
Thidiazuron-induced adventitious shoot regeneration of sweetpotato ( Ipomoea batatas

Annals of Botany 87: 371±378, 2001doi:10.1006/anbo.2000.1346, available online at http://www.idealibrary.com on
Cryopreservation of Shoot Tips from Six Endangered Australian Species using a Modi®edVitri®cation Protocol
S. R. TURNER*{{, T. SENARATNA{, E. BUNN{, B. TAN{, K. W. DIXON{and D. H. TOUCHELL}
{Kings Park and Botanic Garden, West Perth, WA 6005, Australia, {Curtin University of Technology, Bentley,WA 6102, Australia and }USDA-ARS National Seed Storage Laboratory, 1111 South Mason St, Fort Collins,
Colorado 80521, USA
Received: 15 September 2000 Returned for revision: 3 November 2000 Accepted: 21 November 2000 Published electronically: 26 January 2001
somaclonal
0305-7364/0
* For corre
An e�cient vitri®cation procedure was developed and successfully applied to cryopreserve six endangered WestAustralian species ( family Haemodoraceae: Anigozanthos humilis ssp. chrysanthus Hopper; A. kalbarriensis Hopper;A. viridis ssp. terraspectans Hopper; Conostylis dielsia ssp. teres Hopper; C. micrantha Hopper and C. wonganensisHopper). Species were initially evaluated for cryostorage using a basic vitri®cation protocol involving: culturingplantlets in vitro for 21 d; excision of shoot apices; preculture of apical tips on 0.4 M sorbitol for 2 d, followed byincubation in PVS2 (plant vitri®cation solution 2) for 25 min at 0 8C, then direct immersion in liquid nitrogen (LN).Warming of retrieved material was for 1 min in a 40 8C water bath. Using this protocol ®ve of the six speciesexhibited low post-storage survival, while the sixth species, A. viridis ssp. terraspectans posted higher survival(61.1%). Using A. viridis ssp. terraspectans as an indicator species, the initial protocol was modi®ed to include: 3 dpreculture on 0.80 M glycerol, loading treatment with 2.0 M glycerol plus 0.4 M sucrose solution for 20 min, followedby 25 min exposure to a modi®ed PVS2. Survival was signi®cantly improved in the test species, and in furtherexperiments three other species also showed signi®cant improvements with the new protocol. Key ®ndings include:e�ectiveness of glycerol in the preculture medium; the e�ect of preculture duration; the importance of a loading stagefor these species; and the successful use of modi®ed PVS2 solutions with reduced or zero dimethyl sulfoxide(DMSO). # 2001 Annals of Botany Company
Key words: A. humilis ssp. chrysanthus, A. kalbarriensis, A. viridis ssp. terraspectans, Conostylis dielsia ssp. teres,C. micrantha, C. wonganensis, kangaroo paws, Haemodoraceae, vitri®cation, cryopreservation, rare and endangered,
conservation.INTRODUCTION
The Haemodoraceae (or `bloodroots') have representativetaxa on several continents, including Africa, South Americaand North America (Hopper, 1993). In the south west ofWestern Australia (WA) this family shows considerablediversi®cation with 77 species belonging to seven genera.The majority of these genera i.e. Macropidia (blackkangaroo paw), Anigozanthos (kangaroo paws), Blancoa,Conostylis, Phlebocarya and Tribonanthes are endemic tothis region (Hopper, 1993).
Amongst the endemic species of WA Haemodoraceae,many are classi®ed as rare and endangered, including sevenspecies of Conostylis and three subspecies of Anigozanthos(Brown et al., 1998).
Cryopreservation is currently being developed byagencies such as Kings Park and Botanic Garden (WestPerth, Australia) to conserve threatened species. Theadvantages of cryostorage over conventional forms ofgermplasm storage include: reduction in maintenancecosts associated with living collections; reduction in storagespace; minimization of losses due to diseases and con-tamination (with living collections); and minimization of
variation (Bajaj, 1995).
1/030371+08 $35.00/00
spondence. E-mail [email protected]
For cryopreservation of endangered species, shoot apicesare the preferred tissue source as they allow for theconservation of genetically stable tissues, which areessential for future re-introduction programs. Furthermore,cells found in the apical region of shoot apices are muchmore amenable to cryostorage protocols as they consist ofsmall thin-walled actively dividing cells that have smallvacuoles and a high nucleo-cytoplasmic ratio (Engelmann,2000).
Among several cryopreservation protocols, vitri®cation isa relatively simple method requiring no special coolingequipment (Benson, 1999). Most vitri®cation protocolsutilized are based on plant vitri®cation solution 2 (PVS2)developed by Sakai et al. (1990), which involve thefollowing steps: preculturing shoot apices on a sucroseenriched medium, loading treatment and dehydration withPVS2 prior to a plunge into liquid nitrogen (LN).Previously developed protocols for endangered Australianspecies involved preculturing shoot apices on a mediumcontaining sorbitol, followed by incubation in PVS2 beforedirect immersion in LN (Touchell and Dixon, 1999;Touchell et al., 2001).
In a previous study, Turner et al. (2000) achieved post-LN survival of 36% for Anigozanthos viridis ssp. viridis
when shoot apices were precultured on a 0.4 M sorbitol# 2001 Annals of Botany Company

at
medium for 2 d, before exposure to PVS2 for 25 min,followed by direct immersion in LN. This protocol was alsofound to be applicable to other Haemodoraceae species,although survival in all taxa tested was very low.
