
Visan miospore biostratigraphy and correlation of the Poti Formation (Parnaba Basin, northern Brazil
19
Review of Palaeobotany and Palynology 112 (2000) 147–165 www.elsevier.nl/ locate/revpalbo Visan miospore biostratigraphy and correlation of the Poti Formation (Parnaba Basin, northern Brazil ) J.H.G. Melo a, S. Loboziak b a Petrobras/Cenpes/Divex/Sebipe, Cid. Univ., Q.7, I. Fundao, 21949-900 Rio de Janeiro RJ, Brazil b U.S.T.L., UPRESA 8014 du CNRS, Sciences de la Terre, 59655 Villeneuve d’Ascq Cedex, France Received 30 March 2000; received in revised form 26 June 2000; accepted for publication 27 June 2000 Abstract The Poti Formation, which consists mainly of sandstones with minor proportions of carbonaceous shales and other siliciclastic lithologies, represents all the Vise ´an strata thus far recorded in the Parnaı ´ba Basin, northern Brazil. Well-preserved miospores featuring species with both Southern Euramerican and Gondwanan a nities have been recovered from this formation in four well sections. The most characteristic species are listed in this paper, and brief systematic descriptions are presented for the most significant species, along with comments on their biostratigraphy. A new generic combination is proposed: Cordylosporites magnidictyus (Playford and Helby) Loboziak and Melo comb. nov. Comparisons with miospores illustrated from the Grand Erg Occidental, Algerian Sahara, are tentatively proposed. In terms of the Western European Carboniferous palynozonation, miospore assemblages from the Poti Formation are assignable to the Perotrilites tessellatus–Schulzospora campyloptera (TC)–Raistrickia nigra–Triquitrites marginatus (NM ) zonal range. This corresponds to the upper part of the Holkerian and the whole Asbian, which are British regional stages for the lower to middle parts of the upper Vise ´an. The Vise ´an age formerly attributed to biostratigraphic interval XII of Petrobras’ regional palynostratigraphic scheme is therefore confirmed. As already noted in our recent investigations of the Faro Formation in the Amazon Basin and equivalent strata of the Solimo ˜ es Basin, latest Tournaisian and early to middle Vise ´an sections are either absent or barren of characteristic miospores in the Parnaı ´ba Basin as well. © 2000 Elsevier Science B.V. All rights reserved. Keywords: biostratigraphy; Brazil; Miospores; Parnaı ´ba Basin; systematics; Vise ´an 1. Introduction basins of northern Brazil ( Daemon and Contreiras, 1971; Andrade and Daemon, 1974; Daemon, Recently concluded palynological investigations 1974, 1976). of the upper, shalier part of the Faro Formation In the Parnaı ´ba Basin (Fig. 1), interval XII of the Amazon Basin (Melo and Loboziak, 1997; corresponds to the Poti Formation. However, Loboziak et al., 1998) confirmed the occurrence while reviewing the palynostratigraphy of some of Vise ´an miospore assemblages characteristic of Petrobras wells in terms of the Western European biostratigraphic interval XII, an informal biozone miospore zonation as defined in the British Isles defined by Petrobras palynologists in Paleozoic (Higgs et al., 1988a), Loboziak et al. (1992, figs. 2–5; 1993, fig. 5) attributed the Poti Formation to the Spelaeotriletes pretiosus –Raistrickia clavata * Corresponding author. E-mail address: [email protected] (PC ) Zone, which ranges from the late Middle to 0034-6667/00/$ - see front matter © 2000 Elsevier Science B.V. All rights reserved. PII: S0034-6667(00)00043-9
Transcript of Visan miospore biostratigraphy and correlation of the Poti Formation (Parnaba Basin, northern Brazil

Review of Palaeobotany and Palynology 112 (2000) 147–165www.elsevier.nl/locate/revpalbo
Visan miospore biostratigraphy and correlation of the PotiFormation (Parnaba Basin, northern Brazil )
J.H.G. Melo a, S. Loboziak ba Petrobras/Cenpes/Divex/Sebipe, Cid. Univ., Q.7, I. Fundao, 21949-900 Rio de Janeiro RJ, Brazilb U.S.T.L., UPRESA 8014 du CNRS, Sciences de la Terre, 59655 Villeneuve d’Ascq Cedex, France
Received 30 March 2000; received in revised form 26 June 2000; accepted for publication 27 June 2000
Abstract
The Poti Formation, which consists mainly of sandstones with minor proportions of carbonaceous shales andother siliciclastic lithologies, represents all the Visean strata thus far recorded in the Parnaıba Basin, northern Brazil.
Well-preserved miospores featuring species with both Southern Euramerican and Gondwanan affinities have beenrecovered from this formation in four well sections. The most characteristic species are listed in this paper, and briefsystematic descriptions are presented for the most significant species, along with comments on their biostratigraphy. Anew generic combination is proposed: Cordylosporites magnidictyus (Playford and Helby) Loboziak and Melo comb.nov. Comparisons with miospores illustrated from the Grand Erg Occidental, Algerian Sahara, are tentatively proposed.
In terms of the Western European Carboniferous palynozonation, miospore assemblages from the Poti Formationare assignable to the Perotrilites tessellatus–Schulzospora campyloptera (TC )–Raistrickia nigra–Triquitrites marginatus(NM) zonal range. This corresponds to the upper part of the Holkerian and the whole Asbian, which are Britishregional stages for the lower to middle parts of the upper Visean. The Visean age formerly attributed to biostratigraphicinterval XII of Petrobras’ regional palynostratigraphic scheme is therefore confirmed. As already noted in our recentinvestigations of the Faro Formation in the Amazon Basin and equivalent strata of the Solimoes Basin, latestTournaisian and early to middle Visean sections are either absent or barren of characteristic miospores in the ParnaıbaBasin as well. © 2000 Elsevier Science B.V. All rights reserved.
Keywords: biostratigraphy; Brazil; Miospores; Parnaıba Basin; systematics; Visean
1. Introduction basins of northern Brazil (Daemon and Contreiras,1971; Andrade and Daemon, 1974; Daemon,
Recently concluded palynological investigations 1974, 1976).of the upper, shalier part of the Faro Formation In the Parnaıba Basin (Fig. 1), interval XIIof the Amazon Basin (Melo and Loboziak, 1997; corresponds to the Poti Formation. However,Loboziak et al., 1998) confirmed the occurrence while reviewing the palynostratigraphy of someof Visean miospore assemblages characteristic of Petrobras wells in terms of the Western Europeanbiostratigraphic interval XII, an informal biozone miospore zonation as defined in the British Islesdefined by Petrobras palynologists in Paleozoic (Higgs et al., 1988a), Loboziak et al. (1992, figs.
2–5; 1993, fig. 5) attributed the Poti Formation tothe Spelaeotriletes pretiosus–Raistrickia clavata* Corresponding author.
E-mail address: [email protected] (PC) Zone, which ranges from the late Middle to
0034-6667/00/$ - see front matter © 2000 Elsevier Science B.V. All rights reserved.PII: S0034-6667 ( 00 ) 00043-9

148 J.H.G. Melo, S. Loboziak / Review of Palaeobotany and Palynology 112 (2000) 147–165
Fig. 1. Location map of selected wells in the Parnaıba Basin.
the early Late Tournaisian (Higgs et al., 1992). results of our new palynological assessment of thePoti Formation are presented herein.Loboziak et al. (1992, 1993) failed to recognize
the Visean age of the Poti Formation because ofthe absence, in the samples investigated at thattime, of Lycospora pusilla (regarded by them as 2. Geological settingsthe main guide-species for the Visean), and alsobecause they did not take Gondwanan index The term Poti Formation was first used by
Paiva and Miranda (1937) for subsurface strataspecies into consideration.Since then, however, more Petrobras wells in including sandstones, carbonaceous shales and
other siliciclastic lithologies below 219 m in SGMthe Parnaıba Basin have been investigated in thecourse of a comprehensive Petrobras/Universite core-drill n° 125, Teresina city, State of Piauı.
According to Goes and Feijo (1994) the formationdes Sciences et Technologies de Lille (U.S.T.L.)project aiming at the re-evaluation and biostrati- represents the highest subdivision of the Caninde
Group, which comprises mostly Devonian tographic refinement of Devonian and LowerCarboniferous miospore successions in northern Tournaisian rock units of marine origin.
Throughout much of the Parnaıba Basin, the PotiBrazil. In the light of these newly concluded andongoing studies, several previous local identifica- Formation overlies the latest Famennian–
Tournaisian Longa Formation and is unconform-tions and occurrences of Western Europeanmiospore taxa can now be reinterpreted. The main ably overlain by the Westphalian and younger

