
Effects of acute and prolonged naphthalene exposure on brain monoaminergic neurotransmitters in...
11
Effects of acute and prolonged naphthalene exposure on brain monoaminergic neurotransmitters in rainbow trout (Oncorhynchus mykiss) Manuel Gesto, Adrián Tintos, José L. Soengas, Jesús M. Míguez ⁎ Departamento de Biología Funcional y CC. de la Salud, Facultad de Biología, Universidad de Vigo, 36310-Vigo, Spain Received 14 February 2006; received in revised form 3 August 2006; accepted 14 August 2006 Available online 30 August 2006 Abstract We have shown previously that acute (1 to 6 h) and prolonged (1 to 5 days) exposure of rainbow trout to naphthalene resulted in decreased plasmatic cortisol and 17-β-estradiol levels. In order to elucidate the mechanisms through which naphthalene might disrupt endocrine regulation, the present study investigated whether brain monoaminergic neurotransmitters are altered by the action of this polycyclic aromatic hydro- carbon. In a first experiment, immature rainbow trout were injected with vegetable oil alone or containing naphthalene (10 and 50 mg/kg, i.p.), and sacrificed 1, 3 and 6 h after treatment. In a second experiment, slow-coconut oil implants alone or containing naphthalene (doses of 10 and 50 mg/kg) were i.p. located and fish sacrificed 1, 3 and 5 days after treatment. Levels of dopamine (DA), 3,4-dihydroxyphenylacetic acid (DOPAC), serotonin (5-HT), 5-hydroxyindoleacetic acid (5-HIAA) and noradrenaline (NA) were measured in several brain regions by HPLC. The results show that short-term naphthalene increases DA and 5-HT contents in hypothalamus and telencephalon, but differentially alter contents of the acid metabolites. Implants with naphthalene reduced DA content in hypothalamus and preoptic region but increased in telencephalon. 5-HT metabolism was decreased in hypothalamus, preoptic region, pituitary and brain stem after 3 to 6 days of treatment. In addition, the levels of NA were increased in hypothalamus and telencephalon after acute treatment and in hypothalamus and preoptic area after several days of exposure to naphthalene. These data suggest that brain neurotransmitter systems are sensitive to polycyclic aromatic hydrocarbons and could represent a target of the naphthalene-induced neuroendocrine disruption. © 2006 Elsevier Inc. All rights reserved. Keywords: Brain neurotransmitters; Dopamine; Monoamines; Naphthalene; Noradrenaline; Polycyclic aromatic hydrocarbons; Serotonin; Trout 1. Introduction Polycyclic aromatic hydrocarbons (PAHs) are the main toxic and persistent compounds present in most crude oils. As a result of the release of petroleum oils to the sea PAHs are now ubiquitous contaminants of aquatic ecosystems (Leonard and Hellou, 2001). Some of these chemicals have been demonstrat- ed to have mutagenic/carcinogenic (Hendricks et al., 1985; Malins et al., 1987), genotoxic (Pacheco and Santos, 1997, 2001) and cytotoxic (Schirmer et al., 1998) properties. In dif- ferent vertebrate species, environmental PAHs might act as potent endocrine disruptors (Cooper and Kavlock, 1997; Stahl- schmidt-Allner et al., 1997), in particular by binding to and activating nuclear estrogen receptors, resulting in alterations in the transcription rates of estrogen-regulated genes (Hutz et al., 1999; Barron et al., 2004a). In fish, PAHs have been shown to impair cellular steroidogenesis that occurs in gonad and in- terrenal gland, and also to influence systemic steroid regulation via the hypothalamus–pituitary–gonadal (HPG) and hypothal- amus–pituitary–interrenal (HPI) axis (Hontela et al., 1992, 1997; Vijayan et al., 1997; Wilson et al., 1998; Monteiro et al., 2000a,b; Evanson and Van der Kraak, 2001; Navas et al., 2004). In consequence, PAHs could interfere with reproductive activity and with the ability of fish to respond to stress (Al-Kindi et al., 2000, for a review). In addition, PAHs can affect several phy- siological functions in fish, such as energy metabolism, growth and immunity (Hontela et al., 1992; Pacheco and Santos, 2001; 2002; Teles et al., 2005; Tintos et al., in press). Many environmental pollutants have been shown to act as neurotoxicants (Tilson and Kodavanti, 1998; Bemis and Seegal, 1999) and potentially could impair the function of Comparative Biochemistry and Physiology, Part C 144 (2006) 173 – 183 www.elsevier.com/locate/cbpc ⁎ Corresponding author. Tel.: +34 986812386; fax: +34 986 812556. E-mail address: [email protected] (J.M. Míguez). 1532-0456/$ - see front matter © 2006 Elsevier Inc. All rights reserved. doi:10.1016/j.cbpc.2006.08.002
Transcript of Effects of acute and prolonged naphthalene exposure on brain monoaminergic neurotransmitters in...

gy, Part C 144 (2006) 173–183www.elsevier.com/locate/cbpc
Comparative Biochemistry and Physiolo
Effects of acute and prolonged naphthalene exposure on brainmonoaminergic neurotransmitters in rainbow trout (Oncorhynchus mykiss)
Manuel Gesto, Adrián Tintos, José L. Soengas, Jesús M. Míguez ⁎
Departamento de Biología Funcional y CC. de la Salud, Facultad de Biología, Universidad de Vigo, 36310-Vigo, Spain
Received 14 February 2006; received in revised form 3 August 2006; accepted 14 August 2006Available online 30 August 2006
Abstract
We have shown previously that acute (1 to 6 h) and prolonged (1 to 5 days) exposure of rainbow trout to naphthalene resulted in decreasedplasmatic cortisol and 17-β-estradiol levels. In order to elucidate the mechanisms through which naphthalene might disrupt endocrine regulation,the present study investigated whether brain monoaminergic neurotransmitters are altered by the action of this polycyclic aromatic hydro-carbon. In a first experiment, immature rainbow trout were injected with vegetable oil alone or containing naphthalene (10 and 50 mg/kg, i.p.),and sacrificed 1, 3 and 6 h after treatment. In a second experiment, slow-coconut oil implants alone or containing naphthalene (doses of 10 and50 mg/kg) were i.p. located and fish sacrificed 1, 3 and 5 days after treatment. Levels of dopamine (DA), 3,4-dihydroxyphenylacetic acid(DOPAC), serotonin (5-HT), 5-hydroxyindoleacetic acid (5-HIAA) and noradrenaline (NA) were measured in several brain regions by HPLC. Theresults show that short-term naphthalene increases DA and 5-HT contents in hypothalamus and telencephalon, but differentially alter contents ofthe acid metabolites. Implants with naphthalene reduced DA content in hypothalamus and preoptic region but increased in telencephalon. 5-HTmetabolism was decreased in hypothalamus, preoptic region, pituitary and brain stem after 3 to 6 days of treatment. In addition, the levels of NAwere increased in hypothalamus and telencephalon after acute treatment and in hypothalamus and preoptic area after several days of exposure tonaphthalene. These data suggest that brain neurotransmitter systems are sensitive to polycyclic aromatic hydrocarbons and could represent a targetof the naphthalene-induced neuroendocrine disruption.© 2006 Elsevier Inc. All rights reserved.