The objectives of this study were to examine post-LNsurvival of Haemodoraceae species in response to: (a)alterations to preculture duration; (b) sugars/polyalcoholsin the preculture medium; (c) vitri®cation solutions; and (d)
372 Turner et al.ÐCryopreserv
the addition of a loading step.
recovery medium.
improve the above protocol.
warmed, washed, and placed on recovery medium.
MATERIALS AND METHODS
Species and in vitro culture
The six species used in this study were: Conostylis dielsia ssp.teres,C. micrantha,C. wonganensis,Anigozanthos viridis ssp.terraspectans, A. humilis ssp. chrysanthus and A. kalbar-riensis. With the exception of A. humilis ssp. chrysanthus, allspecies were obtained from in vitro shoot culture stocksmaintained at the Plant Science Laboratory, Kings Park andBotanic Garden (Perth) and derived from ®eld collectedshoots. In the case of A. humilis ssp. chrysanthus, shootcultures were initiated from zygotic embryos.
In vitro plantlets were subcultured at 3 to 4 weeklyintervals on a basal medium (BM) containing half strengthMurashige and Skoog (MS) salts (Murashige and Skoog,1962) supplemented with 3.0 mM thiamine hydrochloride,2.5 mM pyridoxine hydrochloride, 4.0 mM niacin, 0.5 mM
MES bu�er, 0.5 mMmyo-inositol, 60 mM sucrose, 0.5 mM 6-benzylaminopurine (BAP) and solidi®ed with 0.8% (w/v)agar. The pH of the medium was adjusted to 5.9 prior to theaddition of agar, then autoclaved at 121 8C for 20 min.Plant cultures were incubated at 22±25 8C and illuminatedwith 16 h light with photosynthetic photon ¯ux density of
30 mmol mÿ2 sÿ1.same as in the previous experiment.
Evaluation of the previously developed protocol
In the ®rst experiment the e�cacy of the protocoldeveloped by Turner et al. (2000) was evaluated using thesix study species. Turner et al.'s protocol consisted of sixphases: culture, preculture, PVS2 exposure, LN immersion,warming and recovery. Shoot apices were harvested andevaluated following four key stages in the protocol: (a)immediately after harvest (placed directly onto recoverymedium without any treatment); (b) after preculture; (c)after preculture and PVS2 exposure; and (d) after preculture,PVS2 exposure, LN immersion, warming and washing.
In the preculture stage, shoot apices 1.0±1.5 mm longwhich were excised from 19±23-d-old shoots were pre-cultured on BM supplemented with 0.4 M sorbitol for 2 d.These shoot apices were then placed onto BM supple-mented with 2 mM choline chloride recovery medium (indarkness for 7 d) (Touchell et al., 2001), with or withoutincubation in PVS2 (and for a sub-sample, LN immersion).
Shoot apices at this stage were exposed to PVS2 at 0 8Cfor 25 min. PVS2 comprised 30% w/v glycerol, 15% w/vethylene glycol and 15%w/v dimethyl sulfoxide (DMSO) inhalf-strength MS liquid medium supplemented with 0.4 M
sucrose (Sakai et al., 1990). The medium was ®lter-sterilized
using a 0.2 mMAcrodisc1 32 ®lter prior to use. Treated shootapices were then washed three times for 4 min per rinse, inhalf-strength MS liquid basal medium containing 1.0 M suc-rose. After washing, shoot apices were placed onto recoverymedium to assess the survival prior to LN immersion.
Apices to be cooled were harvested, precultured, incu-bated in PVS2 as described above, plunged into LN andstored for at least 30 min prior to warming in a 40 8C waterbath for 1 min. After warming, shoot apices were removedfrom the PVS2 medium and washed as described pre-viously. After washing, shoot apices were placed onto
ion of Endangered Species
Comparison of shoot apex responses ( for six di�erentHaemodoraceae species) after direct exposure to PVS2
Shoot apices were harvested from all species and imme-diately incubated in PVS2. These apices were then rinsed in1 M sucrose solution and placed on recovery medium.
