
Influence of vitrification on mouse metaphase II oocyte spindle dynamics and chromatin alignment
9
Influence of vitrification on mouse metaphase II oocyte spindle dynamics and chromatin alignment Claudia Messias Gomes, M.D., a Cristine Ane Silva E. Silva, M.S., b Nicole Acevedo, Ph.D., c Edmund Baracat, M.D., Ph.D., a Paulo Serafini, M.D., d and Gary D. Smith, Ph.D. b,c,e,f a Department of Obstetrics and Gynecology, University of Sao Paulo, Sao Paulo, Brazil; Departments of b Obstetrics and Gynecology; and c Molecular and Integrated Physiology, University of Michigan, Ann Arbor, Michigan; d Huntington Center for Reproductive Medicine of Brazil, Sao Paulo, Brazil; e Department of Urology; and f Reproductive Sciences Program, University of Michigan, Ann Arbor, Michigan Objective: To evaluate influences of vitrification and warming of metaphase II (MII) mouse oocytes on survival, spindle dynamics, spindle morphology, and chromatin alignment on metaphase plates. Design: Experimental animal study. Setting: University animal laboratory. Animal(s): Eight-week-old B6D2F1 mice. Intervention(s): Denuded MII oocytes were used fresh (control), exposed to vitrification/warming solutions (Sol Expos), or vitrified and warmed (Vitr). Main Outcome Measure(s): Oocyte recovery and survival after warming and the influence of solution exposure and cryopreservation on spindle dynamics and chromatin alignment. Result(s): Cryopreservation of two or 10 oocytes per straw resulted in recovery (100% 0% and 95% 4%, re- spectively; mean SE) and survival (95% 2% and 98% 2%, respectively). Immediately after warming (Vitr), significantly fewer oocytes assessed with immunocytochemistry contained spindles, compared with control and Sol Expos. When oocytes were placed into a 37 C environment for 2 hours after exposure or warming, the ability to recognize spindles by immunocytochemistry was not significantly different between groups. Using live-cell time-lapse imaging with LC-Polscope, similar time-dependent spindle formation dynamics were observed. At 2 hours after collection or treatment, spindle morphology and length were not significantly different between the groups, nor was the incidence of aberrant alignment of chromatin on metaphase plates. Conclusion(s): Immediately after warming of vitrified MII oocytes, b-tubulin is depolymerized and chromatin re- mains condensed on the metaphase plate. Within a 2-hour period, b-tubulin repolymerizes, forming morphologi- cally normal metaphase spindles with properly aligned chromatin. (Fertil Steril Ò 2008;90:1396–404. Ó2008 by American Society for Reproductive Medicine.) Key Words: Oocyte vitrification, spindle, chromatin alignment According to 2003 Society for Assisted Reproductive Tech- nologies data, there are currently approximately 400,000 fro- zen human embryos in the United States. Routine oocyte freezing could greatly benefit assisted reproductive technol- ogy (ART) as it would provide an ethically acceptable alter- native to embryo freezing. The ovarian stimulation regimens routinely adopted for IVF remain relatively imprecise as the number of follicles that develop in response to these treat- ments varies greatly between individuals. As it is routine practice to collect oocytes from all stimulated follicles, these procedures may result in the recovery of large numbers of oo- cytes. The common clinical practice in the United States is to fertilize all the oocytes and select only two or three of the best-quality embryos for immediate transfer. The majority of the remaining embryos are frozen for subsequent frozen ET cycles. The ability to store oocytes rather than embryos from controlled ovarian stimulation would allow greater flex- ibility in the treatment of the oocytes and offer the possibility of establishing an oocyte banking system in the near future. Stored oocytes could potentially be used for several purposes: improving the efficiency of IVF, as an alternative to embryo freezing, oocyte preservation for patients with ovarian hyper- stimulation syndrome, oocyte donation programs, treatment of congenital infertility disorders, prevention of fertility loss postoperative, and treatment of premature ovarian fail- ure. Oocyte freezing could also be used to conserve fertility in women diagnosed with cancer who are too young to have either started or completed their families. Oocyte freez- ing therefore has the potential to be an important adjunct to ART in humans. Metaphase oocytes are collected as part of an existing practice in assisted reproduction. At this stage in develop- ment, oocytes have undergone nuclear and have begun cyto- plasmic maturation, the first polar body has been expelled, and the chromosome chromatins are condensed and arranged on the metaphase II (MII) spindle. A spindle apparatus is Received June 27, 2007; revised and accepted August 9, 2007. Gary D. Smith is a member of the Scientific Advisory Board for Irvine Scientific. Supported by National Institutes of Health Training Grant in Reproductive Sciences no. T32-HD07048 (Douglas Foster, to NA). Reprint requests: Gary D. Smith, Ph.D., 6428 Medical Sciences Building I, 1301 East Catherine Street, Ann Arbor, MI 48109-0617 (FAX: 734-936-8617; E-mail: [email protected]). Fertility and Sterility â Vol. 90, Suppl 2, October 2008 0015-0282/08/$34.00 Copyright ª2008 American Society for Reproductive Medicine, Published by Elsevier Inc. doi:10.1016/j.fertnstert.2007.08.025 1396
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Influence of vitrification on mouse metaphase II oocyte spindle dynamics and chromatin alignment

139
Influence of vitrification on mouse metaphase II oocytespindle dynamics and chromatin alignmentClaudia Messias Gomes, M.D.,a Cristine Ane Silva E. Silva, M.S.,b Nicole Acevedo, Ph.D.,c
Edmund Baracat, M.D., Ph.D.,a Paulo Serafini, M.D.,d and Gary D. Smith, Ph.D.b,c,e,f
a Department of Obstetrics and Gynecology, University of Sao Paulo, Sao Paulo, Brazil; Departments of b Obstetrics and
Gynecology; and c Molecular and Integrated Physiology, University of Michigan, Ann Arbor, Michigan; d Huntington Center
for Reproductive Medicine of Brazil, Sao Paulo, Brazil; e Department of Urology; and f Reproductive Sciences Program,
University of Michigan, Ann Arbor, Michigan
Objective: To evaluate influences of vitrification and warming of metaphase II (MII) mouse oocytes on survival,spindle dynamics, spindle morphology, and chromatin alignment on metaphase plates.Design: Experimental animal study.Setting: University animal laboratory.Animal(s): Eight-week-old B6D2F1 mice.Intervention(s): Denuded MII oocytes were used fresh (control), exposed to vitrification/warming solutions(Sol Expos), or vitrified and warmed (Vitr).Main Outcome Measure(s): Oocyte recovery and survival after warming and the influence of solution exposureand cryopreservation on spindle dynamics and chromatin alignment.Result(s): Cryopreservation of two or 10 oocytes per straw resulted in recovery (100% � 0% and 95% � 4%, re-spectively; mean� SE) and survival (95%� 2% and 98%� 2%, respectively). Immediately after warming (Vitr),significantly fewer oocytes assessed with immunocytochemistry contained spindles, compared with control andSol Expos. When oocytes were placed into a 37�C environment for 2 hours after exposure or warming, the abilityto recognize spindles by immunocytochemistry was not significantly different between groups. Using live-celltime-lapse imaging with LC-Polscope, similar time-dependent spindle formation dynamics were observed. At 2hours after collection or treatment, spindle morphology and length were not significantly different between thegroups, nor was the incidence of aberrant alignment of chromatin on metaphase plates.Conclusion(s): Immediately after warming of vitrified MII oocytes, b-tubulin is depolymerized and chromatin re-mains condensed on the metaphase plate. Within a 2-hour period, b-tubulin repolymerizes, forming morphologi-cally normal metaphase spindles with properly aligned chromatin. (Fertil Steril� 2008;90:1396–404. �2008 byAmerican Society for Reproductive Medicine.)
