
Vestibular rehabilitation in elderly patients with Parkinson
Upload
independentCategory
view
0download
0
Vestibular neuritis: Clinical-pathologic correlation ROBERT W. BALOH, MD, IVAN LOPEZ, PhD, AKIRA ISHIYAMA, MD, PHILLIP A. WACI(YM, MD,
and VICENTE HONRUBIA, MD, Los Angeles, California
Postmortem examination of the brain and temporal bones of a patient with well-documented vestibular neuritis showed selective neuronal loss in Scarpa's ganglia on the side with absent caloric response. There was loss of hair cells and an "epithelialization" of the utricular macule and semicircular canal cristae on the deafferented side, and synaptic density in the vestibular nuclei on the deafferented side was decreased compared with that on the normal side. All findings were consistent with an isolated viral infection of Scarpa's ganglia. This is the first description of the effects of chronic deafferentation on the vestibular sensory epithelia and the vestibular nuclei in a human being. [OTOLARYNGOL HEAD NECK SUR® 1996;114:586-92.]
Vest ibu la r neuritis is a common clinical syndrome thought to result from a selective viral inflammation of the vestibular nerve. 1 It is characterized by the acute onset of vertigo that gradually resolves over days. There are no associated auditory or neurologic syl~nptoms. The terms vestibular neuronitis, vestibular labyrinthitis, and vestibular neurolabyrinthitis are of- ten used interchangeably with vestibular neuritis. 24 Epidemiologic studies support a viral cause for ves- tibular neuritis, but viruses have rarely been iden- tified in individual cases. 5 Serologic studies can dem- onstrate that a virus infected the patient but cannot prove that the infectious agent caused the disorder. Furthermore, isolation of a virus from the nasophar- ynx or tissue other than the membranous labyrinth does not prove a causal relationship between the virus and the disease. 5 Recently, contrast enhance- ments of the membranous labyrinth and/or the ves- tibular nerve have been identified on magnetic reso- nance imaging scans, but these are nonspecific find- ings consistent with inflammation of any cause. 6
Possibly the most convincing evidence for a viral cause of vestibular neuritis comes from the temporal
From the Departments of Neurology (Dr: Baloh) and Surgery (Head and Neck) (Drs. Baloh, Lopez, Ishiyama, Wackym, and Honrubia), University of California, Los Angeles, School of Medicine.
Supported by National Institutes of Health grants AG 09693 and DC 01404.
Received for publication Aug. 24, 1995; accepted Aug. 25, 1995. Reprint requests: Robert W. Baloh, MD, Department of Neu-
rology, UCLA School of Medicine, Los Angeles, CA 90095- 1769.
Copyright © 1996 by the American Academy of Otolaryngology- Head and Neck Surgery Foundation, Inc.
0194-5998Y96/$5.00 + 0 23/1/68823
586
bone studies of Schuknecht and Kitamura. 7 They identified isolated atrophy of the vestibular nerve or parts of the vestibular nerve in four patients who reported a prolonged attack of vertigo suggestive of vestibular neuritis. In some cases there Was addi- tional atrophy of the vestibular end-organ, and in others the atrophy was confined to the vestibular nerve. The atrophy of the nerve and end-organs was similar to that associated with well-documented vi- ral disorders such as mumps or measles. A limitation of these studies was the retrospectively obtained history. Each patient had multiple episodes of ver- tigo, and some had associated hearing loss or a systemic illness such as diabetes mellitus. Further- more, only the temporal bones were evaluated, so there was no information regarding the proximal vestibular nerve and the brain stem.
We present a clinical-pathologic correlation of a patient with vestibular neuritis who was followed up prospectively from the time of the vertigo episode to the time of death 9 years later. The autopsy was performed within 3 hours of death, and the vestib- ular end-organs, the vestibular nerves, Scarpa's gan- glia, and the brain stem were studied.