Subsequent experiments were designed to modify and
Comparison of preculture duration time
Shoot apices of A. viridis were precultured for 0, 1, 2, 3and 7 d on BM containing 0.4 M sorbitol, incubated inPVS2 at 0 8C, washed and placed on recovery medium(without placing in LN) or plunged directly into LN,
Comparison of di�erent sugars and polyalcohols in thepreculture medium
Seven di�erent sugars and polyalcohols were evaluated inpreculture media using Anigozanthos viridis shoot apices. Inthe ®rst experiment, shoot apices were precultured on BMsupplemented with either 0.4 M glucose, sucrose, trehalose,ra�nose, glycerol, inositol or sorbitol for 3 d, thenincubated in PVS2, cooled in LN, warmed and placed onrecovery medium as noted above. In the second experiment,shoot apices were placed on BM supplemented withdi�erent molarities of these compounds. Molarities wereformulated to provide the same equimolar concentrations ofhydroxyl (OH) groups as 0.4 M sorbitol (14.448 � 1023 lÿ1of preculture medium). This was formulated by taking intoaccount the total number of OH groups per molecule (e.g.sorbitol has six, glycerol has three) then adjusting theconcentration to provide the same number of OH groups as0.4 M sorbitol. Using this procedure the following concen-trations were determined: 0.480 M glucose, 0.300 M sucrose,0.300 M trehalose, 0.218 M ra�nose, 0.8 M glycerol, 0.400 M
inositol and 0.400 M sorbitol. Subsequent steps were the
Evaluation of di�erent cryoprotectant solutions
Harvested shoot apices from A. viridis were ®rstprecultured on BM supplemented with 0.8 M glycerol for3 d then incubated in di�erent cryoprotectant solutions for
25 min at 0 8C. The cryoprotectant solutions were: (1) PVS1
at
(19% w/v glycerol, 13% w/v ethylene glycol, 13% w/vpropylene glycol, 6%w/v DMSO in half-strengthMS liquidmedium �0.5 M sorbitol) (Uragami et al., 1989), (2) PVS2(30% w/v glycerol, 15% w/v ethylene glycol and 15% w/vDMSO in half-strengthMS liquid medium � 0.4 M sucrose)(Sakai et al., 1990), (3) Mod-1 PVS2 (30% w/v glycerol,15% w/v ethylene glycol, 7.5% w/v DMSO, 7.5%propylene glycol in half-strength MS liquid � 0.4 M
sucrose), (4) Mod-2 PVS2 (30% w/v glycerol, 15% w/vethylene glycol, 15% propylene glycol in half-strength MSliquid medium � 0.4 M sucrose), (5) PVS3 (50% w/vglycerol, 50% w/v sucrose in water) (Nishizawa et al.,1993), (6) Mod PVS3 (50% w/v glycerol, 50% w/v sucrose,5% DMSO in water) (Nishizawa et al., 1993).
The material was then rinsed and placed on recoverymedium, or cooled in LN, then warmed, rinsed and placed
Turner et al.ÐCryopreserv
on recovery medium.
observed for all species.
apices appeared `bleached' 2 to 3 d after treatment.
Comparison between the previous protocol and the modi®edprotocol
Using the optimal combination from the precultureduration experiment, preculture sugar/polyalcohol experi-ments and cryoprotectant solution experiment, all six studyspecies were evaluated using this modi®ed protocol. Shootapices were again evaluated after: (a) harvest (placeddirectly onto recovery medium without any treatment); (b)preculture; (c) preculture, PVS2 exposure and rinsing; and(d) preculture, PVS2 exposure, LN immersion, warmingand rinsing. In the modi®ed protocol, 0.8 M glycerolreplaced 0.4 M sorbitol, 3 d preculture replaced 2, andshoot apices were exposed to Mod-1 PVS2 for 25 min at0 8C. These results were then compared with results from
the previous protocol of Turner et al. (2000).reduced survival (P 5 0.05) compared to 3 d preculture.
Evaluation of a loading stage
For selected species following preculture, shoot apiceswere placed in 2 ml loading solution consisting of 2 M
glycerol, plus half-strength MS liquid culture medium (BM)supplemented with 0.4 M sucrose (Matsumoto et al., 1994;Charoensub et al., 1999). Shoot apices were incubated inthis medium for 20 min at room temperature before placingin Mod-1 PVS2. Shoot apices were then rinsed and placedon recovery medium or immersed in LN, warmed, rinsed
and placed on recovery medium.(P 5 0.05) (Fig. 2A).
Scoring of survival and regeneration
Each experimental treatment was repeated three times(three replicates) and each replicate consisted of 15 shootapices. Survival was determined as detectable growth of allor part of apical tissues 1±28 d after treatment. Recoverywas determined as continued growth of apices into shoots.