Keywords: Brain neurotransmitters; Dopamine; Monoamines; Naphthalene; Noradrenaline; Polycyclic aromatic hydrocarbons; Serotonin; Trout
1. Introduction
Polycyclic aromatic hydrocarbons (PAHs) are the main toxicand persistent compounds present in most crude oils. As a resultof the release of petroleum oils to the sea PAHs are nowubiquitous contaminants of aquatic ecosystems (Leonard andHellou, 2001). Some of these chemicals have been demonstrat-ed to have mutagenic/carcinogenic (Hendricks et al., 1985;Malins et al., 1987), genotoxic (Pacheco and Santos, 1997,2001) and cytotoxic (Schirmer et al., 1998) properties. In dif-ferent vertebrate species, environmental PAHs might act aspotent endocrine disruptors (Cooper and Kavlock, 1997; Stahl-schmidt-Allner et al., 1997), in particular by binding to andactivating nuclear estrogen receptors, resulting in alterations in
⁎ Corresponding author. Tel.: +34 986812386; fax: +34 986 812556.E-mail address: [email protected] (J.M. Míguez).
1532-0456/$ - see front matter © 2006 Elsevier Inc. All rights reserved.doi:10.1016/j.cbpc.2006.08.002
the transcription rates of estrogen-regulated genes (Hutz et al.,1999; Barron et al., 2004a). In fish, PAHs have been shown toimpair cellular steroidogenesis that occurs in gonad and in-terrenal gland, and also to influence systemic steroid regulationvia the hypothalamus–pituitary–gonadal (HPG) and hypothal-amus–pituitary–interrenal (HPI) axis (Hontela et al., 1992,1997; Vijayan et al., 1997; Wilson et al., 1998; Monteiro et al.,2000a,b; Evanson and Van der Kraak, 2001; Navas et al., 2004).In consequence, PAHs could interfere with reproductive activityand with the ability of fish to respond to stress (Al-Kindi et al.,2000, for a review). In addition, PAHs can affect several phy-siological functions in fish, such as energy metabolism, growthand immunity (Hontela et al., 1992; Pacheco and Santos, 2001;2002; Teles et al., 2005; Tintos et al., in press).
Many environmental pollutants have been shown to actas neurotoxicants (Tilson and Kodavanti, 1998; Bemis andSeegal, 1999) and potentially could impair the function of

174 M. Gesto et al. / Comparative Biochemistry and Physiology, Part C 144 (2006) 173–183
neurotransmitter systems and then influence pituitary hormonalsecretion. Despite of all adverse effects PAHs have on aquaticliving animals, there are very few studies about PAHs effects onnervous system. Indeed, PAHs have been shown to accumulatehighly in fish brain (Varanasi et al., 1979; Collier et al., 1980). Ina pioneer study, Fingerman and Short (1983) showed that naph-thalene (a two-ring PAH) alone or in combination with otherpollutants like benzo-a-pyrene (a five-ringed-PAH) and Aroclor1254 (a polychlorinated biphenyl, PCB, mixture) affected brainneurotransmitters levels in Channel catfish (Ictalurus puncta-tus). In addition, several other environmental pollutants havebeen shown to alter the levels of biogenic amines in discretebrain regions of channel catfish (Fingerman and Russell, 1980),trout (Rozados et al., 1991; Aldegunde et al., 1999) and theAtlantic croaker (Khan and Thomas, 1997, 2001; Khan et al.,2001).
Naphthalene and naphthalene derivatives like alkyl-naphtha-lenes are the most abundant PAHs present in most crude oils(Burns et al., 1997; Aas et al., 2000) and are frequently found inpolluted environments (Lee and Anderson, 2005). Exposure offish to naphthalene has been shown to induce a variety ofresponses at both cellular and systemic level in teleost fish(Thomas and Budiantara, 1995; Navas and Segner, 2000;Evanson and Van der Kraak, 2001). In trout, we have recentlyshown that naphthalene exposure induced a marked dose- andtime-dependent decrease in plasma levels of cortisol and 17β-estradiol, and also affected the energy metabolism in severalorgans, including brain (Tintos et al., in press). As an underlyingmechanism(s) mediating endocrine effects of naphthalene, it isplausible that it might interfere with brain neurotransmittersystems involved in regulating the activity of the hypothalam-ic–pituitary–hormonal axis. In this complementary report, wedetermine the effects of naphthalene on the content of dopamine(DA), noradrenaline (NA) and serotonin (5-HT), as well as theirrelative metabolites in discrete brain regions of rainbow trout(Oncorhynchus mykiss).
2. Materials and methods
2.1. Animals and experiments
Immature rainbow trout (O. mykiss, about 60 g) were ob-tained from a local hatchery and transferred to the laboratoriesat Faculty of Marine Sciences in Vigo, Spain. Fish were accli-mated for at least two weeks in well-aerated 150 l tanks, withan open water system. During the experiments, fish weremaintained under natural photoperiod (October 2004) andtemperature (18 °C). Fish were fed daily with 1% body masscommercial dry pellets (Dibaq-Diprotg S.A., Segovia, Spain).They were fasted 24 h before sampling. The experimentsdescribed comply with the guidelines of the European UnionCouncil (86/609/EU) for the use of laboratory animals.
In a first experiment, trout were caught by netting, anes-thetized with MS-222 (50 mg/l) buffered with sodium bicar-bonate, weighed, intraperitoneally injected (2 μl/g body mass,bm) with sunflower oil alone (control) or containing naphtha-lene (10 mg/kg and 50 mg/kg bm) and returned to their tanks. 24
fishes per group were used. Eight fish were sampled from eachgroup 1, 3 and 6 h after injection.
In a second experiment, anesthetized trout were weighed andintraperitoneally implanted (10 μl/g bm) with slow-releasecoconut oil implants alone (control) or containing naphthaleneat doses of 10 mg/kg bm and 50 mg/kg bm, and returned to theirtanks. Eight fish from each group were sampled 1, 3 and 5 daysafter injection.
On each experiment, non-injected fish were included toassess the possible vehicle effect. At each sampling time, fishwere quickly caught by netting, anesthetized with MS-222(50 mg/l) and sacrificed by decapitation. Brains were imme-diately dissected out and the following regions were sampled:telencephalon (excluding olfactory bulbs), hypothalamus,pituitary gland, preoptic area (including the optic tract) andbrain stem (mainly including the medulla oblongata). All tissueswere stored without delay at −80 °C until assay.
2.2. Analysis of monoamines and metabolites
The contents of NA, DA, 3,4-dihydroxyphenylacetic acid(DOPAC, a major DAmetabolite), 5-HTand 5-hydroxyindole-3-acetic-acid (5-HIAA, a major 5-HTmetabolite) were analyzed byhigh performance liquid chromatography with electrochemicaldetection (HPLC-EC) according to the method described byMíguez et al. (1999) with modifications. The HPLC system wasequipped with a Jasco PU-2080 Plus pump, a 5 μm analyticalcolumn (Phenomenex, Nucleosil C18, 150 mm length×4.6 mmdiameter) and a ESACoulochem II detector. The detection systemincluded a double analytical cell (M5011) with oxidationpotentials set at +40 mV (first electrode) and +340 mV (secondelectrode). The mobile phase was composed of 63.9 mMNaH2PO4 (Merck), 0.1 mM Na2EDTA (Sigma), 1.63 mM sodi-um 1-octanesulfonate (Fluka) and 14.9% (v/v) methanol. Its pHwas adjusted to 2.79 with ortho-phosphoric acid (Merck), andwas filtered (0.20 μm filter, Millipore, Bedford, USA) anddegasified at vacuum before use. Tissues were homogenized byultrasonic disruption in 0.1 ml (pituitary), 0.5 ml (hypothalamus,telencephalon, preoptic region) and 1 ml (brain stem) of mobilephase. After extracting an aliquot for protein measurement(Bradford, 1976), homogenates were centrifuged (16,000×g,10 min) and supernatants were diluted with mobile phase prior toHPLC analysis. Dilutions were 1:1 (supernatant/mobile phase)for telencephalon, medulla oblongata and preoptic region, and1:2 for hypothalamus. Pituitary supernatant was not diluted andwas injected directly into the HPLC system. The levels of NAwere not measured in the pituitary gland since they were belowthe detection limit of our technique and in mesencephalon due tointerferences in the analytical assay.
