
Glimpses on Nematode Pests of Groundnut in West Bengal
100
Transcript of Glimpses on Nematode Pests of Groundnut in West Bengal






1
I find words inadequate to express my deepest sense of gratitude and heart-feltregards to my honourable teacher Dr. Kusal Roy, Assistant Professor inNematology & Agricultural Entomology, AICRP on Plant Parasitic Nematodes, Directorate of Research, Bidhan Chandra Krishi Viswavidyalaya, Kalyani, Nadia, West Bengal for his scholarly advice, sagacious guidance, pains takingand valuable help during the course of investigation and in the preparation ofthis manuscript. I avail myself of the opportunity to express my deepest sense of gratitude, regards and sincere indebtedness to Professor A. K. Mukhopadhyay, Dr. A. Pramanik and Dr. M. R. Khan, Department of Agricultural Entomology for theirvaluable suggestions and encouragement throughout the entire period ofinvestigation and providing me all sorts of help whenever required. I alsoexpress my gratefulness to Dr. D. K. Mishra, Department of Plant Pathology forhis support and inspiration throughout the entire period of my research work. I would like to convey my honour to Professor. S. K. Mondal, Head, Departmentof Agricultural Entomology, for his valuable suggestion, constant inspirationand help whenever required. I wish to express my sincere gratitude to Dr. S. Jha, and all the other professors of our department for their valuable suggestionduring the course of investigation. I would also like to express my deepest sense of gratitude to the non teachingstaff members of the Department of Agricultural Entomology especially, Benuda, Gurupada da, Ranjit da and Bhashan kaku for their constant help as and
when required.
My heartily thanks and earnest gratefulness are due to my friends and seniors, Peenaz, Devajani, Gouri, Julitha, Priyanka,Dipankar, Amit, Dombewarisa, Tanmay, Pemba, Ananti, Arun bhaiya, Amitbhaiya, Pramod bhaiya, Tina di, Veronica di, and many others for theirinspiration and constant help for completing this work. At last, but not the least, I am very much indebted to my beloved parents, SriGobinda Chandra Sarkar, Gopa Sarkar, whose blessings are the secrets of mysuccess. Elder brother Srijeet Sarkar and other friends and family members fortheir constant assistance, compassion and unending inspiration during thetenure of my present investigation.

2
My acknowledgement will remain incomplete without thanking GOD whosecountless blessings are our inheritance and without his help execution of mywork would not have been possible.

3

4
Tylenchorhynchus mashhoodi
Tylenchorhynchus mashhoodi
µµTylenchorhynchus mashhoodi

5
Figure No.
Title PageNo.
1. Map of the surveyed area for the study on nematode pests ofgroundnut
49
2. Drawing of Tylenchorhynchus mashhoodi; a. entire femalebody; b. anterior region of female showing stomatostylet, andesophagus; c. female tail showing position of phasmid andlateral lines. ext.d-excretory duct, V-vulva, A-anus, hem-hemizonid
63
3. Drawing of Tylenchorhynchus mashhoodi; (a-c) anteriorregion of female
64
4. Drawing of Tylenchorhynchus mashhoodi; a. lateral lines; b. part of female reproductive system showing uterus, vagina andvulva
65
5. Drawing of different populations of Tylenchorhynchusmashhoodi; (a-d) variations in tail shape and position ofphasmid in female
66
6. Drawing of Tylenchorhynchus mashhoodi; a. entire male body;b. anterior region of male; c. male tail. ext.d-excretory duct, hem-hemizonid
67
7. Drawing of Tylenchorhynchus mashhoodi; a. entire juvenilebody; b. anterior region of juvenile; c. juvenile tail. ext.d-excretory duct, A-anus, hem-hemizonid
68

6
8. Light micrographs of Tylenchorhynchus mashhoodi from WestBengal. A. Anterior region of female; B. Anterior region ofmale
69
9. Light micrographs of Tylenchorhynchus mashhoodi from WestBengal. A. Anterior region of female showing stomatostylet, esophagus, excretory duct and hemizonid; B. Anterior regionof female showing posteriorly directed stylet knob; C. Anteriorregion of male
70
10. Light micrographs of Tylenchorhynchus mashhoodi from WestBengal. A. Vulval region showing spermatheca (arrow); B.Female tail showing phasmid (arrow) and lateral lines; C. Maletail showing bursa, spicule and gubernaculum
71

7
per that is
percentage number
and North
smaller than East
greater than degree centigrade
second stagejuvenile
Inch
micron centimetre
hour Feet
cultivar mean sea level
et al et alia (= and others) millimetre
figure species
gram viz. Namely
kilogram Variety
hectare cubic centimetre
juvenile cubic centimetre
meter millilitre

8

9
The groundnut, Arachis hypogaea L. is a member of the oilseed crops ofleguminosae family. The word Arachis hypogaea L. (groundnut) has beenderived from two Greek words, Arachis meaning a legume and hypogaeameaning below ground (referring to the formation of pods in the soil). It is anherbaceous annual with a more or less upright central stem and with numerousbranches that vary from prostrate to almost erect depending upon the variety.
Groundnut (Arachis hypogaea L.) is an annual legume which is alsoknown as peanut, earthnut, monkeynut and goobers. It is the 13th most importantfood crop and 4th most important oilseed crop of the world. Groundnut seeds(kernels) contain 40-50% fat, 20-50 % protein and 10-20 % carbohydrate. Groundnut seeds are nutritional source of vitamin E, niacin, falacin, calcium, phosphorus, magnesium, zinc, iron, riboflavin, thiamine and potassium. Groundnut kernels are consumed directly as raw, roasted or boiled kernels or oilextracted from the kernel is used as culinary oil). It is also used as animal feed(oil pressings, seeds, green material and straw) and industrial raw material (oilcakes and fertilizer). These multiple uses of groundnut plant make it an excellentcash crop for domestic markets as well as for foreign trade in several developingand developed countries (Stigter, 2006).
Cultivated groundnut originates from South America (Wiess, 2000). It isone of the most popular and universal crops cultivated in more than 100countries in six continents. It is grown in 25.2 million hectares with a totalproduction of 35.9 million metric tons (FAO, 2006). Its cultivation is mostlyconfined to the tropical countries ranging from 40º N to 40º S. Major groundnutproducing countries are China (40.1%), India (16.4%), Nigeria (8.2%), USA(5.9%) and Indonesia (4.1%) (Stigter, 2006).
Among oilseeds crops in India, groundnut accounts for about 50% of thearea and 45% of oil production. In India, about 75% of the groundnut area lies ina low to moderate rainfall zone (parts of peninsular region and western andcentral regions) with a short period of distribution (90-120 days). Total area ofgroundnut in West Bengal is 65,826 hectares with an annual production of1, 13,018 tonnes and yield 1717 kg/ha (Anon., 2010). Based on rainfall pattern,

