
A linkage map for the B-genome of Arachis (Fabaceae) and its synteny to the A-genome

ORIGINAL ARTICLE
Photosynthetic characteristics of groundnut (Arachis hypogaea L.)
under water deficit stress
K. A. Kalariya • A. L. Singh • K. Chakraborty •
P. V. Zala • C. B. Patel
Received: 25 January 2012 / Accepted: 13 March 2013
� Indian Society for Plant Physiology 2013
Abstract In a field experiment during summer season,
the chlorophyll fluorescence and net photosynthetic rate
were studied in six Spanish groundnut cultivars under
water deficit (WD) condition between beginning bloom to
beginning seed (WD I) and beginning seed to beginning
maturity (WD II), by withholding irrigation 31–62 DAS
and 62–87 DAS, respectively. The severity of drought was
so that the soil moisture content of the stressed field
depleted to 9.4 and 6.2 % at 0–15 cm soil depth in WD I
and WD II at 62 and 87 DAS, respectively, as against 17
and 19 % in control. The mean RWC of leaves decreased
from 92 in control to 88 in WDI and from 91 in control to
84 in WD II with the least decrease in ICGS 44 under WD I
and in TAG 24 under WD II. The WD condition signifi-
cantly increased non-photochemical quenching but
decreased the maximum quantum yield of PS II (Fv/Fm)
from 0.81 in control to 0.77 under WD I condition which
was again resumed to 0.80 after 48 h of withdrawal of
stress. The rate of photosynthesis which was 29 and
36 lmol m-2 s-1 in well irrigated plots decreased to 26
and 28 lmol m-2 s-1 with a deduction of 11 and 30 %
under WD I and WD II, respectively. Cultivar TAG 24
showed better stress recovering capacity with high photo-
synthesis under both control as well as WD condition
whereas, data on chlorophyll fluorescence parameters
showed that cultivar ICGS 44 was least affected to damage
via photoinhibitory action.
Keywords Chlorophyll fluorescence � Groundnut �
Leaf temperature � Photosynthesis � Water deficit
stress
Introduction
Groundnut (Arachis hypogaea L.) is an important food
legume and oilseed crop of the world grown on about 24
million ha of land in about 120 countries mostly in tropics
and subtropics of arid and semi-arid regions where the
availability of water is a major constraint on yield and crop
frequently suffers drought of various spells and intensities.
As a result, the groundnut productivity is less than
1,000 kg ha-1 in more than 50 % of groundnut growing
countries. In India, the groundnut is cultivated on about 6
million ha with productivity of about 1,400 kg ha-1. Of all
the abiotic stresses, drought is most damaging one in
groundnut and it has a broad physiological spectrum
affecting many metabolic processes and it is difficult to
assess the contribution of individual processes to the final
damage done to the plants. Drought affects the groundnut
plants at all the stages, with varied sensitivity and crop
cultivars that show differential responses (Singh 2004,
2011; Nautiyal et al. 2012).
The photosynthetic rate (PN) of leaves decreases as
relative water content (RWC) and water potential (W)
decreases and limitation of net photosynthetic rate in low
moisture stressed plant is mainly through stomatal closure
(Cornic and Massacci 1996; Singh 2004) and/or by meta-
bolic impairment. The relative magnitude of stomatal and
non-stomatal factors limiting photosynthesis depends on
the severity of drought. Photosynthesis, the most important
process influencing crop production, is inhibited by
drought stress and high PN is one of the most important
K. A. Kalariya (&) � A. L. Singh � K. Chakraborty �
P. V. Zala � C. B. Patel
Directorate of Groundnut Research, PB 5, Junagadh 362 001,
Gujarat, India
e-mail: [email protected]
123
Ind J Plant Physiol.
DOI 10.1007/s40502-013-0027-x

breeding strategies for crop improvement (Richards 2000).
Chlorophyll fluorescence is a very sensitive probe of
physiological status of leaves and plant performance in a
wide range of situations and successfully applied in crop
improvement programs by carefully selecting and analys-
ing fluorescence parameters (Daniele et al. 2006). Though
the initial phase of drought stress in plants due to stomatal
limitation of photosynthesis is not perceived by the fluo-
rescence parameters that can only monitor the efficiency of
primary photochemical processes of photosynthesis. Araus
et al. (1998) proposed chlorophyll fluorescence as a
selection criteria for improvement of grain yield of durum
wheat in Mediterranean conditions, where frequent drought
accompanied by high temperatures is common. Since then
chlorophyll fluorescence continues in mainstay in studies
of photosynthetic regulation and plant responses to the
environment due to its sensitivity, convenience, and non-
intrusive quality. In recent years, the chlorophyll fluores-
cence technique has become ubiquitous in plant eco-
physiological studies.