Analytical run time for each sample was 15 min at anisocratic flow rate of 1.1 ml/min at room temperature. Acqui-sition and integration of chromatograms were performed usingthe Biocrom XP computer-assisted software (Micron Analítica,Madrid, Spain). Quantification of sample peaks was estimatedcomparing peak areas with those of appropriate externalstandards. Detection limits for indoles examined were between0.5 and 3 pg per injection, with a signal-to-noise ratio of 2.

175M. Gesto et al. / Comparative Biochemistry and Physiology, Part C 144 (2006) 173–183
2.3. Data analyses
Data values are presented as mean plus standard error of themean (mean±S.E.M.) of each experimental group. The ratiosDOPAC/DA and 5-HIAA/5-HT contents were obtained by
Fig. 1. Levels of dopamine (DA) and 3,4-dihydroxyphenylacetic acid (DOPAC) andregion and brain stem in vehicle- (sunflower oil; white bars) and naphthalene-treatedjuvenile rainbow trout. The X-axis indicates the time (1, 3 and 6 h) after the intrape⁎: Significantly different (Pb0.05) from control group (Student–Newman–Keuls te
using the respective values of the amine and metabolite contentfor each sample and data were grouped according to thetreatment. The SPSSv14.0 statistical package was used to thestatistical analysis. The Levene test was used to ascertain thehomogeneity of variances between groups of data. The effects
values of DOPAC/DA ratio in hypothalamus, pituitary, telencephalon, preoptic(10 mg/kg body weight: light grey bars; 50 mg/kg body weight: dark grey bars)ritoneal injection. Each value represents the mean±S.E.M. of six to eight fish.st).
176 M. Gesto et al. / Comparative Biochemistry and Physiology, Part C 144 (2006) 173–183
of different doses of naphthalene and time of exposure wereanalyzed using a two-way ANOVA followed by Student–Newman–Keuls test. In cases of nonhomogenous variances forgroups, the Kruskal–Wallis nonparametric one-way ANOVAwas used followed by the Mann–Whitney U-test. Differencesbetween means were considered significant when Pb0.05.
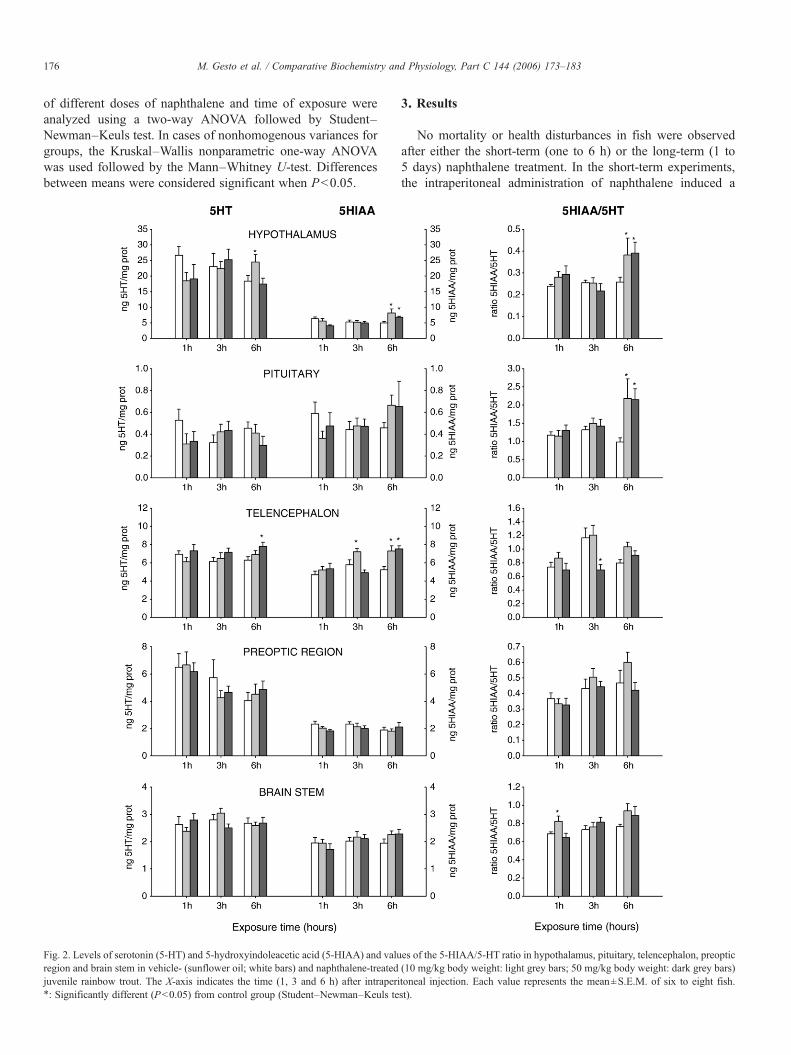
Fig. 2. Levels of serotonin (5-HT) and 5-hydroxyindoleacetic acid (5-HIAA) and valuregion and brain stem in vehicle- (sunflower oil; white bars) and naphthalene-treatedjuvenile rainbow trout. The X-axis indicates the time (1, 3 and 6 h) after intraperi⁎: Significantly different (Pb0.05) from control group (Student–Newman–Keuls te
3. Results
No mortality or health disturbances in fish were observedafter either the short-term (one to 6 h) or the long-term (1 to5 days) naphthalene treatment. In the short-term experiments,the intraperitoneal administration of naphthalene induced a
es of the 5-HIAA/5-HT ratio in hypothalamus, pituitary, telencephalon, preoptic(10 mg/kg body weight: light grey bars; 50 mg/kg body weight: dark grey bars)toneal injection. Each value represents the mean±S.E.M. of six to eight fish.st).

177M. Gesto et al. / Comparative Biochemistry and Physiology, Part C 144 (2006) 173–183
significant increase in the content of DA in the hypothalamus(10 mg/kg, 6 h after treatment) and telencephalon (50 mg/kg,3 h after treatment). In the last region, the highest naphthalenedose also increased DOPAC content, as well as decreasedDOPAC/DA ratio (Fig. 1). The levels of the DA metabolitewere also increased in preoptic region 1 h after the injection of
Fig. 3. Levels of dopamine (DA) and 3,4-dihydroxyphenylacetic acid (DOPAC) andregion and brain stem in vehicle- (coconut oil; white bars) and naphthalene-treated (juvenile rainbow trout. The X-axis indicates the time (1, 3 and 5 days) after intraperit⁎: Significantly different (Pb0.05) from control group (Student–Newman–Keuls te
naphthalene 10 mg/kg. At the hypothalamic level, naphthaleneincreased the content of 5-HT (10 mg/kg, 6 h after treatment)and 5-HIAA (10 and 50 mg/kg, 6 h after treatment), as well asthe 5-HIAA/5-HT ratio (both doses, 6 h). In addition, increasesin 5-HT and 5-HIAA content occurred in the telencephalon withthe higher dose of naphthalene (50 mg/kg, 6 h after treatment),
values of DOPAC/DA ratio in hypothalamus, pituitary, telencephalon, preoptic10 mg/kg body weight: light grey bars; 50 mg/kg body weight: dark grey bars)oneal implantation. Each value represents the mean±S.E.M. of six to eight fish.st).

