
Physiological effects and tissue accumulation of copper in freshwater rainbow trout (Oncorhynchus...
16
AOUATIC TOXICOLgGY ELSEVIER Aquatic Toxicology 29 (1994) 197 212 Physiological effects and tissue accumulation of copper in freshwater rainbow trout (Oncorhynchusmykiss) under normoxic and hypoxic conditions Lone Pilgaard ~'u, Hans Malte *'b, Frank B. Jensen a ~Institute of Biology, Odense University, Campusvej 55, DK-5230 Odense M, Denmark b Institute of Biological Sciences, Department ofZoophysiology, Aarhus University, Building 131, Universitetsparken, DK-8000 Aarhus C, Denmark (Received 15 February 1994; revised 30 November 1994; accepted 8 February 1994) Abstract Rainbow trout (Oncorhynchus mykiss) were exposed to a sublethal copper concentration (0.5 ppm) for 9 days in normoxic (Po2 > 130 mmHg) and hypoxic (70 mmHg), hard water ([Ca 2+] -- 2.15 raM). There were no signs of respiratory dysfunction in any of the exposed fish. A small, transient extracellular alkalosis was induced, which was of metabolic origin in normoxia and of respiratory origin in hypoxia. During copper exposure the plasma concentra- tions of Na +, K + and Ca 2÷ decreased. In hypoxia the decrease in [Na +] continued throughout the experiment, whereas in normoxia [Na ÷] recovered towards the control values. The concentra- tion of copper increased drastically in mucous and gill, liver and kidney tissues, whereas the copper concentration of white muscle, heart, spleen and whole blood was only marginally affected by copper exposure. There were no significant differences in the degree of tissue copper accumulation between copper/normoxia- and copper/hypoxia-exposed fish. Key words." Copper; Copper accumulation; Hypoxia; Acid base status; Ion regulation 1. Introduction Copper occurs naturally in the aquatic environment as a result of weathering and land drainage, but the use of various metal-containing pesticides and fungicides and the use of copper in water pipes, antifouling paints and other human devices have introduced additional quantities of copper into the environment. Copper can be acutely toxic to freshwater fish, the toxicity depending on species, age, size, water temperature and ionic composition (e.g. Demayo et al., 1982; Luoma, 1983). The physiological effects of elevated concentrations of waterborne copper have *Corresponding author. 0166-445X/94/$07.00 © 1994 Elsevier Science B.V. All rights reserved SSDI 0166-445X(94)00014-H
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Physiological effects and tissue accumulation of copper in freshwater rainbow trout (Oncorhynchus...

AOUATIC TOXICOLgGY
ELSEVIER Aquatic Toxicology 29 (1994) 197 212
Physiological effects and tissue accumulation of copper in freshwater rainbow trout (Oncorhynchus mykiss) under
normoxic and hypoxic conditions
Lone Pilgaard ~'u, Hans Malte *'b, Frank B. Jensen a ~Institute of Biology, Odense University, Campusvej 55, DK-5230 Odense M, Denmark
b Institute of Biological Sciences, Department ofZoophysiology, Aarhus University, Building 131, Universitetsparken, DK-8000 Aarhus C, Denmark
(Received 15 February 1994; revised 30 November 1994; accepted 8 February 1994)
Abstract
Rainbow trout (Oncorhynchus mykiss) were exposed to a sublethal copper concentration (0.5 ppm) for 9 days in normoxic (Po2 > 130 mmHg) and hypoxic (70 mmHg), hard water ([Ca 2+] -- 2.15 raM). There were no signs of respiratory dysfunction in any of the exposed fish. A small, transient extracellular alkalosis was induced, which was of metabolic origin in normoxia and of respiratory origin in hypoxia. During copper exposure the plasma concentra- tions of Na +, K + and Ca 2÷ decreased. In hypoxia the decrease in [Na +] continued throughout the experiment, whereas in normoxia [Na ÷] recovered towards the control values. The concentra- tion of copper increased drastically in mucous and gill, liver and kidney tissues, whereas the copper concentration of white muscle, heart, spleen and whole blood was only marginally affected by copper exposure. There were no significant differences in the degree of tissue copper accumulation between copper/normoxia- and copper/hypoxia-exposed fish.
Key words." Copper; Copper accumulation; Hypoxia; Acid base status; Ion regulation
1. Introduction
Copper occurs natural ly in the aquatic environment as a result o f weathering and land drainage, but the use o f various metal-containing pesticides and fungicides and the use o f copper in water pipes, antifouling paints and other human devices have in t roduced addit ional quantities o f copper into the environment. Copper can be acutely toxic to freshwater fish, the toxicity depending on species, age, size, water temperature and ionic composi t ion (e.g. D e m a y o et al., 1982; Luoma, 1983).
The physiological effects o f elevated concentra t ions o f waterborne copper have
*Corresponding author.
0166-445X/94/$07.00 © 1994 Elsevier Science B.V. All rights reserved SSDI 0166-445X(94)00014-H

