Molecular cloning of metaphase chromosome protein 1 (MCP1), a novel human autoantigen that...
11
Molecular cloning of metaphase chromosome protein 1 (MCP1), a novel human autoantigen that associates with condensed chromosomes during mitosis Elsa Bronze-da-Rocha, Jose ´ A. Catita & Claudio E. Sunkel Received 17 September 1997; received in revised form 5 November 1997; accepted for publication by A. Sumner 6 November 1997 Systemic lupus erythematosus autoantibodies were used to identify and to characterize new human chromosome-associated proteins. Previous immuno- localization studies in human and murine tissue culture cells showed that some of these monoclonal antibodies recognize nuclear antigens that associate with condensed chromosomes during mitosis. One antibody was selected for screening a human HeLa S3 cDNA expression library, and cDNAs that code for an antigen of 31–33 kDa were isolated. Immuno- logical, biochemical and cell fractionation data in- dicate that the 31- to 33-kDa antigen corresponds to the chromosome-associated protein recognized by the original monoclonal antibody. Sequence analysis shows that we isolated a novel human gene. Immunolocalization to human tissue culture cells shows that during interphase the antigen is dis- persed in the nucleus and that during mitosis it associates exclusively with condensed chromo- somes. A similar pattern of localization was also observed in mouse fibroblasts, suggesting that the antigen is conserved among different species. Final- ly, we show that part of the antigen remains bound to the scaffold/matrix component, even after high salt extraction. Key words: autoantigens, chromosome, mitosis, systemic lupus erythematosus Introduction In most higher eukaryotes, mitosis is characterized by the presence of condensed chromosomes. The condensa- tion of chromatin into discrete chromosomes is essential for the proper segregation of chromatids into the two daughter cells. The mitotic chromosome is a complex structure in which the DNA nucleosome fibre is thought to form loops that are attached to a proteinaseous component called scaffold (Jackson 1991). A number of proteins have been identified that are constituents of this structure of which topoisomerase II is the best characterized (Saitoh & Laemmli 1994). More recently the SMC ( stability of mini chromosomes) family of proteins has been described and at least some of its members are also thought to be part of the scaffold (Saitoh et al. 1995). Human autoimmune sera from patients with various diseases, including several forms of sclerodermy; calci- nosis, Raynaud syndrome, oesophageal dismotility, sclerodactyly and telangiectasia (CREST); and systemic lupus erythematosus (SLE), have been used to identify chromosomal associated proteins. A number of these proteins have been subsequently cloned (Tan et al. 1988; Tan 1989; Balczon 1993). Some of these proteins are confined to specific chromosomal locations such as the centromere (Earnshaw et al. 1987; Cooke et al., 1987; 1990; Saitoh et al. 1992; Balczon 1993) and others appear to be distributed either at the perichromosomal regions (Hernandez-Verdun & Gautier 1994) or throughout the condensed chromosomes (Earnshaw & Bernat 1991; Rattner 1992). Functional studies of many of these proteins indicated that they are essential for several aspects of chromosomal behaviour (Balczon 1993). SLE in humans and animals is characterized by the presence of high levels of serum antibodies that react with a variety of self antigens, including membrane molecules, cytoplasmic proteins and nuclear determi- nants (Tan 1989; 1991). MRL/ lpr mice develop an equivalent disease and have been used to produce autoreactive monoclonal antibodies (MAbs) (Cohen & Eisenberg 1991; Steinberg et al. 1991). The DNA-binding antibodies produced in mice with SLE have some unusual properties. They recognize at least five major groups of epitopes (Morgan et al. 1985a; Bronze-da- Rocha et al. 1992) and express cross-reactive idiotypes that are shared between humans and mice (Morgan et Chromosome Research 1998, 6, 85–95 # 1998 Rapid Science Publishers E. Bronze-da-Rocha is at the Laborato ´rio de Gene ´tica Molecular, Instituto de Biologia Molecular e Celular da Universidade do Porto, Rua do Campo Alegre 823, 4150 Porto, and the Faculdade de Farma ´cia da Universidade do Porto, Rua Anibal Cunha, 4000 Porto, Portugal. J. A. Catita is at the Laborato ´rio de Gene ´tica Molecular, Instituto de Biologia Molecular e Celular da Universidade do Porto, Rua do Campo Alegre 823, 4150 Porto. C. E. Sunkel (corresponding author) is at the Laborato ´rio de Gene ´tica Molecular, Instituto de Biologia Molecular e Celular da Universidade do Porto, Rua do Campo Alegre 823, 4150 Porto, and the Instituto de Cie ˆncias Biome ´dicas Abel Salazar da Universidade do Porto, 4000 Porto, Portugal. Fax: (351) 2 6099157; Email: [email protected] Chromosome Research Vol 6 1998 85
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Molecular cloning of metaphase chromosome protein 1 (MCP1), a novel human autoantigen that...

Molecular cloning of metaphase chromosome protein 1 (MCP1), anovel human autoantigen that associates with condensedchromosomes during mitosis
Elsa Bronze-da-Rocha, Jose A. Catita & Claudio E. Sunkel
Received 17 September 1997; received in revised form 5 November 1997; accepted for publication by A. Sumner 6November 1997
Systemic lupus erythematosus autoantibodies wereused to identify and to characterize new humanchromosome-associated proteins. Previous immuno-localization studies in human and murine tissueculture cells showed that some of these monoclonalantibodies recognize nuclear antigens that associatewith condensed chromosomes during mitosis. Oneantibody was selected for screening a human HeLaS3 cDNA expression library, and cDNAs that codefor an antigen of 31±33 kDa were isolated. Immuno-logical, biochemical and cell fractionation data in-dicate that the 31- to 33-kDa antigen corresponds tothe chromosome-associated protein recognized bythe original monoclonal antibody. Sequence analysisshows that we isolated a novel human gene.Immunolocalization to human tissue culture cellsshows that during interphase the antigen is dis-persed in the nucleus and that during mitosis itassociates exclusively with condensed chromo-somes. A similar pattern of localization was alsoobserved in mouse ®broblasts, suggesting that theantigen is conserved among different species. Final-ly, we show that part of the antigen remains boundto the scaffold/matrix component, even after highsalt extraction.
Key words: autoantigens, chromosome, mitosis, systemiclupus erythematosus
Introduction
In most higher eukaryotes, mitosis is characterized bythe presence of condensed chromosomes. The condensa-tion of chromatin into discrete chromosomes is essentialfor the proper segregation of chromatids into the twodaughter cells. The mitotic chromosome is a complexstructure in which the DNA nucleosome ®bre is thoughtto form loops that are attached to a proteinaseouscomponent called scaffold (Jackson 1991). A number of
proteins have been identi®ed that are constituents ofthis structure of which topoisomerase II is the bestcharacterized (Saitoh & Laemmli 1994). More recentlythe SMC (stability of minichromosomes) family ofproteins has been described and at least some of itsmembers are also thought to be part of the scaffold(Saitoh et al. 1995).