Therefore, a study was aimed at unravelling the changes
in chlorophyll fluorescence and net photosynthetic rate
under water deficit condition at different phenophases in
groundnut cultivars of different maturity period.
Materials and methods
A field experiment was conducted during summer 2011
(January–June) using six Spanish groundnut varieties at the
research farm of Directorate of Groundnut Research,
Junagadh, Gujarat (lat 21.0 310N, Long 70.0360E), India in
Vertic Ustochrept (pH 7.5) soil. The experiment was laid in
a split plot design taking three irrigation treatments viz.
control, water deficit (WDI) and WD II in main plot, and
six varieties in sub plots. The net plot size was 4 9 3 m
with 9 rows per plot at 45 cm row to row and 10 cm plant
to plant spacing. To synchronize phenophases the
groundnut cultivars SG 99, ICGS 44 and ICGV 86031 of
130 days duration were sown during last week of January
and cultivars TAG 24, AK 159 and DRG 1 of 120 days
duration were sown during first week of February.
The water deficit condition was created by with-holding
irrigation from beginning bloom to beginning of seed i.e.
31–61 DAS (WD I) and from beginning seed to beginning
maturity, i.e. 62–87 DAS (WD II) while adequate moisture
was maintained in the control. The RWC, chlorophyll
fluorescence, photosynthesis and leaf temperature were
recorded from the third leaf at 62 and 87 DAS in WD I,
WD II, respectively and in control and 48 h after relief of
water stress. Maximum efficiency of PSII (Fv/Fm) of the
dark adapted leaves were recorded after 30 min dark
adaptation by leaf clips and actual quantum yield of PSII
and photosynthesis (at 1,650 PPFD artificial light) were
recorded between 08:00 and 10:00 h by LI-COR 6400,
Portable Photosynthesis system (LI-COR Inc. Lincon, NE,
USA) with modulated fluorescence measurement. This
system also simultaneously measures the net rate of pho-
tosynthesis (PN), stomatal conductance (gs) leaf tempera-
ture (Tleaf) and transpiration rate (E) etc. The RWC was
calculated using formula: RWC = 100*((FW - DW)/
(TW - DW)), Where, FW is fresh weight, TW is turgid
weight and DW is dry weight.
Results and discussion
Soil water plant relations
The soil moisture content of the field at 0–15 cm soil depth
was 17.0 and 19.5 % in control plots which, by with-holding
irrigation decreased to 9.4 and 6.2 % under water deficit
condition at the end of WD I and WD II, respectively.
However, the soil moisture content in 15–30 cm soil depth
was 18.0 and 19.0 % in control plots which decreased to 10.1
and 9.4 % in WD I and WD II, respectively. The soil mois-
ture content depleted by 45 and 44 % in WD I and 68 and
51 % in WD II as compared with control at 0–15 cm and
15–30 cm depth, respectively. The RWC of groundnut
leaves which was 93 and 91 % in control decreased to 88 and
84 % by imposition of water deficit at the end of WD I and
WD II, respectively with the highest value in SG 99
(Table 1). Stomatal conductance (gs) and transpiration rate
also decreased under water deficit condition (Table 1). The
gs in control at 62 and 87 DAS was 0.40 and 0.67 m s-1
which decreased to 0.32 and 0.30 m s-1 in WD I andWD II,
respectively which after relieving of stress by irrigation
further increased gs to 0.35 and 0.40 mol m-2 s-1, respec-
tively. Highest stomatal conductance was found in cultivar
ICGS 44 under WD I and in DRG 1 under WD II. The
varieties with minimum transpiration were SG 99, AK 159
and DRG 1 because of corresponding reduction in stomatal
conductance whereas, ICGV 86031 and TAG 24 showed
higher transpiration rates.
The RWC, leaf water potential, stomatal resistance, rate
of transpiration, leaf temperature and canopy temperature
are important parameters that influence water relations in
groundnut (Nautiyal et al. 1995, 2002, 2012; Singh 2004).
