
Extreme diel horizontal migrations by a tropical nearshore resident micronekton community

Global and Planetary Change xxx (2010) xxx–xxx
GLOBAL-01583; No of Pages 16
Contents lists available at ScienceDirect
Global and Planetary Change
j ourna l homepage: www.e lsev ie r.com/ locate /g lop lacha
ARTICLE IN PRESS
Pliocene–Pleistocene diatom biostratigraphy of nearshore Antarctica from theAND-1B drillcore, McMurdo Sound
D. Winter a,⁎, C. Sjunneskog b, R. Scherer c, P. Maffioli d, C. Riesselman e, D. Harwood a
a University of Nebraska–Lincoln, Lincoln, NE, USAb Antarctic Core Facility, Florida State University, Tallahassee, FL, USAc Northern Illinois University, DeKalb, IL, USAd Dipartimento di Scienze Geologiche e Geotecnologie, Università Milano-Biocca, Italye Department of Geological and Environmental Sciences, Stanford University, CA, USA
⁎ Corresponding author. 214 Bessey Hall, DepartmenNebraska-Lincoln, Lincoln, NE 68588-0340, USA. Tel.: +472 4917.
E-mail address: [email protected] (D. Winter).
0921-8181/$ – see front matter © 2010 Elsevier B.V. Aldoi:10.1016/j.gloplacha.2010.04.004
Please cite this article as: Winter, D., et al.,McMurdo Sound, Glob. Planet. Change (20
a b s t r a c t
a r t i c l e i n f oArticle history:Received 29 October 2008Accepted 3 April 2010Available online xxxx
Keywords:ANDRILLAND-1BdiatombiostratigraphyRoss SeaAntarcticaLate NeogenePliocenePleistocene
The near-shore open-marine diatom record recovered in the ANtarctic geological DRILLing (ANDRILL)McMurdo Ice Shelf Project (MIS) AND-1B drillcore, McMurdo Sound, Antarctica, advances our understandingof the marine conditions present in the southern Ross Sea during the Pliocene and early Pleistocene. Thisdiatom history is recorded within alternating diamictite and diatomite that reflect alternating glacial activityand high marine primary productivity. The diatomite units were deposited in a continental shelf open-marine setting during periods of reduced ice cover in West Antarctica. A new diatom biostratigraphic schemespanning the last ca. 5 Ma is proposed for the Antarctic near-shore area, based on prior work from highlatitude drillcores. Four new zones are proposed for the Pliocene/Pleistocene, with eight in total for the newzonal scheme, utilizing Actinocylus fasciculatus, Actinocyclus maccollumii, Fragilariopsis bohatyii, Rouxiaantarctica, and Thalassiosira fasciculata as new zonal markers. The early Pliocene shares the most assemblagecommonality with that of the Southern Ocean with greater numbers of endemic species observed in the latePliocene and early Pleistocene; a group of related Fragilaripsis species characterizes much of this later part ofthe time column. Two new species are proposed, Fragilariopsis tigris sp. nov. Riesselman and Thalassiosirateres sp. nov. Winter; a formal name is also proposed for another species, Rhizosolenia harwoodii sp. nov.Winter. The new zonation is tied to a robust chronology utilizing diatom biostratigraphy, volcanic 40Ar/39Arages and magnetostratigraphy.
t of Geosciences, University of1 402 472 2663; fax: +1 402
l rights reserved.
Pliocene–Pleistocene diatom biostratigraphy10), doi:10.1016/j.gloplacha.2010.04.004
© 2010 Elsevier B.V. All rights reserved.
1. Introduction
The ANDRILL McMurdo Ice Shelf Project drillcore AND-1Brecovered 1284 m of sediment from beneath the McMurdo SoundIce Shelf. Of this, the upper 585 m provides the most completeproximal sedimentary glacial–interglacial record for the high latitudePliocene (4.9–2 Ma) from the Antarctic continental shelf (Naish et al.,2009). Initial field-based analysis recognized that the Southern Ocean(SO) biostratigraphic zonation was not fully applicable to the materialrecovered (Harwood and Maruyama, 1992; Zielinski and Gersonde,2002; Scherer et al., 2007). This resulted from low abundance of somekey marker species and truncated age ranges for other species. Toestablish a reliable biostratigraphic framework for the continentalshelf, the Southern Ocean biostratigraphic zonation must then bemodified. Here we present the biostratigraphy of the AND-1B drillcore
employing a scheme using both established Southern Ocean zonaldiatom datums as well as new marker species.
The Southern Ocean diatom biostratigraphic framework has beendeveloped and refined through core recovery from multiple DSDP andODP Legs. Deep Sea Drilling Project (DSDP) Leg 28 recovered the firstdrillcores from the Antarctic continental shelf in the Ross Sea, but withlimited success (Hayes et al., 1975). This Leg was the starting point formany successive Antarctic drilling seasons. The Ocean Drilling Program(ODP) continued with Antarctic drilling, initially focusing on off-shoremarine sites, including Legs 113, 114, 119, 120, 177, 178, 183 and 188 inthe Southern Ocean and Legs 178 and 188 which recovered cores fromthe Antarctic continental shelf. Cores drilled on the continental shelfduringODP Leg 188 in Prydz Bay had slightly better recovery than thoseof Leg 178 (Leg 188 Site 1166–18.6% vs. Leg 178 Site 1100–15.95%, Site1102–22.04%, Site 1103–12.3%, respectively), but none approached the98% attained by both ANDRILL drilling seasons (Barker et al., 1999;O'Brien et al., 2001; Naish et al., 2007; Harwood et al., 2008–09) andprior Cape Roberts Project (CRP) drilling (Davey et al., 2000) in thewestern Ross Sea.
Biostratigraphic work from the deep ocean has resulted in differentbiostratigraphic schemes applied north and south of the Polar Frontal
of nearshore Antarctica from the AND-1B drillcore,

2 D. Winter et al. / Global and Planetary Change xxx (2010) xxx–xxx
ARTICLE IN PRESS
Zone (Zielinski and Gersonde, 2002), due to species ecologicalpreferences and habitat differences. Furthermore, in the modernSouthern Ocean it is evident that environmental differences exert astrong influence on diatom distribution; assemblages occupying theneritic sea-ice zone are very different from those observed in the openocean (Kellogg and Truesdale, 1979; Cunningham and Leventer, 1998;Armand et al., 2005; Crosta et al., 2005). The rapidly changingdepositional environment reflected in the alternating diatomite/diamictite sequences preserves different assemblages of diatom speciesthan are observed in the Southern Ocean during the mid-Pliocenethrough early Pleistocene. The environmental control on diatomdistribution suggests the need for separate (or modified) biostrati-graphic schemes for the continental shelf versus open ocean. Magne-tostratigraphy from the AND-1B drillcore suggests good correlationwith known age ranges for some species present in the Southern Ocean(Wilson et al., 2007).
At sites closer to the continent, a series of drillcores recovered withland and sea-ice drilling rigs along the southern Victoria Landmargin inthe McMurdo Sound contain a very proximal record for the Cenozoic ofthe western Ross Sea. These include cores recovered during the DryValley Drilling Project (DVDP) (Powell, 1981), McMurdo SoundSediment and Tectonic Studies (MSSTS) (Barrett, 1986), CenozoicInvestigations in the western Ross Sea (CIROS)-1 and -2 (Barrett, 1989;Barrett and Hambrey, 1992) and the Cape Roberts Project (CRP)-1, -2and -3 (Cape Roberts Science Team, 1998, 1999, 2000). Previouslyrecovered sediments of correlative age from this area have generallybeen thin intervals of diatom-rich muds averaging 7 m in thickness,quite different from the diatomite in AND-1B. The new biostratigraphyof this recordwill help improve age constraints for these previous cores.
Prior to the recovery of the AND-1B drillcore, development andapplication of Antarctic diatom biostratigraphy from coastal andcontinental-shelf deposits was restricted to these punctuated andbrief records exposed on land and recovered in near shore drill cores.The strata recovered in ANDRILL's inaugural season preserve anunprecedented record of diatom populations and evolution on thecontinental shelf, upon which a biostratigraphic scheme for coastalAntarctic regions can be based. An excellent record of Late Neogenediatoms is present in the upper 585 m of the AND-1B drillcore, oftenin sufficient abundance to qualify as diatomite (Scherer et al., 2007).The diatomaceous and diatomite units are interbedded within a
Fig. 1. Map of the southern Ross Sea and McMurdo Sound region. The circle indicates th
Please cite this article as: Winter, D., et al., Pliocene–Pleistocene diatomMcMurdo Sound, Glob. Planet. Change (2010), doi:10.1016/j.gloplacha.
cyclical sedimentary sequence of diamictite, mudstone, sandstoneand volcaniclastic sediments (Krissek et al., 2007). Many glacialsurfaces of erosion are present within these sequences, commonly atthe base of and within diamictite units. AND-1B contains more than13 intervals of diatomaceous mud and diatomite representing open-marine deposition in the southern Victoria Land Basin. These diatom-bearing units vary in thickness from less than 1 m in the youngestPleistocene interval to N60 m of continuous lower to mid-Pliocenediatomite, averaging 20 m in thickness.
2. Regional setting
The ANDRILL McMurdo Ice Shelf AND-1B drill site is located insouthern McMurdo Sound at 77.89°S, 167.09°E, in ∼850 m waterdepth, in the southern end of Victoria Land Basin and on its easternflank (Fig. 1). The core was drilled from the McMurdo Ice Shelf to thesoutheast of McMurdo Station/Scott Base on Ross Island and to thenorth–northwest of White Island. The targeted sediments accumu-lated in a basin surrounding Ross Island, thought to result from crustalloading and flexure associated with the growth of Mt. Erebus and theother Ross Island volcanoes (Horgan et al., 2005).
Diatomaceous sediments of this age exist in other areas around theAntarctic continent, some preserved in ice-free areas on land andsome in other drillcore material. Several other cores exist in the sameregion as AND-1B, Dry Valley Drilling Project cores DVDP-10 andDVDP-11 both drilled in 1974 at the mouth of Taylor Valley andCenozoic Investigations in the Western Ross Sea drillcore CIROS-2drilled in 1984 from Ferrar Fjord (McGinnis, 1981; Barrett, 1985).These cores combined contain the near-shore section of the climatichistory recorded in AND-1B. Their sediments are, at best, diatom-richmud units but they document the same interglacial intervalsrepresented in the AND-1B diatomite units (Winter and Harwood,1997; Winter et al., this volume).
On the other side of the continent, subaerially exposed marinesediments of the Sørsdal Formation in the Vestfold Hills region of EastAntarctica represent the time interval equivalent to Diatom Unit 11b(DU-11b) (Whitehead et al., 2001; Scherer et al., 2007). A younger setof strata in the Bardin Bluffs Formation in the Amery Oasis of thenorthern Prince Charles Mountains record ice-distal and ice-freesedimentation in the late Pliocene and early Pleistocene (Whitehead
e location of the ANDRILL MIS AND-1B drillsite (locate in or near Regional Setting).
biostratigraphy of nearshore Antarctica from the AND-1B drillcore,2010.04.004

Fig. 2. Illustration of new zonal scheme based on AND-1B core for the Antarctic continental shelf and near-shore areas. Also shown are previous Southern Ocean zonations describedby Harwood and Maruyama (1992; left) and Zielinski and Gersonde (2002; middle). Together these represent a simple latitudinal cross-section of diatom biostratigraphy for theSouthern Ocean and now the Antarctic continental shelf (locate on or near page 7, first part of Results section). Paleomagnetic time scale follows Ogg and Smith (2004).
3D. Winter et al. / Global and Planetary Change xxx (2010) xxx–xxx
ARTICLE IN PRESS
and McKelvey, 2001). A terrestrially exposed marine section is alsopresent on Cockburn Island, near the tip of the Antarctic Peninsula.This material is suggested by diatom biostratigraphy and 40Ar/39Ardating to be late Pliocene in age, corresponding to diatomite intervalsbetween 200 and 250 mbsf in AND-1B (Harwood, 1986a; Jonkers andKelley, 1998). Subsequent geochemical work on bivalve remainsrecovered in this same material suggests an earlier age of 4.6 Ma,correlating this formation with an older part of the AND-1B corebetween 453 and 459 mbsf (McArthur et al., 2006).
Several till formations preserved in both the Transantarctic as wellas the northern Prince Charles Mountains are interpreted to representa late Pliocene or early Pleistocene advance of the East Antarctic IceSheet. The dating of these till units is based on diatoms present within,the youngest ones being mid-Pliocene in age (Harwood, 1986b;Whitehead et al., 2004). The nature of these sediments, represented
Please cite this article as: Winter, D., et al., Pliocene–Pleistocene diatomMcMurdo Sound, Glob. Planet. Change (2010), doi:10.1016/j.gloplacha.
by the Sirius Group in the Transantarctic Mountains and thePagodroma Tillite in the Prince Charles Mountains suggests theywere emplaced by expanding wet-based glaciers (Webb et al., 1996;Whitehead et al., 2004). Certain parts of the Sirius Group also containswood, plant and invertebrate remains as well (Webb and Harwood,1993; Francis and Hill, 1996, Hill et al., 1996; Ashworth and Cantrill,2004) indicating more temperate conditions than exist today in thispart of Antarctica.
3. Materials and methods
Diatom occurrence and abundance data generated during theAND-1B drilling season (Scherer et al., 2007) is utilized for thiszonation along with data from subsequently analyzed samples. Smearand strewn slides were analyzed for diatom abundance estimation
biostratigraphy of nearshore Antarctica from the AND-1B drillcore,2010.04.004