PATIENT AND METHODS Patient
A 73-year-old man awoke in the middle of the night with acute vertigo, nausea, and vomiting. He tried to get out of bed to go to the bathroom but fell back down onto the bed because of severe imbal- ance. His wife helped him to the bathroom, and he vomited repeatedly and diarrhea developed. The paramedics were called, and he was taken to a local emergency department and hospitalized because of dehydration caused by recurrent vomiting. Neuro-
by P Ashley Wackym on September 13, 2015oto.sagepub.comDownloaded from

Oto aryngo ogy - Head and Neck Surgery Volume 114 Number 4 BALOH et al. 587
logic examination revealed a spontaneous right- 120 beating nystagmus and severe imbalance. His hear- ing was normal, and there were no other focal neurologic findings. A magnetic resonance imaging 0 scan of the', brain was normal. The severe vertigo, nausea, and vomiting gradually subsided over a pe- riod of 5 days, but it was more than a month before he felt nearly back to normal. He did continue to have occasional brief dizzy spells with quick head movements or rapid position changes for the next several years. Caloric examination during the initial hospitalization and 2 years later documented a com- plete absence of caloric response on the left side. Regular follow-up examinations, including quanti- tative rotational testing on three occasions, docu- mented typical findings of a compensated unilateral ~_ 90 peripheral vestibular lesion (Fig. 1). An audiogram o ~ performed within 3 months of death remained nor- ~ 60 mal for his age. Nine years after the episode of vertigo, the patient died of a massive myocardial 30 infarction. The general autopsy was otherwise un-
0 remarkable, and gross examination of the brain was normal.
Tissue Preparat ion
The temporal bones and brain were removed within 3 hours of death. The eighth nerves and internal auditory arteries were sectioned just out- side the internal auditory canal. The brain was then removed with preservation of the blood vessels at the base of the brain. The temporal bones were removed with a circular bone saw. After 10% buffered formalin was injected into the tympanic cavity, both temporal bones and the brain stem were immersed in 10% paraformaldehyde in so- dium potassium phosphate 0.1 mol/L (pH 7.4) for 48 hours. Subsequently the end-organs and Scar- pa's ganglia from both sides were dissected, post- fixed with a mixture of 4% paraformaldehyde and 0.1 glutaraldehyde in 0.1 mol/L phosphate-buffered saline (PBS), pH 7.4. Scarpa's ganglia were de- hydrated and embedded in paraffin. Longitudinal 10 ~xm thick serial sections were obtained, mounted on Superfrost Plus slides (Fisher Scientific Co., Pittsburgh, Pa.), and counterstained with toluidine blue 0.1% in 0.1 mol/L borate buffer. Selected sections were processed for immunohistochemistry or electron microscopy. The end-organs from each side (the three crista ampullaris and macula utricle) were dehydrated and embedded in resin (Araldite). The brain stem was postfixed in 10% formalin for 2 weeks, dehydrated, and embedded in paraffin (Paraplast).
-120 "6" I1)
i1) -o
o o I1) >
-80 m 0 m
/ ' , 4 I
0 20 40
80
o q \ 1 ,,J
5 10
cw
0
10
ccw
10
T i m e (sec)
0.05 Hz 120°/s
0.4 Hz 60o/s
impulse 120°Is
Fig. I. Horizontal VOR in patient I year before death; plots of slow-phase velocity vs. time for sinusoidal and step changes in angular velocity. Low acceleration sinusoidal responses are symmetric, but the high-acceleration step responses remain asymmetric (decreased with rotation toward the lesion side). There is an increase in low-frequency (0.05 Hz) phase lead (vertical dashed line) and a decrease in the VOR time con- stant (in both directions) compared with normals.
Electron Microscopy
Selected sections of Scarpa's ganglia were depar- affinized with 100% xylol, rehydrated by use of descending alcohols (100%, 70%, 50%, 30%), and placed in PBS. Thereafter the sections were im- mersed in a solution of 1% osmium tetroxide for 30 minutes in PBS and dehydrated for plastic embed- ding. Ultrathin sections (80 to 100 nm) were ob- tained with a diamond knife (Du Pont Co., Wil- mington, Del.). Sections were examined with a Zeiss (10/CR, Oberkochen, Germany) transmission elec- tron microscope.
