oligonucleotideprobes that provided low-medium resolutionfor HLA-A alleles (Dynal Biotech, LafayetteHill, PA). This study was performed followingthe guidelines of the Helsinki Declaration of theWorld Medical Association and approved bythe Institutional Review Board of WashingtonUniversity School of Medicine. Informed writtenconsent was obtained from all patients and con-trols.
Mice
C57BL/6 mice carrying the HLA-A*0201 (HLA-A2+) and human CD8 (hCD8+) genes werekindly provided by Dr. Victor H. Engelhard(University of Virginia, Charlottesville, VA) andDr. Linda A. Sherman (The Scripps ResearchInstitute, La Jolla, CA), respectively [27, 28]. F1mice from a cross between these two mousetransgenic lines (HLA-A2+/hCD8+) were used inthis study.
Breast cancer cell lines
Breast cancer cell lines used in this study wereobtained from the American Type Culture Col-lection (Manassas, VA) (Table 1). HLA typing ofthe breast cancer cell lines was performed as de-scribed above. Mammaglobin-A expression wasdetermined by reverse transcriptase-polymerasechain reaction and western blotting as previouslydescribed [19, 24].
Peptides
Mammaglobin-A-derived peptides that bind theHLA-A2 molecule were identified using the HLAclass I-peptide binding computer algorithm fromthe Bioinformatics & Molecular Analysis Sectionof the National Institutes of Health at http://bi-mas.dcrt.nih.gov/molbio/hla_bind/ [29]. Sevenmammaglobin-A-derived peptides with varying
A Jaramillo et al.30

levels of affinity for the HLA-A2 molecule wereused in our study (Table 2). Peptides were synthe-sized by Research Genetics (Hunsville, AL). Thepurity of peptides was determined by high-perfor-mance liquid chromatography and mass spec-trometry. The peptides were dissolved in dimethylsulphoxide (Sigma, St. Louis, MO) at a concen-tration of 10 mg/ml and stored at )70 �C until use.
HLA-A2 membrane stabilization assay
The HLA-A2-binding ability of the peptides wasconfirmed by cell membrane stabilization of the
HLA-A2 molecule in the TAP-deficient T2 cells.Briefly, T2 cells (1 · 106/ml) were incubated inflat-bottom 96-well plates at 25 �C in the presenceof each peptide (40 lg/ml) in 200 ll of RPMI-1640medium (Gibco, Grand Island, NY) supplementedwith 10% defined fetal bovine serum (HyClone,Logan, UT), 100 lM non-essential amino acids,2 mM LL-glutamine, 25 mM HEPES, 1 mM so-dium pyruvate, 100 units/ml penicillin, and100 lg/ml streptomycin. Three micrograms permillilitre of human b2m (Sigma) was also added tothe cultures. After 24 h, the T2 cells were washed(3x) and the levels of HLA-A2 expression weredetermined by flow cytometric analysis. Briefly,the T2 cells were incubated for 30 min at 4 �C withthe BB7.2 anti-HLA-A2 monoclonal antibody(mAb, 10 lg/ml) in phosphate-buffered saline(PBS) supplemented with 2% bovine serum albu-min (BSA), 25 mM HEPES, and 0.02% sodiumazide. The cells were then washed (3x) and incu-bated for 30 min at 4 �C with FITC-conjugatedgoat anti-mouse IgG (10 lg/ml, Becton Dickin-son, Franklin Lakes, NJ). The cells were thenwashed (3x), fixed in 1% paraformaldehyde, andused in a single-color flow cytometric analysis in aFACScan flow cytometer (Becton Dickinson). T2cells cultured in the presence of the HLA-A2-re-stricted GILGFVFTL influenza-derived epitopewere used as positive control [30]. Results ex-pressed as the mean fluorescence shift corre-sponded to the difference between the mean
Table 1. Breast cancer cell lines
Breast cancer cell line Tumor classification HLA-A genotypea Mammaglobin-A
expression (mRNA)bMammaglobin-A
expression (protein)c
UACC-812 Adenocarcinoma HLA-A2,68 + +
AU-565 Adenocarcinoma HLA-A2,3 + +
HBL-100 SV40-transformed HLA-A1,2 + +
MCF-7 Adenocarcinoma HLA-A2, ) ) )
MDA-MB-231 Adenocarcinoma HLA-A2, ) ) )
DU-4475 Adenocarcinoma HLA-A3,31 + NDd
MDA-MB-361 Adenocarcinoma HLA-A1,24 + +
MDA-MB-415 Adenocarcinoma HLA-A30,33 + +
ZR-75-1 Adenocarcinoma HLA-A11, ) + ND
aHLA typing was performed using sequence-specific oligonucleotide probes that provided low-medium resolution for HLA-A alleles
(Dynal Biotech).bDetermined by reverse transcriptase-polymerase chain reaction as previously described [19].cDetermined by western blotting as previously described [24].dNot determined.
Table 2. HLA-A2-restricted mammaglobin-A-derived candi-
date epitopes
Peptide Amino acid
position
Peptide
sequence
HLA-A2-
binding scorea
Mam-A2.1 83–92 LIYDSSLCDL 150.7
Mam-A2.2 2–10 KLLMVLMLA 148.0
Mam-A2.3 4–12 LMVLMLAAL 60.3
Mam-A2.4 66–74 FLNQTDETL 48.2
Mam-A2.5 73–81 TLSNVEVFM 27.3
Mam-A2.6 81–89 MQLIYDSSL 13.6
Mam-A2.7 32–40 TINPQVSKT 2.4
a Estimate of half time of HLA-A2-peptide disassociation based
on the HLA-peptide binding prediction software from the
Bioinformatics & Molecular Analysis Section of the National
Institutes of Health at http://bimas.dcrt.nih.gov/molbio/hla_
bind/ [29].
CD8+ CTLs to mammaglobin-A in breast cancer patients 31

fluorescence obtained with T2 cells cultured in thepresence of the peptide (experimental) and themean fluorescence obtained with ‘empty’ T2 cellscultured in the absence of peptides (negative con-trol). No mean fluorescence shift was observedwith the MOPC-11 mAb used as isotype control(data not shown).