Data generated were analysed statistically by analysis ofvariance (ANOVA). The original data expressed as a pro-portion (%) were transformed (arcsin
p) to conform to
ANOVA assumptions. Mean comparisons were then made
using Fisher's protected LSD, at the 95% con®dence level.RESULTS
Responses of shoot apices ( for six di�erent Haemodoraceaespecies) to di�erent steps in the protocol
The response of shoot apices to each step in the protocolwas evaluated using the original protocol developed forA.viridis ssp. viridis by Turner et al. (2000). Across all sixspecies there was very high survival prior to the incorpora-tion of the preculture step (84.0±100.0%) (data not shown)but survival decreased signi®cantly with the addition of apreculture stage, PVS2 exposure and LN immersion(P 5 0.05). Although some survival following LN immer-sion was recorded for all six species, only A. viridis ssp.terraspectans showed acceptable levels of survival (61.1%)(data not shown). Shoot apices exposed to preculture orpreculture and PVS2, but not immersed in LN, displayedmore rapid growth rates (approximately twice those ofshoot apices cooled with LN) (data not shown). This was
ion of Endangered Species 373
Comparison of shoot apex responses for six di�erentHaemodoraceae species exposed to PVS2 directly afterexcision
For all six species, shoot apices that were excised andthen incubated in PVS2 without any preculture treatmentshad very low survival (0±2%) (data not shown), and all
Comparison of preculture duration time
Shoot apices of A. viridis ssp. terraspectans exposed to0 d preculture, treated with PVS2, and LN immersionshowed no survival, while non-LN immersed shoot apiceswith the same preculture duration (0 d) had very lowsurvival (3.0%) (Fig. 1). However following 1 d preculture,survival of both treated (non-cooled) and treated/cooledshoot apices increased signi®cantly (P 5 0.05) (Fig. 1). Forboth treatments, survival peaked after 3 d preculture.However, for treated shoot apices (non-cooled) andtreated/cooled shoot apices, preculture for 7 d signi®cantly
Comparison of di�erent sugars and polyalcohols at the samemolarity (0.4 M)
When compared at the same molarity, most of the sugarsand polyalcohols examined gave similar levels of survival;the highest survival was obtained with glycerol (77.6%),while the trehalose treatment resulted in signi®cantly lowersurvival (34.0%) than all other compounds examined
Comparison of di�erent sugars and polyalcohols with thesame number of hydroxyl groups
Molarity of the sugars in the preculture media wasadjusted in this experiment to yield a similar total numberof hydroxyl groups (14.448 � 1023) of all sugars and
polyalcohols. The number of hydroxyl groups was based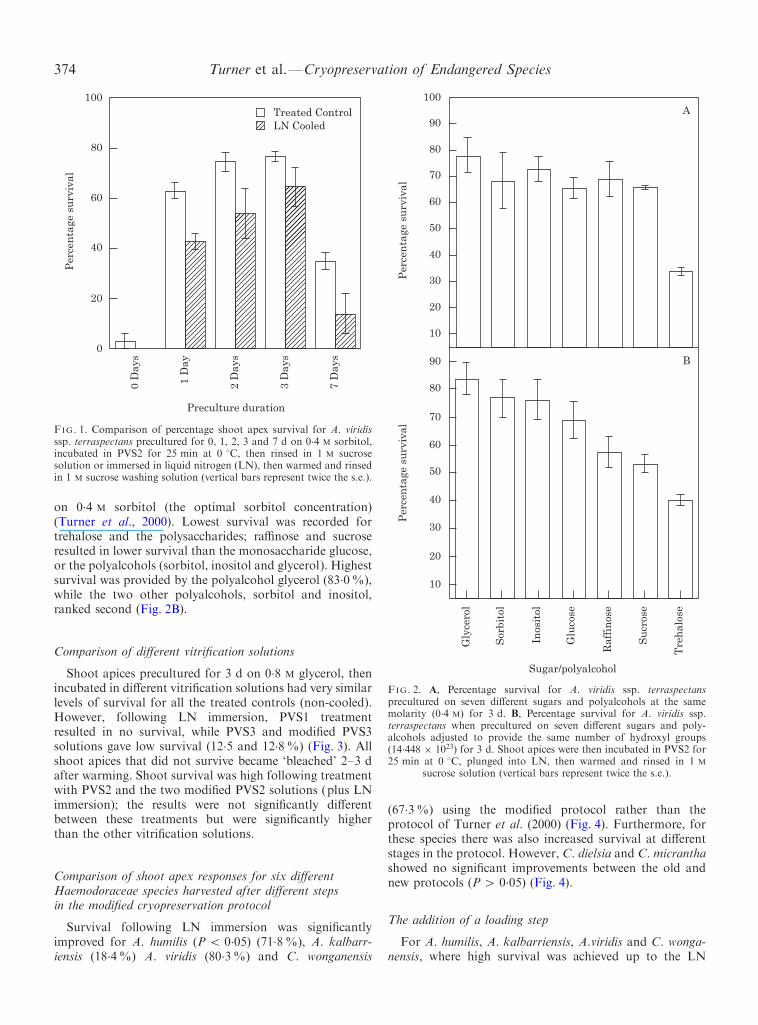
100
80
60
40
20
0
Treated ControlLN Cooled
Preculture duration
0 D
ays
1 D
ay
2 D
ays
3 D
ays
7 D
ays
Per
cen
tage
su
rviv
al
FIG. 1. Comparison of percentage shoot apex survival for A. viridisssp. terraspectans precultured for 0, 1, 2, 3 and 7 d on 0.4 M sorbitol,incubated in PVS2 for 25 min at 0 8C, then rinsed in 1 M sucrosesolution or immersed in liquid nitrogen (LN), then warmed and rinsed
A
B
100
90
80
70
60
50
40
30
20
10
90
80
70
60
50
40
30
20
10
Per
cen
tage
su
rviv
alP
erce
nta
ge s
urv
ival
374 Turner et al.ÐCryopreservation of Endangered Species
on 0.4 M sorbitol (the optimal sorbitol concentration)(Turner et al., 2000). Lowest survival was recorded fortrehalose and the polysaccharides; ra�nose and sucroseresulted in lower survival than the monosaccharide glucose,or the polyalcohols (sorbitol, inositol and glycerol). Highestsurvival was provided by the polyalcohol glycerol (83.0%),while the two other polyalcohols, sorbitol and inositol,
in 1 M sucrose washing solution (vertical bars represent twice the s.e.).
ranked second (Fig. 2B).
Sugar/polyalcohol
Gly
cero
l
Sor
bito
l
Inos
itol
Glu
cose
Raf
fin
ose
Su
cros
e
Tre
hal
ose
FIG. 2. A, Percentage survival for A. viridis ssp. terraspectansprecultured on seven di�erent sugars and polyalcohols at the samemolarity (0.4 M) for 3 d. B, Percentage survival for A. viridis ssp.terraspectans when precultured on seven di�erent sugars and poly-alcohols adjusted to provide the same number of hydroxyl groups(14.448 � 1023) for 3 d. Shoot apices were then incubated in PVS2 for25 min at 0 8C, plunged into LN, then warmed and rinsed in 1 M
sucrose solution (vertical bars represent twice the s.e.).