10
soil factors, diseases and pest situations, groundnut-growing area in India hasbeen divided into five zones.
In India, most of the groundnut production is concentrated in five statesviz., Gujarat, Andhra Pradesh, Tamil Nadu, Karnataka, and Maharashtra. Thesefive states account for about 86% of the total area under peanut cultivation. Theremaining peanut producing area is scattered in the states of Madhya Pradesh, Uttar Pradesh, Rajasthan, Punjab, and Orissa. Although the crop can be grown inall the seasons, it is grown mainly in rainy season (Kharif; June-September). Thekharif season accounts for about 80% of the total groundnut production. In thesouthern and south-eastern regions, groundnut is grown in rice fallows duringpost-rainy season (Rabi; October to March). If irrigation facilities are available, groundnut can be grown during January to May as a spring or summer crop. Monsoon variations cause major fluctuations in groundnut production in India. Groundnut is grown in different cropping systems like sequential, multiple, andintercropping (Basu and Ghosh, 1995).
As to production constraints, because groundnut is grown mainly asrainfed crop there is a high level of fluctuation in the production depending onthe rainfall. The productivity is restrained by drought stress, the use of lowlevels of inputs by small and marginal farmers in dryland areas, high incidenceof foliar fungal diseases, and attack by insect pests and nematodes.
Major nematode pests of groundnut include dagger nematode (Xiphinemaspp.), pod lesion nematode (Tylenchorhynchus brevilineatus, Tylenchorhynchusbrevicaudatus), ring nematode (Criconemoides ornatus), root-knot nematodes(Meloidogyne arenaria, Meloidogyne javanica, Meloidogyne hapla), root lesionnematode (Pratylenchus brachyurus, Pratylenchus coffeae), seed and podnematode (Ditylenchus destructor), spiral nematode (Scutellonema cavenessi), sting nematode (Belonolaimus gracilis, Belonolaimus longicaudatus) andgroundnut testa nematode (Aphelenchoides arachidis) (http://en.wikipedia.org).
The most serious nematode pest of peanut is Meloidogyne arenaria race1, the peanut root-knot nematode. It can be expected to occur wherever peanutsare grown. Pod rot, white mold, and other soil-borne diseases may increasewhen the peanut plant is infected with this root-knot nematode. The lesionnematode, Pratylenchus brachyurus, is less troublesome, but it can reduce yields

11
and seriously disfigure the peanut hulls with unattractive brown lesions that leadto pod rotting.
A severe nematode disease of groundnut, popularly called 'Kalahastimalady', is caused by Tylenchorhynchus brevilineatus. Peanut yellow diseaseoccasionally appeared in the field due to infestation of Criconemoides ornatus.
Areas of root-knot nematode infected peanuts are usually round to oblongin shape. The plants are stunted, less green, and will wilt more readily in the heatof the day than plants in less infested areas of the field. Rows of infected plantsmay never, or not so quickly, meet as those of healthy plants. Above-groundsymptoms in a lesion nematode-infected crop are exhibited as a dull yellowingin oval spots in the field. These symptoms may be similar to nutrientdeficiencies; so care must be taken in diagnosis. Above-ground symptoms ofsuspected nematode problems on peanut should be verified by soil, root, and podassays to properly identify a nematode causal agent (Rich and Kinloch, 2009). All the nematodes affecting peanut reduce feeder roots and produce rootstunting, but nematodes differ in specific symptoms on roots and pods.
Symptoms of peanut root-knot nematodes include galls (knots) on bothroots and pods. The presence of galls on the pods appears as single or multiplewart-like growths that may or may not be discoloured. As root-knot nematodeinfection progresses, secondary root and pod rotting causes further damage andeventual death of the plants. Lesion nematode damage is most easily seen onpods, which show distinct light brown lesions. As infection and diseaseprogresses, the lesions become less distinct and turn black in colour. Presence ofhigh numbers of lesion nematodes results in extensive root and pod rotting (Richand Kinloch, 2009).
Sasser and Freckman (1987) estimated that the plant parasitic nematodescause an annual yield loss of 12.3 % on global basis, while in India nematodescause 5 % loss in oilseed crop production (Seghal and Gaur, 1999). An annualyield loss of groundnut by 21.6% due to infestation of Meloidogyne arenariaand M. javanica in India had been reported by Jain et al. (2007).
In southern part of India, several studies pertaining to plant parasiticnematode community (Mani and Kumar, 1992; Reddy et al.,1993; Senthamizhand Sivakumar, 2004), their pathogenicity (Kalaiarasan et al., 2006) andmanagement (John et al, 2006; Kalaiarasan et al., 2006) in groundnut havealready been made but still there is hardly any information with regard to the

12
occurrence and distribution of phytonematodes on groundnut in West Bengalthough the crop has been growing since twenty years in some areas of the state.
The present investigation was therefore undertaken to study theoccurrence, distribution and community of the phytonematodes andmorphometric variability of Tylenchorhynchus species associated withgroundnut.
Following are the objectives of the present investigation-• To study the occurrence and distribution of phytonematodes associated with
groundnut. • Community study of the prevailing rhizospheric nematodes associated with
the crop. • To confirm the identity of the major nematode pests associated with the crop. • To study the morphological and morphometric variations of
Tylenchorhynchus species associated with the crop.

13
The legume Arachis hypogaea L. commonly known as peanut orgroundnut is a very important food crop throughout the tropics and sub-tropics. The genus is endemic to South America being mostly associated with thesavannah-like Cerrado. All species in the genus are unusual among legumes inthat they produce their fruit below the ground (Bertioli et al., 2011). Groundnutis grown mainly as rainfed crop. The productivity of groundnut is constrained bydrought stress, the use of low levels of inputs, incidence of diseases as well asattack by insect pests and parasitic nematodes.
Major nematode pests of groundnut include dagger nematode (Xiphinemaspp.), pod lesion nematode (Tylenchorhynchus brevilineatus, Tylenchorhynchusbrevicadatus), ring nematode (Criconemoides ornatus), root-knotnematodes(Meloidogyne arenaria, Meloidogyne javanica, Meloidogyne hapla), root lesion nematode (Pratylenchus brachyurus, Pratylenchus coffeae), seed andpod nematode (Ditylenchus destructor), spiral nematode (Scutellonemacavenessi), sting nematode (Belonolaimus glacilis, Belonolaimus longicaudatus)and groundnut testa nematode (Aphelenchoides arachidis)(http://en.wikipedia.org).
Among phytonematodes Meloidogyne, Pratylenchus, Criconemella andBelonolaimus spp. caused diseases of groundnut pods; chlorosis caused byAphasmatylenchus straturatus; yellow patch disease caused by Scutellonemaspp. and seed-borne diseases caused by Aphelenchoides arachidis andDitylenchus destructor (Sharma and McDonald, 1992).
A survey by Mani and Kumar (1992) on groundnuts from the Chittoorand Nellore districts of Andhra Pradesh revealed the presence of 28 plantparasitic nematode genera with a higher occurrence and density in rabi thankharif season. Bitylenchus brevilineatus, Pratylenchus spp., Basirolaimus spp. and Tylenchorhynchus spp. were predominant. The highest frequency ofoccurrence in kharif was for Basirolaimus spp. and Pratylenchus spp. at 68.8and 64.5% respectively and in rabi for Pratylenchus spp. and Bitylenchusbrevilineatus at 74.1 and 70.4%, respectively. Groundnut was recorded as newhost for Basiria, Ecphyadophora, Nothotylenchus, Sakia and Tenunemellus spp.

