
Promotion of asparagus shoot and root growth by growth retardants
14
Plant Cell, Tissue and Organ Culture 11:97-110 (1987) © Martinus Nijhoff Publishers, Dordrecht - Printed in the Netherlands 97 Promotion of asparagus shoot and root growth by growth retardants ALISA KHUNACHAK, CHEE-KOK CHIN,* TRANG LE & TOM GIANFAGNA Department of Horticulture and Forestry, Rutgers University, Cook College, P.O. Box 231, Blake Hall, New Brunswick, NJ 08903, USA (*addressee for correspondence) Received 18 February 1986; in revised form 1 June i987; accepted 3 June i987 Key words: asparagus, growth retardants, gibberellins, ancymidol Abstract. Plantlets regenerated from shoot-tip culture of Asparagus officinalis L. possessed weak shoots and roots. Various combinations of auxins and cytokinins did not improve the plantlets. Incorporation of a number of growth retardants, viz. ancymidol, B-995, phosfon, Amo 1618, cycocel and paclobutrazol, promoted growth of stronger shoots and roots. The effectiveness of the growth retardants varied, with ancymidol being most effective and cycocel least effective. The response to ancymidol was prevented by exogenous GA 3 and GA4! 7. GA~/3 and GA4/7-1ike activities were detected in asparagus shoot-tip culture and these activities were reduced by the presence of the growth retardants ancymidol, Amo-1618, and cycocel. Introduction Asparagus plantlets can be regenerated from shoot-tip culture on Murashige and Skoog's medium containing kinetin and NAA [1]. However, these cul- tures often develop excessive callus and produce thin, weak roots, or even no roots at all. Only plantlets with vigorous roots but not those with thin roots survive transfer to soil [2, 3]. Thus, unlike propagation of many other plants, propagation of asparagus by tissue culture is critically dependent upon formation of thick vigorous roots by the cultures. Usually auxin and cytokinin are used to manipulate regeneration of shoots and roots in tissue culture. Recently we observed that ancymidol, a plant growth retardant, promoted growth of vigorous shoots and roots and suppressed callus formation [2]. In addition to ancymidol a number of other growth retardants, compounds which affect plants by reducing stem elonga- tion without producing abnormalities, are available. The purposes of this investigtion were to find out: (1) whether the effect of ancymidol on asp- aragus culture development was replaceable by auxin/cytokinin combina-
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Promotion of asparagus shoot and root growth by growth retardants

Plant Cell, Tissue and Organ Culture 11:97-110 (1987) © Martinus Nijhoff Publishers, Dordrecht - Printed in the Netherlands 97
Promotion of asparagus shoot and root growth by growth retardants
ALISA KHUNACHAK, CHEE-KOK CHIN,* TRANG LE & TOM GIANFAGNA Department of Horticulture and Forestry, Rutgers University, Cook College, P.O. Box 231, Blake Hall, New Brunswick, NJ 08903, USA (*addressee for correspondence)
Received 18 February 1986; in revised form 1 June i987; accepted 3 June i987
Key words: asparagus, growth retardants, gibberellins, ancymidol
Abstract. Plantlets regenerated from shoot-tip culture of Asparagus officinalis L. possessed weak shoots and roots. Various combinations of auxins and cytokinins did not improve the plantlets. Incorporation of a number of growth retardants, viz. ancymidol, B-995, phosfon, Amo 1618, cycocel and paclobutrazol, promoted growth of stronger shoots and roots. The effectiveness of the growth retardants varied, with ancymidol being most effective and cycocel least effective.
The response to ancymidol was prevented by exogenous GA 3 and GA4! 7. GA~/3 and GA4/7-1ike activities were detected in asparagus shoot-tip culture and these activities were reduced by the presence of the growth retardants ancymidol, Amo-1618, and cycocel.
Introduction
Asparagus plantlets can be regenerated from shoot-tip culture on Murashige and Skoog's medium containing kinetin and NAA [1]. However, these cul- tures often develop excessive callus and produce thin, weak roots, or even no roots at all. Only plantlets with vigorous roots but not those with thin roots survive transfer to soil [2, 3]. Thus, unlike propagation of many other plants, propagation of asparagus by tissue culture is critically dependent upon formation of thick vigorous roots by the cultures.