149J.H.G. Melo, S. Loboziak / Review of Palaeobotany and Palynology 112 (2000) 147–165
Piauı Formation (datings after Melo et al., 1998), Oriximina and Faro Formations, as well as in theSolimoes Basin, within the highly heterogeneouswhich represents the basal subdivision of the Balsas
Group (Goes and Feijo, 1994). The unit, up to Jandiatuba Formation.320 m thick, is extensively represented both in thesubsurface of the basin and along its marginaloutcrop belts. Remainders of two major deposi-
3. Miospore taxonomytional axes trending NW–SE (the Marajo-Parnaıba Lineament, after Goes et al., 1992) and
Of the numerous miospore taxa identified by usNE–SW (the Transbrasiliano Lineament), noticed
in samples from the Poti Formation, the mostin isopach maps, testify to formerly continuous
characteristic are listed below in alphabeticalLower Carboniferous sedimentation beyond the
order. Many of them conform to the originalpresent-day limits of the Parnaıba Basin, possibly
diagnoses and therefore do not require any addi-connecting this region to the Amazon Basin and
tional information. Brief descriptive comments areWestern African basins respectively (Goes, 1995,
given only for the most significant miosporefig. 12).
species. One new combination is proposed, andThe lithofacies characteristics and sedimentary
two species have their specific epithet modified.evolution of the Poti Formation were discussed
Tentative comparisons with taxa illustrated butand successively updated in several previous
only classified in open nomenclature by Lanzonistudies (Lima and Leite, 1978; Caputo, 1984; Della
and Magloire (1969) from the Grand ErgFavera, 1990; Goes, 1995). According to the latest
Occidental, Algerian Sahara, are proposed.review by Goes (1995), the Poti Formation com-prises sandstone, conglomerate and pelite making Apiculiretusispora multiseta (Luber) Butterworthup shelf, nearshore and fluvial deposits arranged and Spinner, 1967 (Plate I, 1)in a prograding succession. This development cul-
Apiculiretusispora semisenta (Playford) Massaminated in more or less steady deltaic-estuarineet al., 1980 (Plate I, 2)sedimentation during the unit’s latest depositional
phases, only occasionally interrupted by brief Aratrisporites saharaensis Loboziak, Clayton andflooding episodes. Tempestite facies are well repre- Owens, 1986 (Plate II, 6)sented in both offshore and shoreface settings. This monolete camerate miospore was firstHigher parts of the formation, of more continental illustrated as ‘Spore monolete zonale n° 2874’ byaspects, are particularly rich in carbonized land Lanzoni and Magloire (1969, pl. III, figs. 11–13).plant megafossils and may also include thin coal In several previous papers (Coquel et al., 1977, pl.seams. 3, 13; Massa et al., 1980, pl. 6, figs. 7, 8; Attar
Goes (1995) interprets the upper Longa et al., 1980, pl. 3, figs. 8, 9; Clayton and Loboziak,Formation and the Poti Formation as parts of the 1985, pl. 34, figs. 19, 20), it was incorrectly assignedsame depositional succession (regressive or progra- to Spelaeotriletes balteatus (Playford) Higgs, 1975,dational coast system), with the more distal lithofa- a species that possesses a similar sculpture of small,cies corresponding to the former unit and the more densely distributed spines but which is trilete.proximal lithofacies to the latter. However, ourpalynological re-evaluations of the Lower Bascaudaspora submarginata (Playford) Higgs,
Clayton and Keegan, 1988 (Plate II, 1)Carboniferous in the Parnaıba Basin (Melo et al.,1998 and the present paper) fail to identify any This species displays a reticulate, subreticulate
to rugulate sculpture on the distal surface, formedlatest Tournaisian–early Visean sections bridgingthe apparent biostratigraphic gap, which, accord- by narrow smooth sinuous muri. It is further
characterized by an undifferentiated cinguluming to our current belief, separates the Longa andPoti Formations. According to Loboziak et al. composed of the expansion or partial fusion of
muri at the equator. Bascaudaspora canipa Owens,(1998) and Melo et al. (1998), similar gaps arealso recorded in the Amazon Basin, between the 1983, described from the Namurian of northern
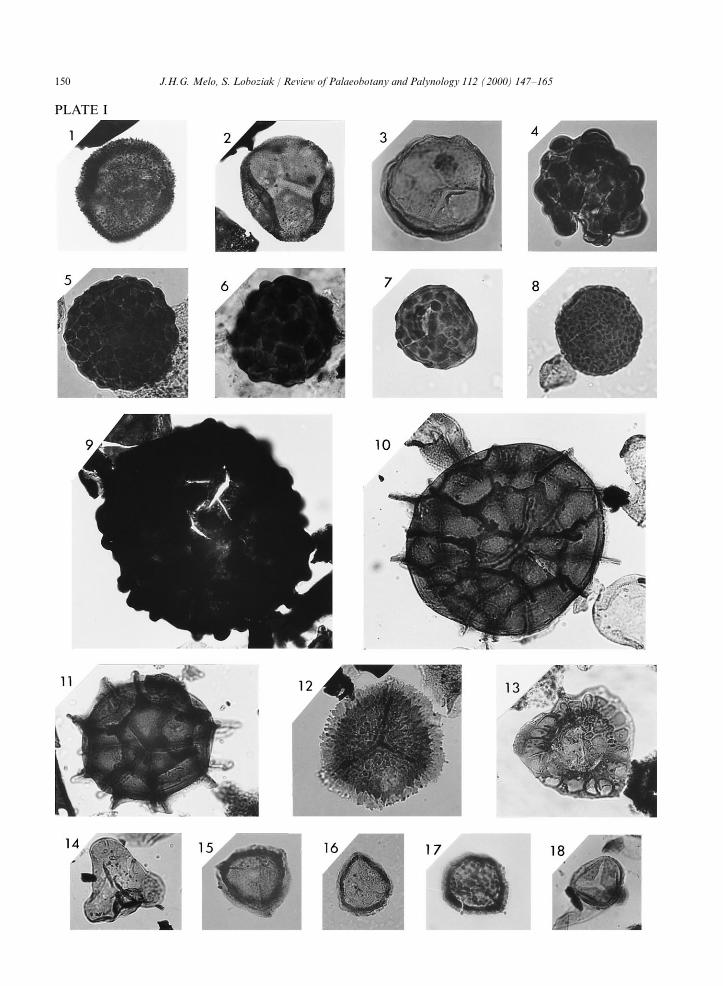
150 J.H.G. Melo, S. Loboziak / Review of Palaeobotany and Palynology 112 (2000) 147–165
PLATE I

