
Differential Expression of Activation Epitopes of β1 Integrins in Psoriasis and Normal Skin

A
awpaoP
K
1
dmtrhwd
ta
Mf
(
0d
Vaccine 25 (2007) 5330–5342
Immunological validation of the EpitOptimizer program forstreamlined design of heteroclitic epitopes
Colin S.B. Houghton a, Manuel E. Engelhorn a, Cailan Liu a, Da Song b, Polly Gregor a,Phillip O. Livingston a, Francesca Orlandi a, Jedd D. Wolchok a, James McCracken c,
Alan N. Houghton a, Jose A. Guevara-Patino a,c,∗a The Swim Across America Laboratory, Memorial Sloan-Kettering Cancer Center, New York, NY 10021, United States
b Research Computing Department, Memorial Sloan-Kettering Cancer Center, New York, NY 10021, United Statesc Laboratory of Tumor Immunology and Cancer Vaccine Development, The University of Chicago, Chicago, IL 60637, United States
Received 26 March 2007; received in revised form 4 May 2007; accepted 9 May 2007Available online 4 June 2007
bstract
One strategy to generate T-cell responses to tumors is to alter subdominant epitopes through substitution of amino acids that are optimalnchors for specific MHC molecules, termed heteroclitic epitopes. This approach is manually error-prone and time-consuming. In here,e describe a computer-based algorithm (EpitOptimizer) for the streamlined design of heteroclitic epitopes. Analysis of two cancer-relatedroteins showed that EpitOptimizer-generated peptides have enhanced MHC-I binding compared with their wild-type counterparts; and were
ble to induce stronger CD8+ T-cell responses against their native epitope. These data demonstrate that this approach can serve as the basisf epitope-engineering against cancer and intracellular pathogens.ublished by Elsevier Ltd.eywords: T Cells; Antigens/peptides/epitopes; Tumor immunity; Vaccination
n[d[con
. Introduction
Development of cancer vaccines faces the fundamentalifficulty that cancer arises from the host’s tissue. Most com-only, cancer antigens are products of unaltered self-genes
hat are typically incapable of inducing primary immuneesponses. These self-proteins may contain epitopes with
igh affinity for self MHC-I molecules, but immature T cellsith high affinity receptors for these self-peptides usuallyie during thymic development, and those that escape thymicAbbreviations: MHC-I, major histocompatibility complex I; TYRP1,yrosinase-related protein 1; PSMA, human prostate-specific membranentigen; PSMAee, epitope enriched PSMA; APCs, antigen presenting cells∗ Corresponding author at: Department of Surgery, 5841 S. Maryland Ave.C 7114, Chicago, IL 60637-1470, United States. Tel.: +1 773 702 5438;
ax: +1 773 834 8140.E-mail address: [email protected]
J.A. Guevara-Patino).
snoirrmCp
p
264-410X/$ – see front matter. Published by Elsevier Ltd.oi:10.1016/j.vaccine.2007.05.008
egative selection are anergized or deleted in the periphery1]. However, T cells with receptors with low or interme-iate affinity for self-peptide/MHC survive thymic selection1]. These subliminal peptide epitopes are unable to elicit T-ell activation and differentiation. Several groups, includingurs, have studied strategies for generating effective immu-ity against self-antigens expressed by cancer. One suchtrategy is to design heteroclitic peptides that trigger recog-ition of subliminal epitopes presented by MHC-I moleculen the surface of tumor cells [1–7]. Using this approach, its possible to alter the magnitude and specificity of T-cellesponses by selectively changing (optimizing) amino acidesidues involved in binding to MHC-I molecules. These opti-ized epitope variants can prime a subset of naıve cognate
D8+ T cells to cross-react against corresponding wild-typeeptide/MHC complexes.Computational algorithms to predict MHC binding ofeptides were pioneered by the groups of Parker and Ram-

Vaccin
mpasaTcMdabTcmoaahtg
fhae[bpTaedespoht
iagtAfita
2
2
C
ASmaaA
2
wcccFawmava
2
amcSldhHt
2
tceaRwbTfliAc
C.S.B. Houghton et al. /
ensee and have been routinely used to predict tumor-derivedeptides that bind MHC-I and II molecules [8–10]. Theselgorithms are based on the isolation and amino acidequences of naturally processed and presented MHC-I lig-nds and the MHC binding properties of synthetic peptides.he predictive capability of these computational algorithmsomes from the identification of primary and secondaryHC-binding anchor residues. The binding score is pre-
icted using quantitative matrices in which every aminocid in a peptide is assigned a coefficient. The overallinding value is calculated from amino acid coefficients.hese algorithms have significantly facilitated the identifi-ation of tumor-derived peptides that bind MHC-I and IIolecules, increasing the epitope arsenal of vaccine devel-
pers. Although EpitOptimizer builds on and extends theselgorithms, the innovative aspect of this computer-basedpproach is that, it is designed to create multiple potentialeteroclitic peptides with enhanced MHC-I binding alonghe entire length of polypeptides to induce greater immuno-enicity.
Current efforts in cancer vaccine development have mainlyocused on inducing CD8+ T-cell responses using native oreteroclitic epitopes [11–19]. Recently, we demonstrated thatnon-immunogenic mouse self-protein is converted into an
ffective immunogen by an epitope optimization approach7]. We manually designed multiple epitopes with improvedinding to MHC-I molecules in mouse tyrosinase-relatedrotein 1 (TYRP1) [30] (using site-directed mutagenesis).his approach combined MHC-I epitope optimization (cre-ting heteroclitic epitopes), epitope enrichment (multiplepitopes), and enhancement of antigen processing throughestabilization by mutations. DNA immunization with thengineered TYRP1 vaccine led to rejection of melanoma andignificantly increased survival. The main caveat for theserevious studies was the time-consuming and tedious analysisf the full-length protein and the laborious manual design ofeteroclitic epitopes. EpitOptimizer was created to overcomehese obstacles.
Here, we present EpitOptimizer and describe initial val-dation studies using a mouse melanoma differentiationntigen TYRP1, human prostate-specific membrane anti-en (PSMA) [31] and five other self-molecules. Usinghis approach, we have discovered three novel HLA-*0201-restricted human PSMA epitopes that are candidates
or further evaluation in clinical studies. These datallustrate the capabilities of EpitOptimizer for selec-ive epitope-engineering of poorly and non-immunogenicntigens.
. Materials and methods
.1. Mice
Female C57BL/6 mice were purchased from the Nationalancer Institute breeding program (Frederick, MD). HLA-
ClCv
e 25 (2007) 5330–5342 5331
*0201/Kb transgenic mice were obtained from Dr. Lindaherman, The Scripps Research Institute (La Jolla, CA). Allice entered the study between 8 and 10 weeks of age. Care of
ll mice used in experiments was under an approved protocolnd in accordance with the guidelines of the Institutionalnimal Care and Use Committee of MSKCC.
.2. Peptides and peptide vaccination
The peptides used in this study were synthesized crudeith >80% purity by GeneMed Synthesis, Inc. (San Fran-
isco, CA). They were resuspended in 100% DMSO at aoncentration of 40 mg/ml, then diluted with PBS (finaloncentration 2 mg/ml) and frozen in aliquots at −20 ◦C.or vaccination, peptides were used at 1:1 with Titermaxdjuvant (Sigma–Aldrich, St. Louis, MO). Recipient miceere vaccinated in the footpad with 20 �g peptide:adjuvantixture. Five days after vaccination, mice were euthanized
nd draining inguinal lymph nodes harvested to assess exivo T-cell responses against wild-type peptides by ELIspotssay.
.3. Antibodies and cell lines
Cell surface expression of Db and HLA-A*0201 wasssessed by staining with FITC-conjugated mAb anti-Db
Ab 28-14-8 or mAb A2.1-specific mAb BB7.2 or a FITC-onjugated isotype control Ab (BD Biosciences Pharmingen,an Jose, CA). The EL4 cell line is a C57BL/6 mouse
ymphoma. RMA-S is a mouse transporter antigen peptide-eficient (TAP2) cell line [32]. K562/HLA-A*0201 is auman leukemia cell line transfected with HLA-A*0201 [37].uman T2 cells are deficient in the transporter antigen pep-
ide molecules (TAP1 and 2) [33].
.4. MHC stabilization assay
To determine the relative ability of heteroclitic pep-ides to bind to MHC class I molecules, synthetic peptidesorresponding to the wild-type and optimized predicted het-roclitic variants were tested by MHC class I stabilizationssay [32]. Briefly, ∼2 × 105 cells TAP2-deficient Kb/Db
MA-S cells were incubated overnight at 23 ◦C. Peptidesere added at 1 �g/ml final concentration, followed by incu-ation for 2 h at 37 ◦C. TAP2-deficient human HLA-A*02012 cells [32] were incubated with and without peptides
or 16 h at 37 ◦C. Cells were then washed, and surfaceevels of Db and HLA-A*0201 were assessed by stain-ng with FITC-conjugated, mAb-Db mAb 28-14-8 or mAb2.1-specific mAb BB7.2 or an FITC-conjugated isotype
ontrol Ab (BD Biosciences Pharmingen, San Jose, CA).
ells were fixed at 4 ◦C in 4% paraformaldehyde and ana-yzed on a FACSCalibur (Becton Dickinson, Mountain View,A) using CellQuest software. Results are expressed as MFIalues.