Immunohistochemistry for Viral Antigens
Sections from Scarpa's ganglion were deparaf- finized, rehydrated with PBS, and sequentially in- cubated with the following solutions: 0.1% of hy- drogen peroxide in PBS for 10 minutes after three washes with PBS for 10 minutes each, blocking solution containing 10% normal horse serum
by P Ashley Wackym on September 13, 2015oto.sagepub.comDownloaded from

588 BALOH ef al.
Otolaryngology - Head and Neck Surgery
April 1996
(NHS), and 0.05% Triton X-100 in PBS for 30 minutes on a rotating shaker. Four mouse antivirus monoclonal antibodies were tested (all from Chemi- con, Temecula, Calif.): antiherpes simplex virus types I and II, antivaricella zoster virus, anticyto- megalovirus (antibody cocktail), and anti-Epstein- Barr virus. Each antibody was diluted 1:5 in PBS containing 1% NHS for 48 hours at 4 ° C, biotinylated secondary antibody for 1 hour at 22 ° C, avidin-biotin- complex (ABC) for 1 hour at 22 ° C (1:50), and 3,3'-diamino-benzidine (DAB; Vector Laborato- ries, Inc., Burlingame, Calif.). At the end of the incubation the tissue was washed with PBS, air- dried, and mounted with Permount. Scarpa's gan- glion sections from a normal subject were processed as described above, but the primary antibody was substituted with NHS.
Immunohistochemistry for Synaptic Density
The brain stem was serially sectioned at 20 txm, and every tenth section was stained with toluidine blue. The boundaries of the four vestibular nuclei were identified according to the criteria of Sadjad- pour and Brodal. 8 The morphology of neurons and the neuronal density on the two sides were qualita- tively compared. Synaptophysin (synaptic) density was assessed by immunostaining paraffin sections with a commercially available antibody to synapto- physin. After the sections were deparaffinized and rehydrated, nonspecific antibody binding was de- creased by incubating the sections with 10% NHS for 30 minutes at room temperature. The primary antibody, diluted 1:50, was applied and allowed to incubate for 24 hours at 4 ° C. Visualization of t h e antigen-antibody complex was achieved with a mouse ABC Elite Kit (Vector Laboratories). The chromagen used was DAB, and hydrogen peroxide was used to oxidize the DAB. After counterstaining with toluidine blue, the synaptophysin density was measured in 10 comparable sections from each side with computer-based microscopy and image-analy- sis software (Image 1.52; National Institutes of Health, Bethesda, Md.).
RESULTS Scarpa's Ganglia and Vestibular Nerves
There was a clear difference in the size and appearance of Scarpa's ganglia on the two sides (Fig. 2). The number of neurons on the left side was markedly decreased, and some of the remaining neurons were darkly staining and shrunken. The neuronal loss was greatest in the inferior division of Scarpa's ganglia, but both divisions were severely
affected. Neurons on both sides exhibited lipofuscin inclusions seen on both light and electron micros- copy. No viral inclusion bodies were seen in the nuclei or cytoplasm of the primary vestibular neu- rons either on routine light microscopy or on elec- tron microscopy. Furthermore, immunohistochemi- cal staining for antigens of herpes simplexvirus I and II, varicella zoster virus, cytomegalovirus, and Ep- stein-Barr virus was negative on both sides. Immu- nohistochemical staining was present in positive controls and absent in the normal control.
Serial cross-sections of the vestibular nerves be- ginning at the inner ear and including the root entry zone at the brain stem showed a marked loss of nerve fibers on only the left side. There was no evidence of prior infarction.
Inner Ear
Of the vestibular end-organs, the most prominent changes were seen in the utricular macule (Fig. 3). There was a marked drop out in hair cells with only a few remaining. Also, compared with the normal side, there were few afferent nerve terminals and a markedly decreased number of nerve fibers at the base. Despite this dramatic loss of hair cells, the sensory epithelium on the left was not atrophic, and there were no signs of fibrosis (Fig. 3). The center of the macule consisted mainly of rectangular cells with tight junctions typical of supporting cells normally seen in the periphery. Similar changes were seen in the semicircular canal cristae on the left, although they were less prominent than in the utricular macule.