Generation of epitope-reactive human CD8+CTL lines
After monocyte and dendritic cell depletion byadherence to plastic for 90 min at 37 �C, periph-eral blood lymphocytes (2 · 106) were cultured in2 ml of RPMI-1640 medium supplemented as de-scribed above in 24-well plates in the presence of apool of irradiated (10,000 rads) T2 cells (1 · 106)individually loaded with the mammaglobin-A-de-rived peptides as described above. At this time,b2m (3 lg/ml) and the CD28.2 anti-CD28 mAb(500 ng/ml, BD Biosciences, San Diego CA) werealso added to the cultures. In addition, re-combinant human IL-2 (20 U/ml, Chiron,Emeryville, CA) and human T-STIM� withoutphytohemagglutinin (10%, BD Biosciences) wereadded to the cultures after 24 h. The T cells(2 · 105) were restimulated every 8–10 days withirradiated, peptide-loaded T2 cells (1 · 106) in thepresence of irradiated (2000 rads) autologousperipheral blood lymphocytes (2 · 106) in 24-wellplated in 2 ml of culture medium supplementedwith IL-2, b2m, and anti-CD28. After three stim-ulations, the CD8+ CTLs were purified by nega-tive selection in a MiniMacs separation columnusing anti-CD4-coated microbeads (Miltenyi Bio-tec, Auburn, CA). The CD3+ cells purified by thismethod were >95% CD8+ as determined by flowcytometric analysis (data not shown). The cyto-toxic activities of the resulting CD8+ CTL lineswere analyzed after four to six stimulations.
DNA constructs
Two DNA constructs containing the coding se-quences for the Mam-A2.1 peptide, murine b2m,and HLA-A*0201 molecules (Mam-A2.1-b2m-HLA-A2) or the coding sequences for the GIL-GFVFTL influenza-derived epitope, murine b2m,and HLA-A*0201 molecules (Influenza-b2m-HLA-A2) were generated as previously described[31, 32]. Briefly, the constructs encoded the leader
sequence of murine b2m followed by the corre-sponding peptide sequence (Mam-A2.1 or Influ-enza), then a linker of 10–15 residues [(G4S)2–3].This first linker was followed by the murine b2msequence, a second linker of 15–20 residues[(G4S)3–4], and the HLA-A2 a chain sequence. Theconstructs were expressed using the pIRES.neovector (Clontech Laboratories, Palo Alto, CA).
DNA vaccination
DNA was prepared and immunizations wereconducted as previously described [31, 32]. Briefly,HLA-A2+/hCD8+ mice were injected intramus-cularly in the quadriceps with 100 lg of the Mam-A2.1-b2m-HLA-A2 or the Influenza-b2m-HLA-A2(control) DNA constructs. The mice were inocu-lated four times separated by 2-week intervals.Eight weeks later, spleen cells from vaccinatedmice were harvested, separated from erythrocytesby means of centrifugation through Lympholyte-M� (Cedarlane Laboratories, Hornby, Ontario,Canada), and tested for CD8+ CTL activityagainst peptide-loaded T2 cells and breast cancercells. This protocol has been previously shown toinduce high levels of epitope-specific CD8+ CTLsagainst a similar construct coding for an ovalbu-min-derived epitope, b2m, and H-2Kb molecules[31, 32].
ELISPOT assay
MultiScreen� 96-well filtration plates (Millipore,Bedford, MA) were coated overnight at 4 �C with5.0 lg/ml of a capture human IFN-c-specific mAb(BD Biosciences) in 0.05 M carbonate-bicarbonatebuffer (pH: 9.6). Then, the plates were blockedwith 1% BSA for 1 h and washed (3x) with PBS.Subsequently, 3 · 105 peripheral blood mononu-clear cells were cultured ex vivo in quadruplicatewells in the mAb-coated plates in 200 ll of RPMI-1640 medium supplemented as described above inthe presence of individual peptides (40 lg/ml).Cells cultured in the presence of the GILGFVFTLinfluenza-derived epitope were used as positivecontrol. Cells cultured in culture medium alonewere used as negative control. After 48 h, theplates were washed with PBS (3x) and PBS sup-plemented with 0.05% Tween-20 (3x). Then,2.0 lg/ml of a biotinylated human IFN-c-specificmAb (BD Biosciences) in PBS/BSA/Tween-20 was
A Jaramillo et al.32

added to the wells. After an overnight incubationat 4 �C, the plates were washed (3x) and horse-radish peroxidase-labeled streptavidin (BD Bio-sciences), diluted 1:2000 in PBS/BSA/Tween-20,was added to the wells. After 2 h, 3-amino-9-eth-ylcarbazole substrate reagent (BD Biosciences)was added to the wells for 5–10 min. The plateswere washed with tap water to stop the reactionand air-dried. Then, the spots were analyzed in anImmunoSpot Series I analyzer (Cellular Technol-ogy, Cleveland, OH) that was designed to detectspots with pre-determined criteria for size, shape,and colorimetric density. A test was consideredpositive if the number of spots in the experimentalcultures was significantly different from the num-ber of spots in the negative control cultures (Stu-dent t-test with the a set at P<0.01). The numberof spots in the negative control cultures wereconsistently £5 spots/well. The number of spots inthe negative control cultures was subtracted fromthe number of spots in the experimental cultures.Results are expressed as the number of IFN-c-positive cells/1 · 106 peripheral blood mononu-clear cells.
To test the functional activity of murine CD8+CTLs from DNA-vaccinated mice, 5 · 105 spleencells were cultured ex vivo in quadruplicate wells inMultiScreen� 96-well filtration plates coated witha capture mouse IFN-c-specific mAb (BD Bio-sciences) in 200 ll of RPMI-1640 medium sup-plemented as described above in the presence ofirradiated (10,000 rads) peptide-loaded T2 cells(5 · 105). Cells cultured in the presence of ‘empty’T2 cells (no peptide) were used as negative control.After 48 h, the cells were washed and the plateswere coated with a biotinylated mouse IFN-c-specific mAb (BD Biosciences), horse radish per-oxidase-labeled streptavidin, and substrate reagentas described above. Then, the plates were analyzedas described above. Results are expressed as thenumber of IFN-c-positive cells/1 · 106 spleencells.