4 D. Winter et al. / Global and Planetary Change xxx (2010) xxx–xxx
ARTICLE IN PRESS
during the on-ice phase of work; slides used for additionalbiostratigraphic and taxonomic information are from various inter-vals. Sample spacing varied from 0.5 m to 3 m within the diatomiteintervals, 665 samples were analyzed for this work. Subsequentsample analysis was carried out on strewn slides prepared using thefollowing method: approximately 0.5 g of each sample was broken upand treated with 15–20 ml 30% H2O2 to disaggregate the sedimentfurther and remove organic material then the residue was settled 3–5times in 50 cc of 5% Calgon solution for 3 h each to remove excess clayand other fine material from the samples. Finally, each sample wasrinsed 4 timeswith Ropurewater to remove all trace of chemicals. Theresulting material was agitated and allowed to settle for 30 s beforetransferring 1–3 drops of the supernatant collected by pipette throughthe suspendedwater column onto a coverslip. This was allowed to dryon a hotplate and affixed to a microscope slide with Norland OpticalAdhesive #61 (refractive index 1.56) and cured under a UV light.Counts and taxonomic work were completed on an Olympus CHtransmitted light microscope. A minimum of 300 valves were countedin each sample at a magnification of 600×, the resulting species totalswere changed to percent for analysis and sample comparison. Amagnification of 1000× was used to confirm species identifications.Taxa are identified to the species level when possible. A valve wascounted as ‘one’ if the central raphe and one terminal end of a pennatespecies is preserved, or consisted of at least one half of the valve of acentric species including the central area. A single Chaetoceros restingspore cell is counted as ‘one’ in this method, as well as individualgirdle bands ofDactyliosolen antarcticus. Counts of 300 valves per slidewere performed for subsequent sample analysis in addition to initialdescriptive abundance estimation. Quantitative analysis allowed adetailed assessment to be made of the assemblages in the differentdiatomite units throughout the core. Comparison of this quantitativedata with the original qualitative abundance estimates follows themethod described by Zielinski and Gersonde (2002), in which the rawcounts are changed to percentages and assigned to abundancecategories as follows: N30%=Abundant (A), 15–30%=Common (C),3–15%=Few (F), 3–1%=Rare (R), b1%=Scarce (X).
The biostratigraphic scheme presented here is based on zonationsdeveloped from various DSDP and ODP cruises (Fig. 2). SouthernOcean zonations were applied with little modification to several otherdrillcores within McMurdo Sound (Winter and Harwood, 1997). Theearly and mid-Pliocene diatom biostratigraphy of drillcores CIROS-2,DVDP-10 and DVDP-11 is discussed along with their comparativesections in the AND-1B core in this volume (Winter et al., this vol.).
4. Results
Here we describe a zonal scheme for the near-shore continentalshelf of the Ross Sea. The early Pliocene shares two zones in commonwith several Southern Ocean zonations (Fig. 2), while the late Plioceneand early Pleistocene differ more significantly. We propose four newzones reflecting regional assemblage characteristics specific to thisarea, defined by species with greater biostratigraphic utility in thisnear-shore setting; Thalassiosira fasciculata Zone, Fragilariopsis boha-tyi Zone, Actinocyclus fasciculatus–Actinocyclus maccollumii Zone andthe Rouxia antarctica Zone. This work, along with the continental shelfdiatom biostratigraphy for the Oligocene–lower Miocene from theCape Roberts Project (Olney et al., 2009) and recent Miocenebiostratigraphy from Southern McMurdo Sound (AND-2A, Harwood
Fig. 3. Lithologic log of the upper 600 m of the AND-1B drillcore, with the proposed near-shrepresent diatom units used in initial core description. Abundance data for biostratigraphicaDepth intervals for Diatom Units are as follows: DU-I (58.15–58.90 mbsf), DU-II (86.92–97180.73 mbsf), DU-VI (183.42–201.59 mbsf), DU-VII (209.96–224.44 mbsf), DU-VIII (250(363.37–459.24 mbsf), DU-XII (503.42–511.56 mbsf), DU-XIII (550.78–586.45 mbsf) (locate
Please cite this article as: Winter, D., et al., Pliocene–Pleistocene diatomMcMurdo Sound, Glob. Planet. Change (2010), doi:10.1016/j.gloplacha.
et al., 2008–09) will continue the development and improvement of acomposite diatom biostratigraphy for the Antarctic continental shelf.
Fig. 3 illustrates the zonal scheme developed and applied to theAND-1B drillcore with the four new zones, two further divided intosub-zones. The species used have well-constrained ages at othercircum-Antarctic sites and appear to express regionally correlativefirst occurrence (FO) and last occurrence (LO) datums according tothe age/depth model developed for this core (Wilson et al., 2007).Three of the four new zones are within the late Pliocene and earlyPleistocene, with one new zone in the early Pliocene. The diatoma-ceous units (DU) applied in the initial results (Scherer et al., 2007) areretained here. Each unit comprises a single diatomite intervalbounded above and below by diamictite or several diatomite unitsthat share a common assemblage indicating that any glacial activityrecorded between each diatomite interval was short-lived and did notsignificantly alter the environment affecting assemblage character.The upper 585 m of AND-1B are comprised of alternating diamictiteand diatomite, with volcanic sand and mud units in lower abundance.Themajority of these diatom units define diatomite intervals boundedby diamictite, while DU-V comprises several diatomite intervals of thesame age. Abundance data for biostratigraphically useful species isillustrated as individual graphs for each species. The extent of the greyboxes, denoting the inferred in situ ranges of species illustrated, donot extend above DU-II; DU-I is interpreted to comprise reworkedassemblages containing both contemporaneous and older Miocenespecies. Fig. 4 illustrates FO and LO datums occurring within AND-1B(Table 1). Species datums truncated by glacial surfaces of erosioncluster together at similar depths, such as occurs at 150 mbsf. Theearly to mid-Pliocene Fragilariopsis interfrigidaria Zone is expandedwithin the core, compared to its absolute age. Other zones, such as theThalassiosira fasciculata Zone, are preserved within a limited strati-graphic interval. The shaded box to the left of the correlation lineindicating the FO of F. interfrigidaria represents the approximate timeinterpreted to be missing at this unconformable boundary, confinedby the base of C2An.3n and the reported age for the FO of this species(Cody et al., 2008).
The youngest modern Southern Ocean zone, Thalassiosira lentigi-nosa Zone, is not preserved in diatom-bearing sediments of AND-1B.The youngest sediments recovered in the core are composed of aseries of diamictite units N80 m in thickness following the loss of theuppermost ∼24 m of sediment in the process of embedding the seariser (Falconer et al., 2007). It is known from previous samplesrecovered under the ice shelf at this site that this youngest zone isrepresented within these lost surface sediment samples as short corestaken prior to initiation of drilling contained assemblages reflectingdeposition during this time period (Dunbar et al., 2007). Diamictiteunits are generally barren of diatoms and do not provide usefulcorrelative reference to open-ocean zonations.
The details of the biostratigraphic zones identified within theAND-1B core are presented below and discussed further in thefollowing section. The ages presented for the boundaries of each zoneare drawn from new refined average age model estimates derived bythe CONstrained OPtimization (CONOP) method (Cody et al., 2008).This method developed a dataset comprising all reported FO and LOfor Antarctic diatom species, allowing for a detailed and morecomplete assessment of the entire biostratigraphic ranges of thesespecies. The noted paleomagnetic correlations below correspond tothe mean of the average age model age estimates from Cody et al.(2008). Both the published age range and average-model CONOP ages
ore zonation to the left and key biostratigraphic species to the right. Roman numeralslly useful species is plotted by depth, grey boxes denote their in situ range in AND-1B..08 mbsf), DU-III (116.75–118.70 mbsf), DU-IV (150.87–159.33 mbsf), DU-V (164.10–.02–258.32 mbsf), DU-IX (283.35–292.66 mbsf), DU-X (295.46–346.94 mbsf), DU-XIon or near page 7, first part of Results section).
biostratigraphy of nearshore Antarctica from the AND-1B drillcore,2010.04.004

5D. Winter et al. / Global and Planetary Change xxx (2010) xxx–xxx
ARTICLE IN PRESS
Please cite this article as: Winter, D., et al., Pliocene–Pleistocene diatom biostratigraphy of nearshore Antarctica from the AND-1B drillcore,McMurdo Sound, Glob. Planet. Change (2010), doi:10.1016/j.gloplacha.2010.04.004

Fig. 4.Diatom events plotted by depth and age. Lithologic log and proposed zonation for AND-1B are illustrated on the left. Paleomagnetic time scale is across the topwith near-shorezones indicated above. Biostratigraphically important diatom datums indicated with triangles; left-pointing triangles are first occurrence datums, right-pointing triangle lastoccurrence datums. Wavy lines indicate glacial surfaces of erosion and interpreted intervals of missing time. Grey box indicates interpreted hiatus at base of F. interfrigidaria Zone.Paleomagnetic time scale on horizontal axis follows Ogg and Smith (2004) (locate on or near page 7, first part of Results section).
6 D. Winter et al. / Global and Planetary Change xxx (2010) xxx–xxx
ARTICLE IN PRESS
of biostratigraphically significant species are presented in Table 1along with the depths at which each of these events occur in AND-1B.
4.1. Near-shore Ross Sea Zonation
Shionodiscus tetraoestrupii Partial Range Zone (renamed herein)Authors: Baldauf and Barron (1991), renamed here based on
application of species name change and transfer to genus Shionodiscus(Bodén, 1993; Alverson et al., 2006).
Top: FO Thalassiosira inura (507.90 mbsf)Base: FO Shionodiscus tetraoestrupii (583.64 mbsf)Stratigraphic position and age: early Pliocene, 4.74 to 4.95 Ma
Please cite this article as: Winter, D., et al., Pliocene–Pleistocene diatomMcMurdo Sound, Glob. Planet. Change (2010), doi:10.1016/j.gloplacha.
Paleomagnetic correlation: early Gilbert (top of the zone atC3n.2r)
Discussion: The presence of S. tetraoestrupii through the oldestsamples containing well-preserved diatomaceous material at 583.64indicates the base of this zone is not preserved in AND-1B. Below thisdepth diatoms are only very rarely present, quite dissolved and/orovergrown. Two diatom units, DU-XIII and DU-XII, preserve this zonewith the upper boundary well constrained in DU-XII. Actinocyclusoctonarius and Actinocyclus karstenii are common and occur inrelatively equal proportions in assemblages of this zone. Other speciesrepresenting a significant part of the assemblage include: Chaetocerosspecies, Coscinodiscus species, Dactyliozolen antarcticus, Denticulopsis
biostratigraphy of nearshore Antarctica from the AND-1B drillcore,2010.04.004