14
Senthamizh and Sivakumar (2004) collected soil and root samples fromthe rhizosphere of groundnut from 51 localities in Namakkal district, processedit and then analysed. The results revealed the presence of eight nematode speciesbelong to several genera. The occurrence of Criconemoides curvatum, Helicotylenchus dihystera, Hoplolaimus (=Basirolaimus) seinhorsti andTylenchorhynchus brevilineatus in soil samples. Pratylenchus zeae, Meloidogyne arenaria, M. incognita and Rotylenchulus reniformis in rootsamples were also observed. P. zeae was found most frequently with an absolutefrequency of 67%. M. incognita recorded the prominence values of 34.50 and21.80 in soil and roots, respectively, followed by M. arenaria.
Khan and Misari (1992) studied 47 groundnut-growing localities in the 4ecological zones viz., Sahel savanna, Sudan savanna, Northern Guinea savannaand Southern Guinea savanna, of Nigeria and had found the presence of 22genera of plant parasitic nematodes in the rhizosphere, roots and pods of thiscrop in low to high populations. Based on their high frequency of occurrence, high populations encountered and direct and indirect evidence of pathogenicity, Aphelenchoides arachidis, Pratylenchus brachyurus, P. coffeae andScutellonema cavenessi are considered the most important nematode pests ongroundnut in these zones. Chlorotic symptoms, observed on groundnut foliage incertain localities, are attributed to the infection of S. cavenessi.
Distribution of plant-parasitic nematodes in coastal, mid land hilly, andriver bed delta in Nghe A province in North Vietnam was appraised in April1993. Thirty-one nematode species within 21 genera were identified associatedwith groundnut in 25 villages in 5 districts. Macroposthonia ornata, Tylenchorhynchus annulatus, and Pratylenchus spp were the most frequentlyassociated nematodes. M. ornata was considered most important because of itsincidence in 93% of the surveyed region; it was predominant in 32% of thelocations. Swelling of the apical parts of groundnut roots was common and M. ornata was suspected of being linked with 'apical gall' symptoms. T. annulatuswas present in 80% of the samples but its population density was normally low(less than 0.5 nematodes per cm 3 soil). Pratylenchus brachyurus and P. zeaevaried in their distribution; P. brachyurus was widespread in sandy coastal soilswhile P. zeae populations preferred loamy soils in the midland hilly region. Cysts of Heterodera spp were found in soil samples in 43% of locations, butgroundnut roots were free from cysts. Meloidogyne graminicola, M. javanica, and Meloidogyne sp. were found in 77% of locations surveyed. Populations of

15
these species were suspected to affect the rice production. Incidentally, 21 of thespecies identified are being reported for the first time in association withgroundnut in Vietnam, and 8 of them have not previously been reported inassociation with groundnut in any country (Sharma et al., 1994).
Kepenekci and Ozturk (2002) carried out a nematode survey on groundnut(A. hypogaea) growing areas in the Mediterranean region of Turkey, 15 specieswere determined as belonging to ten genera of eight families within thesuperfamilies Tylenchoidea, Dolichodoroidea, Hoplolaimoidea andAnguinoidea. Each of them constitutes a new record on groundnut in Turkey, and Scutylenchus tumensis and Pratylenchus brachyurus were reported for thefirst time in the nematode fauna of Turkey.
Mukhopadhyay and Haque (1974) reported the occurrence ofRotylenchulus reniformis from castor growing areas of Burdwan, Hooghly andNorth 24 Parganas districts of West Bengal indicating the population range of1600-2340/100g of soil. Later in 1978, Singh and Khera revealed that R. reniformis was one of the commonest and dominating nematode parasitizingvegetable crops (brinjal, okra and tomato) of West Bengal.
A roving survey was carried out by Mukherjee and Dasgupta (1983) infive locations (Chandan Nagar, Hooghly, Chinsurah, Mankundu andAdisaptagram) of Hooghly districts covering banana plantations and noticed thatRotylenchulus reniformis was most frequently present in every locations andplaced 2nd after Tylenchorhynchus coffeae. Other species of nematodes viz., Meloidogyne incognita, Helicotylenchus multicinctus, H. indicus, Pratylenchuszeae, P. brachyurus, Hoplolaimus indicus, H. columbus, Scutellonema simense, Hemicriconemoides mangiferae, Xiphenema insigne, Aphelenchus avenae, Sakiaindica, Cephalenchus leptus and Boleodorus sp were also encountered.
Survey was undertaken by Mukhopadhyay and Roy (2006) to determinethe frequency and abundance of plant parasitic nematodes associated withvegetable crops in the eastern and north eastern regions of India. Species of thegenera Meloidogyne, Rotylenchulus, Tylenchorhynchus, Hoplolaimus, Pratylenchus, Helicotylenchus, Hirschmanniella and Criconemoides were foundto be the most important ones frequently encountered in the sample from WestBengal, Orissa, Tripura, Arunachal Pradesh and Nagaland. Members of thelongidoridae were also recorded from Arunachal Pradesh but in small numbers. Community analysis of these nematodes revealed that with regard to the cropgroups prominence value and density, the genus Meloidogyne (M. incognita and