Key Words: Oocyte vitrification, spindle, chromatin alignment
According to 2003 Society for Assisted Reproductive Tech-nologies data, there are currently approximately 400,000 fro-zen human embryos in the United States. Routine oocytefreezing could greatly benefit assisted reproductive technol-ogy (ART) as it would provide an ethically acceptable alter-native to embryo freezing. The ovarian stimulation regimensroutinely adopted for IVF remain relatively imprecise as thenumber of follicles that develop in response to these treat-ments varies greatly between individuals. As it is routinepractice to collect oocytes from all stimulated follicles, theseprocedures may result in the recovery of large numbers of oo-cytes. The common clinical practice in the United States is tofertilize all the oocytes and select only two or three of thebest-quality embryos for immediate transfer. The majority
Received June 27, 2007; revised and accepted August 9, 2007.
Gary D. Smith is a member of the Scientific Advisory Board for Irvine
Scientific.
Supported by National Institutes of Health Training Grant in Reproductive
Sciences no. T32-HD07048 (Douglas Foster, to NA).
Reprint requests: Gary D. Smith, Ph.D., 6428 Medical Sciences Building I,
1301 East Catherine Street, Ann Arbor, MI 48109-0617 (FAX:
734-936-8617; E-mail: [email protected]).
Fertility and Sterility� Vol. 90, Suppl 2, October 2008Copyright ª2008 American Society for Reproductive Medicine
6
of the remaining embryos are frozen for subsequent frozenET cycles. The ability to store oocytes rather than embryosfrom controlled ovarian stimulation would allow greater flex-ibility in the treatment of the oocytes and offer the possibilityof establishing an oocyte banking system in the near future.Stored oocytes could potentially be used for several purposes:improving the efficiency of IVF, as an alternative to embryofreezing, oocyte preservation for patients with ovarian hyper-stimulation syndrome, oocyte donation programs, treatmentof congenital infertility disorders, prevention of fertilityloss postoperative, and treatment of premature ovarian fail-ure. Oocyte freezing could also be used to conserve fertilityin women diagnosed with cancer who are too young tohave either started or completed their families. Oocyte freez-ing therefore has the potential to be an important adjunct toART in humans.
Metaphase oocytes are collected as part of an existingpractice in assisted reproduction. At this stage in develop-ment, oocytes have undergone nuclear and have begun cyto-plasmic maturation, the first polar body has been expelled,and the chromosome chromatins are condensed and arrangedon the metaphase II (MII) spindle. A spindle apparatus is
0015-0282/08/$34.00, Published by Elsevier Inc. doi:10.1016/j.fertnstert.2007.08.025

a dynamic conglomerate of microtubules, comprised of a-and b-tubulin and associated structural proteins, acting to co-ordinate cyto- and karyokinetic events essential for normalchromosome segregation. Microtubules persist in phases ofelongation or shortening, where subunits are added or sub-tracted from tubule ends. As oocytes transition into meta-phase, microtubules change from radial arrays to anorganized barrel-shaped bipolar structure containing a blendof dense material at either pole known as microtubule orga-nizing centers (MTOCs) (1). The main structural proteinscomprising oocyte MTOCs include g-tubulin and pericentrin(2). Numerous reports have demonstrated that exposure ofoocytes to cooling (3–5), cryoprotectants (6, 7), or the cryo-preservation process (7) can cause depolymerization and dis-organization of spindle microtubules. This event could leadto chromosomal scattering and development of aneuploidy.
Currently, there are two methods that are used to cryo-preserve mammalian oocytes: slow-rate freezing and vitrifi-cation (8). Independent of the methodology used forcryopreservation, effects on oocyte cellular functions cancompromise abilities to develop normally after the cryo-preservation process. Compromised cellular events can becollectively termed oocyte cryodamage. Documented and/or theoretical specific cellular structures and functions thatare/may be compromised by cryopreservation as well as sub-sequent effects on oocyte and embryonic developmentalcompetence have been reviewed elsewhere (9).
During cryopreservation, cells are exposed to numerousstresses, including mechanical, thermal, and chemicalstresses (10, 11), which may lead to compromised cell func-tion and cell death. In general, it has been demonstrated thatoocytes are more sensitive to cryodamage than are later em-bryonic stages (12). Slow-rate freezing attempts to controlthe biophysical properties of freezing, like cooling and warm-ing rates, in conjunction with cryoprotectants to minimizeadverse cellular events. This method allows cells to be cooledto very low temperatures while minimizing intracellular icecrystal formation and at the same time attempting to minimizethe detrimental influences of increased solute concentrationsand osmotic stress (12). Conversely, vitrification, a form ofrapid cooling, uses very high concentrations of cryoprotectantthat solidify without forming ice crystals. Vitrification can beconsidered a nonequilibrium approach to cryopreservation,which was originally developed for cryopreservation ofmammalian sperm (13) and embryos (14). In this technique,oocytes are dehydrated by brief exposure to a concentratedsolution of cryoprotectant before the samples are plunged di-rectly into liquid nitrogen. Human oocytes (15), pronuclearzygotes (16), cleavage-stage embryos (17), and blastocysts(18–20) have been successfully vitrified. Excellent reviewsof vitrification history, use, and potential advantages are avail-able (21, 22). Live births have been obtained from both slow-rate freezing and vitrification of human oocytes. However,large-scale studies with detailed observations of potential in-tracellular perturbations after human oocyte cryopreservationare impractical and have not been performed.
Fertility and Sterility�
The objectives of this study were to use the mouse MII oo-cyte model system to evaluate oocyte vitrification and warm-ing influences on oocyte survival; spindle presence, absence,dynamics, and spindle morphology; and incidence of normalalignment of chromatin on the MII spindle plate. Informationgained by such studies with animal oocyte model systemsprovides insight into intracellular structural events occurringin human oocytes after cryopreservation by vitrification.Such knowledge is critical in determining the proper han-dling of cryopreserved human oocytes in attempt to obtainsubsequent fertilization, embryo development, and the estab-lishment of healthy live births.
