
Influence of vitrification on mouse metaphase II oocyte spindle dynamics and chromatin alignment

Ftwa©
E
Available online at www.sciencedirect.com
Theriogenology 77 (2012) 1741–1753
0d
Potential applications of sheep oocytes as affected by vitrificationand in vitro aging
S.M. Hosseinia, V. Asgaria, S. Ostadhosseinia, M. Hajiana, A. Piryaeib, M. Najaraslb,M.H. Nasr-Esfahania,c,*
a Department of Reproduction and Development, Reproductive Biomedicine Centre, Royan Institute for Animal Biotechnology, ACECR,Isfahan, Islamic Republic of Iran
b Department of Stem Cells and Developmental Biology, Cell Science Research Center, Royan Institute for Stem Cell Biology andTechnology, ACECR, Tehran, Islamic Republic of Iran
c Department of Embryology, Reproductive Biomedicine Centre, Royan Institute for Reproductive Biomedicine, ACECR, Tehran, IslamicRepublic of Iran
Received 28 August 2011; received in revised form 28 November 2011; accepted 4 December 2011
Abstract
The present study was carried out to investigate how the interactions between aging, vitrification and post-warming intervalaffect the credibility of sheep MII-oocytes for in vitro fertilization (IVF), intracytoplasmic injection (ICSI), and parthenogeneticactivation (PA). According to our results, aged oocytes had significantly higher rates of chromosome and spindle abnormalitiescompared to young oocytes. However after vitrification-warming, the total rates of these abnormalities were not significantlydifferent between aged and young oocytes. Unvitrified-aged, and vitrified young and aged oocytes had comparable ultrastructuralcharacteristics, whereas they were completely dissimilar in compared with unvitrified-young oocytes. Although mRNA abundancewas reduced during vitrification-warming in both aged and young oocytes, the post-warming interval could improve the relativemRNA abundance. Aged oocytes had lower capacity for IVF and ICSI in compared with young oocytes, but had similar patternfor PA process. The vitrification process decreased developmental competence of both aged and young oocytes in compared withyoung ones, particularly when warmed oocytes were rested for 2 h before IVF, ICSI and PA. The results of the present studyshowed that in vitro aged oocytes had higher capacity to be used for parthenogenetic studies rather than IVF and ICSI.
urthermore, it was shown that vitrified oocytes had a time-dependent decline in quality and developmental potential. Notably,he speed of this decline was higher in vitrified-young oocytes, indicating that the vitrified oocytes do not require to be rested postarming. Conclusively, the results of this study can be useful in preserving in vitro aged oocytes to provide a valuable and easy
ccess source of oocytes for research purposed studies.2012 Elsevier Inc. All rights reserved.
Keywords: Vitrification; Oocyte aging; ICSI; IVF; Parthenogenetic activation
www.theriojournal.com
1. Introduction
In vivo or in vitro matured, metaphase II (MII)oocyte has a limited time-window for optimal fertiliza-
* Corresponding author. Tel.: �983112600446; fax: �983112605525.E-mail address: [email protected] (M.H. Nasr-
sfahani).
093-691X/$ – see front matter © 2012 Elsevier Inc. All rights reserved.oi:10.1016/j.theriogenology.2011.12.005
tion with a progressive deterioration in quality; whichnamed oocyte aging [1,2]. A considerable volume ofstudies suggest that cellular and molecular changesduring aging may render oocytes unfavorable for as-sisted reproductive technique (ART) (for review, see[1]). An increasing body of evidence suggests that agedoocytes may serve as a valuable source for several
research purposes, including production of embryonic
btcu
esign.
1742 S.M. Hosseini et al. / Theriogenology 77 (2012) 1741–1753
stem cells (ESCs) using somatic cell nuclear transfer(SCNT) and parthenogenetic embryos [2–4]. Interest-ingly, it has been reported that it is possible to inhibitand delay molecular pathways of oocyte aging usingchemical agents, such as caffeine, MG-132, and nitricoxide, respectively. These approaches may improve thepotential applications of in vitro aged oocytes in diversefields [1,5]. Therefore, establishing an efficient cryo-ank of vitrified-aged oocytes would provide an ethicalool for objective assessment of oocyte developmentalompetence in such area and also for preclinical eval-ation of experimental procedures [2,4].
Upon cryopreservation, oocyte undergoes a wide rangeof injuries at cellular and molecular levels which mayhave important consequences on embryo development[6,7]. Given the acknowledged importance of initialoocyte quality on ART outcomes [8], most cryopreserva-tion studies have been focused on invariably freshly ovu-lated oocytes to provide a platform for successful results[9–11]. To date, however, it is poorly understood how theinteractions between aging and vitrification affect thecredibility of MII-oocytes for further development.
Through several studies, time-dependent capability ofvitrified-warmed oocytes for recovering the cryoassoci-ated injuries has been established, particularly for spindleand chromosome disorganization [12]. Accordingly, vit-
Fig. 1. Diagrammatic representation of experimental d
rified-warmed matured oocytes are commonly rested for
up to 3 h before further treatment [13]. Through thepresent study, we observed that post-warming intervaladversely affected the meiotic progression and partheno-genetic activation of vitrified-warmed sheep oocytes ma-tured for 20 to 22 h in vitro [7]. Moreover, using confocalmicroscopy, high resolution optical sectioning, and 3-Dimage analysis Bromfield, et al. [1] provided evidencesindicating the importance of subjecting cryopreservedoocytes to in vitro fertilization (IVF) or intracytoplasmicinjection (ICSI) sooner than is routinely practiced in ARTclinics [14]. Importantly, considering that the agedoocytes have established lower capacity to maintain theirintegrity in compared with the young ones [14,15], thepossible advantages vs. disadvantages of this time intervalin vitrified-aged oocytes needs to be assessed. Therefore,as depicted in Fig. 1, this study was accomplished toinvestigate the interaction effects of aging, vitrificationand post-warming interval on the credibility of sheepMII-oocytes to be used for IVF, ICSI, and parthenoge-netic activation (PA).
2. Materials and methods
Unless otherwise mentioned, all chemicals and mediawere obtained from Sigma, Chemical Co. (St. Louis, MO,
hpm: hour post maturation, hpw: hour post warming.
USA) and Gibco (Grand Island, NY, USA), respectively.

M
egBUcwB(1vE
AW
1743S.M. Hosseini et al. / Theriogenology 77 (2012) 1741–1753
2.1. Oocyte in vitro maturation
Abattoir-derived sheep ovaries were used for in vitrooocyte maturation (IVM) as described previously [7].In brief, cumulus-oocyte complexes (COCs) were as-pirated of antral follicles (2–6 mm) using 20-gaugeneedles attached to a vacuum pump (80 mm Hg). Next,COCs with more than three layers of cumulus cells andhomogenous cytoplasm were selected and washedthrice in HEPES-supplemented tissue culture medium -199 (H-TCM199) �10% estrus sheep serum and thenin maturation medium (MM). Eventually, COCs werecultured in groups of 20 to 25 and 100 �l droplets of
M covered with mineral oil at 39 °C, 5% CO2 andhumidified air. The MM was comprised of TCM-199containing Na-pyruvate (2.5 mM), L-glutamine (1 mM),penicillin (100 IU/mL), streptomycin (100 �g/mL),sheep serum (10% v/v), EGF (100 �M), IGF (100 �M),ovine FSH (10 �g/mL), ovine LH (10 �g/mL), estra-diol-17� (1 �g/mL) and cysteamine (0.1 mM).