16
M. javanica) ranked first in solanaceous and leguminous vegetables, second incucurbitaceous and third in cruciferous vegetables in West Bengal. The samewas the most important nematode group in the vegetable crops of Orissa andArunachal Pradesh too. The Rotylenchulus reniformis was recorded as the mostimportant nematode in vegetable crops of Tripura and Nagaland andcucurbitaceous vegetables of West Bengal while second important insolanaceous and third in leguminous vegetables in West Bengal. However, incruciferous vegetable crops Tylenchorhynchus and Hirschmanniella were themost predominant over other nematodes in West Bengal.
A roving survey was conducted to the leguminous vegetable cropsgrowing areas of West Bengal during 2001-2004. Soil samples from the sevendistricts of West Bengal under survey had Rotylenchulus, Meloidogyne, Tylenchorhynchus, Pratylenchus and Helicotylenchus nematodes population. Among four leguminous vegetable crops (cowpea, dolichosbean, french beanand pea) surveyed, only cowpea, Vigna unguiculata (L.) Walp. and pea, Pisumsativum L. var. hortense roots showed occurrence of Rotylenchulus reniformisbut all supported Meloidogyne incognita and M. javanica population. Community analysis revealed 17 species of plant parasitic nematodes belongingto eight genera, which remained in close association with the crops. Rotylenchulus reniformis was most predominant based on prominence value, frequency and density. Meloidogyne spp. and Tylenchorhynchus spp. wereprevalent next to Rotylenchulus reniformis with ranking 2nd and 3rd, respectively(Roy et al., 2007).
Roy and Mukhopadhyay (2011) carried out a roving survey onsolanaceous vegetable crops (Solanum melongena L., Lycopersicon esculentumMill. and Capsicum annum L.) and collected 213 soil samples from 11 districts[24 Parganas (South and North), Nadia, Burdwan, Hooghly, Midnapore (Eastand West), Murshidabad, Malda, South Dinajpur and Purulia] of West Bengal. Community analysis of soil nematodes associated with the crops revealedprevalence of twelve species of plant parasitic nematodes under eight genera. Those were Meloidogyne incognita, M. javanica, Rotylenchulus reniformis, Tylenchorhynchus mashhoodi, Tylenchorhynchus sp., Hoplolaimus indicus, Pratylenchus sp., Helicotylenchus dihystera, Helicotylenchus sp., Hirschmanniella oryzae, H. mucronata and Criconemoides onoensis. Frequencyof occurrence of Meloidogyne spp. were 83% as compared to 59% in case of R.reniformis. M. incognita, M. javanica and R. reniformis were identified as

17
dominant pathogenic nematodes, causing crop damage in the state. Saprozoicnematodes were also encountered in huge number from soil samples collectedfrom different areas of the state.
Meloidogyne Three major Meloidogyne spp. parasitize groundnuts and each is capable
of causing severe suppression of groundnut yields and fruit quality. Two of thespecies, M. arenaria (groundnut root knot nematode) and M. javanica (Javaneseroot knot nematode), are highly virulent pathogens of groundnut, whereas M. hapla (northern root knot nematode) causes less damage but none the less is animportant disease-inducing agent of groundnut. All three species occur ongroundnut worldwide (Sasser, 1977).
M. arenaria and M. javanica are common in warm and hot regions of theworld, whereas M. hapla occurs only in cooler regions. A new species of rootknot nematode causing disease on groundnut in Texas, USA was recentlydescribed as M .haplanaria (Eisenback et al., 2003).In the USA, M. arenaria isthe dominant Meloidogyne species parasitizing groundnut in Alabama, Florida, Georgia and Texas, while patchy occurrences have been reported in NorthCarolina, South Carolina and Virginia. In other regions of the world, M. arenaria is reported damaging groundnut in Zimbabwe (Martin, 1958), Israel(Orion and Cohn, 1975), Egypt (Ibrahim and El-Saedy, 1976a), India (Sharmaet al., 1978; Dhurj and Vaishnav, 1981; Sakhuja and Sethi , 1985b), Taiwan (Cheng and Tu , 1980; Cheng et al., 1981) and China (Zhang, 1985). M. javanica was first reported parasitizing groundnut in Zimbabwe (Martin, 1958). Although the species is highly virulent on groundnut; it is less frequentlyencountered on groundnut in the USA than M. arenaria. It is only reportedparasitizing groundnut in Florida (Cetintas et al., 2003), Georgia (Minton et al., 1969) and Texas (Tomaszewski et al., 1994). The species is also known to occuron groundnut in Egypt (Ibrahim and El-Saedy, 1976b), Brazil (Lordello andGerin, 1981; Carneiro et al., 2003) and India (Sakhuja and Sethi, 1985a). M. hapla is encountered more frequently on groundnut in the more northernlatitudes. However, the nematode may be encountered at higher elevations intropical regions (Eisenback and Triantaphyllou, 1991). M. hapla has beenreported infecting groundnut in all groundnut producing states in the USA withthe exception of Florida (Dickson, 1998). It is frequently encountered infectinggroundnut in North Carolina, Virginia and Oklahoma. The species parasitizesgroundnut in Israel (Minz, 1956), South Africa (Van der Linde, 1956), Australia

18
(Colbran, 1958; Saint-Smith et al., 1972), Zimbabwe (Martin, 1961), Japan(Mitsui et al., 1976), Korea (Choi, 1981) and China (Yin and Feng, 1981; Yang, 1984; Zhang, 1985).
Noticeable above and below ground symptoms of root knot nematodes ongroundnut can be observed as early as 45–75 days after planting, but mostsevere symptoms are observed after 90–120 days. Above-ground symptoms ofroot knot disease may be subtle or very conspicuous, especially as the crop nearsmaturity. The degree of symptoms depends on the growing environment as wellas the population density of root knot nematode juveniles at the time of planting. In some cases, stunting of young plants may be severe. The generalcharacteristics of diseased groundnut plants are typical of other plantsinfected by these nematodes. As the crop nears maturity, heavily infected plantsmay be severely stunted, showing symptoms of chlorosis, incipient wilting, nutrient deficiencies, or even death when conditions are hot and dry. Symptomsare distributed in patches of varying sizes. Infected plants exhibit a rusty, yellowish and mottled appearance. If drought occurs near the end of the season, the severity of root knot disease is accentuated and weakened plants die. Earlyseason symptoms include stunted plants that fail to cover the soil between rows.The slowly dying and browning plants present a mottled effect among thegreener plants, but even when such plants show conspicuous galling theirneighbouring plants in apparent vigour are usually also infected (Machmer, 1951). In China, M. arenaria infected plants may become yellow and stunted asearly as 40 days after planting (Zhang, 1985).Second stage juveniles infectgroundnut plants soon after they germinate, but noticeable galling and eggmasses are not apparent on the roots until 55–90 days after planting. Thecharacteristic symptom on roots is the abnormal swelling (galls or knots)however, these are often difficult for the novice to see. Galled tissue on rootsmay attain a diameter larger than that of normal adjacent roots, but, because ofthe abundance of nodules containing nitrogen-fixing bacteria (Bradyrhizobium(Arachis) sp.), the amount of galling is difficult to determine. This is distinctfrom what happens on tomato or cucumber where galling is evident on roots 2–3weeks after planting. Groundnut root galls are small and generally discrete, whereas galls on other host crops may be largeand become coalesced. Nematode galls can be distinguished from nodulescontaining nitrogen-fixing bacteria. Nodules are distinctive, round swellingsattached to the root and are easily detached, whereas nematode galls are