Table 1Biostratigraphically useful species depths and as observed in AND-1B. Cody et al. (2008) ages are from the average CONOP model, the published ages are an average for each datumbased on previously published Southern Ocean sites. Species names in bold type are used as zonal marker species. An asterisk (*) next to a FO or LO indicates this datum is truncatedby an unconformity. The age in parenthesis for T. torokina is the average of previously published ages for this datum, the second age represents a new youngest age for this datum inAND-1B (discussed in text) (locate on or near page 7–8 — end of Results section).
Datum Species Depth Published age(min–max)
Cody et al., 2008(average min–max)
(mbsf)
LO* Rouxia leventerae Bohaty, Scherer et Harwood 52.98 0.13–0.14 0.07–0.35LO* Thalassiosira elliptipora (Donahue) Fenner 58.15 0.3–1.81 0.64–0.71LO Thalassiosira fasciculata Harwood er Maruyama 58.49 0.75–1.81 0.89LO* Thalassiosira torokina Brady 86.90 1.88–2.18 2.2–2.27LO* Actinocyclus ingens Rattray 86.90 0.3–1.99 0.5–0.57LO* Rouxia antarctica Heiden, in Heiden and Kolbe 116.93 1.35–1.57 1.48–1.51LO* Actinocyclus karstenii Van Heurck 150.70 1.81–2.91 2.13–2.16LO* Thalassiosira inura Gersonde 150.70 1.88–3.1 2.53–2.55LO* Thalassiosira kolbei (Jousé) Gersonde 150.70 1.61–2.7 1.98LO Shionodiscus tetraoestrupii var. reimeri Mahood et Barron 150.80 1.32–1.61 1.31–1.34LO Rhizosolenia harwoodii sp. nov. 150.80 – –
LO Rouxia diploneides Schrader 151.21 1.62–3.23 2.55–2.69LO Fragilaripsis bohatyi sp. nov. 152.80 – –
FO Actinocyclus actinochilus (Ehrenberg) Simonsen 155.80 2.2–3.19 2.72–2.81LO* Actinocyclus fasciculatus Harwood et Maruyama 164.10 1.81–2.81 2.05–2.27LO Actinocyclus maccollumii Harwood et Maruyama 176.20 1.81–2.91 2.4–2.45LO Thalassiosira vulnifica (Gombos) Fenner 169.80 0.6–3.1 2.15–2.19FO Rouxia leventerae Bohaty, Scherer et Harwood 176.20 – 2–2.08FO Shinodiscus tetraoestrupii var. reimeri Mahood et Barron 176.70 2.4–2.66 2.35–2.37FO Actinocyclus fasciculatus Harwood et Maruyama 191.95 1.72–1.81 2.65–2.77LO Thalassiosira webbi Harwood et Maruyama 191.95 2.91 2.65–2.77LO Creania lacyaeOlney 175.70 – –
FO Actinocyclus maccollumii Harwood et Maruyama 201.20 2.5–3.3 2.79–2.84LO Fragilariopsis barronii (Gersonde) Gersonde et Bárcena 251.74 0.8–2.6 1.19–1.29FO* Thalassiosira vulnifica (Gombos) Fenner 251.76 2.2–3.51 3.12–3.18LO* Fragilariopsis praeinterfrigidaria (McCollum) Gersonde et Bárcena 251.76 2.09–4.61 3.45–3.49FO Thalassiosira webbi Harwood et Maruyama 253.10 3.41 3.17–3.44LO Fragilariopsis tigris sp. nov. 284.90 – –
LO Thalassiosira complicata Gersonde 284.90 2.61–4.51 3.36–3.44LO Thalassiosira striata Harwood et Maruyama 285.36 1.81–3.51 2.89–2.96FO Thalassiosira elliptipora (Donahue) Fenner 288.76 1.07–3.51 2–2.06FO* Fragilariopsis tigris sp. nov. 291.50 – –
FO* Fragilariopsis bohatyi sp. nov. 291.50 – –
LO Proboscia barboi (Brun) Jordan et Priddle 330.90 1.61–4.21 1.6–1.73LO* Fragilariopsis interfrigidaria (McCollum) Gersonde et Bárcena 364.38 1.81–3.3 2.4–2.45LO* Denticulopsis delicata Yangasawa et Akiba 364.38 – –
FO Fragilariopsis barronii (Gersonde) Gersonde et Bárcena 413.60 4.01–4.8 4.28–4.52FO Thalassiosira kolbei (Jousé) Gersonde 437.00 2.23–4.8 3.8–4.02FO Fragilariopsis interfrigidaria (McCollum) Gersonde et Bárcena 437.59 3.3–4.01 3.93–4.19FO Rhizosolenia harwoodii sp. nov. 438.00 4.5 3.08–3.58LO Denticulopsis sp. cf. D. delicata 440.12 – –
FO Thalassiosira fasciculata Harwood et Maruyama 450.83 4.49 4.25–4.42FO Thalassiosira striata Harwood et Maruyama 452.25 3.48–4.49 4.3–4.64FO Thalassiosira inura Gersonde 507.90 4.8–6.83 4.71–4.77FO Fragilariopsis praeinterfrigidaria (McCollum) Gersonde et Bárcen 508.95 4.51–5.8 4.72–4.78FO Rouxia antarctica Heiden, in Heiden and Kolbe 509.10 – 4.43–4.57FO Fragilariopsis curta (Van Heurck) Hustedt 570.13 3.72 3.54–3.57FO Rouxia diploneides Schrader 581.84 3.72–4.49 4.61–4.7FO Denticulopsis sp. cf. D. delicata 581.84 – –
FO* Thalassiosira complicata Gersonde 583.64 4.51–4.8 4.64–4.71FO* Shinodiscus tetraoestrupii (Bodén) Alverson, Kang et Theriot 583.64 4.31–5.96 4.8–4.95
7D. Winter et al. / Global and Planetary Change xxx (2010) xxx–xxx
ARTICLE IN PRESS
delicata, Denticulopsis sp. cf. D. delicata, Eucampia antarctica, Rouxiaspecies, Stellarima microtrias, Thalassionema/Thalassiothrix species,S. tetraoestrupii and Thalassiosira torokina. The FO of F. praeinterfrigi-daria (508.95 mbsf), Fragilariopsis praecurta (568.34 mbsf), Fragilar-iopsis curta (570.13 mbsf) occur within this zone in AND-1B.
Thalassiosira inura Partial Range ZoneAuthors: Gersonde and Burckle (1990), upper boundary redefined
hereinTop: FO Thalassiosira fasciculata (450.83 mbsf)Base: FO Thalassiosira inura (507.90 mbsf)Stratigraphic position and age: middle early Pliocene, 4.34 to
4.74 MaPaleomagnetic correlation: early to middle Gilbert (C3n.1r to
C3n.2r)
Please cite this article as: Winter, D., et al., Pliocene–Pleistocene diatomMcMurdo Sound, Glob. Planet. Change (2010), doi:10.1016/j.gloplacha.
Discussion: Both the upper and lower boundaries of this zone arewell constrained within diatomite intervals. The upper boundary isredefined here as the FO of T. fasciculata. This zone is expressed in DU-XIIthroughDU-XIb. The FOof Thalassiosira striata (452.25 mbsf) occurs in theupper part of this zone. Thalassiosira torokina is one of themore abundantspecies in this zone. Shionodiscus tetraoestrupii and D. antarcticus havegreater abundance toward the base of the zonewhile E. antarctica ismoreabundance in the upper part. Thalassionema/Thalassiothrix fragments areoften very abundant, with Thalassiothrix antarcticamaking up the greaterproportion of these fragments and Thalassionema nitzschioides in lowabundance. Denticulopsis delicata and D. sp. cf. D. delicata are observed inlow to moderate abundance in this zone. Actinocyclus karstenii andA. octonaris are a prominent part of the assemblage, with the formerobserved consistently and in higher abundance.
biostratigraphy of nearshore Antarctica from the AND-1B drillcore,2010.04.004
8 D. Winter et al. / Global and Planetary Change xxx (2010) xxx–xxx
ARTICLE IN PRESS
Thalassiosira fasciculata Partial Range Zone (new zone)Top: FO Fragilariopsis interfrigidaria (437.59 mbsf)Base: FO Thalassiosira fasciculata (450.83 mbsf)Stratigraphic position and age: middle early Pliocene, 4.21 to
4.49 Ma (4.25 to 4.34 Ma)Paleomagnetic correlation: middle to late Gilbert (C3n.1r)
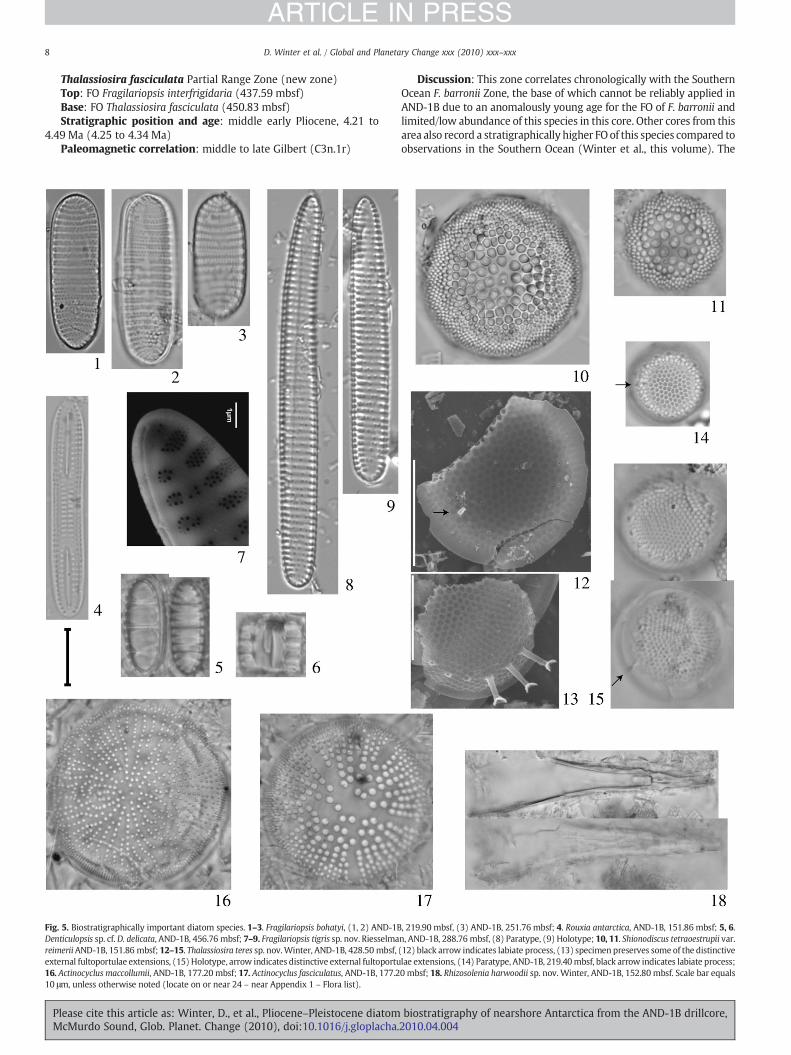
Fig. 5. Biostratigraphically important diatom species. 1–3. Fragilariopsis bohatyi, (1, 2) AND-1BDenticulopsis sp. cf. D. delicata, AND-1B, 456.76 mbsf; 7–9. Fragilariopsis tigris sp. nov. RiesselmareimeriiAND-1B, 151.86 mbsf; 12–15. Thalassiosira teres sp. nov.Winter, AND-1B, 428.50 mbsf,external fultoportulae extensions, (15) Holotype, arrow indicates distinctive external fultoportu16. Actinocyclus maccollumii, AND-1B, 177.20 mbsf; 17. Actinocyclus fasciculatus, AND-1B, 177.210 μm, unless otherwise noted (locate on or near 24 – near Appendix 1 – Flora list).
Please cite this article as: Winter, D., et al., Pliocene–Pleistocene diatomMcMurdo Sound, Glob. Planet. Change (2010), doi:10.1016/j.gloplacha.
Discussion: This zone correlates chronologically with the SouthernOcean F. barronii Zone, the base of which cannot be reliably applied inAND-1B due to an anomalously young age for the FO of F. barronii andlimited/low abundance of this species in this core. Other cores from thisarea also record a stratigraphically higher FOof this species compared toobservations in the Southern Ocean (Winter et al., this volume). The
, 219.90 mbsf, (3) AND-1B, 251.76 mbsf; 4. Rouxia antarctica, AND-1B, 151.86 mbsf; 5, 6.n, AND-1B, 288.76 mbsf, (8) Paratype, (9) Holotype; 10, 11. Shionodiscus tetraoestrupii var.(12) black arrow indicates labiate process, (13) specimen preserves some of the distinctivelae extensions, (14) Paratype, AND-1B, 219.40mbsf, black arrow indicates labiate process;0 mbsf; 18. Rhizosolenia harwoodii sp. nov. Winter, AND-1B, 152.80 mbsf. Scale bar equals
biostratigraphy of nearshore Antarctica from the AND-1B drillcore,2010.04.004