Comparison of di�erent vitri®cation solutions
Shoot apices precultured for 3 d on 0.8 M glycerol, thenincubated in di�erent vitri®cation solutions had very similarlevels of survival for all the treated controls (non-cooled).However, following LN immersion, PVS1 treatmentresulted in no survival, while PVS3 and modi®ed PVS3solutions gave low survival (12.5 and 12.8%) (Fig. 3). Allshoot apices that did not survive became `bleached' 2±3 dafter warming. Shoot survival was high following treatmentwith PVS2 and the two modi®ed PVS2 solutions (plus LNimmersion); the results were not signi®cantly di�erentbetween these treatments but were signi®cantly higher
than the other vitri®cation solutions.new protocols (P 4 0.05) (Fig. 4).
Comparison of shoot apex responses for six di�erentHaemodoraceae species harvested after di�erent stepsin the modi®ed cryopreservation protocolSurvival following LN immersion was signi®cantlyimproved for A. humilis (P 5 0.05) (71.8%), A. kalbarr-
iensis (18.4%) A. viridis (80.3%) and C. wonganensis(67.3%) using the modi®ed protocol rather than theprotocol of Turner et al. (2000) (Fig. 4). Furthermore, forthese species there was also increased survival at di�erentstages in the protocol. However, C. dielsia and C. micranthashowed no signi®cant improvements between the old and
The addition of a loading step
For A. humilis, A. kalbarriensis, A.viridis and C. wonga-
nensis, where high survival was achieved up to the LN
100
80
60
40
20
0
Per
cen
tage
su
rviv
al
PV
S1
PV
S2
PV
S3
Mod
PV
S3
(5%
DM
SO
)
Mod
PV
S2
(7. 5
% P
ropa
ndi
ol/7
. 5%
DM
SO
)
Mod
PV
S2
(15%
Pro
pan
diol
)
Treated ControlLN Cooled
FIG. 3. Percentage survival of A. viridis ssp. terraspectans shoot apicesexposed to di�erent vitri®cation solutions. Material was preculturedfor 3 d on 0.8 M glycerol, then incubated for 25 min at 0 8C in one ofsix di�erent vitri®cation solutions (PVS1, PVS2, Mod-1 PVS2, Mod-2PVS2, PVS3 and Mod PVS3), then rinsed in 1 M sucrose solution orimmersed in LN, warmed and rinsed in 1 M sucrose solution (vertical
bars represent twice the s.e.).
Turner et al.ÐCryopreservat
immersion stage, a loading phase was added. For three ofthese species post-LN survival was higher (P 5 0.05) withthe use of a loading phase (Fig. 5). Furthermore, signs ofsurvival were ®rst noticed 1±2 d earlier for the loadedsamples, while the initial rate of growth for these plants wasalso more rapid (data not shown). This was observed for all
species.DISCUSSION
The results presented in this study demonstrate the e�cacyof using an indicator species (in this case Anigozanthosviridis ssp. terraspectans) to optimize cryopreservationprotocols that can later be successfully applied to other
related species. A. viridis ssp. terraspectans was chosen asthe indicator species due to its high multiplication rate, andbecause the initial protocol was developed on the closelyrelated species A. viridis ssp. viridis (Turner et al., 2000). Incomparison, the other species in this study had lowmultiplication rates, or insu�cient numbers which impededlarge scale experimentation.
All species examined with the original protocol had post-LN survival; however, survival was low for most species,indicating that the protocol was sub-optimal. This wascon®rmed when the protocol steps were examined, with allspecies exhibiting a sharp decline in survival prior to LNimmersion. Therefore, to increase post-LN survival, adjust-ments were needed to produce higher survival in thepreculture and PVS incubation phases.
The importance of preculture duration is obvious whenshoot apices from all species were excised and incubated inPVS2 for 25 min without preculture. Four of the six speciesdid not survive, and survival was only 2% for the other twospecies. These observations suggest that shoot apices needsu�cient preculture (at least 24 h) to acquire osmotoleranceto PVS2 prior to being plunged into LN. Furthermore, forA. viridis ssp. terraspectans an increasing trend in post-LNsurvival was seen from 1 to 3 d preculture on sorbitol.Optimal preculture duration may vary for di�erent species.For example, Grevillea scapigera (Touchell, 1995) andMacropidia fuliginosa (another Haemodoraceae species)achieved maximum post-LN survival following 2 d pre-culture, while Hopkinsia anoectocolea, another nativemonocot species, had the highest post-LN survival(46.4%) after preculturing shoot apices on 0.2 M sorbitolfor 3 d before incubation in PVS2 for 25 mins at 0 8C(unpubl. obs.).
Preculture on di�erent sugars and polyalcohols of thesame molarity resulted in similar survival patterns exceptfor the trehalose treatment which produced signi®cantlylower survival. This was unexpected as trehalose has beenextensively found in a diverse range of desiccation-tolerantorganisms (Crowe and Crowe, 1986). However, its use inplant systems has been limited (Bhandal et al., 1985; Reed,1996).