Usually auxin and cytokinin are used to manipulate regeneration of shoots and roots in tissue culture. Recently we observed that ancymidol, a plant growth retardant, promoted growth of vigorous shoots and roots and suppressed callus formation [2]. In addition to ancymidol a number of other growth retardants, compounds which affect plants by reducing stem elonga- tion without producing abnormalities, are available. The purposes of this investigtion were to find out: (1) whether the effect of ancymidol on asp- aragus culture development was replaceable by auxin/cytokinin combina-

98
tions, and/or other growth retardants, and (2) to examine whether ancy- midol exerted its effect through the inhibition of gibberellin (GA) synthesis.
Materials and Methods
Plant materials
Young spears from greenhouse-grown asparagus, Asparagus officinalis L., Clone 56, a pistillate selection, were used as the source of explants.
Surface sterilization
The explants were sterilized with 15% Clorox for 15 minutes, followed by 4 rinses with autoclaved distilled water. Shoot tips, 0.1-0.3 mm in length with a few leaf primordia, were excised aseptically.
Culture medium
The basal medium used in all experiments contained inorganic salts and vitamins according to Murashige and Skoog [4], 3% sucrose, and 0.5% Difco Bacto agar. The media were adjusted to pH 5.7 + 0.1 with either 1N NaOH or 1N HC1, and autoclaved at 15 psi, 121°C for 15 minutes. In experiments with growth regulators, combinations of auxins indoleacetic acid (IAA), indolebutyric acid (IBA), naphthaleneacetic acid (NAA), and cytokinins dimethylallyladenine (DMAA), kinetin, benzyladenine (BA), at 0, 0.125, 0.25, 0.5, 1.0/tM were used. In experiments with growth retardants, the media contained 0.5 #M NAA, 0.5/tM DMAA, and various concentra- tions of ancymidol, (from Elanco, Indianapolis, Indiana) paclobutrazol, (from Sandoz, Chicago, Illinois), phosphon, (from Mobil Chem. Co., Ash- land, Virginia) B-995, (from Uniroyal, Naugatuck, Connecticut), Amo- 1618 (from Calbiochem, Los Angeles, California) and cycoeel (from American Cyanamid, Wayne, New Jersey). The toxic level of each growth retardant was established in the preliminary experiments. The concentrations below the toxic levels were used in this investigtion. Growth regulators and growth retardants were added to the media prior to autoclaving.
Thirty ml of medium was dispensed in sterile plastic petri dishes (15 x 100mm). Shoots tips were placed on the surface of the solidified media and the petri dishes were sealed with Parafilm and maintained at 26°C under Cool White fluorescent light at 19/~mol m-2s -l

99
Determination of Growth Response: After 6 weeks, the length and thick- ness of shoot and roots were measured. Each treatment consisted of 4 dishes, each with 10 shoot tips. Very little variations were found among cultures in a dish. All experiments were repeated twice. The data collected were analyzed by an analysis of variance and the test of significance was deter- mined by Duncan's Multiple Range Test at the 5% level. Other characteris- tics such as callus formation, evidence of chlorosis, and general vigor of plants were also observed.
Extraction and purifiation of GA
GA-like compounds were extracted from 6-week old cultures. The extrac- tion and purification of gibberellin-like compounds from the plant material was adapted from Powell and Tautvydas [5]. In every treatment, 10 grams of fresh tissue were homogenized in about 100ml of cold 80% methanol (v/v) in a Waring blender. The slurry was stirred in a flask overnight at 5 °C, then vacuum-filtered through Celite (Fisher, Fairlawn N J). The filtrate was evaporated to the aqueous phase in vacuo at 35 °C with a Buchi rotary evaporator. Phosphate buffer (0.5M at pH 8.5) was added to the aqueous extract to make up the volume to 150 ml. The extract was then partitioned against 4 x 75 ml of hexane. The hexane was discarded and the aqueous phase, after adjusting the pH to 3.0 with 3N HC1, was loaded on a charcoal (Sigma, St. Louis, MO) celite (1:2) column (25 x 300 ram). The column was then washed with 30 ml distilled water and the GA-like activity was eluted with 120ml 80% acetone. The eluate was evaporated in vacuo at 35 °C to the aqueous phase and partitioned 3 times against equal-volumes of ethyl acetate. The combined ethyl acetate extracts was dried over anhydrous sodium sulfate, and concentrated in vacuo at 35 °C. 1-2 ml of ethyl acetate was added to the residue which was then spotted on a disc containing 6 layers of fiberglass filter paper (Whatman brand 934-AH, 2.4 CM dia.) and dried. The discs were placed on top of a silicic acid column prepared from 4.3 g silicic acid (Fisher, Fairlawn, N J) powder hydrated with 2.7 ml of 0.5 M formic acid. The column was eluted with hexane saturated with 0.5 M formic acid followed by saturated hexane containing ethyl acetate increased in 3 % v/v steps under pressurized nitrogen gas. Each fraction (1 ml) was collected in optically clear glass specimen vial and dried overnight in a fume hood. The next day 0.5 ml ethyl acetate was added to each vial, shaken and dried again. Samples were then subjected to the lettuce hypocotyl bioassay [6]. GA-like activity was quantified using a standard curve generated by measur- ing hypocotyl length in the presence of known amounts of authentic GA3 or GA4/7 (Sigma, St. Louis, MO).