2.2. Experimental design
At 18 to 20 (young) or 30 to 32 (aged) h post-matura-tion, the oocytes were used for the designed experiments.As shown in Fig. 1, the vitrified-warmed oocytes, eitherimmediately (immediate group: IG) or two h post-warm-ing (delayed group: DG) along with their correspondingunvitrified controls were used for assessment of (i) sur-vival rate, (ii) meiotic spindle and chromosomal organi-zation, (iii) ultrastructural changes, (iv) gene expression,(v) cortical granule release, and zona hardening and (vi)embryo development using IVF, ICSI, and PA.
2.2.1. Vitrification-warmingThe process of oocyte vitrification-warming was as
described previously [7]. In brief, the oocytes wereincubated in 100-�L basic solution (BS) containingphosphate buffer saline (PBS) plus 20% serum for 1min. Meanwhile, three separate 100-�L droplets ofquilibration solution [ES: containing 7.5% ethylenelycol (EG) and 7.5% dimethyl sulfoxide (DMSO) inS] were placed in vicinity of each other and BS.nder stereomicroscope, the first BS droplet was gently
onnected to its nearest ES drop, and the BS-ES dropas connected to the next ES drop. Ultimately, theS-ES-ES drop was connected to the third ES drop
BS-ES-ES-ES). The time taken between each step wasmin. The equilibrated oocytes were then incubated in
itrification solution (VS) comprised of BS plus 15%G 15% DMSO and 0.5 M sucrose for 20 to 25 s. In the
next step, five oocytes were loaded on the tip of the
cryotop (Cryologic; CVM, Fibreplug and Sleeve, Aus-tralia) and quickly plunged in liquid nitrogen (LN2). Allincubations were performed at room temperature (RT).For the warming process, cryotops were removed fromLN2 and then the applied tip quickly plunged in warm-ing solution 1 (WS1: 1 M sucrose in BS at 38.5 °C).
fter 1 min, the warmed oocytes were transferred intoS2 for 3 min (0.5 M sucrose in BS) and then another
3 min into WS3 (0.25 M sucrose in BS). Afterward, theoocytes were incubated in the washing solution (PBS �20% serum) for 5 min and finally transferred to TCM-199 plus 10% serum.
2.2.2. SurvivalThe survival rate of oocytes was assessed as de-
scribed by Succu, et al. (2007) [16]. Accordingly,oocytes with morphologic evidences of shrinkage, cy-toplasmic vacuolization, damaged membrane or zonapellucida were considered as non-viable.
2.2.3. Immunoflourescence microscopyThe oocyte microtubules were immunostained with
anti-�-tubulin monoclonal primary antibody (1:100) inPBS and fluorescein isothiocyanate (FITC)-labeled sec-ondary antibody (1:100) as described previously [17].Chromosomes were counterstained with 10 �g/mL ofH33342. Then, oocytes were washed and mounted onglass slides containing a glycerol droplet for usingepifluorescence microscopy (Olympus BX51, Japan).Digital images of each oocyte were taken with a highlysensitive camera at 400� magnification (DP-72 Olym-pus, Japan) using DP2-BSW software. Spindle andchromosome configurations (Fig. 2) were categorizedaccording to the previous reports [7,18,19].
2.2.4. Transmission electron microscopyThe oocytes were washed three times in PBS and
fixed in Karnovsky fixative at RT for 20 min. After that,they were subsequently rinsed in PBS for 10 min andpost-fixed with 1% osmium tetroxide in PBS for 3 h.The prepared samples were dehydrated in ascendingconcentrations of ethanol solutions (50–100%), substi-tuted in propylene oxide, embedded in araldite resinand eventually serially semithin-sectioned (2 mm).Semithin sections were stained with toluidine blue andexamined under light microscope. The selected sampleswere subsequently reembedded, and ultrathin sectionswere stained with uranyl acetate, and lead citrate andthen examined by transmission electron microscope
(Philips CM 10, Netherlands).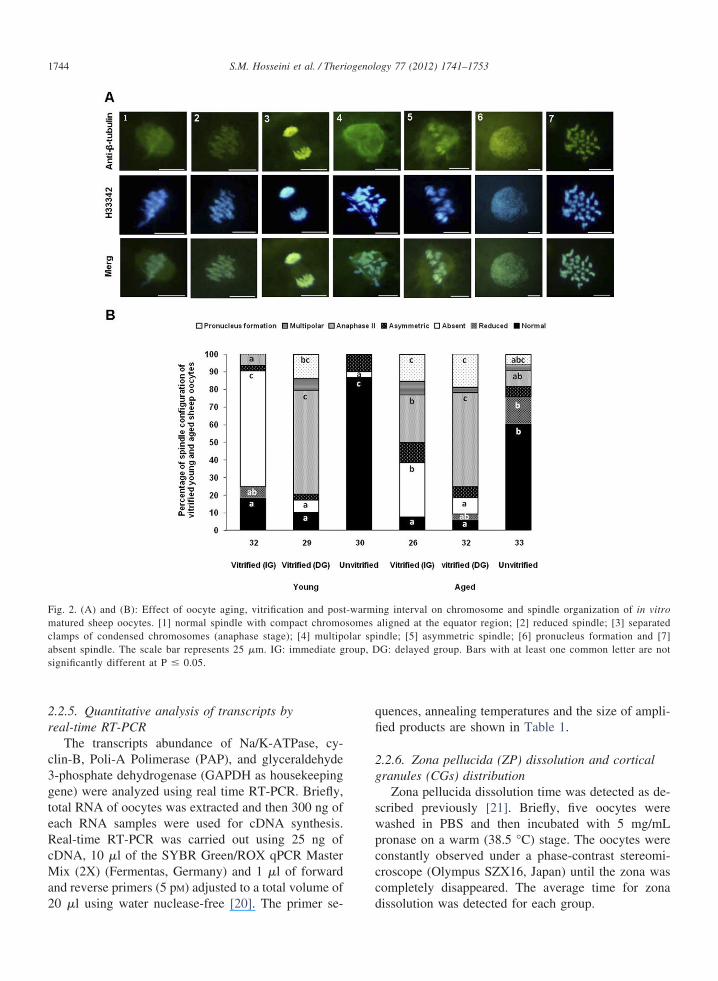
2
1744 S.M. Hosseini et al. / Theriogenology 77 (2012) 1741–1753
2.2.5. Quantitative analysis of transcripts byreal-time RT-PCR
The transcripts abundance of Na/K-ATPase, cy-clin-B, Poli-A Polimerase (PAP), and glyceraldehyde3-phosphate dehydrogenase (GAPDH as housekeepinggene) were analyzed using real time RT-PCR. Briefly,total RNA of oocytes was extracted and then 300 ng ofeach RNA samples were used for cDNA synthesis.Real-time RT-PCR was carried out using 25 ng ofcDNA, 10 �l of the SYBR Green/ROX qPCR MasterMix (2X) (Fermentas, Germany) and 1 �l of forwardand reverse primers (5 pM) adjusted to a total volume of
Fig. 2. (A) and (B): Effect of oocyte aging, vitrification and postmatured sheep oocytes. [1] normal spindle with compact chromoclamps of condensed chromosomes (anaphase stage); [4] multipabsent spindle. The scale bar represents 25 �m. IG: immediate gsignificantly different at P � 0.05.