151J.H.G. Melo, S. Loboziak / Review of Palaeobotany and Palynology 112 (2000) 147–165
England, seems to be very similar in appearance Convolutispora stigmoidea Bharadwaj andVenkatachala, 1961 (Plate I, 4)to B. submarginata but is distinctly camerate.
Convolutispora sp. (Plate I, 5)Brochotriletes diversifoveatus Playford andIdentical miospores, bearing flat and broadSatterthwait, 1985 (Plate II, 3)
verrucae separated by irregular, sinuous channels,Reticulatisporites sp. n° 2571, figured byhave been illustrated as Verrucosisporites sp. n°Lanzoni and Magloire (1969, pl. II, figs. 4, 5),2904 by Lanzoni and Magloire (1969, pl. IV, figs.probably belongs to this species.3, 4).
Cingulizonates bialatus (Waltz) Smith andCordylosporites magnidictyus (Playford and Helby)Butterworth, 1967 (Plate II, 12)Loboziak and Melo comb. nov. (Plate I, 10, 11)Cingulizonates sp. n° 3282, figured by LanzoniBasionym: Reticulatisporites magnidictyus Playfordand Magloire (1969, pl. III, figs. 1, 2, ? nonand Helby, 1968, p. 110, pl. 10, figs. 7–10.Plate II, 20), is probably assignable to this species.
This azonate, reticulate trilete spore with atypically circular amb was first described from aColatisporites decorus (Bharadwaj and
Venkatachala) Williams in Neves et al., 1973 Carboniferous section in the Hunter Valley, NewSouth Wales, Australia. It bears a coarse reticulum(Plate I, 3)
PLATE ISlides from wells 2-IZ-1-MA, 1-RB-1-MA, 1-CA-1-MA and slide 9507131 from well 2-PM-1-MA are housed in the palynologicalslide collection of the Biostratigraphy and Paleoecology Section of Petrobras Research Centre (Cenpes/Divex/Sebipe), Rio de Janeiro,Brazil. Slides P.B.99.29(2) and P.B.99.31(2) from well 2-PM-1-MA are housed in the palynological collection of the Laboratory ofPaleobotany, University of Lille, France. Miospore locations on slides are based on England Finder graticules. Magnification ofillustrated specimens: ×500.
1. Apiculiretusispora multiseta (Luber) Butterworth and Spinner, 1967. Slide 9405930: E44/3, well 2-IZ-1-MA, core 335/336at 1395/1401 m.
2. Apiculiretusispora semisenta (Playford) Massa et al., 1980. Slide 9405930: V32/3, well 2-IZ-1-MA, core 335/336 at1395/1401 m.
3. Colatisporites decorus (Bharadwaj and Venkatachala) Williams in Neves et al., 1973. Slide 940024 (bis): Z59/3, well 2-IZ-1-MA, core 330 at 1380/1383 m.
4. Convolutispora stigmoidea Bharadwaj and Venkatachala, 1961. Slide 9405929: G34, well 2-IZ-1-MA, core 334/335 at1392/1398 m.
5. Convolutispora sp. Slide 9405929: X28/4, well 2-IZ-1-MA, core 334/335 at 1392/1398 m.6. Verrucosisporites congestus Playford, 1964. Slide Q 1412: K46/4, well 2-IZ-1-MA, core 335 at 1395/1398 m.7. Verrucosisporites gobbettii Playford, 1962. Slide 940026 (bis): 067/2, well 2-IZ-1-MA, core 332 at 1386/1389 m.8. Verrucosisporites depressus Winslow, 1962. Slide 940073: K50/1, well 2-IZ-1-MA, core 334/335 at 1395/1398 m.9. Raistrickia ponderosa Playford, 1964. Slide 9405930: E28/2, well 2-IZ-1-MA, core 335/336 at 1395/1401 m.10, 11. Cordylosporites magnidictyus (Playford and Helby) comb. nov.10. Slide 9405928: B43, well 2-IZ-1-MA, core 334(?)/335 at 1392/1398 m.11. Slide 940069: K50/3, well 2-IZ-1-MA, core 332 at 1386/1389 m.12. Cristatisporites echinatus Playford, 1964. Slide 9405929: R29/3, well 2-IZ-1-MA, core 334/335 at 1392/1398 m.13. Radiizonates arcuatus Loboziak, Playford and Melo, 2000. Slide 9507589: P46, well 1-RB-1-MA, cuttings sample at 623 m.14. Waltzispora polita (Hoffmeister, Staplin and Malloy) Butterworth and Williams, 1967. Slide 940070: U56/2, well 2-IZ-
1-MA, core 333 at 1389/1392 m.15, 16. Lycospora pusilla Butterworth and Williams, 1958.15. Slide 940078 (bis): L38, well 2-IZ-1-MA, core 347/348 at 1429/1435 m.16. Slide 940078: T50/4, well 2-IZ-1-MA, core 347/348 at 1429/1435 m.17. Lycospora noctuina Butterworth and Williams, 1958. Slide 9405930: P31/2, well 2-IZ-1-MA, core 335/336 at 1395/1401 m.18. Rotaspora ergonulii (Agrali) Sullivan and Marshall, 1966. Slide P.B.99.29(2): N42/3, well 2-PM-1-MA, core 29 at
793.4/794.4 m.

152 J.H.G. Melo, S. Loboziak / Review of Palaeobotany and Palynology 112 (2000) 147–165
PLATE II

153J.H.G. Melo, S. Loboziak / Review of Palaeobotany and Palynology 112 (2000) 147–165
consisting of straight to broadly curved muri, from the intersections of muri as seen inCordylosporites spathulatus ( Winslow) Playfordwhich enclose large polygonal lumina. The sides
of the muri taper abruptly and regularly from and Satterthwait 1985, an assignment of R. mag-nidictyus to the genus Cordylosporites seems mostrelatively broad bases to sharp diaphanous edges
and thus have a spine-like profile. The reticulum appropriate.Identical specimens from the Algerian Saharapresents a clear differential development on the
two hemispheres. It is more coarsely meshed proxi- were illustrated as Dictyotriletes sp. n° 2908 byLanzoni and Magloire (1969, pl. II, figs. 2, 3), amally where the muri are radially elongate. On the
distal surface, the muri are more or less of the designation also applied to Syrian forms by Ravnet al. (1994, fig. 2.3, pl. 2.A.2 fig. 2). Additionalsame size and enclose more numerous, more equi-
lateral polygonal lumina. occurrences from North Africa were referred toReticulatisporites cancellatus ( Waltz) Playford,Much like Owens (1983, p. 46), we do not
accept Playford and Helby’s (1968, p. 110) inter- 1962 by Massa et al. (1980, pl. V, fig. 1), Attaret al., 1980 (table I, pl. 2, fig. 1) and Abdesselam-pretation implying that miospores previously
assigned to the genus Reticulatisporites could be Rouighi and Coquel (1997, pl. III, fig. 4), as wellas to Dictyotriletes sp. A by Clayton and Loboziakazonate. For that reason, and also because of the
presence of distinct supramural projections rising (1985, pl. 33, fig. 21) and Dictyotriletes sp. by
PLATE II
1. Bascaudaspora submarginata (Playford) Higgs, Clayton and Keegan, 1988. Slide 940024 (bis): F39, well 2-IZ-1-MA, core 330at 1380/1383 m.
2. Foveosporites appositus Playford, 1971. Slide 940026 (bis): X57, well 2-IZ-1-MA, core 332 at 1389 m.3. Brochotriletes diversifoveatus Playford and Satterthwait, 1985. Slide 9405928: J20/3, well 2-IZ-1-MA, core 334(?)/335 at
1392/1398 m.4. Grandispora maculosa Playford and Helby, 1968. Slide 9405070: M54/3, well 2-IZ-1-MA, core 333 at 1386/1389 m.5. Grandispora spiculifera Playford, 1976. Slide P.B.99.31(2): C24/4, well 2-PM-1-MA, core 31 at 864.4/867.4 m.6. Aratrisporites saharaensis Loboziak, Clayton and Owens, 1986. Slide 940078: L62/3, well 2-IZ-1-MA, core 347/348 at
1429/1435 m.7. Perotrilites tessellatus (Staplin) Neville in Neves et al., 1973. Slide 9405930: D28, well 2-IZ-1-MA, core 335/336 at 1395/1401 m.8. Diatomozonotriletes rarus Playford, 1963. Slide 940068 (bis): J65/4, well 2-IZ-1-MA, core 331 at 1383/1386 m.9. Diatomozonotriletes fragilis Clayton in Neves et al., 1973. Slide Q 1412: R38, well 2-IZ-1-MA, core 335 at 1395/1398 m.10. Densosporites claytonii Ravn, McPhilemy, Rutherford, Talli and Bahra, 1994. Slide 9405928: V20/4, well 2-IZ-1-MA, core
334(?)/335 at 1392/1398 m.11. Densosporites spitsbergensis Playford, 1963. Slide 9405930: L23/4, well 2-IZ-1-MA, core 335/336 at 1395/1401 m.12. Cingulizonates bialatus (Waltz) Smith and Butterworth, 1967. Slide 940024 (bis): M49/2, well 2-IZ-1-MA, core 330 at
1380/1383 m.13. Vallatisporites verrucosus Hacquebard, 1957. Slide 940068: M56/3, well 2-IZ-1-MA, core 331 at 1383/1386 m.14. Vallatisporites ciliaris (Luber) Sullivan, 1964. Slide 940065: W41, well 2-IZ-1-MA, core 323/324 at 1365/1370.7 m.15. Vallatisporites vallatus Hacquebard, 1957. Slide 940070: O55/1, well 2-IZ-1-MA, core 333 at 1389/1392 m.16. Vallatisporites splendens Staplin and Jansonius, 1964. Slide 940073: O52/2, well 2-IZ-1-MA, core 334/335 at 1392/1398 m.17. Vallatisporites agadesensis Loboziak and Alpern, 1978 nom. corr. Slide 940026 (bis): M41/3, well 2-IZ-1-MA, core 332 at
1386/1389 m.18. Vallatisporites banffensis Staplin and Jansonius, 1964. Slide 940026 (bis): R55/2, well 2-IZ-1-MA, core 332 at 1386/1389 m.19. Indotriradites dolianitii (Daemon) Loboziak, Melo and Streel, 1999. Slide 9507131: M54/3, well 2-PM-1-MA, core 31 at
864.4/867.4 m.20. Indotriradites daemonii Loboziak, Melo and Streel, 1999. Slide 9507455: N42/3, well 1-CA-1-MA, cuttings sample at 444 m.21. Indotriradites tedantus (Playford and Satterthwait) Playford, 1991. Slide 9405929: R46, well 2-IZ-1-MA, core 334/335 at
1392/1398 m.22. Spelaeotriletes arenaceus Neves and Owens, 1966. Slide 9507456: E42, well 1-CA-1-MA, cuttings sample at 450 m.23. Spelaeotriletes triangulus Neves and Owens, 1966. Slide 9507455: X63, well 1-CA-1-MA, cuttings sample at 444 m.24. Spelaeotriletes owensii Loboziak and Alpern, 1978 nom. corr. Slide 940065 (1503): B52, well 2-IZ-1-MA, core 323/324 at
1365/1370.7 m.