198 L. Pilgaard et al./Aquatic Toxicology 29 (1994) 197 212
been subject to numerous studies. In teleost fishes copper has been demonstrated to affect branchial ion transport (Laur6n and McDonald, 1987a), plasma electrolyte concentrations (Stagg and Shuttleworth, 1982a), haematological parameters (Mckim et al., 1970) and enzyme activities (Jackim et al., 1970; Donaldson and Dye, 1975). In addition, copper can cause lysis of the red blood cells (Smith et al., 1988), immuno- suppression (Anderson and Dixon, 1989), vertebral deformities (Bengtsson and Lars- son, 1986), and it seems to affect the nervous system (Baatrup, 1991). Histological studies (Baker, 1969) have shown that copper induces morphological changes in the liver, kidney and gills. Measurements of copper content in selected tissues (Benoit, 1975; Stagg and Shuttleworth, 1982a) have revealed copper accumulation in several organs of copper-exposed fish.
Only few studies have addressed the physiological effects and toxicity of a combi- nation of stresses in aquatic organisms. Discharge of wastes from industrial and agricultural activities can contaminate aquatic environment with both organic matter and heavy metals, making the combination of low oxygen content and increased heavy metal levels a likely phenomenon.
Spicer and Weber (1992) found a higher gill diffusion barrier thickness in the marine crab Cancer pagurus exposed to both copper and hypoxia than in animals exposed to copper or hypoxia alone. This synergistic effect of copper and hypoxia was also demonstrated in the shorecrab, Carcinus maenas (Boitel and Weber, unpublished results), where the thickening of the gill epithelium was associated with an increased mortality. Reduced oxygen tensions in the ambient water may also increase the toxic- ity of metals, including copper, to fish (Lloyd, 1961).
In the present study, the physiological effects of copper exposure was investigated in rainbow trout, Oncorhynchus mykiss, both under normoxic and hypoxic condi- tions, and the degree of copper accumulation in several tissues was assessed.
2. Materials and Methods
Experimental animals Rainbow trout (Oncorhynchus mykiss; weight 400 +_ 20 g) were obtained from a
local fish farm and maintained for at least two weeks in 1000-liter aquaria supplied with running, normoxic (Po2 > 130 mmHg) tap water at 15 +_ 1 °C, under a 12 h light/12 h darkness photoperiod. The fish were fed a commercial trout diet, but were starved for 48 h before and during the experiments.
Experimental protocol The fish were anaesthetized in ethyl p-aminobenzoate (benzocaine) (1 : 10 000) and
cannulated with PE 50 tubing in the dorsal aorta (Soivio et al., 1975), while the gills were irrigated with normoxic water containing benzocaine (1:30 000). After surgery the fish were allowed to recover for at least 48 h in the experimental aquaria. These were individual 55-liter tanks, where the fish were allowed unrestrained swimming. The temperature was controlled at 15 + 1 °C throughout the experiments. Half the volume of water was renewed each day.

L. Pilgaard et al./Aquatic Toxicology 29 (1994) 197-212 199
The laboratory water used in the study had the following ionic composition (mM): NH4 < 0.01; NO2 < 0.01; NO~ < 0.05; C1 = 0.69_+ 0.05; SO24 - = 0.16 + 0.03; HCO 3 = 4 . 2 + 0 . 1 ; Na + = 1 . 1 2 + 0 . 0 9 ; K ÷ = 0 . 1 6 + 0 . 0 4 ; Ca 2+=2 .15+0 .05 ; Mg 2+ = 0.44 + 0.05; pH = 8.179 + 0.039.
One control and two experimental series were run:
N O R M O X I A (Control) Six fish were kept in copper-free water throughout the experiment and the oxygen tension was maintained close to air saturation ( > 130 mmHg) by constantly bubbling air through the aquarium water.
N O R M O X I A P L U S COPPER Eight fish was exposed to 0.5 ppm copper (added as CuSO4 '5H20 ) in normoxic water. Water samples were taken at the beginning and at the end of periods between water renewal and analyzed for copper to de- tect any displacement of copper from the waterphase to e.g. the walls of the aquar- ium.
H Y P O X I A P L U S COPPER Seven fish were exposed to 0.5 ppm copper in hy- poxic water. The oxygen tension was lowered to 70 + 5 mmHg by means of feed-back controlled bubbling of nitrogen through the water. The Po2 was monitored by a Radiometer E4047 electrode connected to a PHM 71 monitor. Water samples were analysed for copper as above.
Blood (0.7 ml) was sampled on day 0 (control) and at 3 h, 1 day, 2 days, 4 days, 7 days and 9 days after addition of copper.
At the 9th day of exposure an additional bloodsample was collected (1 ml) before the fish were killed by a sharp blow to the head. Hereafter the fish was weighed. A mucous sample from the gills was collected, dissolved in 0.6 M perchloric acid (1:2) and frozen for later analysis of Cu concentration. The second gill arch on the left side of the fish was dissected out and washed in saline (0.9%) to free it from mucous. The heart was dissected out and injected with a heparin solution (Na-heparinate in 0.9% NaC1) to wash out blood. Further tissue samples (0.5 1 g) were taken from the spleen, the liver, kidney and white muscle (epaxial muscle immediately posterior to the dorsal fin). All the tissue samples were weighed and stored at -80°C.
The tissue samples were freeze-dried to constant weight and digested in concen- trated HNO3 and H 2 0 2. The remaining salts were redissolved in 10 ml 0.2% HNO3 and used for analysis for copper concentration.
Measuremen ts
Blood gas tensions, acid-base status and haematological parameters were deter- mined immediately after sampling of blood.
Arterial blood pH and oxygen tension (Paoz) were measured at 15°C on a BMS 3 Mk 2 electrode assembly linked to a PHM 71 Mk 2 Acid-base analyser (Radiometer, Copenhagen). The plasma total carbon dioxide content (Cco2) and the whole blood oxygen content (Co2) was measured by the methods of Cameron (1971) and Tucker (1967), respectively.
Haematocrit (Hct) was determined by centrifugation of blood (3 min at 12 000