Cytotoxicity assay
Target cells (T2 or breast cancer cells) were labeledwith 51Cr (Na51CrO4, 250 lCi, ICN Pharmaceu-ticals, Costa Mesa, CA) in 100 ll of culture med-ium. After 1 h, 5 · 103 labeled cells were plated inquadruplicate cultures in round bottom 96-wellplates in the presence of varying numbers of hu-
man CD8+ CTLs (E:T ratios: 50, 25, and 12.5)and incubated at 37 �C overnight. A 20-fold excess(1 · 105) of the HLA class I-negative K562 cellline was added to each well to inhibit non-specificlysis by NK cells. Epitope-specificity of breastcancer cell lysis was further determined in a coldtarget inhibition assay by analyzing the capacity ofpeptide-loaded unlabeled T2 cells (20-fold excess)to block the lysis of breast cancer cells. For anti-body blocking experiments, the target cells wereincubated with the BB7.2 (anti-HLA-A2, 10 lg/ml) or the MOPC-11 (isotype control, 10 lg/ml)mAbs for 30 min before addition of the effectorCD8+ CTLs. To test the functional activity ofCD8+ CTLs from DNA-vaccinated HLA-A2+/hCD8+ mice, 5 · 103 labeled breast cancer cellswere plated in quadruplicate cultures in roundbottom 96-well plates in the presence of 2.5 · 105
spleen cells (E:T ratio: 50) and incubated at 37 �Covernight. Control wells for determining sponta-neous 51Cr release contained labeled target cellsalone. Maximal release was determined by adding1% Triton X-100 to the target cells. The percentagespecific lysis was calculated as follows: [(experi-mental 51Cr release) ) (spontaneous 51Cr release)/(maximum 51Cr release) ) (spontaneous 51Cr re-lease)] · 100.
Results
Identification of HLA-A2-restrictedmammaglobin-A-derived CD8+ T cell epitopes
Using a HLA class I-peptide binding predictioncomputer program [29], we screened the mamma-globin-A protein sequence and identified sevencandidate epitopes with different binding affinitiesto the HLA-A2 molecule (Table 2). The actualbinding affinity of the peptides was determined bymeans of a HLA-A2 membrane stabilization assayin T2 cells. As shown in Figure 1, all the peptidesshowed a significant affinity for the HLA-A2molecule comparable to the one observed for theGILGFVFTL influenza-derived epitope. The dis-crepancies between the predicted affinities of thepeptides and their corresponding membrane sta-bilization abilities of the HLA-A2 molecule may bedue to the fact that the binding affinity of a givenpeptide is determined by its amino acid sequence(binding motifs) as well as the 3-dimensional
CD8+ CTLs to mammaglobin-A in breast cancer patients 33

structure of the peptide in the MHC class I groove[33, 34]. In this regard, it has been previouslyshown that the affinity of a given peptide to aMHC class I molecule does not necessarily corre-late with its ability to generate CD8+ CTLs [35,36]. For this reason, we used all the peptides toexamine the profile of CD8+ CTL reactivity ineight HLA-A2+ breast cancer patients and fiveHLA-A2+ healthy individuals by means of anIFN-c ELISPOT assay. As shown in Table 3,CD8+ CTL reactivity was observed against pep-tides Mam-A2.1, Mam-A2.2, Mam-A2.3, Mam-A2.4, and Mam-A2.7. Six patients (75%) showedCD8+ CTL reactivity against peptide Mam-A2.2,five patients (62.5%) showed CD8+ CTL reactiv-ity against peptides MamA-2.1, Mam-A2.3, andMam-A2.7, and four patient showed reactivityagainst peptides Mam-A2.4 (50%). Different pat-terns of HLA-A2-restricted epitope recognitionwas not detected in these patients as previouslyobserved against HLA-A3-restricted epitopes [25].These epitopes are different from a previously de-tected HLA-A2-restricted mammaglobin-A-de-rived epitope (80–89, FMQLIYDSSL) [26]. It is
Figure 1. Membrane stabilization of the HLA-A2 molecule
by mammaglobin-A-derived candidate epitopes. The com-
puter predicted HLA-A2-binding ability of the candidate
epitopes was confirmed by membrane stabilization of the
HLA-A2 molecule in T2 cells. Toward this, T2 cells were
incubated in the presence of each peptide. After 24 h, the
levels of HLA-A2 expression was determined by means of
flow cytometric analysis using the BB7.2 anti-HLA-A2 mAb.
T2 cells incubated in the presence of the GILGFVFTL
influenza-derived epitope were used as positive control. Re-
sults expressed as the mean fluorescence shift represent to
the difference between the mean fluorescence obtained with
T2 cells cultured in the presence of peptides and the mean
fluorescence obtained with ‘empty’ T2 cells cultured in the
absence of peptides.
Table 3. Recognition of HLA-A2-restricted mammaglobin-A-derived epitopes by CD8+ CTLs from breast cancer patientsa
Mam-A2.1 Mam-A2.2 Mam-A2.3 Mam-A2.4 Mam-A2.5 Mam-A2.6 Mam-A2.7 Influenza
Patients
1 0b 27c 40 0 0 0 0 24
2 300 0 15 307 0 0 73 83
3 40 20 0 3 0 0 0 27
4 77 233 0 107 0 3 257 50
5 0 0 0 0 0 0 37 9
6 50 14 89 74 0 6 60 53
7 3 33 30 3 0 3 9 106
8 19 24 16 19 0 3 15 19
Controls
1 0 40 0 0 0 0 0 37
2 0 128 6 0 0 0 0 14
3 0 0 9 3 0 0 0 21
4 0 85 0 0 6 0 0 119
5 9 49 6 0 6 6 0 33
a IFN-c-positive cells/1 · 106 peripheral blood mononuclear cells.b 0: Experimental cultures £ negative control cultures.c Bold: P<0.01 (student t-test). Experimental cultures versus negative control cultures.