Human autoimmune sera from patients with variousdiseases, including several forms of sclerodermy; calci-nosis, Raynaud syndrome, oesophageal dismotility,sclerodactyly and telangiectasia (CREST); and systemiclupus erythematosus (SLE), have been used to identifychromosomal associated proteins. A number of theseproteins have been subsequently cloned (Tan et al. 1988;Tan 1989; Balczon 1993). Some of these proteins arecon®ned to speci®c chromosomal locations such as thecentromere (Earnshaw et al. 1987; Cooke et al., 1987;1990; Saitoh et al. 1992; Balczon 1993) and others appearto be distributed either at the perichromosomal regions(Hernandez-Verdun & Gautier 1994) or throughout thecondensed chromosomes (Earnshaw & Bernat 1991;Rattner 1992). Functional studies of many of theseproteins indicated that they are essential for severalaspects of chromosomal behaviour (Balczon 1993).
SLE in humans and animals is characterized by thepresence of high levels of serum antibodies that reactwith a variety of self antigens, including membranemolecules, cytoplasmic proteins and nuclear determi-nants (Tan 1989; 1991). MRL/ lpr mice develop anequivalent disease and have been used to produceautoreactive monoclonal antibodies (MAbs) (Cohen &Eisenberg 1991; Steinberg et al. 1991). The DNA-bindingantibodies produced in mice with SLE have someunusual properties. They recognize at least ®ve majorgroups of epitopes (Morgan et al. 1985a; Bronze-da-Rocha et al. 1992) and express cross-reactive idiotypesthat are shared between humans and mice (Morgan et
Chromosome Research 1998, 6, 85±95
# 1998 Rapid Science Publishers
E. Bronze-da-Rocha is at the LaboratoÂrio de GeneÂtica Molecular, Instituto de Biologia Molecular e Celular da Universidade doPorto, Rua do Campo Alegre 823, 4150 Porto, and the Faculdade de FarmaÂcia da Universidade do Porto, Rua Anibal Cunha,4000 Porto, Portugal. J. A. Catita is at the LaboratoÂrio de GeneÂtica Molecular, Instituto de Biologia Molecular e Celular daUniversidade do Porto, Rua do Campo Alegre 823, 4150 Porto. C. E. Sunkel (corresponding author) is at the LaboratoÂrio deGeneÂtica Molecular, Instituto de Biologia Molecular e Celular da Universidade do Porto, Rua do Campo Alegre 823, 4150 Porto,and the Instituto de CieÃncias BiomeÂdicas Abel Salazar da Universidade do Porto, 4000 Porto, Portugal. Fax: (�351) 2 6099157;Email: [email protected]
Chromosome Research Vol 6 1998 85

al. 1985b; Watts et al. 1990). Not all of these antibodiesare immunochemically speci®c for DNA because theyalso recognize nuclear, cytoplasmic and membraneantigens (Eilat, 1985; Bronze-da-Rocha et al. 1992). Onthe basis of the immuno¯uorescence staining pattern ofHEp-2 tissue culture cells during interphase, SLE mouseMAbs were classi®ed in ®ve groups. In mitotic cells,only two different staining patterns were observed(Bronze-da-Rocha et al. 1992). A group of antibodiesrecognized perichromosomal antigens staining the sur-face of mitotic chromosomes, whereas the other groupgives staining throughout condensed chromosomes. Inaddition, immunoblotting and immunoprecipitation ex-periments demonstrated that antibodies belonging to all®ve groups reacted with a wide range of phosphopro-teins with molecular weights from 25 kDa to 210 kDa(Bronze-da-Rocha et al. 1992). Although most of thereactivity was lost after dephosphorylation, some anti-bodies were still able to recognize speci®c nuclearantigens. The cross-reaction of these antibodies offersthe possibility for the identi®cation, isolation andmolecular cloning of chromosomal associated proteins.
Here we describe the isolation of a new human genethat encodes a chromosome-associated protein. cDNAswere isolated using a SLE mouse MAb that waspreviously shown to stain mitotic chromosomes. Theantigen encoded by the cDNAs is 31±33 kDa, and thesequence shows no homology to known sequences. Apolyclonal serum was raised against the bacteriallyexpressed protein and used for subcellular localizationin tissue culture cells. The results indicate that duringmitosis the 31±33-kDa antigen associates exclusivelywith condensed chromosomes. During other stages ofthe cell cycle it shows a speckle distribution within thenucleus, excluding the nucleolus. The localization of theantigen to interphase nuclei and mitotic chromosomeswas con®rmed after subcellular fractionation. Theseassays also showed that part of the antigen remainstightly bound to the scaffold/matrix component evenafter high salt extraction.
Materials and methods
Antibodies
MAbs selected for their reactivity with DNA were preparedfrom spleen cells of unstimulated MRL/Mp± lpr/lpr (MRL/lpr)according to the method described previously (Lake et al. 1987;Ravirajan & Staines 1991). MAb 402 was obtained from anadult MRL/lpr mouse. Polyclonal antibody (Rb-7) was pre-pared against the bacterially expressed maltose-binding:MCP33fusion protein injected intramuscularly into a New Zealandwhite rabbit.
Screening of a human cDNA expression library
A human HeLa S3 cDNA ëgt11 expression library (59 stretch,Clontech laboratories, Palo Alto, CA, USA) was screened withMAb 402. A total of 600±000 recombinant phages werescreened with MAb 402 in a 1:100 dilution. Phage DNA wasprepared according to previously published methods
(Sambrook et al., 1989). Positive clones were grouped by dotblot using radiolabelled DNA probes.
Expression of fusion proteins
Fusion proteins were prepared using ëgt11 (Clontech labora-tories, Palo Alto, CA, USA) and pMAL-cRI (New EnglandBiolabs, MA, USA), according to the published protocols(Sambrook et al., 1989). Maltose-binding fusion protein waspuri®ed using a maltose resin column according to themanufacturer's instructions (New England Biolabs). Polyclonalantibody was af®nity puri®ed using nitrocellulose membranecontaining puri®ed maltose-binding fusion protein (Harlow &Lane 1988). Puri®ed maltose-binding fusion protein wascleaved with Xa factor (New England Biolabs).
Electrophoresis and immunoblottingElectrophoresis was carried out according to Laemmli (1970).Total HEp-2 cell extracts or puri®ed proteins were solubilizedin Laemmli sample buffer and electrophoretically transferred tonitrocellulose (Hybond-C, Amersham, UK) as described pre-viously (Towbin et al. 1979). For immunoblotting, MAb 402(1:100 or 1:250 dilution) and Rb-7 (1:200 dilution) were used asprimary antibodies. Goat anti-rabbit IgG (H and L) and horseanti-mouse IgG (H and L) conjugated to peroxidase (VectorLaboratoreis, UK) were used as second antibodies in a 1:100dilution. The immune signal was detected using the ECLWestern blot detection system (Amersham, UK).