MATERIALS AND METHODS
Oocyte Collection, Preparation, and Treatments
Eight-week-old B6D2F1 female mice were injected IP with7.5 IU of equine chorionic gonadotropin (eCG; Sigma, St.Louis) followed by injection with 7.5 IU of hCG (Sigma)48 hours later. Oocyte-cumulus complexes were collectedin HEPES-buffered human tubal fluid (HTF; Irvine Scien-tific, Santa Ana, CA) mediumþ 0.4% synthetic serum substi-tute (H-HTF-0.4SSS; Irvine Scientific) 14 hours after hCGinjection. Cumulus cells were removed by repeated pipettingin H-HTF medium containing 80 IU/mL hyaluronidase(Sigma). Oocytes were collected from numerous females,pooled, placed into bicarbonate-buffered HTF medium con-taining 0.4% SSS in an atmosphere of 5% CO2 in air at37�C and 100% humidity, and subsequently randomly allo-cated to treatment groups.
Denuded MII oocytes were used after collection withoutfurther treatment (control), exposed to vitrification andwarming solutions without actual cryopreservation (Sol Ex-pos), or vitrified and warmed (Vitr). All solution exposuresand actual vitrification and warming was performed by oneindividual. Vitrification and warming solutions were pre-pared as described elsewhere (15, 23). For Expos and Vitr,oocytes were initially placed into a 20-mL drop of modifiedHTF (H-HTF; Irvine Scientific) þ 0.4% serum substitutesupplement (SSS; Irvine Scientific) for 1 minute beforemerging with an adjacent 20-mL drop of equilibrationsolution (ES; 7.5% [vol/vol] ethylene glycol, 7.5% [vol/vol]dimethylsulfoxide [DMSO], 20% [vol/vol] SSS inM199-Hepes). After 2 minutes, a second 20-mL drop of ESwas merged with drops containing oocytes. Two minuteslater, oocytes were removed and pipetted into a fresh 20-mLdrop of ES for 3 minutes. Oocytes were subsequently pipettedinto separate 20-mL drops of vitrification solution (VS; 15%[vol/vol] ethylene glycol, 15% [vol/vol] DMSO, 0.5 M su-crose, 20% [vol/vol] SSS in M199-H) for 5, 5, 10, and 90 sec-onds. All solution exposures for both Sol Expos and Vitr wereperformed at 22�C. For the Sol Expos group, oocytes wereimmediately transferred through warming solutions as de-scribed below for cryopreserved oocytes. During the final90 seconds in VS, oocytes to be cryopreserved were loadedinto pulled straws with VS and heat sealed at the thin end;protective metal jackets were positioned over the thin portion
1397

of the straw, heat sealed at the large end (Fig. 1), and sub-merged in liquid nitrogen. Closed pulled straws were main-tained in liquid nitrogen for 1–24 hours before warming.For warming, straws containing oocytes were rapidly trans-ferred from liquid nitrogen into a 37�C water bath for 3 sec-onds. After straws were wiped with sterile gauze, both endswere opened and the contents expelled as an �1 mL drop.Straws were rinsed with �1 mL of initial warming solution(IWS; 1.0 M sucrose, 20% [vol/vol] SSS in M199-H), whichwas expelled as a drop and merged with the straw contentdrop for 1 minute. Oocytes were transferred into a 20-mLdrop of IWS for an additional minute followed by movementthrough two 20-mL drops of dilution solution (0.5 M sucrose,20% [vol/vol] SSS in M199-H) for 2 minutes each and finallythrough three 20-mL drops of wash solution (WS; 20% [vol/vol] SSS in M199-H) for 2 minutes per drop. All solutionexposures were again performed at 22�C.
Microscopic Observations and Immunocytochemistry (ICC)
After warming or solution exposure, oocytes were evaluatedfor recovery and normal morphology as assessed with an in-verted bright-field microscope with Hoffman ModulationContrast optics (Leica DMIRB; Leica Microsystems, Wet-zlar, Germany) at�400 magnification. Oocytes were catego-rized as having survived if they contained an intact zonapellucida and plasma membrane, a refractive and relativelyhomogeneous cytoplasm, and minimal space between theplasma membrane and zona pellucida except in the positionof the first polar body.
For characterization of MII spindles and condensed chro-matin on the metaphase plate, oocytes were fixed andmounted on polylysine-coated coverslips as described else-where (24) either immediately upon removal from the finalWS drop (T0) or after 2 hours of incubation in HTF (IrvineScientific) þ 0.4% SSS (vol/vol) at 37�C in 5% CO2 andair (T2). Briefly, oocytes were fixed and permeabilized in2% (wt/vol) paraformaldehyde with 0.5% (vol/vol) Triton-X-100 for 45 minutes at 37�C. Oocytes were washed 3 timesthen blocked overnight at 4�C with a solution of PBS contain-ing 2% bovine serum albumin, 2% powdered milk, 0.1 M gly-cine, and 0.5% Triton-X-100. Oocytes were incubated witha monoclonal antibody for b-tubulin (Sigma; diluted 1:200)for 1 hour at 37�C in a humidified chamber. Nonimmunemouse serum was substituted for primary antibodies as a neg-ative control. Staining was visualized using an anti-mouseAlexa Fluor 488 (Molecular Probes, Eugene, OR; diluted1:750). Chromatin was counterstained with mounting solu-tion containing 10 mg/mL Hoescht 33342 (blue) or PropidiumIodide (Sigma; red). Fluorescence was evaluated with a LeicaDMR fluorescent microscope at �400–1000 magnification,and images were captured and processed with SPOT RTSlider digital camera and software (Diagnostic Instruments,Sterling Heights, MI). In some experiments, spindle pole-to-pole length was measured at �400 magnification andused to determine average spindle length. Importantly, spin-dle length was only evaluated in oocytes that had spindles
1398 Gomes et al. Vitrification and oocyte normalcy
positioned with poles and the metaphase plate in a perpen-dicular orientation in relation to the microscopic field ofview. Finally, a cohort of oocytes in each treatment wasevaluated for proper chromatin alignment on the metaphaseplate. Oocytes were categorized as either having normalor abnormal chromatin alignment as observed at �1000magnification.
FIGURE 1
The closed pulled straw (CPS) method for oocytevitrification. (A) The tip of the pulled straw has fourmarked lines. To load the pulled straw, VS isaspirated to the first line, and then oocytes and VSare aspirated to reach the second line; additional VSis aspirated so that the fluid meniscus reaches thethird line. The relevant characteristics of CPS are itssmall, fine, and delicate nature. To show this, CPS iscompared with the size of a penny in the figure. Thestraw is attached to a 1-mL syringe. The metal end iswhere the oocytes will be placed. (B) A CPScontaining seven fresh oocytes. (C) CPS is heatsealed just below the first mark and then above thefourth mark. At warming, CPS is taken out of theliquid nitrogen and is cut at the large end near thefourth mark.