100
Results
Combinations of auxins and cytokinins
The effects of various combinations of auxins and cytokinins on shoot formation are shown in Table 1 and on root formation in Table 2. Shoot formation took place in almost every treatment. Multiple shoots ranging from 3 to 10 were found in most media containing NAA and IBA in combination with DMAA. Root development was found in media contain- ing NAA in combination with kinetin or DMAA. The quality of the roots was poor; they were thin with average diameter less than 0.Smm and yellowish. Among all the combinations the best plantlets were obtained with 0.5 #M NAA and 0.5/~M DMAA (Table 1), but the shoots and roots of these plantlets were still not vigorous; the diameters of the roots and shoots were approximately 0.5 mm.
Effect of ancymidol (~-cyclopropyl-~-(p-methoxyphenyl)-5-pyrimidine methylalcohol)
The incorporation of ancymidol greatly promoted stronger shoot and root development and suppresed callus formation (Fig. 1). At low range of concentrations (0.125-0.5 mg L -1 , ancymidol stimulated shoot thickening. Roots were slightly longer and much thicker than the control. In the range of 1.'0 to 2.0 mg L -~ , shoots were very short and thick and roots were long and fleshy in appearance. Callus formation was completely suppressed at these concentrations. At 4 mg L 1, shoot length was severely reduced, and only a few short stunted roots were produced. At 8 mg L -1, ancymidol suppressed the growth of both shoot and root totally.
Effect of paclobutrazol (PP-333, 2-RS,3-RS)-4-chlorophenyl-4-4-dimethyl- 2- (1,2 ,3-triazol- l-yl ) pentan-3-O1
Paclobutrazol at low concentrations (0.125-0.25 mg L -1) did not show any apparent effect on shoot or root growth (Fig. 2). The thickness of the shoots increased slightly while the length remained the same as the concentrations of paclobutrazol increased from 0.5 to 1.0 mg L -~ . The roots, however, were longer and thicker and the number increased over the control. Optimum effects were obtained between 2.0 to 4.0 mg L -1 where the shoots and roots were much thicker. Higher concentrations (8.0-16.0mg L -~) severely in- hibited shoot and root growth. All treatments reduced callus production.

101
o
&
0
0 0
, .0
,-~
0
<
n~
<
0
o
e~
< < Z
<
.4
~ o
~-~
0
0
c5
0
,q.