154 J.H.G. Melo, S. Loboziak / Review of Palaeobotany and Palynology 112 (2000) 147–165
Clayton (1995, pl. I, fig. 10). In more recent publi- Diatomozonotriletes rarus Playford, 1963(Plate II, 8)cations, Brazilian representatives of the species
were also recorded as Dictyotriletes sp. cf. R. Conspecific forms were illustrated asDiatomozonotriletes sp. n° 2911 by Lanzoni andreticulatus by Loboziak et al. (1998, fig. 2, pl. I,
fig. 16) and Melo et al. (1998, fig. 2, pl. 3, fig. 5). Magloire (1969, pl. II, figs. 12, 13).
Foveosporites appositus Playford, 1971 (Plate II, 2)Cristatisporites echinatus Playford, 1964 (Plate I,12)
Grandispora maculosa Playford and Helby, 1968(Plate II, 4)Densosporites anulatus (Loose) Butterworth and
Williams, 1967Grandispora spiculifera Playford, 1976 (Plate II, 5)
Densosporites claytonii Ravn et al., 1994Indotriradites daemonii Loboziak et al., 1999(Plate II, 10)(Plate II, 20)This cavate species, described from the Visean
A trilete, cavate miospore belonging to theof northeastern Syria, presents a cingulum modi-Indotriradites dolianitii Morphon, a distinctivefied by narrow channel-like invaginations, whichgroup of miospore species that appears to bemay break the zona into a series of large verrucategeographically restricted to Gondwanan areas andprojections. These invaginations may extend ontostratigraphically to the Lower Carboniferous. Itthe distal surface as large verrucae, which usuallyhas so far been recorded from Visean strata ofbecome less distinct towards the pole and gradeNorth Africa, the Middle East and Brazil.into an irregular rugulate–foveolate sculpture.
The exoexine is equatorially extended as a zonaRepresentatives of this species were illustratedof more or less uniform width and slightly darkeras Anulatisporites cf. anulatus (Loose) Potonie andaround its inner margin. The distal surface, includ-Kremp, 1954 n° 3241 by Lanzoni and Magloireing that of zona, is sculptured with minute coni(1969, pl. IV, figs. 9, 10). Similar specimens fromand spinae, not wider but longer on or around thethe Agades Coal Basin, Niger were recorded asinner margin of zona, which presents a dense radialDensosporites variomarginatus, Playford 1963 byplication.Loboziak and Alpern (1978, pl. 1, fig. 28). This
Similar specimens were illustrated assame denomination was also used in subsequentRadiizonates sp. n° 2922 by Lanzoni and Magloirepalynological assessments of North Africa and(1969, pl. II, figs. 14, 15), as well as by Ravn et al.adjacent regions (Attar et al., 1980, pl. 2, fig. 6;(1994, pl. 2A.2, fig. 6). They were referred to asClayton and Loboziak, 1985, pl. 34, figs. 8, 14;Radiizonates sp. cf. Hymenozonotriletes dolianitiiLoboziak and Clayton, 1988, pl. 24, fig. 4;Daemon, 1974 by Loboziak et al. (1998, pl. 1,Grignani et al., 1991, pl. 26, figs. 15, 16; Coquelfig. 1, non fig. 2) and Melo et al. (1998, pl. 3, figs.et al., 1995, pl. II, fig. 10; Abdesselam-Rouighi1, 2).and Coquel, 1997, pl. II, fig. 16).
Indotriradites dolianitii (Daemon) Loboziak et al.,Densosporites spitsbergensis Playford, 19631999 (Plate II, 19)(Plate II, 11)
This species differs from I. daemonii by itsThe forms recorded as Densosporites sp. n° 2913thicker, wider-based sculptural elements and theby Lanzoni and Magloire (1969, pl. III, figs. 5, 6)absence or poor development of plication on theprobably belong to this species.zona.
The specimen illustrated as ‘Spore trilete zonaleDiatomozonotriletes fragilis Clayton in Neves et al.,1973 (Plate II, 9) n° 3203’ by Lanzoni and Magloire (1969, pl. III,
fig. 10, ? non fig. 9) probably belongs to thisIdentical specimens were illustrated asDiatomozonotriletes sp. n° 2997 by Lanzoni and species. The form referred to as Radiizonates sp.
cf. Hymenozonotriletes dolianitii Daemon, 1974 byMagloire (1969, pl. III, fig. 20; pl. IV, fig. 1).