A Jaramillo et al.34

noteworthy that four (80%) healthy individualsalso showed a significantly high CD8+ CTLreactivity against peptide Mam-A2.2 comparableto the one observed in the breast cancer patients.This peptide has been previously shown to inducethe generation of CD8+ CTLs from healthyindividuals [26]. Overall, these results clearlyidentify peptides MamA2.1, Mam-A2.3, Mam-A2.4, and Mam-A2.7 as HLA-A2-restrictedCD8+ CTL epitopes presented by breast tumors.
Immunodominance of the HLA-A2-restrictedCD8+ CTL response against mammaglobin-A-derived epi- topes in vitro
To determine whether the HLA-A2-restrictedmammaglobin-A-derived epitopes identified abovecould induce the growth of CD8+ CTLs in vitro,peripheral blood lymphocytes from two HLA-A2+ healthy individuals were stimulated in vitro inthe presence of a pool of T2 cells individuallyloaded with all the HLA-A2-binding mammaglo-bin-A-derived peptides (Table 2). After fourstimulations, the specificity of both CD8+ CTLlines was evaluated against T2 cells individuallyloaded with the peptides. As shown in Figure 2,both CD8+ CTL lines showed a similar profile ofpolyclonal cytotoxic activity against peptides
Mam-A2.1, Mam-A2.2, Mam-A2.3, and Mam-A2.4 but not against peptides Mam-A2.5, Mam-A2.6, and Mam-A2.7. These results indicate thatCD8+ CTL lines generated in vitro show a com-parable reactivity profile as the one generated bybreast cancer patients in vivo. Interestingly, noneof the CD8+ CTL lines shown any reactivity tothe Mam-A2.7 peptide despite the presence ofMam-A2.7-reactive CD8+ CTL precursors inHLA-A2+ human individuals (Table 3).
Lysis of breast cancer cells by HLA-A2-restrictedCD8+ CTLs reactive against mammaglobin-A-derived epitopes
To determine whether the HLA-A2-restrictedCD8+ CTL lines reactive against the computer-predicted mammaglobin-A-derived epitopes wereable to recognize endogenously produced mam-maglobin-A-derived peptides, we determined thereactivity of the CD8+ CTL lines against a panelof eight different breast cancer cell lines (Table 1).As shown in Figure 3, both CD8+ CTL linesshowed a similar pattern of reactivity against thepanel of breast cancer cell lines. Both CD8+CTL lines showed a significant cytotoxic activityagainst one of three HLA-A2+/mammaglobin-A+ breast cancer cell lines (UACC-812) but not
Figure 2. Immunodominance of the in vitro CD8+ CTL immune response against mammaglobin-A-derived epitopes. Two CD8+
CTL lines were generated from two HLA-A2-positive healthy individuals by in vitro stimulations with pooled T2 cells individually
loaded with the HLA-A2-binding mammaglobin-A-derived peptides (Mam-A2.1-7). Then, the immunodominance of the CD8+ CTL
lines was evaluated against T2 cells individually loaded with the candidate epitopes by means of a standard 51Cr release assay. T2 cells
loaded with the GILGFVFTL influenza-derived epitope and ‘empty’ T2 cells (no peptide) were used as negative controls. The assay
was performed at E:T ratios of 50, 25, and 12.5. Only the results obtained at the E:T ratio of 50 are shown. Results are presented as the
mean ± standard deviation of quadruplicate cultures.
CD8+ CTLs to mammaglobin-A in breast cancer patients 35

against HLA-A2+/mammaglobin-A) (MCF-7,MDA-MB-231) and HLA-A2)/mammaglobin-A+ (DU-4475, MDA-MB-361, MDA-MB-415,ZR-75-1) breast cancer cell lines. No cytotoxicactivity was observed against the natural killercell-sensitive K562 cell line (data not shown). It isinteresting to note that both CD8+ CTL lines didnot show any cytotoxic activity against twoHLA-A2+/mammaglobin-A+ breast cancer celllines (AU-565, HBL-100) previously demon-strated to be recognized by HLA-A2-restrictedCD8+ CTLs generated in vitro against mamma-globin-A-transfected dendritic cells [37]. Theseresults suggest that the Mam-A2.1, Mam-A2.2,Mam-A2.3, and Mam-A2.4 epitopes are not ex-pressed by the AU-565 and HBL-100 breastcancer cell lines.
The epitope specificity and MHC-restriction ofthe cytotoxic activity of the CD8+ CTL line No. 1was confirmed in a cold target inhibition assay. Asshown in Figure 4, lysis of the UACC-812 breastcancer cell line was blocked by the addition of T2cells loaded with the Mam-A2.1 peptide, whereasT2 cells loaded with the Mam-A.2.2, Mam-A2.3,and Mam-A2.4 peptides showed no effect on thecytotoxic activity against this breast cancer cellline. These results show that the UACC-812 breast
Figure 3. Recognition of breast cancer cells by CD8+ CTLs generated in vitro against mammaglobin-A-derived epitopes. Two CD8+
CTL lines were generated from two HLA-A2-positive healthy individuals by in vitro stimulations with pooledT2 cells individually
loaded with the HLA-A2-binding mammaglobin-A-derived peptides (Mam-A2.1-7). The mammaglobin-A-specificity and HLA-A2-
restriction of the CD8+ CTL lines was then evaluated against HLA-A2+/mammaglobin-A+ (UACC-812, HBL-100, AU-565), HLA-
A2+/mammaglobin-A) (MCF-7, MDA-MB-231), and HLA-A2)/mammaglobin-A+ (DU-4475, MDA-MB-361, MDA-MB-415,
ZR-75-1) breast cancer cell lines by means of a standard 51Cr release assay. The assay was performed at E:T ratios of 50, 25, and 12.5.
Only the results obtained at the E:T ratio of 50 are shown. Results are presented as the mean ± standard deviation of quadruplicate
cultures.
Figure 4. Recognition of the Mam-A2.1 epitope on HLA-
A2+/mammaglobin-A+ breast cancer cells by CD8+ CTLs
generated in vitro against mammaglobin-A-derived epitopes.