9D. Winter et al. / Global and Planetary Change xxx (2010) xxx–xxx
ARTICLE IN PRESS
more abundant T. fasciculata has greater local utility in this near-shoreAntarctic setting. Its FO is reported as only slightly older than that ofF. barronii in other circum-Antarctic cores. The top of this zone istruncated in AND-1B by an interval of non-deposition, representedsedimentologically by several debris flow deposits and finely laminatedmudstone. The lower boundary iswell constrained. This zone is entirelywithin Diatom Unit DU-XIb which is bounded above and below by twodebris flow units. A distinct assemblage is present within this zone;Thalassionema nitzschioides is often very abundant in these samples.Proboscia barboi, Stephanopyxis species, S. tetraoestrupii and T. inura arealso observed in high abundance, though not equal to T. nitzschioides.Shionodiscus tetraoestrupii is consistently more abundant than T. inura.Silicoflagellates occur in low abundance in half of the samples from thisinterval, and are absent from the rest. The LO of D. sp. cf. D. delicata iscoincidentwith the upper boundary of this zone and diatomite interval.
Fragilariopsis interfrigidaria Partial Range ZoneAuthors: McCollum (1975), upper boundary redefined hereinTop: FO Fragilariopsis bohatyi (291.50 mbsf)Base: FO Fragilariopsis interfrigidaria (437.59 mbsf)Stratigraphic position and age: early late Pliocene to late early
Pliocene, 3.2 to 4.06 MaPaleomagnetic correlation: lower Gauss (C2An.2n to C2An.3n)Discussion: The upper boundary for this zone is redefined by the
FO of F. bohatyi; a new zonal marker species abundant in thiscontinental shelf site. Other datums that have used to identify the topof this zone in open ocean zonations are the FO of T. vulnifica(Harwood and Maruyama, 1992) and the LO of T. insigna (Baldauf andBarron, 1991). Themorphological representation of the first species inAND-1B material reflects an early form not often observed at othersites which is difficult to separate from T. fasciculata, making it lessuseful for biostratigraphy. The second is not present in AND-1Bsamples. This zone is represented in DU-XIa and DU-X. The upper55 m of this zone contains the unit DU-X, a diatom-bearing diamictiteinterval. Some uncertainty exists as to whether the diatoms present inDU-X reflect incorporation of in situ local production or are reworkedinto the diamictite. There is no evidence of older species in DU-X andthe diatom assemblage present indicates a relatively well-constrainedage for this unit. The lower boundary for this zone is truncated,removing the basal range of F. interfrigidaria. Fragilariopsis inter-frigidaria is rare with sporadic occurrence in the lower part of its rangein AND-1B. Silicoflagellates are an abundant component of theassemblage from 398–417 mbsf.
Fragilariopsis bohatyi Partial Range Zone (new zone)Top: FO Actinocyclus maccollumii (201.20 mbsf)Base: FO Fragilariopsis bohatyi (291.50 mbsf)Stratigraphic position and age: early late Pliocene, 2.82 to 3.2 MaPaleomagnetic correlation: upper Gauss (C2An.1n to C2An.2n)Discussion: The lower boundary of this zone, defined by the FO of
F. bohatyi, lies unconformably on the F. interfrigidaria Zone. DiatomUnitsDU-IX throughDU-VII preserves sediments fromthisnewzone. Thebasalage of this zone is somewhat ambiguous due to the glacial surface oferosion at the lower boundary of this zone and the use of a new specieswhose FOage isnotwell constrainedat this time. Though the interpretivework of Naish et al. (2009) suggest an age of 3.2 Ma for this stratigraphicinterval. The base of this zone at 291.50 mbsf is marked by a change inassemblage reflecting a distinct alteration in species dominance. Acomplex of Fragilariopsis species is abundant within this zone, as areChaetoceros spores in some samples. Rare to common Rouxia species arepresent in most samples. Thalassiosira inura and S. tetraoestrupii are alsooften observed in high abundance, their changes in abundance occurboth coincidentally aswell as alternatively. The LOof F. praeinterfrigidaria(251.76 mbsf) occurs within this zone, though the LCO of this species(330.9 mbsf) is slightly older andobserved in the zonebelowat the topofDU-XIa. A new species, Fragilariopsis tigris sp. nov. Riesselman, isdescribed from samples within this zone. It occurs within a restrictedstratigraphic interval, ranging from 284.90–291.50 mbsf (Fig. 5, Appen-
Please cite this article as: Winter, D., et al., Pliocene–Pleistocene diatomMcMurdo Sound, Glob. Planet. Change (2010), doi:10.1016/j.gloplacha.
dix 1). The LO of F. barronii is observed at 251.74 mbsf. The youngestforms range morphologically between F. barronii and Fragilariopsisritscherii, a phenomenon also observed in ODP Leg 188 Site 1165 inPrydz Bay, causing difficulty in determining the true LO for F. barronii(Whitehead and Bohaty, 2003). Harwood and Maruyama (1992)illustrated transitional forms of this species from ODP Leg 120 on theKerguelen Plateau. The FO of T. elliptipora is observed at 288.76 mbsf,extending the lower range for this species on the continental Antarcticshelf. An early form of T. vulnifica is present in this zone, with its FOoccurring at 288.76 mbsf. This species is unusually well preserved at thissite and these early forms exhibit morphologic similarities toT. fasciculata.
Actinocyclus fasciculatus–Actinocyclus maccollumii ConcurrentRange Zone (new zone)
Top: LO Actinocyclus fasciculatus (164.10 mbsf)Base: FO Actinocyclus maccollumii (201.20 mbsf)Stratigraphic position and age: late Pliocene, 2.16 to 2.82 MaPaleomagnetic correlation: upper Gauss to themiddleMatuyama
(C2r.2r to C2An.1n)Subzones: This zone is divided into two subzones. The boundary
between subzones a and b is defined by the FO of Shionodiscustetraoestrupii var. reimerii at 176.70 mbsf (∼2.36 Ma).
Discussion: The entire ranges of A. fasciculatus and A. maccollumiiare confined to this zone; the FO of the first occurring at 191.95 mbsfand the LO of the latter at 176.20 mbsf. The stratigraphic interval forthis zone includes DU-V through DU-VI. The FO of S. tetraoestrupii var.reimerii is observed at 176.70 mbsf, with consistent occurrences up to167.80 mbsf. Its distinct morphology and ease of identification lead toits use as a subzonal marker species. The LO of Creania lacyae isrecorded within subzone b in two distinct peaks at 175.70 and 178.20mbsf. An interval of abundant T. vulnifica is restricted to this zone andthe LO appears to be constricted at the top as compared to other sitesaround the Antarctic continent. Scattered occurrences of T. vulnificathat occur higher in the core are considered reworked due to thecondition of these specimens and presence of other reworked species.An “acme” or last common abundance of T. fasciculata is observed inthis zone between 201.20 and 192.42 mbsf, this event correspondswith the last high abundance of S. tetraoestrupii. Significant changes inthe diatom assemblage include a shift in dominance of species withinthe Fragilariopsis complex, from F. bohatyi to Fragilariopsis robusta, andincreased abundance of Rouxia antarctica. A form intermediatebetween T. inura and T. insigna was observed in subzone a. Anotherreport of this form from ODP Leg 188 Site 1165B is within theT. insigna–T.inura Zone, the lower boundary of which is defined by theFO of T. vulnifica (Whitehead and Bohaty, 2003). The AND-1B sampleat 189.00 mbsf in which this form is common occurs within theinterval of abundant T. vulnifica. Harwood and Maruyama (1992)report and illustrate a similar intermediate form near the samestratigraphic level within their T. insigna–T. vulnifica Zone, its rangeextending into the slightly older F. interfrigidaria Zone.
Rouxia antarctica Partial Range Zone (new zone)Top: LO Rouxia antarctica (116.93 mbsf)Base: LO Actinocyclus fasciculatus (165.10 mbsf)Stratigraphic position and age: early Pleistocene to latest
Pliocene, 1.5 to 2.16 MaPaleomagnetic correlation: middle Matuyama (C1r.3r to C2r.2r)Subzones: Two subzones are present in this zone; the transition
between subzones a and b is represented by the LO of T. kolbei at150.70 mbsf (∼2.2 Ma).
Discussion: The top of subzone a is the upper boundary of unitDU-IV and coincidentwith a glacial surfaceof erosion. The LOof T. kolbei,defining the subzonal boundary, represents a truncated upper range forthis species. Naish et al. (2009) suggested up to 400 k.y. may have beenlost at this erosional surface, though the agemodel for this core (Wilsonet al., 2007) suggests that the LO of this species is near the truestratigraphic position at which it has been observed in other sites
biostratigraphy of nearshore Antarctica from the AND-1B drillcore,2010.04.004

10 D. Winter et al. / Global and Planetary Change xxx (2010) xxx–xxx
ARTICLE IN PRESS
around Antarctica. The FO of Actinocyclus actinochilus is observed at155.80 mbsf in subzone a. This zone encompasses two Diatom Units(DU-III and DU-IV), the uppermost ofwhich is a thin diatomite between116.75 and 118.10 mbsf, in which reworking is evident in the presenceof Denticulopsis species. T. torokina is present consistently throughoutthis zone, with greatest abundance in subzone a. Rouxia species areabundant within this zone, Rouxia diploneides appear in a distinct peaktogether with S. tetraoestrupii var. remerii in the lower part of DU-IVbetween 150.70 and 159.34 mbsf. Alongwith T. kolbei, the upper rangesof several species are truncated at the boundary between subzones aand b at 150.7 mbsf, these include Rhizosolenia harwoodii sp. nov.Winter, R. diploneides, S. tetraoestrupii, S. tetraoestrupii var. reimerii.
Actinocyclus ingens Partial Range ZoneAuthors: Gersonde and Burckle (1990), lower boundary redefined
hereinTop: LO Actinocyclus ingens (86.90 mbsf)Base: LO Rouxia antarctica (116.93 mbsf)Stratigraphic position and age: middle to early Pleistocene, 0.54
to 1.5 MaPaleomagnetic correlation: lower Bruhnes to middle Matuyama
(C1r.1n to C1r.3r).Discussion and remarks: The upper boundary for this zone is
placed at 86.90 mbsf in the AND-1B core, coincident with the top ofthe youngest in situ diatomaceous–rich mudstone interval. This singleinterval, DU-II, is the only diatom-rich unit in which this zone isrepresented. The small unit above, DU-I, is considered to contain areworked assemblage. DU-II, from 96.02–86.90 mbsf, is interpreted tocorrelate with Marine Isotope Stage 31, the last known warminterglacial recording open-marine sedimentation in the southernRoss Sea (Scherer et al., 2008). The T. torokina present in this unit areconsidered in place, thus extending the overall range of this speciesand recording a new youngest occurrence for the Antarctic region.Reworking and glacially-induced fragmentation is evident in theabundant Thalassionema/Thalassiothrix fragments and presence ofDenticulopsis species. Thalassiosira elliptipora is abundant in thesesamples, with a known correlative Acme in this interval in CapeRoberts Project drillcore CRP-1 (Bohaty et al., 1998).
5. Discussion
Diatom assemblages preserved in the AND-1B drillcore vary inresponse to both environmental and evolutionary influences. Speciespresent within diatomite deposited in the early and mid-Plioceneshare assemblage characteristics with Southern Ocean core materialof the same age. Whereas the younger diatomite units in the latePliocene and early Pleistocene record distinct assemblages unique tothis setting. Long-ranging modern sea-ice related diatom species arepresent in low to moderate abundances (D. antarcticus, E. antarctica,F. curta, Porosira spp., S. microtrias), even in assemblages containing ahigher abundance of species associated today with more northerlyoceanic areas (S. stellaris, T. nitzschioides, S. tetraoestrupii). Fig. 2 inWinter et al. (this volume) illustrates abundance changes in of someof the species discussed below for the early to mid-Pliocene.
The diatom biostratigraphy of the Southern Ocean is continuallyrefined with each new drillcore recovered. While the first occurrence(FO) and last occurrence (LO) datums of some species are well definedandnearly isochronous across latitude,manyothers reflect evolutionarymodifications of species lineages in response to environmental changesspecific todifferent geographic andecological settings.Near-shorehigh-latitude locations are perhaps the most likely to contain provincialassemblages and exhibit diachrony within individual species' evolu-tionary events. Comparison of the AND-1B core with the ODP Leg 188Site 1166, drilled on the Prydz Bay shelf, illustrates the regional variancepossible between assemblages. Site 1166 recovered upper Pliocenesediments at its base, with the oldest samples unzoned but below theThalassiosira kolbei Zone. Actinocyclus fasciculatus is rare to common in
Please cite this article as: Winter, D., et al., Pliocene–Pleistocene diatomMcMurdo Sound, Glob. Planet. Change (2010), doi:10.1016/j.gloplacha.
Site 1166 samples, and Thalassiosira oliverana rare to few in theassemblages. In contrast, AND-1B material of the same age withconsistent A. fasciculatus have fewer and less consistent abundances ofT. oliverana, reaching only scarce to rare abundance when present.Previous work in the McMurdo Sound area on three drill-cores (CIROS-2, DVDP-10 and DVDP-11) provide a starting point with which tointegrate the diatom record from AND-1B material (Winter, 1995;Winter and Harwood, 1997; Scherer et al., 2007; Winter et al., thisvolume) with other Antarctic diatom biostratigraphies (McCollum,1975; Schrader, 1976; Weaver and Gombos, 1981; Ciesielski, 1983,1986; Gersonde and Burckle, 1990; Baldauf and Barron, 1991; Harwoodand Maruyama, 1992; Zielinski and Gersonde, 1997).
It was noted in these three cores from the Southern Victoria Landmargin (DVDP-10, DVDP-11, CIROS-2) that many open-ocean markerspecies defining zonal boundaries are present in the near-shore shelfenvironment, and these species can be used to correlate with openocean sequences and infer similar ages for the sediment in which theyare occurring (Winter and Harwood, 1997). A limitation with thesecores is the many unconformities truncating sedimentary unitsreflecting open marine conditions. While sea-level highstands duringwarm periods led to intervals of marine incursion at these veryproximal sites, subsequent episodes of glacial erosion removed muchof the time represented by these near-shore marine deposits. Newmaterial from AND-1B enables us to recognize that the near-shorebiostratigraphic ranges of some of these species do not reflect theirentire range, being more completely recorded in cores from theSouthern Ocean. Some species appear chronologically earlier in thisnear-shore setting (e.g. Thalassiosira complicata, Thalassiosira ellipti-pora), while others have a limited presence expressing only part oftheir entire range (e.g. Fragilariopsis barronii, Proboscia barboi).Several taxa common in the Pliocene Southern Ocean are not presentin AND-1B material (e.g. Fragilariopsis weaverii, Thalassiosira insigna).Specific environmental conditions influencing species temporal andspatial distribution are currently under investigation.
The diatom-rich sediments of the AND-1B drillcore are dividedroughly into four time intervals by alterations in sedimentationpatterns. The biostratigraphic record in the core is discussed in relationto these sedimentary divisions below.
5.1. Early Pliocene
The oldest sediments in AND-1B containing well-preserveddiatoms, present at and above 583.64 mbsf, were deposited in theearly Pliocene during the S. tetraoestrupii zonal interval. Four diatomzones represent the early Pliocene, the youngest of which is discussedseparately in the following section. Four diatomite intervals (DU-XIIIthrough DU-XIb) preserve the oldest three biostratigraphic zones.DU-XIII and DU-XII are a sequence of interbedded diatomite anddiamictite; DU-XIc and DU-XIb are the base of an extended enigmaticdiatomite unit which continues through into the mid-Pliocene. Thisextended diatomite is separated into three distinct time intervals bytwo debris flow sequences (Fig. 3).
The overall character of deposition in the early Pliocene is one ofrelatively brief open-marine interglacial conditions alternating withglacial conditions recorded by deposition of diamictite. The assem-blages preserved within these diatomite units share many character-istics in common with those observed in the Southern Ocean. Ages ofdatums defining zonal boundaries are interpreted to be equivalent inboth near-shore and open-ocean settings.
Early Pliocene assemblages contain greater numbers of A. karstenii,A. octonarius,D. delicata, andD. sp. cf.D. delicata than are observed in theyounger sections of the core. The range of the latter extends only to thetop of the T. fasciculata Zone. Chaetoceros spores are often common insamples from DU-XIII between 552.47 and 583.64 mbsf, as areE. antarctica, S. microtrias and T. torokina. A small peak of T. nitzschioidesis observed at the base of DU-XIII (577.95–581.84 mbsf), followed by an
biostratigraphy of nearshore Antarctica from the AND-1B drillcore,2010.04.004