178 M. Gesto et al. / Comparative Biochemistry and Physiology, Part C 144 (2006) 173–183
whereas 5-HIAA content increased with the two doses tested(10 mg/kg after 3 and 6 h of treatment; 50 mg/kg after 6 h oftreatment). The 5-HIAA/5-HT ratio decreased in the telenceph-alon 6 h after treatment, but it was increased in brain stem(50 mg/kg, 1 h) and pituitary gland (10 and 50 mg/kg, 6 h)(Fig. 2).
Fig. 4. Levels of serotonin (5-HT) and 5-hydroxyindoleacetic acid (5-HIAA) and thand brain stem of vehicle- (coconut oil; white bars) and naphthalene-treated (10 mg/krainbow trout. The X-axis indicates the time (1, 3 and 5 days) after intraperitone⁎: Significantly different (Pb0.05) from control group (Student–Newman–Keuls te
To investigate longer-term effects of naphthalene, fish wereimplanted with slow-release coconut oil implants containing thehydrocarbon or adequate vehicle. This procedure has been usedto administrate PAHs in fish species, like rainbow trout (Vijayanet al., 1997) and eel (Pacheco and Santos, 1998). After thenaphthalene implants, a decrease in DA content was found in
e ratio 5-HIAA/5-HT in hypothalamus, pituitary, telencephalon, preoptic regiong body weight: light grey bars; 50 mg/kg body weight: dark grey bars) juvenileal implantation. Each value represents the mean±S.E.M. of six to eight fish.st).

Table 1Noradrenaline (NA) levels in hypothalamus, telencephalon and preoptic region of naphthalene (NAP)-injected (h) or naphthalene-implanted (days) juvenile rainbowtrout, at different times after treatment
Time after treatment (h) Time after treatment (days)
1 3 6 1 3 5
HypothalamusControl 7.99±1.17 5.92±0.58 7.18±0.87 6.96±0.91 8.75±0.68 9.28±1.08NAP 10 mg/kg 6.69±0.52 7.70±0.97 9.36±0.91 6.45±1.16 9.23±1.20 8.22±0.46NAP 50 mg/kg 9.18±1.84 9.12±1.21 8.52±1.17 8.11±0.31 8.41±0.93 6.84±0.59
TelencephalonControl 8.39±0.77 10.43±1.38 9.46±0.45 6.08±0.42 6.62±0.63 8.51±0.88NAP 10 mg/kg 9.00±0.45 10.41±0.61 11.40±0.54⁎ 6.14±0.47 9.70±0.56⁎ 6.93±0.66NAP 50 mg/kg 8.16±0.75 12.57±0.63 11.58±0.87⁎ 9.64±1.50⁎ 9.99±1.01⁎ 7.84±0.70
Preoptic regionControl 1.22±0.38 0.93±0.31 0.76±0.16 0.52±0.14 1.03±0.35 0.79±0.08NAP 10 mg/kg 1.09±0.32 0.86±0.09 0.94±0.26 1.78±0.16⁎ 1.18±0.27 0.76±0.11NAP 50 mg/kg 1.14±0.31 1.26±0.27 1.10±0.29 1.28±0.34⁎ 0.98±0.11 0.59±0.12
Control group received vehicle treatment.Each value represents the mean±S.E.M. of six to eight fish.⁎ Significantly different (Pb0.05) from control group.
179M. Gesto et al. / Comparative Biochemistry and Physiology, Part C 144 (2006) 173–183
the hypothalamus (50 mg/kg, 5 days) and preoptic region(50 mg/kg, 3 days). This last area also exhibited a significantdecrease in DOPAC content in fish exposed for 5 days to bothdoses of naphthalene (Fig. 3). A different effect of the hydro-carbon was observed in the telencephalon since it inducedincreases in DA content (10 mg/kg at 5 days; 50 mg/kg at 1 and5 days of exposure) and decreases in DOPAC levels (10 and50 mg/kg, 1 day of exposure). The DOPAC/DA ratio wasdecreased by treatment in telencephalon (1 and 3 days of ex-posure), but increased in the hypothalamus (after 5 days) andthe pituitary (after 1 day of exposure).
Changes observed in indoleamines after chronic naphthaleneimplants attained all regions, except the telencephalon (Fig. 4).In the hypothalamus, the dose of 50 mg/kg induced a significantdecrease in the content of 5-HIAA (3 days of exposure) and 5-HT (5 days of exposure), which resulted in increases (3 days)and decreases (5 days) in the 5-HIAA/5-HT ratio. Also in thepreoptic region naphthalene treatment induced a decrease in thecontent 5-HT (10 mg/kg, 3 days) and its metabolite (50 mg/kg,5 days of exposure), whereas a drop in 5-HIAA level wasobserved in the pituitary (both naphthalene doses, 3 days ofexposure) and brain stem (50 mg/kg, 5 days of exposure). Inaddition, the 5-HIAA/5-HT ratio was reduced in brain stem withthe higher naphthalene dose (3 and 5 days of treatment).
Changes in NA levels after acute or chronic naphthalenetreatment are shown in Table 1. We observe a short-term stimula-tory effect of naphthalene on the content of NA in the hypothal-amus (50 mg/kg, 3 h) and telencephalon (10 and 50 mg/kg, 6 h).The long-term exposure to naphthalene also caused significantincreases in the levels of this neurotransmitter in the preopticregion (1 day of exposure) and telencephalon (1 and 3 days ofexposure), without significant changes in the hypothalamus.
4. Discussion
The present study describes for the first time detailed changesin catecholaminergic and indoleaminergic neurotransmitters in
discrete brain regions of immature rainbow trout exposed from1 h to several days to naphthalene. In general, changes induced bynaphthalene were very dependent upon the type of neurotrans-mitter and exhibited amarked brain regional selectivity, being themajor effects located in the hypothalamus, telencephalon andpreoptic area. The effects observedwere inconsistent over time ofexposure and the dose of naphthalene tested.
Various studies have shown that when fish were exposed tothese hydrocarbons brains accumulate relatively high levels ofnaphthalene and its methyl-substituted derivatives (Varanasiet al., 1979; Collier et al., 1980). However, the effects ofenvironmental pollutants on brain function were poorly studiedin fish. Decreased levels of 5-HT and DA and increased lev-els of their main metabolites were found in Atlantic croaker(Micropongias undulates L.) after exposure to PCBs-mixtureAroclor 1254 indicating that PCBs may affect brain neuro-transmitter activity (Fingerman and Russell, 1980; Khan andThomas, 1997, 2000). The pesticide lindane has also beenshown to change brain serotonergic metabolism in rainbowtrout O. mykiss (Rozados et al., 1991). With respect to naph-thalene, there is only a preliminary report of Fingerman andShort (1983) describing a variety of changes in whole brainmonoamine content of channel catfish which varied largely overtime of exposure. Our present results confirm the lack of aconsistent temporal trend in the effects of naphthalene, as wellas a clear dose–response following the two doses of naph-thalene examined (10 and 50 mg/kg). These data are in contrastwith other studies addressing dose- and time-related effects ofnaphthalene and related PAHs derivatives on several tissues andcellular processes, i.e., oxidative stress in brain and liver,hepatic intermediary metabolism and steroid secretion fromendocrine glands (Hutz et al., 1999; Evanson and Van derKraak, 2001; Bagchi et al., 2002; Tintos et al., in press).