0 �l using water nuclease-free [20]. The primer se-
quences, annealing temperatures and the size of ampli-fied products are shown in Table 1.
2.2.6. Zona pellucida (ZP) dissolution and corticalgranules (CGs) distribution
Zona pellucida dissolution time was detected as de-scribed previously [21]. Briefly, five oocytes werewashed in PBS and then incubated with 5 mg/mLpronase on a warm (38.5 °C) stage. The oocytes wereconstantly observed under a phase-contrast stereomi-croscope (Olympus SZX16, Japan) until the zona wascompletely disappeared. The average time for zona
ing interval on chromosome and spindle organization of in vitroaligned at the equator region; [2] reduced spindle; [3] separatedindle; [5] asymmetric spindle; [6] pronucleus formation and [7]G: delayed group. Bars with at least one common letter are not
-warmsomesolar sproup, D
dissolution was detected for each group.

0t
zo
t
B
2tmTmoro
2pb
1745S.M. Hosseini et al. / Theriogenology 77 (2012) 1741–1753
The pattern of CGs distribution in the vitrified-warmed and control oocytes was assessed as describedby Tian, et al. (2007) [22]. In brief, after the removal ofzona, the oocytes were washed in PBS before beingfixed with 3.7% paraformaldehyde in PBS at RT for 30min. Fixed oocytes were then washed three times inPBS containing 0.3% bovine serum albumin (BSA) and100-mM glycine. The oocytes were then incubated in.1% Triton X-100 in PBS for 5 min, washed threeimes in PBS and then incubated in 100 �g/mL FITC-
labeled PNA in PBS for 30 min in a dark place. Afterremoving the residual unbound dyes through washingwith PBS containing 0.3% BSA and 0.01% TritonX-100, the oocytes were washed and mounted on non-fluorescent slides and observed under an epifluores-cence microscope (Olympus BX51, Japan) at 400�magnification. The fluorescence emission of eachoocyte was detected under 450 to 490-nm excitation byusing 490 nm filter for CGs. Upon exposure, a digitalimage of each oocyte provided with a high sensitivecamera (DP-72 Olympus, Japan) operated on DP2-BSW software.
2.2.7. Embryo development
2.2.7.1. In vitro fertilization (IVF). The process of IVFwas carried out as described previously [23]. In brief,
Table 1Sequence specific primers used for qRT-PCR.
Gene Primer sequence (5=–3=)
GAPDH F: TGCCGCCTGGAGAAACCR: TGAAGTCGCAGGAGACAAC
Na/K-ATPase F: GCTGACTTGGTCATCTGCAAR: CATTCCAGGGCAGTAGGAA
Cyclin-B F: GATTGGAGAGGTTGATGTTGR: AGGTAATGCTGTAGAGTTG
Poli A Polimerase F: GAACTTGCCAGCTTTATCCAR: TTGTGGGTATGCTGGTGTAA
Table 2Survival rate of young and in vitro aged sheep oocytes as affectedby vitrification and post warming interval. IG: immediate group,DG: delayed group.
Age of oocytes Group Oocyte no. Viability (%)
Young Vitrified (IG) 38 92.0 � 0.5Vitrified (DG) 35 91.3 � 0.6Unvitrified 28 100 � 0
Aged Vitrified (IG) 32 94.8 � 5.1Vitrified (DG) 30 89.6 � 5.7
Unvitrified 35 100 � 0frozen ram semen straws were thawed in a warm waterbath (37 °C) for 40 s and then centrifuged at 700 g for10 min at RT to remove cryoprotectants. Then, highlymotile sperm were retrieved via centrifugation on a60:40 gradient of Pure Sperm (Nidacon, Gothenburg,Sweden) at 700 g for 15 min at RT. Meanwhile, thematured oocytes were washed trice in fertilization me-dium (Fert-TALP) and a total of 4 � 105 sperm wasadded to each 200-�L droplet of IVF medium contain-ing 40 to 50 oocytes under oil. IVF was performed at39 °C, 5% CO2 in humidified air for 24 h. Presumptiveygotes were then washed trice in modified syntheticviductal fluid (mSOF), cultured in groups of 10 in 20
�L droplet of mSOF without serum and glucose for 3days, and subsequently in the presence of charcoalstripped serum (5%) and glucose (1.5 mM) for 5 days.The cleavage, blastocyst and hatching rates were deter-mined on the day 3, 7 and 8 post embryo culture,respectively. The mSOF was based on Tervit, et al.(1972) [24], and was comprised of NaCl (107.7 mM),KCl (7.15 mM), KH2PO4 (0.3 mM), NaHCO3 (5 mM),Na-lactate (3.32 mM), penicillin-G (100 IU/mL), strep-omycin (100 �g/mL), Na-pyruvate (0.33 mM),
CaCl2.2H2O (1.71 mM), fatty acid free BSA (3 mg/mL),ME amino acids (2%) and MEM amino acids (1%).
.2.7.2. PA. As described by Hosseini, et al. (2008) [25],he matured oocytes were first activated with 5 �M iono-ycin prepared in H-TCM199 (devoid of FCS) for 1 min.hey were then thoroughly washed and incubated for 5in in H-TCH199 � 30 mg/mL BSA. Afterward, the
ocytes were incubated with 2 mM 6-dimethyl aminopu-ine (6-DMAP) for 2 h and eventually the activatedocytes were washed and cultured as for IVF.
.2.7.3. Intracytoplasmic Sperm Injection (ICSI). Therocess of ICSI was as described previously [26]. Inrief, oocytes were transferred to 15-�L droplets of
Length (bp) Annealing temperature (°C)
121 60
129 58
174 62
203 54
C
AAG
GTG
H-TCM plus 10% serum prepared in ICSI petri dish (50 �

tH
epe
3
tTu
3
i
woivam(
1746 S.M. Hosseini et al. / Theriogenology 77 (2012) 1741–1753
9 mm2, BD Falcon, USA). Highly motile sperm (pre-pared as described for IVF) were introduced into poly-vinylpyrolidone medium (Cook, Australia) prepared inH-TCM199. The motile sperm with normal morphol-ogy were immobilized by tail scoring using an invertedmicroscope (Nikon Eclipse TE-2000-U, Japan) equippedwith manipulators (Eppendrof, Germany). Then, thesperm aspirated into the injection pipettes tail first, andthen injected into the oocyte. Two h post injection, theoocytes were activated using ionomycin (5 �M, 1 min),horoughly washed and incubated for 5 min in-TCH199 � 30 mg/mL BSA before being cultured as
described for IVF process.
2.3. Statistical analysis
All the experiments in the present study were re-peated at least three times. The Percentages data weremodeled to the binomial model of parameters byArcSin transformation. The transformed data in de-velopmental competency, and the data related togene expression and zona dissolution time were an-alyzed by one way ANOVA model of SPSS 17. Thexisting differences were compared by Tukey multi-le comparison post hoc test. To determine differ-nces in tubulin and chromatin organization, the �2
test, or Fisher exact test when appropriate, were used. Itis noteworthy to mention that differences consideredsignificant at P � 0.05.
. Results
For both unvitrified young and aged oocytes, 2 hime interval did not affect the oocyte characteristics.herefore, these data were pooled and represented asnited controls.
.1. Survival rate
Morphologic assessments of vitrified-warmed oocytesndicated that neither oocyte aging, nor the vitrification
process significantly affected the survival rate of theoocytes (Table 2).