Northern and Southern analysisTotal RNA from HEp-2 and HeLa cells (1 3 106 cells=ml) wasextracted with guanidinium thiocyanate followed by centrifu-gation in caesium chloride solution. Poly(A)� RNA wasisolated by af®nity chromatography on oligo(dT)-cellulose(Sigma) according to the method described previously(Sambrook et al. 1989). Total RNA and Poly(A)� RNA weredenatured with glyoxal and dimethyl sulphoxide, separated byelectrophoresis on a 1.5% agarose gel and blotted onto nylonmembranes. Probes for hybridization were labelled with[32P]dCTP. Genomic DNA from HEp-2 cells, human blood,mouse NIH3T3 cells and adult Drosophila ¯ies was isolated,digested with restriction enzymes, separated by electrophoresison a 0.8% agarose gel and transferred to a nylon membraneunder alkaline conditions (Sambrook et al. 1989). DNA probeswere labelled with [32P]-á-dCTP and hybridization was carriedout as described previously (Sambrook et al. 1989).
DNA sequencing
Complete cDNA inserts were subcloned into the phagemidspBS (�=ÿ) and pBluescript II SK (�=ÿ). Unidirectional dele-tions were made using the Erase-a-base system (Promega,USA). Subclones were sequenced with Sequenase (UnitedStates Biochemical, USA). The ®nal sequence was assembledand analysed with PC/gene software (Intelligenetics, USA).
In vitro transcription and translation
The complete cDNA was subcloned into the transcriptionvectors pSPT18 and pSPT19 (Boehringer Mannheim, UK).Transcription was carried out according to the manufacturersinstructions and transcripts were analysed on a 1% agarose gel.In vitro translation was performed with a rabbit reticulocytelysate (Amersham, UK) in the presence of [35S]methionine(Amersham, UK). The translated product was analysed using15% SDS±PAGE, and immunoprecipitation was carried outwith immunopuri®ed polyclonal serum (Rb-7) and monoclonal
86 Chromosome Research Vol 6 1998
E. Bronze-da-Rocha et al.

(MAb 402) antibodies, as described previously (Bronze-daRocha et al. 1992).
Cell culture and preparation of protein extractsHuman HEp-2 and HeLa, and mouse NIH3T3 cell lines werecultured in Dulbecco's modi®ed Eagle medium (DMEM)supplemented with 10% fetal calf serum (FCS), 2 mM glutha-mine, 100 i:u:=ml penicillin, and 100 ìg=ml streptomycin, at378C in a humidi®ed atmosphere in a 7% CO2 incubator. TotalHEp-2 and HeLa cell extracts were prepared as describedpreviously (Bronze-da Rocha et al. 1992). Interphase nuclearextracts were prepared according to published protocols(Pankov et al. 1990). For mitotic chromosomes, HeLa or HEp-2cells were arrested with 0:4 ìg=ml of colchicine for 24 h. Mitoticchromosomes were obtained according to the method of Bernatet al. (1990). Preparation of nuclear matrix extracts wasperformed according to published protocols (Staufenbiel &Deppert 1984).
Indirect immuno¯uorescence microscopy
Antigens were localized by indirect immuno¯uorescence onHEp-2, HeLa and NIH cells grown on glass coverslips. Cellswere ®xed with 3% paraformaldehyde in phosphate-bufferedsaline (PBS) for 10 min. MAb 402 was used at a 1:50 dilutionand Rb7 at 1:20 dilution. Second antibodies were anti-mouseIgG ¯uorescein-linked whole antibody from sheep (Amersham,UK) and an anti-rabbit IgG (H and L) ¯uorescein-linkedantibody from goat (Vector Laboratories,UK); both were usedat 1:1000 dilution in PBS containing 10% FCS and 0.1% TritonX-100 for 1 h at room temperature. Preparations were treatedwith 50 ìg=ml RNAase during antibody incubation to reducebackground staining of propidium iodide. Coverslips weremounted in Vector Shield (Vector Laboratories, UK) andobserved using a confocal laser scanning microscope BioR-adMRC-600. Image analysis was performed using COMOSsoftware (Bio-Rad Laboratories, USA).
Results
Isolation and characterization of cDNAclonesPreviously (Bronze-da-Rocha et al. 1992), we had shownthat SLE MAb 402 stained condensed chromosomesduring mitosis and recognized a 33-kDa antigen inHEp-2 cell extracts. MAb 402 was then used to screen ahuman HeLa S3 cDNA ëgt11 expression library. TwocDNA clones (XT-2 and XT-5) were isolated and shownto contain a 1.2-kb insert that hybridized to each otherby dot blot. The nucleotide sequence of the XT-2 cDNAclone was determined for both strands after unidirec-tional deletions (Figure 1). Analysis of the sequenceshows that the XT-2 cDNA has an open reading frameof 870 bp, extending from the ®rst ATG at position 105to position 974 (Figure 1). The coding sequence isfollowed by a 39 untranslated region characterized bythe presence of several stop codons in all readingframes and a putative polyadenylation signal. The openreading frame encodes for a polypeptide of 290 aminoacids of 33 kDa. A total of 41 acidic (Asp � Glu) and 28basic (Arg � Lys) amino acids were found, predicting anet negative charge with an isoelectric point (IP) of 4.56.The IP of the bacterially expressed protein was 4.8 (data
not shown). A number of potential phosphorylationsites were identi®ed for protein kinase C, casein kinaseII and for p34cdc2 kinase (Figure 1). A search throughdatabases revealed that the open reading frame presentin the XT-2 cDNA has no signi®cant homology withother eukaryotic proteins described to date. Given thatthe 31- to 33-kDa antigen is present in nuclear extractsand in isolated chromosome Preparations (see later), wehave called the protein MCP1 (metaphase chromosomeprotein).
The cDNA XT-2 encodes the nuclear antigenrecognized by MAb 402To show that XT-2 cDNA encodes for the nuclearantigen recognized by MAb 402, a number of experi-ments were performed. The coding region of the XT-2cDNA was expressed as different fusion proteins, itsmolecular weight determined and its reactivity to MAb402 and the new antibody Rb7 ascertained (Figure 2).Fusion proteins from the original XT-2 and XT-5 phagelysogens and a control phage were expressed in Escher-ichia coli, separated by SDS±PAGE and stained (Figure2A). The results indicate that both XT-2 and XT-5express a 145-kDa protein that is not present in thecontrol phage. In this case, the â-galactosidase part offusion proteins accounts for approximately 110 kDa,suggesting that the cDNA encodes a 30- to 35-kDaprotein. The 1.2-kb insert from XT-2 was also subclonedinto a pMAL expression vector and the fusion proteinisolated by af®nity chromatography through a maltosecolumn. The results indicate that after cleavage of thefusion protein with factor Xa to release the maltosebinding part, a band of 33 kDa is present in thepreparation (Figure 2B).