The time course of the effect of chemical pollutants on brainneurochemistry is probably due to the rate of uptake of thepollutant and their effects on the different mechanisms regulatingthe content of individual neurotransmitters (i.e., synthesis,

180 M. Gesto et al. / Comparative Biochemistry and Physiology, Part C 144 (2006) 173–183
storage, uptake/release and degradation) in discrete regions of thenervous tissue. DAergic neuronal bodies in teleost fish brain arelocated mainly in hypothalamus, telencephalon and pretectalareas, and in medulla oblongata. Fibres of these neurons arewidely distributed in teleostean brain with the highest densityof DAergic terminals in ventral telencephalon, and preopticand hypothalamic regions (Hornby and Piekut, 1990; Meek,1995). 5-HTergic cells bodies are mainly confined to discretediencephalic areas and the mesencephalon, whereas 5-HT-containing fibres are particularly dense in the hypothalamus(Frankenhuis-van den Heuvel and Nieuwenhuys, 1984; Rodrí-guez-Gomez et al., 2000). In our study, the administration ofnaphthalene produced a short-term increase in the contents of DAand 5-HT in the hypothalamus, but inhibited the level of thesecompounds after a 5-day exposure. The increases in hypothalamicDA levels took place without any alteration in its main acidmetabolite, the DOPAC, suggesting that acute naphthalenetreatment increased synthesis rather than catabolism of thisneurotransmitter. However, the increases in 5-HT content in thehypothalamus were accompanied by parallel alterations in thelevels of 5-HIAA and also in the 5-HIAA/5-HT ratio, which isconsidered to be a more direct index of the neurotransmitteractivity than absolute values of the monoamines (Winberg et al.,1992, 1997). The 5-HIAA/5-HT ratio was increased after 6 h anddecreased after 5 days of naphthalene exposure, indicating thatshort-term naphthalene treatment increased both synthesis andturnover of the neurotransmitter, whereas both processes wereinhibited by a several-day exposure. This differential effect ofnaphthalene over time of exposure agrees with previous studiesshowing a reversal of the effects of naphthalene and otherchemicals pollutants on monoamine levels in whole brain of fish(Taylor and DiStefano, 1976; Fingerman and Russell, 1980;Fingerman and Short, 1983).
Savolainen (1977) has postulated that acute neurotoxiceffects of low molecular weight hydrocarbons in mammals aredue to accumulation of the parent hydrocarbons in the brain andthat chronic effects may be due to the formation of reactiveintermediates. Studies of Collier et al. (1980) found that severalhours after exposure to naphthalene, trout brain accumulatesnaphthalene at concentrations similar to those found at the sametime in liver, but brain capacity to take up, metabolize and flushthem out is very different from liver. In fact, trout brain has beenshown to display very little potential for converting aromatichydrocarbons to oxidized metabolites and a limited capacity toacquire metabolites from blood (Collier et al., 1980). Therefore,it seems that differences between short-term (hours) and long-term (days) effects of naphthalene on hypothalamic monaminesare not due to formation of intermediate naphthalene metabo-lites, but rather to changes in sensitivity of the neuronalmechanisms regulating the levels of aminergic compounds.
The effects of naphthalene on telencephalic DA metabolismwere similar after hours and days of treatment, with increasesin DA and decreases in DOPAC contents. In addition, theDOPAC/DA ratio was decreased by treatment, suggesting a lowutilization of the neurotransmitter which caused an elevation ofthe amine content. In the same way, treatment with naphthalenedecreased DA content in the preoptic region after 3 days and
DOPAC content after a 5-day exposure. Taking together theseresults and those obtained the hypothalamus, it seems as thatnaphthalene exposure induced a predominant low DAergicactivity in diencephalic (hypothalamus and preoptic area) andtelencephalic regions of trout brain, which have high density ofdopaminergic somas and terminals.
Similarly to that found for DA, the content of 5-HT in thehypothalamus was increased by short-term treatment and de-creased after long-term exposure to naphthalene. However,differentially to DA, changes in 5-HT content were accompa-nied by similar effects on the levels of its main acid metabolite,the 5-HIAA. This suggests that several hours of exposureto naphthalene increases 5-HT synthesis and turnover whilethe contrary takes place after several days of exposure. In-creased 5-HT and 5-HIAA contents were also found intelencephalon after short-term, but not long-term, naphthaleneexposure. In addition, in the preoptic region implants ofnaphthalene inhibited 5-HT and 5-HIAA contents respectivelyat 3 and 5 days of exposure to the hydrocarbon. These resultspoint to a predominant rise of 5-HT turnover in diencephalicand telencephalic regions after acute naphthalene treatmentand a decrease after an exposure to the hydrocarbon for severaldays.
There are immunohistochemical studies in teleosts thatconfirmed the presence of fibres containing monoamines andmonoamine-synthesizing enzymes in the pituitary (Rodríguez-Gomez et al., 2000). These fibres originate likely in the ventralhypothalamus and belong to the neuronal complex involved inmodulating pituitary hormone secretion (Yu et al., 1997). In thepresent study, we detect very low levels of DA, 5-HT andrelated acid metabolites in trout pituitary, which agrees withprevious neurochemical studies (Saligaut et al., 1990; Hernán-dez-Rauda et al., 2000; Hernández-Rauda and Aldegunde,2002). The effect of naphthalene on pituitary monoaminesevidences inconsistent changes over time with increases in the5-HIAA/5-HT ratio at 6 h and 1 day, and decreases in 5-HIAAcontents 3 days after naphthalene treatment. These changesfollow a similar trend as those described for hypothalamicmonoamines at these specific times, suggesting that changesobserved in the hypothalamus are immediately translated intochanges in pituitary monoamine levels.
The mesencephalic brainstem that contains the raphe nucleiis particularly rich in neuronal serotonergic soma (Frankenhuis-van den Heuvel and Nieuwenhuys, 1984; Meek et al., 1989;Aldegunde et al., 2000). In this region, effects of naphthalenemodified 5-HT turnover, and changes followed a biphasic trendsince the 5-HIAA/5-HT increased at 1 h but decreased at 3 and6 days of naphthalene treatment. This decrease is explainedmainly by decreased 5-HIAA content more than alteration inthe levels of the amine, which suggests a lower transformationof 5-HT to 5-HIAAwithout apparent changes in 5-HTsynthesis.Thus, it seems that neural mechanisms mediating naphthaleneeffects on 5-HT are simpler in brain areas containing pre-dominantly neuronal 5-HT bodies (brain stem) than thosehaving both neuronal somas and terminals (i.e., hypothalamusand telencephalon), in which alterations in both synthesis andturnover of the neurotransmitter coexist.