3.2. Spindle and chromosome organization
Figure 2 represents sample pictures of different chro-mosomal and spindle organizations observed in oocytes ofdifferent groups. According to the results, the chromo-some and spindle abnormalities of unvitrified-agedoocytes were significantly higher than young ones (39.3vs. 13.3%, respectively). Vitrification significantly in-
creased the chromosome abnormalities in both young andaged oocytes in compared with unvitrified ones. Immedi-ately after warming, the major patterns of spindle abnor-malities in vitrified-young oocytes were absent (65.6%,Fig. 2A7 and B) and AII (6.2%, Fig. 2A3 and B) which
ere significantly different from that of the vitrified-agedocytes (30.7 and 26.9%, respectively). The post-warm-ng interval had no effect on the total abnormalities initrified young and aged oocytes, though the pattern ofbnormalities differed. Accordingly, most spindle abnor-alities in the rested vitrified-young oocytes were AII
58.6%) and PN (13.7%, Fig. 2A6 and B) which wereslightly different in vitrified-aged oocytes (53.1 and18.7%, respectively).
3.3. Transmission electron microscopy
As displayed in Fig. 3, the unvitrified-young oocytesexhibited typical ultrastructural features of MII-oocytes,including obvious perivitelline space (Fig. 3A), numerousvacuoles (Fig. 3B), homogenous lipid droplet (Fig. 3D),regular oolemma with numerous microvilli, and withelectron-dense cortical granules that aligned in a single(and somewhere double) layer beneath the oolemma(Fig. 3A). Mitochondria showed a relatively electrondense matrix and occasionally was crossed by smalltransverse cristae (Fig. 3C). Aside from minor changesand irrespective of post-warming interval, unvitrified-aged, vitrified-warmed young and aged oocytes hadcomparable ultrastructural characteristics which werecompletely different in the unvitrified-young oocytes.According to our results, irregular oolemma (Fig. 3I, K,M), shorter and thicker microvilli (Fig. 3I), less abun-dant CGs (Fig. 3I, K, M), non-homogenous lipid drop-let (Fig. 3E, N), more cytoplasm vacuolization (Fig. 3F,G), degenerating mitochondria (Fig. 3H) and mitochon-dria with decreased matrix electron density (Fig. 3L, N)were observed in these groups in compared with unvit-rified-young oocytes. However post-warming intervalwas not a matter of distinction between the groups.Noteworthy, irrespective of post-warming interval, acharacteristic organelle-free area was developed be-neath the oolemma of unvitrified- and vitrified-agedoocytes (Fig. 3I, J, K, M).
3.4. Real-time RT-PCR
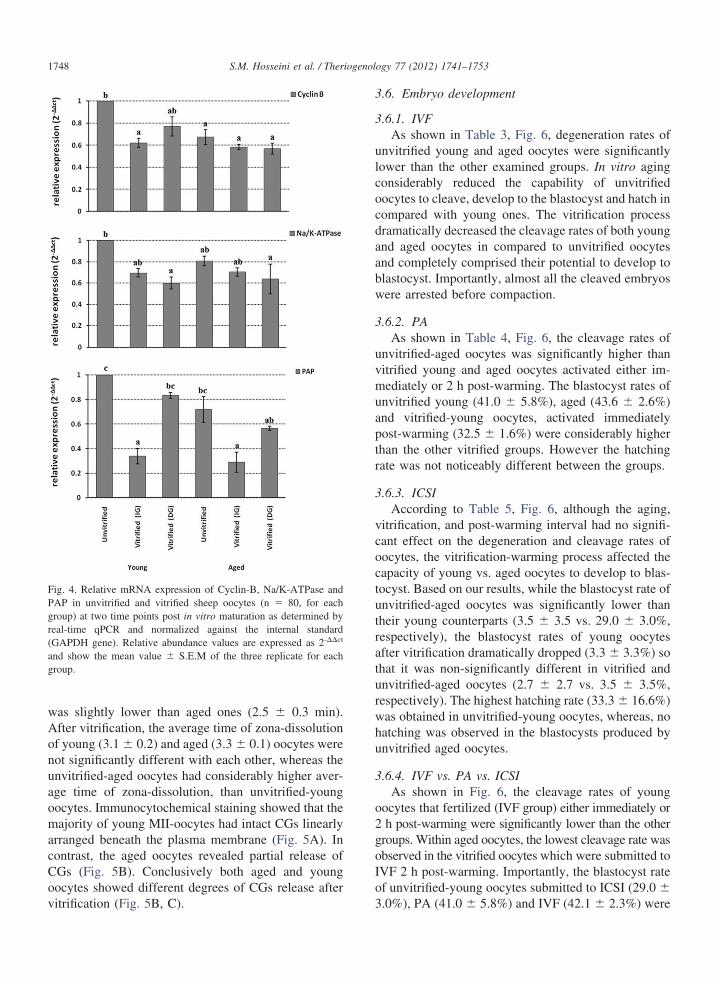
The aged oocytes had lower mRNA abundance forcyclin-B, Na/K-ATPase and Poli-A Polimerase, whereastheir young counterparts had significant mRNA abun-dance only for cyclin-B. Immediately after warming,mRNA abundance of cyclin-B in vitrified-young, andPoli-A in vitrified young and aged oocytes were signifi-
cantly lower than unvitrified oocytes. Interestingly, after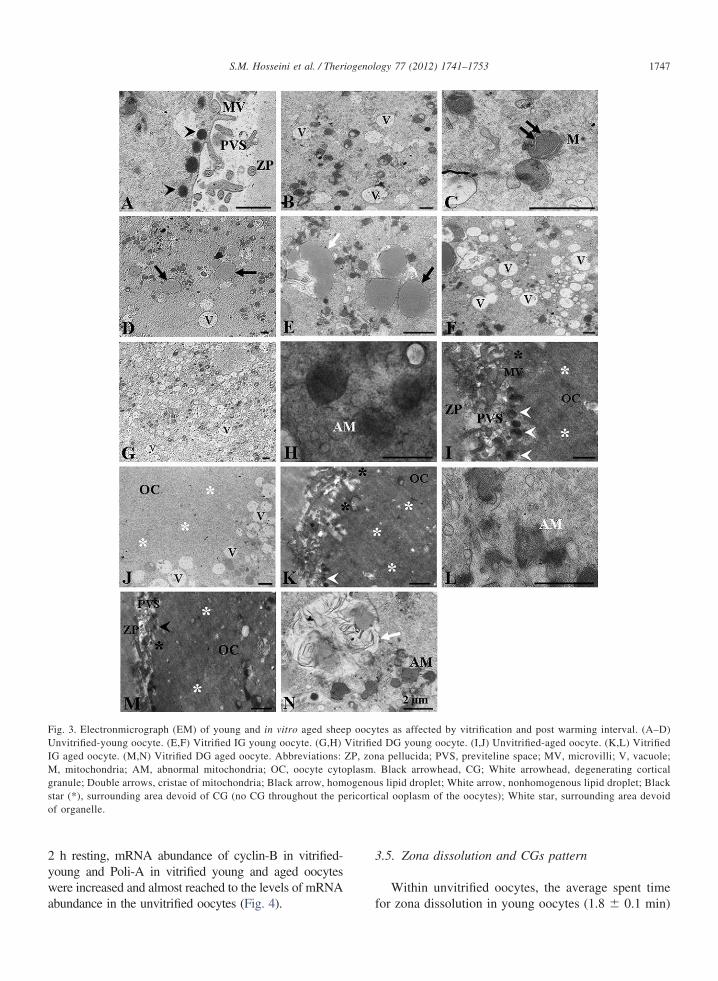
UIMgso
1747S.M. Hosseini et al. / Theriogenology 77 (2012) 1741–1753
2 h resting, mRNA abundance of cyclin-B in vitrified-young and Poli-A in vitrified young and aged oocyteswere increased and almost reached to the levels of mRNA
Fig. 3. Electronmicrograph (EM) of young and in vitro aged sheenvitrified-young oocyte. (E,F) Vitrified IG young oocyte. (G,H)
G aged oocyte. (M,N) Vitrified DG aged oocyte. Abbreviations:, mitochondria; AM, abnormal mitochondria; OC, oocyte cyto
ranule; Double arrows, cristae of mitochondria; Black arrow, homtar (*), surrounding area devoid of CG (no CG throughout the pf organelle.
abundance in the unvitrified oocytes (Fig. 4).