11D. Winter et al. / Global and Planetary Change xxx (2010) xxx–xxx
ARTICLE IN PRESS
increase in abundance of S. tetraoestrupii. Greater numbers ofT. nitzschioides are associated today with locations north of the PolarFrontal Zone (Pichon et al., 1992). S. oestrupii, descendant ofS. tetraoestrupii, is a subantarctic species indicating surface waterconditions of 7–10 °C, when occurring in abundances of 10% or greater(Zielinski andGersonde, 1997). Creania lacyae is mostly absent from thebasal assemblage with high abundance of T. nitzschioides, it thenabruptly increases in the same samples upcore as S. tetraoestrupii.Creania lacyaehas been suggested to be associatedwith sea ice (Olney etal., 2009). The presence of Chaetoceros spores could indicate increasednutrients and/or stable water conditions as well as warmer overallwater temperatures.
The next younger diatomite (DU-XII) from 503.43–511.09 mbsf isvery similar to the underlying one, with a slightly greater number ofT. oliverana and Thalassiothrix antarctica. The first of these species isassociated today with waters south of the Polar Front, and the secondindicates high productivity when observed in larger numbers and ageneral association with cooler waters. The uppermost part of theT. inura zone is observed in DI-XIc and XIb. Early Pliocene occurrencesof A. ingens are confined to DU-XII through DU-XIc.
5.2. The early to mid-Pliocene diatomite unit
While the lower part of the mid-Pliocene section is confined to theuppermost diatomite unit DU-XIa, between 364.38 and 439.00 mbsf,the entire DU-XI diatomite (364.38–459.24 mbsf) represents anextended period of open-marine productivity uninterrupted by glacialsurfaces of erosion and diamictite recording proximal glacial activity.Two debris flow intervals separate this diatomite into three distinctparts. Three zones are expressed within the whole diatomite interval,the top of the T. inura Zone, the new T. fasciculata Zone, and theF. interfrigidaria Zone (Figs. 3 and 4). Chronostratigraphically, there isan interpreted hiatus of 50.3–80.5 ky between DU-XIa and DU-XIbwith a shorter hiatus between DU-XIb and DU-XIc (Naish et al., 2009).Bioturbation is prominent in some intervals of this diatomite,indicating slower sedimentation rates; laminated intervals recordincreased biogenic productivity and sedimentation rates. The ex-panded depth interval of the diatomite part of the F. interfrigidariaZone is interpreted to result from increased sedimentation due to highproductivity and terrigenous input at the top. Species present withinthe assemblages of this diatomite reflect marine conditions notcurrently present at any location in such proximity to the modernAntarctic coast.
Each interval of DU-XI contains a distinct diatom assemblage. Theassemblages present in DU-XIc (455.27–459.24 mbsf), share morecharacteristics with DU-XIII and DU-XII than the overlying DU-XIband DU-XIa. DU-XIb, containing the T. fasciculata Zone and the veryuppermost portion of the T. inura Zone, is markedly different fromboth DU-XIc and DU-XIa. Thalassionema nitzschioides is oftendominant in assemblages of this interval. The modern cold form ofthis species (Zielinski and Gersonde, 1997) indicates water tempera-tures between 0 and 3 °C. Maximum numbers of the nominate varietyof this species (5%) are today associated with the Polar Frontal Zone,with numbers declining to the north and south of this area. Samplesfrom the very upper part of DU-XIb record abundances of T.nitzschioides greater than 40%.
This interval from 440.12–453.85 mbsf (DU-XIb) that is oftendominated by T. nitzschioides, also records increased abundances ofP. barboi. Other sites around the continent have also reported distinctintervals with both species in the early Pliocene. The former is oftennot observed in corematerial from sites south of the Polar Front due toits ecological preferences, thus its presence in other cores formsdiscrete patterns within species data. Sites 747, 748 and 751, fromODP Leg 120, all report these two species co-occurring in lowerPliocene sections (Harwood and Maruyama, 1992). The samples withboth species from Site 747 are from the entire early Pliocene. In Site
Please cite this article as: Winter, D., et al., Pliocene–Pleistocene diatomMcMurdo Sound, Glob. Planet. Change (2010), doi:10.1016/j.gloplacha.
751 this pair begins to be observed together in the late MioceneNitzschia denticulopsis Zone and extends up to the Nitzschia barroniiZone (now F. barronii). Site 748 records the tightest age constraint ontheir co-occurence, as they are observed only in the N. barronii Zoneand at the base of the N. interfrigidaria Zone. The T. fasciculata Zone inAND-1B is coeval with the N. barronii Zone. Site 1165 in Prydz Bay,ODP Leg 188, reports higher abundances of T. nitzschioides withP. barboi from the lower N. interfrigidaria Zone to the T. inura Zone(Whitehead and Bohaty, 2003). The record of these two species inmaterial from Leg 178 along the Antarctic Peninsula contains onlyinconsistent and often rare occurrences of these two species, so acomparison is not possible (Winter and Iwai, 2002). It would seemtheir co-occurrence in sediments of this age from various locationsaround the continent represents an incursion of warmer water intoAntarctic near-shore areas and the southern Ross Sea, persisting for alonger period of time at sites further from the continent. Terrestrialmarine deposits recording increased sea level and near-shore marineproductivity exist in the Vestfold Hills region in East Antarctica andare temporally coincident with this unit. Many of the same speciesobserved in these assemblages in AND-1B are present inmaterial fromthe Sørsdal Formation alongwith a non-cryophillic cetacean; togetherthey are interpreted to suggest summer sea surface temperatureswere at times 4–5 °C (Whitehead et al., 2001).
DU-XIa, extending from 364.38–439.00 mbsf, does not have adominant assemblage; rather different assemblages are present overdifferent depth intervals. One noticeable alteration in the overallbiogenic assemblage occurs between 364.38 and 418.90 mbsf wheresilicoflagellates are abundant, with the greatest numbers present inthe interval between 400.60 and 418.90 mbsf. Occurrences ofF. barronii are also confined to the upper part of DU-XIa but not inequal abundance or as consistently as the silicoflagellates, occurringfrom 401.10–413.60 mbsf as scarce occurrences. All of the silico-flagellates observed belong to the genus Distephanus, and as suchcannot be used to infer water temperature as has been done in otherareas where genus Dictyocha is noted (Bohaty and Harwood, 1998).Variations in assemblages for this upper interval record a dynamic andrapidly fluctuating glacial history of the region.
The uppermost part of the F. interfrigidaria Zone (DU-X) is a uniquediamictite unit in the often abundant diatoms preserved within it. Allother diamictite units in this core are either barren of diatoms orcontain only a rare reworked few. This diamictite has a very diverseand contemporaneous assemblage of diatoms. The sedimentologicalinterpretation for this unit is one of subglacial emplacement (Krisseket al., 2007). Diatomaceous material in this diamictite is thought tohave several possible sources. It may be derived from glacialincorporation of underlying diatomite, and it also may also reflectinclusion of contemporary open-water production. The latter may beindicated by some species, such as F. barronii, occurring in slighterhigher abundance in this unit than are observed in the diatomitebelow. Any reworked diatomite in this diamictite unit appears to besourced only from the underlying diatomite as biostratigraphicallyolder species are not present in the assemblage; this assemblage alsodoes not have evident alteration by glacial transport.
5.3. Late mid-Pliocene to late Pliocene
The upper mid-Pliocene and late Pliocene of AND-1B is contained indiatomite units occurring between 150.70 and 292.70 mbsf (DU-IIIthrough DU-IX). The pattern of interbedded diatomite and diamictitereappears in the late mid-Pliocene and continues through to the earlyPleistocene. Reworked older diatom species are not present inassemblages from these intervals. We interpret this to reflect highbioproductivity during intervals of diatomite deposition and anytransport and redeposition of older material is either not occurring, orrepresents a negligible contribution to sedimentation. The assemblagesin this interval are often dominated by a Fragilariopsis group that has not
biostratigraphy of nearshore Antarctica from the AND-1B drillcore,2010.04.004

12 D. Winter et al. / Global and Planetary Change xxx (2010) xxx–xxx
ARTICLE IN PRESS
been reported in this complexity and abundance previously. This groupdisplays a rapid diversification in the latemid-Pliocene and the differentvarieties shift in relative abundance through the late Pliocene. SouthernOcean species defining zonal boundaries for the late Pliocene are oflimited utility in the near-shore setting of McMurdo Sound due to theirunderrepresentation in the assemblage and the higher abundance ofother species.
Southern Ocean zonal marker species T. vulnifica is present inmoderate abundance from 169.80–201.40 mbsf, with sporadic occur-rences of low abundance down to 251.76 mbsf (Table 1, Figs. 3 and 4).Early forms of T. vulnifica share morphological similarities withT. fasciculata, but have a reduced number ofmarginal strutted processes,placing them within T. vulnifica (Mahood and Barron, 1996). The FO ofT. elliptipora is observed near the base of DU-IX, in low but consistentnumbers. This species occurs in moderate abundance and is a fairlyconsistent part of assemblages in the late Pliocene, slightly higherabundances are observed in the early Pleistocene interval discussedbelow. Mahood and Barron (1996) and Baldauf and Barron (1991) bothobserved the FO of T. ellilptipora to occur younger than the FO ofT. vulnifica; the opposite order of these two events in AND-1B extendsthe known lower range of the T. elliptipora for the Southern Ocean. Thespecies F. weaverii was reported from initial on-ice work at the base ofthe F. bohatyi Zone, from 284.90–291.50 mbsf, but was later found to bea new Fragilariopsis species, potentially endemic to the Ross Sea, and isdescribed herein as a new species (see F. tigiris in Appendix 1 – Florallist) (Scherer et al., 2007). Rouxia species are very common in DU-VIthrough DU-IV, the upper part of the late Pliocene interval (∼150–200 mbsf). Rouxia antarctica and R. diploneides are present throughoutmost of the AND-1B drillcore, but reach higher abundances in theseuppermost diatomite units (Fig. 3). The environmental interpretation ofR. antarctica is uncertain but the co-occurrence of this species withF. curta and other sea-ice species suggest that R. antarctica is possiblyindicative of colder conditionswith increased sea ice. Thalassiosira inuraand S. tetraoestrupii increase in abundance as sample depth increases;A.karstenii occur in higher numbers from ∼150–200 mbsf with lowerabundance observed below. Stellarima microtrias and T. torokina arepresent in moderate numbers throughout. The species present in theassemblages suggest a variable environment of deposition for the latePliocene,with relativelywarmer conditionspreservedbetween210 and225 mbsf (though not as warm as those existing during the early mid-Pliocene) and cooler conditions above, up to 150 mbsf.
5.4. Early Pleistocene
The early Pleistocene diatom record is represented in the upper150 m of the AND-1B drillcore. Diatom-rich sediments in this intervalare mudstone units at 58.15–58.88, 86.90–92.24 and 116.93–118.10 mbsf, DU-I through DU-III. DU-I is considered an entirelyreworked assemblage, offering no biostratigraphic utility. Theassemblages in DU-II and DU-III are thought to represent in situproductivity though they also contain common evidence of reworkeddiatoms. McCollum (1975) noted that Cores 1–6 of DSDP Site 274contained obviously reworked Miocene to Oligocene diatom species,in an inverted sequence. These cores extend into his T. kolbei/P. barboiZone, roughly equivalent with the R. antarctica Zone of AND-1B; theupper part of which contains DU-III mentioned above. This earlyPleistocene section of the core contains the greatest number ofreworked diatoms in the drillcore; the older diatomite assemblages inthe core contain very few reworked species. The A. ingens Zone andSubzone b of the R. antarctica Zone are within this time period.
Species indicative of reworking in the upper 150 m of the AND-1Bdrillcore are Denticulopsis lauta, Denticulopsis maccollumii, Denticulopsissimonsenii, Paralia sulcata, Stephanopyxis species, and Trinacria species;these species are all robust and likely to preserve in transported andreworked sediments. The first three are restricted to the Miocene; theothers are long-ranging species. Thalassiosira elliptipora is abundant in
Please cite this article as: Winter, D., et al., Pliocene–Pleistocene diatomMcMurdo Sound, Glob. Planet. Change (2010), doi:10.1016/j.gloplacha.
these upper diatom-rich units. Actinocyclus actinochilus is an importantmember of the assemblages from the Pleistocene interval of AND-1B; itcontinues downcore into DU-IV, below the unconformity at150.70 mbsf, but in lower abundance. Actinocyclus actinochilus isobserved in even older sediments at other sites around the continent,so this represents an abbreviated lower range for this species, likelyresponding to local environmental differences at this site. This species isassociated today with colder waters (−2 to 1 °C; Zielinski andGersonde, 1997) with only a fewmonths of ice-free conditions (Pichonet al., 1987). Its abbreviated range here is unexpected for such asoutherly location. It is possible other environmental variables thantemperature and ice cover play a role in its temporal and geographicdistribution. Other species present in higher numbers in earlyPleistocene assemblages are A. karstenii, E. antarctica, S. microtrias,T. torokinaand T. vulnifica. Smaller and lighter species suchas F. curta andT. antarctica are rare in these sediments suggesting that contempora-neous primary productivity is accompanied by some winnowing ortransport. The sample interval between 86.90 and 92.24 mbsf, DU-II,was deposited during Marine Isotope Stage 31, recording the youngestinterval of open-marine productivity in the McMurdo Sound and arelated significant reduction in sea-ice cover. The age of DU-II is∼1.06 Ma, as derived fromorbital tuning. This age is further constrainedby the position of this diatomite in the lower part of the Jaramillosubchron (Fig. 4); a time of unusually warm interglacial conditionscharacterized by very high solar insolation (Scherer et al., 2008). Several40Ar/39Ar ages bounding this diatom-rich interval support this age; oneat 84.97 mbsf and another at 91.13 mbsf have associated ages of0.988 Ma and 1.072 Ma, respectively.
6. Conclusions
This new drillcore from southern McMurdo Sound provides anextended glimpse into the previously unknown record of the Plio-Pleistocene open-marine environment of near-shore Antarctica. Diatomassemblages of the early Pliocene share many similarities with thosefrom the Southern Ocean. Those in the middle/late Pliocene containfewer species common in open-ocean cores, replaced by a moreendemic group of species reflecting the differences between the coolingdynamic coastal environment and the cool but still stable open ocean.
Incomplete ranges of well-established Southern Ocean markerspecies, and the unique assemblages observed in the late mid-Pliocene to early Pleistocene, result in the proposal of three newzones, two of which are further subdivided into to two subzones each(Fig. 2). These are the F. bohatyi Zone, the A. fasciculatus–A.maccollumiiZone and the R. antarctica Zone. The fourth new zone proposed here,the T. fasciculata Zone is in the early Pliocene and utilizes the FO of thenominative taxon as its base. Comparison of assemblages present inthis time period with sites from other locations around the continentsuggest that this new zonation will be a useful tool for subsequentdiatom biostratigraphic work in other continental shelf areas.
Distinct ‘glacial’ diamictite alternates with ‘interglacial’ diatomitereflecting open-marine diatom production interspersed betweenglacial intervals from 459–584 mbsf. A diatomite/diatom-rich intervalextending nearly 100 m was deposited during the early late to mid-Pliocene, extending across three diatom zones. This interval reflects aperiod of extended open-marine conditions in the southern Ross Seawith no indication of sediment processes directly related to glacialactivity. The time represented could be up to 1.5 Ma in duration, but islikely less as two debris flow units subdivide the diatomite into threesections. Each of these is interpreted to be associated with a hiatus,though the amount of time contained in each is presently uncertain.The middle section of the long diatomite interval contains a distinctassemblage often dominated by T. nitzschioides, distinct from eithersection of the diatomite above and below. This interval correlates withevidence of warmer open-marine productivity at other locationsaround the continent suggesting the impact of the environment
biostratigraphy of nearshore Antarctica from the AND-1B drillcore,2010.04.004