o
~L
4 - 4 - 4 - 1 I
+ + + ÷ ÷
+ ÷ + + ÷ + +
+ + ÷ ÷ + ÷ + + +
+ + + ÷ +
÷ + + + + ÷ +
÷ ÷ + + + +
+ + + + +
+ + + + +
+ ÷ ÷ + +
+ + ÷ + +
+ + ÷ + +
+ + + + +
+ + ÷ + +
+ + + + +
<
I I
+ I
+ +
+ ÷ ÷ + +
+
+ + + I I
+ + ÷ ÷ + + + + + +
+ + + + +
÷ + + + +
÷ + + + +
+ + + + +
+ + + + + + + +
+ + + + +
÷ + + + + + +
÷ + + ÷ +
+ + + + + ÷ ÷ + + + ÷ ÷
+ ÷
+ t + + ÷ + + + ÷ ÷
+ + + + ÷
< < <
Z
I l l
+ + l
+ + +
+ ÷ l
÷ ÷ ÷
÷ + + + ÷ ÷ +
÷ + + ÷ + ÷ +
+ ÷ + + ÷ +
0 0
+ + + ÷ +
+ + + ÷ +
0
E + II + + + ÷
+ + + ÷ + + ÷ +
+ + + ÷ o
+ + + + +
+ + + + II + + + + + +
÷
0 + + ÷ o
÷ ÷ ÷ ÷ +
I
I + + + + II ÷
~ ° o
II
I
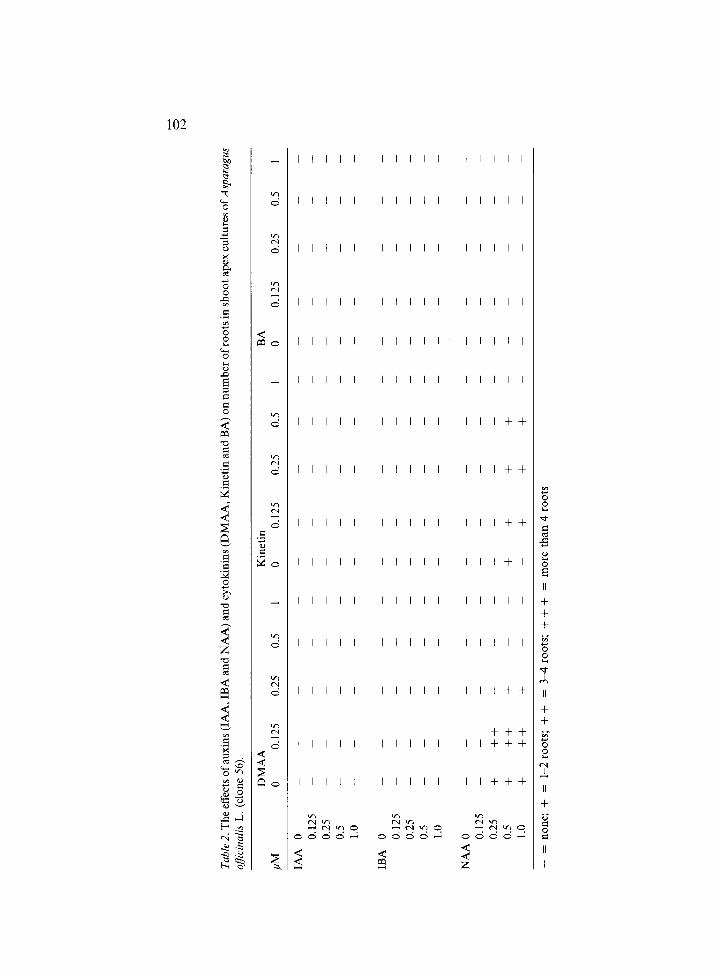
102
o
0
0 o
,.o
0
<
<
< < Z
<
<
0
0
0
c5
0
c5
0
+ +
+ +
0 o
+ l o
II I I +
+ +
0 o
+ + II + +
+ + +
+ + + o o
I + + +
II +
~. ~. ~. o. o
"~ II
Z t

103
Fig. 1. Effects of ancymidol on asparagus shoot-apex culture.
Fig. 2. Effects of paclobutrazol 333 on asparagus shoot-apex culture.

104
Effect of phosfon (chlorphonium chloride, tributyl-2, 4-dichlorobenzyl phosphonium chloride)
At concentrations of 0.0007 and 0.0015mg L -~, phosfon reduced shoot length (Fig. 3). Increasing the concentrations from 0.0015 to 0.006 mg L- resulted in an increase in shoot length. Further increase in phosfon con- centrations reduced the shoot length again. Root development was sti- mulated by phosfon from 0.006 to 0.025 mg L -1 . From 0.006 to 0.05 mg L -~ callus formation was suppressed. Optimal growth was obtained with 0.025 mg L -~ of phosfon.
Effect of B995 (Daminozide, N-dimethylamino succinamic acid)
At low concentrations (0.125-0.5 mg L -1) (Fig. 4) B-995 stimulated callus formation without any apparent effect on the length of shoots or roots. Increasing B-995 levels from 0.5 to 1.0mg L -1 resulted in a stimulation of shoot elongation without any change in root growth. At 2.0 mg L- i shoot length was reduced significantly (Table 3), but more roots were formed. Inhibition of shoot and root was observed at concentrations higher than 2mg L -~. Callus formation was reduced at 2mg L -1 .