3.5. Zona dissolution and CGs pattern
Within unvitrified oocytes, the average spent time
tes as affected by vitrification and post warming interval. (A–D)d DG young oocyte. (I,J) Unvitrified-aged oocyte. (K,L) Vitrifieda pellucida; PVS, previteline space; MV, microvilli; V, vacuole;Black arrowhead, CG; White arrowhead, degenerating corticals lipid droplet; White arrow, nonhomogenous lipid droplet; Black
cal ooplasm of the oocytes); White star, surrounding area devoid
p oocyVitrifieZP, zonplasm.ogenou
ericorti
for zona dissolution in young oocytes (1.8 � 0.1 min)
ulcocdaabw
3
uvmu
g
1748 S.M. Hosseini et al. / Theriogenology 77 (2012) 1741–1753
was slightly lower than aged ones (2.5 � 0.3 min).After vitrification, the average time of zona-dissolutionof young (3.1 � 0.2) and aged (3.3 � 0.1) oocytes werenot significantly different with each other, whereas theunvitrified-aged oocytes had considerably higher aver-age time of zona-dissolution, than unvitrified-youngoocytes. Immunocytochemical staining showed that themajority of young MII-oocytes had intact CGs linearlyarranged beneath the plasma membrane (Fig. 5A). Incontrast, the aged oocytes revealed partial release ofCGs (Fig. 5B). Conclusively both aged and youngoocytes showed different degrees of CGs release after
Fig. 4. Relative mRNA expression of Cyclin-B, Na/K-ATPase andPAP in unvitrified and vitrified sheep oocytes (n � 80, for eachgroup) at two time points post in vitro maturation as determined byreal-time qPCR and normalized against the internal standard(GAPDH gene). Relative abundance values are expressed as 2-��ct
and show the mean value � S.E.M of the three replicate for eachroup.
vitrification (Fig. 5B, C).
3.6. Embryo development
3.6.1. IVFAs shown in Table 3, Fig. 6, degeneration rates of
nvitrified young and aged oocytes were significantlyower than the other examined groups. In vitro agingonsiderably reduced the capability of unvitrifiedocytes to cleave, develop to the blastocyst and hatch inompared with young ones. The vitrification processramatically decreased the cleavage rates of both youngnd aged oocytes in compared to unvitrified oocytesnd completely comprised their potential to develop tolastocyst. Importantly, almost all the cleaved embryosere arrested before compaction.
.6.2. PAAs shown in Table 4, Fig. 6, the cleavage rates of
nvitrified-aged oocytes was significantly higher thanitrified young and aged oocytes activated either im-ediately or 2 h post-warming. The blastocyst rates of
nvitrified young (41.0 � 5.8%), aged (43.6 � 2.6%)and vitrified-young oocytes, activated immediatelypost-warming (32.5 � 1.6%) were considerably higherthan the other vitrified groups. However the hatchingrate was not noticeably different between the groups.
3.6.3. ICSIAccording to Table 5, Fig. 6, although the aging,
vitrification, and post-warming interval had no signifi-cant effect on the degeneration and cleavage rates ofoocytes, the vitrification-warming process affected thecapacity of young vs. aged oocytes to develop to blas-tocyst. Based on our results, while the blastocyst rate ofunvitrified-aged oocytes was significantly lower thantheir young counterparts (3.5 � 3.5 vs. 29.0 � 3.0%,respectively), the blastocyst rates of young oocytesafter vitrification dramatically dropped (3.3 � 3.3%) sothat it was non-significantly different in vitrified andunvitrified-aged oocytes (2.7 � 2.7 vs. 3.5 � 3.5%,respectively). The highest hatching rate (33.3 � 16.6%)was obtained in unvitrified-young oocytes, whereas, nohatching was observed in the blastocysts produced byunvitrified aged oocytes.
3.6.4. IVF vs. PA vs. ICSIAs shown in Fig. 6, the cleavage rates of young
oocytes that fertilized (IVF group) either immediately or2 h post-warming were significantly lower than the othergroups. Within aged oocytes, the lowest cleavage rate wasobserved in the vitrified oocytes which were submitted toIVF 2 h post-warming. Importantly, the blastocyst rateof unvitrified-young oocytes submitted to ICSI (29.0 �
3.0%), PA (41.0 � 5.8%) and IVF (42.1 � 2.3%) were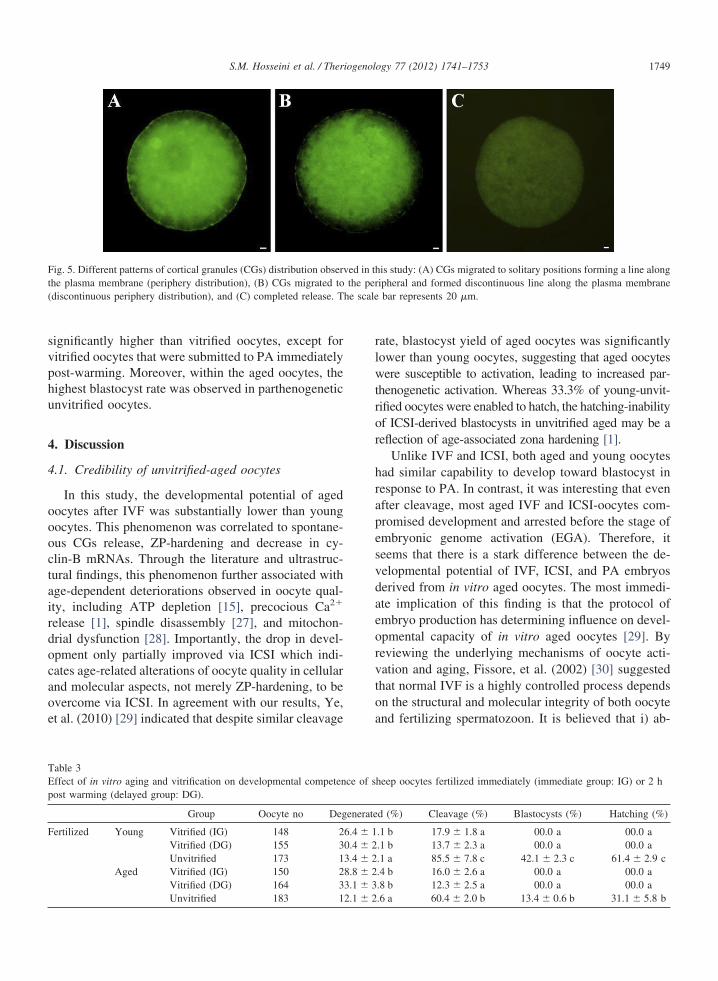
aeorvtoa
he scale
p
F
1749S.M. Hosseini et al. / Theriogenology 77 (2012) 1741–1753
significantly higher than vitrified oocytes, except forvitrified oocytes that were submitted to PA immediatelypost-warming. Moreover, within the aged oocytes, thehighest blastocyst rate was observed in parthenogeneticunvitrified oocytes.