Two CD8+ CTL lines were generated from two HLA-A2-
positive healthy individuals by in vitro stimulations with
pooled T2 cells individually loaded with the HLA-A2-
binding mammaglobin-A-derived peptides (Mam-A2.1-7).
Then, the epitope-specificity of the CD8+ CTL line No. 1
was evaluated against the HLA-A2+/mammaglobin-A+
UACC-812 breast cancer cell line by means of a standard51Cr release assay in the presence of 20-fold excess of
unlabeled T2 cells individually loaded with the Mam-A2.1,
Mam-A2.2, Mam-A2.3, and Mam-A2.4 epitopes. T2 cells
loaded with the GILGFVFTL influenza-derived epitope and
‘empty’ T2 cells (no peptide) were used as negative con-
trols. The assay was performed at an E:T ratio of 50.
Results are presented as the mean ± standard deviation of
quadruplicate cultures.
A Jaramillo et al.36

cancer cell line only expresses epitope Mam-A2.1.Overall, these results indicate the heterogenicnature of breast cancer antigenicity and under-score the importance of a polyclonal CD8+ CTLresponse in regards to the design of immunother-apy for breast cancer.
To confirm the presentation of the Mam-A2.1peptide by the HLA-A2 molecule on the UACC-812 breast cancer cell line, the cytotoxic activityof the CD8+ CTL line No. 1 was also deter-mined in the presence of a mAb directed againstthe HLA-A2 molecule. As shown in Figure 5,addition of the BB7.2 anti-HLA-A2 mAb inhib-ited the lysis of the UACC-812 breast cancer cellline whereas addition of the MOPC-11 controlmAb showed no effect on the cytotoxic activityagainst this breast cancer cell line. These resultsfurther confirmed the HLA-A2-restriction of theMam-A2.1 peptide recognition by this CD8+CTL line.
Induction of a CD8+ CTLs reactivity against breastcancer cells by vaccination with DNA encoding theMam-A2.1 epitope and the HLA-A2 molecule
To determine the potential use of this epitope inthe prevention for breast cancer, we designed a
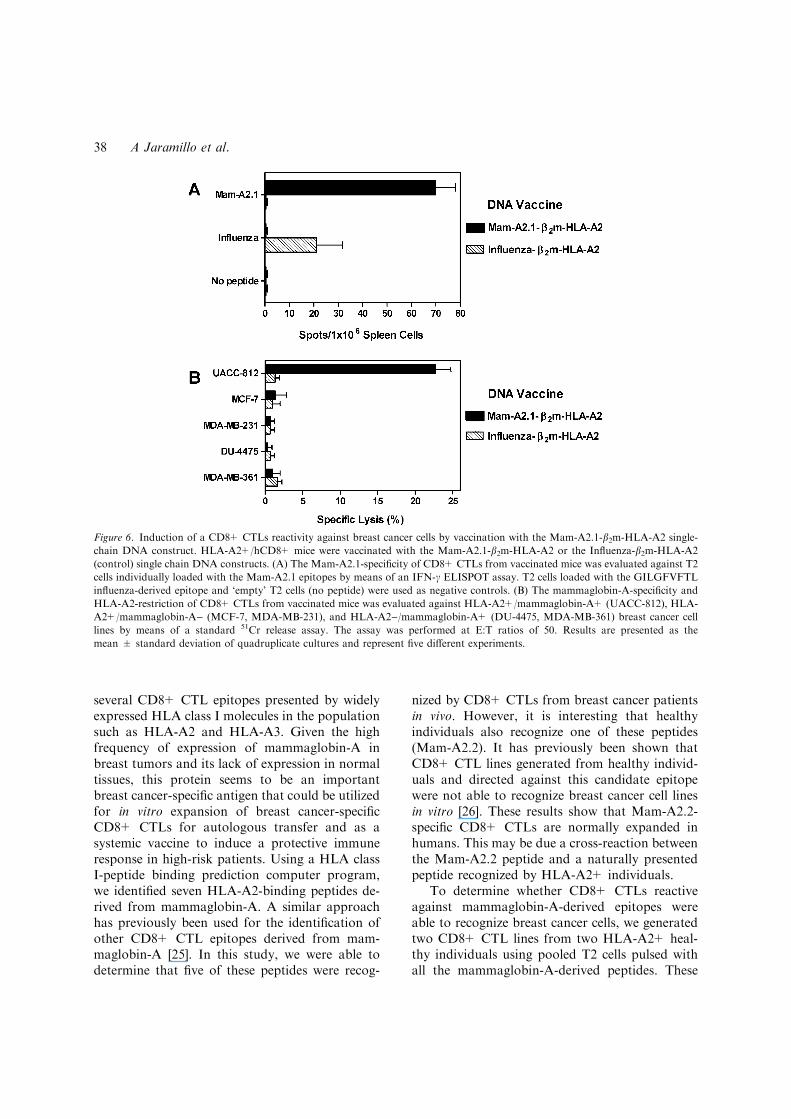
vaccination protocol using a DNA construct thatencoded the Mam-A2.1 epitope presented in thecontext of the HLA-A2 molecule (Mam-A2.1-b2m-HLA-A2). As shown in Figure 6A, vaccination ofHLA-A2+/hCD8+ mice with the Mam-A2.1-b2m-HLA-A2 construct induced the specific expansionof Mam-A2.1-reactive CD8+ CTLs. On the con-trary, as also shown in Figure 6A, vaccination ofHLA-A2+/hCD8+ mice with the control Influ-enza-b2m-HLA-A2 construct induce the specificexpansion of Influenza epitope-reactive CD8+CTLs. These results indicate that this protocol ofDNA vaccination induced the expansion of epi-tope-specific CD8+ CTLs.