200 L. Pilgaard et al./Aquatic Toxicology 29 (1994) 197-212
rev/min) in glass capillaries, and the blood haemoglobin concentration was measured by the cyanmethaemoglobin method.
Plasma for cation analyses was obtained by centrifugation of blood, and was stored at -80°C until analysed. For plasma lactate and chloride measurements, 70 ¢tl of plasma was deproteinized with 140 oil of PCA, and the supernatant was stored at -80°C until use. Lactate was measured according to the lactate dehydrogenase enzy- matic method (Boehringer-Mannheim). Chloride was measured with a Radiometer CMT 10 chloride titrator. Plasma sodium and potassium concentrations were deter- mined by a FLM3 flamephotometer (Radiometer, Copenhagen). Measurements of calcium, magnesium and copper were carried out by atomic absorbtion spectro- photometry (Perkin-Elmer 2380).
Calculations The arterial CO2 tension, Pco2, was calculated from the measured values of blood
pH and plasma total CO2:
Pc02 = Cc02/(ac02 " (10(pH-pKa) + 1))
using the solubility coefficient of CO2 in plasma, (C~co 2 (15°C) = 53.5/IM/mmHg) and the apparent dissociation constant of carbonic acid (pK~ = 6.781-(0.089.pH)) in trout plasma (Boutilier, 1984). The concentration of HCO3 was calculated as the difference between Cco2 and the concentration of free CO2:
[HCO3] = Cco2 -Ctco2"Pco2
The 02 content was corrected for dissolved 02 using a Bunsen solubility coefficient (~o2) of 1.8 ¢tM/mmHg (Christoforides and Hedley-Whyte, 1969).
The haemoglobin concentration was calculated using a millimolar extinction coef- ficient of 10.99 mM 1 cm-i (monomeric) for cyanmethaemoglobin at 540 nm. The mean cellular haemoglobin concentration (MCHC) was estimated as:
MCHC = 100. CHb]HCt
Presentation of the data Data are presented as means _+ SEM, unless otherwise stated. Paired, two-tailed
t-tests were used to evaluate the level of significance of differences relative to control values (each fish serving as its own control). Unpaired t-tests were used for comparing datasets of exposed and unexposed fish.
3. Results
Blood respiratory properties Arterial Po2 increased slightly with time in the control group and in the copper-

L. Pilgaard et al./Aquatic Toxicology 29 (1994) 197-212 201
1 5 0
" ~ 1 2 5 .
E 100- E 7 5
5 0
2 5 ¸
5
Normoxia Hypoxia
o ~ O
4
3"
2 "
1
0 ; i ) ; ~ ; o l l i -) ;
days days
Fig. 1. Time-dependent changes of dorsal aortic oxygen tension and oxygen concentration in rainbow trout in normoxic water (c~), in normoxic, copper-containing water (w) and in hypoxic, copper-containing water (e). All values are means + SEM. Significant differences between the exposed fish and their own control value (day 0) are denoted by *, significant differences between copper-exposed and control fish of the individual timepoints are denoted by +, and significant differences between copper/hypoxia exposed and copper exposed fish are denoted by *. The significance levels are indicated by: *P < 0.05, **P < 0.01, ***P < 0.001.
exposed group, whereas it fell significantly to half the initial value in copper /hypoxia- exposed fish (Fig. 1). This decrease in oxygen tension caused an immediate decrease in the arterial oxygen concentra t ion to values that stayed more or less constant at subsequent sampling times (Fig. l, lower right panel).
In control fish and in fish exposed to copper under normoxic conditions, the arteri- al Co2 decreased gradual ly during the experimental period (Fig. 1). This was due to the repeated blood sampling, as reflected by the decrease in haematocr i t and blood haemoglobin concentra t ion (Fig. 2). The effect o f b lood sampling was more modest in copper /hypoxia-exposed fish, p robab ly as the result o f a hypoxia- induced release o f red cells f rom the spleen, which caused an immediate increase in b lood [Hb] and limited the [Hb] decrease at subsequent sampling times (Fig 2). The mean cellular haemoglobin concentra t ion ( M C H C ) did not change significantly in any o f the exper- imental groups (Fig. 2), showing that red cell swelling was not induced.
Blood acid-base status
Exposure to copper resulted in a small but significant extracellular alkalosis both