4. Discussion
4.1. Credibility of unvitrified-aged oocytes
In this study, the developmental potential of agedoocytes after IVF was substantially lower than youngoocytes. This phenomenon was correlated to spontane-ous CGs release, ZP-hardening and decrease in cy-clin-B mRNAs. Through the literature and ultrastruc-tural findings, this phenomenon further associated withage-dependent deteriorations observed in oocyte qual-ity, including ATP depletion [15], precocious Ca2�
release [1], spindle disassembly [27], and mitochon-drial dysfunction [28]. Importantly, the drop in devel-opment only partially improved via ICSI which indi-cates age-related alterations of oocyte quality in cellularand molecular aspects, not merely ZP-hardening, to beovercome via ICSI. In agreement with our results, Ye,et al. (2010) [29] indicated that despite similar cleavage
Fig. 5. Different patterns of cortical granules (CGs) distribution obserthe plasma membrane (periphery distribution), (B) CGs migrated to(discontinuous periphery distribution), and (C) completed release. T
Table 3Effect of in vitro aging and vitrification on developmental competenost warming (delayed group: DG).
Group Oocyte no Deg
ertilized Young Vitrified (IG) 148 2Vitrified (DG) 155 3Unvitrified 173 1
Aged Vitrified (IG) 150 2Vitrified (DG) 164 3
Unvitrified 183 12.1 � 2rate, blastocyst yield of aged oocytes was significantlylower than young oocytes, suggesting that aged oocyteswere susceptible to activation, leading to increased par-thenogenetic activation. Whereas 33.3% of young-unvit-rified oocytes were enabled to hatch, the hatching-inabilityof ICSI-derived blastocysts in unvitrified aged may be areflection of age-associated zona hardening [1].
Unlike IVF and ICSI, both aged and young oocyteshad similar capability to develop toward blastocyst inresponse to PA. In contrast, it was interesting that evenafter cleavage, most aged IVF and ICSI-oocytes com-promised development and arrested before the stage ofembryonic genome activation (EGA). Therefore, itseems that there is a stark difference between the de-velopmental potential of IVF, ICSI, and PA embryosderived from in vitro aged oocytes. The most immedi-te implication of this finding is that the protocol ofmbryo production has determining influence on devel-pmental capacity of in vitro aged oocytes [29]. Byeviewing the underlying mechanisms of oocyte acti-ation and aging, Fissore, et al. (2002) [30] suggestedhat normal IVF is a highly controlled process dependsn the structural and molecular integrity of both oocytend fertilizing spermatozoon. It is believed that i) ab-
is study: (A) CGs migrated to solitary positions forming a line alongipheral and formed discontinuous line along the plasma membranebar represents 20 �m.
heep oocytes fertilized immediately (immediate group: IG) or 2 h
d (%) Cleavage (%) Blastocysts (%) Hatching (%)
.1 b 17.9 � 1.8 a 00.0 a 00.0 a
.1 b 13.7 � 2.3 a 00.0 a 00.0 a
.1 a 85.5 � 7.8 c 42.1 � 2.3 c 61.4 � 2.9 c
.4 b 16.0 � 2.6 a 00.0 a 00.0 a
.8 b 12.3 � 2.5 a 00.0 a 00.0 a
ved in ththe per
ce of s
enerate
6.4 � 10.4 � 23.4 � 28.8 � 23.1 � 3
.6 a 60.4 � 2.0 b 13.4 � 0.6 b 31.1 � 5.8 b

1750 S.M. Hosseini et al. / Theriogenology 77 (2012) 1741–1753
Fig. 6. Effect of vitrification, aging and post-warming interval on in vitro development of sheep oocytes derived by intracytoplasmic sperminjection (ICSI), parthenogenetic activation (PA) and in vitro fertilization (IVF). A–F, bars with at least one common letter are not significantly
different at P � 0.05. hpm: hour post maturation: IG Immediate group: DG Delayed group: UN Unvitrified.
(
P
2.1
TE(
1751S.M. Hosseini et al. / Theriogenology 77 (2012) 1741–1753
normal calcium homeostasis, ii) decreased amounts ofantiapoptotic proteins, and iii) spontaneous meiosis re-sumption are likely to play an important role in limitingdevelopmental potential, and may signal arrest in de-velopment [1]. Therefore, these changes could explainthe poor efficiency of aged oocyte when they were usedfor IVF and ICSI.
Cyclin-B is the regulatory subunit of meiosis pro-moting factor (MPF) which is responsible for G2-toM-phase transition. Cyclin-B degradation results inMPF depression which in turn triggers meiotic matu-ration events [31]. A sharp decrease in MPF is a po-tential crisis for matured oocyte cryobiology linked toprecocious calcium release which may have importantconsequences on developmental potential of vitrifiedoocytes [32]. For example, precocious MPF reductionmay result in a-synchronization of oocyte and spermpronuclei formation [33]. Importantly, it was observedthat relative abundance of Poli-A polimerase (whichinvolved in constitutive cell functions [22]) was recov-ered when aged and young oocytes were rested for twoh post-warming. Similar trend was also observed forcyclin-B in young oocytes. Although it is unexpectedfor completely condensed chromatin to undergo mRNAexpression, and therefore MII-oocytes is generally con-sidered transcriptionally inactive [34]. This phenome-non may be related to partial chromatin decondensationthat was observed in vitrified-warmed oocytes [18].
Table 4Effect of in vitro aging and vitrification on developmental competenimmediate group: IG) or 2 h post warming (delayed group: DG).
Group Oocyte no
arthenogeneticallyactivated
Young Vitrified (IG) 167Vitrified (DG) 190Unvitrified 173
Aged Vitrified (IG) 182Vitrified (DG) 159Unvitrified 241
able 5ffect of in vitro aging and vitrification on developmental competen
immediate group: IG) or 2 h post warming (delayed group: DG).
Group Oocyteno.
Degen
ICSI Young Vitrified (IG) 59 19Vitrified (DG) 58 19Unvitrified 78 14
Aged Vitrified (IG) 99 24Vitrified (DG) 94 18
Unvitrified 94 22.3 � 3.7In conclusion, stark differences between cellular andmolecular characteristics of aged vs. young oocytesconvinced us that aged oocytes should have substan-tially lower capability than young ones. Although thishypothesis was confirmed by IVF and ICSI, develop-mental potential of parthenogenetic embryos clearlyindicates that aged oocytes have great potential to beused for production of ESCs using parthenogeneticactivation. Therefore, to preserve these oocytes in anegg-bank, it was necessary to investigate the freezeabil-ity of aged oocytes.