To further characterize the protein encoded by theXT-2 cDNA, a polyclonal serum was raised in rabbitsagainst the pMAL±MCP1 fusion protein. The reactivityof the immunopuri®ed polyclonal sera (Rb-7) was testedagainst puri®ed fusion protein and total HEp-2 cellextracts (Figure 2C). Immunoblotting indicates thatpolyclonal serum Rb-7 recognizes speci®cally thepMAL±MCP1 fusion protein extracts (Figure 2C, lane6), and no reactivity was detected when preimmuneserum was used (not shown). Furthermore, the immu-nopuri®ed polyclonal serum Rb-7 identi®es a faint 33-kDa polypeptide in whole extracts of HEp-2 cells(Figure 2C, lane 7). MAb 402 also recognizes thepMAL±MCP1 fusion protein (Figure 2C, lane 8) but intotal HEp-2 cell extracts recognizes a number of pro-teins. Nevertheless, a strongly reactive band of 33 kDacan be observed (Figure 2C, lane 9). However, if nuclearextracts from HeLa cells are used, MAb 402 recognizesa polypeptide of 33 kDa and a faint band of 31 kDa(Figure 2C, lane 10).
Finally, we wished to determine whether the XT-2cDNA clone could encode the full MCP1 open readingframe. The complete insert from XT-2 was subcloned inboth orientations into the pSPT19 expression vector. Invitro translation of the RNA produced from the pSPT-
Chromosome Research Vol 6 1998 87
Mitotic chromosome-associated protein

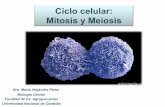
19.1 (the 1.2 kb insert from XT-2 cloned in the senseorientation) was performed in rabbit reticulocyte lysatesin the presence of [35S]Met and the products resolvedby SDS±PAGE (Figure 3). The results show that whenthe vector pSPT-19.1 is used, two speci®c polypeptidesof 31 and 33 kDa are produced (Figure 3A, lane 2).Neither band is observed when the control plasmidpSPT-18.1 is used (Figure 3A, lane 1). Immunoprecipi-tation with either MAb 402 (Figure 3B, lane 3) orimmunopuri®ed Rb-7 polyclonal serum (Figure 3B,lanes 4) shows that the two polypeptides of 31 and33 kDa are speci®cally recognized by both antibodies.No immunoprecipitation was detected when preim-mune Rb7 serum was used (Figure 3B, lane 5). The
results indicate that two immunologically related trans-lation products can be obtained from the XT-2 insert.The same two antigens are detected in nuclear extractsfrom HeLa cells (see Figure 2C, lane 10). As thenucleotide sequence of XT-2 shows a number of in-frame ATGs near the 59 end, it is possible that at leastone of them, apart from the ®rst ATG, could be usedduring translation both in vivo and in vitro.
MCP1 is encoded by a single locus thatexpresses a 1.3-kb transcriptTo determine the size and number of transcripts derivedfrom the MCP1 locus, Northern blot analysis wasperformed using Poly(A)� RNA isolated from exponen-
Figure 1. Nucleotide sequence and conceptual translation of the XT-2 cDNA. The amino acid sequence of the proteinencoded by cDNA XT-2 is shown above the nucleic acid sequence using a single-letter code. Nucleotide sequencepositions are numbered to the left of the ®gure and the amino acid sequence is numbered to the right. The ®rstinitiation codon, the stop codon and the putative polyadenylation signal are underlined. One potential glycosylation (#)and several potential sites for protein kinase C (·), casein kinase II (�) and p34cdc2 kinase (°) are also shown. TheEMBL accession number for this sequence is X94629.
88 Chromosome Research Vol 6 1998
E. Bronze-da-Rocha et al.

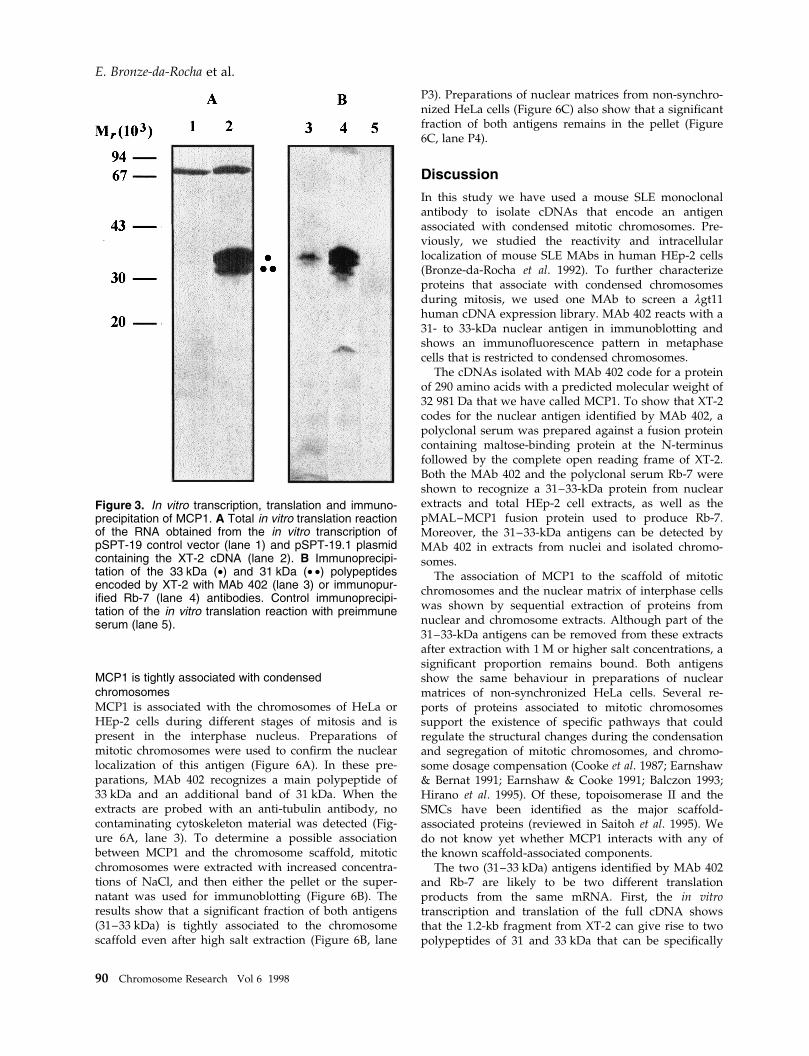
tially growing HeLa cells and hybridized with [32P]XT-2cDNA probe (Figure 4). The results shows that a singleband of 1.3 kb is obtained. The size of the transcriptidenti®ed by Northern analysis is in good agreementwith the size of the XT-2 cDNA, suggesting that wehave cloned a full size transcript.