Gomes. Vitrification and oocyte normalcy. Fertil Steril 2008.
Vol. 90, Suppl 2, October 2008

Results obtained from ICC studies on dynamics of spindlepolymerization after cryopreservation were confirmed withthe use of live-cell imaging of oocytes after vitrificationand warming. Immediately after removal from the finalWS, oocytes were placed in a 5-mL drop of H-HTF þ 0.4%SSS covered with prewarmed paraffin oil (Sigma) at 37�Cin a thermostatic system to maintain 37�C during image cap-turing. Oocytes were examined under an inverted microscope(Olympus I � 81; Olympus America, Inc., Melville, NY)equipped with an analog video camera (OlympusDP30BW) and LC-PolScope (Cambridge Research and In-strumentation, Woburn, MA) retardance imaging systemthat combines liquid crystal–based polarizing light technol-ogy and data acquisition/analysis software to display bire-fringence, an inherent optical property of highly orderedmolecules such as spindle microtubules in the oocyte. Thisnoninvasive technology has been previously employed to in-dicate the presence of birefringent spindles in human oocytesused for intracytoplasmic sperm injection (ICSI) as well as toidentify oocytes with abnormal spindles (25–28); birefrin-gent images were captured every 10 minutes for 5 secondsduring a 120-minute period of time.
Statistical Analysis
Parametric and nonparametric statistical comparisons wereperformed with Student’s unpaired t-test and c2 analysis, re-spectively. P<.05 was considered statistically significant.
RESULTS
Oocytes were vitrified either two per straw (n¼ 40) or 10 perstraw (n ¼ 1040; Fig. 2A). After warming, the recovery ratewas 100% (two per straw) and 95% � 4% (10 per straw;mean � SE) and was not significantly different between oo-cyte number groups (Fig. 2A). After warming and washingoocytes in dilution solution and WS, oocytes were assessedfor cryosurvival. Survival rates of recovered oocytes were ex-tremely high and not significantly different between two oo-cytes per straw (95%� 3%) and 10 oocytes per straw (98%�3%). MII oocytes after vitrification and warming were mor-phologically indistinguishable compared with those beforecryopreservation (Fig. 2B). At the light-microscope level, vit-rified and warmed oocytes had normal-appearing zona pellu-cidae, intact polar bodies, normal perivitellin spaces, integralplasma membranes, and normal-appearing evenly granularcytoplasm.
After vitrification and warming, oocytes were fixed andsubjected to ICC and chromatin counterstain to visualizepolymerized b-tubulin and condensed chromatin withinMII spindles and the metaphase plate, respectively. The abil-ity to recognize the spindle was inconsistent and appeared tobe related to time of fixation after warming. Thus experi-ments were performed to investigate the influence of vitri-fication/warming solution alone, cryopreservation withvitrification/warming, and time of assessment after warmingon spindle dynamics. If oocytes were fixed immediately afterremoval from the final WS (T0), 100% of Sol Expos had rec-
Fertility and Sterility�
ognizable MII spindles, whereas significantly fewer Vitr oo-cytes (21%; P<.01) had identifiable spindles at the same time(Table 1). However, if after vitrification oocytes were placedinto 37�C culture media and maintained at 37�C for 2 hours,b-tubulin repolymerized and 100% of the oocytes had MIIspindles, similar to Sol Expos oocytes at T0 and T2 (100%;P>.05; Fig. 3).
While ICC allows evaluation of many oocytes within a sin-gle treatment at a single time, it does not allow continualobservations of dynamic changes. To confirm this time-dependent influence on spindle dynamics after vitrificationand warming, time-lapse live-cell imaging at 37�C with po-larizing light was used to observe birefringence. Similar tofindings from ICC, MII oocytes after vitrification/warminghad a high degree of b-tubulin depolymerization (Fig. 4;T ¼ 0 minutes). However, over time at 37�C, b-tubulinrepolymerizes to form normal-appearing MII spindles(Fig. 4; T ¼ 120 minutes).
With b-tubulin depolymerizing during vitrification, but re-polymerizing over time after warming, the question arises,Are spindles that re-form of normal morphology? To addressthis question, MII oocytes were fixed and subjected to ICCfor b-tubulin as freshly isolated controls, after solution expo-sure only or after vitrification/warming and 2 hour culture at37�C. Pole-to-pole spindle length measures were only per-formed when pole-to-pole observations were perpendicularto the line of microscopic view (Fig. 5A). Using these strictcriteria of spindle morphology, spindle length was not signif-icantly different among control (n¼ 30; 32.5� 0.7 mm; mean� SE), Sol Expos (n ¼ 17; 31.8 � 0.6 mm), and Vitr oocytes(n ¼ 32; 32.2 � 1.2 mm; Fig. 5B).
Considering that MII spindles depolymerize and repoly-merize after vitrification/warming and culture, the subsequentalignment of chromatin on the MII plate was investigated.Using ICC for spindle identification and chromatin count-erstained with propidium iodide, the incidence of aberrantalignment of chromatin on metaphase plates was not signifi-cantly different among control (1/37; 2.7%), Sol Expos (1/47;2.1%), and Vitr (3/49; 6.1%; P¼.61) oocytes.
DISCUSSION
The incorporation of oocyte cryopreservation into the clinicalpractice of assisted reproduction has long been a goal, and re-cent advances in laboratory practices, including fertilizationwith ICSI, improved culture media, and optimization of cryo-protectants, have begun to make this a viable reality (29, 30).Vitrification is an ultrarapid cooling technique that producesa glasslike solidification of living cells that completely avoidsice crystal formation during cooling and, equally important,during warming to recover cells for biological applications(22, 23, 29). A major advantage of vitrification is eliminationof mechanical damage caused by intra- or extracellular icecrystals and potential reduction of chilling damage by short-ening the exposure to suboptimal temperatures. However, vit-rification is a result of high cooling rates associated with high
1399

FIGURE 2
(A) Recovery and survival rates of vitrified oocytes after warming. The rates are not significantly different betweenstraws with two or 10 oocytes. Recovery after warming: 100% (two oocytes per straw) and 95% � 4% (10oocytes per straw). Survival after warming: 95% � 3% (two oocytes per straw) and 98% � 3% (10 oocytes perstraw). (B) Assessment of oocyte survival using light-microscope analysis with Hoffman Modulation Contrastoptics (Leica DMIRB; Leica Microsystems) at �400 magnification.
Gomes. Vitrification and oocyte normalcy. Fertil Steril 2008.
concentrations of cryoprotectants. If used incorrectly, thesecryoprotectants can be toxic. Therefore vitrification necessi-tates reduced and precise exposure times and rapid move-ment of cells through solutions. Because of these precise
TABLE 1Identification of MII oocyte spindles after notreatment (control), vitrification/warmingsolution exposure only (Sol expos), orvitrification/warming at times T0 and T2 afterwarming.