202 L. Pilgaard et al./Aquatic Toxicology 29 (1994) 197-212
"1-
3 0
20
1 0
Normoxia Hypoxia
E
"1-
1.0 ¸
0 . 8
0.6
0.4
0.2
0
6.
A 5.
0
3
0 1 1 ~ + ; 0 1 1 i + ; days days
Fig. 2. Changes in haematocrit (Hct), blood haemoglobin concentration (Hb) and mean cellular haemo- globin concentration (MCHC) in rainbow trout in normoxic water ([]), in normoxic, copper-containing water (w) and in hypoxic, copper-containing water (e). Other symbols as in Fig. 1.
under normoxic and hypoxic conditions. The alkalosis lasted for about two days whereafter pH returned towards control values (Fig. 3).
The arterial Pco2 in the copper/normoxia-exposed fish (Fig. 3) was slightly higher than in control fish at day 0 but increased significantly at 3 h and at day 1, which was not the case in control fish. In copper/hypoxia-exposed fish the arterial Pco2 de- creased significantly (Fig. 3). The plasma bicarbonate concentration increased and stayed elevated for two days in copper-exposed fish under normoxic conditions, whereas it decreased significantly under hypoxic conditions (Fig. 3). Plasma lactate concentrations were low and unaffected by copper or copper/hypoxia stress (Fig. 3).
The extracellular acid-base changes following copper and copper/hypoxia expo-
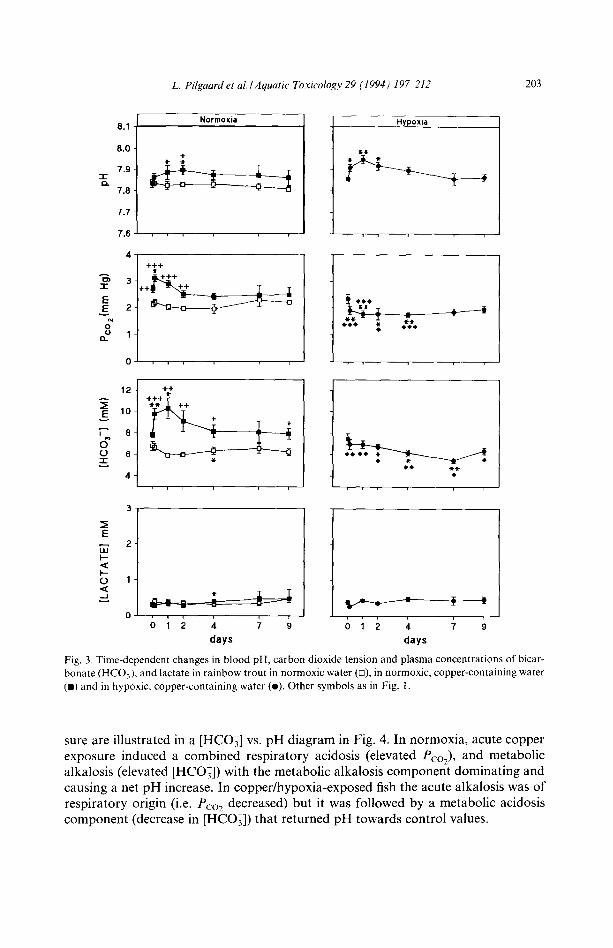
L. Pilgaard et al./Aquatic' Toxicology 29 (1994) 19~212 203
8.1
8.0
7.9 "I- Q.
7.8
7.7
7.6
4"
"~ 3. T
E E 2
g
12-
E lO-
O (9 6 -i-
4
Norrnoxia
+ -I(-
+~*
+ +
+.+.÷ ÷ . ÷
F4
Hypoxia
E
LLI t---
p- (9
3.
2 .
days days
Fig. 3. Time-dependent changes in blood pH, carbon dioxide tension and plasma concentrations of bicar- bonate (HCOj) and lactate in rainbow trout in normoxic water (D), in normoxic, copper-containing water (m) and in hypoxic, copper-containing water (e). Other symbols as in Fig. 1.
sure are illustrated in a [HCO3] vs. pH diagram in Fig. 4. In normoxia, acute copper exposure induced a combined respiratory acidosis (elevated Pco2), and metabolic alkalosis (elevated [HCO3]) with the metabolic alkaiosis component dominating and causing a net pH increase. In copper/hypoxia-exposed fish the acute alkalosis was of respiratory origin (i.e. Pco2 decreased) but it was followed by a metabolic acidosis component (decrease in [HCO3]) that returned pH towards control values.

204 L. Pilgaard et al./Aquatic' Toxicology 29 (1994) 197 212
12
E ~ 1 0
7-"
cr 8 + I
G 6 0 -r
/ '3 /
1 / /
7
i i
7.8 7.9
pH 8.0
Fig. 4. Plasma [HCO3] versus pH diagram including carbon dioxide tension isopleths and the changes in extracellular acid base status in rainbow trout in normoxic water (•), in normoxic, copper-containing water (1) and in hypoxic, copper-containing water (e). 0 denotes the control value. 3 h, 1,2, 4, 7, 9 denotes three hours, one day, two days, etc. after start of the exposure. Other symbols as in Fig. 1. The shown buffer line was calculated from the blood haemoglobin concentration using the relationship given by Wood, McDonald and McMahon (1982).
Extracellular electrolyte balance Plasma [Na +] (Fig. 5) decreased significantly in both copper-exposed groups. Un-
der normoxic conditions [Na +] returned to control values at day 7, whereas no com- pensation was seen under hypoxic conditions, resulting in a significantly lower [Na +] in copper/hypoxia-exposed fish than in copper/normoxia-exposed fish at day 7 and 9. Plasma [C1-] stayed constant in control fish but was slightly decreased in copper- exposed fish.
The concentration of potassium in plasma was reduced by about 20% at day 9 in both copper-exposed groups when compared to the control values at day 0 (Fig. 5).
The plasma calcium concentration (Fig. 5) also decreased significantly by about 20% in both copper exposed groups during the 9 days exposure period.
Plasma magnesium did not change significantly with time in control and copper/ normoxia-exposed rainbow trout, whereas a small but significant decrease occurred in copper/hypoxia-exposed fish.
Tissue copper concentration The copper concentration of mucous collected at the gill surface after nine days of
copper exposure was about 1.5/~g/g wet weight in both the experimental groups, which was an increase by 700% compared to the control group (Fig. 6a). The copper concentration of the mucous-free gill tissue was raised by 570% in the copper-exposed group and by 490% in the copper/hypoxia-exposed group, reaching values of 33 and 29 ~g/g dry weight, respectively, after 9 days of exposure (Fig. 6b). In the liver, the Cu concentration was some 10 times higher than in other tissues, and it increased by 110% and 165% upon exposure to copper under normoxic and hypoxic conditions,

L. Pilgaard et al./Aquatic" Toxicology 29 (1994) 19~212 205
160
150
140 "4-
:~ 130
120
Norrnoxia
+
I I
Hypoxia
150
140 E
130 i
~-. 120
11(1
÷+
+ + ÷
2.8
3; 2.6
E 2.4
~,~ 2.2
2.0
1.8
3.0
~E 2.5 E
2.0
1.5
1.0
1.0
* 41-
0.8
0.6 ~E
0.4
-I-
4P
days days
Fig. 5. Time-dependent changes in plasma sodium [Na+], chloride [C1-], potassium [K+], calcium [Ca 2+] and magnesium [Mg 2+] of rainbow trout in normoxic water ([]), in normoxic, copper-containing water ( i ) and in hypoxic, copper-containing water (o). Other symbols as in Fig. l. Dashed lines are biexponential curves fitted to the calcium data for copper exposed fish. The rate constants are 6.61 day -~ and 0.018 day n under normoxic conditions, and 8.51 day i and 0.012 day -t under hypoxic conditions.
respectively. The copper concentration of the kidney increased by 100% in the copper/ normoxia-exposed group and by 137% in the copper/hypoxia-exposed group (Fig. 6b). In contrast to mucous and to gill, liver and kidney tissues, the copper concentra-