155J.H.G. Melo, S. Loboziak / Review of Palaeobotany and Palynology 112 (2000) 147–165
Loboziak et al. (1998, pl. 1, fig. 2, non fig. 1) is This is a large, radial, trilete, cameratemiospore. Several species of Spelaeotriletes havecertainly conspecific with I. dolianitii.been identified in this study. They differ from each
Indotriradites tedantus (Playford and Satterthwait)other in that they have an exinal sculpture on the
Playford, 1991 (Plate II, 21)distal and equatorial surface and that portion of
The specimens illustrated as ‘Spore trilete zonalethe proximal surface adjacent to the curvaturae.
n° 2920’ and ‘n° 2920 forme b’ in Lanzoni andIn S. arenaceus, the sculpture consists of bacula,
Magloire (1969, pl. III, figs. 17–19) resemble thispointed or flat topped coni and variably shaped
species.verrucae, 1–2 mm high and 1–2.5 mm in diameter.
Identical specimens were illustrated asLycospora noctuina Butterworth and Williams,1958 (Plate II, 17) Pustulatisporites cf. pretiosus Playford 1964 n°
2902 by Lanzoni and Magloire (1969, pl. 1, figs.Lycospora pusilla Butterworth and Williams, 1958
18, 19, non fig. 17).(Plate II, 15, 16)
Spelaeotriletes owensii Loboziak and Alpern, 1978Perotrilites tessellatus (Staplin) Neville in Neves(Plate II, 24)et al., 1973 (Plate II, 7)
This species bears a reticulate to vermiculateThis is a cavate Visean zonal species of thesculpture. Muri and rugulae are often surmountedWestern European (British Isles) miospore zona-by small spinae or coni.tion. The exoexine is intensively wrinkled and
Identical specimens were illustrated as ?folded. The incised equatorial margin has a lobateHymenozonotriletes sp. n° 2900 by Lanzoni andappearance due to these infoldings.Magloire (1969, pl. II, figs. 8, 9).
Radiizonates arcuatus Loboziak et al., 2000Spelaeotriletes triangulus Neves and Owens, 1966(Plate I, 13)(Plate II, 23)This is a radially symmetrical, trilete zonate
The sculpture of this species consists of discrete,species recently established for Earlylarge, subcircular, low verrucae, up to 4 mm highCarboniferous forms from Western Gondwanaand up to 8 mm in basal diameter. These verrucaehitherto mistakenly ascribed to Radiizonates genui-may be mammillate, i.e. surmounted by smallnus (Jushko) Loboziak and Alpern, 1978. Theapiculate elements.zona is bipartite, consisting of an inner, thicker
Conspecific forms were illustrated aspart slightly raised above the central proximal areaPustulatisporites cf. pretiosus Playford 1964 n°and an outer, broader, membranous part. Curved2902 by Lanzoni and Magloire (1969, pl. 1, fig. 17,ribs radiate from the inner part to the outer partnon figs. 18, 19). According to Loboziak andof the zona and interconnect to form arches almostClayton (1988), Spelaeotriletes ybertii (Marques-at the equatorial margin.Toigo) Playford and Powis, 1979 is a probableRadiizonates sp. n° 3212, illustrated by Lanzonisenior synonym of S. triangulus.and Magloire (1969, pl. II, fig. 16, ? non fig. 17),
seems to correspond to this species.Vallatisporites agadesensis Loboziak and Alpern,1978 (Plate II, 17)Raistrickia ponderosa Playford, 1964 (Plate I, 9)
Specimens illustrated as Verrucosisporites sp. n° This is a radial, trilete, camerate miospore,which presents a vacuolate zona with a cuniculus2905 by Lanzoni and Magloire (1969, pl. I, figs.7,
8) conform closely to this species’ description and adjacent to the equatorial margin of intexine.Several species belonging to the same genus havecould be attributed to it.been identified. They differ from each other in the
Rotaspora ergonulii (Agrali) Sullivan andsculpture of the distal surface and the proximal
Marshall, 1966 (Plate I, 18)region of the equatorial margin. The sculpture ofV. agadesensis is vermiculate to crudely reticulate,Spelaeotriletes arenaceus Neves and Owens, 1966
(Plate II, 22) being formed by vermiculate sinuous ridges and

156 J.H.G. Melo, S. Loboziak / Review of Palaeobotany and Palynology 112 (2000) 147–165
irregular verrucae and coni, which may be fused between the years 1958 and 1966 in the western,central and northern parts of the Parnaıba Basinat their bases.
Identical specimens are illustrated as (Fig. 1).The dominant fraction of the organic matter isVallatisporites n° 3324 by Lanzoni and Magloire
(1969, pl. III, figs. 7, 8). of continental character. Miospores are present inmost of the studied samples. They are well pre-
Vallatisporites banffensis Staplin and Jansonius,served and are often yellow. However their distri-
1964 (Plate II, 18)bution is uneven in terms of both relative
Conspecific forms were illustrated asabundance and diversity. A large proportion of
Vallatisporites cf. banffensis n° 2891 by Lanzonithe Poti Formation miospore assemblages is com-
and Magloire (1969, pl. V, figs. 15, 16).posed of morphologically simple forms with verypoor biostratigraphic resolution. IdentifiableVallatisporites ciliaris (Luber) Sullivan, 1964
(Plate II, 14) miospores with stratigraphic significance are rela-tively less numerous. These fall into two major
Vallatisporites splendens Staplin and Jansonius,biogeographic categories: those with a Southern
1964 (Plate II, 16)Euramerican affinity, the ranges of which can bedirectly calibrated in terms of CarboniferousVallatisporites vallatus Hacquebard, 1957
(Plate II, 15) miospore successions from Britain (Neves et al.,1972, 1973; Clayton et al., 1978), and those with
Vallatisporites verrucosus Hacquebard, 1957a Gondwanan affinity, comprising only endemic
(Plate II, 13)forms with regional biostratigraphic usefulness.
Figs. 2–5 give the vertical distribution of theVerrucosisporites congestus Playford, 1964(Plate I, 6) main characteristic miospore species identified in
productive samples from the Poti Formation inThe specimens illustrated as Verrucosisporitescf. congestus n° 2887 by Lanzoni and Magloire the four wells investigated.(1969, pl. I, figs. 11, 12) probably belong tothis species. 4.1. Well 1-CA-1-MA (Fig. 2)
Verrucosisporites depressus Winslow, 1962This well is located at 07° 32∞ 30◊S and 45° 56∞
(Plate I, 8)57◊W. The sampled section between 408 and 645 mrepresents most of the Poti Formation (397–Verrucosisporites gobbettii Playford, 1962 (Plate I,
7) 655 m). In most of the 25 productive cuttingssamples investigated, miospores are abundant,The specimens illustrated as Verrucosisporites
sp. n° 2889 by Lanzoni and Magloire (1969, pl. I, diverse and relatively well preserved. Most of thembelong to smooth (Retusotriletes spp.) or apiculatefigs. 13, 14) can be attributed to this species.(Apiculiretusispora spp.) retusoid forms, to verru-
Verrucosisporites nitidus Playford, 1964.cate (Verrusosisporites spp.) and convolute-sculp-tured (Convolutispora spp.) species, as well as to aWaltzispora polita (Hoffmeister, Staplin and
Malloy) Butterworth and Williams, 1967 (Plate I, variety of genera of the densospore group(Cristatisporites, Densosporites, Radiizonates and14)Vallatisporites).
There are also erratic or more regular occur-rences of several indisputable Visean zonal or4. Miospore stratigraphic distribution in the Poti
Formation characteristic taxa. These include forms sharedwith Western Europe (Lycospora pusilla,Perotrilites tessellatus, Vallatisporites ciliaris, someThe results presented herein are based on
detailed palynological investigations of core representratives of the genus Waltzispora sensustricto) and Gondwanan species such assamples and cuttings from four wells drilled

157J.H.G. Melo, S. Loboziak / Review of Palaeobotany and Palynology 112 (2000) 147–165
Fig
.2.
Dis
trib
utio
nch
art
ofse
lect
edm
iosp
ore
taxa
inw
ell
1-C
A-1
-MA
.

158 J.H.G. Melo, S. Loboziak / Review of Palaeobotany and Palynology 112 (2000) 147–165
Fig
.3.
Dis
trib
utio
nch
art
ofse
lect
edm
iosp
ore
taxa
inw
ell
2-IZ
-1-M
A.

159J.H.G. Melo, S. Loboziak / Review of Palaeobotany and Palynology 112 (2000) 147–165
Fig
.4.
Dis
trib
utio
nch
art
ofse
lect
edm
iosp
ore
taxa
inw
ell
2-P
M-1
-MA
.