Fig. 3. Effects of phosphon on asparagus shoot-apex culture.

105
Fig. 4. Effects of B-995 on asparagus shoot-apex culture.
Table 3. The effects of optimal concentration of growth retardants on shoot and root growth in shoot apex cultures of Asparagus offieinalis L.
Growth retardants Shoot Root (at optimum conc.) length thickness length thickness
mm mm
None 16.10a 0.45c 5.43c 0.64c 1.0 mg L- ~ Ancymidol 7.27d 1.10a 8.16a 1.18a 4.0 mg L t Paclobutrazol 7.08d 1.01a 7.80b 1.03a 0.025mg L -~ Phosfon 7.46d 0.98a 8.61a 0.83b 2.0rag L ~ B-995 8.86bc 0.67bc 6.87bc 0.81b 0.125 mg L -~ Amo-1618 16.70a 0.75b 7.39b 0.81b 0.25 mg L ~ Cycocel 13.75b 0.50c 7.66b 0.70bc
The quantitative data for the effects of growth retardants on shoot and root growth were analysed for all concentrations tested. However, for each growth retardant only the results for the concentration producing the maximal effects are presented in this table. Mean separation in columns by Duncan's multiple range test, 5% level.
Effect of amo-1618 (2-isopropyl-4-dimethylamino-5-methylphenyl-1- piperidine carboxylate methyl chloride)
The th ickness o f bo th , shoots a n d roots were s t imula t ed by A m o - 1 6 1 8 (Fig.
5) Cal lus f o r m a t i o n was f o u n d in every t r e a t m e n t even at the highest
c o n c e n t r a t i o n (1.0 mg L -1) tested.

106
Fig. 5. Effects of Amo-1618 on asparagus shoot-apex culture.
Effect of cycocel (chlormequat chloride, (2-chloroethyl) trimethyl ammonium chloride)
Very limited improvement was obtained with cycocel treatments (Fig. 6, Table 3). No concentrations of cycocel tested suppressed callus formation.
Effects of ancymidol, GA 3, and GA4/7.
Table 4 shows that thickness of shoot was increased by ancymidol. How- ever, this effect was reversed by GA 3 or GA4/7. Incorporation of GA 3 or GA4/7 completely inhibited root formation.
Effects of ancymidol, amo-1618, and cycocel on GA-like activities.
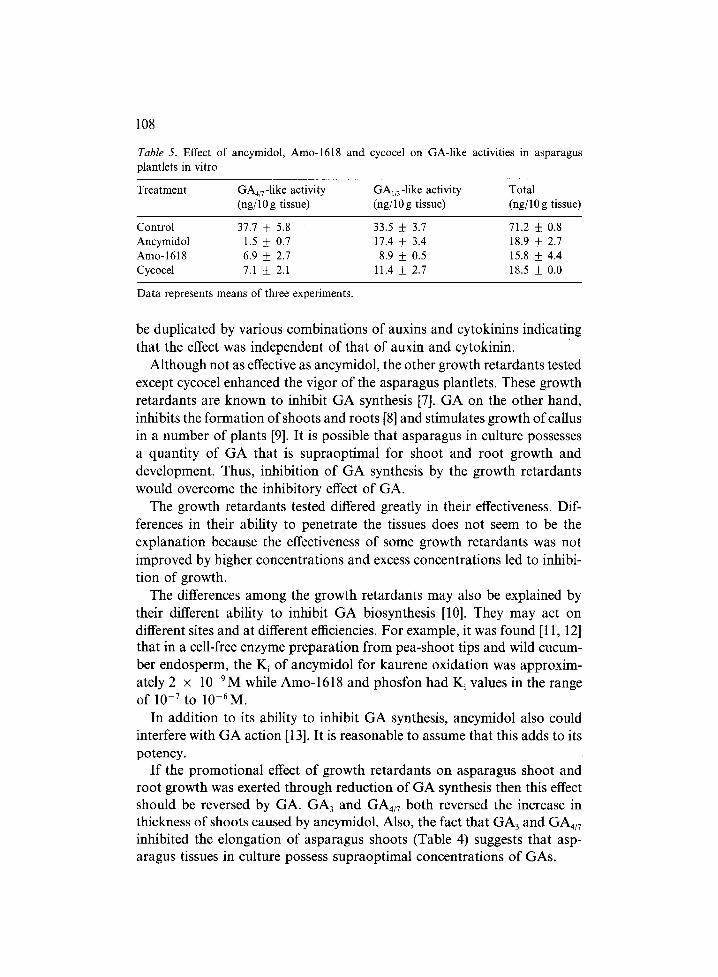
Two distinct zones of GA-like activities as determined by the lettuce hypo- cotyl test (6) were resolved from the silicic acid column (Table 5). According to the study of Powell and Tautvydas [5], these two zones represent GA1/3 an d GA4/7-1ike activities. Ancymidol, Amo-1618 and cycocel all reduced GA1/3 and GAa/7-1ike activities. Ancymidol was much more effective than Amo-1618 and cycocel in inhibiting the GA4/7,1ike activity, whereas Amo- 1618 and cycocel were more effective than ancymidol in inhibiting the GA1/3-like activity. The total GA-like activities in plantlets treated with the three growth retardants were similar.