206 L. Pilgaard et al./Aquatic Toxicology 29 (1994) 197 212
o ~ ~ **
o E m ~ 6 u
o mucus
TISSUE
"o
-I w
0 == o
e~ I--
[] =k
=k =k
gill muscle heart
=k
liver
TISSUE
500
~ control 1 copper [ _r:. hypoxia/ 4 0 0 < copper
300 -~ o
200 ~
__.
r ~ 100 ¢=
0 kidney spleen blood
Fig. 6. Copper concentration in mucus (Fig. 6a) (measured as,ug copper/g wet weight), and in gill, muscle, heart, liver, kidney, spleen and whole blood (Fig. 6b) (measured as pg copper/g dry weight) after 9 days of exposure. Note that the value for the liver should be read at the right axis. Significant differences between
exposed and control fish are denoted by: *P < 0.05, **P < 0.01, ***P < 0.001.
tion of whole blood and white muscle, heart and spleen tissues was not elevated to any appreciable extent by copper exposure (Fig. 6b).
4. Discussion
Respiratory changes Metal exposure in fish can cause structural changes in the gills that may disturb
both ionoregulatory and respiratory functions (McDonald and Wood, 1993). In the

L. Pilgaard et al./Aquatic Toxicology 29 (1994) 197-212 207
present experiments on copper exposure in rainbow trout there was clear evidence of ionoregulatory disturbances (cf. below) whereas oxygen transport was not seriously compromised. Thus, exposure to 0.5 ppm copper in normoxic water did not reduce arterial Po2 and even though arterial Po2 decreased in hypoxia, the presence of 0.5 ppm copper did not exacerbate the situation to an extent that anaerobiosis (lactic acid production) or red cell swelling was induced. These findings suggest that copper exposure did not induce structural or functional changes of the gill that affected the gill 02 diffusion conductance. This is in line with the idea that for many metals the toxicant threshold for respiratory effects is higher than the threshold for ionoregula- tory effects (McDonald and Wood 1993).
Blood acid-base s tatus The changes in acid-base status during copper exposure in the rainbow trout were
small but of different nature in normoxia and hypoxia. In normoxic fish an acute, simultaneous respiratory acidosis and metabolic alkalosis was induced, with the met- abolic alkalosis (mediated by an increase in strong ion difference) dominating and causing an increase in bicarbonate concentration and pH. Copper/hypoxia-exposed fish responded with an acute respiratory alkalosis, which is the classic response to moderate hypoxia (e.g. Thomas and Hughes, 1982). The increase in P,co2 observed in copper/normoxia-exposed fish was not parallelled by a lowered Pao2 suggesting that copper transiently inhibited the excretion of carbonic dioxide specifically. This could be due to a transient inhibition of the gill epithelial carbon anhydrase, since a small part the total carbon dioxide excretion is mediated by H + and HCO3 ions generated from the hydration of CO2. In copper/hypoxia exposed fish, this effect apparently was overridden by hyperventilation, that decreased arterial Pco 2.
In contrast to the small and transient alkalosis in rainbow trout (Figs. 3 and 4) the euryhaline decapod Carcinus maenas (shore crab) exposed to copper in brackish wa- ter (where it hyperosmoregulates like freshwater fish) gradually develops a large and persistent non-lactic metabolic acidosis with major decreases in haemolymph pH and bicarbonate concentration and only minor changes in carbon dioxide tension (Boitel and Truchot, 1990; Weeks et al., 1993). Thus, the size and nature of the acid base changes during copper exposure appears to differ between freshwater fish and hyper- osmoregulating crustaceans.
Extracel lular electrolyte balance Exposure to copper decreased the plasma concentration of several major ions un-
der normoxic as well as hypoxic conditions (Fig. 5). Since plasma ion concentrations stay constant with time in normoxic (Fig. 5) and hypoxic (Soivio et al., 1981) rainbow trout in the absence of copper, the extracellular ion decrease was clearly a copper- induced effect.
Several investigators have measured the concentration of monovalent ions in plas- ma of fish exposed to copper. In general, plasma ion concentrations increase in hy- poosmotic marine teleosts (Stagg and Shuttleworth, 1982a; Cardeilhac et al., 1979) and decrease in hyperosmotic freshwater fish (Christensen et al., 1972; McKim et al., 1970). Lorz and McPherson (1976) showed that the gill Na+/K+-ATPase activity in