To determine whether the Mam-A2.1-reactiveCD8+ CTLs generated in vivo by means of DNAvaccination were able to kill breast cancer cellsexpressing this epitope, we determined the reac-tivity of CD8+ CTLs present in the spleen ofvaccinated HLA-A2+/hCD8+ mice against a pa-nel of five different breast cancer cell lines (Ta-ble 1). As shown in Figure 6B, CD8+ CTLs fromHLA-A2+/hCD8+ mice vaccinated with theMam-A2.1-b2m-HLA-A2 construct showed a sig-nificant cytotoxic activity against the HLA-A2+/mammaglobin-A+ UAC-812 breast cancer cellline but not against HLA-A2+/mammaglobin-A)(MCF-7, MDA-MB-231) and HLA-A2)/mam-maglobin-A+ (DU-4475, MDA-MB-361) breastcancer cell lines. As also shown in Figure 6B,CD8+ CTLs from HLA-A2+/hCD8+ mice vac-cinated with the Influenza-b2m-HLA-A2 constructdid not showed a significant cytotoxic activityagainst the HLA-A2+/mammaglobin-A+ UAC-812 breast cancer cell line. These results indicatethe potential use of DNA vaccination with mam-maglobin-A-derived epitopes as a preventivetherapeutic alternative in high-risk patients forprimary breast cancer development.
Discussion
It is well established that CD8+ CTLs are a crit-ical component of the immune response to tumors.Thus, the identification of CD8+ CTL epitopesderived from breast cancer-specific antigens iscrucial for the successful development of breastcancer immunotherapies. A clinically feasiblevaccine for breast cancer would need to have
Figure 5. Inhibition of the cytotoxic activity of the mamma-
globin-A-reactive CD8+ CTL line by anti-HLA-A2 antibodies.
Two CD8+ CTL lines were generated from two HLA-A2-po-
sitive healthy individuals by in vitro stimulations with pooled T2
cells individually loaded with the HLA-A2-binding mamma-
globin-A-derived peptides (Mam-A2.1-7). Then, the HLA-A2-
restriction of the CD8+ CTL line No. 1 was evaluated against
the HLA-A2+/mammaglobin-A+ UACC-812 breast cancer
cell line in the presence of the BB7.2 (anti-HLA-A2) or the
MOPC-11 (isotype control) mAbs. The assay was performed at
an E:T ratio of 50. Results are presented as the mean ± stan-
dard deviation of quadruplicate cultures.
CD8+ CTLs to mammaglobin-A in breast cancer patients 37
several CD8+ CTL epitopes presented by widelyexpressed HLA class I molecules in the populationsuch as HLA-A2 and HLA-A3. Given the highfrequency of expression of mammaglobin-A inbreast tumors and its lack of expression in normaltissues, this protein seems to be an importantbreast cancer-specific antigen that could be utilizedfor in vitro expansion of breast cancer-specificCD8+ CTLs for autologous transfer and as asystemic vaccine to induce a protective immuneresponse in high-risk patients. Using a HLA classI-peptide binding prediction computer program,we identified seven HLA-A2-binding peptides de-rived from mammaglobin-A. A similar approachhas previously been used for the identification ofother CD8+ CTL epitopes derived from mam-maglobin-A [25]. In this study, we were able todetermine that five of these peptides were recog-
nized by CD8+ CTLs from breast cancer patientsin vivo. However, it is interesting that healthyindividuals also recognize one of these peptides(Mam-A2.2). It has previously been shown thatCD8+ CTL lines generated from healthy individ-uals and directed against this candidate epitopewere not able to recognize breast cancer cell linesin vitro [26]. These results show that Mam-A2.2-specific CD8+ CTLs are normally expanded inhumans. This may be due a cross-reaction betweenthe Mam-A2.2 peptide and a naturally presentedpeptide recognized by HLA-A2+ individuals.
To determine whether CD8+ CTLs reactiveagainst mammaglobin-A-derived epitopes wereable to recognize breast cancer cells, we generatedtwo CD8+ CTL lines from two HLA-A2+ heal-thy individuals using pooled T2 cells pulsed withall the mammaglobin-A-derived peptides. These
Figure 6. Induction of a CD8+ CTLs reactivity against breast cancer cells by vaccination with the Mam-A2.1-b2m-HLA-A2 single-
chain DNA construct. HLA-A2+/hCD8+ mice were vaccinated with the Mam-A2.1-b2m-HLA-A2 or the Influenza-b2m-HLA-A2
(control) single chain DNA constructs. (A) The Mam-A2.1-specificity of CD8+ CTLs from vaccinated mice was evaluated against T2
cells individually loaded with the Mam-A2.1 epitopes by means of an IFN-c ELISPOT assay. T2 cells loaded with the GILGFVFTL
influenza-derived epitope and ‘empty’ T2 cells (no peptide) were used as negative controls. (B) The mammaglobin-A-specificity and
HLA-A2-restriction of CD8+ CTLs from vaccinated mice was evaluated against HLA-A2+/mammaglobin-A+ (UACC-812), HLA-
A2+/mammaglobin-A) (MCF-7, MDA-MB-231), and HLA-A2)/mammaglobin-A+ (DU-4475, MDA-MB-361) breast cancer cell
lines by means of a standard 51Cr release assay. The assay was performed at E:T ratios of 50. Results are presented as the
mean ± standard deviation of quadruplicate cultures and represent five different experiments.
A Jaramillo et al.38

CD8+ CTL lines recognized the Mam-A2.1,Mam-A2.2, Mam-A2.3, and Mam-A2.4 peptidesbut not the Mam-A2.7 peptide. This may be due tothe significantly lower predicted affinity of peptideMam-A2.7 for the HLA-A2 molecule as comparedto the other peptides recognized in vivo. Theresulting CD8+ CTL lines displayed a significantcytotoxic activity against one of three HLA-A2+/mammaglobin-A+ breast cancer cell lines andfurther analysis demonstrated that this breastcancer cell line expresses only one epitope (Mam-A2.1). The lack of reactivity of the CD8+ CTLlines against two other HLA-A2+/mammaglobin-A+ breast cancer cell lines suggest that not all theprimary breast tumors and breast cancer cell linesexpress all the epitopes that can possibly be gen-erated from mammaglobin-A. In this regard, thefact that seven of the eight (87.5%) breast cancerpatients included in this study showed differentepitope reactivity profiles indicate the heteroge-nous nature of breast tumors. These epitopes maybe differentially expressed by breast tumors in vivodue to several reasons including differential en-zyme cleavage of the mammaglobin-A protein inthe proteosome [38] and/or selectively loss themembrane expression of a given HLA class I allele[25]. This also indicates that studies with a singlebreast cancer cell line in vitro can not reproducethe complex antigenic nature of breast tumors invivo. These results underscore the importance ofinducing a multi-specific CD8+ CTL immune re-sponse for the effective development of preventiveimmune therapies for breast cancer.