Of the other sugars/polyalcohols examined, glyceroltreatments resulted in higher survival both at the samemolarity (0.4 M) and when used with the same number ofhydroxyl groups (0.8 M) as sorbitol (Fig. 2), although thisdi�erence was not signi®cant. When treatments contain thesame number of hydroxyl groups, glycerol is present at aconcentration of 0.8 M which is twice the concentration ofsorbitol, the polyalcohol used in the original protocol. Theoriginal study on A. viridis ssp. viridis found that 0.6 M
sorbitol resulted in signi®cantly lower levels of survival(Turner et al., 2000).
The mode of superior action for glycerol is not clear. It isthought that the hydroxyl groups present in sugars/polyalcohols interact by hydrogen bonding with membranephospholipids, imparting membrane stabilization duringdehydration (Crowe and Crowe, 1986). Phunchindawanet al. (1996) and Thammasiri (1999) achieved high survivalusing a mixture of glycerol and sucrose in the preculturemedium for horseradish hairy root cultures (at 1.0 M) and
ion of Endangered Species 375
for embryonic axes of jackfruit (at 0.5 M).

A B C DA B C DTreatment Treatment
100
80
60
40
20
0
100
80
60
40
20
0
100
80
60
40
20
0
100
80
60
40
20
0A B C D A B C D* *
Treatment Treatment
100
80
60
40
20
0
100
80
60
40
20
0A B C DA B C D
Treatment Treatment
A B
C D
E F
Per
cen
tage
su
rviv
al
Per
cen
tage
su
rviv
alP
erce
nta
ge s
urv
ival
Per
cen
tage
su
rviv
al
Per
cen
tage
su
rviv
al
Per
cen
tage
su
rviv
al
FIG. 4. Comparison of percentage shoot apex survival between the original and modi®ed protocol for A. humilis ssp. chrysanthus (A),A. kalbarriensis (B) and A. viridis ssp. terraspectans (C), C. dielsia ssp. teres (D), C. micrantha (E), and C. wonganensis (F) after di�erent steps inthe cryopreservation protocol: A, no treatment ( freshly excised shoot apices were placed directly onto recovery medium); B, preculture on 0.4 M
sorbitol or 0.8 M glycerol for 2 or 3 d; C, preculture on 0.4 M sorbitol or 0.8 M glycerol for 2 or 3 d, then incubation in 25 min PVS2 or 25 minMod-1 PVS2, then rinsed in 1 M sucrose solution; D, preculture on 0.4 M sorbitol or 0.8 M glycerol for 2 or 3 d, then incubation in 25 min PVS2or 25 min Mod-1 PVS2, then immersion in LN and ®nally warmed and rinsed in 1 M sucrose solution (vertical bars represent twice the s.e.). h,
i
376 Turner et al.ÐCryopreservation of Endangered Species
Successful utilization of di�erent plant vitri®cationsolutions such as PVS1 (Uragami et al., 1989), PVS2(Sakai et al., 1990) and PVS3 (Nishizawa et al., 1993) arereported in the literature, but PVS2 is the most commonlyused (Charoensub et al., 1999; Thammasiri, 1999; Thinhet al., 1999; Sakai et al., 2000).
Results achieved from the PVS experiment demonstratedthat before LN immersion very high survival was achieved
original; F, mod
for apices vitri®ed in the vitri®cation solutions tested.
However, following LN immersion only the PVS2 andmodi®ed PVS2 solutions resulted in high post-LN survival.In fact, plants cryopreserved in the three PVS2 andmodi®ed PVS2 solutions had very similar survival(Fig. 3)Ðeven in the Mod-2 PVS2 solution withoutDMSO. The replacement component propylene glycol hasbeen reported to have similar properties to DMSO in thatcell membranes are highly permeable to it and it has a low
®ed; *, no data.
molecular weight (Boucaud and Brison, 1995), but it has
Australia and elsewhere.
Hopper SD. 1993. Kangaroo paws and cats paws. Perth: Department of
100
80
60
40
20
0
Per
cen
tage
su
rviv
al
Non-LoadedLoaded
Species
A. h
um
ilis
A. k
alba
rrie
nsi
s
A. v
irid
is
C. w
onga
nen
sis
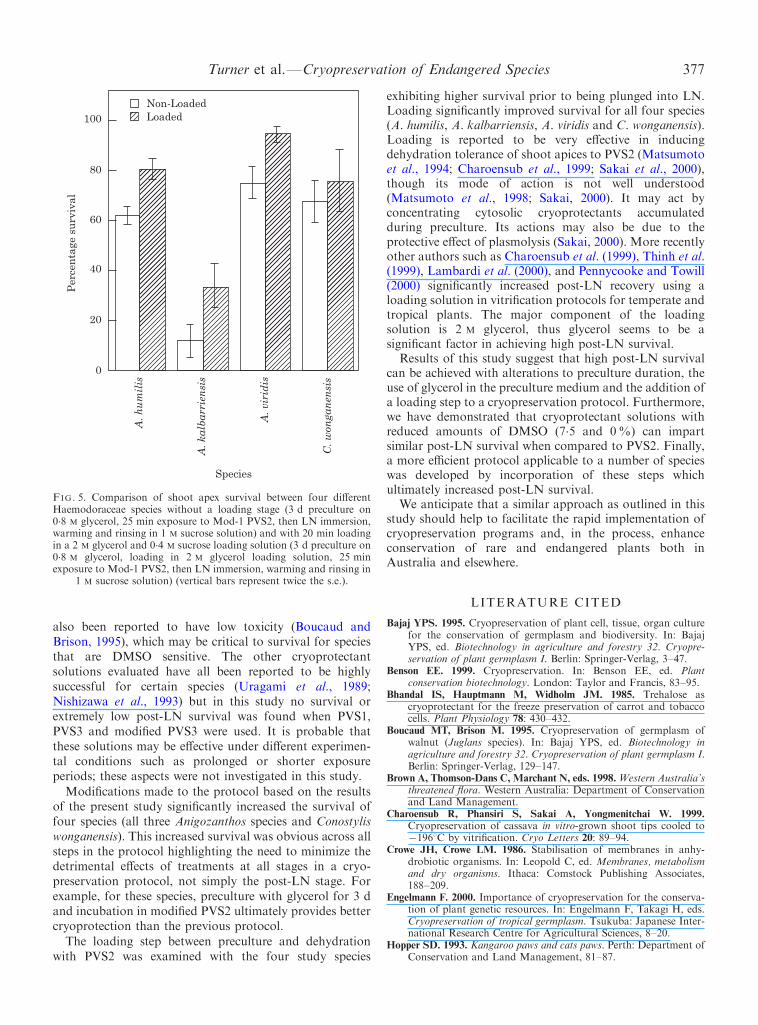
FIG. 5. Comparison of shoot apex survival between four di�erentHaemodoraceae species without a loading stage (3 d preculture on0.8 M glycerol, 25 min exposure to Mod-1 PVS2, then LN immersion,warming and rinsing in 1 M sucrose solution) and with 20 min loadingin a 2 M glycerol and 0.4 M sucrose loading solution (3 d preculture on0.8 M glycerol, loading in 2 M glycerol loading solution, 25 minexposure to Mod-1 PVS2, then LN immersion, warming and rinsing in