4.2. Credibility of vitrified aged oocytes
Irrespective of oocytes age, vitrification significantlyreduced oocyte survival and developmental compe-tence after IVF in compared with their unvitrified coun-terparts. This finding agrees with previous studies withhuman and non-human oocytes, and has been related tocryoassociated changes occurred in vitrified oocytes[35]. Despite the importance of ICSI technique to over-come cryoassociated ZP hardening, blastocyst yield ofvitrified-warmed oocytes was not affected by this tech-nique which is in accordance with the results of Rho, etal. (2004) [36]. Interestingly, yield of blastocyst invitrified-warmed oocytes submitted to ICSI or IVF wassimilar, irrespective of post-warming interval. There-fore, it seems that vitrification and aging result in wide
heep oocytes submitted to parthenogenetic activation immediately
erated (%) Cleavage (%) Blastocysts (%) Hatching (%)
� 1.1 ab 85.3 � 1.7 a 32.5 � 1.6 b 31.4 � 2.7� 0.3 ab 81.8 � 1.2 a 14.5 � 0.5 a 14.2 � 8.2� 1.1 ab 92.8 � 2.5 ab 41.0 � 5.8 b 35.5 � 2.0� 3.9 b 83.1 � 3.2 a 14.5 � 1.5 a 36.6 � 16.2� 4.5 b 84.3 � 2.3 a 8.5 � 2.7 a 10.0 � 10� 0.7 a 97.9 � 0.7 b 43.6 � 2.6 b 28.3 � 5.7
heep oocytes submitted to intracytoplasmic injection immediately
(%) Cleavage (%) Blastocysts (%) Hatching (%)
76.0 � 13.0 3.3 � 3.3 a 00.0 a71.6 � 5.8 3.0 � 3.0 a 00.0 a74.0 � 6.8 29.0 � 3.0 b 33.3 � 16.6 b63.9 � 4.6 2.7 � 2.7 a 00.0 a65.7 � 10.1 1.3 � 1.3 a 00.0 a
ce of s
Degen
9.19.48.3
16.015.9
ce of s
erated
.6 � 1.8
.4 � 5.0
.1 � 3.3
.6 � 3.8
.6 � 2.8
77.2 � 8.7 3.5 � 3.5 a 00.0 a
ti
1752 S.M. Hosseini et al. / Theriogenology 77 (2012) 1741–1753
cellular changes, among them, only ZP-hardening canbe overcame via ICSI.
Importantly, post-warming interval differently af-fected the pattern of spindle and chromosome abnor-malities in both young and aged oocytes, so that im-mediately after warming, absent spindle (65.6%) andAII (6.2%) were the major patterns of abnormalities invitrified-young oocytes which were significantly differ-ent from aged oocytes (30.7% and 26.9%, respec-tively). Also, most abnormalities in the rested vitrified-young oocytes were AII (58.6%) and PN (13.7%)which were slightly different in vitrified-aged oocytes(53.1 and 18.7%, respectively). These findings implythat vitrification-warming leads to spindle depolymer-ization which can be then repolymerized within 2 h; thefinding that has been previously reported by the otherstudies [12,37,38]. However, it was observed that therest time enables oocytes to resume meiosis and prog-ress toward AII and even PN formation. The dynamismof meiosis progression in the aged and young restedoocytes (Fig. 2B) indicates that although most vitrifiedyoung and aged oocytes could restore microtubules duringrest time, the speed of progression to AII in vitrified-young oocytes was higher than aged oocytes during thislimited time-window. Based on the structural, ultrastruc-tural and molecular observations, aging and vitrificationinduced spontaneous activation and probably improperembryo development after ICSI in vitrified young, agedand unvitrified-aged oocytes was related to the asynchro-nous development of pronuclei in ICSI-embryos [33,39].Interestingly, this study demonstrates that although vitri-fication-warming process induced spontaneous activation,it is not possibly strong enough to derive a complete roundof early embryonic development [7]. Therefore, the rela-tively low blastocyst rate of ICSI-activated vitrified-warmed and aged oocyte could be related to the inappro-priate activation stimulus and probable harmful effect ofICSI procedure [40].
The activation time-point also differently affected par-henogenetic development of young and aged oocytes;mmediate activation post warming resulted in higher
blastocyst development of vitrified young in comparedwith aged oocytes, whereas delayed activation resultedin similar blastocyst rates in both vitrified young andaged oocytes. These findings suggest that although vit-rified-young oocytes had higher developmental poten-tial than aged oocytes, delayed activation results inirreversible changes in nuclear status of young oocyteswhich are probably detrimental for parthenogenetic em-bryo development. Moreover, gene expression pattern
of vitrified oocyte indicates that only young oocytes areable restore MPF activity within 2 h. Therefore, whilesharp drop of MPF in vitrified young oocytes signalsmeiosis resumption and nuclear progression toward TIIand PN, the recovered MPF inhibits this progressionand possibly leading to severe chromosome abnormal-ities. Considering the molecular, cellular and develop-mental characteristics of vitrified and unvitrifiedoocytes from two different age categories, and at twotime-points post warming, one may conclude that vit-rified young oocytes endure premature aging during ashort time post warming. Although this problem ismarginal in vitrified aged oocytes, it seems that oocytesdo not require to be rested post warming.
4.3. Conclusion
The present study demonstrated that in vitro agedoocytes have reasonable capacity to be used for parthe-nogenetic studies however not for IVF and ICSI. Fur-thermore, it was determined that the vitrified oocytesshowed a time-dependent decline in quality and devel-opmental potential. Accordingly, the vitrified-youngoocytes had higher speed quality decline suggestingthat there is no requirement for further rest post warm-ing. The results of the present study may be valuable inpreserving in vitro aged oocytes to provide egg-bankfor research studies.
Acknowledgments
This study was funded by a grant from Royan Instituteof IRI. The authors would like to thank Mansouri andJafarpour for statistical analysis. The authors are alsograteful to Heidari and Khajeh for ovaries preparation.
References
[1] Miao Y, Kikuchi K, Sun QY, Schatten H. Oocyte aging: cellularand molecular changes, developmental potential and reversalpossibility. Hum Reprod Update 2009;1:1–13.
[2] Tatone C, Di Emidio G, Barbaro R, Vento M, Ciriminna R,Artini PG. Effects of reproductive aging and postovulatoryaging on the maintenance of biological competence after oocytevitrification: insights from the mouse model. Theriogenology2011;76:864–73.
[3] Hall VJ, Compton D, Stojkovic P, Nesbitt M, Herbert M,Murdoch A, et al. Developmental competence of human in vitroaged oocytes as host cells for nuclear transfer. Hum Reprod2007;22:52–62.
[4] Ono T, Mizutani E, Li C, Yamagata K, Wakayama T. Offspringfrom intracytoplasmic sperm injection of aged mouse oocytestreated with caffeine or MG132. Genesis 2011;49:460–71.
[5] Kikuchi K, Naito K, Noguchi J, Kaneko H, Tojo H. Maturation/M-phase promoting factor regulates aging of porcine oocytes
matured in vitro. Cloning Stem Cells 2002;4:211–22.
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
1753S.M. Hosseini et al. / Theriogenology 77 (2012) 1741–1753
[6] Ghetler Y, Yavin S, Shalgi R, Arav A. The effect of chilling onmembrane lipid phase transition in human oocytes and zygotes.Hum Reprod 2005;20:3385–9.
[7] Asgari V, Hosseini SM, Ostadhosseini S, Hajian M, Nasr-Esfahani MH. Time dependent effect of post warming intervalon microtubule organization, meiotic status, and parthenoge-netic activation of vitrified in vitro matured sheep oocytes.Theriogenology 2011;75:904–10.