13D. Winter et al. / Global and Planetary Change xxx (2010) xxx–xxx
ARTICLE IN PRESS
producing this diatomite was extensive and continent-wide. DU-XIarecords a dynamic glacial history for this time interval throughchanges in diatom assemblages. The pattern of alternating diatomiteand diamictite resumes above this long diatomite interval, indicatinga return of periodic glacial activity proximal to the AND-1B site.
The late Pliocene preserves assemblages dominated by a group ofrelated Fragilariopsis species. This group of species has not beenobserved before in such complexity or abundance from other sitesaround the Antarctic continent. Subsequent work notes members ofthis group existing in lower numbers at some sites, previouslyidentified as other species. The upper 150 m of the AND-1B drillcorerecords a change in the pattern of preservation of diatom-richmaterial. One small diatomite is preserved from 116–118 mbsf,above this the two short intervals containing diatoms are mudstoneunits. Most of the Pleistocene is represented by a series of diamictiteunits barren of diatomaceous material.
Acknowledgements
The reviewers and editor provided many excellent comments,greatly improving this paper. The basis for figure three is the result ofwork by Dr. Richard Levy, his kindness in sharing it is muchappreciated. This material is based upon work supported by theNational Science Foundation under Cooperative Agreement No.0342484 through subawards administered by the ANDRILL ScienceManagement Office at the University of Nebraska–Lincoln, and issuedthrough Northern Illinois University (for the MIS Project), as part ofthe ANDRILL U.S. Science Support Program. Any opinions, findings,and conclusions or recommendations expressed in this material arethose of the author(s) and do not necessarily reflect the views of theNational Science Foundation.
The ANDRILL (ANtarctic geologic DRILLing) Program is a multina-tional collaboration between the Antarctic Programs of Germany,Italy, New Zealand and the United States. Antarctica New Zealand isthe project operator and has developed the drilling system incollaboration with Alex Pyne at Victoria University of Wellingtonand Webster Drilling and Exploration. The U.S. Antarctic Program(USAP) and Raytheon Polar Services Corporation (RPSC) supportedthe science team at McMurdo Station and in the Crary Science andEngineering Laboratory, while Antarctica New Zealand supported thedrilling team at Scott Base. Scientific studies are jointly supported bythe US National Science Foundation, NZ Foundation for Research, theItalian Antarctic Research Program, the German Science Foundationand the Alfred Wegener Institute.
Appendix 1. Floral list
Achnanthes brevipes var. angustata (Greville) CleveActinocyclus actinochilus (Ehrenberg) SimonsenActinocyclus dimorphus (Castracane) Harwood et MaruyamaActinocyclus fasciculatus Harwood et Maruyama (Fig. 5, No. 16)Actinocyclus ingens RattrayActinocyclus ingens var. nodus BaldaufActinocyclus karstenii Van HeurckActinocyclus maccollumii Harwood et Maruyama (Fig. 5, No. 15)Actinocyclus octonarius EhrenbergActinocyclus octonarius v. tenella (Brebisson) HustedtActinocyclus sp. A Scherer, 1991Actinocyclus sp. C Harwood et Maruyama, 1992Actinocyclus spp.Actinoptychus spp.Amphora helenensis GiffenAulacodiscus browneii McCollumBacillaria paradoxa GmelinBiddulphia aurita (Lyngbye) Brébisson et GodeyChaetoceros bulbosum (Ehrenberg) Heiden, in Heiden and Kolbe
Please cite this article as: Winter, D., et al., Pliocene–Pleistocene diatomMcMurdo Sound, Glob. Planet. Change (2010), doi:10.1016/j.gloplacha.
Chaetoceros mitra (Bailey) CleveChaetoceros spp.Cocconeis costata GregoryCorethron criophilum CastracaneCoscinodiscus asteromphalus EhrenbergCoscinodiscus marginatus EhrenbergCoscinodiscus oculus-iridus EhrenbergCoscinodiscus radiatus EhrenbergCoscinodiscus spp.Creania lacyae OlneyDactyliosolen antarcticus CastracaneRemarks: This species is present in low numbers sporadically
throughout AND-1B, except in the highly laminated interval at166.80–167.90 mbsf (Krissek et al., 2007). Within these laminationsthere are some that are nearly monospecific for this species, smallerpercentages of Actinocyclus, Corethron and Rouxia species are presentas well.
Denticulopsis sp. cf. D. delicataRemarks: This species is distinct morphologically, but has yet to be
given a formal name. Iwai and Winter (2002) referred to it as D. sp. cf.D. delicata andwe continue this herein. It is often observed in associationwith D. delicata, when occurring without this species it is usuallyobserved in low numbers. The biostratigraphic range for D. sp. cf.D. delicata extends from near the lowest diatom-bearing sediments, at581.84 mbsf, and persists in greater numbers until 455.27 mbsf. DU-XIbcontains inconsistent and generally lower numbers of this species, withonly rare occurrences above 440.12 mbsf (Fig. 5, Nos. 5, 6).
Denticulopsis delicata Yangasawa et AkibaDenticulopsis dimorpha (Schrader) SimonsenDenticulopsis hustedtii (Simonsen et Kanaya) SimonsenDenticulopsis lauta (Bailey) SimonsenDenticulopsis maccollumii SimonsenDenticulopsis simonsenii Yanagisawa et AkibaDenticulopsis spp.Diploneis bombus EhrenbergDiploneis spp.Eucampia antarctica var recta (Mangin) Fryxell et PrasadFragilariopsis aff. sublinearis MISFragilariopsis angulata (O Meara) HasleFragilariopsis arcula (Gersonde) Gersonde et BárcenaFragilariopsis aurica (Gersonde) Gersonde et BárcenaFragilariopsis barronii (Gersonde) Gersonde et BárcenaRemarks: At the upper range of this species, this species begins to
resemble F. ritscherii. Whitehead and Bohaty (2003) discuss thedifficulties in determining the true LOof this species due to the presenceof a F. sp. cf. barronii that appears to be a transitional form between thetwo from ODP Leg 188 Site 1165. Harwood and Maruyama (1992)illustrate these same transitional forms in Plate 17, Figures 27 and 28.
Fragilariopsis bohatyi Sjunneskog (Fig. 5, Nos. 1–3)Fragilariopsis curta (Van Heurck) HustedtFragilariopsis interfrigidaria (McCollum) Gersonde et BárcenaFragilariopsis clementia (Gombos) Zielinski et GersondeFragilariopsis obliquecostata (Van Heurck) HasleFragilariopsis peragallii (Hasle) CremerFragilariopsis praecurta (Gersonde) Gersonde et BárcenaFragilariopsis praeinterfrigidaria (McCollum) Gersonde et BárcenaFragilariopsis ritscheri HustedtFragilariopsis sp. in Bohaty et al 1998Fragilariopsis spp.Fragilariopsis sublinearis (Van Heurck) HeidenFragilariopsis vanheurckii (Peragallo) HustedtFragilariopsis tigris sp. nov. RiesselmanDescription: Valve outline linear–lanceolate to oval, heteropolar.
One apex obtusely rounded and the other tapering to an acutelyroundedor pointed tip. Rapheeccentric, located in the junction betweenvalve faceandmantle. In some specimens, the taperedapex curves away
biostratigraphy of nearshore Antarctica from the AND-1B drillcore,2010.04.004

14 D. Winter et al. / Global and Planetary Change xxx (2010) xxx–xxx
ARTICLE IN PRESS
from the raphidmargin (Fig. 4, No. 7). Apical axis 38.0–83.9 µm(median53.0 µm), transapical axis 5.6–7.1 µm at widest point (median 6.6 µm).Transapical costae parallel and straight to slightly oblique in themiddleof the valve, becoming curved toward the blunt apex, 7–10 (median 9)in 10 µm. Valve face incompletely striated by punctae; observed underSEM, these form clusters near the valve margin and decrease toirregularly spaced single or paired punctae in a linear arrangementacross themiddle portion of the valve.Marginal clusters of punctae givethe margin a ribbed appearance under light microscopy.
Synonomy: Nitzschia sp. A of Scherer, 1991, p. 405, pl.III, fig. 8Type Locality: AND-1B drillcore, 77.89° S, 167.09° E, McMurdo Ice
Shelf in the southern McMurdo Sound, Ross Sea, AntarcticaType Level and Age: early late Pliocene, AND-1B, 288.76 mbsfStratigraphic Range: Restricted to early late Pliocene in the lower
portion of the F. bohatyi diatom zone (this paper). In AND-1B, F. tigrisis limited to DU IX (284.90–291.50 mbsf), with peak abundance at288.33–291.22 mbsf.
Discussion and remarks: Fragilariopsis tigris has affinities to thePliocene Antarctic evolutionary lineage F. praeinterfrigidaria (McCol-lum) Gersonde et Bárcena – F. interfrigidaria (McCollum) Gersonde etBárcena – F. weaverii (Ciesielski) Gersonde et Bárcena in thearrangement of costae and marginal ribs, but differs from these threespecies in its distinctive heteropolar valve outline. F. tigris is probablyconspecific with a taxon labeledNitzschia (?) sp. A by Scherer (1991). Asingle broadapexof Scherer'sN. sp.Awasdescribed froma subglacial tillsample collected beneath Ice Stream B at 83°28′40″ S, 138°05′49″ W(UpB). The occurrence of F. tigris in AND-1B and in the UpB samplesuggests that this specieswas likely endemic to the Ross Sea continentalshelf in the late Pliocene. Based on its stratigraphic position in AND-1Band onmorphologic similarities, it is possible that F. tigris is a near-shorecorollary to the open-ocean species F. weaverii. In fact, tapered-apexfragments of F. tigris were initially reported as F. weaverii from on-icework. Its narrow stratigraphic range in AND-1B may provide a usefulbiostratigraphic constraint for future Ross Sea drilling.
Etymology: From the Latin tigris, in reference to the tiger-likenature of valve face striation, particularly evident under SEM;originally from the Old Persian tigr, translated as “fast” or “arrow-like” (Fig. 5, Nos. 7, 8).
Grammatophora spp.Gyrosigma spp.Hyalodiscus spp.Isthmia spp.Navicula spp.Nitzschia fossilis (Frenguelli) Kanaya, in Kayana and KoizumiNitzschia sicula (Castracane) HustedtNitzschia sp. 17 of Schrader, 1976Nitzschia spp.Nitzschia stellata ManginOdontella weissflogii (Janisch) GrunowParalia architecturalis (Brun) FennerParalia sulcata (Ehrenberg) ClevePinnularia spp.Pleurosigma spp.Porosira glacialis (Grunow) JörgensenPorosira pseudodenticulata (Hustedt) Jousé, in KozlovaProboscia barboi (Brun) Jordan et PriddleRemarks: The LO of this species in AND-1B is truncated and thus
not the true last occurrence of this species, as it ranges up to 1.6–1.7 Ma (Cody et al, 2008). Zielinski and Gersonde (1997) have the LOof this species at 1.8 Ma in their Southern Ocean zonations.
Proboscia truncata (Karsten) Nothig et Ligowski, ex Jorden et al.Pseudogomphonema spp. MedlinRhizosolenia antennata f. antennata SundströmRhizosolenia antennata f. semispina SundströmRhizosolenia barboi (Brun) Tempère and PeragalloRhizosolenia crassa Schimper ex Karsten 1905
Please cite this article as: Winter, D., et al., Pliocene–Pleistocene diatomMcMurdo Sound, Glob. Planet. Change (2010), doi:10.1016/j.gloplacha.
Rhizosolenia harwoodii sp. nov. WinterDescription: Valve heavily silicified; linear rows of fine punctae
parallel to valve axis; separated by thickened silica ridges, also orientedparallel to valve axis; terminal end of process blunt and rounded.
Synonomy: Rhizosolenia sp. D Harwood et Maruyama, 1992, Pl. 18,Figs. 7–10; Rhizosolenia sp. A. of Akiba, 1986, Pl. 18, Fig. 11; Rhizosoleniasp. A of Koizumi,1968, Pl. 34, Figs. 2a–b.
Type Locality: AND-1B drillcore, 77.89° S, 167.09° E, McMurdo IceShelf in the southern McMurdo Sound, Ross Sea, Antarctica
Type Level and Age: late PlioceneStratigraphic Range: early Pliocene to PleistoceneDiscussion and remarks: This species is very robust and can be
easily reworked.Etymology: This species is named for Dr. David Harwood, whose
informal designation has been used in many publications for thisspecies (Fig. 5, No. 17).
Rhizosolenia sima f. silicea SundströmRhizosolenia spp.Rhoicosphenia spp.Rouxia antarctica Heiden, in Heiden and Kolbe, (Fig. 5, No. 4)Rouxia constricta Zielinski et GersondeRemarks: There are many specimens throughout AND-1B that
have elongate, tapered ends that do fit into the definitions of neitherR. constricta nor R. leventerae. As R. isopolica was synonomized withR. constricta, we group all of this general morphology into this speciesthroughout core.
Rouxia diploneides SchraderRemarks: There are high abundances of this species (acme) from
154.76–156.3 mbsf, max 20%, ave 6.5%, median 4.7%. For the samplesapart from these between 151.86 and 166.7 mbsf this species occursas 2–5% within the assemblage.
Rouxia heteropolara GombosRouxia leventerae Bohaty, Scherer et Harwood 1998Rouxia naviculoides SchraderRouxia spp.Sellaphora spp.Shionodiscus frenguelliopsis (Fryxell et Johansen) Alverson, Kang et
TheriotShionodiscus gracilis var. expectus (Van Landingham) Alverson,
Kang et TheriotShionodiscus gracilis var. gracilis (Karsten) Alverson, Kang et
TheriotShionodiscus poroseriatus (Ramsfjell) Alverson, Kang et TheriotShionodiscus ritscherii (Hustedt) Alverson, Kang et TheriotShionodiscus tetraoestrupii (Bodén) Alverson, Kang et TheriotRemarks: The modern descendent in this lineage, S. oestrupii, is
only found in subantarctic waters today with highest abundancesoccurring where surface water temperatures are above 7–10 °C(Zielinski and Gersonde, 1997). Due to the presence of S. tetraoestrupiithroughout the AND-1B core, and its co-occurrence with other speciesthat have modern ecological constraints of much cooler conditionsand in some cases strong association with sea ice, the exactenvironmental associations of this fossil species are as of yet unclear.The species, S. frenguelliopsis, which is very similar to S. tetraoestrupii,is often found co-occurring in assemblages together. The assemblageswith both species will usually have other species with modernecological constraints of more northerly, warmer conditions; such asS. stellaris and T. nitzschioides. This species is not illustrated here due tothe existing illustrations in other biostratigraphic publications, pleaserefer to Harwood and Maruyama (1992; Pl. 16, Figs. 5–7) andJohansen and Fryxell (1985; Figs. 75, 76).
Shionodiscus tetraoestrupii var. reimeri (Mahood et Barron) Alverson,Kang et Theriot (Fig. 5, Nos. 9, 10)
Stellarima microtrias (Ehrenberg) Hasle et P.A. SimsStellarima stellaris (Roper) Hasle et SimsStephanopyxis dimorpha Schrader
biostratigraphy of nearshore Antarctica from the AND-1B drillcore,2010.04.004