208 L. Pilgaard et al./Aquatic Toxicology 29 (1994) 197-212
yearling coho salmon (Oncorhynchus kisutch) decreased in proportion to the concen- tration of copper, and that this reduced the Na + uptake across the gill epithelium. Laur6n and McDonald (1985, 1986) showed that a net loss of Na ÷ during copper exposure was the result of a concentration-dependent inhibition of influx as well as a concentration-dependent stimulation of efflux, and that the efflux component was strongly dependent on water hardness and alkalinity. Under the experimental condi- tions of the present study (hard water, high alkalinity), Na + efflux was most likely not affected suggesting that inhibition of active sodium uptake accounted for the ob- served plasma Na + depletion.
Following the decline in Na + and C1- during the first days of copper exposure under normoxic conditions the plasma concentrations of these ions recovered to the pre-exposure level (Fig. 5). An acclimational response of branchial Na + transport kinetics to copper exposure has previously been shown to involve recovery of an increased Km and decreased Jm,x toward preexposure levels (Lauren and McDonald, 1987a). The biochemical basis for this recovery may be an increase in the number of Na+/K+-ATPase units as has been shown in metal-stressed fish (e.g. Stagg and Shut- tleworth, 1982b) or be caused by proliferation of chloride cells. In contrast to the situation in normoxic water, plasma Na + did not recover towards control values during copper exposure in hypoxic water. The lack of an acclimational response in the copper/hypoxia-exposed fish may be due to an added metabolic cost, which can not be meet at the imposed level of hypoxia.
Calcium uptake from the water requires active transport by a Ca2+-ATPase situat- ed in the basolateral membrane of the chloride cells (Perry and Flik, 1988). In vitro studies by Shephard and Simkiss (1978) on the roach Rutilus rutilus have shown, that several heavy metals, including copper, inhibits the CaZ+-ATPase. The reduced calci- um concentration in both groups of copper-exposed fish (Fig. 5) may therefore result from an inhibition of the branchial Ca2+-ATPase as well as from an increased passive efflux. The plasma Ca 2+ concentration vs. time curves for both the copper-exposed and copper/hypoxia-exposed fish fit very nicely to a biexponential decay curve (Fig. 5), suggesting that there is a rapid loss to the environment and a slow gain from another calcium pool, acting as a buffer. This buffer pool could be white muscle, which has a large pool of calcium compared to the plasma. It is noteworthy, that there is a continual loss of calcium throughout the experiment, despite that the concentra- tion gradient for calcium runs down, or is reversed (water [Ca 2+] = 2.15 mM). Thus the net outwards flux is largely driven by the transepithelial potential difference across the gill. Also, in contrast to sodium and chloride concentrations under copper/ normoxia exposure, there were no sign of recovery in plasma Ca 2+ during the nine days of copper exposure.
Tissue copper concentrations The uptake of copper by the fish is considered to be passive, involving diffusion
down gradients made by adsorption or binding of the metal to tissues or cell surfaces. The gill is an obvious site of copper uptake, because of its great surface area and
intimate contact with the external environment. The mucous covering the gills accu- mulated copper to a great extent (Fig. 6a). This binding of copper to mucous might

L. Pilgaard et al./Aquatic Toxicology 29 (1994) 197 212 209
hinder the most toxic states of copper, the copper ion and the copper hydroxides (Meador, 1991), in binding to the apical membrane of the gill epithelium. An elevated rate of mucus secretion appears to be a defense mechanism against several toxic substances, including acid (McDonald, 1983), aluminium (Handy et al., 1989) and zinc (Sellers et al., 1975). In the present experiments, however, this response was not observed. Similarly, Sellers et al. (1975) observed no excessive mucous production, when rainbow trout were exposed to copper concentrations ranging downwards from the 48-h LC50. Baker (1969) reported a decreased number of mucous cells in the gills of flounder (Pseudopleuronectes americanus) exposed to low levels of copper. In carp, Cyprinus carpio, exposure caused an apparently immediate ejection of the mucous cell content, after which the cells remained empty, suggesting a possible decrease in mucus secretion (Labat et al., 1974). Thus, the chelation of copper by mucus seen in the copper-exposed trout (Fig. 6a) may only provide limited protection against copper toxicity.
Gill tissue freed of mucous was the tissue showing the greatest increase in copper concentration (Fig 6b), which agrees with the idea that the gill initially is the primary target organ of copper toxicity (Brungs et al., 1973; Buckley et al., 1982; Stagg and Shuttleworth, 1982a). Lauren and McDonald (1987b), however, observed only a modest increase in gill copper concentration of juvenile rainbow trout during 28 days of copper exposure. The higher gill Cu accumulation observed in the present study may be due to a more efficient metallothionein induction in adult fish than in juve- niles.
Apart from the gill, the liver and the kidney showed large increases in copper concentration, whereas Cu-accumulation in muscle, heart, spleen and whole blood was modest or absent (Fig. 6b). This corroborates the view, that the gills, liver and kidney are the organs in aquatic animals in which metallothioneins are mostly con- centrated (Roesijadi, 1992). Induction of metallothioneins in these organs leads to metal accumulation in the cells, but the specific binding to the metallothionein mole- cules keeps the free concentration of metal extremely low, which protects cellular proteins from metal-induced damages.
The much higher copper concentration in the liver than in the other tissues (Fig. 6b), may be ascribed to its role as the major storage and regulatory organ in copper homeostasis. Olsson et al. (1987) reported that the hepatic copper levels were higher in rainbow trout than in other teleosts. Stagg and Shuttleworth (1982a) and Buckley et al. (1982) did, however, find copper levels in the liver of flounder (Platichthys flesus) and coho salmon (Oncorhynchus kitsutch) that were comparable to those here observed. The increase in liver copper concentration quantitatively accounts for the major part of the total copper accumulation during the experiment. Accumulation in tissues like white muscle, seems to become important only when the maximal storage capacity of the liver is reached (Lauren and McDonald, 1987b). In the present exper- iment the maximal storage capacity of the liver was probably not reached.
The accumulation of copper in tissues such as the gill, the liver and the kidney (Fig. 6b), may cause histological changes (e.g. Baker, 1969). This will occur, however, only when de novo synthesis of metallothioneins is insufficient to meet the sum of net influx of heavy metal and the metal that is released from degraded metallothionein