T0a T2b
Group n % n %
Control 15 100 47 100Vitrification/warming solution
exposure only24 100 49 100
Vitrification/warming 39 21 24 100
Note: n ¼ number of oocytes assessed in each group.a T0 represents identification of MII oocyte spindles
after removal of oocytes from the final WS.b T2 represents identification of MII oocytes spindles
after 2 hours of incubation in HTF-0.4% SSS (vol/vol) at 37�C in 5% CO2 and air.
Gomes. Vitrification and oocyte normalcy. Fertil Steril 2008.
1400 Gomes et al. Vitrification and oocyte normalcy
cell-handling requirements, this technique has been labeledby some as technically difficult (22, 29). Because of the pre-cise nature of vitrification, it does require practice and cancarry a technical fingerprint that is dependent on the experi-ence of the technician. With practice, very high recovery andsurvival rates after oocyte vitrification and warming can beobtained (22, 29).
The recovery rate is the number of remaining oocytes afterexpulsion of solution from the container during warming. Itsanalysis is important because cells must tolerate a sequenceof volumetric contractions and expansions because of dif-ferent concentrations of solutions, and they can be lost orrupture during this process. Moreover, this rate can beinfluenced by the number of cells per straw as increased num-bers enhance the time required for movement of cells throughsolutions, making overexposure or loss of cells possible be-cause of time constraints. Because of the precise attentionto timing required for vitrification, the cryopreservation ofmore cells per container equates to a need for experienceand practice. Thus, vitrification could be considered to beless forgiving of technical flaws compared with other cryo-preservation techniques. Consideration of these requirementsleads one to conclude that the technique is challenging. How-ever, with practice, the difficulty is greatly reduced. Survivalrates, as determined by oocyte morphological integrity, weresimilar in this study to those cited in other mouse oocyte vit-rification experiments (31–33) and better than those related to
Vol. 90, Suppl 2, October 2008
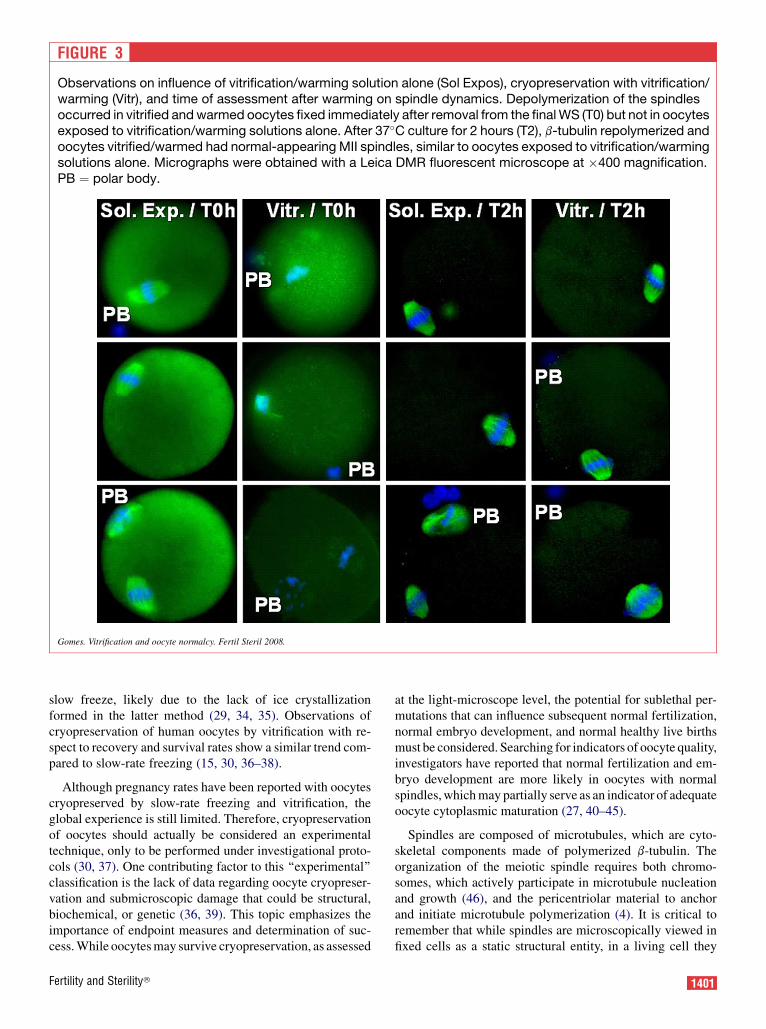
FIGURE 3
Observations on influence of vitrification/warming solution alone (Sol Expos), cryopreservation with vitrification/warming (Vitr), and time of assessment after warming on spindle dynamics. Depolymerization of the spindlesoccurred in vitrified and warmed oocytes fixed immediately after removal from the final WS (T0) but not in oocytesexposed to vitrification/warming solutions alone. After 37�C culture for 2 hours (T2), b-tubulin repolymerized andoocytes vitrified/warmed had normal-appearing MII spindles, similar to oocytes exposed to vitrification/warmingsolutions alone. Micrographs were obtained with a Leica DMR fluorescent microscope at �400 magnification.PB ¼ polar body.
Gomes. Vitrification and oocyte normalcy. Fertil Steril 2008.
slow freeze, likely due to the lack of ice crystallizationformed in the latter method (29, 34, 35). Observations ofcryopreservation of human oocytes by vitrification with re-spect to recovery and survival rates show a similar trend com-pared to slow-rate freezing (15, 30, 36–38).
Although pregnancy rates have been reported with oocytescryopreserved by slow-rate freezing and vitrification, theglobal experience is still limited. Therefore, cryopreservationof oocytes should actually be considered an experimentaltechnique, only to be performed under investigational proto-cols (30, 37). One contributing factor to this ‘‘experimental’’classification is the lack of data regarding oocyte cryopreser-vation and submicroscopic damage that could be structural,biochemical, or genetic (36, 39). This topic emphasizes theimportance of endpoint measures and determination of suc-cess. While oocytes may survive cryopreservation, as assessed
Fertility and Sterility�
at the light-microscope level, the potential for sublethal per-mutations that can influence subsequent normal fertilization,normal embryo development, and normal healthy live birthsmust be considered. Searching for indicators of oocyte quality,investigators have reported that normal fertilization and em-bryo development are more likely in oocytes with normalspindles, which may partially serve as an indicator of adequateoocyte cytoplasmic maturation (27, 40–45).