15D. Winter et al. / Global and Planetary Change xxx (2010) xxx–xxx
ARTICLE IN PRESS
Stephanopyxis spp.Stephanopyxis superbus (Greville) GrunowStephanopyxis turris (Greville et Arnott) Ralfs, in PritchardSynedropsis cheethamii OlneySynedropsis hyperboreoides Hasle, Syvertsen et MedlinSynedropsis laevis (Heiden) Hasle, Medlin et SyvertsenSynedropsis recta Hasle, Medlin et SyvertsenSynedropsis spp.Thalassionema /Thalassiothrix spp.Thalassionema nitzschioides (Grunow) Van HeurckThalassiosira antarctica ComberThalassiosira antarctica type 1Thalassiosira australis PeragalloThalassiosira complicata GersondeThalassiosira convexa MuchinaThalassiosira delicatula HustedtThalassiosira eccentrica (Ehrenberg) CleveThalassiosira elliptipora (Donahue) FennerThalassiosira fasciculata Harwood et MaruyamaRemarks: This species is not illustrated here due to the existing
illustrations in other biostratigraphic publications, please refer toHarwood and Maruyama (1992; Pl. 15, Figs. 4–6).
Thalassiosira ferelineata Hasle et FryxellThalassiosira fraga Schrader, in Schrader and FennerThalassiosira insigna (Jousé) Harwood et MaruyamaThalassiosira insigna/inuraThalassiosira inura GersondeThalassiosira jacksonii Koizumi et BarronThalassiosira kolbei (Jousé) GersondeThalassiosira lentiginosa (Janisch) FryxellThalassiosira lineata JouséThalassiosira nativa Sheshukova-PoretzkayaThalassiosira nidulus (Tempère & Brun) JouséThalassiosira oliverana (O Meara) SourniaThalassiosira oliverana v. sparsa Harwood et MaruyamaThalassiosira praefraga Gladekov et BarronThalassiosira spp.Thalassiosira striata Harwood et MaruyamaThalassiosira symmetrica SimonsenThalassiosira teres sp. nov. WinterDescription: valvediameter10–25 μm,16.5 μmaverage(n=35);12–
14 areolae in 10 μm arranged in a somewhat tangential pattern; centralareolae can be oddly shaped and enlarged in comparison to others onvalve face; ring of fultoportulae on shoulder of valve face 3–5 areolae fromvalvemantlewith 4 satellite pores;mantle flattened, especially in smallervalves; single rimoportula process present either in line with ring offultoportulae or slightly inside, oriented radially towards the center of thevalve; external extensions of fultoportulae and rimoportula often broken.
Holotype: Deposited in the California Academy of Sciences, acc.#627361 slide #222083, AND-1B, 428.5 mbsf.
Type Locality: AND-1B drillcore, 77.89° S, 167.09° E, McMurdo IceShelf in the southern McMurdo Sound, Ross Sea, Antarctica
Type Level and Age: early Pliocene, sample AND-1B, 428.50 mbsfParatype: Deposited in the California Academy of Sciences, acc.
#627359 slide #222081, AND-1B, 219.40 mbsf.Stratigraphic Range: early Pliocene to early PleistoceneDiscussion and remarks: The two valves are often together,
perhaps due to its small size or general morphological properties. Thisspecies is very delicate, often broken and universally difficult toimage. Some valves begin to have a more irregular pattern of areaolaeon the valve face, similar to T. antarctica.
Etymology: The species name teres was chosen due to the fine,elegant structureof thevalves in the lightmicroscope (Fig. 5,Nos. 11–14).
Thalassiosira torokina BradyRemarks: This species is present throughout AND-1B. Its early
Pliocenemorphology tends to have a flatter shape across the valve face.
Please cite this article as: Winter, D., et al., Pliocene–Pleistocene diatomMcMurdo Sound, Glob. Planet. Change (2010), doi:10.1016/j.gloplacha.
As it ranges up towards it new youngest LO, the valve face shapebecomes domedwith a high pervalvar axis. The occurrenceswithin DU-II are considered to be in place, associated with warm MIS stage 31.
Thalassiosira tumida (Janisch) HasleThalassiosira vulnifica (Gombos) FennerRemarks: The early part of the stratigraphic range of this species
has been preserved in this core. This site has recorded the earliestknown occurrence of this species. These early forms exhibit striaecounts similar to T. fasciculata, being around 10 in 10 μm (shared byboth), as well as more numerous strutted processes on the valve facethan occurs in later forms. The distinct dissolution of these processesobserved in many Southern Ocean sites is lacking in these early forms.
Thalassiosira webbi Harwood et MaruyamaThalassiothrix antarctica Schimper ex KarstenThalassiothrix longissima Cleve and GrunowThalassiothrix spp.Trachyneis aspera (Ehrenberg) CleveTriceratium groningensis ReinholdTrichotoxon reinboldii (Van Heurck) Reid et RoundTrinacria excavata HeibergTrinacria spp.Chrysophyte spp.Silicoflagelate spp.
References
Alverson, A.J., Kang, S.-H., Theriot, E.C., 2006. Cell wall morphology and systematicimportance of Thalassiosira ritscheri (Hustedt) Hasle, with a description ofShionodiscus gen. nov. Diatom Research 21 (2), 251–262.
Armand, L.K., Crosta, X., Romero, O., Pichon, J.-J., 2005. The biogeography of major diatomtaxa in Southern Ocean sediments: 1. Sea ice related species. Palaeogeography,Palaeoclimatology, Palaeoecology 223, 93–126.
Ashworth, A.C., Cantrill, D.J., 2004. Neogene vegetation of the Meyer Desert Formation(Sirius Group) Transantarctic Mountains, Antarctica. Palaeogeography, Palaeocli-matology, Palaeoecology 213 (1–2), 65–82.
Baldauf, J.G., Barron, J.A., 1991. Diatom biostratigraphy: Kerguelen Plateau and PrydzBay regions of the Southern Ocean. In: Barron, J., Larsen, B., et al. (Eds.), Proc. ODP,Sci. Results, 119. Ocean Drilling Program, College Station, TX, pp. 547–598.
Barker, P.F., Camerlenghi, A., Acton, G.D., et al., 1999. Proc. ODP, Init. Repts., 178. OceanDrilling Program, College Station, TX10.2973/odp.proc.ir.178.1999.
Barrett, P.J., 1985. Plio-Pleistocene glacial sequence cored at CIROS 2, Ferrar Fjord,western McMurdo Sound. New Zealand Antarctic Record 6 (2), 8–19.
Barrett, P.J. (Ed.), 1986. Antarctic Cenozoic history from the MSSTS-1 drillhole, McMurdoSound, Antarctica: NZ DSIR Bulletin, 237. Science Information Publishing Centre,Wellington. 174 pp.
Barrett, P.J. (Ed.), 1989. Antarctic Cenozoic history from the CIROS-1 drillhole,McMurdo Sound, Antarctica: NZ DSIR Bulletin, 245. Science Information PublishingCentre, Wellington. 254 pp.
Barrett, P.J., Hambrey, M.J., 1992. Plio-Pleistocene sedimentation in Ferrar Fjord,Antarctica. Sedimentology 39, 109–123.
Bodén, P., 1993. Taxonomy and stratigraphic occurrence of Thalassiosira tetraoestrupiisp. nov. and related species in the upper Miocene and lower Pliocene sedimentsfrom the Norwegian Sea, North Atlantic and northwest Pacific. Terra Nova 5, 61–75.
Bohaty, S.M., Harwood, D.M., 1998. Southern Ocean Pliocene paleotemperature variationfrom high-resolution silicoflagellate biostratigraphy. Marine Micropaleontology 33(3–4), 241–272.
Bohaty, S.M., Scherer, R.P., Harwood, D.M., 1998. Quaternary diatom biostratigraphyand palaeoenvironments of the CRP-1 drillcore, Ross Sea, Antarctica. TerraAntarctica 5 (3), 431–453.
Cape Roberts Science Team, 1998. Initial Report on CRP-3. Terra Antartica 5, 1–187.Cape Roberts Science Team, 1999. Studies from the Cape Roberts Project, Ross, Sea,
Antarctica – Initial Report on CRP-2/2A, Terra Antartica, 6, 1-173, with supplement.Cape Roberts Science Team, 2000. Studies from the Cape Roberts Project, Ross, Sea,
Antarctica, Initial Report on CRP-3. Terra Antartica, 7, 1–209 with supplement.Ciesielski, P.F., 1983. The Neogene and Quaternary diatom biostratigraphy of
subantarctic sediments, Deep Sea Drilling project Leg 71. In: Ludwig, W.J.,Krasheninnikov, V.A., et al. (Eds.), Init. Repts. DSDP, 71 (Pt. 2). U.S. Govt. PrintingOffice, Washington, pp. 635–665.
Ciesielski, P.F., 1986. Middle Miocene to Quaternary diatom biostratigraphy of Deep SeaDrilling Project Site 594, Chatham Rise, southwest Pacific. In: Kennett, J.P., von derBorch, C.C., et al. (Eds.), Init. Repts. DSDP, 90. U.S. Govt. Printing Office, Washington,pp. 863–885.
Cody, R.D., Levy, R.H., Harwood, D.M., Sadler, P.M., 2008. Thinking outside the zone:high-resolution quantitative diatom biochronology for the Antarctic Neogene.Palaeogeography, Palaeoclimatology, Palaeoecology 260, 92–121.
Crosta, X., Romero, O., Armand, L.K., Pichon, J.-J., 2005. The biogeography of majordiatom taxa in Southern Ocean sediments: 2. Open ocean related species.Palaeogeography, Palaeoclimatology, Palaeoecology 223, 66–92.
biostratigraphy of nearshore Antarctica from the AND-1B drillcore,2010.04.004