210 L. Pilgaard et al./Aquatic Toxicology 29 (1994) 197 212
(Hogs t rand and Haux, 1991). This state was probably no t reached in the present experiments. Even the 6 7 fold increase in gill copper concentra t ion , did not cause major histological changes, as judged from the absence of respiratory disturbances.
Lloyd (1961) has suggested, that a greater toxicity of copper in hypoxic waters than
in normoxic waters could be related to the increased volume of water coming into contact with the gills as a result of the hypoxia- induced hypervent i la t ion. This argu- ment will, however, only apply, if a significant a m o u n t of the copper enter ing the inter lamellar spaces of the gill is taken up as the water passes the secondary lamellae. A rough est imation, based on the measured increase in liver copper concent ra t ion , and using an (unrealistically high) liver weight of 50 g kg ~ bodyweight yields a total net influx of copper of abou t 2 mg copper kg -~ dur ing the n ine days of Cu exposure. In the same time the estimated a m o u n t of copper coming into contact with the gills (using a vent i la t ion volume of 200 ml rain ~ kg -~ and the nomina l water copper
concent ra t ion of 0.5 mg 1 -~) is ca. 1300 mg copper kg -~. With a m a x i m u m of only 0.15% of the copper removed from the venti lated water, an increased vent i la t ion
minu te volume canno t increase the diffusion gradient for copper and thereby copper uptake. This agrees with the results of the present study, where there were no statisti- cally significant differences between the accumula t ion of copper in fish exposed to copper under hypoxic and normoxic condi t ions (Fig. 6b). A similar result was report- ed in an invest igat ion of the effects of z inc /normoxia and zinc/hypoxia on ra inbow trout (Hughes and Flos, 1978), where there were no significant differences between the zinc concen t ra t ion of the gills in the two groups of fish. Thus the observed differ-
ences in the response to copper exposure between the hypoxic and the normoxic group canno t be related to differences in the rate of accumula t ion of copper.
References
Anderson, D.R and O.W. Dixon (1989) Suppression of antibody-producing cells in rainbow trout spleen sections exposed to copper in vitro. J. Aquat. Anita. Health I, 57 61.
Baatrup, E. (1991) Structural and functional effects of heavy metals on the nervous system, including sense organs offish. Comp. Biochem. Physiol. 100C, 253 257.
Baker, J.T.R (1969) Histological and electron microscopical observations on copper poisoning in the Winter Flounder (Pseudopleuronectes americanus). J. Fish. Res. Board Can. 26, 2785 2793.
Bengtsson, B. and A. Larsson (1986) Vertebral deformities and physiological effects in fourhorn Sculpin (Myoxocephalus quadricornis) after long-term exposure to a simulated heavy metal containing effluent. Aquat. Toxicol. 9, 215 229.
Benoit D.A. (1975) Chronic effects of copper on survival, growth, and reproduction of the Bluegill (Lc- pomis macrochirus). Trans. Am. Fish. Soc. 104, 353 358.
Boitel, F. and J.l~ Truchot (1990) Comparative study of the effects of copper on haemolymph ion concen- trations and acid base balance in the shore crabs Carcinus maenas acclimated to full-strength or dilute seawater. Comp. Biochem. Physiol. 95C, 307 312.
Boutilier. R.G., T.A. Heming and G.K. lwama (1984) Appendix: Physiochemical parameters tbr use in lish respiratory physiology. In: Fish Physiology XA, edited by W.S. Hoar and D.J. Randall. Academic Press, New York, pp. 403430.
Brungs, W.A., E.N. Leonard and J.M. McKim (1973) Acute and long-term accumulation of copper by tile brown bullhead lctalurus nebulosus. J. Fish. Res. Board Can. 3(I, 583 586.
Buckley, J.T., M. Roch, J.A. McCarter, C.A. Rendell and A.T. Matheson (1982) Chronic exposure of coho