181M. Gesto et al. / Comparative Biochemistry and Physiology, Part C 144 (2006) 173–183
A great deal of studies have demonstrated that the dopa-minergic and serotonergic systems of the telencephalon, pre-optic region and hypothalamus are involved in the control ofbehavior in fish, including feeding, swimming activity, stressresponse and social interactions, among others (Winberg et al.,1992, 1997; Overli et al., 1998; Höglund et al., 2005). TheDAergic and 5-HTergic neurons of the hypothalamic–pituitarycomplex are also implicated as a major component of the vasteneural regulation affecting pituitary hormone secretion. Aprevious study of our group has shown that both acute andprolonged treatment with naphthalene to immature troutinhibited drastically the levels of cortisol and 17β-estradiol inblood (Tintos et al., in press). The i.p. administration ofnaphthalene (10 mg/kg and 50 mg/kg) induced an acute dose-dependent decrease in plasma cortisol levels, which was evenmore evident after several days of exposure to naphthaleneimplants. In addition, short-term naphthalene induced aconsistent decrease in plasma 17β-estradiol levels, whilelong-term treatment was followed by a clear dose-dependentreduction in the levels of this steroid. The data of the presentstudy showing parallel naphthalene-induced effects in thehypothalamic aminergic neurotransmitters point to that someof these endocrine effects of naphthalene could be mediatedthrough the hypothalamic–pituitary DAergic and 5-HTergicsystems, which are major factors influencing the HPI and HPGaxis in fish (Winberg et al., 1997; Yu et al., 1997; Lepage et al.,2002; Höglund et al., 2002). However, there was not a clearcorrelation among the dose- and time-dependent effects ofnaphthalene on plasma hormonal levels and those observed inbrain neurotransmitters. This is consistent with the fact that amajor part of PAHs effects on circulating cortisol and 17β-estradiol might be mediated by directly acting the hydrocarbonon the interrenal glands and gonads, affecting either thesensitivity to regulation by pituitary hormones or the steroido-genesis process (Stahlschmidt-Allner et al., 1997; Vijayan et al.,1997; Wilson et al., 1998; Hutz et al., 1999; Khan et al., 2001;Khan and Thomas, 2001; Monteiro et al., 2000a,b; Evanson andVan der Kraak, 2001; Navas et al., 2004). In addition, it is wellknown that in teleost fish there is an important steroid feedbackon hypothalamo-hypophysial activity (Peter and Yu, 1983).Endogenous steroids participate in the up- and down-regulationof pituitary hormone secretion at least in part by exertingpositive and negative effect on brain inputs to the pituitary,including the neurotransmitters DA and 5-HT (Mommsen et al.,1999; Hernández-Rauda and Aldegunde, 2002; DiBattista et al.,2005). Considering that naphthalene-exposed trout displaysaltered levels of gonadal and interrenal steroids it is not unlikelythat the variable effect of naphthalene over time on brainneurotransmitters is due, at least in part, to disturbances in thesteroid feedback action on hypothalamic monoaminergicsystems. Whether this hormonal action on brain neurotransmit-ters might interfere with a local effect of the hydrocarbon inspecific brain centres remains to be elucidated.
Significant increases in the content of NA in the hypothal-amus and telencephalon occur after acute naphthalene treatmentto trout, while a similar effect was observed in telencephalonand preoptic region following implants. All these regions re-
ceive noradrenergic inputs from outer brain areas, mainly fromthe locus coeruleous complex of the mesencephalon (Meek,1995). The effect of naphthalene on NA content follows a welldifferent trend from those on DA in spite of the closed meta-bolic relationship between both monoamines. This suggeststhat different mechanisms are mediating the action of naphtha-lene on the metabolism of each catecholamine at the neuronalendings.
In summary, the present study shows that catecholaminergicand indoleaminergic systems in trout brain are very sensitive toacute and prolonged exposure to naphthalene and might be a cuefor understanding the disruption of endocrine status followingexposure to polycyclic aromatic hydrocarbons present in pol-luted environments. Recent research indicates that high doses ofPAHs are likely to induce acute toxicity through a narcosismechanism of action caused by hydrophobic chemical partition-ing into cell membranes and nervous tissues that result indisruption of central nervous system function (Barron et al.,2001, 2004b). Such a nonspecific action on neuronal membranesmay explain the widely spread changes in neurotransmittersaffecting different brain regions after exposure to naphthalene.In addition, the mechanism of neural toxicity of PAHs mayinvolve events which entail an oxidative stress and production ofreactive oxygen species (Bagchi et al., 2002), and several studieshave already demonstrated PAH-induced effects on intracellularcalcium in mammalian cells (Mounho et al., 1997; Tannheimeret al., 1999). Changes in calcium homeostasis have been alsosuggested to be involved in PCBs-induced alterations in brainneurotransmission (Tilson and Kodavanti, 1998). Future studiesshould determine whether a similar mechanism could mediatethe neuronal action of naphthalene and simultaneously estimatetheir implication as endocrine disruptor.
Acknowledgements
This work was partly supported by grants VEM2003-20062(Ministerio de Ciencia y Tecnologia and FEDER, Spain),PGIDT04PXIC31208PN (Xunta de Galicia, Spain) andAGL2004-8137-c04-03/ACU (Ministerio de Educacion yCiencia and FEDER, Spain). M. Gesto is recipient of a Xuntade Galicia predoctoral fellowship.
References
Aas, E., Baussant, T., Balk, L., Liewenborg, B., Andersen, O.K., 2000. PAHmetabolites in bile, cytochrome P4501A and DNA adducts as environmentalrisk parameters for chronic oil exposure: a laboratory experiment withAtlantic cod. Aquat. Toxicol. 51, 241–258.
Al-Kindi, A.Y.A., Brown, J.A., Waring, C.P., 2000. Endocrine, physiologicaland histopathological responses of fish and their larvae to stress withemphasis on exposure to crude oil and various petroleum hydrocarbons.Science and Technology, Special Review, pp. 1–30.
Aldegunde, M., Soengas, J.L., Rubial, C., Andrés, M.D., 1999. Effects ofchronic exposure to γ-HCH (lindane) on brain serotonergic and gabaergicsystems, and serum cortisol and thyroxine levels of rainbow trout, Oncor-hynchus mykiss. Fish Physiol. Biochem. 20, 325–330.
Aldegunde, M., Soengas, J.L., Rozas, G., 2000. Acute effects of L-tryptophan ontryptophan hydroxylation rate in brain regions (hypothalamus and medulla)of rainbow trout (Oncorhynchus mykiss). J. Exp. Zool. 286, 131–135.

182 M. Gesto et al. / Comparative Biochemistry and Physiology, Part C 144 (2006) 173–183
Bagchi, D., Balmoori, J., Bagchi, M., Ye, X.M., Williams, C.B., Stohs, S.J.,2002. Comparative effects of TCDD, endrin, naphthalene and chromium(VI) on oxidative stress and tissue damage in the liver and brain tissues ofmice. Toxicology 175, 73–82.
Barron, M.G., Hansen, J., Lipton, J., 2001. Association between contaminanttissue residues and adverse effects in aquatic organisms. Rev. Environ.Contam. Toxicol. 173, 1–37.
Barron, M.G., Heintz, R., Rice, S.D., 2004a. Relative potency of PAHs andheterocycles as aryl hydrocarbon receptor agonists in fish. Mar. Environ.Res. 58, 95–100.
Barron, M.G., Carls, M.G., Heintz, R., Rice, S.D., 2004b. Evaluation of fishearly life-stage toxicity models of chronic embryonic exposures to complexpolycyclic aromatic hydrocarbon mixtures. Toxicol. Sci. 78, 60–67.
Bemis, J.C., Seegal, R.F., 1999. Polychlorinated biphenyls and methylmercuryact synergistically to reduce rat brain dopamine content in vitro. Environ.Health Perspect. 107, 879–885.
Bradford, M.M., 1976. A rapid and sensitive method for the quantitation ofmicrogram quantities of protein utilizing the principle of protein-dyebinding. Anal. Biochem. 72, 248–254.