19
swellings that constitute a part of the fibrous root system and cannot be removedwithout destroying the integrity of the root. Also, because second stagenematodes can infect nodules in some instances, galls may appear on thenodules, and vice versa. Root knot juveniles may also infect pegs and podsafter blooming and initiation of pod set (generally ~45 days after planting). Galling on pegs and pods is distinctive and more easily seen than that on rootsbut, interestingly, it does not always appear even though roots are galled. Situations where galling on pegs and pods is extensive generally result in a largereduction in potential yield. Yield potential is lost because pegs are weakenedand easily fall off during harvest, pod formation is aborted, or damaged pods failto produce seeds. Galling induced by M. hapla is distinctively different fromthat caused by M. arenaria or M. javanica. The former results in smaller gallswith some root proliferation above galls that results in a denser root system(Sasser, 1954).The most obvious sign of root knot nematodes is theobservation of female nematodes in galled roots, pegs or pods. Females areglobose, pearly white in colour, and have sharp pointed necks and heads off toone side that are generally visible. An egg mass is generally extruded from thevulva end of each female at or near the root surface. This positions the egg masson the outside surface of galled tissue, which facilitates both egg hatchand secondary infection of roots by the freshly hatched juveniles. Egg massesare about 1mm in width, appear as brownish masses adhering to galled tissueand contain up to 300–500 eggs each. They are generally plentiful along galledtissue. Staining with food colouring (Thies et al., 2002) or Phloxine B(Dickson and Struble, 1965) enables one to see them more readily. M. haplasymptoms are generally less severe than those caused by either M. arenaria orM. javanica. Above-ground symptoms of M. hapla may be difficult todetect because this pathogen causes less stunting or chlorosis. The most severesymptoms generally indicate large population densities of infective juveniles inthe soil. Severity over a field varies, depending on the variations in soilpopulation densities and soil type. Sandy areas within a field often show themost severe symptoms. Plants with light infections usually do not show stuntingor chlorosis. Typically, the only indication of root knot disease on such plants isgalls on roots, pegs and pods.
Biochemical changes induced by Meloidogyne javanica pathotype 2 ingroundnut cv. GG 20 revealed that peroxidase and polyphenol oxidase activity, and total phenol contents of groundnut progressively increased with an increase

20
Pratylenchus brachyurus
The lesion nematode, Pratylenchus brachyurus, is a major nematodeparasite of groundnut, with a distribution mainly in the warmer groundnutproduction regions of the world (Loof, 1964). The species was first reported ongroundnut in Alabama, USA in 1942 (Steiner, 1949), and is now known toparasitize groundnut in most of the groundnut producing states in the USA, andseveral other countries of the world including Egypt (Oteifa, 1962), Australia(Colbran, 1968) and Zimbabwe (Anon., 1973). One other lesion nematodespecies, P. coffeae, was reported parasitizing groundnut in India (Chabra andMahajan, 1976). Reddy et al. (1993) reported Pratylenchus brachyurus from 3groundnut growing districts in Karnataka, India. This is the first report of thisspecies on groundnut in this area. Taxonomic separation of species of
in nematode inoculum levels. Highest inoculum level of 10,000 J2/plantrecorded maximum activity of peroxidase and polyphenol oxidase, and totalphenol contents of roots over control. Significant reduction in chlorophyll a, band total chlorophyll contents was observed at all the inoculum levels. Maximum reduction in chlorophyll content was noticed at the 10,000 J2/plantlevel (Patel et al., 1999).
Patel and Patel (1999) conducted an experiment on the pathogenicity ofMeloidogyne javanica pathotype 2 on groundnut cv. GG 20 during the kharif1996 and 1997 seasons, it showed that an inoculum level of 100 J2 andabove/plant significantly reduced plant growth parameters viz., plant height, fresh and dry shoot weight. Fresh and dry root weights progressively increasedwith an increase in nematode inoculum level from 10 to 10,000 J2/plant due toheavy root galling and profuse egg sac formation on roots. A nematodeinoculum level of 10,000 J2/plant showed significantly minimal plant height, fresh and dry shoot weights. The total nematode population increased with anincrease in nematode inoculum levels while nematode reproduction ratedecreased with a corresponding increase in inoculum levels. It was maximum atthe level of 10 J2/plant, and minimum at the highest level of 10,000 J2/plant. M.arenaria was inoculated on groundnut cv. CO 3 at 10, 100, 1000 and 10,000J/kg soil in Tamil Nadu, India. Plants with 100 J2/plot showed 28.19% reductionin pod yield compared to the untreated control, while 100% yield reduction wasobtained under the 2 highest inoculum levels (Kalaiarasan et al., 2006).

21
Pratylenchus is difficult because they exhibit little morphological diversity(Roman and Hirschmann, 1969).
Distinct field symptoms of P. brachyurus damage on groundnut aredifficult to discern. Severely infected groundnut plants may be stunted andchlorotic, but this is rare. Heavy infection by the lesion nematode is reported tocause extensive discoloration of below-ground plant parts and reduced rootsystems and pod weights. Above-ground symptoms may include slight stuntingwith unthrifty, yellow-green foliage (Miller and Duke, 1961; Boswell, 1968). The most obvious symptom of lesion nematode damage on groundnut is small, purplish-brown to black lesions that form on the groundnut shell (Good et al.,1958; Boswell, 1968). The plant nematode-induced lesions are described asgiving pods a speckled appearance, and are conspicuous to the trained eye(Miller and Duke, 1961). These lesions have distinct boundaries and first appearas small brown tunnels in the shell and have the potential later to coalesce toform larger lesions. When coalesced, they are difficult to separate from thoseinduced by other soil microbes. Secondary soil bornepathogens may enter these lesions causing them to increase in size, or theinfected pegs and pods may rot. Infection of pegs by P. brachyurus has beencorrelated with a peg rot condition resembling the peg rot disease caused bySclerotium rolfsii (Good et al., 1958). Combinations of fungal- and nematode-induced lesions may occur, but this has received little study (Good et al, 1958).P. brachyurus can be found in roots and pegs, as well as shells of mature pods, but the nematode is more numerous in shell tissue. The lesions on mature podsare ‘purplish-brown’ and have a somewhat darker colour withdistinct boundaries as compared with those induced by a soil–microbial complex(Good et al., 1958).Symptoms of pod lesions may vary depending on type ofgroundnut or cultivar, e.g. they may be less conspicuous on Virginia-typegroundnut than on Spanish and Runner types (Good et al., 1958; Minton et al., 1970). P. brachyurus feeding within the pegs weakens them, resulting in podloss at harvest (Good et al., 1958; Boswell, 1968; Jackson and Sturgeon, 1973). Also, microorganisms that colonize damaged pods may penetrate the shell anddamage the seed, thus the yield; as well as the quality and value of the crop maybe reduced (Good et al., 1958).
Criconemoides ornatus
Criconemoides ornatus (commonly called the groundnut ring nematode)was first reported associated with groundnut in Georgia (Boyle, 1950; Machmer,