To determine the feasibility of vaccination withmammaglobin-A DNA to induce a breast cancer-specific CD8+ CTL reactivity, we use a novelapproach with a single-chain DNA construct thathas ben shown to induce potent peptide-specificCD8+ CTL and B cell responses [31, 32]. The datapresented herein clearly demonstrate that DNAvaccination with mammaglobin-A-derived epi-topes could represent a novel therapeutic alterna-tive for breast cancer and, more importantly, as apreventive treatment of patients at high-risk forprimary disease development.
In summary, we have identified four HLA-A2-restricted CD8+ CTL epitopes from a breastcancer-specific antigen, mammaglobin-A. Inaddition, we show herein that CD8+ CTLs gen-erated in vitro by multiple stimulations and in vivoby DNA vaccination against computer-predicted
epitopes have the ability to recognize breast cancercells. Current studies in our laboratory are aimedtoward identifying patients at high risk for pri-mary disease development or recurrence aftersurgical removal of the tumor based on theirreactivity profile against mammaglobin-A epi-topes. These results further show that identifica-tion of mammaglobin-A-derived epitopes couldprovide a novel approach for designing newimmunotherapy protocols for the treatment andprevention of breast cancer.
Acknowledgement
This work was supported by a grant from TheSusan G. Komen Breast Cancer Foundation.
References
1. Finke JH, Rayman P, Alexander J, Edinger M, Tubbs RR,
Connelly R, Pontes E, Bukowski R: Characterization of the
cytolytic activity of CD4+ and CD8+ tumor-infiltrating
lymphocytes in human renal cell carcinoma. Cancer Res 50:
2363–2370, 1990
2. Peoples GE, Schoof DD, Andrews JV, Goedegebuure PS,
Eberlein TJ: T cell recognition of ovarian cancer. Surgery
114: 227–234, 1993
3. Topalian S, Solomon D, Rosenberg SA: Tumor-specific
cytolysis by lymphocytes infiltrating human melanomas. J
Immunol 142: 3714–3725, 1989
4. Linehan DC, Goedegebuure PS, Peoples GE, Rogers SO,
Eberlein TJ: Tumor-specific and HLA-A2-restricted cytol-
ysis by tumor associated lymphocytes in human metastatic
breast cancer. J Immunol 155: 4486–4491, 1995
5. Peoples GE, Goedegebuure PS, Smith R, Linehan DC,
Yoshino I, Eberlein TJ: Breast and ovarian cancer-specific
cytotoxic T lymphocytes recognize the same HER2/neu
derived peptide. Proc Natl Acad Sci USA 92: 432–436, 1995
6. Boon T, van der Bruggen P: Human tumor antigens
recognized by T lymphocytes. J Exp Med 183: 725–729,
1996
7. Boon T, Coulie PG, Van den Eynde B: Tumor antigens
recognized by T cells. Immunol Today 18: 267–268, 1997
8. Apostolopoulos V, Sandrin MS, McKenzie IF: Carbohy-
drate/peptide mimics: effect on MUC1 cancer immunother-
apy. J Mol Med 77: 427–436, 1999
9. Finn OJ, Jerome KR, Henderson RA, Pecher G, Domenech
N, Magarian-Blander J, Barratt-Boyes SM: MUC1 epithe-
lial tumor-mucin-based immunity and cancer vaccines.
Immunol Rev 145: 61–89, 1995
10. Brossart P, Heinrich KS, Stuhler G, Behnke L, Reichardt
VL, Stevanovic S, Muhm A, Rammensee H-G, Kanz L,
Brugger W: Identification of HLA-A2-restricted T cell
epitopes derived from the MUC1 tumor antigen for
CD8+ CTLs to mammaglobin-A in breast cancer patients 39

broadly applicable vaccine therapies. Blood 93: 4309–
4317, 1999
11. Domenech N, Henderson RA, Finn OJ: Identification of an
HLA-A11-restricted epitope from the tandem repeat
domain of the epithelial tumor antigen mucin. J Immunol
155: 4766–4774, 1995
12. Brossart P, Wirths S, Stuhler G, Reichardt VL, Kanz L,
Brugger W: Induction of cytotoxic T-lymphocyte responses
in vivo after vaccinations with peptide-pulsed dendritic
cells. Blood 96: 3102–3108, 2002
13. Slamon DJ, Godolphin W, Jones LA, Holt JA, Wong SG,
Keith DE, Levin WJ, Stuart SG, Udove J, Ulrich A, Press
MF: Studies of the Her-2/neu protooncogene in human
breast and ovarian cancer. Science 244: 707–712, 1989
14. Lohrisch C, Piccart M: HER2/neu as a predictive factor in
breast cancer. Clin Breast Cancer 2: 129–135, 2001
15. Graham RA, Burchell JM, Taylor-Papadimitriou J: The
polymorphic epithelial mucin: potential as an immunogen
for a cancer vaccine. Cancer Immunol Immunother 42: 71–
80, 1996
16. Apostolopoulos V, Pietersz GA, McKenzie IF: MUC1 and
breast cancer. Curr Opin Mol Ther 1: 98–103, 1999
17. Rahn JJ, Dabbagh L, Pasdar M, Hugh JC: The impor-
tance of MUC1 cellular localization in patients with
breast carcinoma: an immunohistologic study of 71
patients and review of the literature. Cancer 91: 1973–
1982, 2001
18. Watson MA, Fleming TP: Isolation of differentially
expressed sequence tags from human breast cancer. Cancer
Res 54: 4598–4602, 1994
19. Watson MA, Fleming TP: Mammaglobin, a mammary
specific member of the uteroglobin gene family, is overex-
pressed in human breast cancer. Cancer Res 56: 860–865,
1996
20. Watson MA, Darrow C, Zimonjic DB, Popescu NC,
Fleming TP: Structure and transcriptional regulation of