The RWC of leaves is reported to be higher during the
initial stages of development and declines as the dry matter
accumulates and leaf matures (Jain et al. 1997). In non-
stressed groundnut plants the RWC in leaves ranges from
85 to 90 %, while under drought stress it goes down
heavily (Babu and Rao 1983). Daniele et al. (2006) found
that in groundnut, the genotypic discrimination of RWC
trait depends on the water regime and genetic background
Ind J Plant Physiol.
123

and this trait has no value as selection criteria but serves to
characterize finely the water status of plants. Water uptake
was reported to be maximised by increasing root depth.
Allen et al. (1976) measured soil water extraction during
water stress and found that groundnut roots effectively
extracted soil water to a depth of at least 180 cm in fine
sand soil as roots in lower depths continue to grow deeper
even though vegetative growth appears to stop.
Simmonds and Ong (1987) found that the cultivar Robut
33–1 extracted water more rapidly from deeper layers
when grown at high vapour pressure deficits than grown in
more humid air. Devries et al. (1989) reported that cultivar
Florunner had higher root length density in deeper layers
(60–150 cm) during drought periods. All these traits con-
tribute to groundnut’s ability to avoid drought stress in this
study. Drought stressed plants transpire less than unstressed
plants, as a result decreased rate of transpiration was
observed in this study. The same pattern was also observed
for stomatal conductance. Subramaniam and Maheswari
(1990) reported that leaf water potential, transpiration rate
and photosynthetic rate decreased progressively with
increasing duration of water stress in groundnut.
Maximum efficiency of PS II (Fv/Fm) and quantum
yield of PS II (UPSII)
Maximum efficiency of PS II (Fv/Fm) decreased signifi-
cantly from 0.81 in control to 0.77 under WD condition and
again reached to 0.80 after 48 h of withdrawal of stress
showing a good stress recovery indication under WD I
(Table 2). The recovery was highest in cultivar ICGS 44
and AK 159. Under WD II also the Fv/Fm decreased from
Table 1 Leaf relative water content (RWC), stomatal conductance (gs), transpiration rate and leaf temperature (Tleaf) in groundnut varieties at
62 DAS in WD I and 87 DAS in WD II as affected by water deficit conditions during summer 2011
Varieties Relative leaf water content (%) gs (m s-1)
62 DAS 87 DAS 62 DAS 87 DAS
Control WDI Mean Control WDII Mean Control WDI SR Mean Control WDII SR Mean
AK 159 91.3 87.7 89.5 91.0 83.9 87.5 0.41 0.30 0.36 0.35 0.68 0.25 0.41 0.45
DRG 1 91.8 86.4 89.1 92.4 82.6 87.5 0.42 0.28 0.41 0.37 0.66 0.43 0.42 0.50
TAG 24 91.2 88.1 89.6 90.5 83.1 86.8 0.41 0.34 0.36 0.37 0.62 0.38 0.50 0.50
ICGS 44 90.9 90.3 90.6 92.0 83.9 87.9 0.42 0.38 0.34 0.38 0.62 0.22 0.40 0.41
ICGV 86031 91.7 85.8 88.7 91.2 84.1 87.6 0.39 0.31 0.32 0.34 0.70 0.24 0.34 0.43
SG 99 94.2 91.8 93.0 90.8 86.5 88.6 0.38 0.31 0.35 0.34 0.72 0.29 0.33 0.45
Mean 91.8 88.4 90.1 91.3 84.0 87.7 0.40 0.32 0.35 0.36 0.67 0.30 0.40 0.46
CD (p = 0.05)
Water deficit 1.7 2.09 0.02 0.09
Varieties 2.4 NS NS NS
Interactions NS NS 0.06 0.12
Varieties Transpiration rate (mmol m-2 s-1) Tleaf (�C)
62 DAS 87 DAS 62 DAS 87 DAS
Control WDI SR Mean Control WDII SR Mean Control WDI SR Mean Control WDII SR Mean
AK 159 10.6 5.6 10.1 8.8 9.9 5.4 7.9 7.7 32.1 34.7 31.9 32.9 33.0 35.5 34.0 34.2
DRG 1 10.6 5.8 11.2 9.2 10.5 7.4 7.4 8.4 32.4 37.9 32.8 34.4 32.7 38.7 33.6 35.0
TAG 24 10.1 7.5 9.7 9.1 11.0 7.2 8.1 8.8 31.2 37.0 32.3 33.5 31.9 37.8 34.3 34.6
ICGS 44 10.4 6.0 9.2 8.5 10.6 5.7 7.9 8.1 32.4 36.9 32.8 34.0 32.9 37.2 34.1 34.7
ICGV 86031 10.1 7.8 9.8 9.3 12.3 6.6 7.1 8.7 32.9 36.6 32.8 34.1 33.2 38.5 34.6 35.5
SG 99 10.5 5.2 10.1 8.6 9.7 6.1 7.8 7.8 32.1 36.4 32.2 33.6 32.3 37.2 33.8 34.4
Mean 10.4 6.3 10.0 8.9 10.9 6.4 7.7 8.3 32.2 36.6 32.5 33.7 32.7 37.5 34.1 34.73
CD (p = 0.05)
Water deficit 0.74 1.60 1.4 1.3
Varieties NS NS NS NS
Interactions 1.57 1.14 NS NS
SR represents 48 h after stress relief
Ind J Plant Physiol.