1 M sucrose solution) (vertical bars represent twice the s.e.).
Turner et al.ÐCryopreservat
also been reported to have low toxicity (Boucaud andBrison, 1995), which may be critical to survival for speciesthat are DMSO sensitive. The other cryoprotectantsolutions evaluated have all been reported to be highlysuccessful for certain species (Uragami et al., 1989;Nishizawa et al., 1993) but in this study no survival orextremely low post-LN survival was found when PVS1,PVS3 and modi®ed PVS3 were used. It is probable thatthese solutions may be e�ective under di�erent experimen-tal conditions such as prolonged or shorter exposureperiods; these aspects were not investigated in this study.
Modi®cations made to the protocol based on the resultsof the present study signi®cantly increased the survival offour species (all three Anigozanthos species and Conostyliswonganensis). This increased survival was obvious across allsteps in the protocol highlighting the need to minimize thedetrimental e�ects of treatments at all stages in a cryo-preservation protocol, not simply the post-LN stage. Forexample, for these species, preculture with glycerol for 3 dand incubation in modi®ed PVS2 ultimately provides bettercryoprotection than the previous protocol.
The loading step between preculture and dehydration
with PVS2 was examined with the four study speciesexhibiting higher survival prior to being plunged into LN.Loading signi®cantly improved survival for all four species(A. humilis, A. kalbarriensis, A. viridis and C. wonganensis).Loading is reported to be very e�ective in inducingdehydration tolerance of shoot apices to PVS2 (Matsumotoet al., 1994; Charoensub et al., 1999; Sakai et al., 2000),though its mode of action is not well understood(Matsumoto et al., 1998; Sakai, 2000). It may act byconcentrating cytosolic cryoprotectants accumulatedduring preculture. Its actions may also be due to theprotective e�ect of plasmolysis (Sakai, 2000). More recentlyother authors such as Charoensub et al. (1999), Thinh et al.(1999), Lambardi et al. (2000), and Pennycooke and Towill(2000) signi®cantly increased post-LN recovery using aloading solution in vitri®cation protocols for temperate andtropical plants. The major component of the loadingsolution is 2 M glycerol, thus glycerol seems to be asigni®cant factor in achieving high post-LN survival.
Results of this study suggest that high post-LN survivalcan be achieved with alterations to preculture duration, theuse of glycerol in the preculture medium and the addition ofa loading step to a cryopreservation protocol. Furthermore,we have demonstrated that cryoprotectant solutions withreduced amounts of DMSO (7.5 and 0%) can impartsimilar post-LN survival when compared to PVS2. Finally,a more e�cient protocol applicable to a number of specieswas developed by incorporation of these steps whichultimately increased post-LN survival.
We anticipate that a similar approach as outlined in thisstudy should help to facilitate the rapid implementation ofcryopreservation programs and, in the process, enhanceconservation of rare and endangered plants both in
ion of Endangered Species 377
LITERATURE CITED
Bajaj YPS. 1995. Cryopreservation of plant cell, tissue, organ culturefor the conservation of germplasm and biodiversity. In: BajajYPS, ed. Biotechnology in agriculture and forestry 32. Cryopre-servation of plant germplasm I. Berlin: Springer-Verlag, 3±47.
Benson EE. 1999. Cryopreservation. In: Benson EE, ed. Plantconservation biotechnology. London: Taylor and Francis, 83±95.
Bhandal IS, Hauptmann M, Widholm JM. 1985. Trehalose ascryoprotectant for the freeze preservation of carrot and tobaccocells. Plant Physiology 78: 430±432.