Genomic Southern analysis was performed to deter-mine whether this protein is encoded by a single gene.Genomic human DNA was digested with severalrestriction enzymes, blotted and probed with the com-plete XT-2 cDNA. Digestion with BamHI gave a singleband of 4.2 kb (Figure 4B, lane 2). Digestion with EcoRIgave two bands of 4.2 kb and 4.0 kb (Figure 4B, lane 3),and with HindIII gave a single band of 2.1 kb (Figure4B, lane 4). Genomic DNA from other species includingMus musculus and Drosophila melanogaster was digestedwith restriction enzymes, blotted and probed with thefull cDNA at moderate stringency. The results indicatethat both species contain sequences homologous toMCP1 (Figure 4C & D).
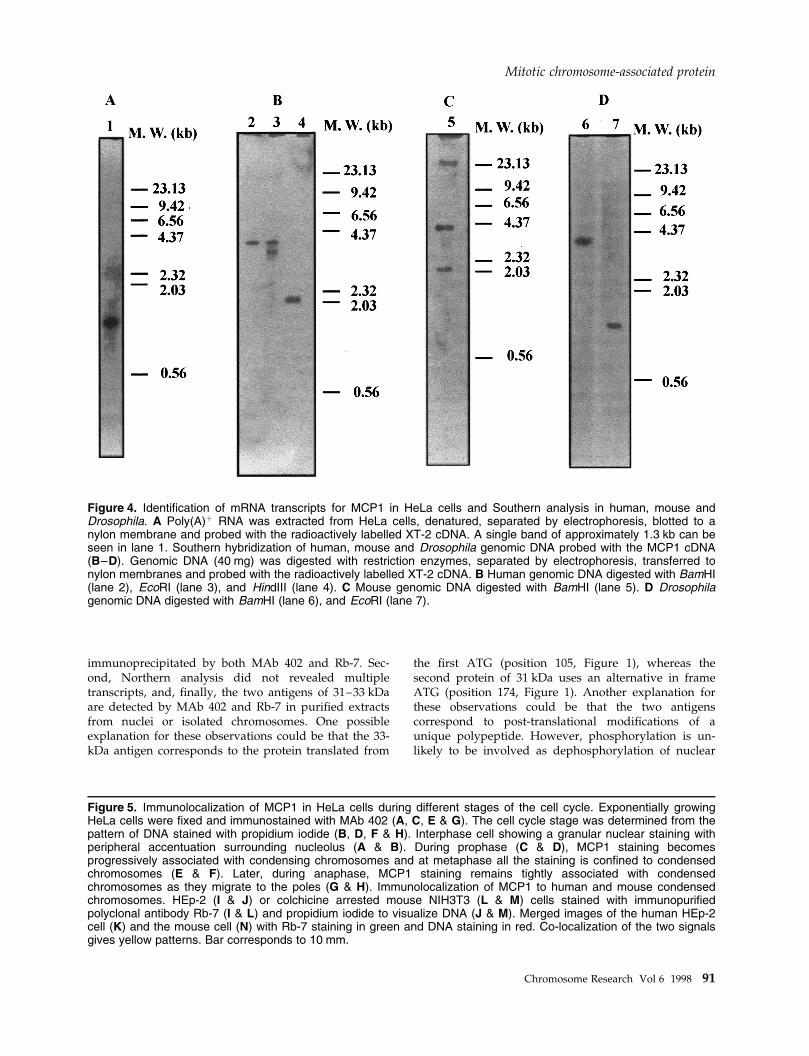
MCP1 associates with condensedchromosomes during mitosisThe intracellular distribution of MCP1 in HeLa cellsduring different cell cycle stages was determined usingMAb 402 (Figure 5A±H) and con®rmed using immuno-puri®ed RB-7 antibody (Figure 5I±N). Cells in inter-phase showed a nuclear granular staining withperipheral accentuation surrounding the nucleolus (Fig-ure 5A & B). During prophase the staining becomesexclusively associated with condensing chromatin(Figure 5C & D) and at metaphase strong staining ofchromosomes is observed (Figure 5E & F) that remainsuntil anaphase (Figure 5G & H). Immunopuri®ed Rb-7shows a strong staining of condensed chromosomes inHEp-2 cells (Figure 5I) and in mouse ®broblasts NIH3T3cells arrested at metaphase with colchicine (Figure 5L).The cell cycle distribution of MCP1 did not depend onthe ®xation methods used and no staining of chromo-somes was detected with RB-7 preimmune sera (datanot shown).
Figure 2. Characterization of the protein encoded by the XT-2 cDNA and production of the polyclonal serum Rb-7. ACoomassie blue stained SDS±PAGE gel showing the ëgt11 fusion proteins from XT-2 DNA (lane 1), lgt11 phage withno insert (lane 2) and XT-5 cDNA (lane 3). B Coomassie blue-stained SDS±PAGE gel of the puri®ed MCP1 proteinobtained by cleavage of the pMAL±MCP1 fusion protein with Xa factor (lane 4) and the complete pMAL±MCP1 fusionprotein (lane 5). C Immunoblotting assay showing the reactivity of the immunopuri®ed polyclonal antibody Rb-7 (lane 6)and MAb 402 (lane 8) against the pMAL±MCP1 fusion protein. The immunoblotting assay also shows the reactivity ofthe immunopuri®ed polyclonal antibody Rb-7 (lane 7) and MAb 402 (lane 9) against total HEp-2 cell extracts or nuclearextracts from HeLa cells probed with MAb 402 (lane 10). MAb 402 and immunopuri®ed Rb-7 serum were used at 1:100dilution.