Vestibular Nuclei
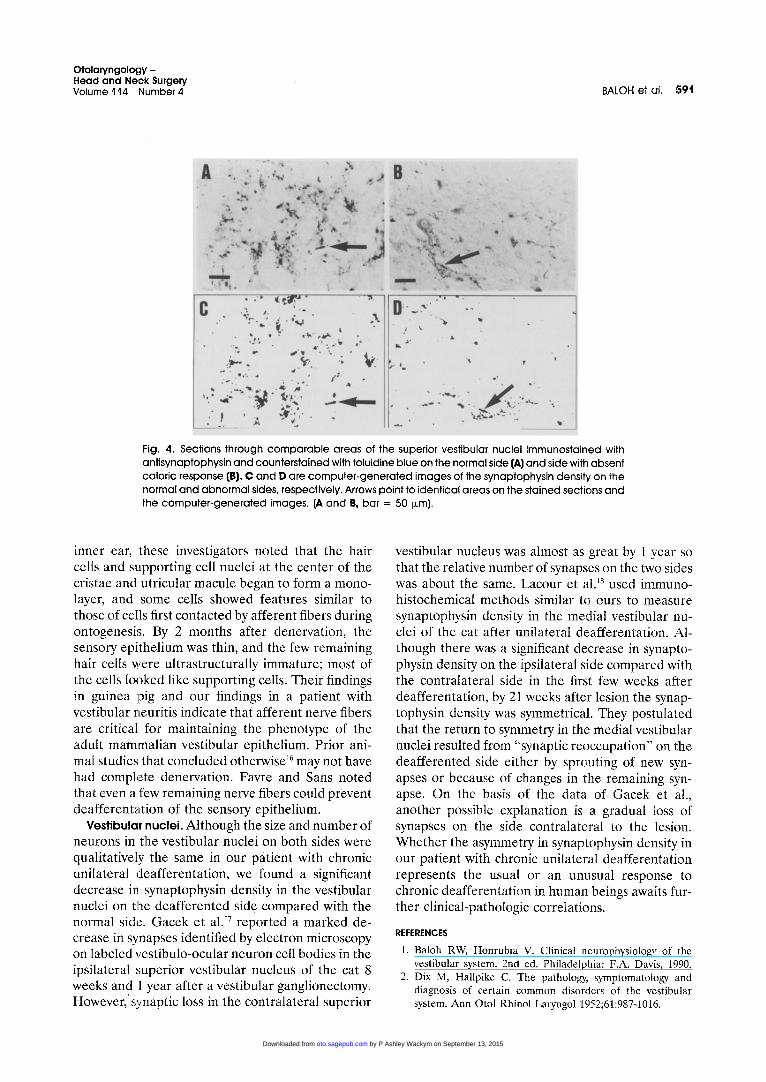
The number of neurons and the morphology of neurons in the four vestibular subnuclei on the two sides were qualitatively similar. As in Scarpa's gan- glia, lipofuscin deposits were common in neurons on both sides of the brain stem. Antisynaptophysin immunoreactivity was significantly less on the left side than on the right (analysis of variance, p < 0.001) (Fig. 4). The percentage decrease on the left side in the four subnuclei was as follows: supe- rior, 59%; medial, 45%; lateral, 35%; and descend- ing, 51%.
DISCUSSION Clinical Features
This patient initially had the characteristic clinical profile of vestibular neuri t is- acute vertigo, nausea, and vomiting gradually resolving over weeks and unassociated with auditory or other neurologic
by P Ashley Wackym on September 13, 2015oto.sagepub.comDownloaded from

Otolaryngology - Head and Neck Surgery Volume t14 Number 4 BALOH et al. 589
J
)
B . /
-" 2
I . . . .
Fig. 2. Sections through c o m p a r a b l e areas of the inferior division of Scarpa's gang l ia on the normal side (A) and the side with absent calor ic response (B) stained with toluidine blue. Only a few small neurons remain in B. [Bar = 100 i~m.]
symptoms. 1 The absent caloric responses on the left side (acutely and 2 years later) documented a per- sistent fixed unilateral peripheral vestibular deficit. Some patients with typical vestibular neuritis will have a return to normal caloric responses. In these cases, presumably the inflammation resolves without leading to any permanent damage. Regardless of whether the caloric response returns, patients re- cover clinically as the brain readjusts and compen- sates for the peripheral vestibular loss. Vestibulo- ocular responses (VORs) to rotational stimuli are markedly asymmetric during the acute phase of vestibular neuritis, but as clinical recovery occurs, symmetry of rotational responses returns. However, the gain of the VOR to high-amplitude step accel-
erations remains asymmetric, and the VOR time constant remains short (as in Fig. 1). 9 Models have been developed to explain these dynamic changes, but their cellular basis is unknown. 9,1°
P a t h o p h y s i o l o g y
Pathologic findings in our patient with vestibular neuritis are most consistent with an isolated viral infection of Scarpa's ganglia on the left. Viruses can be selective for a subpopulation of sensory" neu- rons.l~ The atrophy of individual neurons intermixed with normal-appearing neurons and the absence of fibrosis is not consistent with a vascular cause. That viral inclusions were absent on light and electron microscopy and immunohistochemical stains for
by P Ashley Wackym on September 13, 2015oto.sagepub.comDownloaded from