5 Vaccin
2
tJoctiu
LVGNGRSSEKSMII
2
fbaapldTa
2
ccabTwswiwbRaaET
2
Ba
3
3
tgaMoD(oiNIptYrf(av(oatea
3
tgmlmHiaidn
332 C.S.B. Houghton et al. /
.5. Plasmid constructs and site-directed mutagenesis
The human PSMA cDNA was mutated at 14 different posi-ions using reiterations of the Quickchange (Stratagene, Laolla, CA) protocol. Bulk plasmid miniprep from one roundf mutagenesis was used as template for the next. Candidatelones for the fully altered construct were sequenced afterhe last round. The confirmed insert was finally subclonednto pCDNA3 plasmid. The following oligonucleotides weresed site directed mutagenesis.
4Y GCCGAGATGTGGAATTACCTTCACGAAACCGA27L TGCGCTGGGGCGCTGCTGCTGGCGGGTGGCT36L GGCTTCTTTCTCCTCCTCTTCCTCTTCGGGTG76L AAGAAGTTCTTATATCTTTTTACACAGATACC150V AACCACCTCCTCCAGTATATGAAAATGTTTC181L ATGTTAACTATGCACTAACTGAAGACTTCTT241L GCTCCTGGGGTGAAGCTCTATCCAGATGGTTG312V GAAAAAATGGGTGGCGTAGCACCACCAGATAG436V TACTGAGTGGGCAGTGGAGAATTCAAGACTC500V GAAAGTTGGACTAAAGTAAGTCCTTCCCCAGA628L AAGACATACAGTGTACTATTTGATTCACTTTT664L ATAGTATTAAGAATGCTGAATGATCAACTCA
708L GAGTCATTCCCAGGACTTTATGATGCTCTGT732L GAAGTGAAGAGACAGCTTTATGTTGCAGCCT
.6. Gene gun immunization
Immunization using DNA-coated gold particles was per-ormed as previously described, using a gene gun providedy PowderMed, Inc. (Oxford, UK) [36]. Four injectionst 400 pounds/in.2 were delivered to each mouse, one perbdominal quadrant, for a total of 4 �g plasmid DNAer mouse weekly for 4 weeks. Five days after theast vaccination, mice were euthanized and spleens orraining lymph nodes were harvested to assess ex vivo-cell responses against wild-type peptides in ELIspotssays.
.7. IFN-γ ELIspot assay
IP-Multiscreen plates (Millipore, Burlington, MA) wereoated with 100 �l anti-mouse IFN-� antibody (10 �g/ml;lone AN18, MabTech, Sweden) in PBS, incubated overnightt 4 ◦C, washed with PBS to remove unbound antibody, andlocked with RPMI/FCS for 2 h at 37 ◦C. Purified CD8+
cells were plated at a concentration of (1–2) × 105 perell. For antigen presentation, 5 × 104 irradiated antigen pre-
enting cells per well were pulsed with 10 �g/ml peptide,hich was added to a final volume of 100 �l/well. After
ncubation for 20 h at 37 ◦C, plates were extensively washedith PBS/0.05% Tween and were incubated with 100 �l/welliotinylated-antibody against mouse IFN-� (2 �g/ml; clone4-6A2, MabTech, Sweden). Plates were incubated for an
dditional 2 h at 37 ◦C and spot development was performeds described [34]. Spots were counted with an AutomatedLIspot Reader System with KS 4.3 software (Carl Zeiss,hornwood, NY).btam
e 25 (2007) 5330–5342
.8. Software
EpitOptimizer code was developed using Microsoft Visualasic (Microsoft, Redmond, WA). Web-based version isvailable at http://skiweb.mskcc.org/MHC/select.aspx.
. Results
.1. Automated design of heteroclitic peptides
The EpitOptimizer computer algorithm (Fig. 1A) goeshrough a set of steps to create optimized peptides. The pro-ram searches for single amino acid residues that can serves major or minor anchor residues for products of a specificHC class I allele [10,20–29] in any polypeptide sequence
f any length. For example, nonapeptides are created for H2-b motifs following two rules: (1) if position 5 is asparagine
N), then position 9 is changed to M or I (which are knownptimal anchors at position 9 for this MHC allele), and (2)nversely, if position 9 is M or I, then position 5 is changed to. For HLA-A*0201 nonapeptides, if position 2 is M, V, L or
, then position 6 is changed to V, L, or I (positions 2 and 9 arerimary anchors, position 6 is a secondary anchor). In addi-ion, position 6 is changed to V and position 1 is changed to. In summary, EpitOptimizer: (1) identifies potential anchor
esidues in a protein sequence and creates multiple mutatedorms to generate candidate heteroclitic epitopes (Fig. 1B);2) calculates estimated steady-state binding as a measure offfinity of each peptide for its cognate MHC molecule usingirtual quantitative matrices, as described by Parker et al. [22]Fig. 1B); (3) performs a BLAST search to check specificityf epitopes, specifically mutated sequences, by verifying thatnew candidate heteroclitic epitope does not mimic self pep-
ides encoded by other genes (e.g., mimic a peptide sequencexpressed in a crucial organ); (4) organizes all peptides intohierarchical list with predicted binding affinities (Fig. 1B).
.2. Computer generated heteroclitic peptides
To demonstrate the biological relevance of this approach,wo cancer-related proteins were initially used as test anti-ens, mouse TYRP1 [30] and human PSMA [31]. Theouse TYRP1 amino acid sequence (Fig. 1A) was ana-
yzed using EpitOptimizer for potential anchor residues thatight bind to H2-Db and human PSMA was analyzed forLA-A*0201-binding residues [22,24]. EpitOptimizer then