Boucaud MT, Brison M. 1995. Cryopreservation of germplasm ofwalnut (Juglans species). In: Bajaj YPS, ed. Biotechnology inagriculture and forestry 32. Cryopreservation of plant germplasm I.Berlin: Springer-Verlag, 129±147.
Brown A, Thomson-Dans C, Marchant N, eds. 1998.Western Australia'sthreatened ¯ora. Western Australia: Department of Conservationand Land Management.
Charoensub R, Phansiri S, Sakai A, Yongmenitchai W. 1999.Cryopreservation of cassava in vitro-grown shoot tips cooled toÿ1968C by vitri®cation. Cryo Letters 20: 89±94.
Crowe JH, Crowe LM. 1986. Stabilisation of membranes in anhy-drobiotic organisms. In: Leopold C, ed. Membranes, metabolismand dry organisms. Ithaca: Comstock Publishing Associates,188±209.
Engelmann F. 2000. Importance of cryopreservation for the conserva-tion of plant genetic resources. In: Engelmann F, Takagi H, eds.Cryopreservation of tropical germplasm. Tsukuba: Japanese Inter-national Research Centre for Agricultural Sciences, 8±20.
Conservation and Land Management, 81±87.

at
Lambardi M, Fabbri A, Caccavale A. 2000. Cryopreservation of whitepoplar (Populus alba L.) by vitri®cation of in vitro-grown shoottips. Plant Cell Reports 19: 213±218.
Matsumoto T, Sakai A, Nako Y. 1998. A novel preculture forenhancing the survival of in vitro-grown meristems of wasabi(Wasabi japonica) cooled to ÿ1968C by vitri®cation. Cryo-Letters19: 27±36.
Matsumoto T, Sakai A, Yamada K. 1994. Cryopreservation of in vitro-grown apical meristems of wasabi (Wasabia japonica) by vitri®ca-tion and subsequent high plant regeneration. Plant Cell Reports13: 442±446.
Murashige T, Skoog F. 1962. A revised medium for rapid growth andbio-assay with tobacco tissue cultures. Physiologia Plantarium 15:473±497.
Nishizawa S, Sakai A, Amano Y, Matsuzawa T. 1993. Cryopreservationof asparagus (Asparagus o�cinalis L.) embryonic suspension cellsand subsequent plant regeneration by vitri®cation. Plant Science91: 67±73.
Pennycooke JC, Towill LE. 2000. Cryopreservation of shoot tips fromin vitro plants of sweet potato (Ipomoea batatas (L) Lam.) byvitri®cation. Plant Cell Reports 19: 733±737.
Phunchindawan M, Hirata K, Sakai A, Miyamoto K. 1996. Cryopre-servation of encapsulated shoot primordia induced in horseradish(Armoracia rusticana) hairy root cultures. Plant Cell Reports 16:469±473.
Reed BM. 1996. Pre-treatment strategies for cryopreservation of planttissues. In: Normah MN, Narimah MH, Clyde MM, eds.Techniques in in vitro conservation. Proceedings of the inter-national workshop on in vitro conservation of plant geneticresources, July 4±6 1995, Kuala Lumpur. Universiti Kebangsaan
378 Turner et al.ÐCryopreserv
Malaysia Publishers, 73±87.
Sakai A. 2000. Development of cryopreservation techniques. In:Engelmann F, Takagi H, eds. Cryopreservation of tropicalgermplasm. Tsukuba: Japanese International Research Centrefor Agricultural Sciences, 1±7.
Sakai A, Kobayashi S, Oiyama I. 1990. Cryopreservation of nucellarcells of naval orange (Citrus sinesis Osb. var. brasiliensis Tanaka)by vitri®cation. Plant Cell Reports 9: 30±33.
Sakai A, Matsumoto T, Hirai D, Niino T. 2000. Newly developedencapsulation-dehydration protocol for plant cryopreservation.Cryo-Letters 21: 53±62.
Thammasiri K. 1999. Cryopreservation of embryonic axes of jackfruit.Cryo-Letters 20: 21±28.
Thinh NT, Takagi H, Yashima S. 1999. Cryopreservation of in vitrogrown shoot tips of banana (Musa sp) by vitri®cation method.Cryo-Letters 20: 163±174.
Touchell DH. 1995. Principles of Cryobiology for Conservation ofThreatened Species. PhD thesis, University of Western Australia,Perth.
Touchell DH, Dixon KW. 1999. In vitro preservation. In: Bowes BG, ed.A colour atlas of plant conservation and propagation. London:Manson Publishing Ltd, 108±118.
Touchell DH, Turner SR, Bunn E, Dixon KW. 2001. Cryostorage ofsomatic tissues of endangered Australian species. In: Bajaj YPS,ed. Biotechnology in agriculture and forestry 32. Cryopreservationof plant germplasm II. Berlin: Springer-Verlag.
Turner SR, Touchell DH, Dixon K, Tan B. 2000. Cryopreservation ofAnigozanthos viridis ssp. viridis and related taxa from the southwest of Western Australia. Australian Journal of Botany 48:739±744.
Uragami A, Sakai A, Nagai M, Takahash T. 1989. Survival of cells andsomatic embryos of Asparagus o�cinalis cryopreserved by
ion of Endangered Species
vitri®cation. Plant Cell Reports 8: 418±421.
Copyright © 2022 FDOKUMEN




