123

0.84 in control to 0.80 in WD condition which recovered
after 48 h of withdrawal of stress. Significant varietal dif-
ference for UPSII with the high value in TAG 24, ICGS 44
and SG 99 was observed under WD I. Also, there was
significant interaction of varieties and treatment for UPSII
where the reduction in these parameters was lower in TAG
24, ICGS 44 and SG 99 than other varieties under WD I.
This was also reflected in terms of higher rate of photo-
synthesis in these varieties under WD I. After 48 h of stress
recovery, increase in the proportion of absorbed energy
utilised for photochemistry (UPSII) with the highest value in
cultivar TAG 24 and SG 99 showed the better stress
recovering capacity. Groundnut has the ability to recover
from prolonged desiccation indicative of moisture stress
endurance (Babu and Rao 1983). Bogalea et al. (2011)
showed that the F0 has increased while variable fluores-
cence (Fv), maximum fluorescence (Fm) and optimum
quantum yield fluorescence (Fv/Fm) decreased under water
deficit condition in durum wheat. The increase in F0 is a
characteristic of PSII inactivation whereas a decline in Fv
under stress may indicate the increase in non-photochem-
ical quenching process at or close to reaction centres.
Similarly, the increase in F0 and the decrease in Fm under
water deficit with concomitant decrease in Fv/Fm, indicate
the occurrence of chronic photo inhibition due to photo
inactivation of PSII centres (Baker and Horton 1987).
Leaf temperature (Tleaf) and non-photochemical
quenching (NPQ)
There was increase in leaf temperature by 4.4 and 4.8 �C
due to WD I and WD II, respectively compared to control
but, there was no significant difference between Tleaf
among groundnut cultivars (Table 1). The correlation
(Table 3) between transpiration and Tleaf being r = -0.87
and -0.71 under WD I and WD II, respectively shows the
cooling effect of leaf under higher transpiration. The NPQ
also significantly increased under water deficit condition.
The correlation between Tleaf and NPQ was r = 0.80 under
WD I and r = 0.71 under WD II. Likewise increase in
UPSII with a corresponding decrease in NPQ have been
observed for stress recovered plant also indicates that after
stress recovery, energy was not utilised for heat dissipation,
but for photochemistry. The highest decrease in NPQ in
TAG 24 variety again confirms its better stress recovering
capacity. Most of the plants adapt themselves to water
stress by dissipating the excess excitation energy thermally
with the down regulation of PSII activity to protect pho-
tosynthetic apparatus from photo-damaging effect under
water stress often coinciding with high leaf temperature
(Bilger and Bjorkman 1990). Shahenshah and Isoda (2010)
reported that under green-house condition, Tleaf in
groundnut was increased due to water deficit stress. The
increase in volume of NPQ coupled with a negative effect
on rate of photosynthesis (r = -0.78 and -0.80 at WD I
and WD II, respectively) confirms that the incident energy
was diversified towards heat dissipation instead of photo-
chemistry under water deficit stress (Table 3). Excess light
is harmlessly dissipated in the antennae complexes of PSII
as heat is involved in a process of xanthophyll cycle, a
reversible conversion of the carotenoid violaxanthin (V) to
zeaxanthin (Z) via antherozanthin (A) as well as low thy-
lakoid pH. The xanthophyll cycle pigments Z and A are
formed from V during conditions of excess light and are
both thought to be involved in the photo-protective dissi-
pation process (Bjorkman and Demmig-Adams 1994).