nserted mutated residues at candidate major and minornchor sites to design peptides for enhanced binding to thendicated MHC-I molecule. For TYRP1, a total of 537 can-idate wild-type peptides and 1591 optimized peptides (dataot shown) were identified based on the predicted peptide
inding to MHC-I molecules, calculated using virtual quan-itative matrices for the H2-Db motif (Fig. 1B). Human PSMAnalysis revealed 750 wild-type peptides and 2244 opti-ized peptides (data not shown). Wild-type peptides with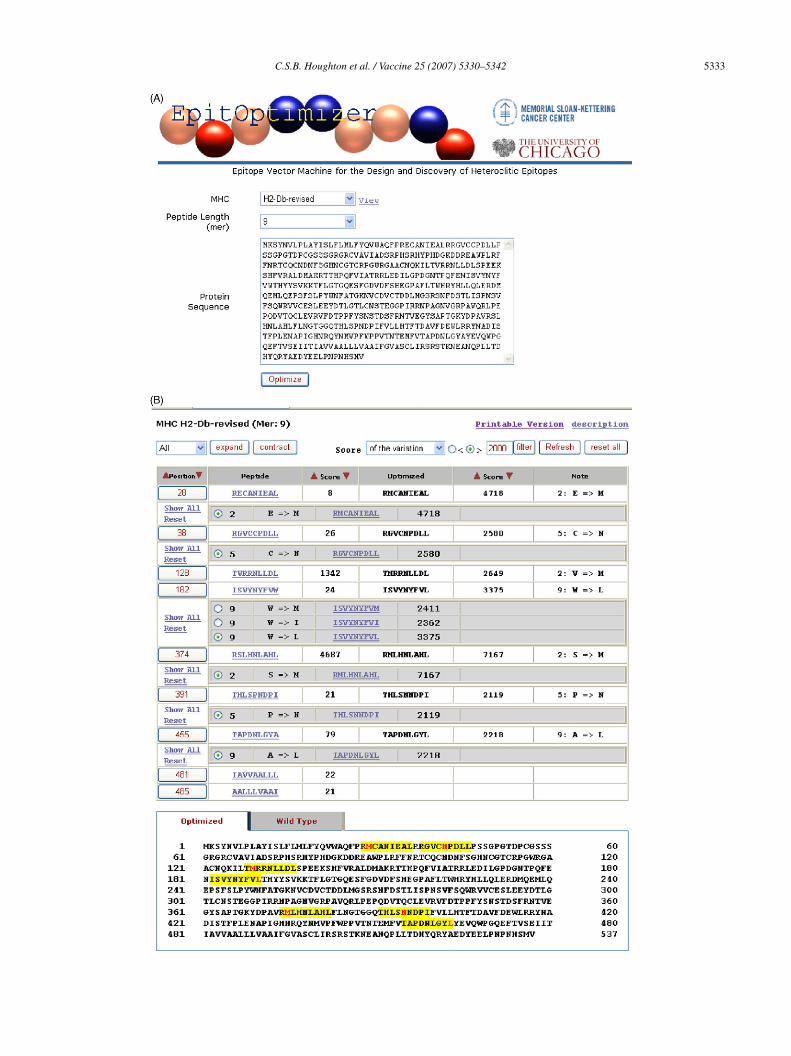
C.S.B. Houghton et al. / Vaccine 25 (2007) 5330–5342 5333
5334 C.S.B. Houghton et al. / Vaccine 25 (2007) 5330–5342
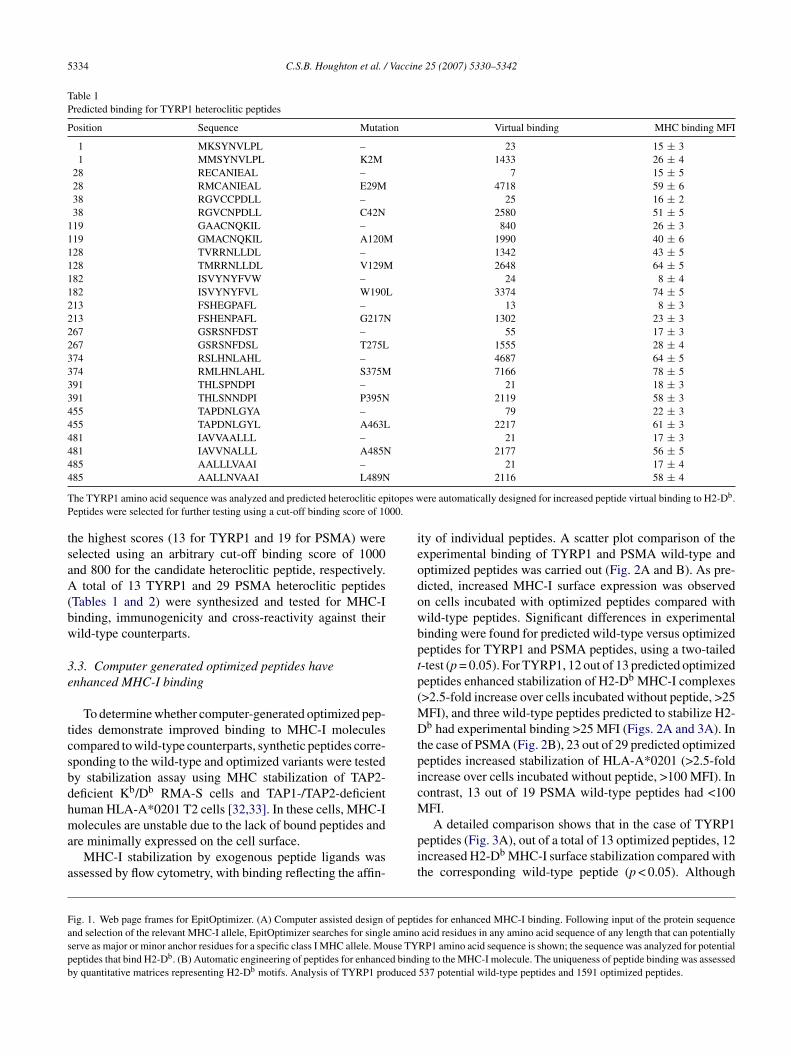
Table 1Predicted binding for TYRP1 heteroclitic peptides
Position Sequence Mutation Virtual binding MHC binding MFI
1 MKSYNVLPL – 23 15 ± 31 MMSYNVLPL K2M 1433 26 ± 4
28 RECANIEAL – 7 15 ± 528 RMCANIEAL E29M 4718 59 ± 638 RGVCCPDLL – 25 16 ± 238 RGVCNPDLL C42N 2580 51 ± 5
119 GAACNQKIL – 840 26 ± 3119 GMACNQKIL A120M 1990 40 ± 6128 TVRRNLLDL – 1342 43 ± 5128 TMRRNLLDL V129M 2648 64 ± 5182 ISVYNYFVW – 24 8 ± 4182 ISVYNYFVL W190L 3374 74 ± 5213 FSHEGPAFL – 13 8 ± 3213 FSHENPAFL G217N 1302 23 ± 3267 GSRSNFDST – 55 17 ± 3267 GSRSNFDSL T275L 1555 28 ± 4374 RSLHNLAHL – 4687 64 ± 5374 RMLHNLAHL S375M 7166 78 ± 5391 THLSPNDPI – 21 18 ± 3391 THLSNNDPI P395N 2119 58 ± 3455 TAPDNLGYA – 79 22 ± 3455 TAPDNLGYL A463L 2217 61 ± 3481 IAVVAALLL – 21 17 ± 3481 IAVVNALLL A485N 2177 56 ± 5485 AALLLVAAI – 21 17 ± 44
T itopes wP 00.
tsaA(bw
3e
tcsbdhma
a
ieodowbptp(MDtpicM
Faspb
85 AALLNVAAI L489N
he TYRP1 amino acid sequence was analyzed and predicted heteroclitic epeptides were selected for further testing using a cut-off binding score of 10
he highest scores (13 for TYRP1 and 19 for PSMA) wereelected using an arbitrary cut-off binding score of 1000nd 800 for the candidate heteroclitic peptide, respectively.
total of 13 TYRP1 and 29 PSMA heteroclitic peptidesTables 1 and 2) were synthesized and tested for MHC-Iinding, immunogenicity and cross-reactivity against theirild-type counterparts.
.3. Computer generated optimized peptides havenhanced MHC-I binding
To determine whether computer-generated optimized pep-ides demonstrate improved binding to MHC-I moleculesompared to wild-type counterparts, synthetic peptides corre-ponding to the wild-type and optimized variants were testedy stabilization assay using MHC stabilization of TAP2-eficient Kb/Db RMA-S cells and TAP1-/TAP2-deficientuman HLA-A*0201 T2 cells [32,33]. In these cells, MHC-I
olecules are unstable due to the lack of bound peptides andre minimally expressed on the cell surface.MHC-I stabilization by exogenous peptide ligands was
ssessed by flow cytometry, with binding reflecting the affin-
pit
ig. 1. Web page frames for EpitOptimizer. (A) Computer assisted design of peptind selection of the relevant MHC-I allele, EpitOptimizer searches for single aminoerve as major or minor anchor residues for a specific class I MHC allele. Mouse TYeptides that bind H2-Db. (B) Automatic engineering of peptides for enhanced bindiy quantitative matrices representing H2-Db motifs. Analysis of TYRP1 produced
2116 58 ± 4
ere automatically designed for increased peptide virtual binding to H2-Db.
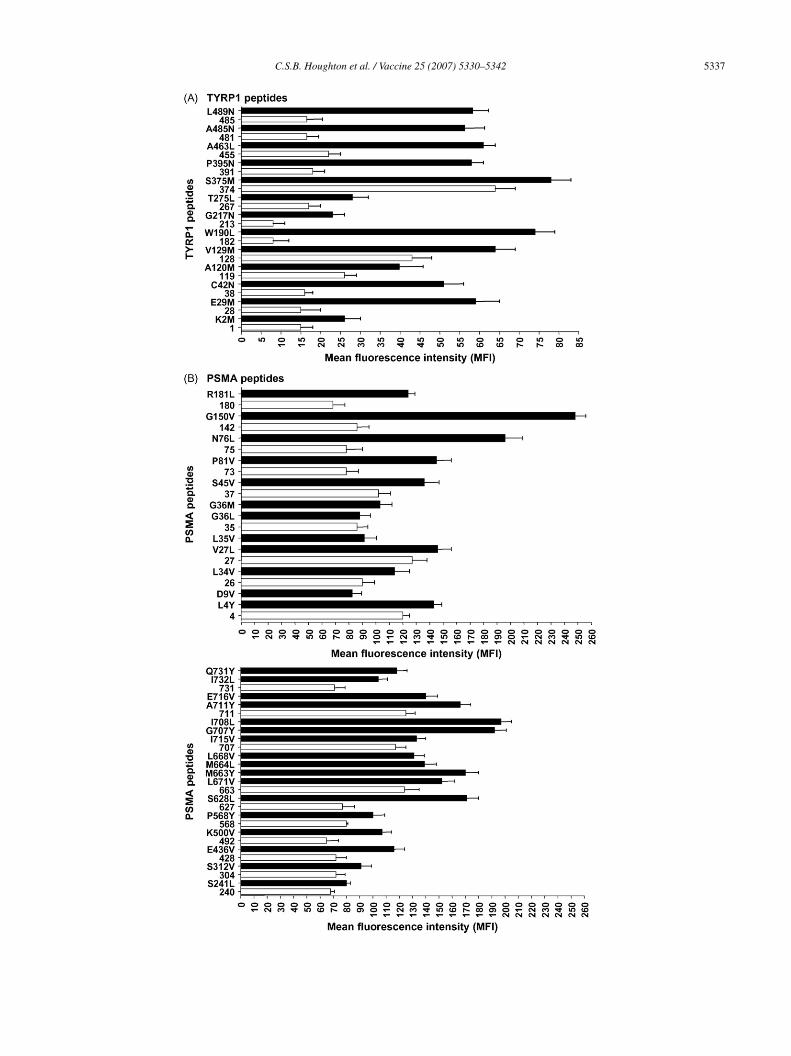
ty of individual peptides. A scatter plot comparison of thexperimental binding of TYRP1 and PSMA wild-type andptimized peptides was carried out (Fig. 2A and B). As pre-icted, increased MHC-I surface expression was observedn cells incubated with optimized peptides compared withild-type peptides. Significant differences in experimentalinding were found for predicted wild-type versus optimizedeptides for TYRP1 and PSMA peptides, using a two-tailed-test (p = 0.05). For TYRP1, 12 out of 13 predicted optimizedeptides enhanced stabilization of H2-Db MHC-I complexes>2.5-fold increase over cells incubated without peptide, >25
FI), and three wild-type peptides predicted to stabilize H2-b had experimental binding >25 MFI (Figs. 2A and 3A). In
he case of PSMA (Fig. 2B), 23 out of 29 predicted optimizedeptides increased stabilization of HLA-A*0201 (>2.5-foldncrease over cells incubated without peptide, >100 MFI). Inontrast, 13 out of 19 PSMA wild-type peptides had <100FI.
A detailed comparison shows that in the case of TYRP1eptides (Fig. 3A), out of a total of 13 optimized peptides, 12ncreased H2-Db MHC-I surface stabilization compared withhe corresponding wild-type peptide (p < 0.05). Although
des for enhanced MHC-I binding. Following input of the protein sequenceacid residues in any amino acid sequence of any length that can potentially
RP1 amino acid sequence is shown; the sequence was analyzed for potentialng to the MHC-I molecule. The uniqueness of peptide binding was assessed537 potential wild-type peptides and 1591 optimized peptides.