22
1953). It is now known to occur in a large percentage of the groundnutproduction regions of the USA (Alexander, 1963; Minton and Bell, 1969;Motsinger et al., 1976; Ingram and Rodríguez-Kábana, 1980; Wheeler and Starr, 1987). Criconemoides spp. has been reported in Burkina Faso (Germani andDhéry, 1973), Egypt (Ibrahim and El-Saedy, 1976a) and Gambia (Merny et al., 1974).
A chlorotic condition of groundnut, called ‘groundnut yellows’, wasreported in Georgia in a soil heavily infested with a species of Criconemoides(Machmer, 1953). Although the species involved was never identified, it wasprobably C. ornatus. The ‘yellows disease’ symptom was duplicated in microplots by using freshly extracted, greenhouse grown inoculums of C. ornatus(Barker et al., 1982). Although yellowing of groundnut has been observed andduplicated in micro plots, this is not a common symptom. In fact, it is probablyrarely seen. As few as 178 freshly introduced C. ornatus/500cm3of soil will stuntgroundnut. Roots, pegs and pods of plants growing in micro plots in soil heavilyinfested with C. ornatus are severely discoloured with brown necrotic lesions(Minton and Bell, 1969). Small necrotic lesions caused bythis nematode are often superficial, but necrosis in large lesions usually extendsdeep into the tissues. The primordial of many lateral roots and young roots arekilled, which results in reduced numbers of lateral roots. Pod yields fromnematode infected plants are reduced by about one-half.
Tylenchorhynchus brevilineatusTylenchorhynchus brevilineatus was first observed causing damage to
groundnut in1976 in the Kalahasti area of Andhra Pradesh State, India (Reddy etal., 1984).The disease caused by this nematode is known as ‘Kalahasti malady’. Since 1976, the disease has been widespread in the Kalahasti area, but has alsobeen observed in Nellore District in Andhra Pradesh (Reddy et al., 1984). InChittoor area of Andhra Pradesh (India), ‘Kalahasti malady’ was also reported tocause severe reduction in groundnut yields (John et al., 2006).
Disease symptoms are characterized by small pods and a brownish-blackdiscoloration of the pod surfaces (Reddy et al., 1984). Small, brownish-yellowlesions appear on the pegs and pod stalks and on young, developing pods. Lesion margins are slightly elevated because of host cell proliferation aroundthem. The length of pod stalks is greatly reduced, and in advanced stages of thedisease the pod surface becomes completely discoloured, but seeds fromdiseased pods are healthy. Discoloration is also observed on roots but is less

23
severe than on pods. Pathogenicity tests in the greenhouse corroborated fieldobservations (Reddy et al., 1984). Groundnut plants inoculated with 500 T.brevilineatus /12cm diameter pot were severely stunted and had reduced rootsystems. Lesions were present on the roots but were not extensive. Pods wereseverely discoloured and small, but seeds from the discoloured pods werehealthy. Brownish-yellow lesions were observed on individually inoculated podsafter 15 days. The number of lesions increased and extensive discoloration wasobserved by 30days after inoculation. Out of 1599 groundnut germplasmaccessions, 14 were confirmed to be resistant in advanced screening trials in the1986 rainy and the 1985-866 and 1986-87 post-rainy season. Most of theresistant genotypes have undesirable pod/seed characteristics. One of theresistant genotypes is a high-yielding breeding line (TCG 1518) and this is beingreleased for use in disease-affected areas of Andhra Pradesh (Mehan et al., 1993). A total of 39 groundnut genotypes, including three control varieties werescreened for resistance to the nematode disease i.e. ‘Kalahasti malady’ atSrikalahasti, Andhra Pradesh during 1992-94. Three test genotypes, TCGS-307, TCGS-313 and TCGS-320 and a resistant check variety TPT-3 exhibitedresistant reaction to the nematode disease with a disease score of 2.0 (Naidu andMoses, 2000). Thirty groundnut genotypes were screened for resistance toTylenchorhynchus brevilineatus in a field experiment in Andhra Pradesh, Indiaduring the 1998-99 post-rainy seasons. Among these resistant genotypes, TCGS-320, TCGS-303, TCGS-597, TCGS-337 and TCGS-428 had pods with hardshells, prominent ribbing and kernels with pink colour, indicating that nematoderesistance was associated with morphological characters. (Sreenivasulu et al., 2001). Infestation of JL-24 (susceptible) groundnut pods with T. brevilineatusresulted in a slight increase in total protein (8.8%), total soluble sugars (1%), and total phenolic compound (21.2%) and a decrease in total crude fibres(21.3%) (Naidu et al., 2000).
Ditylenchus africanus
Ditylenchus africanus, the groundnut pod nematode, was originallydescribed as D. destructor, the potato rot nematode. It was first reporteddamaging groundnut in the Transvaal Province of South Africa in 1987(Jonesand De Waele, 1988). A subsequent survey revealed the presence of thisnematode in seven major groundnut-producing regions (De Waele et al., 1988), where 75% of 877 seed samples that graded ‘damaged’ were infected. An

24
average of 160 nematodes/seed was recovered. This nematode has not beenreported on groundnut in other parts of the world.
D. africanus was isolated from roots, pegs, shells and groundnut seeds(De Waele et al., 1988). Visible symptoms are not apparent on roots, but seedshow blemishes and premature germination before harvest. Infected pods of cv. Sellie are black, resembling black hull caused by Chalara elegans Nag Raj&Kendrick. Approximately 40–60% of the pods and seeds are destroyed inheavily infested fields. D. africanus is presentin both hulls and seeds, which results in a lower quality grade and reducedgroundnut yield (Mc Donald et al., 2005). In greenhouse pathogenicity tests (DeWaele et al., 1988), nematodes were pre-sent in the peg, exocarp and endocarp, testa and embryo, and on the cotyledons. The first symptom to develop wasbrown necrotic tissue at the pod base at the juncture of the peg and pod. Thesurface of infected tissue was dark brown and had a corky appearance. The mostdistinct symptom of advanced disease was dark brownto black discoloration of veins that extended longitudinally in the exocarp justbeneath the pod surface. Infected pods lacked the lustre of healthy pods andappeared dead. Infected seeds were usually shrunken and the micropyles weredark brown to black. The testa were flaccid, had dark vascular strands and wereeasily removed. The inner layer of the testa had a distinct yellow discoloration. Infected embryos were usually olive green to brown instead of having thenormal colourless to yellow appearance. The extent of potential yield lossescaused by this nematode and research relative to its management has not beenreported. Steenkamp et al. (2011) found that Ditylenchus africanus causescellular breakdown in pod tissue of susceptible groundnut cultivars. Thehistopathology of this nematode on a resistant genotype was studied using lightmicroscopy and compared with the histopathology of D. africanus on asusceptible genotype. In contrast to the susceptible genotype, only a smallnumber of nematodes were observed in restricted areas of the pod tissue of theresistant genotype. Furthermore, the resistant genotype showed neither externalsymptoms nor cellular breakdown in reaction to D. africanus. According toresults of this study, the mechanism of resistance involved may be the inhibitionof proper development, migration and reproduction of this nematode, thuspreventing it building up to damaging population levels.
Tylenchorhynchus
Phyllum-Nematoda Rudolphi, 1808 (Lankester, 1877)