590 BALOH et al.
Otolaryngology - Head and Neck Surgery
April 1996
Fig. 3. Sections through the center of the utricular macule on the normal side (A) and the side with absent caloric response (B) stained with toluidine blue. There is loss of hair cells and nerve fibers in B. Arrow points to a single remaining hair cell. [A, bar = 50 l~m; B, bar = 35 l~m.]
specific viral antigens were negative does not rule out the many potential viruses implicated in vestib- ular neuritis. 5 Furthermore, the viral antigens that are identified with these immunohistochemical tech- niques may no longer be present 9 years after the acute infection. Other techniques such as poly- merase chain reaction and in situ hybridization are more sensitive because they identify residual viral nucleic acid, 12 but results of these techniques may be positive in normal subjects. 13
Friedman and House 14 found lipofuscin inclusions in Scarpa's ganglia surgically removed from a patient with persistent dizziness caused by vestibular neu- ritis. Because their patient was 47 years old, they
concluded that the lipofuscin inclusions in the ves- tibular neurons represented a degenerative change caused by viral infection. Our patient was older and showed similar amounts of lipofuscin inclusions on both sides, so this finding was probably caused by normal aging.
Chronic Effects of Deafferentat ion
End-organs. The changes in the vestibular end- organs in our patient with vestibular neuritis are likely caused by denervation. Favre and Sans 15 showed similar changes in the sensory epithelium of the cristae and utricular macule in the guinea pig after denervation. Within 2 weeks of denervating the
by P Ashley Wackym on September 13, 2015oto.sagepub.comDownloaded from
Otolaryngology - Head and Neck Surgery Volume '114 Number 4 BALOH et al. 591
C a ,, : a
'.~ ,~ ~,.~ ~ ' . \ .
I D I . " . .
• A
J m w
!
I
Fig. 4. Sections through c o m p a r a b l e areas of the superior vestibular nuclei immunostained with ant isynaptophysin and counterstained with toluidine blue on the normal side (,6,) and side with absent calor ic response (B). C and D are compute r -genera ted images of the synaptophysin density on the normal and abnormal sides, respectively. Arrows point to ident ical areas on the stained sections and the compute r -genera ted images. (A and B, bar = 50 i~m).
inner ear, these investigators noted that the hair cells and supporting cell nuclei at the center of the cristae and utricular macule began to form a mono- layer, and some cells showed features similar to those of cells first contacted by afferent fibers during ontogenesis. By 2 months after denervation, the sensory epithelium was thin, and the few remaining hair cells were ultrastructurally immature; most of the cells looked like supporting cells. Their findings in guinea pig and our findings in a patient with vestibular neuritis indicate that afferent nerve fibers are critical for maintaining the phenotype of the adult mammalian vestibular epithelium. Prior ani- mal studies that concluded otherwise 16 may not have had complete denervation. Favre and Sans noted that even a few remaining nerve fibers could prevent deafferentation of the sensory epithelium.
Vestibular nuclei. Although the size and number of neurons in the vestibular nuclei on both sides were qualitatively the same in our patient with chronic unilateral deafferentation, we found a significant decrease in synaptophysin density in the vestibular nuclei on tl~e deafferented side compared with the normal side. Gacek et all 7 reported a marked de- crease in synapses identified by electron microscopy on labeled vestibulo-ocular neuron cell bodies in the ipsilateral superior vestibular nucleus of the cat 8 weeks and 1 year after a vestibular ganglionectomy. However, synaptic loss in the contralateral superior
vestibular nucleus was almost as great by 1 year so that the relative number of synapses on the two sides was about the same. Lacour et al. 18 used immuno- histochemical methods similar to ours to measure synaptophysin density in the medial vestibular nu- clei of the cat after unilateral deafferentation. Al- though there was a significant decrease in synapto- physin density on the ipsilateral side compared with the contralateral side in the first few weeks after deafferentation, by 21 weeks after lesion the synap- tophysin density was symmetrical. They postulated that the return to symmetry in the medial vestibular nuclei resulted from "synaptic reoccupation" on the deafferented side either by sprouting of new syn- apses or because of changes in the remaining syn- apse. On the basis of the data of Gacek et al., another possible explanation is a gradual loss of synapses on the side contralateral to the lesion. Whether the asymmetry in synaptophysin density in our patient with chronic unilateral deafferentation represents the usual or an unusual response to chronic deafferentation in human beings awaits fur- ther clinical-pathologic correlations.