Chromosome Research Vol 6 1998 89
Mitotic chromosome-associated protein
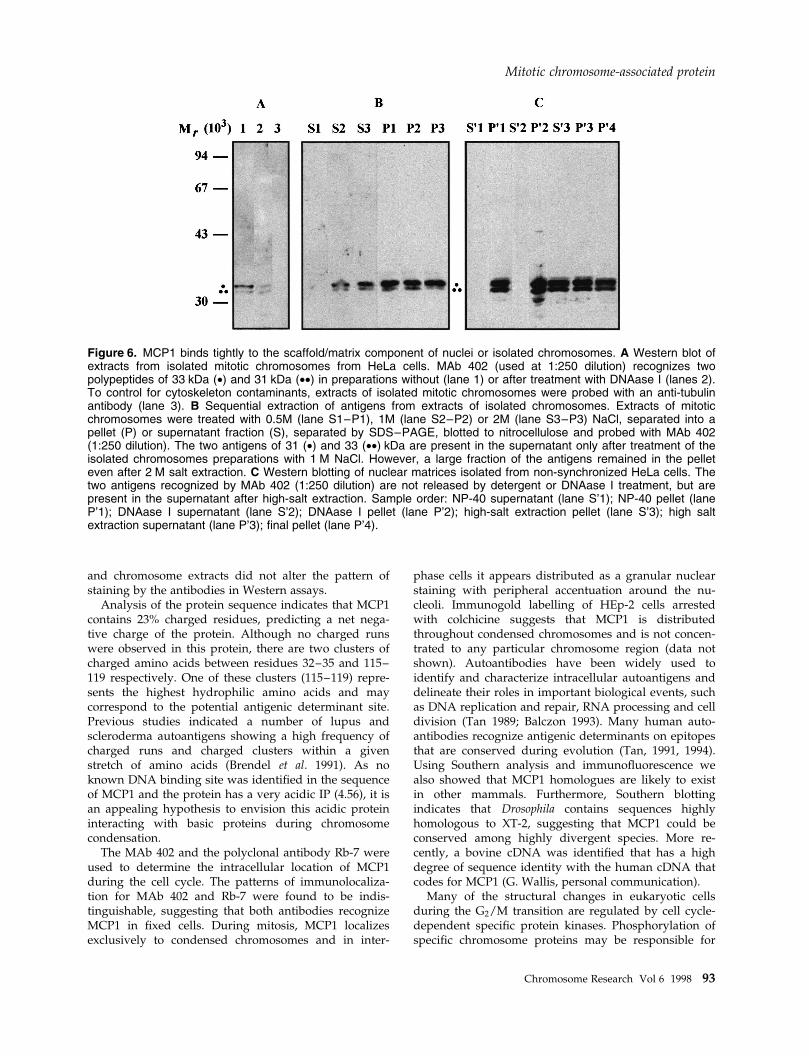
MCP1 is tightly associated with condensedchromosomesMCP1 is associated with the chromosomes of HeLa orHEp-2 cells during different stages of mitosis and ispresent in the interphase nucleus. Preparations ofmitotic chromosomes were used to con®rm the nuclearlocalization of this antigen (Figure 6A). In these pre-parations, MAb 402 recognizes a main polypeptide of33 kDa and an additional band of 31 kDa. When theextracts are probed with an anti-tubulin antibody, nocontaminating cytoskeleton material was detected (Fig-ure 6A, lane 3). To determine a possible associationbetween MCP1 and the chromosome scaffold, mitoticchromosomes were extracted with increased concentra-tions of NaCl, and then either the pellet or the super-natant was used for immunoblotting (Figure 6B). Theresults show that a signi®cant fraction of both antigens(31±33 kDa) is tightly associated to the chromosomescaffold even after high salt extraction (Figure 6B, lane
P3). Preparations of nuclear matrices from non-synchro-nized HeLa cells (Figure 6C) also show that a signi®cantfraction of both antigens remains in the pellet (Figure6C, lane P4).
Discussion
In this study we have used a mouse SLE monoclonalantibody to isolate cDNAs that encode an antigenassociated with condensed mitotic chromosomes. Pre-viously, we studied the reactivity and intracellularlocalization of mouse SLE MAbs in human HEp-2 cells(Bronze-da-Rocha et al. 1992). To further characterizeproteins that associate with condensed chromosomesduring mitosis, we used one MAb to screen a ëgt11human cDNA expression library. MAb 402 reacts with a31- to 33-kDa nuclear antigen in immunoblotting andshows an immuno¯uorescence pattern in metaphasecells that is restricted to condensed chromosomes.
The cDNAs isolated with MAb 402 code for a proteinof 290 amino acids with a predicted molecular weight of32 981 Da that we have called MCP1. To show that XT-2codes for the nuclear antigen identi®ed by MAb 402, apolyclonal serum was prepared against a fusion proteincontaining maltose-binding protein at the N-terminusfollowed by the complete open reading frame of XT-2.Both the MAb 402 and the polyclonal serum Rb-7 wereshown to recognize a 31±33-kDa protein from nuclearextracts and total HEp-2 cell extracts, as well as thepMAL±MCP1 fusion protein used to produce Rb-7.Moreover, the 31±33-kDa antigens can be detected byMAb 402 in extracts from nuclei and isolated chromo-somes.
The association of MCP1 to the scaffold of mitoticchromosomes and the nuclear matrix of interphase cellswas shown by sequential extraction of proteins fromnuclear and chromosome extracts. Although part of the31±33-kDa antigens can be removed from these extractsafter extraction with 1 M or higher salt concentrations, asigni®cant proportion remains bound. Both antigensshow the same behaviour in preparations of nuclearmatrices of non-synchronized HeLa cells. Several re-ports of proteins associated to mitotic chromosomessupport the existence of speci®c pathways that couldregulate the structural changes during the condensationand segregation of mitotic chromosomes, and chromo-some dosage compensation (Cooke et al. 1987; Earnshaw& Bernat 1991; Earnshaw & Cooke 1991; Balczon 1993;Hirano et al. 1995). Of these, topoisomerase II and theSMCs have been identi®ed as the major scaffold-associated proteins (reviewed in Saitoh et al. 1995). Wedo not know yet whether MCP1 interacts with any ofthe known scaffold-associated components.
The two (31±33 kDa) antigens identi®ed by MAb 402and Rb-7 are likely to be two different translationproducts from the same mRNA. First, the in vitrotranscription and translation of the full cDNA showsthat the 1.2-kb fragment from XT-2 can give rise to twopolypeptides of 31 and 33 kDa that can be speci®cally
Figure 3. In vitro transcription, translation and immuno-precipitation of MCP1. A Total in vitro translation reactionof the RNA obtained from the in vitro transcription ofpSPT-19 control vector (lane 1) and pSPT-19.1 plasmidcontaining the XT-2 cDNA (lane 2). B Immunoprecipi-tation of the 33 kDa (·) and 31 kDa (· ·) polypeptidesencoded by XT-2 with MAb 402 (lane 3) or immunopur-i®ed Rb-7 (lane 4) antibodies. Control immunoprecipi-tation of the in vitro translation reaction with preimmuneserum (lane 5).
90 Chromosome Research Vol 6 1998
E. Bronze-da-Rocha et al.
immunoprecipitated by both MAb 402 and Rb-7. Sec-ond, Northern analysis did not revealed multipletranscripts, and, ®nally, the two antigens of 31±33 kDaare detected by MAb 402 and Rb-7 in puri®ed extractsfrom nuclei or isolated chromosomes. One possibleexplanation for these observations could be that the 33-kDa antigen corresponds to the protein translated from
the ®rst ATG (position 105, Figure 1), whereas thesecond protein of 31 kDa uses an alternative in frameATG (position 174, Figure 1). Another explanation forthese observations could be that the two antigenscorrespond to post-translational modi®cations of aunique polypeptide. However, phosphorylation is un-likely to be involved as dephosphorylation of nuclear
Figure 4. Identi®cation of mRNA transcripts for MCP1 in HeLa cells and Southern analysis in human, mouse andDrosophila. A Poly(A)� RNA was extracted from HeLa cells, denatured, separated by electrophoresis, blotted to anylon membrane and probed with the radioactively labelled XT-2 cDNA. A single band of approximately 1.3 kb can beseen in lane 1. Southern hybridization of human, mouse and Drosophila genomic DNA probed with the MCP1 cDNA(B±D). Genomic DNA (40 mg) was digested with restriction enzymes, separated by electrophoresis, transferred tonylon membranes and probed with the radioactively labelled XT-2 cDNA. B Human genomic DNA digested with BamHI(lane 2), EcoRI (lane 3), and HindIII (lane 4). C Mouse genomic DNA digested with BamHI (lane 5). D Drosophilagenomic DNA digested with BamHI (lane 6), and EcoRI (lane 7).