25
Class-Secernentea von Linstow, 1905; Andrassy, 1976Sub class-Tylenchia Inglis, 1983
According to Allen and Sher (1967) Tylenchorhynchus belongs to thesuper family-Tylenchoidea, family-Tylenchidae and sub family-Tylenchorhynchinae. Andrassy (1976) proposed super family-Hoplolaimoideainstead of Tylenchoidea of Allen and Sher (1967) keeping the similar family andsub family for the genus Tylenchorhynchus. Siddiqi (1986) placedTylenchorhynchus under order –Tylenchida (Thorne, 1949), sub order-Tylenchina (Chitwood, 1950), super family- Dolichodoroidea (Chitwood, 1950), family- Dolichodoroidae and sub family-Tylenchorhynchinae (Eliava, 1964).
The cosmopolitan genus Tylenchorhynchus was established by Cobb(1913) for T. cylindricus found in southern California. Currently this genuscontains 111 species that parasitize a wide variety of plants. Allen (1955)enhanced taxonomic criteria for Tylenchorhynchus. Tarjan in 1964 prepared acompendium of the genus Tylenchorhynchus and later in1973 he gave asynopsis, key, and diagnostic data of the genera and species belonged toTylenchorhynchinae. The history of the genus was discussed by Hooper (1978), Golden et al. (1987), and Anderson and Potter (1991). Siddiqi (1986) regarded71 total species as being in Tylenchorhynchus. Fortuner and Luc (1987), in theirreappraisal of Tylenchina, included Tylenchorhynchus under the familyBelonolaimidae, subfamily Telotylenchinae, and recognized 129 valid species, defining Tylenchorhynchus as having 2 to 5 lines in the lateral field, which wassometimes areolated. Mahajan (1988) gave a diagnostic compendium to speciesof Tylenchorhynchus and included 89 valid species in the genus. Esser (1991)listed 257 nominal species in his checklist of the genus. Brzeski and Dolinski(1998) compiled a compendium containing 177 species with 2 to 5 lines in thelateral field. The taxonomy of stunt nematodes has been advanced throughscanning electron microscopy (Fortuner and Luc, 1987; Powers, 1983; Powers etal., 1983; Siddiqi, 1986). As a result of the inevitable taxonomic changes, development of a dichotomous key to Tylenchorhynchus spp. has becomeincreasingly difficult. Many species previously included in Tylenchorhynchushave been placed in newer genera, and numerous species within related generahave been either shifted to Tylenchorhynchus or synonymized with otherspecies. The most important character used in distinguishing these genera is thenumber of lateral lines or incisures, which can range from three to six. Thehistory of the genus and the taxonomic changes to stunt nematodes

26
(Tylenchorhynchus spp.) were proposed by different workers and aresummarized by Handoo (2000) and Handoo et al. (2007). Handoo (2000)considered those species of Tylenchorhynchus having only four lines in thelateral field to prepare a diagnostic compendium for identification of 111 validspecies.
1. Female tail subdigitate, phasmids enlarged-----------------------ScutylenchusJairajpuri, 1971
Female tail not subdigitate, phasmids normal------------------------------------------------------2
2. Female tail acute or subacute, terminus finely rounded------------TetylenchusFilipjev, 1936
Female tail bluntly conical to cylindrical, terminus bluntly pointed tohemispherical-------3
3. Stylet knobs characteristically angular, protractor muscles of stylet attached toinner wall of labial cavity, epitigma present at vulva-------------------Nagelus Thorne and Malek, 1968
Stylet knobs rounded, protractor muscles of stylet attached to base ofcephalic framework, vulva seldom with epitigma--------------------------------------------------------------------------4
4. Labial region with prominent perioral disc, stylet characteristically slender----------------------------------------------------------------------------------GeocenamusThorne and Malek, 1968
Labial region without perioral disc, stylet moderately robust to heavy-------------------------5
5. Lateral field with 3-4 incisures-----------------------------------------------------------------------6
Lateral field with 5-6 incisures-----------------------------------------------------------------------7
6. Lateral field areolated, with 3 incisures-------------------------UliginotylenchusSiddiqi, 1971
Lateral field usually with 4 incisures, never areolated when with 3 incisures-------------------------------------------------------------------------------------------Tylenchorhynchus Cobb, 1913
7. Lateral field with 5 incisures-------------------------------------------QuinisulciusSiddiqi, 1971

27
Lateral field with 6 incisures----------------------------------------------MerliniusSiddiqi, 1970
TylenchorhynchusType species:Tylenchorhynchus cylindricus Cobb, 1913Other species:T. aduncus de Guiran, 1967T. aerolatus Tobar Jime´nez, 1970T. agri Ferris, 1963T. allii Khurma & Mahajan, 1987T. alami Shaw & Khan, 1996T. amgi Kumar, 1981T. ancorastyletus Ivanova, 1983T. annulatus (Cassidy, 1930) Golden, 1971T. antarcticus Wouts & Sher, 1981T. aspericutis Knobloch, 1975T. badliensis Saha & Khan, 1982T. bicaudatus Khakimov, 1973T. bohrrensis Gupta & Uma, 1980T. brassicae Siddiqi, 1961T. brevilineatus Williams, 1960T. bryobius Sturhan, 1966T. canalis Thorne & Malek, 1968T. clarus Allen, 1955T. clavicaudatus Seinhorst, 1963T. clavus Khan, 1990T. claytoni Steiner, 1937T. coffeae Siddiqi & Basir, 1959T. contractus Loof, 1964T. crassicaudatus Williams, 1960T. cristatus Ivanova, 1983T. crotoni Pathak & Siddiqi, 1997T. cuticaudatus Ray & Das, 1983T. cynodoni Kumar, 1981T. delhiensis Chawla, Bhamburkar, Khan &Prasad, 1968

28
T. depressus Jairajpuri, 1982T. dewaeli Kleynhans, 1992T. dubius (Butschli, 1873) Filipjev, 1936T. ebriensis Seinhorst, 1963T. elegans Siddiqi, 1961T. eremicolus Allen, 1955T. eroshenkoi Siddiqi, 1986T. estherae Kleynhans, 1992T. ewingi Hopper, 1959T. georgiensis Eliashvili, 1971T. goffarti Sturhan, 1966T. goldeni Rashid & Singh, 1982T. gossypii Nasira & Maqbool, 1996T. graciliformis Siddiqi & Siddiqui, 1983T. haki Fotedar & Mahajan, 1971T. hordei Khan, 1972T. huesingi Paetzold, 1958T. ibericus Mahajan & Nombela, 1986T. iphilus (Minagawa, 1995) n. comb. syn. Bitylenchus iphilus Minagawa, 1995T. irregularis Wu, 1969T. ismaili Azmi & Ahmad, 1989T. kamlae Shaw & Khan, 1996T. kashmirensis Mahajan, 1974T. kegasawai Minagawa, 1995T. kegenicus Litvinova, 1946T. kidwaii Rashid & Heyns, 1990T. lamilliferus (de Man, 1880) Filipjev, 1936T. latus Allen, 1955T. leucaenus Azmi, 1991T. leviterminalis (Siddiqi, Mukherjee & Dasgupta,1982) Siddiqi, 1986T. malinus Lin, 1992T. manubriatus Litvinova, 1946T. mashhoodi Siddiqi & Basir, 1959T. maximus Allen, 1955T. mexicanus Knobloch & Laughlin, 1973