At the leaf level, the dissipation of the excitation energy
through processes other than photosynthetic carbon
metabolism is an important defence mechanism under
water stress conditions and is accompanied by down-reg-
ulation of photochemistry in the long term carbon metab-
olism (Chaves et al. 2002). Also the energy dissipation in
closed stomata can occur via ATP and NADPH, which are
used for other metabolic processes, and are obviously
important mechanisms of tolerance and protection against
water stress and photo-oxidative damage. Daniele et al.
(2006) reported that fluorescence parameters showed
strong sensitivity to drought and good genotypic discrim-
ination in groundnut genotypes.
Net photosynthetic rate (PN)
The rate of photosynthesis significantly decreased due to
water deficit condition at both the stress periods (Table 2).
The mean PN was 29 and 36 lmol m-2 s-1 which due to
water stress decreased to 26 and 28 lmol m-2 s-1 at WD I
and WD II, respectively. However, the relief of stress
further increased PN to 28 and 29 lmol m-2 s-1, respec-
tively. The decrease in PN compared to control under WD I
and WD II by 11 and 30 %, respectively showed that the
growth period as defined by Boote (1982) between R5 and
R7 is more susceptible to water deficit condition for carbon
partitioning. The highest decrease was in cultivar DRG 1
under WD I, but interestingly the least decrease also in the
same cultivar under WD II shows stage-specificity in tol-
erance and susceptibility to water deficit condition. Canopy
photosynthesis is reduced by moisture stress mainly due to
reduction in stomatal conductance and leaf area, and as
moisture stress increases, stomata start closing as a
mechanism to reduce transpiration. As a consequence, the
entry of carbon dioxide is also reduced (Singh 2011). The
decrease in conductance of mesophyll cells due to moisture
stress results in low conductance of carbon dioxide and a
reduction in photosynthesis. During pod development
stage, both the vegetative and reproductive sinks
operate concurrently in groundnut because of relatively
Ind J Plant Physiol.
123
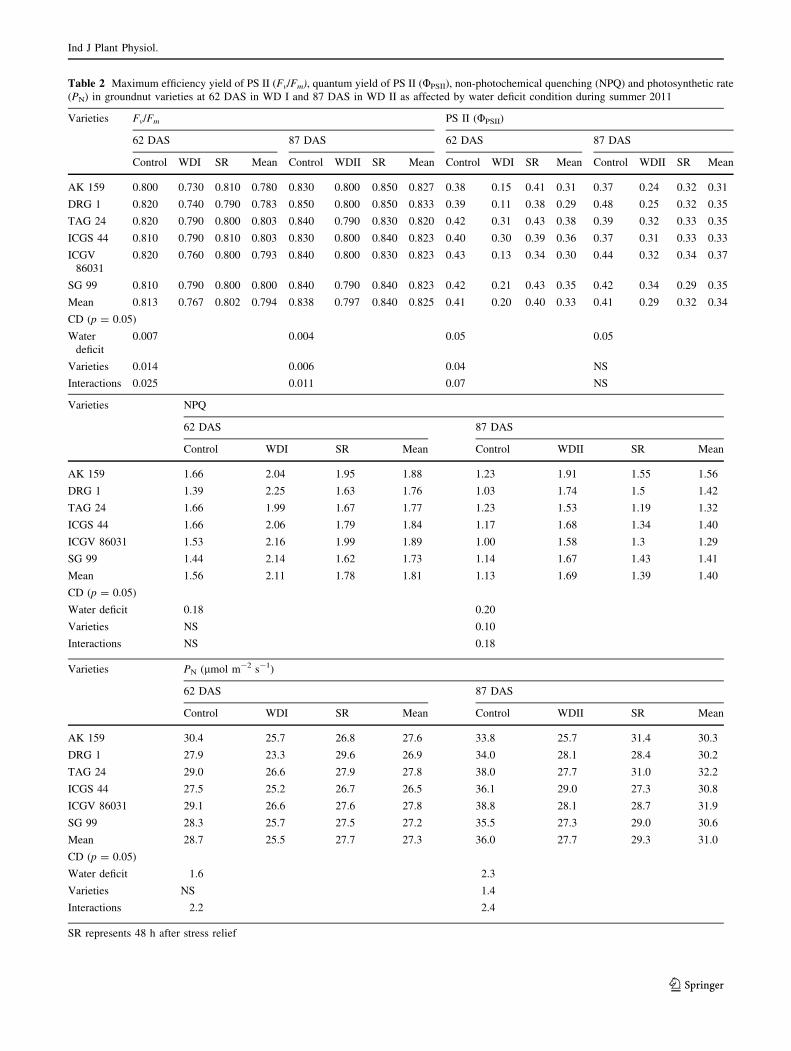