the human mammaglobin gene, a breast cancer associated
member of the uteroglobin gene family localized to chro-
mosome 11q13. Oncogene 16: 817–824, 1998
21. Fleming TP, Watson MA: Mammaglobin, a breast-specific
gene, and its utility as a marker for breast cancer. Ann NY
Acad Sci 923: 78–89, 2000
22. Marchetti A, Buttitta F, Bertacca G, Zavaglia K, Bevilac-
qua G, Angelucci D, Viacava P, Naccarato A, Bonadio A,
Barassi F, Felicioni L, Salvatore S, Mucilli F: mRNA
markers of breast cancer nodal metastases: comparison
between mammaglobin and carcinoembryonic antigen in
248 patients. J Pathol 195: 186–190, 2001
23. Grunewald K, Haun M, Urbanek M, Fiegel M, Muller-
Holzner E, Gunsilius E, Dunser M, Marth C, Castle G:
Mammaglobin gene expression: a superior marker of breast
cancer cells in peripheral blood in comparison to epidermal
growth factor receptor and cytokeratin-19. Lab Invest 80:
1071–1077, 2000
24. Watson MA, Dintzis S, Darrow CM, Voss LE, DiPersio J,
Jensen R, Fleming TP: Mammaglobin expression in
primary, metastatic and occult breast cancer. Cancer Res
59: 3028–3031, 1999
25. Jaramillo A, Majumder K, Manna PP, Fleming TP,
Doherty G, Dipersio JF, Mohanakumar T: Identification
of HLA-A3-restricted CD8+ T cell epitopes derived from
mammaglobin-A, a tumor-associated antigen of human
breast cancer. Int J Cancer 102: 499–506, 2002
26. Tanaka Y, Amos KD, Fleming TP, Eberlein TJ, Goed-
egebuure PS: Mammaglobin-A is a tumor-associated anti-
gen in human breast carcinoma. Surgery 133: 74–80,
2003
27. Man S, Newberg MH, Crotzer VL, Luckey CJ, Williams
NS, Chen Y, Huczko EL, Ridge JP, Engelhard VH:
Definition of a human T cell epitope from influenza A
non-structural protein 1 using HLA-A2.1 transgenic mice.
Int Immunol 7: 597–605, 1995
28. Lustgarten J, Theobald M, Labadie C, LaFace D, Peterson
P, Disis ML, Cheever MA, Sherman LA: Identification of
Her-2/Neu CTL epitopes using double transgenic mice
expressing HLA-A2.1 and human CD.8. Hum Immunol 52:
109–118, 1997
29. Parker KC, Bednarek MA, Coligan JE: Scheme for ranking
potential HLA-A2 binding peptides based on independent
binding of individual peptide side-chains. J Immunol 152:
163–175, 1994
30. Bednarek MA, Sauma SY, Gammon MC, Porter G,
Tamhankar S, Williamson AR, Zweerink HJ: The mini-
mum peptide epitope from the influenza virus matrix
protein. Extra and intracellular loading of HLA-A2. J
Immunol 147: 4047–4053, 1991
31. Yu YYL, Netuschil N, Lybarger L, Connolly JM, Hansen
TH: Single-chain trimer of MHC class I molecules form
stable structures that potently stimulate antigen-specific T
cells and B cells. J Immunol 168: 3145–3149, 2002
32. Lybarger L, Yu YY, Miley MJ, Fremont DH, Myers N,
Primeau T, Truscott SM, Connolly JM, Hansen TH:
Enhanced immune presentation of a single-chain major
histocompatibility complex class I molecule engineered to
optimize linkage of a C-terminally extended peptide. J Biol
Chem 278: 27105–27111, 2003
33. Kuhns JJ, Batalia MA, Tan S, Collins EJ: Poor binding of a
HER-2/neu epitope (GP2) to HLA-A2.1 is due to a lack of
interactions with the center of the peptide. J Biol Chem 274:
36422–36427, 1999
34. Tanaka Y, Amos KD, Joo HG, Eberlein TJ, Goedegeburre
PS: Modification of the HER-2/neu-derived tumor antigen
GP2 improves induction of GP2-reactive cytotoxic T
lymphocytes. Int J Cancer 94: 540–544, 2001
35. Bullock TN, Mullins DW, Colella TA, Engelhard VH:
Manipulation of avidity to improve effectiveness of adop-
tively transferred CD8(+) T cells for melanoma immuno-
therapy in human MHC class I-transgenic mice. J Immunol
167: 5824–5831, 2001
36. Nijman HW, Houbiers JG, van der Burg SH, Vierboom
MP, Kenemans P, Kast WM, Melief CJ: Characterization
of cytotoxic T lymphocyte epitopes of a self-protein, p 53,
and a non-self-protein, influenza matrix: relationship
between major histocompatibility complex peptide binding
affinity and immune responsiveness to peptides. J Immun-
other 14: 121–126, 1993
37. Manna PP, Jaramillo A, Majumder K, Campbell LG,
Fleming TP, Dietz JR, DiPersio JF, Mohanakumar T:
Generation of CD8+ cytotoxic T lymphocytes against
breast cancer cells by stimulation with mammaglobin-A-
pulsed dendritic cells. Breast Cancer Res Treat 79: 133–136,
2003
A Jaramillo et al.40

38. Luckey CJ, Marto JA, Partridge M, Hall E, White FM,
Lippolis JD, Shabanowitz J, Hunt DF, Engelhard VH:
Differences in the expression of human class I MHC alleles
and their associated peptides in the presence of proteosome
inhibitors. J Immunol 167: 1212–1221, 2001
Address for offprints and correspondence: Dr. Andres Jaramillo,
Department of Surgery, Washington University School of
Medicine, Box 8109-3328, CSRB, 660 South Euclid Avenue, St.
Louis, MO 63110, USA; Tel.: +1-314-362-8458; Fax: +1-314-
747-1560; E-mail: [email protected]
CD8+ CTLs to mammaglobin-A in breast cancer patients 41



