Figure 5. Immunolocalization of MCP1 in HeLa cells during different stages of the cell cycle. Exponentially growingHeLa cells were ®xed and immunostained with MAb 402 (A, C, E & G). The cell cycle stage was determined from thepattern of DNA stained with propidium iodide (B, D, F & H). Interphase cell showing a granular nuclear staining withperipheral accentuation surrounding nucleolus (A & B). During prophase (C & D), MCP1 staining becomesprogressively associated with condensing chromosomes and at metaphase all the staining is con®ned to condensedchromosomes (E & F). Later, during anaphase, MCP1 staining remains tightly associated with condensedchromosomes as they migrate to the poles (G & H). Immunolocalization of MCP1 to human and mouse condensedchromosomes. HEp-2 (I & J) or colchicine arrested mouse NIH3T3 (L & M) cells stained with immunopuri®edpolyclonal antibody Rb-7 (I & L) and propidium iodide to visualize DNA (J & M). Merged images of the human HEp-2cell (K) and the mouse cell (N) with Rb-7 staining in green and DNA staining in red. Co-localization of the two signalsgives yellow patterns. Bar corresponds to 10 mm.
Chromosome Research Vol 6 1998 91
Mitotic chromosome-associated protein
E. Bronze-da-Rocha et al.
and chromosome extracts did not alter the pattern ofstaining by the antibodies in Western assays.
Analysis of the protein sequence indicates that MCP1contains 23% charged residues, predicting a net nega-tive charge of the protein. Although no charged runswere observed in this protein, there are two clusters ofcharged amino acids between residues 32±35 and 115±119 respectively. One of these clusters (115±119) repre-sents the highest hydrophilic amino acids and maycorrespond to the potential antigenic determinant site.Previous studies indicated a number of lupus andscleroderma autoantigens showing a high frequency ofcharged runs and charged clusters within a givenstretch of amino acids (Brendel et al. 1991). As noknown DNA binding site was identi®ed in the sequenceof MCP1 and the protein has a very acidic IP (4.56), it isan appealing hypothesis to envision this acidic proteininteracting with basic proteins during chromosomecondensation.
The MAb 402 and the polyclonal antibody Rb-7 wereused to determine the intracellular location of MCP1during the cell cycle. The patterns of immunolocaliza-tion for MAb 402 and Rb-7 were found to be indis-tinguishable, suggesting that both antibodies recognizeMCP1 in ®xed cells. During mitosis, MCP1 localizesexclusively to condensed chromosomes and in inter-
phase cells it appears distributed as a granular nuclearstaining with peripheral accentuation around the nu-cleoli. Immunogold labelling of HEp-2 cells arrestedwith colchicine suggests that MCP1 is distributedthroughout condensed chromosomes and is not concen-trated to any particular chromosome region (data notshown). Autoantibodies have been widely used toidentify and characterize intracellular autoantigens anddelineate their roles in important biological events, suchas DNA replication and repair, RNA processing and celldivision (Tan 1989; Balczon 1993). Many human auto-antibodies recognize antigenic determinants on epitopesthat are conserved during evolution (Tan, 1991, 1994).Using Southern analysis and immuno¯uorescence wealso showed that MCP1 homologues are likely to existin other mammals. Furthermore, Southern blottingindicates that Drosophila contains sequences highlyhomologous to XT-2, suggesting that MCP1 could beconserved among highly divergent species. More re-cently, a bovine cDNA was identi®ed that has a highdegree of sequence identity with the human cDNA thatcodes for MCP1 (G. Wallis, personal communication).
Many of the structural changes in eukaryotic cellsduring the G2/M transition are regulated by cell cycle-dependent speci®c protein kinases. Phosphorylation ofspeci®c chromosome proteins may be responsible for
Figure 6. MCP1 binds tightly to the scaffold/matrix component of nuclei or isolated chromosomes. A Western blot ofextracts from isolated mitotic chromosomes from HeLa cells. MAb 402 (used at 1:250 dilution) recognizes twopolypeptides of 33 kDa (·) and 31 kDa (··) in preparations without (lane 1) or after treatment with DNAase I (lanes 2).To control for cytoskeleton contaminants, extracts of isolated mitotic chromosomes were probed with an anti-tubulinantibody (lane 3). B Sequential extraction of antigens from extracts of isolated chromosomes. Extracts of mitoticchromosomes were treated with 0.5M (lane S1±P1), 1M (lane S2±P2) or 2M (lane S3±P3) NaCl, separated into apellet (P) or supernatant fraction (S), separated by SDS±PAGE, blotted to nitrocellulose and probed with MAb 402(1:250 dilution). The two antigens of 31 (·) and 33 (··) kDa are present in the supernatant only after treatment of theisolated chromosomes preparations with 1 M NaCl. However, a large fraction of the antigens remained in the pelleteven after 2 M salt extraction. C Western blotting of nuclear matrices isolated from non-synchronized HeLa cells. Thetwo antigens recognized by MAb 402 (1:250 dilution) are not released by detergent or DNAase I treatment, but arepresent in the supernatant after high-salt extraction. Sample order: NP-40 supernatant (lane S'1); NP-40 pellet (laneP'1); DNAase I supernatant (lane S'2); DNAase I pellet (lane P'2); high-salt extraction pellet (lane S'3); high saltextraction supernatant (lane P'3); ®nal pellet (lane P'4).
Mitotic chromosome-associated protein
Chromosome Research Vol 6 1998 93

the change of chromosome structure seen at the onset ofmitosis. Some of the antigens that had been describedshowed a cell cycle-dependent phosphorylation anddephosphorylation (Vandre et al. 1984; 1991; Davis et al.1983; Rao et al. 1989). MCP1 contains a large number ofputative phosphorylation sites in its sequence. Althoughpreliminary evidence shows that MCP1 is phosphory-lated in vivo (E. Bronze-da-Rocha & C. E. Sunkel,unpublished results), the sites have not yet beenmapped and their functional relevance has not beendetermined. Further in vivo experiments are in progressto determine the role of MCP1 during mitosis.
Acknowledgements
We are grateful to Adelaide Santos for general technicalassistance, FundacËaÄo Gomes Teixeira da Universidadedo Porto for their ®nancial contribution towards thisproject and Gillian Wallis for providing results beforepublication. The Molecular Genetics Laboratory is®nanced by grants from the Junta Nacional de Investi-gacËaÄo Ciente®ca e TecnoloÂgica (JNICT) of Portugal andthe European Union.
References
Balczon R (1993) Autoantibodies as probes in cell and molecu-lar biology Proc Soc Exp Biol Med 204: 138±154.
Bernat BL, Borisy GG, Roth®eld NF, Earnshaw WC (1990)Injection of anticentromere antibodies in interphase disruptsevents required for chromosome movement at mitosis. J CellBiol 111: 1519±1533.