Spindles are composed of microtubules, which are cyto-skeletal components made of polymerized b-tubulin. Theorganization of the meiotic spindle requires both chromo-somes, which actively participate in microtubule nucleationand growth (46), and the pericentriolar material to anchorand initiate microtubule polymerization (4). It is critical toremember that while spindles are microscopically viewed infixed cells as a static structural entity, in a living cell they
1401
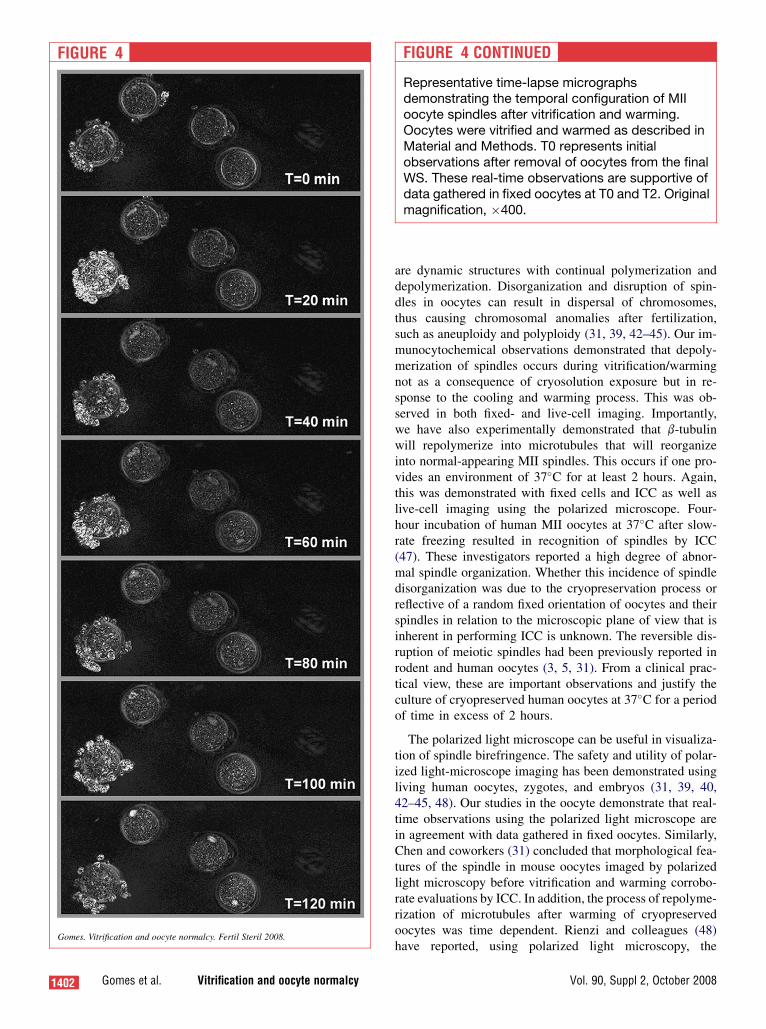
FIGURE 4
Gomes. Vitrification and oocyte normalcy. Fertil Steril 2008.
1402 Gomes et al. Vitrification and oocyte normalcy
are dynamic structures with continual polymerization anddepolymerization. Disorganization and disruption of spin-dles in oocytes can result in dispersal of chromosomes,thus causing chromosomal anomalies after fertilization,such as aneuploidy and polyploidy (31, 39, 42–45). Our im-munocytochemical observations demonstrated that depoly-merization of spindles occurs during vitrification/warmingnot as a consequence of cryosolution exposure but in re-sponse to the cooling and warming process. This was ob-served in both fixed- and live-cell imaging. Importantly,we have also experimentally demonstrated that b-tubulinwill repolymerize into microtubules that will reorganizeinto normal-appearing MII spindles. This occurs if one pro-vides an environment of 37�C for at least 2 hours. Again,this was demonstrated with fixed cells and ICC as well aslive-cell imaging using the polarized microscope. Four-hour incubation of human MII oocytes at 37�C after slow-rate freezing resulted in recognition of spindles by ICC(47). These investigators reported a high degree of abnor-mal spindle organization. Whether this incidence of spindledisorganization was due to the cryopreservation process orreflective of a random fixed orientation of oocytes and theirspindles in relation to the microscopic plane of view that isinherent in performing ICC is unknown. The reversible dis-ruption of meiotic spindles had been previously reported inrodent and human oocytes (3, 5, 31). From a clinical prac-tical view, these are important observations and justify theculture of cryopreserved human oocytes at 37�C for a periodof time in excess of 2 hours.
The polarized light microscope can be useful in visualiza-tion of spindle birefringence. The safety and utility of polar-ized light-microscope imaging has been demonstrated usingliving human oocytes, zygotes, and embryos (31, 39, 40,42–45, 48). Our studies in the oocyte demonstrate that real-time observations using the polarized light microscope arein agreement with data gathered in fixed oocytes. Similarly,Chen and coworkers (31) concluded that morphological fea-tures of the spindle in mouse oocytes imaged by polarizedlight microscopy before vitrification and warming corrobo-rate evaluations by ICC. In addition, the process of repolyme-rization of microtubules after warming of cryopreservedoocytes was time dependent. Rienzi and colleagues (48)have reported, using polarized light microscopy, the
FIGURE 4 CONTINUED
Representative time-lapse micrographsdemonstrating the temporal configuration of MIIoocyte spindles after vitrification and warming.Oocytes were vitrified and warmed as described inMaterial and Methods. T0 represents initialobservations after removal of oocytes from the finalWS. These real-time observations are supportive ofdata gathered in fixed oocytes at T0 and T2. Originalmagnification, �400.
Vol. 90, Suppl 2, October 2008
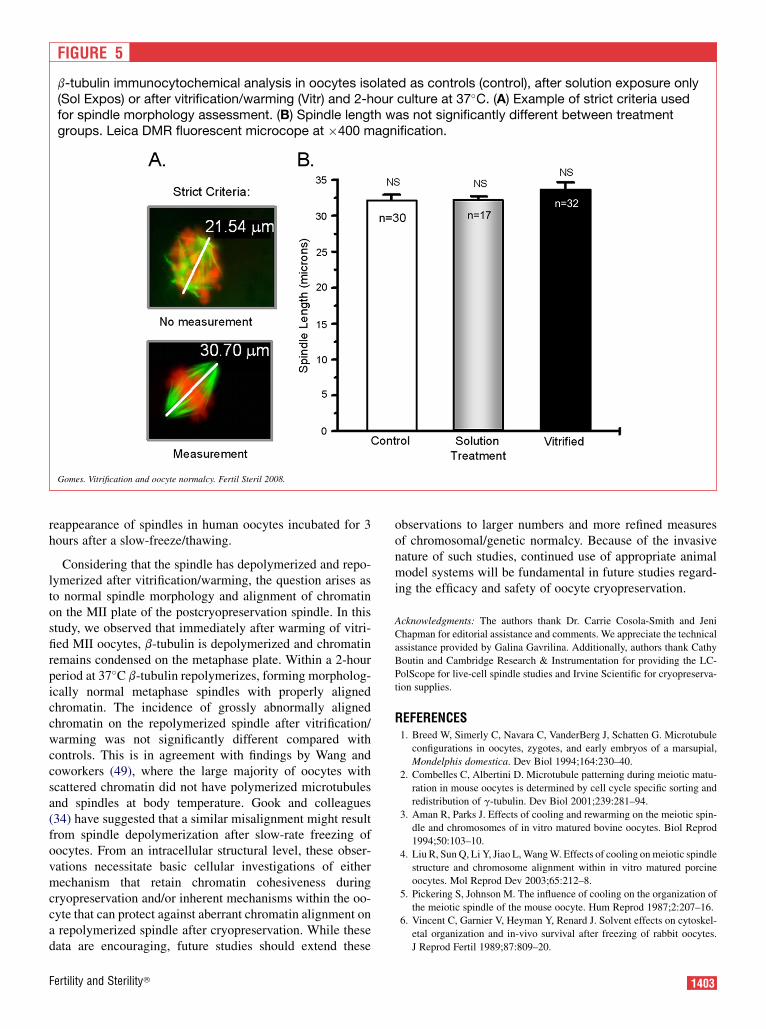
FIGURE 5
b-tubulin immunocytochemical analysis in oocytes isolated as controls (control), after solution exposure only(Sol Expos) or after vitrification/warming (Vitr) and 2-hour culture at 37�C. (A) Example of strict criteria usedfor spindle morphology assessment. (B) Spindle length was not significantly different between treatmentgroups. Leica DMR fluorescent microcope at �400 magnification.