C.S.B. Houghton et al. / Vaccine 25 (2007) 5330–5342 5335
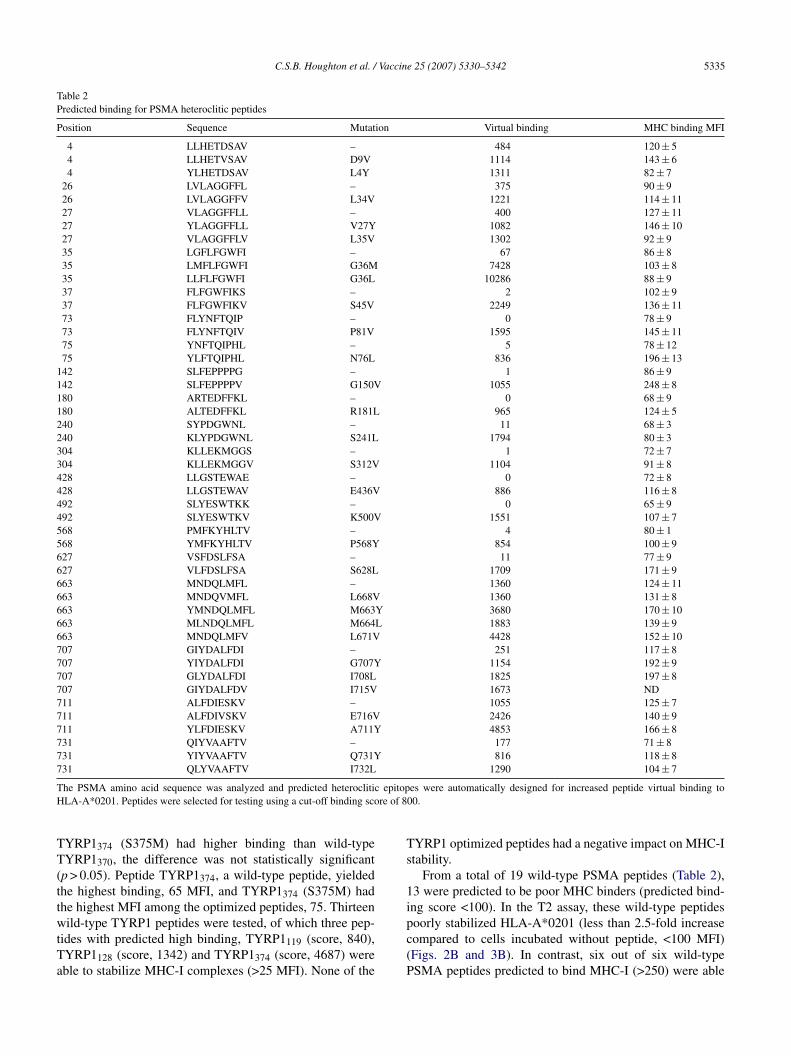
Table 2Predicted binding for PSMA heteroclitic peptides
Position Sequence Mutation Virtual binding MHC binding MFI
4 LLHETDSAV – 484 120 ± 54 LLHETVSAV D9V 1114 143 ± 64 YLHETDSAV L4Y 1311 82 ± 7
26 LVLAGGFFL – 375 90 ± 926 LVLAGGFFV L34V 1221 114 ± 1127 VLAGGFFLL – 400 127 ± 1127 YLAGGFFLL V27Y 1082 146 ± 1027 VLAGGFFLV L35V 1302 92 ± 935 LGFLFGWFI – 67 86 ± 835 LMFLFGWFI G36M 7428 103 ± 835 LLFLFGWFI G36L 10286 88 ± 937 FLFGWFIKS – 2 102 ± 937 FLFGWFIKV S45V 2249 136 ± 1173 FLYNFTQIP – 0 78 ± 973 FLYNFTQIV P81V 1595 145 ± 1175 YNFTQIPHL – 5 78 ± 1275 YLFTQIPHL N76L 836 196 ± 13
142 SLFEPPPPG – 1 86 ± 9142 SLFEPPPPV G150V 1055 248 ± 8180 ARTEDFFKL – 0 68 ± 9180 ALTEDFFKL R181L 965 124 ± 5240 SYPDGWNL – 11 68 ± 3240 KLYPDGWNL S241L 1794 80 ± 3304 KLLEKMGGS – 1 72 ± 7304 KLLEKMGGV S312V 1104 91 ± 8428 LLGSTEWAE – 0 72 ± 8428 LLGSTEWAV E436V 886 116 ± 8492 SLYESWTKK – 0 65 ± 9492 SLYESWTKV K500V 1551 107 ± 7568 PMFKYHLTV – 4 80 ± 1568 YMFKYHLTV P568Y 854 100 ± 9627 VSFDSLFSA – 11 77 ± 9627 VLFDSLFSA S628L 1709 171 ± 9663 MNDQLMFL – 1360 124 ± 11663 MNDQVMFL L668V 1360 131 ± 8663 YMNDQLMFL M663Y 3680 170 ± 10663 MLNDQLMFL M664L 1883 139 ± 9663 MNDQLMFV L671V 4428 152 ± 10707 GIYDALFDI – 251 117 ± 8707 YIYDALFDI G707Y 1154 192 ± 9707 GLYDALFDI I708L 1825 197 ± 8707 GIYDALFDV I715V 1673 ND711 ALFDIESKV – 1055 125 ± 7711 ALFDIVSKV E716V 2426 140 ± 9711 YLFDIESKV A711Y 4853 166 ± 8731 QIYVAAFTV – 177 71 ± 8731 YIYVAAFTV Q731Y 816 118 ± 8731 QLYVAAFTV I732L 1290 104 ± 7
T epitopH re of 8
TT(ttwtTa
Ts
1i
he PSMA amino acid sequence was analyzed and predicted heterocliticLA-A*0201. Peptides were selected for testing using a cut-off binding sco
YRP1374 (S375M) had higher binding than wild-typeYRP1370, the difference was not statistically significant
p > 0.05). Peptide TYRP1374, a wild-type peptide, yieldedhe highest binding, 65 MFI, and TYRP1374 (S375M) hadhe highest MFI among the optimized peptides, 75. Thirteen
ild-type TYRP1 peptides were tested, of which three pep-ides with predicted high binding, TYRP1119 (score, 840),YRP1128 (score, 1342) and TYRP1374 (score, 4687) wereble to stabilize MHC-I complexes (>25 MFI). None of the
pc(P
es were automatically designed for increased peptide virtual binding to00.
YRP1 optimized peptides had a negative impact on MHC-Itability.
From a total of 19 wild-type PSMA peptides (Table 2),3 were predicted to be poor MHC binders (predicted bind-ng score <100). In the T2 assay, these wild-type peptides
oorly stabilized HLA-A*0201 (less than 2.5-fold increaseompared to cells incubated without peptide, <100 MFI)Figs. 2B and 3B). In contrast, six out of six wild-typeSMA peptides predicted to bind MHC-I (>250) were able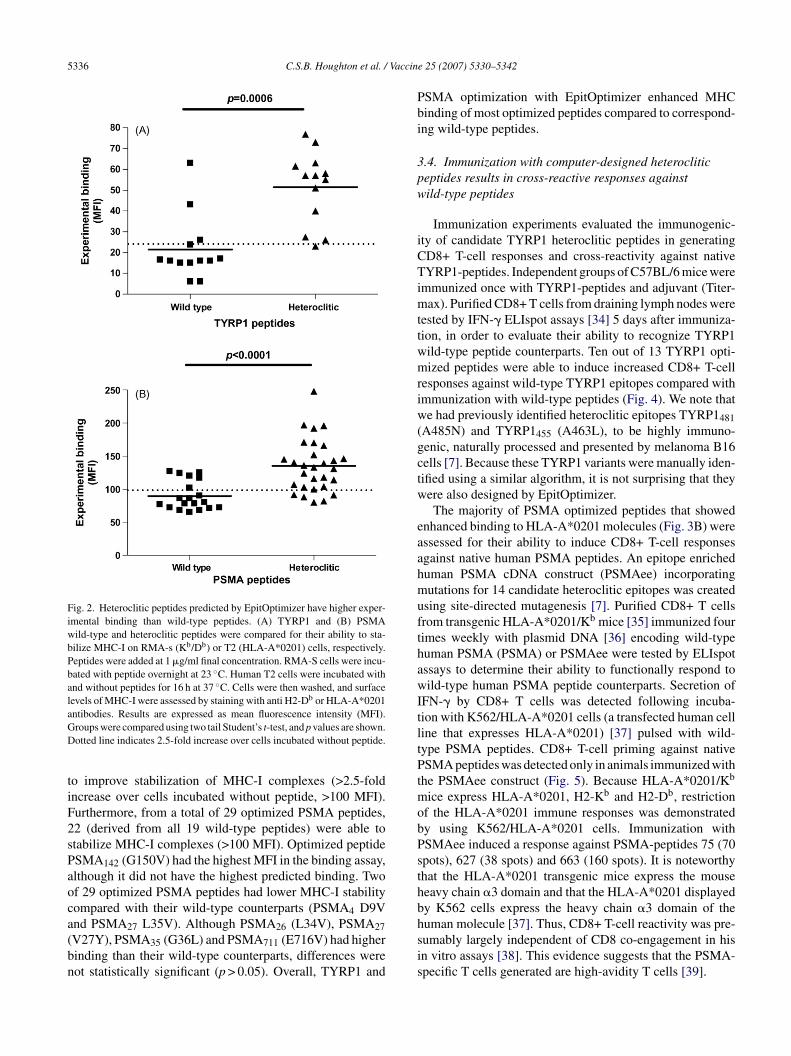
5336 C.S.B. Houghton et al. / Vaccin
Fig. 2. Heteroclitic peptides predicted by EpitOptimizer have higher exper-imental binding than wild-type peptides. (A) TYRP1 and (B) PSMAwild-type and heteroclitic peptides were compared for their ability to sta-bilize MHC-I on RMA-s (Kb/Db) or T2 (HLA-A*0201) cells, respectively.Peptides were added at 1 �g/ml final concentration. RMA-S cells were incu-bated with peptide overnight at 23 ◦C. Human T2 cells were incubated withand without peptides for 16 h at 37 ◦C. Cells were then washed, and surfacelevels of MHC-I were assessed by staining with anti H2-Db or HLA-A*0201aGD
tiF2sPaoca(bn
Pbi
3pw
iCTimttwmriw(gctw
eaahmufthawItltPtmobPsthby K562 cells express the heavy chain �3 domain of thehuman molecule [37]. Thus, CD8+ T-cell reactivity was pre-
ntibodies. Results are expressed as mean fluorescence intensity (MFI).roups were compared using two tail Student’s t-test, and p values are shown.otted line indicates 2.5-fold increase over cells incubated without peptide.