Table 2 Maximum efficiency yield of PS II (Fv/Fm), quantum yield of PS II (UPSII), non-photochemical quenching (NPQ) and photosynthetic rate
(PN) in groundnut varieties at 62 DAS in WD I and 87 DAS in WD II as affected by water deficit condition during summer 2011
Varieties Fv/Fm PS II (UPSII)
62 DAS 87 DAS 62 DAS 87 DAS
Control WDI SR Mean Control WDII SR Mean Control WDI SR Mean Control WDII SR Mean
AK 159 0.800 0.730 0.810 0.780 0.830 0.800 0.850 0.827 0.38 0.15 0.41 0.31 0.37 0.24 0.32 0.31
DRG 1 0.820 0.740 0.790 0.783 0.850 0.800 0.850 0.833 0.39 0.11 0.38 0.29 0.48 0.25 0.32 0.35
TAG 24 0.820 0.790 0.800 0.803 0.840 0.790 0.830 0.820 0.42 0.31 0.43 0.38 0.39 0.32 0.33 0.35
ICGS 44 0.810 0.790 0.810 0.803 0.830 0.800 0.840 0.823 0.40 0.30 0.39 0.36 0.37 0.31 0.33 0.33
ICGV
86031
0.820 0.760 0.800 0.793 0.840 0.800 0.830 0.823 0.43 0.13 0.34 0.30 0.44 0.32 0.34 0.37
SG 99 0.810 0.790 0.800 0.800 0.840 0.790 0.840 0.823 0.42 0.21 0.43 0.35 0.42 0.34 0.29 0.35
Mean 0.813 0.767 0.802 0.794 0.838 0.797 0.840 0.825 0.41 0.20 0.40 0.33 0.41 0.29 0.32 0.34
CD (p = 0.05)
Water
deficit
0.007 0.004 0.05 0.05
Varieties 0.014 0.006 0.04 NS
Interactions 0.025 0.011 0.07 NS
Varieties NPQ
62 DAS 87 DAS
Control WDI SR Mean Control WDII SR Mean
AK 159 1.66 2.04 1.95 1.88 1.23 1.91 1.55 1.56
DRG 1 1.39 2.25 1.63 1.76 1.03 1.74 1.5 1.42
TAG 24 1.66 1.99 1.67 1.77 1.23 1.53 1.19 1.32
ICGS 44 1.66 2.06 1.79 1.84 1.17 1.68 1.34 1.40
ICGV 86031 1.53 2.16 1.99 1.89 1.00 1.58 1.3 1.29
SG 99 1.44 2.14 1.62 1.73 1.14 1.67 1.43 1.41
Mean 1.56 2.11 1.78 1.81 1.13 1.69 1.39 1.40
CD (p = 0.05)
Water deficit 0.18 0.20
Varieties NS 0.10
Interactions NS 0.18
Varieties PN (lmol m-2 s-1)
62 DAS 87 DAS
Control WDI SR Mean Control WDII SR Mean
AK 159 30.4 25.7 26.8 27.6 33.8 25.7 31.4 30.3
DRG 1 27.9 23.3 29.6 26.9 34.0 28.1 28.4 30.2
TAG 24 29.0 26.6 27.9 27.8 38.0 27.7 31.0 32.2
ICGS 44 27.5 25.2 26.7 26.5 36.1 29.0 27.3 30.8
ICGV 86031 29.1 26.6 27.6 27.8 38.8 28.1 28.7 31.9
SG 99 28.3 25.7 27.5 27.2 35.5 27.3 29.0 30.6
Mean 28.7 25.5 27.7 27.3 36.0 27.7 29.3 31.0
CD (p = 0.05)
Water deficit 1.6 2.3
Varieties NS 1.4
Interactions 2.2 2.4
SR represents 48 h after stress relief
Ind J Plant Physiol.