Gomes. Vitrification and oocyte normalcy. Fertil Steril 2008.
reappearance of spindles in human oocytes incubated for 3hours after a slow-freeze/thawing.
Considering that the spindle has depolymerized and repo-lymerized after vitrification/warming, the question arises asto normal spindle morphology and alignment of chromatinon the MII plate of the postcryopreservation spindle. In thisstudy, we observed that immediately after warming of vitri-fied MII oocytes, b-tubulin is depolymerized and chromatinremains condensed on the metaphase plate. Within a 2-hourperiod at 37�C b-tubulin repolymerizes, forming morpholog-ically normal metaphase spindles with properly alignedchromatin. The incidence of grossly abnormally alignedchromatin on the repolymerized spindle after vitrification/warming was not significantly different compared withcontrols. This is in agreement with findings by Wang andcoworkers (49), where the large majority of oocytes withscattered chromatin did not have polymerized microtubulesand spindles at body temperature. Gook and colleagues(34) have suggested that a similar misalignment might resultfrom spindle depolymerization after slow-rate freezing ofoocytes. From an intracellular structural level, these obser-vations necessitate basic cellular investigations of eithermechanism that retain chromatin cohesiveness duringcryopreservation and/or inherent mechanisms within the oo-cyte that can protect against aberrant chromatin alignment ona repolymerized spindle after cryopreservation. While thesedata are encouraging, future studies should extend these
Fertility and Sterility�
observations to larger numbers and more refined measuresof chromosomal/genetic normalcy. Because of the invasivenature of such studies, continued use of appropriate animalmodel systems will be fundamental in future studies regard-ing the efficacy and safety of oocyte cryopreservation.
Acknowledgments: The authors thank Dr. Carrie Cosola-Smith and Jeni
Chapman for editorial assistance and comments. We appreciate the technical
assistance provided by Galina Gavrilina. Additionally, authors thank Cathy
Boutin and Cambridge Research & Instrumentation for providing the LC-
PolScope for live-cell spindle studies and Irvine Scientific for cryopreserva-
tion supplies.
REFERENCES1. Breed W, Simerly C, Navara C, VanderBerg J, Schatten G. Microtubule
configurations in oocytes, zygotes, and early embryos of a marsupial,
Mondelphis domestica. Dev Biol 1994;164:230–40.
2. Combelles C, Albertini D. Microtubule patterning during meiotic matu-
ration in mouse oocytes is determined by cell cycle specific sorting and
redistribution of g-tubulin. Dev Biol 2001;239:281–94.
3. Aman R, Parks J. Effects of cooling and rewarming on the meiotic spin-
dle and chromosomes of in vitro matured bovine oocytes. Biol Reprod
1994;50:103–10.
4. Liu R, Sun Q, Li Y, Jiao L, Wang W. Effects of cooling on meiotic spindle
structure and chromosome alignment within in vitro matured porcine
oocytes. Mol Reprod Dev 2003;65:212–8.
5. Pickering S, Johnson M. The influence of cooling on the organization of
the meiotic spindle of the mouse oocyte. Hum Reprod 1987;2:207–16.
6. Vincent C, Garnier V, Heyman Y, Renard J. Solvent effects on cytoskel-
etal organization and in-vivo survival after freezing of rabbit oocytes.
J Reprod Fertil 1989;87:809–20.
1403

7. Aigner S, Van der Elst J, Siebzehnrubl E, Wildt L, Lang N, Van
Steirteghem A. The influence of slow and ultra-rapid freezing on the or-
ganization of the meiotic spindle of the mouse oocyte. Hum Reprod
1992;7:857–64.
8. Bernard A, Fuller B. Cryopreservation of human oocytes: a review of
current problems and perspectives. Hum Reprod Update 1996;2:
193–207.
9. Smith G, Silva E, Silva C. Developmental consequences of cryopreser-
vation of mammalian oocytes and embryos. Reprod Biomed Online
2004;9:171–8.
10. Meryman H. Cryoprotective Agents. Cryobiology 1971;8:173–83.
11. Mazur P, Leibo S, Chu E. A two-factor hypothesis of freezing injury: ev-
idence from Chinese hamster tissue-culture cells. Exp Cell Res 1972;71:
345–55.
12. Friedler S, Giudice L, Lamb E. Cryopreservation of embryos and ova.
Fertil Steril 1988;49:743–64.
13. Polge E, Smith A, Parkes A. Revival of spermatozoa after vitrification
and dehydration at low temperatures. Nature 1949;164:666.
14. Rall W, Fahy G. Ice-free cryopreservation of mouse embryos at�196�Cby vitrification. Nature 1985;313:573–5.
15. Kuwayama M, Vajta G, Kato O, Leibo S. Highly efficient vitrification
method for cryopreservation of human oocytes. Reprod BioMed Online
2005;11:300–8.
16. Isachenko V, Montag M, Isachenko E, Van der Ven H. Vitrification of
mouse pronuclear embryos after polar body biopsy without direct contact
with liquid nitrogen. Fertil Steril 2005;84:1011–6.
17. El-Danasouri I, Selman H. Successful pregnancies and deliveries after
a simple vitrification protocol for day 3 human embryos. Fertil Steril
2001;76:400–2.
18. Liebermann J, Tucker M, Graham J, Han T, Davis A, Levy M. Blastocyst
development after vitrification of multipronuclear zygotes using Flexipet
denuding pipette. Reprod Biomed Online 2002;4:146–50.
19. Lane M, Schoolcraft W, Gardner D. Vitrification of mouse and human
blastocysts using a novel cryo-loop container-less technique. Fertil Steril
1999;72:1073–8.