107
Fig. 6. Effects of cycocel on asparagus shoot-apex culture.
Table 4. The effects of ancymidol and GA 3 on shoot and root growth in shoot apex cultures of Asparagus officinalis L.
Concentration (rag L- ~) Clone 56
ancymidol GA3 GA4/7 shoot (mm) root (ram)
length thickness length thickness
0 0 0 25.14a ~ 0.52b 6.15a 0.77b 0 1 0 3.63c 0.45b 0.00c 0.00c 2 0 0 6.20b 1.14a 0.68b 1.27a 2 1 0 1.40c 0.23c 0.00c 0.00c 2 2 0 1.46c 0.23c 0.00c 0.00c 2 3 0 1.37c 0.22c 0.00c 0.00c 2 4 0 1.15c 0.22c 0.00c 0.00c 2 5 0 0.00d 0.00d 0.00c 0.00c 0 0 1 6.4b 0.34bc 0.00c 0.00c 2 0 1 8.7b 0.54b 0.00c 0.00c 2 0 2 6.16b 0.36c 0.00c 0.00c
ZMean separation in columns by Duncan's multiple range test, 5% level.
Discussion
A n c y m i d o l great ly e n h a n c e d the g rowth o f v igorous shoots a n d roots , as
m e a s u r e d by th ickness of a spa ragus shoo t - t ip cul ture . This effect cou ld n o t
108
Table 5. Effect of ancymidol, Amo-1618 and cycocel on GA-like activities in asparagus plantlets in vitro
Treatment GA4/7-1ike activity GAin-like activity Total (ng/10 g tissue) (ng/10 g tissue) (ng/10 g tissue)
Control 37.7 _ 5.8 33.5 + 3.7 71.2 ___ 0.8 Ancymidol 1.5 _ 0.7 17.4 + 3.4 18.9 + 2.7 Amo-1618 6.9 _ 2.7 8.9 + 0.5 15.8 + 4.4 Cycocel 7.1 + 2.1 11.4 + 2.7 18.5 + 0.0
Data represents means of three experiments.
be duplicated by various combinations of auxins and cytokinins indicating that the effect was independent of that of auxin and cytokinin.
Although not as effective as ancymidol, the other growth retardants tested except cycocel enhanced the vigor of the asparagus plantlets. These growth retardants are known to inhibit GA synthesis [7]. GA on the other hand, inhibits the formation of shoots and roots [8] and stimulates growth of callus in a number of plants [9]. It is possible that asparagus in culture possesses a quantity of GA that is supraoptimal for shoot and root growth and development. Thus, inhibition of GA synthesis by the growth retardants would overcome the inhibitory effect of GA.
The growth retardants tested differed greatly in their effectiveness. Dif- ferences in their ability to penetrate the tissues does not seem to be the explanation because the effectiveness of some growth retardants was not improved by higher concentrations and excess concentrations led to inhibi- tion of growth.