123

indeterminate growth habit. Under such situation, parti-
tioning of photosynthates into developing pods is most
important physiological trait in yield determination. Single
leaf PN has been studied for several groundnut genotypes
and genetic variation is reported (Nautiyal et al. 1995;
Collino et al. 2001). The photosynthesis is fundamental in
both biomass accumulation and productivity and it could
be best utilized in identifying the efficient cultivars and to
understand the physiological traits of productivity both
under normal and stress conditions. Under increasing
moisture deficit stress, the low SLA type groundnut
genotypes were able to maintain higher RWC, PN and gs(Nautiyal et al. 2002). Nautiyal et al. (2012) reported
variation in PN and gs and based on average PN groundnut
cultivars were divided into high, medium and low groups
based on the cropping seasons and accessed the influence
of environment on PN, gs and leaf temperature. They also
reported that photosynthetic rate and associated traits dur-
ing different reproductive growth stages in summer season
showed wide genetic variability in their response to meet
the extra demand of photosynthates by the developing pods
and cultivar TAG 24 showed higher PN during all the
growth stages and maintained least increase in leaf tem-
perature (1.0 �C), which makes it most suitable for culti-
vation in summer season. Bhagsari et al. (1976) observed
large reductions in photosynthesis and stomatal conduc-
tance as the relative water content of groundnut leaves
decreased from 80 to 75 within 3 days of withholding
water in potted plants. Stomatal responses are more closely
linked to soil moisture content than to leaf water status, as
stomata are responding to chemical signals (ABA) pro-
duced by dehydrating roots (Davies and Zhang 1991).
Stage specificity in tolerance and susceptibility to water
deficit condition in this study clearly indicated that pod
development stage between 62 and 87 DAS (WD II) was
more susceptible to water stress than growth period
between 31 and 62 DAS (WD I) and variety TAG 24
showed better stress recovering capacity with high photo-
synthesis under WD condition. The data on chlorophyll
fluorescence parameters showed that cultivar ICGS 44 was
least affected to damage via photo-inhibitory action.
References
Allen, L. H, Jr., Boote, K. J., & Hammondm, L. C. (1976). Peanut
stomatal diffusive resistance affected by soil water and solar
radiation. Proceedings of Soil and Crop Science Society, 35,
42–46.
Araus, J. L., Amaro, T., Voltas, J., Nakkoul, H., & Nachit, M. M.
(1998). Chlorophyll fluorescence as a selection criterion for
grain yield in durum wheat under Mediterranean conditions.
Field Crops Research, 55, 209–223.
Babu, V. R., & Rao, D. V. M. (1983). Water stress adaptations in the
groundnut (Arachis hypogaea L.) foliar characteristics and
adaptations to moisture stress. Plant Physiology and Biochem-
istry, 10, 64–80.
Baker, N. R., &Horton, P. (1987). Chlorophyll fluorescence quenching
during photoinhibition. In D. J. Kyle, C. B. Osmond, & C.
J. Arntzen (Eds.), Photoinhibition (pp. 145–168). Amsterdam:
Elsevier Science Publishers.
Bhagsari, A. S., Brown, R. H., & Schepers, J. S. (1976). Effect of
moisture stress on photosynthesis and some related physiological
characteristics in peanuts. Crop Science, 16, 712–715.
Bilger, W., & Bjorkman, O. (1990). Role of the xanthophyll cycle in
photoprotection elucidated by measurements of light-induced
absorbance changes, fluorescence and photosynthesis in leaves
of Hedera canariensis. Photosynthesis Research, 25, 85–173.
Bjorkman, O., & Demmig-Adams, B. (1994). Regulation of photo-
synthetic light energy capture, conversion, and dissipation in
leave of higher plants. In E.-D. Schulze & M. M. Caldwell
(Eds.), Ecophysiology of photosynthesis (p. 1747). Berlin:
Springer.
Bogalea, A., Tesfaye, K., & Geleto, T. (2011). Morphological and
physiological attributes associated to drought tolerance of
Ethiopian durum wheat genotypes under water deficit condition.
Journal of Biodiversity and Environmental Sciences, 1, 22–36.
Boote, K. J. (1982). Growth stages of peanut (Arachis hypogaea L.).
Peanut Science, 9, 35–40.
Chaves, M. M., Pereira, J. S., Maroco, J., Rodrigues, M. L., Ricardo,
C. P., Osorio, M. L., et al. (2002). How plants cope with water
stress in the field. Photosynthesis and growth. Annals of Botany,
89, 907–916.