160 J.H.G. Melo, S. Loboziak / Review of Palaeobotany and Palynology 112 (2000) 147–165
Fig. 5. Distribution chart of selected miospore taxa in well 1-RB-1-MA.
Cordylosporites magnidictyus, Indotriradites dae- half of the Poti Formation, the top of which liesat 1283 m.monii and I. dolianitii [all commonly present in
Visean strata of northern Brazilian basins, The palynoflora is apparently homogeneousthroughout the investigated interval. In most ofalthough some also occur occasionally in late
Tournaisian strata (Loboziak et al., 1999)]. The the nine productive core samples, miospores areabundant, diverse and well preserved. A highpresence of Brochotriletes diversifoveatus,
Cingulizonates bialatus, Densosporites claytonii, percentage is of simple morphology and devoid ofstratigraphic significance. Particularly numerousFoveosporites appositus, Spelaeotriletes arenaceus,
S. owensii and S. triangulus contributes to the are verrucate forms of the genus Verrusosisporitesas well as representatives of the densospore group,Visean aspect of the palynoflora. These species are
joined by holdovers with both Western European mainly belonging to the genera Indotriradites andRadiizonates.and Gondwanan affinities, such as Aratrisporites
saharaensis, Grandispora spiculifera, Radiizonates Visean guide-taxa of the Western Europeanbiozonation (Lycospora pusilla, Perotrilites tessella-arcuatus, Vallatisporites verrucosus, V. vallatus and
Verrucosisporites nitidus, all known to first occur tus, Diatomozonotriletes fragilis, representatives ofthe genus Waltzispora sensu stricto) and speciesin the latest Devonian/earliest Carboniferous
interval, and Colatisporites decorus, which appears with Gondwanan affinities (Cordylosporites mag-nidictyus, Indotriradites daemonii and I. dolianitii)much later in the late Middle/early Late
Tournaisian. are less abundant but nevertheless regularly pre-sent. They occur together with Brochotriletesdiversifoveatus, Cingulizonates bialatus,4.2. Well 2-IZ-1-MA (Fig. 3)Densosporites claytonii, Foveosporites appositus,Indotriradites tedantus, Spealeotriletes arenaceus,This well is located at 59° 51∞ 19◊ S and 47° 29∞
46◊W. Miospore analyses are from the cored S. owensii, S. triangulus and Vallatisporites agade-sensis. Of the species that persist in the Potiinterval 347/348 at 1429/1434 m, at the base of the
Poti Formation, up to core 323/324 at Formation palynoflora and that have their earliestoccurrence in pre-Visean rocks, the most common1365/1368 m. They therefore concern the lower

161J.H.G. Melo, S. Loboziak / Review of Palaeobotany and Palynology 112 (2000) 147–165
are Aratrisporites saharaensis, Bascaudaspora tus, Vallatisporites ciliaris (only in core 34) andRotaspora ergonilii (only in core 30–31). Alsosubmarginata, Colatisporites decorus, Grandisporarelevant is the occurrence of the followingspiculifera, Radiizonates arcuatus, VallatisporitesGondwanan species: Cordylosporites magnidictyus,banffensis, V. splendens, V. vallatus and V.Indotriradites daemonii and I. dolianitii. The Viseanverrucosus.character of the Poti Formation palynoflora isfurther enhanced by the presence of Densosporites4.3. Well 2-PM-1-MA (Fig. 4)claytonii, Diatomozonotriletes rarus andIndotriradites tedantus (only in core 34),This well is located at 03° 37∞ 00◊ S andFoveosporites appositus (present in both core45° 23∞25◊ W. Here, the Poti Formation is 195 msamples), and Brochotriletes diversifoveatus,thick, ranging from 855 to 1050 m. The investi-Cingulizonates bialatus, Spelaeotriletes owensii andgated samples are core 34 at 1034.4/1037.4 m andVallatisporites agadesensis (only in core 30–31).core 30–31 at 862.4/867.4 m; they represent the
base and the upper part of the formation,4.4. Well 1-RB-1-MA (Fig. 5)respectively.
Some biostratigraphic results from the PotiThis well is located at 08° 02∞ 30◊ S andFormation in this borehole have already been
46° 05∞01◊ W. The miospore investigations onlypresented in a previous paper (Loboziak et al.,concern four cuttings samples located between 5901992). In the samples studied at that time, becauseand 581 m within the lowermost part of the Potiof the presence of several Tournaisian miosporesFormation. The unit is 274 m thick in this well,diagnostic of Higgs et al.’s (1988a) zonal schemeranging from 344 to 618 m.for the uppermost Devonian and Lower
Miospores are numerous in all preparations,Carboniferous of Western Europe, and becausebut most of them are morphologically simple andVisean datings in earlier phases of our northerndevoid of biostratigraphic significance. VerrucateBrazilian studies relied mainly on identificationsforms of the genus Verrucosisporites are also fre-
of erratic specimens of Lycospora spp., a Middle quent as well as representatives of the densosporeto Late Tournaisian age had been mistakenly group (Cristatisporites spp., Indotriradites spp.,attributed to the Poti Formation (Loboziak et al., Radiizonates spp.).1992, p. 216, fig. 2). The Visean age of the studied section is sup-
Based on the investigation of additional samples ported by the identification of zonal species of theand the examination of supplementary slides, more Western European palynozonation (Perotrilites tes-accurate miospore results were obtained, which no sellatus) and species with Gondwanan affinitieslonger support the Tournaisian dating of the Poti (Cordylosporites magnidictyus), along with theFormation. The new analyses do confirm the pres- presence (in samples 590–584 m) of Foveosporitesence of some taxa recognized in our previous appositus. The most common holdovers identifiedstudy, such as Aratrisporites saharaensis and in this Visean palynoflora are Bascaudaspora subm-Radiizonates arcuatus (=R. genuinus in Loboziak arginata, Colatisporites decorus, Grandisporaet al., 1992), both recorded in core 34 only, and spiculifera, and Verucosisporites nitidus.Colatisporites decorus, Grandispora spiculifera, V.vallatus, V. verrucosus and Verrucosisporites niti-dus, all of which also occur in core 30–31. These 5. Biostratigraphic assignment of the Potitaxa, although mainly restricted to latest Devonian Formation (Fig. 6)and Tournaisian sections elsewhere, are oftenfound in Visean strata of some Gondwanan The present results confirm the occurrence,regions. Once again, no Lycospora has been found, throughout the Poti Formation, of Viseanbut now, our Visean dating is based on the identi- miospores with both Southern Euramerican andfication of other guide-species of the Western Gondwanan affinities. Among them is Perotrilites
tessellatus, an age-diagnostic species of the BritishEuropean palynozonation, viz. Perotrilites tessella-

162 J.H.G. Melo, S. Loboziak / Review of Palaeobotany and Palynology 112 (2000) 147–165
palynozonation. This species is known to occur 6. Comparison with the Faro Formation (AmazonBasin)from the Perotrilites tessellatus–Schulzospora
campyloptera (TC) to the Raistrickia nigra–Triquitrites marginatus (NM) Biozones (Neves The miospore stratigraphy of the Faro
Formation (Amazon Basin’s equivalent of the Potiet al., 1972, fig. 2 and Neves et al., 1973, table 1).According to Clayton (1985, fig. 1) and Higgs Formation, likewise assignable to biostratigraphic
interval XII) was recently reviewed and updatedet al. (1988b, fig. 1), these biozones correspond tothe upper part of the Holkerian (or the Lower by Loboziak et al. (1998), who also provide an
overview of the previous literature on the subject.Asbian according to Riley, 1993, p. 438 and fig. 1)and the whole Asbian, i.e. the British regional By and large, miospore assemblages from the
Faro Formation could be envisaged as slight vari-stages for the lower and middle parts of theUpper Visean. ants of the richer Poti Formation palynofloras.
Some distinctive constituents of the latter, likePerotrilites tessellatus has often been identifiedin the lower part of the Poti Formation (wells Rotaspora ergonulii and Vallatisporites ciliaris, are
still unrecorded in Amazon Basin Visean assem-2-IZ-1-MA, 2-PM-1-MA and 1-RB-1-MA) but isstill present in its highest part (2-PM-1-MA). In blages, which in their turn include such additional
components as Schopfipollenites sp. andcuttings from well 1-CA-1-MA, it has only beenrecorded at 477 m, i.e. within the middle part of Granulatisporites granulatus, both of which are
unknown in the Poti Formation. Nevertheless,the formation. The apparent scarcity of that speciesin this well probably results from regional paleo- miospore assemblages from both formations share
numerous characteristic or index species, whichenvironmental constraints or could just reflectsome sampling artifact. Therefore, a late Visean ensure their correlation and common assigment to
interval XII, e.g. Perotrilites tessellatus,age can be assigned to the whole Poti Formation.This dating is confirmed by the additional presence Spelaeotriletes arenaceus, S. owensii, S. triangulus,
Diatomozonotriletes fragilis, Cirratriradites rarus,of such species as Diatomozonotriletes fragilis,Rotaspora ergonulii and Waltzispora polita (a repre- Foveosporites appositus, Cordylosporites magnidic-
tyus, Vallatisporites agadesensis, components of thesentative of the genus Waltzispora sensu stricto,which first occurs in the late Visean—see W. plani- Indotriradites dolianitii Morphon, representatives
of the genera Lycospora and Waltzispora sensuangulata in Neves et al., 1972, fig. 2 and Neveset al., 1973, table I ). stricto, etc. Other common attributes of Visean
palynofloras from the two formations include theOur review also confirms the assignment of thePoti Formation to Daemon’s (1974) Visean occurrence of several pre-Visean holdovers (see
above) and the apparent absence of truly marineReticulatisporites cf. magnidictyus miospore Zone,which corresponds to biostratigraphic interval XII palynomorphs, even though sparse marine shelly
faunas (Edmondia, Nucula, Lingulidiscina; cf.of Petrobras regional palynozonation. Its epony-mous species (Cordylosporites magnidictyus) and Kegel, 1954 and Mesner and Wooldridge, 1964)
and extensive offshore/shoreface lithofacies (Dellacharacteristic species (Indotriradites dolianiti/I. dae-monii) are common throughout the Poti Favera, 1990; Goes, 1995) are reported from the
Poti Formation.Formation, and relatively numerous comparedwith other Visean guide-species. In their palynological revaluation of the Faro
Formation in well 2-LF-1-AM, Loboziak et al.The regular occurrence, in the Poti Formation,of miospore taxa such as Grandispora spiculifera, (1998) proposed a tentative extension of the unit’s
uppermost part into the latest Visean TripartitesVallatisporites verrucosus, V. vallatus, andVerrucosisporites nitidus, characteristic of pre- vetustus–Rotaspora fracta (VF) Zone because of
the supposed disappearance of Perotrilites tessella-Visean sections in Euramerica, may reflect eitherreworking from latest Famennian and/or tus in that highest section. Our current results
from the Parnaıba Basin demonstrate that P. tes-Tournaisian sediments or disparate Gondwananranges with respect to Euramerican occurrences. sellatus persists up to the topmost Poti Formation