The differences among the growth retardants may also be explained by their different ability to inhibit GA biosynthesis [10]. They may act on different sites and at different efficiencies. For example, it was found [11, 12] that in a cell-free enzyme preparation from pea-shoot tips and wild cucum- ber endosperm, the K i of ancymidol for kaurene oxidation was approxim- ately 2 x 10-9M while Amo-1618 and phosfon had Ki values in the range of 10 -7 to 10-6M.
In addition to its ability to inhibit GA synthesis, ancymidol also could interfere with GA action [13]. It is reasonable to assume that this adds to its potency.
If the promotional effect of growth retardants on asparagus shoot and root growth was exerted through reduction of GA synthesis then this effect should be reversed by GA. GA 3 and GA4/7 both reversed the increase in thickness of shoots caused by ancymidol. Also, the fact that GA3 and GA4/7 inhibited the elongation of asparagus shoots (Table 4) suggests that asp- aragus tissues in culture possess supraoptimal concentrations of GAs.

109
To further examine the relationship between growth retardants and their inhibition of GA synthesis, cultures were treated with ancymidol, Amo- 1618, and cycocel, and after 6 weeks the GA levels in these cultures were determined. These three growth retardants were selected because among the growth retardants tested ancymidol was most effective, Amo-1618 inter- mediately effective, and cycocel least effective in promoting asparagus shoot and root development. Two distinct zones of GA-like activities, which according to the study of Powell and Tautvydas [5] represented GA1/3 and GA4/y-like activities were recovered from asparagus tissue. Ancymidol which was most effective in stimulating the thickening of shoots and roots was also most effective in reducing the level of GA4/v-like activity and Amo- 1618 and cycocel which were less effective in stimulating the thickening of shoots and roots were less effective in reducing the GA4/v-like activity. This suggests that GA4/7 is more involved in the decreased vigor of shoots and roots. The results of this study do not clearly resolve the importance of GA4/7 because it was observed that the GA4/7-1ike activities of Amo-1618 and cycocel treated plants were similar, although the former promoted stronger shoots and roots.
Acknowledgement
This work was supported by the New Jersey Agricultural Experiment Sta- tion and performed as part of New Jersey Agricultural Experiment Station project 12151.
References
1. Murashige T, Shabde MN, Hasegawa PM, Taketori FB, Jones JB (1972) Propagation of asparagus through shoot apex culture. I. Nutrient medium for formation of plantlets. J Am Soc Hort Sci 97(2): 1258-161
2. Chin CK (1982) Promotion of shoot and root formation in asparagus in vitro by ancy- midol. Hort Science 17:590-591
3. Yang HJ, Clore WJ (1974) Improving the survival of aseptically-cultured asparagus plants in transplanting. Hort Science 9:235 236
4. Murashige T, Skoog F (1962) A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol Plant 15:473~497
5. Powell IE, Tautvydas KJ (1967) Chromatography of gibberellins on silica gel partition columns. Nature 213:292-293
6. Frankland B, Wareing PF (1960) Effect of gibberelic acid on hypocotyl growth of lettuce seedlings. Nature 4708:255-256
7. Dennis DT, Upper CD, West CA (1965) An enzymatic site of inhibition of gibberellin biosynthesis by Amo-1618 and other plant growth retardants. Plant Physiol 40:948-952

110
8. Nickell LG (1958) Gibberellin and the growth of plant tissue culture. Nature 181:499-500 9. Murashige T (1964) Analysis of the inhibition of organ formation in tobacco tissue culture
by gibberellin. Physiol Plant 17:636-643 10. Russell GF, West CA (1977) Properties of Kaurene synthetase from Marah macrocarpus.
Plant Physiol 59:22-29 11. Coolbaugh RC, Hamilton, R (1976) Inhibition of ent-kaurene oxidtion and growth by
e-cyclopropyl-c~-(p-methoxyphenyl)-5-pyrimidine methylalcohol. Plant Physiol 57: 245- 248
12. Coolbaugh RC, Hirano SS, West, CA (1978) Studies on the specificity and site of action of c~-cyclopropyl-c~-(p-methoxyphenyl)-5-pyrimidine methylalcohol (ancymidol), a plant growth regulator. Plant Physiol 62:571 576
13. Leopold AC (1971) Antagonism of some gibberellin action by a substituted pyrimidine. Plant Physiol 48:537 540.




