[8] Serhal PF, Ranieri DM. kinis A, Marchant S, Davies M, Kha-dum IM. Oocyte morphology predicts outcome of intracytoplas-mic sperm injection. Hum Reprod 1997;12:1267–70.
[9] Jain JK, Paulson RJ. Oocyte cryopreservation. Fertil Steril2006;86:1037–46.
10] Agarwal AC, Nagy ZP, Agarwal A. Current trends, biologicalfoundations and future prospects of oocyte and embryo cryo-preservation. Reprod Biomed Online 2009;19:126–40.
11] Cutting R, Barlow S, Anderson R. Human oocyte cryopreser-vation: evidence for practice. Hum Fertil 2009;12:125–36.
12] Chen SU, Lien YR, Chao KH, Ho HN, Yang YS, Lee TY.Effects of cryopreservation on meiotic spindles of oocytes andits dynamics after thawing: clinical implications in oocyte freez-ing–a review article. Mol Cell Endocrinol 2003;202:101–7.
13] Chen SU, Yang YS. Slow freezing or vitrification of oocytes:their effects on survival and meiotic spindles, and the timeschedule for clinical practice. Taiwan. J Obstet Gynaecol 2009;48:15–22.
14] Bromfield JJ, Coticchio G, Hutt K, Sciajno R, Borini A, Alber-tini DF. Meiotic spindle dynamics in human oocytes followingslow-cooling cryopreservation. Hum Reprod 2009;24:2114–23.
15] Igarashi H, Takahashi T, Takahashi E, Tezuka N, Nakahara K,Takahashi K, et al. Aged mouse oocytes fail to readjust intra-cellular adenosine triphosphates at fertilization. Biol Reprod2005;72:1256–61.
16] Succu S, Leoni GG, Bebbere D, Berlinguer F, Mossa F,Bogliolo L, et al. Vitrification devices affect structural andmolecular status of in vitro matured ovine oocytes. Mol ReprodDev 2007;74:1337–44.
17] Takahashi K, Matsui H, Takahashi I, Matsumoto H, Fukui E,Mitsuhiro M, et al. Effect of in vitro aging of mouse oocytes onmetaphase II spindle morphology, in vitro fertilization and sub-sequent embryonic development. J Mamma Ova Res 2010;27:42–50.
18] Bogliolo L, Ariu F, Fois S, Rosati I, Zedda MT, Leoni G, et al.Morphological and biochemical analysis of immature ovineoocytes vitrified with or without cumulus cells. Theriogenology2007;68:1138–49.
19] Morató R, Izquierdo D, Paramio MT, Mogas T. Embryo devel-opment and structural analysis of in vitro matured bovineoocytes vitrified in flexipet denuding pipettes. Theriogenology2008;70:1536–43.
20] Succu S, Bebbere D, Bogliolo L, Ariu F, Fois S, Leoni GG, etal. Vitrification of in vitro matured ovine oocytes affects in vitropre-implantation development and mRNA abundance. Mol Re-prod Dev 2008;75:538–46.
21] Nasr-Esfahani MH, Hosseini SM, Hajian M, Forouzanfar M,Ostadhosseini S, Abedi P, et al. Development of an optimizedzona-free method of somatic cell nuclear transfer in the goat.Cell Reprog 2011;13:157–70.
22] Tian SJ, Yan CL, Yang HX, Zhou GB, Yang ZQ, Zhu SE. Vitri-fication solution containing DMSO and EG can induce partheno-genetic activation of in vitro matured ovine oocytes and decrease
sperm penetration. Anim Reprod Sci 2007;101:365–71.23] Forouzanfar M, Sharafi M, Hosseini SM, Ostadhosseini S,Hajian M, Hosseini L, et al. In vitro comparison of egg yolk–based and soybean lecithin–based extenders for cryopreserva-tion of ram semen. Theriogenology 2010;73:480–7.
24] Tervit HR, Whittingham DG, Rowson LEA. Successful culturein vitro of sheep and cattle ova. J Reprod Dev 1972;30:493–7.
25] Hosseini SM, Hajian M, Moulavi F, Shahverdi AH, Nasr-Esfa-hani MH. Optimized combined electrical-chemical parthenoge-netic activation for in vitro matured bovine oocytes. AnimReprod Sci 2008;108:122–33.
26] Shadanloo F, Najafi MH, Hosseini SM, Hajian M, ForouzanfarM, Ghaedi K, et al. Sperm status and DNA dose play key rolesin sperm/ICSI-mediated gene transfer in caprine. Mol ReprodDev 2010;77:868–75.
27] Yin H. Spindles, mitochondria and redox potential in agingoocytes. Reprod Biomed Online 2003;8:45–58.
28] Wilding M, Dale B, Marino M, di Matteo L, Alviggi C, PisaturoML, et al. Mitochondrial aggregation patterns and activity inhuman oocytes and preimplantation embryos. Hum Reprod2001;16:909–17.
29] Ye XF, Chen SB, Wang LQ, Zhao YC, et al. Caffeine anddithiothreitol delay ovine oocyte ageing. Reprod Fertil Dev2010;22:1254–61.
30] Fissore RA, Kurokawa M, Knott J, Zhang M, Smyth J. Mech-anisms underlying oocyte activation and postovulatory ageing.Reproduction 2002;124:745–54.
31] Kikuchi K, Naito K, Noguchi J, Shimada A, Kaneko H, YamashitaM, et al. Maturation/M-phase promoting factor: a regulator ofaging in porcine oocytes. Biol Reprod 2000;63:715–22.
32] Larman MG, Sheehan CB, Gardner DK. Calcium-free vitrifica-tion reduces cryoprotectant-induced zona pellucida hardeningand increases fertilization rates in mouse oocytes. Reproduction2006;131:53–61.
33] Nasr-Esfahani MH, Razavi S, Mardani M, Shirazi R, Javan-mardi S. Effects of failed oocyte activation and sperm prota-mine deficiency on fertilization post-ICSI. Reprod Biomed On-line 2007;14:422–9.
34] Bianchi E, Sette C. Post-transcriptional control of Gene expres-sion in mouse early embryo development: A view from the tipof the Iceberg. Genes 2011;2:345–59.
35] Bernard A, Fuller BJ. Cryopreservation of human oocytes: areview of current problems and perspectives. Hum Reprod Up-date 1996;2:193–207.
36] Rho GJ, Lee SL, Kim YS, Yeo HJ, Ock SA, BalasubramanianS, et al. Intracytoplasmic sperm injection of frozen-thawedbovine oocytes and subsequent embryo development. Mol Re-prod Dev 2004;68:449–55.
37] Aigner S, Van der Elst J, Siebzehnrübl E, Wildt L, Lang N, VanSteirteghem AC. The influence of slow and ultra-rapid freezingon the organization of the meiotic spindle of the mouse oocyte.Hum Reprod 1992;7:857–64.
38] Gook DA, Osborn SM, Johnston WI. Cryopreservation ofmouse and human oocytes using 1,2-propanediol and the con-figuration of the meiotic spindle. Hum Reprod 1993;8:1101–9.
39] Tesarik J. Oocyte activation after intracytoplasmic injection ofmature and immature sperm cells. Hum Reprod 1998;13:117–27.
40] Shirazi A, Ostad-Hosseini S, Ahmadi E, Heidari B, Shams-Esfandabadi N. In vitro developmental competence of ICSI-derived activated ovine embryos. Theriogenology 2009;71:
342–8.Copyright © 2022 FDOKUMEN




