20. Mukaida T, Nakamura S, Tomiyama T, Wada S, Oka C, Kasai M, et al.
Vitrification of human blastocysts using cryoloops: clinical outcome of
223 cycles. Hum Reprod 2003;18:384–91.
21. Kuleshova L, MacFarlane D, Trounson A, Shaw J. Sugars exert a major
influence on the vitrification properties of ethylene glycol-based
solutions and have low toxicity to embryos and oocytes. Cryobiology
1999;38:119–30.
22. Liebermann J, Nawroth F, Isachenko V, Isachenko E, Rahimi G,
Tucker M. Potential importance of vitrification in reproductive medicine.
Biol Reprod 2002;67:1671–80.
23. Tachikawa S, Otoi T, Kondo S, Machida T, Kasai M. Successful vitrifi-
cation of bovine blastocysts, derived by in vitro maturation and fertiliza-
tion. Mol Reprod Dev 1993;34:266–71.
24. Lu Q, Smith G, Chen D, Yang Z, Han Z, Schatten H, et al. Phosphoryla-
tion of mitogen-activated protein kinase is regulated by protein kinase C,
cyclic 30, 50- adenosine monophosphate, and protein phosphatase modu-
lators during meiosis resumption in rat oocytes. Biol Reprod 2001;64:
1444–50.
25. Sun XF, Wang WH, Keefe DL. Overheating is detrimental to meiotic
spindles within in vitro matured human oocytes. Zygote 2004;12:65–70.
26. Wang WH, Keefe DL. Prediction of chromosome misalignment among
in vitro matured human oocytes by spindle imaging with the PolScope.
Fertil Steril 2002;78:1077–81.
27. Wang WH, Meng L, Hackett RJ, Keefe DL. Developmental ability of hu-
man oocytes with or without birefringent spindles imaged by Polscope
before insemination. Hum Reprod 2001;16:1464–8.
28. Wang WH, Meng L, Hackett RJ, Odenbourg R, Keefe DL. The spindle
observation and its relationship with fertilization after intracytoplasmic
sperm injection in living human oocytes. Fertil Steril 2001;75:348–53.
1404 Gomes et al. Vitrification and oocyte normalcy
29. Kuleshova L, Lopata A. Vitrification can be more favorable than slow
cooling. Fertil Steril 2002;78:449–54.
30. Jain J, Paulson R. Oocyte cryopreservation. Fertil Steril 2006;86:
1037–46.
31. Chen C, Wang C, Tsai W, Hsieh L, Wang H, Soong Y. Evaluation of mei-
otic spindles in thawed oocytes after vitrification using polarized light
microscopy. Fertil Steril 2004;82:666–72.
32. Sun H, Hu Y, Chen L, Zhang N, Xu Z. Influence of two different vitrifi-
cation cryopreservation methods on spindle of mouse oocytes. Zhonghua
Nan Ke Xue 2006;12:1076–9.
33. Kim S, Ku S, Sung K, Kang M, Kim S, Kim H, et al. Simplified EM grid
vitrification is a convenient and efficient method for mouse mature
oocyte cryopreservation. Yonsei Med J 2006;47:399–404.
34. Gook D, Osborn S, Johnston W. Cryopreservation of mouse and human
oocyte using 1,2-propanodiol and the configuration of the meiotic spin-
dle. Hum Reprod 1993;8:1101–9.
35. Murayama S, Yamano S, Kobayashi T, Ishikawa H, Kunikata K, Aono T.
Successful freezing of unfertilized mouse oocytes and effect of cocul-
tures in oviducts on development of in vitro fertilized embryos after
thawing. J Assist Reprod Genet 1994;11:156–61.
36. La Sala G, Nicoli A, Villani M, Pescarini M, Gallinelli A, Blickstein I.
Outcome of 518 salvage oocyte-cryopreservation cycles performed as
a routine procedure in an in vitro fertilization program. Fertil Steril
2006;86:1423–7.
37. Oktay K, Cil A, Bang H. Efficiency of oocyte cryopreservation: a meta-
analysis. Fertil Steril 2006;86:70–80.
38. Liebermann J, Tucker M. Effect of carrier system on the yield of human
oocytes and embryos as assessed by survival and developmental potential
after vitrification. Reproduction 2002;124:483–9.
39. Coticchio G, Bonu M, Bianchi V, Flamigni C. Criteria to assess human
oocyte quality after cryopreservation. Reprod Biomed Online 2005;11:
421–7.
40. Wang W, Keefe D. Spindle observation in living mammalian oocytes
with the polarization microscope and its practical use. Cloning Stem
Cells 2002;4:269–76.
41. Wang W, Meng L, Hackett R, Odenbourg R, Keefe D. The spindle obser-
vation and its relationship with fertilization after intracytoplasmic sperm
injection in living human oocytes. Fertil Steril 2001;75:348–53.
42. Rienzi L, Ubaldi F, Iacobelli M, Minasi M, Romano S, Greco E. Meiotic
spindle visualization in living human oocytes. Reprod Biomed Online
2005;10:192–8.
43. Wang W, Meng L, Hackett R, Odenbourg R, Keefe D. Limited recovery
of meiotic spindles in living human oocytes after cooling-rewarming ob-
served using polarized light microscopy. Hum Reprod 2001;16:2374–8.
44. Konc J, Kanyo K, Cseh S. Visualization and examination of meiotic spin-
dle in human oocytes with Polscope. J Assist Reprod Genet 2004;21:
349–53.
45. Keefe D, Liu L, Wang W, Silva C. Imaging meiotic spindles by polariza-
tion light microscopy: principles and applications to IVF. Reprod
Biomed Online 2003;7:24–9.
46. Nam-Hyung K, Funahashi H, Prather R, Schatten G, Day B. Microtubule
and microfilament dynamics in porcine oocytes during meiotic matura-
tion. Mol Reprod Dev 1996;43:248–55.
47. Boiso I, Marti M, Santalo J, Ponsa M, Barri P, Veiga A. A confocal
microscopy analysis of the spindle and chromosome configurations of
human oocytes cryopreserved at the germinal vesicle and metaphase II
stage. Hum Reprod 2002;17:1885–91.
48. Rienzi L, Martinez F, Ubaldi F, Minasi M, Iacobelli M, Tesarik J, et al.
Polscope analysis of meiotic spindle changes in living metaphase II
human oocytes during freezing and thawing procedures. Hum Reprod
2004;19:655–9.
49. Wang W, Keefe D. Prediction of chromosome misalignment among in
vitro matured human oocytes by spindle imaging with the Polscope.
Fertil Steril 2002;78:1077–81.
Vol. 90, Suppl 2, October 2008




![[sp1] oocyte collection from superstimulated disease-free ...](https://static.fdokumen.com/doc/165x107/631dd3361aedb9cd850f788f/sp1-oocyte-collection-from-superstimulated-disease-free-.jpg)