16 D. Winter et al. / Global and Planetary Change xxx (2010) xxx–xxx
ARTICLE IN PRESS
Cunningham, W.L., Leventer, A., 1998. Diatom assemblages in surface sediments of theRoss Sea: relationship to present oceanographic conditions. Antarctic Science 10 (2),134–146.
Davey, F.J., Brancolini, G., Hamilton, R.J., Henrys, S.A., Sorlien, C.C., Bartek, L.R., 2000. Arevised correlation of the seismic stratigraphy at the Cape Roberts drill sites withthe seismic stratigraphy of the Victoria Land Basin, Antarctica. In Studies from theCape Roberts Project, Ross Sea, Antarctica; scientific results of CRP-2/2A; Part I,Geophysics and physical properties studies for CRP-2/2A. Terra Antartica 7 (3),215–220.
Dunbar, G., Niessen, F., Vogel, S., Tulaczyk, S., Mandernack, K., Krissek, L., Carter, L.,Cowen, E., Wilch, T., Peng, C., Strong, C.P., Scherer, R., Sjunneskog, C., Winter, D.,McKay, R., Talarico, F., Pompilio, M., the ANDRILL-MIS Science Team, 2007. LatePleistocene to Holocene Strata from Soft-Sediment Coring at the AND-1B Site,ANDRILL McMurdo Ice Shelf Project, Antarctica. Terra Antartica 14 (2), 141–154.
Falconer, T., Pyne, A., Levy, R., Olney, M., Curren, M., the ANDRILL-MIS Science Team,2007. Operations overview for the ANDRILL McMurdo Ice Shelf Project, Antarctica.Terra Antartica 14 (3), 131–140.
Francis, J.E., Hill, R.S., 1996. Fossil plants from the Pliocene Sirius Group, TransantarcticMountains: evidence for climate from growth rings and fossil leaves. Palaios 11,389–396.
Gersonde, R., Burckle, L.H., 1990. Neogene diatom biostratigraphy of ODP Leg 113,Weddell Sea (Antarctic Ocean). In: Barker, P.F., Kennett, J.P., et al. (Eds.), Proc. ODP,Sci. Results, 113. Ocean Drilling Program, College Station, TX, pp. 761–789.
Harwood, D.M., 1986a. Diatom biostratigraphy, paleoecology, and Cenozoic history ofAntarctica. Unpublished Phd thesis, Ohio State University.
Harwood, D.M., 1986a. Recycled siliceous microfossils from the Sirius Formation.Antarctic Journal of the United States 21 (5), 101–103.
Harwood, D.M., Maruyama, T., 1992. Middle Eocene to Pleistocene diatom biostratig-raphy of Southern Ocean sediments from the Kerguelen Plateau. In: Wise Jr., S.W.,Schlich, R., et al. (Eds.), Proc. ODP: Sci. Results. Ocean Drilling Program, CollegeStation, TX, pp. 683–733. 120.
Harwood, D.M., Florindo, F., Talarico, F., Levy, R.H. (Eds.), 2008–09. Studies from theANDRILL, Southern McMurdo Sound Project, Antarctica: Terra Antartica, 15 (1).233 pp.
Hayes, D.E., Frakes, L.A., et al., 1975. Initial Reports of the Deep Sea Drilling Project,Volume 28. U.S. Government Printing Office, Washington, p. 1017.
Hill, R.S., Harwood, D.M., Webb, P.N., 1996. Nothofagus beardmorensis (Nothofagaceae),a new species based on leaves from the Pliocene Sirius Group, TransantarcticMountains. Antarctica. Review of Palaeobotany and Palynology 94, 11–24.
Horgan, H., Naish, T., Bannister, S., Balfour, N., Wilson, G., 2005. Seismic stratigraphy ofthe Plio-Pleistocene Ross Island flexural moat-fill: a prognosis for ANDRILL Programdrilling beneath McMurdo-Ross Ice Shelf. Global and Planetary Change 45, 83–97.
Iwai, M., Winter, D., 2002. Data report: Taxonomic notes of Neogene diatoms from thewestern Antarctic peninsula: Ocean Drilling Program Leg 178. In: Barker, P.F.,Camerlenghi, A., Acton, G.D., Ramsay, A.T.S. (Eds.), Proc. ODP, Sci. Results, 178, pp.1–57. http://www-odp.tamu.edu/publications/178_SR/VOLUME/CHAPTERS/SR178_35.PDF.
Johansen, J.R., Fryxell, G.A., 1985. The genus Thalassiosira (Bacillariophyceae): studieson species occurring south of the Antarctic Convergence Zone. Phycologia 24,155–179.
Jonkers, H.A., Kelley, S.P., 1998. A reassessment of the age of the Cockburn IslandFormation, northern Antarctic Peninsula, and its palaeoclimatic implications.Journal of the Geological Society (London) 155, 737–740.
Kellogg, T.B., Truesdale, R.S., 1979. Ross Sea Diatoms: modern assemblage distributionsand their relationship to ecologic, oceanographic and sedimentary conditions.Marine Micropaleontology 4, 13–31.
Krissek, L., Browne, G., Carter, L., Cowan, E., Dunbar, G., McKay, R., Naish, T., Powell, R.,Reed, J., Wilch, T., The ANDRILL-MIS Science Team, 2007. Sedimentology andStratigraphy of the AND-1B Core, ANDRILL McMurdo Ice Shelf Project, Antarctica.Terra Antartica 14 (3), 185–222.
Mahood, A.D., Barron, J.A., 1996. Late Pliocene diatoms in a diatomite from Prydz Bay.East Antarctica Micropaleontology 42 (3), 285–302.
McArthur, J.M., Rio, D., Massari, F., Castradori, D., Bailey, T.R., Thirlwall, M., Houghton, S.,2006. A revised Pliocene record for marine-87Sr/86Sr used to date an interglacialevent recorded in the Cockburn Island Formation, Antarctic Peninsula. Palaeogeo-graphy, Palaeoclimatology, Palaeoecology 242, 126–136.
McCollum,D.W., 1975.Diatomstratigraphyof theSouthernOcean. In:Hayes,D.E., Frakes, L.A.,et al. (Eds.), Init. Repts. DSDP, 28. U.S. Govt. Printing Office, Washington, pp. 515–571.
McGinnis, L.D., 1981. Dry Valley Drilling Project. Antarctic Research Series, 33.American Geophysical Union, Washington, DC. 465 pp.
Naish, T., Powell, R., Levy, R., Florindo, F., Harwood, D.M., Kuhn, G., Niessen, F., Talarico, F.,Wilson, G., 2007. A record of Antarctic climate and ice sheet history recovered. Eos,Transactions, American Geophysical Union 88 (50), 557–558.
Naish, T., Powell, R., Levy, R., Krissek, L., Niessen, F., Pompilio, M., Scherer, R., Talarico, F.,Wilson, G., Wilson, T., McKay, R., Ross, J., Winter, D., Barrett, P., Browne, G., Carter, L.,Cody, R., Cowan, E., Crampton, J., DeConto, R., Dunbar, G., Dunbar, N., Florindo, F.,Gebhardt, C., Graham, I., Hannah,M., Harwood, D., Hansaraj, D., Henrys, S., Helling, D.,Kuhn, G., Kyle, P., Läufer, A., Maffioli, P., Magens, D., Mandernack, K., McIntosh, W.,Millan, C., Morin, R., Ohneiser, C., Paulsen, T., Persico, D., Pollard, D., Reed, J., Raine, I.,Schmitt, D., Sagnotti, L., Sjunneskog, C., Strong, P., Taviani, M., Vogel, S., Wilch, T.,Williams, T., 2009. Late CenozoicAntarctic Ice SheetOscillations. Nature 458, 322–329.
Please cite this article as: Winter, D., et al., Pliocene–Pleistocene diatomMcMurdo Sound, Glob. Planet. Change (2010), doi:10.1016/j.gloplacha.
O'Brien, P.E., Cooper, A.K., Richter, C., et al. (Eds.), 2001. Proc. ODP, Init. Repts., 188.Ocean Drilling Program, College Station, TX10.2973/odp.proc.ir.188.2001.
Ogg, J.G. and Smith, A.G., 2004. The geomagnetic polarity time scale. In; Gradstein, F.M.,Ogg, J.G. & Smith, AG (Eds.) A Geological Time Scale 2004. Cambridge UniversityPress, Cambridge UK, pp. 63–95.
Olney, M.P., Bohaty, S.M., Harwood, D.M., Scherer, R.P., 2009. Creania lacyae gen. nov. etsp. nov. and Synedropsis cheethamii sp. nov.: fossil indicators of Antarctic sea ice?Diatom Research 24 (2), 357–375.
Pichon, J.J., Labracherie, M., Labeyrie, L.D., Duprat, J., 1987. Transfer functions betweendiatom assemblages and surface hydrology in the Southern Ocean. Palaeogeo-graphy, Palaeoclimatology, Palaeoecology 61, 79–95.
Pichon, J.-J., Labeyrie, L.D., Bareille, G., Labracherie, M., Duprat, J., Jouzel, J., 1992. Surfacewater temperature changes in the high latitudes of the Southern Hemisphere overthe last glacial–interglacial cycle. Paleoceanography 7 (3), 289–318.
Powell, R.D., 1981. Sedimentation conditions in Taylor Valley inferred from texturalanalyses of DVDP cores. In: McGinnis, L.D. (Ed.), Dry Valley Drilling Project,Antarctic Research Ser., vol. 81. AGU, Washington, D.C, pp. 331–350.
Schrader, H.-J., 1976. Cenozoic planktonic diatom biostratigraphy of the SouthernPacific Ocean. In: Hollister, C.D., Craddock, C., et al. (Eds.), Init. Repts. DSDP, 35. U.S.Govt. Printing Office, Washington, pp. 605–672.
Scherer, R.P., 1991. Quaternary and Tertiary microfossils from beneath Ice Stream B;evidence for a dynamic West Antarctic ice sheet history. Global and PlanetaryChange 4 (4), 395–412.
Scherer, R., Hannah, M., Maffioli, P., Persico, D., Sjunneskog, C., Strong, C.P., Taviani, M.,Winter, D., Science Team, A.N.D.R.I.L.L.-M.I.S., 2007. Palaeontologic characterisationand analysis of the AND-1B Core, ANDRILL McMurdo Ice Shelf Project, Antarctica.Terra Antartica 14 (2), 223–254.
Scherer, R.P., Bohaty, S.M., Dunbar, R.B., Esper, O., Flores, J.-A., Gersonde, R., Harwood, D.M.,Roberts, A.P., Taviani, M., 2008. Antarctic records of precession-paced insolation-driven warming during early Pleistocene Marine Isotope Stage 31. GeophysicalResearch Letters 35, L0350510.1029/2007GL032254.
Weaver, F.M., Gombos, A.M., 1981. Southern high-latitude diatom biostratigraphy. In:Warme, T.E., Douglas, R.G., Winterer, E.L. (Eds.), The Deep Sea Drilling Project: ADecade of Progress: Spec. Publ. Soc. Econ. Paleontol. Mineral., 32, pp. 445–470.
Webb, P.-N., Harwood, D.M., 1993. Pliocene Nothofagus (southern beech) fromAntarctica: phytogeography, dispersal strategies, and survival in high latitudeglacial deglacial environments. In: Alden, J., Mastrantonio, J.C., Odum, S. (Eds.),Forest Development in Cold Climates. Plenum, New York, pp. 135–165.
Webb, P.-N., Harwood, D.M., Mabin, M.G.C., McKelvey, B.C., 1996. Amarine and terrestrialSirius Group succession, middle Beardmore Glacier-Queen Alexandra Range,Transantarctic Mountains, Antarctica. Marine Micropaleontology 27, 273–297.
Whitehead, J.M., McKelvey, B.C., 2001. The stratigraphy of the Pliocene–lowerPleistocene Bardin Bluffs Formation, Amery Oasis, northern Prince CharlesMountains, Antarctica. Antarctic Science 13, 79–86.
Whitehead, J.M., Quilty, P.G., Harwood, D.M., McMinn, A., 2001. Early Pliocenepaleoenvironment of the Sørsdal Formation, Vestfold Hills, based on diatom data.Marine Micropaleontology 41, 125–152.
Whitehead, J.M., Bohaty, S.M., 2003. Data report: Quaternary–Pliocene diatombiostratigraphy of ODP Sites 1165 and 1166, Cooperation Sea and Prydz Bay. In:Cooper, A.K., O'Brien, P.E., Richter, C. (Eds.), Proc. ODP, Sci. Results, 188, pp. 1–25.[Online]. Available from World Wide Web: http://www-odp.tamu.edu/publications/188_SR/VOLUME/CHAPTERS/008.PDF.
Whitehead, J.M., Harwood, D.M., McKelvey, B.C., Hambrey, M.J., McMinn, A., 2004.Diatom biostratigraphy of the Cenozoic glaciomarine Pagodroma Group, northernPrince Charles Mountains, East Antarctica. Australian Journal of Earth Sciences 51,521–547.
Wilson, G., Levy, R., Browne, G., Cody, R., Dunbar, N., Florindo, F., Henrys, S., Graham, I.,McIntosh, W., McKay, R., Naish, T., Ohneiser, C., Powell, R., Ross, J., Sagnotti, L.,Scherer, R., Sjunneskog, C., Strong, C.P., Taviani, M., Winter, D., the ANDRILL-MISScience Team, 2007. Preliminary Integrated Chronostratigraphy of the AND-1BCore, ANDRILL McMurdo Ice Shelf Project, Antarctica. Terra Antarctica 14 (2),297–316.
Winter, D.M., 1995. Upper neogene diatom biostratigraphy from coastal drillcores insouthern Victoria Land, Antarctica. University of Nebraska-Lincoln, M.S. thesis.
Winter, D.M., Harwood, D.M., 1997. Integrated diatom biostratigraphy of late Neogenedrillholes in southernVictoria Landand correlation to SouthernOcean records. In: Ricci,Carlo Alberto (Ed.), The Antarctic Region; Geological Evolution and Processes;Proceedings of the VII international symposium on Antarctic earth sciences,International Symposium on Antarctic Earth Sciences, Vol. 7. Terra AntarcticaPublication, Sienna, Italy, pp. 985–992.
Winter, D., Iwai, M., 2002. Data report: Neogene diatom biostratigraphy, AntarcticPeninsula Pacific margin, ODP Leg 178 rise sites. In: Barker, P.F., Camerlenghi, A.,Acton, G.D., Ramsay, A.T.S. (Eds.), Proc. ODP, Sci. Results, 178, pp. 1–25. Online].Available from World Wide Web: bhttp://www-odp.tamu.edu/publications/178_SR/VOLUME/CHAPTERS/SR178_29.PDF.
Zielinski, U., Gersonde, R., 1997. Diatom distribution in Southern Ocean surfacesediments (Atlantic sector): implications for paleoenvironmental reconstructions.Palaeogeography, Palaeoclimatology, Palaeoecology 129, 213–250.
Zielinski, U., Gersonde, R., 2002. Plio-Pleistocene diatom biostratigraphy from ODP Leg177, Atlantic sector of the Southern Ocean. Marine Micropaleontology 45, 225–268.
biostratigraphy of nearshore Antarctica from the AND-1B drillcore,2010.04.004
Copyright © 2022 FDOKUMEN




