Burns, W.A., Mankiewicz, P.J., Bence, A.E., Page, D.S., Parker, K.R., 1997.A principal-component and least-squares method for allocating polycyclicaromatic hydrocarbons in sediment to multiple sources. Environ. Toxicol.Chem. 16, 1119–1131.
Collier, T.K., Krahn, M.M., Malins, D.C., 1980. The disposition of naphthaleneand its metabolites in the brain of rainbow trout (Salmo gairdneri). Environ.Res. 23, 35–41.
Cooper, R.L., Kavlock, R.J., 1997. Endocrine disruptors and reproductivedevelopment: a weight-of-evidence overview. J. Endocrinol. 152, 159–166.
DiBattista, J.D., Anisman, H., Whitehead, M., Gilmour, K.M., 2005. The effectsof cortisol administration on social status and brain monoaminergic activityin rainbow trout Oncorhynchus mykiss. J. Exp. Biol. 208, 2707–2718.
Evanson, M., Van der Kraak, G.J., 2001. Stimulatory effects of selected PAHson testosterone production in goldfish and rainbow trout and possiblemechanisms of action. Comp. Biochem. Physiol. C 130, 249–258.
Fingerman, S.W., Russell, L.C., 1980. Effects of the polychlorinated biphenylAroclor 1242 on locomotor activity and on the neurotransmitters dopamineand norepinephrine in the brain of the Gulf killifish, Fundulus grandis. Bull.Environ. Contam. Toxicol. 25, 682–687.
Fingerman, S.W., Short, E.C., 1983. Changes in neurotransmitter levels inchannel catfish after exposure to benzo(a)pyrene, naphthalene, and Aroclor-1254. Bull. Environ. Contam. Toxicol. 30, 147–151.
Frankenhuis-van den Heuvel, T.H.M., Nieuwenhuys, R., 1984. Distribution ofserotonin-inmunoreactivity in the diencephalon and mesencephalon of thetrout, Salmo gairdneri. Anat. Embryol. 169, 193–204.
Hendricks, J.D., Meyers, T.R., Shelton, D.W., Casteel, J.L., Bailey, G.S., 1985.Hepatocarcinogenicity of benzo(a)pyrene to rainbow trout by dietaryexposure and intraperitoneal injection. J. Natl. Cancer Inst. 74, 839–851.
Hernández-Rauda, R., Aldegunde, M., 2002. Effects of acute 17α-methyltes-tosterone, acute 17β-estradiol, and chronic 17α-methyltestosterone ondopamine, norepinephrine and serotonin levels in the pituitary, hypothal-amus and telencephalon of rainbow trout (Oncorhynchus mykiss). J. Comp.Physiol. B 172, 659–667.
Hernández-Rauda, R., Míguez, J.M., Ruibal, C., Aldegunde, M., 2000. Effectsof melatonin on dopamine metabolism in the hypothalamus and the pituitaryof the rainbow trout, Oncorhynchus mykiss. J. Exp. Zool. 287, 440–444.
Höglund, E., Balm, P.H.M., Winberg, S., 2002. Stimulatory and inhibitoryeffects of 5-HT1A receptors on adrenocorticotropic hormone and cortisolsecretion in a teleost fish, the Arctic charr (Salvelinus alpinus). Neurosci.Lett. 324, 193–196.
Höglund, E., Weltzien, F.A., Schjolden, J., Winberg, S., Ursin, H., Doving, K.B.,2005. Avoidance behavior and brain monoamines in fish. Brain Res. 1032,104–110.
Hontela, A., Rasmussen, J.B., Audet, C., Chevalier, G., 1992. Impaired cortisolstress response in fish from environments polluted by PAHs, PCBs, andmercury. Arch. Environ. Contam. Toxicol. 22, 278–283.
Hontela,A.,Daniel, C., Rasmussen, J.B., 1997. Structural and functional impairmentof the hypothalamo–pituitary–interrenal axis in fish exposed to bleached kraftmill effluent in the St Maurice River, Quebec. Ecotoxicology 6, 1–12.
Hornby, P.J., Piekut, D.T., 1990. Distribution of catecholamine-synthesizingenzymes in goldfish brains: presumptive dopamine and norepinephrineneuronal organization. Brain Behav. Evol. 35, 49–64.
Hutz, R.J., Wimpee, B.A.B., Dasmahapatra, A., Weber, D.N., Heimler, I.,Chaffin, C.L., 1999. Differential modulation by aromatic hydrocarbonreceptor agonist of circulating estradiol-17β and estrogen-receptor DNA-binding capability in female rainbow trout (Oncorhynchus mykiss). Zool.Sci. 16, 161–166.
Khan, I.A., Thomas, P., 1997. Aroclor 1254-induced alterations in hypothalamicmonoamine metabolism in the Atlantic croaker (Micropogonias undulatas):correlation with pituitary gonadotropin release. Neurotoxicology 18, 553–560.
Khan, I.A., Thomas, P., 2000. Lead and Aroclor 1254 disrupt reproductiveneuroendocrine function inAtlantic croacker. Mar. Environ. Res. 50, 119–123.
Khan, I.A., Thomas, P., 2001. Disruption of neuroendocrine control ofluteinizing hormone secretion by Aroclor 1254 involves inhibition ofhypothalamic tryptophan hydroxylase activity. Biol. Reprod. 64, 955–964.
Khan, I.A., Mathews, S., Okuzawa, K., Kagawa, H., Thomas, P., 2001. Alterationsin the GnRH-LH system in relation to gonadal stage and Aroclor 1254exposure in Atlantic croaker. Comp. Biochem. Physiol. B 129, 251–259.
Lee, R.F., Anderson, J.W., 2005. Significance of cytochrome P450 systemresponses and levels of bile fluorescent aromatic compounds in marinewildlife following oil spills. Mar. Pollut. Bull. 50, 705–723.
Leonard, J.D., Hellou, J., 2001. Separation and characterization of gall bladderbile metabolites from speckled trout, Salvelinus fontinalis, exposed toindividual polycyclic aromatic compounds. Environ. Toxicol. Chem. 20,618–623.
Lepage,O., Tottmar,O.,Winberg, S., 2002. Elevated dietary intake of L-tryptophancounteracts the stress-induced elevation of plasma cortisol in rainbow trout(Oncorhynchus mykiss). J. Exp. Biol. 205, 3679–3687.
Malins, D.C., McCain, B.B., Brown, D.W., Varanasi, U., Krahn, M.M., Myers,M.S., Chan, S.L., 1987. Sediment-associated contaminants and liverdiseases in bottom-dwelling fish. Hydrobiologia 149, 67–74.
Meek, J., 1995. Catecholamines in the brains of Osteichthyes. In: Smeets,W.J.A.J., Reiner, A. (Eds.), Phylogeny and Development of Catechol-amine Systems in the CNS of Vertebrates. Cambridge Univ. Press,London, pp. 49–76.
Meek, J., Joosten, H.W.J., Steinbusch, H.W.M., 1989. Distribution of dopamineimmunoreactivity in the brain of the mormyrid teleost Gnathonemuspetersii. J. Comp. Neurol. 281, 362–383.
Míguez, J.M., Aldegunde, M., Paz-Valñas, L., Sánchez-Barceló, E., 1999.Selective changes in the contents of noradrenaline, dopamine and serotoninin rat brain areas during aging. J. Neural Transm. 106, 1089–1098.
Mommsen, T.P., Vijayan, M.M., Moon, T.W., 1999. Cortisol in teleosts:dynamics, mechanisms of action, and metabolic regulation. Rev. Fish Biol.Fish. 9, 211–268.