163J.H.G. Melo, S. Loboziak / Review of Palaeobotany and Palynology 112 (2000) 147–165
Fig. 6. Miospore range chart and dating of the Poti Formation considering characteristic Visean taxa of the British Isles miosporebiozonation. Chronostratigraphy, biostratigraphy and miospore ranges after Neves et al. (1972, fig. 2); Neves et al. (1973, table I );Clayton in Neves et al. (1973, p. 36); Clayton (1985, table 1) and Higgs et al. (1988b, fig. 3 and p. 65).
in at least well 2-PM-1-MA, thus restricting its Cordylosporites magnidictyus, the eponymousspecies of interval XII (=Reticulatisporites cf. mag-correlation to the TC-NM zonal range only
(Fig. 6). It is presently unclear whether the Faro nidictyus Zone of Daemon, 1974), as well asmiospores of the Indotriradites dolianitii Morphon,Formation could indeed be partly younger than
the Poti Formation or whether the apparent are usually common in the Poti Formation. Theyrepresent the most significant Gondwanan compo-absence of P. tessellatus in the uppermost Faro
Formation could only be ascribed to paleo- nents of interval XII palynofloras, and thereforeconstitute good stratigraphic markers for theecological constraints.Visean in northern Brazilian basins.
The occurrence, throughout the Poti Formation,of Perotrilites tessellatus, an age-diagnostic species7. Conclusionsof the British Isles biozonation, is consistent witha restricted late Holkerian-Asbian age (TC-NMAs recently demonstrated for the Faro
Formation of the Amazon Basin and equivalent zonal range). Hence, a possible extension of theformation into the latest Visean, i.e. into thestrata of the Solimoes Basin (Melo and Loboziak,
1997; Loboziak et al., 1998; Melo et al., 1998), Tripartites vetustus–Rotaspora fracta (VF) Zoneof early Brigantian age, such as tentatively envis-this review confirms the late Visean age previously
attributed by Petrobras palynologists to biostrati- aged by Melo and Loboziak (1997) and Loboziaket al. (1998) for the Faro Formation in well 2-LF-graphic interval XII in northern Brazilian
Paleozoic basins (Daemon and Contreiras, 1971; 1-AM, is now considered most unlikely.Throughout most of the Parnaıba Basin, theDaemon, 1974, 1976; Andrade and Daemon,
1974). Poti Formation rests on the Longa Formation,

164 J.H.G. Melo, S. Loboziak / Review of Palaeobotany and Palynology 112 (2000) 147–165
lycospores dans la serie stratigraphique. Ann. Soc. geol.which apparently lacks any Visean index species.Nord 5, 47–57. 2eme serIn terms of the Western European palynozonation,
Andrade, S.M., Daemon, R.F., 1974. Litoestratigrafia emiospore assemblages from the Longa Formation bioestratigrafia do flanco sudoeste da Bacia do Parnaıbarange from the Retispora lepidophyta– (Devoniano e Carbonfero). In: 28° Congr. Brasil. Geol.,Verrucosisporites nitidus (LN) through the Porto Alegre, Anais, 2, 129–137.
Attar, A., Candilier, A.M., Coquel, R., Fournier, J., 1980.Spelaeotriletes pretiosus–Raistrickia clavata (PC)Etude palynologique du Devonien terminal et du Carboni-spore zones (Melo et al., 1998).fere inferieur du bassin d’Illizi (Fort-Polignac), Algerie.Therefore, our miospore studies of LowerBull. Centres Rech. Expl.-Prod. Elf Aquitaine 17 (1),
Carboniferous strata in the Parnaıba Basin (Melo 79–147.et al., 1998 and the present paper) fail to identify Caputo, M.V., 1984. Stratigraphy Tectonics Paleoclimatology
and Paleogeography of Northern Basins of Brazil. Univer-any palyniferous sections of latest Tournaisian–sity of California, Santa Barbara, CA, 583 pp.early Visean age that could bridge the biostrati-
Clayton, G., Higgs, K., Keegan, J.B., Sevastopulo, G.D., 1978.graphic gap between the Longa and PotiCorrelation of the palynological zonation of the Dinantian
Formations. The former unit spans an age from of the British Isles. Palinologıa, num. extraord. 1, 137–147.latest Famennian to late middle or early late Clayton, G., 1985. Dinantian miospores and inter-continental
correlation. In: C.R. 10e Congr. Internat. Stratigr. Geol.Tournaisian (LN-PC zonal range), whereas theCarbonifere, Madrid, 1983, 4, 9–23.latter is evidently restricted to the late Visean
Clayton, G., Loboziak, S., 1985. Early Carboniferous (early(TC-NM zonal range). Loboziak et al. (1998) andVisean–Serpukhovian) palynomorphs. In: Owens, B.,
Melo et al. (1998) also record similar gaps in the Thusu, B. (Eds.), Palynostratigraphy of north-east Libya,Amazon Basin, between the Oriximina and Faro J. Micropalaeont. 4 (1), 83–101.Formations, as well as in the Solimoes Basin, Clayton, G., 1995. Carboniferous miospore and pollen assem-
blages from the Kingdom of Saudi Arabia. Rev. Palaeobot.within the Jandiatuba Formation. These results doPalynol. 89, 1/2, 115–123.not support Goes’ (1995) interpretation that the
Coquel, R., Loboziak, S., Stampfli, G., Stampfli-Vuille, B.,upper Longa and Poti Formations are parts of one1977. Palynologie du Devonien superieur et du Carbonifere
single depositional succession (regressive or pro- inferieur dans l’Elburz Oriental (Iran Nord-est). Rev.gradational coast system), with the more distal Micropaleont. 20 (2), 59–71.
Coquel, R., Lang, J., Yahaya, M., 1995. Palynologie du Car-lithofacies corresponding to the former unit andbonifere du Nord Niger et de la plate-forme saharienne—the more proximal lithofacies to the latter.implications stratigraphiques et paleogeographiques. Rev.Palaeobot. Palynol. 89, 319–334.
Daemon, R.F., 1974. Palinomorfos-guias do Devoniano Supe-rior e Carbonıfero Inferior das bacias do Amazonas e Par-
Acknowledgements naıba. An. Acad. Brasil. Ci. 46, 3/4, 549–587.Daemon, R.F., 1976. Correlacao bioestratigrafica entre os sedi-
mentos do Siluriano, Devoniano e Carbonıfero Inferior dasThe authors are indebted to Rene Rodriguesbacias do Amazonas, Parnaıba e Parana. In: 29° Congr.and Cecılia Cunha Lana (Biostratigraphy andBrasil. Geol., Ouro Preto, Anais, 2, 189–194.Paleoecology Section of Petrobras Research and
Daemon, R.F., Contreiras, C.J.A., 1971. Zoneamento palino-Development Centre, Cenpes/Divex/Sebipe, Rio logico da Bacia do Amazonas. In: 25° Congr. Brasil. Geol.,de Janeiro, Brazil ) for the constructive criticism Sao Paulo, Anais, 3, 79–88.and review of the manuscript. J.H.G. Melo thanks Della Favera, J.C., 1990. Tempestitos da Bacia do Parnaıba.
Porto Alegre, Univ. Fed. Rio Grande do Sul 2 v. (unpub-Petrobras—Petroleo Brasileiro S.A. for the per-lished D.Sc. thesis).mission to publish this paper.
Goes, A.M.O., Travassos, W.A., Nunes, K.C., 1992. ProjetoParnaıba—reavalicao da bacia e perspectivas exploratorias.Belem, Petrobras Vol. 1, unpublished report.
Goes, A.M.O., Feijo, F.J., 1994. Bacia do Parnaıba. Bol. Geoci.References Petrobras 8 (1), 57–67.
Goes, A.M., 1995. A Formacao Poti (Carbonıfero Inferior) daBacia do Parnaıba. Univ. Sao Paulo, Sao Paulo,Abdesselam-Rouighi, F.F., Coquel, R., 1997. Palynologie duxix+180 pp. (unpublished D.Sc. thesis).Devonien terminal—Carbonifere inferieur dans le sud-est
du bassin d’Illizi (Sahara algerien): position des premieres Grignani, D., Lanzoni, E., Elatrash, H., 1991. Palaeozoic and