REFERENCES
1. Baloh RW, Honrubia V. Clinical neurophysiology of the vestibular system. 2nd ed. Philadelphia: F.A. Davis, 1990.
2. Dix M, Hallpike C. The pathology, symptomatology and diagnosis of certain common disorders of the vestibular system: Ann Otol Rhinol Laryngol 1952;61:987-1016•
by P Ashley Wackym on September 13, 2015oto.sagepub.comDownloaded from

.592 BALOH et al.
Otolaryngology - Head and Neck Surgery
April 1996
3. Burrowed W. Acute labyrinthitis. BMJ 1952;2:1182-3. 4. Schuknecht HF. Neurolabyrinthitis. Viral infections of the
peripheral auditory and vestibular systems. In: Nomura Y, ed. Hearing loss and dizziness. Tokyo: Igaku-Shoin, 1985:1-15.
5. Davis LE, Johnsson L-G. Viral infections of the inner ear: clinical, virologic and pathologic studies in humans and ani- mals. Am J Otolaryngol 1983;4:347-62.
6. Mark AS, Fitzgerald D. MRI of the inner ear. Baillieres Clin Neurol 1994;3:515-35.
7. Schuknecht HF, Kitamura K. Vestibular neuritis. Ann Otol Rhinol Laryngol 1981;90(suppl 78):1-10.
8. Sadjadpour K, Brodal A. The vestibular nuclei in man. A morphological study in light of experimental findings in the cat. J Hirnforsch 1968;10:299-323.
9. Baloh RW, Honrubia V, Yee RD, Hess K. Changes in the human vestibulo-ocular reflex after loss of peripheral sensi- tivity. Ann Neurol 1984;16:222-8.
10. Honrubia V, Garland K, Marco J, Minser J, Baloh RW. Vestibulo-ocular reflex changes following peripheral labyrin- thine lesions. In: Ruben RW, ed. The biology of change in otolaryngology. Amsterdam: Elsevier Science Publishers, 1986:155-70.
11. Morgolis TP, Sedarati F, Dobson AT, Feldman LT, Stevens JG. Pathways of viral gene expression during active neuronal infection with HSV-1. Virology 1992;189:150-60.
12. Wackym PA, Popper P, Kerner MM, Grody WW. Varicella- zoster DNA in temporal bones of patients with Ramsay Hunt syndrome. Lancet 1993;342:1555.
13. Furuta Y, Takasu T, Fukuda S, Inuyama Y, Sato KC, Na- gashima K. Latent herpes simplex virus type 1 in human vestibular ganglia. Acta Otolaryngol (Stockh) Suppl 1993; 503:85-9.
14. Friedman I, House W. Vestibular neuronitis. Electron mi- croscopy of Scarpa's ganglion. J Laryngol Otol 1980;94:877- 83.
15. Favre D, Sans A. Dedifferentiation phenomena after dener- vation of mammalian adult vestibular receptors. Neuroreport 1991;2:501-4.
16. Hirokawa N. Disappearance of afferent and efferent nerve terminals in the inner ear of the chick embryo after chronic treatment with beta-bungarotoxin. J Cell Biol 1977;73:27-46.
17. Gacek R, Lyon M, Schoonmaker J. Ultrastructural changes in vestibulo-ocular neurons following vestibular neurectomy in the cat. Ann Otol Rhinol Laryngol 1988;97:42-57.
18. Lacour M, Ez-Zaher L, Raymond J. Plasticity mechanisms in vestibular compensation in the cat are improved by an extract of Ginkgo biloba (EGb 761). Pharmacol Biochem Behav 1990;40:367-79.
by P Ashley Wackym on September 13, 2015oto.sagepub.comDownloaded from
Copyright © 2022 FDOKUMEN




