Brendel V, Dohlman J, Blaisdell E, Karlin S (1991) Very longcharge runs in systemic lupus erythematosus-associatedautoantigens Proc Natl Acad Sci USA 88: 1634±1540.
Bronze-da-Rocha E, Machado C, Staines NA, Sunkel CE (1992)Systemic lupus erythematosus murine monoclonal DNA±binding antibodies recognize cytoplasmic and nuclear phos-phorylated antigens that display cell cycle redistribution inHEp-2 cells. Immunology 77: 582±591.
Cohen PL, Eisenberg RA (1991) Lpr and gld: single genemodels of systemic autoimmunity and lymphoproliferativedisease. Annu Rev Immunol 9: 234±269.
Cooke CA, Heck MM, Earnshaw WC (1987) The inner centro-mere protein (INCENP) antigens: movement from innercentromere to midbody during mitosis J Cell Biol 105: 2053±2067.
Cooke CA, Bernat RL, Earnshaw WC (1990) CENP-B: a majorhuman centromere protein located beneath the kinetochore. JCell Biol 110: 1475±1488.
Davis FM, Tsao TY, Fowler SK, Rao PN (1983) Monoclonalantibodies to mitotic cells. Proc Natl Acad Sci USA 80: 2926±2930.
Earnshaw WC, Sullivan KF, Machlin PS et al. (1987) Molecularcloning of cDNA for CENP-B, the major human centromereautoantigen. J Cell Biol 104: 817±829.
Earnshaw WC, Bernat RL (1991) Chromosomal passenger:toward an integrated view of mitosis. Chromosoma 100: 139±146.
Earnshaw WC, Cooke CA (1991) Analysis of the distribution ofthe INCENPS throughout mitosis reveals the existence of a
pathway of structural changes in the chromosomes duringmetaphase and early events in the cleavage furrow formationJ Cell Sci 98: 443±461.
Eilat D (1985) Cross-reactions of anti-DNA antibodies and thecentral dogma of lupus nephritis. Immunol Today 6: 123±127.
Harlow E, Lane D (1988) Antibodies a Laboratory Manual, 1stedn. Cold Spring Harbor, USA: Cold Spring Harbor Labora-tory Press.
Hernandez-Verdun D, Gautier T (1994) The chromosomeperiphery during mitosis. Bioessays 16: 179±185.
Hirano T, Mitchison TJ, Swedlow JR (1995) The SMC family:from chromosome condensation to dosage compensation.Curr Opin Cell Biol 7: 329±336.
Jackson AA (1991) Structure±function relationships in eukaryo-tic nuclei. Bioessays 13: 1±10.
Laemmli UK (1970) Cleavage of structural protein duringassembly of the head of bacteriophage T4. Nature 227: 680±685.
Lake R, Morgan A, Staines N (1987) Production of monoclonalantibodies. Soc Appl Microbiol Tech Series 24: 1±19.
Morgan A, Buchanan RC, Lew AM, Oslen I, Staines NA (1985a)Five groups of antigenic determinants on DNA identi®ed bymonoclonal antibodies from (NZB 3 NZW)F1 and MRL/Mp± lpr/lpr mice. Immunology 55: 75±83.
Morgan A, Isenberg DA, Naparstek Y et al. (1985b) Sharedidiotypes are expressed on mouse and human anti-DNAautoantibodies. Immunology 56: 393±399.
Pankov R, Lemieux M, Hancok R (1990) An antigen located inthe kinetochore region in metaphase and polar microtubuleends in midbody region in anaphase, characterized usingmonoclonal antibody. Chromosoma 99: 95±101.
Rao PN, Zhao J, Ganju RK, Ashorn CL (1989) Monoclonalantibody against the centrosome. J Cell Sci 93: 63±69.
Rattner JB (1992) Integrating chromosome structure with func-tion. Chromosoma 101, 259±264.
Ravirajan CT, Staines NA (1991) Involvement in lupus diseaseof idiotypes Id.F-423 and Id.IV-228 de®ned, respectively,upon foetal and adult MRL/Mp-lpr/lpr DNA-binding mono-clonal antibodies. Immunology 74: 342±347.
Saitoh Y, Laemmli UK (1994) Metaphase chromosome structure:bands arise from differential folding path of the highly ATrich scaffold. Cell 76: 1260±1269.
Saitoh H, Tonkiel J, Cooke CA et al. (1992) CENP-C, anautoantigen in scleroderma, is a component of the humaninner kinetochore plate. Cell 70: 115±125.
Saitoh N, Goldberg I, Earnshaw WC (1995) The SMC proteinsand the coming of age of the chromosome scaffold hypoth-esis. Bioessays 17: 759±765.
Sambrook J, Fritsch EF, Maniatis T (1989) Molecular Cloning aLaboratory Manual, 2nd edn. Cold Spring Harbor Laboratory,NY: Cold Spring Harbor Laboratory Press.
Staufenbiel M, Deppert W (1984) Preparation of nuclearmatrices from cultured cells: subfractionation of nuclei insitu. J Cell Biol 98: 1886±1894.
Steinberg AD, Gourley MF, Klinman DM et al. (1991) Systemiclupus erythematosus. Ann Intern Med 115: 548±559.
Tan EM (1989) Antinuclear antibodies: diagnostic markers forautoimmune diseases and probes for cell biology. AdvImmmunol 44: 93±151.
Tan EM (1991) Autoantibodies in pathology and cell biology.Cell 67: 841±842.
Tan EM (1994) Autoimmunity and apoptosis. J Exp Med 179:1083±1086.
Tan EM, Chan KL, Sullivan KF, Rubin RL (1988) Antinuclear
E. Bronze-da-Rocha et al.
94 Chromosome Research Vol 6 1998

antibodies (ANAs): diagnostically speci®c immune markersand clues toward the understanding of systemic autoimmu-nity. Clin Immunol Immunopathol 47: 121±141.
Towbin H, Staehelin T, Gordon J (1979) Electrophoretic transferof proteins from polyacrylamide gels to nitrocellulose sheets:procedure and some applications. Proc Natl Acad Sci USA 76:4350±4354.
Vandre DD, Davis FM, Rao PN, Borisy GG (1984) Phosphopro-teins are components of mitotic microtubule organizing
centers. Proc Natl Acad Sci USA 81: 4439±4443.Vandre DD, Centonze VE, Peloquin J, Tombes RM, Borisy GG
(1991) Proteins of the mammalian mitotic spindle: phosphor-ylations/dephosphorylations of MAP-4 during mitosis. J CellSci 98: 577±588.
Watts RA, Ravirajan CT, Staines NA, Isenberg DA (1990) Ahuman fetal monoclonal DNA-binding antibody shares idio-types with fetal and adult murine monoclonal DNA-bindingantibodies. Immunology 69: 348±354.
Mitotic chromosome-associated protein
Chromosome Research Vol 6 1998 95