o improve stabilization of MHC-I complexes (>2.5-foldncrease over cells incubated without peptide, >100 MFI).urthermore, from a total of 29 optimized PSMA peptides,2 (derived from all 19 wild-type peptides) were able totabilize MHC-I complexes (>100 MFI). Optimized peptideSMA142 (G150V) had the highest MFI in the binding assay,lthough it did not have the highest predicted binding. Twof 29 optimized PSMA peptides had lower MHC-I stabilityompared with their wild-type counterparts (PSMA4 D9Vnd PSMA L35V). Although PSMA (L34V), PSMA
27 26 27V27Y), PSMA35 (G36L) and PSMA711 (E716V) had higherinding than their wild-type counterparts, differences wereot statistically significant (p > 0.05). Overall, TYRP1 andsis
e 25 (2007) 5330–5342
SMA optimization with EpitOptimizer enhanced MHCinding of most optimized peptides compared to correspond-ng wild-type peptides.
.4. Immunization with computer-designed heterocliticeptides results in cross-reactive responses againstild-type peptides
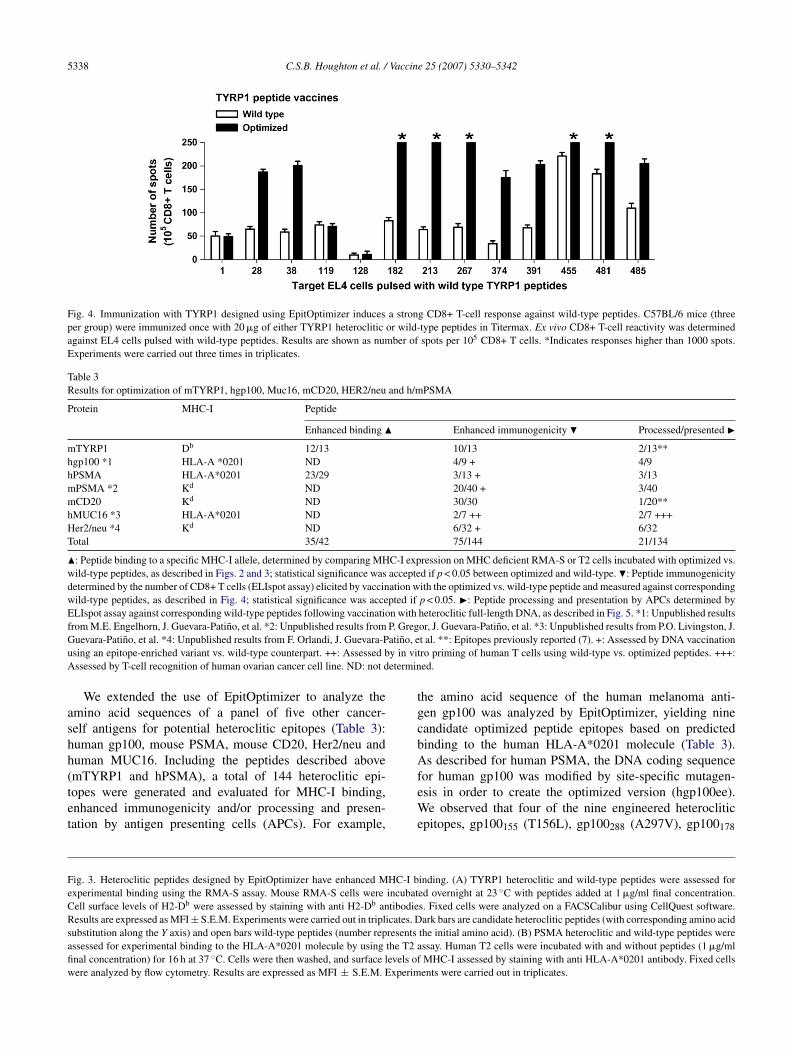
Immunization experiments evaluated the immunogenic-ty of candidate TYRP1 heteroclitic peptides in generatingD8+ T-cell responses and cross-reactivity against nativeYRP1-peptides. Independent groups of C57BL/6 mice were
mmunized once with TYRP1-peptides and adjuvant (Titer-ax). Purified CD8+ T cells from draining lymph nodes were
ested by IFN-� ELIspot assays [34] 5 days after immuniza-ion, in order to evaluate their ability to recognize TYRP1ild-type peptide counterparts. Ten out of 13 TYRP1 opti-ized peptides were able to induce increased CD8+ T-cell
esponses against wild-type TYRP1 epitopes compared withmmunization with wild-type peptides (Fig. 4). We note thate had previously identified heteroclitic epitopes TYRP1481
A485N) and TYRP1455 (A463L), to be highly immuno-enic, naturally processed and presented by melanoma B16ells [7]. Because these TYRP1 variants were manually iden-ified using a similar algorithm, it is not surprising that theyere also designed by EpitOptimizer.The majority of PSMA optimized peptides that showed
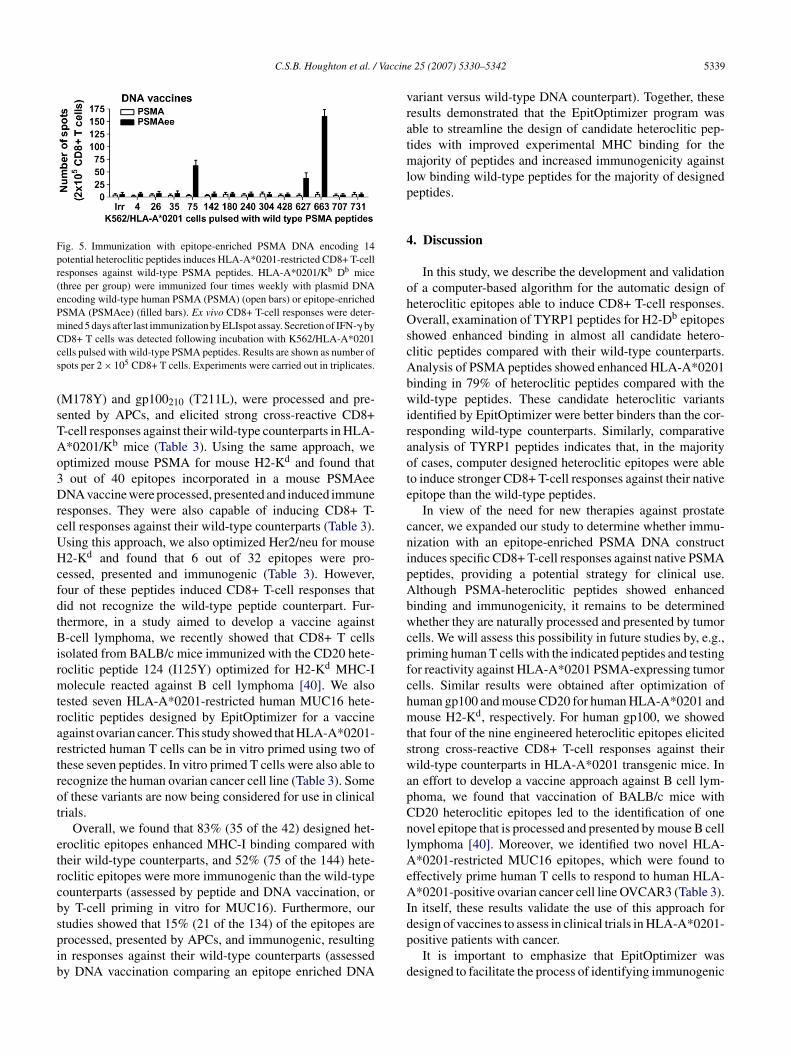
nhanced binding to HLA-A*0201 molecules (Fig. 3B) weressessed for their ability to induce CD8+ T-cell responsesgainst native human PSMA peptides. An epitope enricheduman PSMA cDNA construct (PSMAee) incorporatingutations for 14 candidate heteroclitic epitopes was created
sing site-directed mutagenesis [7]. Purified CD8+ T cellsrom transgenic HLA-A*0201/Kb mice [35] immunized fourimes weekly with plasmid DNA [36] encoding wild-typeuman PSMA (PSMA) or PSMAee were tested by ELIspotssays to determine their ability to functionally respond toild-type human PSMA peptide counterparts. Secretion of
FN-� by CD8+ T cells was detected following incuba-ion with K562/HLA-A*0201 cells (a transfected human celline that expresses HLA-A*0201) [37] pulsed with wild-ype PSMA peptides. CD8+ T-cell priming against nativeSMA peptides was detected only in animals immunized with
he PSMAee construct (Fig. 5). Because HLA-A*0201/Kb
ice express HLA-A*0201, H2-Kb and H2-Db, restrictionf the HLA-A*0201 immune responses was demonstratedy using K562/HLA-A*0201 cells. Immunization withSMAee induced a response against PSMA-peptides 75 (70pots), 627 (38 spots) and 663 (160 spots). It is noteworthyhat the HLA-A*0201 transgenic mice express the mouseeavy chain �3 domain and that the HLA-A*0201 displayed
umably largely independent of CD8 co-engagement in hisn vitro assays [38]. This evidence suggests that the PSMA-pecific T cells generated are high-avidity T cells [39].