L. Pilgaard et aL / Aquatic Toxicology 29 (1994) 19~212 211
salmon to sublethal concentrations of copper I. Effect on growth, on accumulation and distribution of copper, and on copper tolerance. Comp. Biochem. Physiol. 72C, 15 19.
Cameron, J.N. (1971) Rapid method for determination of total carbon dioxide in small blood samples. J. Appl. Physiol. 31,632 634.
Cardeilhac, RT., C.F. Simpson, R.L. Lovelock, S.F. Yosha, H.W. Calderwood and J.C. Gudat (1979) Failure of osmoregulation with apparent potassium intoxication in marine teleosts: a primary toxic effect of copper. Aquaculture 17:231 239.
Christensen, G.M., J.M. McKim, W.A.Brungs and E.R Hunt (1972) Changes in the blood of the Brown Bullhead (h'talurus nebulosus (Lesueur)) following short and long term exposure to copper(It). Toxicol. Appl. Phamacol. 23,417-427.
Christoforides, C. and J. Hedley-Whyte (1969) Effect of temperature and hemoglobin concentration oll solubility ofO 2 in blood. J. Appl. Physiol. 27, 592 596.
Demayo, A., M.C. Taylor and K.W. Taylor (1982) Effects of copper on humans, laboratory and farm animals, terrestrial plants and aquatic life. From the series entitled Guidelines for Surface Water Quality, Vol. 1, Inorganic Chemical Substances. Department of the Environment Ottawa, Ontario, Canada.
Donaldson, E.M. and H.M. Dye (1975) Corticosteroid concentrations in Sockeye Salmon (Oncorhynchus nerka) exposed to low concentration of copper. J. Fish. Res. Board Can. 32, 533 539.
Handy, R.D., F.B. Eddy and G. Romain (1989) In vitro evidence for the ionoregulatory role of rainbow trout mucus in acid, acid/aluminium and zinc toxicity. J. Fish. Biol. 35, 737 747.
Hogstrand, C. and C. Haux (1991) Binding and detoxification of heavy metals in lower vertebrates with reference to metallothionein (minireview). Comp. Biochem. Physiol. 100C, 137 141.
Hughes, G.M. and R. Flos (1978) Zinc content of the gills of rainbow trout (S. gairdneri) after treatment with zinc solutions under normoxic and hypoxic conditions. J. Fish. Biol. 13,717 728.
Jackim, E., J.M. Hamlin and S. Sonis (1970) Effects of metal poisoning on five liver enzymes in the Killifish (Fundulus heteroclitus) J. Fish. Res. Board Can. 27, 383 390.
Labat, R., J. Pequignot and A. Chatelet (1974) Toxic action of copper on the gills of carp (Qvprimts carpio). Ann. Limnol. 10, 109 114.
Laur6n, D.J. and D.G. McDonald (1985) Effects of copper on branchial ionoregulation in the rainbow trout, Salrno Gairdneri Richardson, modulation by water hardness and pH. J. Comp. Physiol. B155,635 644.
Lauren, D.J. and D.G. McDonald (1986) Influence of water hardness, pH, and alkalinity on copper toxicity in juvenile rainbow trout, Salmo gairdneri. Can. J. Fish. Aquat. Sci. 43, 1488 1496.
Laur6n, D.J. and D.G. McDonald (1987a) Acclimation to copper by rainbow trout, Sah~to gairdneri: Physiology. Can. J. Fish. Aquat. Sci. 44, 99 104.
Laur6n, D.J. and D.G. McDonald (1987b) Acclimation to copper by rainbow trout, Salmo gairdneri: Biochemistry. Can. J. Fish. Aquat. Sci. 44, 105 1 t 1.
Lloyd, R. (1961) Effect of dissolved oxygen concentrations on the toxicity of several poisons to Rainbow trout (Salmo Gairdneri Richardson). J. Exp. Biol. 38, 447455.
Lorz, H.W. and B.R McPherson (1976) Effects of copper or zinc in fresh water on the adaptation to sea water and ATPase activity, and the effects of copper on migratory disposition of coho salmon (On- corhynchus kisutch). J. Fish. Res. Board Can. 33, 2023 2030.
Luoma, S.N. (1983) Bioavailability of trace metals to aquatic organisms a review. Sci. Total Environ. 28, 1 22.
McDonald, D.G. (t983) The effects of H + upon the gills of freshwater fish. Can. J. Zool. 61,691 703. McDonald, D.G. and C.M. Wood (1993) Branchial mechanism of acclimation to metals in freshwater fish.
In: Fish Ecophysiology, edited by J.C. Rankin and F.B. Jensen. Chapman & Hall, London, pp. 297 321. McKim, J.M., G.M. Christensen and E.P. Hunt (1970) Changes in the blood of Brook Trout (Salvelinus
Jimtinalis) after short-term and long-term exposure to copper. J. Fish. Res. Board Can. 27, 1883 1889. Meador, J.l~ (1991) The interaction of pH, dissolved organic carbon and total copper in the determination
of ionic copper and toxicity. Aquat. Toxicol. 19, 13 32. Olsson, P., C. Haux and L. F6rlin (1987) Variations in hepatic metallothionein, zinc and copper levels
during an annual reproductive cycle in rainbow trout, Salmo gairdneri. Fish Physiol. Biochem. 3, 39~7.

212 L. Pilgaard et al./Aquatic Toxicology 29 (1994) 197 212
Perry S.F. and G. Flik (1988) Characterization of branchial trans-epithelial calcium fluxes in freshwater trout, Salmo gairdneri. Am. J. Physiol. 254, R491 R498.
Roesijadi, G. (1992) Metallothioneins in metal regulation and toxicity in aquatic animals. Aquat. Toxicol. 22, 81 114.
Sellers, C.M., Jr., A.G. Heath and M.L. Bass (1975) The effect of sublethal concentrations of copper and zinc on ventilatory activity, blood oxygen and pH in rainbow trout (Sahno Gairdneri). Water Res. 9, 401 408.
Shephard, K. and K. Simkiss (1978) The effects of heavy metal ions on Ca=+-ATPase extracted from fish gills. Comp. Biochem. Physiol. 61,69 72.
Smith, H.A., A.K. Dasmahapatra, J.M. Caffrey and E. Frieden, 1988. A comparative study of copper ion induced lysis of vertebrate red blood cells. Comp. Biochem. Physiol. 91C, 301 305.
Soivio, A., K. Nyholm and K. Westman (1975) A technique for repeated sampling of the blood of individ- ual resting fish. J. Exp. Biol. 63,207 217.
Soivio, A., M. Nikinmaa, K. Nyholm and K. Westman (1981) The role of gills in the responses of Salmo Gairdneri during moderate hypoxia. Comp. Biochem. Physiol. 70A, 133 139.
Spicer, J.I. and R.E. Weber (1992) Respiratory impairment by water-borne copper and zinc in the edible crab Cancer pagurus (Crustacea: Decapoda) during hypoxic exposure. Mar. Biol. 112,429~,35.
Stagg, R.M. and T.J. Shuttleworth (1982a) The accumulation of copper in PlatiehthysJtesus L. and its effects on plasma electrolyte concentrations. J. Fish. Biol. 20,491 500.
Stagg, R.M. and T.J. Shuttleworth (1982b) The effects of copper on ionic regulation by the gills of the seawater-adapted Flounder (PlatichthysJtesus L.). J. Comp. Physiol. 149, 83 90.
Thomas, S. and G.M. Hughes (1982) A study of the effects of hypoxia on acid base status of rainbow trout using an extracorporeal blood circulation. Respir. Physiol. 49, 371 382.
Tucker, V.A. (1967) Method for oxygen content and dissociation curves on microliter blood samples. J. Appl. Physiol. 23, 410414.
Weeks, J.M., F.B. Jensen and M.H. Depledge (1993) Acid base status, haemolymph composition and tissue copper accumulation in the shore crab Carcinus maenas exposed to combined copper and salinity stress. Mar. Ecol. Prog. Ser. 97, 91 98.
Wood, C.M., D.G. McDonald and B.R. McM ahon (1982) The i nfl uence of experimental anaemia on blood acid base regulation in vivo and in vitro in the starry flounder (Platichthys stellams) and the rainbow trout (Salmo gairdneri). J. Exp. Biol. 96, 221 237.




