Collino, D. J., Dardanelli, J. L., Sereno, R., & Racca, R. W. (2001).
Physiological response of argentine peanut varieties to water
stress. Light interception, radiation use efficiency and partition-
ing of assimilate. Field Crops Research, 70, 177–184.
Table 3 Correlation matrix between various parameters in groundnut varieties at 62 DAS in WD I and 87 DAS in WDII as affected by water
deficit condition during summer 2011
Fv/Fm NPQ PN Transpiration Tleaf
WD I WD II WD I WD II WD I WD II WD I WD II WD I WD II
NPQ -0.71** -0.73**
PN 0.66** 0.62** -0.78** -0.80**
Transpiration 0.71** 0.67** -0.83** -0.87** 0.84** 0.94**
Tleaf -0.67** -0.87** 0.80** 0.71** -0.77** -0.70** -0.87** -0.71**
gs 0.74** 0.64** -0.80** -0.84** 0.76** 0.88** 0.73** 0.93** -0.64** -0.73**
* significant at 5 % level
** significant at 1 % level
Ind J Plant Physiol.
123

Cornic, G., & Massacci, A. (1996). Leaf photosynthesis under stress.
In R. N. Baker (Ed.), Photosynthesis and the environment
(pp. 66–347). The Netherlands: Kluwer Academic Publishers.
Daniele, C., Omar, D., Jean, L. K., & Serge, B. (2006). Genotypes
variations in fluorescence parameters among closely related
groundnut (Arachis hypogaea L.) lines and their potential for
drought screening programs. Field Crops Research, 96,
296–306.
Davies, W. J., & Zhang, J. (1991). Root signals and the regulation of
growth and development of plant in drying soil. Annual Review
of Plant Physiology and Plant Molecular Biology, 42, 55–76.
Devries, J. D., Bennett, J. M., Albrecht, S. L., & Boote, K. J. (1989).
Water relations, nitrogenase and root development of three grain
legumes in response to soil water deficits. Field Crops Research,
21, 215–216.
Jain, L. L., Panda, R. K., & Sharma, C. P. (1997). Water stress
response function for groundnut (Arachis hypogaea L.). Agri-
cultural Water Management, 32, 197–209.
Nautiyal, P. C., Rachaputi, N. R., & Joshi, Y. C. (2002). Moisture-
deficit induced changes in leaf–water content, leaf carbon
exchange rate and biomass production in groundnut cultivars
differing in specific leaf area. Field Crops Research, 74, 67–79.
Nautiyal, P. C., Ravindra, V., & Joshi, Y. C. (1995). Gas exchange
and leaf water relations in two peanut cultivars of different
drought tolerance. Biological Plantarum, 7, 371–374.
Nautiyal, P. C., Ravindra, V., Rathnakumar, A. L., Ajay, B. C., &
Zala, P. V. (2012). Genetic variations in photosynthetic rate, pod
yield and yield components in Spanish groundnut cultivars
during three cropping seasons. Field Crops Research, 125,
83–91.
Richards, A. (2000). Selectable traits to increase crop photosynthesis
and yield of grain crops. Journal of Experimental Botany, 51,
447–458.
Shahenshah, & Isoda, A. (2010). Effect of water stress on leaf
temperature and chlorophyll fluorescence parameters in cotton
and peanut. Plant Production Science, 13, 269–278.
Simmonds, L. P., & Ong, C. K. (1987). Response to saturation deficit
in a stand of groundnut (Archis hypogaea L.). I.Water use.
Annals of Botany, 59, 113–119.
Singh, A. L. (2004). Growth and physiology of groundnut. In
M. S. Basu & N. B. Singh (Eds.), Groundnut research in India
(Vol. 6, pp. 178–212). Jodhpur: National Research Centre for
Groundnut (ICAR).
Singh, A. L. (2011). Physiological basis for realizing yield potentials
in groundnut. In A. Hemantranjan (Ed.), Advances in plant
physiology (Vol. 12, pp. 131–242). Jodhpur: Scientific Publishers
(India).
Subramaniam, V. B., & Maheswari, M. (1990). Physiological
responses of groundnut to water stress. Indian Journal of Plant
Physiology, 33, 130–135.
Ind J Plant Physiol.
123
Copyright © 2022 FDOKUMEN




