C.S.B. Houghton et al. / Vaccine 25 (2007) 5330–5342 5337
5338 C.S.B. Houghton et al. / Vaccine 25 (2007) 5330–5342
Fig. 4. Immunization with TYRP1 designed using EpitOptimizer induces a strong CD8+ T-cell response against wild-type peptides. C57BL/6 mice (threeper group) were immunized once with 20 �g of either TYRP1 heteroclitic or wild-type peptides in Titermax. Ex vivo CD8+ T-cell reactivity was determinedagainst EL4 cells pulsed with wild-type peptides. Results are shown as number of spots per 105 CD8+ T cells. *Indicates responses higher than 1000 spots.Experiments were carried out three times in triplicates.
Table 3Results for optimization of mTYRP1, hgp100, Muc16, mCD20, HER2/neu and h/mPSMA
Protein MHC-I Peptide
Enhanced binding � Enhanced immunogenicity � Processed/presented �mTYRP1 Db 12/13 10/13 2/13**hgp100 *1 HLA-A *0201 ND 4/9 + 4/9hPSMA HLA-A*0201 23/29 3/13 + 3/13mPSMA *2 Kd ND 20/40 + 3/40mCD20 Kd ND 30/30 1/20**hMUC16 *3 HLA-A*0201 ND 2/7 ++ 2/7 +++Her2/neu *4 Kd ND 6/32 + 6/32Total 35/42 75/144 21/134
�: Peptide binding to a specific MHC-I allele, determined by comparing MHC-I expression on MHC deficient RMA-S or T2 cells incubated with optimized vs.wild-type peptides, as described in Figs. 2 and 3; statistical significance was accepted if p < 0.05 between optimized and wild-type. �: Peptide immunogenicitydetermined by the number of CD8+ T cells (ELIspot assay) elicited by vaccination with the optimized vs. wild-type peptide and measured against correspondingwild-type peptides, as described in Fig. 4; statistical significance was accepted if p < 0.05. �: Peptide processing and presentation by APCs determined byELIspot assay against corresponding wild-type peptides following vaccination with heteroclitic full-length DNA, as described in Fig. 5. *1: Unpublished resultsfrom M.E. Engelhorn, J. Guevara-Patino, et al. *2: Unpublished results from P. Gregor, J. Guevara-Patino, et al. *3: Unpublished results from P.O. Livingston, J.G atino, eu y in vitA etermin
ashh(tet
tgcbA
FeCRsafiw
uevara-Patino, et al. *4: Unpublished results from F. Orlandi, J. Guevara-Psing an epitope-enriched variant vs. wild-type counterpart. ++: Assessed bssessed by T-cell recognition of human ovarian cancer cell line. ND: not d
We extended the use of EpitOptimizer to analyze themino acid sequences of a panel of five other cancer-elf antigens for potential heteroclitic epitopes (Table 3):uman gp100, mouse PSMA, mouse CD20, Her2/neu anduman MUC16. Including the peptides described above
mTYRP1 and hPSMA), a total of 144 heteroclitic epi-opes were generated and evaluated for MHC-I binding,nhanced immunogenicity and/or processing and presen-ation by antigen presenting cells (APCs). For example,feWe
ig. 3. Heteroclitic peptides designed by EpitOptimizer have enhanced MHC-I bxperimental binding using the RMA-S assay. Mouse RMA-S cells were incubatell surface levels of H2-Db were assessed by staining with anti H2-Db antibodieesults are expressed as MFI ± S.E.M. Experiments were carried out in triplicates. D
ubstitution along the Y axis) and open bars wild-type peptides (number representsssessed for experimental binding to the HLA-A*0201 molecule by using the T2 anal concentration) for 16 h at 37 ◦C. Cells were then washed, and surface levels ofere analyzed by flow cytometry. Results are expressed as MFI ± S.E.M. Experim
t al. **: Epitopes previously reported (7). +: Assessed by DNA vaccinationro priming of human T cells using wild-type vs. optimized peptides. +++:ed.
he amino acid sequence of the human melanoma anti-en gp100 was analyzed by EpitOptimizer, yielding nineandidate optimized peptide epitopes based on predictedinding to the human HLA-A*0201 molecule (Table 3).s described for human PSMA, the DNA coding sequence
or human gp100 was modified by site-specific mutagen-sis in order to create the optimized version (hgp100ee).e observed that four of the nine engineered heteroclitic
pitopes, gp100155 (T156L), gp100288 (A297V), gp100178
inding. (A) TYRP1 heteroclitic and wild-type peptides were assessed fored overnight at 23 ◦C with peptides added at 1 �g/ml final concentration.s. Fixed cells were analyzed on a FACSCalibur using CellQuest software.ark bars are candidate heteroclitic peptides (with corresponding amino acid
the initial amino acid). (B) PSMA heteroclitic and wild-type peptides weressay. Human T2 cells were incubated with and without peptides (1 �g/mlMHC-I assessed by staining with anti HLA-A*0201 antibody. Fixed cellsents were carried out in triplicates.
C.S.B. Houghton et al. / Vaccin
Fig. 5. Immunization with epitope-enriched PSMA DNA encoding 14potential heteroclitic peptides induces HLA-A*0201-restricted CD8+ T-cellresponses against wild-type PSMA peptides. HLA-A*0201/Kb Db mice(three per group) were immunized four times weekly with plasmid DNAencoding wild-type human PSMA (PSMA) (open bars) or epitope-enrichedPSMA (PSMAee) (filled bars). Ex vivo CD8+ T-cell responses were deter-mined 5 days after last immunization by ELIspot assay. Secretion of IFN-� byCcs
(sTAo3DrcUHcfdtBirmtrartrot
etrcbspib
vratmlp
4
ohOscAbwiraote
cnipAbwcpfchmtswapCnlAeAI
D8+ T cells was detected following incubation with K562/HLA-A*0201ells pulsed with wild-type PSMA peptides. Results are shown as number ofpots per 2 × 105 CD8+ T cells. Experiments were carried out in triplicates.
M178Y) and gp100210 (T211L), were processed and pre-ented by APCs, and elicited strong cross-reactive CD8+-cell responses against their wild-type counterparts in HLA-*0201/Kb mice (Table 3). Using the same approach, weptimized mouse PSMA for mouse H2-Kd and found that
out of 40 epitopes incorporated in a mouse PSMAeeNA vaccine were processed, presented and induced immune
esponses. They were also capable of inducing CD8+ T-ell responses against their wild-type counterparts (Table 3).sing this approach, we also optimized Her2/neu for mouse2-Kd and found that 6 out of 32 epitopes were pro-
essed, presented and immunogenic (Table 3). However,our of these peptides induced CD8+ T-cell responses thatid not recognize the wild-type peptide counterpart. Fur-hermore, in a study aimed to develop a vaccine against-cell lymphoma, we recently showed that CD8+ T cells
solated from BALB/c mice immunized with the CD20 hete-oclitic peptide 124 (I125Y) optimized for H2-Kd MHC-Iolecule reacted against B cell lymphoma [40]. We also
ested seven HLA-A*0201-restricted human MUC16 hete-oclitic peptides designed by EpitOptimizer for a vaccinegainst ovarian cancer. This study showed that HLA-A*0201-estricted human T cells can be in vitro primed using two ofhese seven peptides. In vitro primed T cells were also able toecognize the human ovarian cancer cell line (Table 3). Somef these variants are now being considered for use in clinicalrials.
Overall, we found that 83% (35 of the 42) designed het-roclitic epitopes enhanced MHC-I binding compared withheir wild-type counterparts, and 52% (75 of the 144) hete-oclitic epitopes were more immunogenic than the wild-typeounterparts (assessed by peptide and DNA vaccination, ory T-cell priming in vitro for MUC16). Furthermore, our
tudies showed that 15% (21 of the 134) of the epitopes arerocessed, presented by APCs, and immunogenic, resultingn responses against their wild-type counterparts (assessedy DNA vaccination comparing an epitope enriched DNAdp
d
e 25 (2007) 5330–5342 5339
ariant versus wild-type DNA counterpart). Together, theseesults demonstrated that the EpitOptimizer program wasble to streamline the design of candidate heteroclitic pep-ides with improved experimental MHC binding for theajority of peptides and increased immunogenicity against
ow binding wild-type peptides for the majority of designedeptides.