165J.H.G. Melo, S. Loboziak / Review of Palaeobotany and Palynology 112 (2000) 147–165
Mesozoic subsurface palynostratigraphy in the Al Kufrah Visean miospore biostratigraphy in the Amazon Basin,northern Brazil. Rev. Palaeobot. Palynol. 104, 143–155.Basin, Libya. In: Salem, M.J., Hammuda, O.S., Eliagoubi,
Loboziak, S., Melo, J.H.G., Playford, G., Streel, M., 1999. TheB.A. (Eds.), The Geology of Libya (Third Symposium onIndotriradites dolianitii Morphon, a distinctive group speciesthe Geology of Libya, Tripoli, 27–30 September, 1987), 4,from the Lower Carboniferous of Gondwana. Rev. Palaeo-1159–1227.bot. Palynol. 107, 17–22.Higgs, K.T., Clayton, G., Keegan, J.B., 1988a. Stratigraphy
Massa, D., Coquel, R., Loboziak, S., Taugourdeau-Lantz, J.,and systematic palynology of the Tournaisian rocks of Ire-1980. Essai de synthese stratigraphique et palynologique duland. Geol. Surv. Ireland Spec. Pap. 7, 1–93.Carbonifere en Libye occidentale. Ann. Soc. geol. NordHiggs, K.T., McPhilemy, B., Keegan, J.B., Clayton, G., 1988b.99, 429–442.New data on palynological boundaries within the Irish
Melo, J.H.G., Loboziak, S., 1997. Visean miospore biostratigra-Dinantian. Rev. Palaeobot. Palynol. 56, 61–68.phy in the Amazon Basin: state-of-the-art. Rev. Univ. Guar-Higgs, K.T., Dreesen, R., Dusar, M., Streel, M., 1992. Palyno-ulhos—Geocienc. 2, 210 no. especial.stratigraphy of the Tournaisian (Hastarian) rocks in the
Melo, J.H.G., Loboziak, S., Streel, M., 1998. Latest DevonianNamur Synclinorium, West Flanders, Belgium. Rev. Palaeo-to early Late Carboniferous biostratigraphy of northernbot. Palynol. 72, 149–158.Brazil: an update. Bull. Centr. Rech. Expl.-Prod. Elf Aqui-Kegel, W., 1954. Lamelibranquios da Formacao Poti (Carbonıf-taine 20 (1), 13–33.ero Inferior) do Piauı. Not. Prelim. Est. D.G.M./D.N.P.M.
Mesner, J.C., Wooldridge, L.C.P., 1964. Maranhao Paleozoic88, 1–14.Basin and Cretaceous coastal basins, North Brazil. Bull.Lanzoni, E., Magloire, L., 1969. Associations palynologiquesAm. Assoc. Petr. Geol. 48 (9), 1475–1512.et leurs applications stratigraphiques dans le Devonien
Neves, R., Gueinn, K.J., Clayton, G., Ioannides, N., Neville,superieur et Carbonifere inferieur du Grand Erg OccidentalR.S.W., 1972. A scheme of miospore zones for the British(Sahara algerien). Rev. Inst. Fr. Petrole 23 (4), 441–465.Dinantian. C.R. 7e Congr. Internat. Statigr. Geol. Carboni-Lima, E.A.M., Leite, J.F., 1978. Projeto estudo global dosfere, Krefeld 1971 (1), 347–353.recursos minerais da Bacia Sedimentar do Parnaıba, integ-
Neves, R., Gueinn, K.J., Clayton, G., Ioannides, N.S., Neville,racao geologico-metalogenetica, relatorio final da etapa III.
R.S.W., Kruszewska, K., 1973. Palynological correlationsRecife, DNPM/CPRM Vol. 16.
within the Lower Carboniferous of Scotland and northernLoboziak, S., Alpern, B., 1978. Le bassin houiller Viseen
England. R. Soc. Edinburgh Trans. 69, 23–76.d’Agades (Niger) III. Les Microspores. Palinologıa, num. Owens, B., 1983. Bascaudaspora gen. nov., a new reticulateextraord. 1, 55–67. miospore genus from the Namurian of Northern England.
Loboziak, S., Clayton, G., 1988. The Carboniferous palyno- Rep. Inst. Geol. Sc. 83 (10), 45–49.stratigraphy of northeast Libya. In: El-Arnauti, A., Owens, Paiva, G., Miranda, J., 1937. Carvao mineral do Piauı. Bol.B., Thusu, B. (Eds.), Subsurface Palynostratigraphy of DNPM/DFPM 20, 1–16.Northeast Libya. Garyounis University/Agoco/CIMP, Ben- Playford, G., Helby, R., 1968. Spores from a Carboniferousghazi, pp. 129–149. section in the Hunter Valley, New South Wales. J. Geol.
Loboziak, S., Streel, M., Caputo, M.V., Melo, J.H.G., 1992. Soc. Australia 15 (1), 103–119.Middle Devonian to Lower Carboniferous miospore stratig- Ravn, R.L., McPhilemy, B., Rutherford, M., Talli, S., Bahra,raphy in the Central Parnaıba Basin, Brazil. Ann. Soc. geol. G., 1994. Late Devonian and Early Carboniferous palyno-Belg. 115 (1), 212–226. stratigraphy and its applications in northeastern Syria. In:
Loboziak, S., Streel, M., Caputo, M.V., Melo, J.H.G., 1993. Simmons, M.D. (Ed.), Micropalaeontology and hydro-Middle Devonian to Lower Carboniferous miospores from carbon exploration in the Middle East. Chapman and Hall,selected boreholes in Amazonas and Parnaıba Basins London, pp. 5–21.(Brazil ): additional data, synthesis and correlation. Doc. Riley, N.J., 1993. Dinantian (Lower Carboniferous) biostratig-Lab. Geol. Lyon 125, 277–289. raphy and chrono-stratigraphy in the British Isles. J. Geol.
Soc. 150 (3), 427–446.Loboziak, S., Melo, J.H.G., Streel, M., 1998. Reassessment of




