Monteiro, P.R.R., Reis-Henriques, M.A., Coimbra, J., 2000a. Plasma steroidlevels in female flounder (Platichthys flesus) after chronic dietary exposureto single polycyclic aromatic hydrocarbons. Mar. Environ. Res. 49,453–467.
Monteiro, P.R.R., Reis-Henriques, M.A., Coimbra, J., 2000b. Polycyclicaromatic hydrocarbons inhibit in vitro ovarian steroidogenesis in theflounder (Platichthys flesus L.). Aquat. Toxicol. 48, 549–559.
Mounho, B.J., Davila, D.R., Burchiel, S.W., 1997. Characterization ofintracellular calcium responses produced by polycyclic aromatic hydro-carbons in surface marker-defined human peripheral blood mononuclearcells. Toxicol. Appl. Pharmacol. 145, 323–330.
Navas, J.M., Segner, H., 2000. Antiestrogenicity ofβ-naphthoflavone and PAHs incultured rainbow trout hepatocytes: evidence for a role of the arylhydrocarbonreceptor. Aquat. Toxicol. 51, 79–92.
Navas, J.M., Zanuy, S., Segner, H., Carrillo, M., 2004. Beta-naphthoflavonealters normal plasma levels of vitellogenin, 17β-estradiol and luteinizinghormone in sea bass broodstock. Aquat. Toxicol. 67, 337–345.
Overli, O., Winberg, S., Damsgard, B., Jobling, M., 1998. Food intake andspontaneous swimming activity inArctic charr (Salvelinus alpinus): role of brainserotonergic activity and social interactions. Can. J. Zool. 76, 1366–1370.
Pacheco, M., Santos, M.A., 1997. Induction of EROD activity and genotoxiceffects by polycyclic aromatic hydrocarbons and resin acids on the juvenileeel (Anguilla anguilla L.). Ecotoxicol. Environ. Saf. 38, 252–259.

183M. Gesto et al. / Comparative Biochemistry and Physiology, Part C 144 (2006) 173–183
Pacheco, M., Santos, M.A., 1998. Induction of liver EROD and erythrocyticnuclear abnormalities by cyclophosphamide and PAHs in Anguilla anguillaL. Ecotoxicol. Environ. Saf. 40, 71–76.
Pacheco, M., Santos, M.A., 2001. Biotransformation, endocrine, and geneticresponses of Anguilla anguilla L. to petroleum distillate products andenvironmentally contaminated waters. Ecotoxicol. Environ. Saf. 49, 64–75.
Pacheco, M., Santos, M.A., 2002. Naphthalene and β-naphthoflavone effects onAnguilla anguilla L. hepatic metabolism and erythrocytic nuclear abnor-malities. Environ. Int. 28, 285–293.
Peter, R.E., Yu, K.L., 1983. In: Hoar, W.S., Randall, D.J., Donaldson, E.M.(Eds.), The brain and neurohormones in teleost reproduction. FishPhysiology, vol. 9. Academic Press, New York, pp. 97–135.
Rodríguez-Gomez, F.J., Rendón-Unceta, C., Sarasquete, C., Muñoz-Cueto, J.A.,2000. Distribution of serotonin in the brain of the senegalese sole, Soleasenegalensis: an immunohistochemical study. J. Chem. Neuroanat. 18,103–115.
Rozados, M.V., Andres, M.D., Aldegunde, M.A., 1991. Preliminary studies onthe acute effect of lindane (γ-HCH) on brain serotoninergic system inrainbow trout Oncorhynchus mykiss. Aquat. Toxicol. 19, 33–40.
Saligaut, C., Bailhache, T., Salbert, G., Jego, P., 1990. Dynamic characteristicsof serotonin and dopamine metabolism in the rainbow trout brain: a regionalstudy using liquid chromatography with electrochemical detection. FishPhysiol. Biochem. 8, 199–205.
Savolainen, H., 1977. Some aspects of mechanisms by which industrial solventsproduce neurotic effects. Chem. Biol. Interact. 18, 1–10.
Schirmer, K., Dixon, D.G., Greenberg, B.M., Bols, N.C., 1998. Ability of 16priority PAHs to be directly cytotoxic to a cell line from the rainbow troutgill. Toxicology 127, 129–141.
Stahlschmidt-Allner, P., Allner, B., Römbke, J., Knacker, T., 1997. Endocrinedisrupters in the aquatic environment. Environ. Sci. Pollut. Res. Int. 4,155–162.
Tannheimer, S.L., Lauer, F.T., Lane, J., Burchiel, S.W., 1999. Factorsinfluencing elevation of intracellular Ca2+ in the MCF-10A human
mammary epithelial cell line by carcinogenic polycyclic aromatic hydro-carbons. Mol. Carcinog. 25, 48–54.
Taylor, L.L., DiStefano, V., 1976. Effects of methylmercury on brain biogenicamines in developing rat pup. Toxicol. Appl. Pharmacol. 38, 489–497.
Teles,M.,Oliveira,M., Pacheco,M., Santos,M.A., 2005. Endocrine andmetabolicchanges in Anguilla anguilla L. following exposure to β-naphthoflavone—a microsomal enzyme inducer. Environ. Int. 31, 99–104.
Thomas, P., Budiantara, L., 1995. Reproductive life-history stages sensitive tooil and naphthalene in Atlantic croaker. Mar. Environ. Res. 39, 147–150.
Tilson, H.A., Kodavanti, P.R.S., 1998. The neurotoxicity of polychlorinatedbiphenyls. Neurotoxicology 19, 517–525.
Tintos, A., Gesto, M., Míguez, J.M., Soengas, J.L., in press. Naphthalenetreatment alters liver intermediary metabolism and levels of steroidhormones in plasma of rainbow trout (Oncorhynchus mykiss). Ecotoxicol.Environ. Saf.
Varanasi, U., Gmur, D.J., Treseler, P.A., 1979. Influence of time and mode ofexposure on biotransformation of naphthalene by juvenile starry flounder(Platichthys stellatus) and rock sole (Lepidopsetta bilineata). Arch. Environ.Contam. Toxicol. 8, 673–692.
Vijayan, M.M., Feist, G., Otto, D.M.E., Schreck, C.B., Moon, T.W., 1997.3,3′,4,4′-Tetrachlorobiphenyl affects cortisol dynamics and hepatic functionin rainbow trout. Aquat. Toxicol. 37, 87–98.
Wilson, J.M., Vijayan, M.M., Kennedy, C.J., Iwama, G.K., 1998. β-Naphtho-flavone abolishes interrenal sensitivity to ACTH stimulation in rainbow trout.J. Endocrinol. 157, 63–70.
Winberg, S., Nilsson, G.E., Olsen, K.H., 1992. Changes in brain serotonergicactivity during hierarchical behavior in arctic charr (Salvelinus alpinus L)are socially induced. J. Comp. Physiol. A 170, 93–99.
Winberg, S., Winberg, Y., Fernald, R.D., 1997. Effect of social rank on brainmonoaminergic activity in a cichlid fish. Brain Behav. Evol. 49, 230–236.
Yu, K.L., Lin, X.W., da Cunha Bastos, J., Peter, R.E., 1997. Neural regulation ofGnRH in teleost fishes. In: Parhar, I.S., Sakuma, Y. (Eds.), GnRH Neurons:Gene to Behavior. Brain Shuppan, Tokyo, pp. 277–312.




