. Discussion
In this study, we describe the development and validationf a computer-based algorithm for the automatic design ofeteroclitic epitopes able to induce CD8+ T-cell responses.verall, examination of TYRP1 peptides for H2-Db epitopes
howed enhanced binding in almost all candidate hetero-litic peptides compared with their wild-type counterparts.nalysis of PSMA peptides showed enhanced HLA-A*0201inding in 79% of heteroclitic peptides compared with theild-type peptides. These candidate heteroclitic variants
dentified by EpitOptimizer were better binders than the cor-esponding wild-type counterparts. Similarly, comparativenalysis of TYRP1 peptides indicates that, in the majorityf cases, computer designed heteroclitic epitopes were ableo induce stronger CD8+ T-cell responses against their nativepitope than the wild-type peptides.
In view of the need for new therapies against prostateancer, we expanded our study to determine whether immu-ization with an epitope-enriched PSMA DNA constructnduces specific CD8+ T-cell responses against native PSMAeptides, providing a potential strategy for clinical use.lthough PSMA-heteroclitic peptides showed enhancedinding and immunogenicity, it remains to be determinedhether they are naturally processed and presented by tumor
ells. We will assess this possibility in future studies by, e.g.,riming human T cells with the indicated peptides and testingor reactivity against HLA-A*0201 PSMA-expressing tumorells. Similar results were obtained after optimization ofuman gp100 and mouse CD20 for human HLA-A*0201 andouse H2-Kd, respectively. For human gp100, we showed
hat four of the nine engineered heteroclitic epitopes elicitedtrong cross-reactive CD8+ T-cell responses against theirild-type counterparts in HLA-A*0201 transgenic mice. In
n effort to develop a vaccine approach against B cell lym-homa, we found that vaccination of BALB/c mice withD20 heteroclitic epitopes led to the identification of oneovel epitope that is processed and presented by mouse B cellymphoma [40]. Moreover, we identified two novel HLA-*0201-restricted MUC16 epitopes, which were found to
ffectively prime human T cells to respond to human HLA-*0201-positive ovarian cancer cell line OVCAR3 (Table 3).
n itself, these results validate the use of this approach for
esign of vaccines to assess in clinical trials in HLA-A*0201-ositive patients with cancer.It is important to emphasize that EpitOptimizer wasesigned to facilitate the process of identifying immunogenic

5 Vaccin
vteaIotwtIaMiIaTrootttCTietbs
masTcoafwbgrpIahhommsi
aii
ciadifTosWattIhuTltaet
mvsFPcaep
A
RHGwTGfJAmbAJaa
340 C.S.B. Houghton et al. /
ariants, but natural processing and presentation of the wild-ype counterparts by tumor cells will still need to be evaluatedxperimentally. The program will be further improved bydding new features, including matrices for additional MHC-alleles of mouse, human and macaque and incorporationf matrices for MHC-II alleles. To facilitate peptide selec-ion, a proteasome algorithm may be useful in conjunctionith EpitOptimizer [41,42]. It is important to acknowledge
hat predictive algorithms are not 100% accurate [8,23,43].n some cases, certain peptides have been shown to lackdefined binding motif, but are capable of binding to theHC molecule. Conversely, peptides with a predicted bind-
ng motif may not bind to the MHC (i.e. PSMA D9V, L35V).n these cases, the reasons for the lack of binding are not clearnd range from peptide solubility to true absence of binding.he prediction power of EpitOptimizer is relative to the accu-
acy of the MHC-I matrices used [10]. The algorithm is basedn described primary and secondary anchor residues, and cannly design epitopes following defined motifs. Other poten-ial obstacles in heteroclitic peptide design are the breadth ofhe T-cell repertoire for each peptide:MHC-I complex andhe number of peptide-specific T cells for a given clone.onsequently, it is still unknown how many cross-reactivecells can be recruited using heteroclitic epitopes. Regard-
ng epitope selection, it is certainly prohibitive to analyzexperimentally all the variants generated for a single pro-ein and MHC molecule, and therefore, the ranking offeredy EpitOptimizer can help in making an educated peptideelection.
We have previously shown that the “selective gene opti-ization” approach, whereby genes are altered with the
im of converting immunologically inert self antigens intotrong anti-tumor vaccines, induced strong anti-tumor CD8+-cell responses, protected mice against melanoma tumorhallenge, and prolonged survival. This method is basedn T-cell epitope optimization, antigen epitope enrichment,nd enhancement of epitope processing, and was success-ully tested in a mouse melanoma model [7]. In this system,e created multiple immunogenic epitopes for enhancedinding to MHC-I molecules and further enhanced immuno-enicity by altering the vaccine’s cellular processing (i.e.,emoving the N-glycosylation sites), as a means to augmentroteolytic processing leading to stronger tumor-immunity.t is important to note that the “selective gene optimization”pproach has three implications for clinical applications: (1)igh-affinity peptides can stimulate low-avidity T cells thatave escaped immune regulation and selection; (2) inductionf simultaneous T-cell responses against multiple epitopesay prevent the immune-escape by tumor variants throughutations [44]; (3) the existence of defined epitopes allows
traightforward immune monitoring of T-cell responses inmmunized patients.
Although it is possible to manually design “selectivelyltered genes” and calculate theoretical binding constants forndividual peptides binding to a number of well character-zed MHC-I molecules, it takes days to weeks to identify and
imam
e 25 (2007) 5330–5342
arefully examine all the candidate peptides for MHC-I bind-ng motifs. Therefore, by developing an immune-informaticspproach (EpitOptimizer) that allows us to streamline theesign and discovery of peptides with enhanced MHC-I bind-ng and immunogenicity, we can survey target polypeptidesor all existing MHC-I binding motifs in a matter of minutes.he program also enables us to calculate the estimated affinityf heteroclitic peptides that may be generated by introducingingle amino acid substitutions in the MHC-I anchor residues.eb-based programs like MHCPred and MMBPred also
llow the computation of T cell epitopes within a larger pro-ein. These programs have several advantages, most notablyhe inclusion of a large panel of mouse and human MHC classand II alleles and tool for the design of promiscuous andigh affinity binders; however these programs only allow theser to input one peptide at a time for epitope enhancement.he utility of EpitOptimizer is its ability to search the entire
ength of a protein for potential epitopes and modify each ofhese epitopes for enhanced MHC binding. The final resultllows for the generation of a large number of potential het-roclitic peptides to choose from in a very short amount ofime.
In this study, we illustrate the potential use of EpitOpti-izer as an instrument that can accelerate heteroclitic-epitope
accine development. The use of this program should greatlytreamline the design of vaccines using heteroclitic epitopes.urthermore, we described three HLA-A*0201-restrictedSMA heteroclitic epitopes that can be further tested forlinical use. The data obtained demonstrate that this broadlypplicable approach can serve as the basis of epitope-ngineering for other antigens against cancer and intracellularathogens.
cknowledgements
Funding for this work was provided by NIH grants01 CA56821, P01 CA33049, and P01 CA59350 (to A.N.oughton); Swim Across America; the Mr. William H.oodwin and Mrs. Alice Goodwin and the Common-ealth Cancer Foundation for Research and the Experimentalherapeutics Center of MSKCC (to A.N. Houghton, J.A.uevara-Patino, M.E. Engelhorn); and a Fellowship award
rom the Cancer Research Institute (to M.E. Engelhorn)..D. Wolchok is supported by a Damon Runyon/Eli Lillyward, and A.N. Houghton has Damon Runyon/Eli Lillyentorship support. We are extremely grateful to the Annen-
erg Hazen Foundation as well as to the Louis & Annebrons Foundation, the T.J. Martell Foundation, the Peter
. Sharp Foundation/Breast Cancer Research Foundation,nd the Mr. and Mrs. Quentin J. Kennedy Fund. Theuthors wish to acknowledge the assistance of Dr. Rod-
ca Stan for her invaluable assistance in preparing theanuscript. We thank Mary Jo Turk for helpful commentsnd Linda Sherman for providing the HLA-A*0201/Kbice.

Vaccin
R
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
C.S.B. Houghton et al. /
eferences
[1] Solinger AM, Ultee ME, Margoliash E, Schwartz RH. T-lymphocyteresponse to cytochrome c. I. Demonstration of a T-cell heterocliticproliferative response and identification of a topographic antigenicdeterminant on pigeon cytochrome c whose immune recognitionrequires two complementing major histocompatibility complex-linkedimmune response genes. J Exp Med 1979;150:830.
[2] England RD, Kullberg MC, Cornette JL, Berzofsky JA. Molecular anal-ysis of a heteroclitic T cell response to the immunodominant epitopeof sperm whale myoglobin. Implications for peptide partial agonists. JImmunol 1995;155:4295.
[3] Dyall R, Bowne WB, Weber LW, LeMaoult J, Szabo P, Moroi Y, etal. Heteroclitic immunization induces tumor immunity. J Exp Med1998;188:1553.
[4] Bakker AB, van der Burg SH, Huijbens RJ, Drijfhout JW, Melief CJ,Adema GJ, et al. Analogues of CTL epitopes with improved MHC class-I binding capacity elicit anti-melanoma CTL recognizing the wild-typeepitope. Int J Cancer 1997;70:302.
[5] Parkhurst MR, Salgaller ML, Southwood S, Robbins PF, Sette A,Rosenberg SA, et al. Improved induction of melanoma-reactive CTLwith peptides from the melanoma antigen gp100 modified at HLA-A*0201-binding residues. J Immunol 1996;157:2539.
[6] Hombach J, Dumrese T, Rammensee HG, Zinkernagel RM, Hengart-ner H, Gallimore A. Hierarchies of antigen-specific cytotoxic T-cellresponses. Eur J Immunol 1998;28:3301.
[7] Guevara-Patino JA, Engelhorn ME, Turk MJ, Liu C, Duan F, RizzutoG, et al. Optimization of a self antigen for presentation of multipleepitopes in cancer immunity. J Clin Invest 2006;116:1382.
[8] Lu J, Celis E. Use of two predictive algorithms of the world wideweb for the identification of tumor-reactive T-cell epitopes. Cancer Res2000;60:5223.
[9] Rammensee H, Bachmann J, Emmerich NP, Bachor OA, Stevanovic S.SYFPEITHI: database for MHC ligands and peptide motifs. Immuno-genetics 1999;50:213.
10] Parker KC, Shields M, DiBrino M, Brooks A, Coligan JE. Peptidebinding to MHC class I molecules: implications for antigenic peptideprediction. Immunol Res 1995;14:34.
11] Restifo NP, Rosenberg SA. Developing recombinant and syntheticvaccines for the treatment of melanoma. Curr Opin Oncol 1999;11:50.
12] Cormier JN, Salgaller ML, Prevette T, Barracchini KC, Rivoltini L, Res-tifo NP, et al. Enhancement of cellular immunity in melanoma patientsimmunized with a peptide from Mart-1/Melan-A. Cancer J Sci Am1997;3:37.
13] Rosenberg SA, Yang JC, Schwartzentruber DJ, Hwu P, Marincola FM,Topalian SL, et al. Immunologic and therapeutic evaluation of a syn-thetic peptide vaccine for the treatment of patients with metastaticmelanoma [see comments]. Nat Med 1998;4:321.
14] Lewis JJ, Janetzki S, Schaed S, Panageas KS, Wang S, Williams L, et al.Evaluation of CD8(+) T-cell frequencies by the Elispot assay in healthyindividuals and in patients with metastatic melanoma immunized withtyrosinase peptide. Int J Cancer 2000;87:391.
15] Schaed SG, Klimek VM, Panageas KS, Musselli CM, Butterworth L,Hwu WJ, et al. T-cell responses against tyrosinase 368–376(370D)peptide in HLA(*)A0201(+) melanoma patients: randomized trialcomparing incomplete Freund’s adjuvant, granulocyte macrophagecolony-stimulating factor, and QS-21 as immunological adjuvants. ClinCancer Res 2002;8:967.
16] Kawashima I, Hudson SJ, Tsai V, Southwood S, Takesako K, Appella E,et al. The multi-epitope approach for immunotherapy for cancer: identi-fication of several CTL epitopes from various tumor-associated antigens
expressed on solid epithelial tumors. Hum Immunol 1998;59:1.17] Celis E, Tsai V, Crimi C, DeMars R, Wentworth PA, Chesnut RW, etal. Induction of anti-tumor cytotoxic T lymphocytes in normal humansusing primary cultures and synthetic peptide epitopes. Proc Natl AcadSci USA 1994;91:2105.
[
[
e 25 (2007) 5330–5342 5341
18] Kobayashi H, Kennedy R, Lu J, Davila E, Celis E. MHC-bindingpeptides as immunotherapeutics for cancer. Immunol Invest 2000;29:105.
19] Alexander RB, Brady F, Leffell MS, Tsai V, Celis E. Specific Tcell recognition of peptides derived from prostate-specific antigen inpatients with prostate cancer. Urology 1998;51:150.
20] Wilson IA, Fremont DH. Structural analysis of MHC class I moleculeswith bound peptide antigens. Semin Immunol 1993;5:75.
21] Parker KC, Bednarek MA, Coligan JE. Scheme for ranking potentialHLA-A2 binding peptides based on independent binding of individualpeptide side-chains. J Immunol 1994;152:163.
22] Parker KC, Biddison WE, Coligan JE. Pocket mutations of HLA-B27show that anchor residues act cumulatively to stabilize peptide binding.Biochemistry 1994;33:7736.
23] Sette A, Sidney J, del Guercio MF, Southwood S, Ruppert J, DahlbergC, et al. Peptide binding to the most frequent HLA-A class I alle-les measured by quantitative molecular binding assays. Mol Immunol1994;31:813.
24] Ruppert J, Sidney J, Celis E, Kubo RT, Grey HM, Sette A. Prominentrole of secondary anchor residues in peptide binding to HLA-A2.1molecules. Cell 1993;74:929.
25] DiBrino M, Parker KC, Shiloach J, Knierman M, Lukszo J, TurnerRV, et al. Endogenous peptides bound to HLA-A3 possess a spe-cific combination of anchor residues that permit identification ofpotential antigenic peptides. Proc Natl Acad Sci USA 1993;90:1508.
26] Falk K, Rotzschke O, Rammensee HG. A self-peptide naturallypresented by both H-2Kb and H-2Kbm1 molecules demonstratesMHC restriction of self-tolerance at the molecular level. Int Immunol1992;4:321.
27] Fremont DH, Stura EA, Matsumura M, Peterson PA, Wilson IA. Crystalstructure of an H-2Kb-ovalbumin peptide complex reveals the interplayof primary and secondary anchor positions in the major histocompat-ibility complex binding groove. Proc Natl Acad Sci USA 1995;92:2479.
28] Grey HM, Ruppert J, Vitiello A, Sidney J, Kast WM, Kubo RT, et al.Class I MHC–peptide interactions: structural requirements and func-tional implications. Cancer Surv 1995;22:37.
29] Tourdot S, Scardino A, Saloustrou E, Gross DA, Pascolo S, CordopatisP, et al. A general strategy to enhance immunogenicity of low-affinityHLA-A2.1-associated peptides: implication in the identification ofcryptic tumor epitopes. Eur J Immunol 2000;30:3411.
30] Vijayasaradhi S, Bouchard B, Houghton AN. The melanoma antigengp75 is the human homologue of the mouse b (brown) locus geneproduct. J Exp Med 1990;171:1375.
31] Israeli RS, Powell CT, Fair WR, Heston WD. Molecular cloning of acomplementary DNA encoding a prostate-specific membrane antigen.Cancer Res 1993;53:227.
32] Hosken NA, Bevan MJ. An endogenous antigenic peptide bypasses theclass I antigen presentation defect in RMA-S. J Exp Med 1992;175:719.
33] Salter RD, Howell DN, Cresswell P. Genes regulating HLA classI antigen expression in T–B lymphoblast hybrids. Immunogenetics1985;21:235.
34] Scheibenbogen C, Lee KH, Mayer S, Stevanovic S, Moebius U, HerrW, et al. A sensitive ELISPOT assay for detection of CD8+ T lym-phocytes specific for HLA class I-binding peptide epitopes derivedfrom influenza proteins in the blood of healthy donors and melanomapatients. Clin Cancer Res 1997;3:221.
35] Theobald M, Biggs J, Hernandez J, Lustgarten J, Labadie C, ShermanLA. Tolerance to p53 by A2.1-restricted cytotoxic T lymphocytes. JExp Med 1997;185:833.
36] Weber LW, Bowne WB, Wolchok JD, Srinivasan R, Qin J, Moroi Y, etal. Tumor immunity and autoimmunity induced by immunization withhomologous DNA. J Clin Invest 1998;102:1258.
37] Britten CM, Meyer RG, Kreer T, Drexler I, Wolfel T, Herr W. The useof HLA-A*0201-transfected K562 as standard antigen-presenting cells

5 Vaccin
[
[
[
[
[
[
342 C.S.B. Houghton et al. /
for CD8(+) T lymphocytes in IFN-gamma ELISPOT assays. J ImmunolMethods 2002;259:95.
38] LaFace DM, Vestberg M, Yang Y, Srivastava R, DiSanto J, Flomen-berg N, et al. Human CD8 transgene regulation of HLA recognition bymurine T cells. J Exp Med 1995;182:1315.
39] Sherman LA, Hesse SV, Irwin MJ, La Face D, Peterson P. SelectingT cell receptors with high affinity for self-MHC by decreasing thecontribution of CD8. Science 1992;258:815.
40] Palomba ML, Roberts WK, Dao T, Manukian G, Guevara-PatinoJA, Wolchok JD, et al. CD8+ T-cell-dependent immunity fol-lowing xenogeneic DNA immunization against CD20 in a tumorchallenge model of B-cell lymphoma. Clin Cancer Res 2005;11:370.
[
e 25 (2007) 5330–5342
41] Altuvia Y, Margalit H. Sequence signals for generation of antigenicpeptides by the proteasome: implications for proteasomal cleavagemechanism. J Mol Biol 2000;295:879.
42] Holzhutter HG, Frommel C, Kloetzel PM. A theoretical approachtowards the identification of cleavage-determining amino acid motifsof the 20 S proteasome. J Mol Biol 1999;286:1251.
43] Schirle M, Weinschenk T, Stevanovic S. Combining computer algo-rithms with experimental approaches permits the rapid and accurate
identification of T cell epitopes from defined antigens. J ImmunolMethods 2001;257:1.44] Restifo NP, Antony PA, Finkelstein SE, Leitner WW, Surman DP, The-oret MR, et al. Assumptions of the tumor ‘escape’ hypothesis. SeminCancer Biol 2002;12:81.
Copyright © 2022 FDOKUMEN




