29
T. microcephalus Siddiqi & Patel, 1990T. microconus Siddiqi, Mukherjee & Dasgupta,1982T. musae Kumar, 1981T. namibiensis Rashid & Heyns, 1990T. natalensis Kleynhans, 1984T. neoclavicaudatus Mathur, Sanwal & Lal, 1979T. nordiensis Khan & Nanjappa, 1974T. novenus Nobbs, 1989T. nudus Allen, 1955T. olereaceae Gupta & Uma, 1981T. pachys Thorne & Malek, 1968T. paracanalis Khan, 1991T. paranudus Phukan & Sanwal, 1982T. paratriversus Brzeski, 1991T. parvus Allen, 1955T. paulettae Bloemers & Wanless, 1998T. penniseti Gupta & Uma, 1980T. projectus Khan, 1990T. punensis Khan & Darekar, 1979T. quaidi Golden, Maqbool & Handoo, 1987T. queirozi Monteiro & Lordello, 1976T. robustus Thorne & Malek, 1968T. rosei Zarina & Maqbool, 1991T. sacchari Sivakumar & Muthukrishnan,1983T. sanwali Kumar, 1982T. siccus Nobbs, 1989T. silvaticus Ferris, 1963T. solani Gupta & Uma, 1982T. spinaceae Singh, 1976T. striatus Allen, 1955T. swarupi Singh & Khera, 1978T. tarjani Andra´ssy, 1969T. teeni Hashim, 1984T. tenuicaudatus Wouts & Sher, 1981T. thermophilus Golden, Baldwin & Mundo-Ocampo, 1995T. tobari Sauer & Annells, 1981

30
T. tritici Golden, Maqbool & Handoo, 1987T. tuberosus Zarina & Maqbool, 1994T. usmanensis Khurma & Mahajan, 1987T. variacaudatus Singh, 1971T. velatus Sauer & Annells, 1981T. ventrosignatus Tobar Jime´nez, 1969T. vishwanathensis Pathak & Siddiqi, 1996T. vulgaris Upadhyay, Swarup & Sethi, 1972T. wilskii Kornobis, 1980T. zambiensis Venditti & Noel, 1995
Tylenchorhynchus
1. Lateral field with 3 incisures---------------------2
Lateral field with 4 incisures---------------------42. Cuticle of lateral fields extended on female tail, stylet 16-17 µ long, lip region
offset-------- -------------------------------------------------------T. divittatusSiddiqi, 1961
Cuticle of lateral fields normal to female tail, stylet 20-28 µ long, lip regioncontinuous------------------------------------------------------------3
3. Tail with 9 annules, cephalic framework heavily sclerotized, anterior surfaceof stylet knobs inclined anteriorly------------------------- T. sculptus Seinhorst, 1963
Tail with 13-15 annules, cephalic framework lightly sclerotized, anteriorsurface of stylet knobs inclined laterally-----------------------T. triglyphusSeinhorst, 1963 (syn. T. chonai Sethi and Swarup, 1968)
4. Body with longitudinal striae---------------------5 Body without longitudinal striae-----------------125. Body with 29 longitudinal striae at midbody, tail with 10 annules--T. claytoni
Steiner, 1937 Body with 20 or less longitudinal striae, tail with 13 or more annules-----------
-------------66. Lip region with 1-2 annules, tail with13 annules, anterior surface of stylet
knobs inclined anteriorly-------------------------------------------- T. pachysThorne and Malek, 1968
Lip region with 5-7 annules, tail with 22 or more annules, anterior surface ofstylet knobs inclined posteriorly or laterally--------------------7

31
7. Tail terminus annulated-----------------------------8 Tail terminus smooth--------------------------------98. Stylet 24-28µm long, T/ABW = 1.9, V% = 46-53---------T. lamelliferus (de
Man, 1880) Filipjev, 1936 Stylet 21µ long, T/ABW = 2.2-2.6, V% = 54-56------------- T. judithae
Andrássy, 19629. Stylet 16µ long, tail shape sub cylindrical, tail terminus hemispherical----------
------------------------------------------------------------------------ T. brevilineatusWilliams, 1960
Stylet 19-27 µ long, tail shape conoid, tail terminus bluntly pointed--------------------------10
10. Body with 16-20 longitudinal striae, tail with 53-55 annules, body 0.77-0.94mm long-------------------------------------------------- T. microphasmis Loof, 1960
Body with 12 longitudinal striae, tail with 22-50 annules, body 0.54-0.77mm long------11
11. Bursa distally recurved, female tail with 22 annules, spicules 24 µ long---------------------------------------------------------------------- T. phaseoli Sethi andSwarup, 1968
Bursa continuing to end of tail, female tail with 30-50 annules, spicules 27 µlong--------------------------------------------------------- T. sulcatus de Guiran, 1967
12. Tail terminus annulated---------------------13 Tail terminus smooth------------------------2313. Stylet 27-38 µ long--------------------------14 Stylet 17-24 µ long--------------------------1514. Tail with 13-14 annules, T/ABW=1.1, stylet 37-38 µ long--------T.
brevicaudatus Hooper, 1959 Tail with 31-33 annules, T/ABW=2.5, stylet 27-30µ long-------T.
magnicauda (Thorne,1935) Filipjev, 193615. Tail with 20-29 annules, lip region with 4 annules--------------------16 Tail with 31-66 annules, lip region with 3 or 5-7 annules------------1716. Cuticular annulations irregular, lip region offset, anterior surface of stylet
knobs inclined posteriorly------------------------------------------------------------------T. irregularis Wu, 1969

32
Cuticular annulations coarse, lip region continuous, nterior surface of styletknobs inclined anteriorly-----------------------------------------------------T.eremicolus Allen, 1955
Tail with 31-48 annules, stylet knobs inclined posteriorly or laterally----------------------18
18. Stylet 21-24 µ long----------------------------------------19 Stylet 17-19 µ long----------------------------------------2019. Tail subcylindrical, tail terminus subhemispherical, body 0.76-0.86 mm
20. Tail shape clavate, lip region with 4 annules, T/ABW=3.8-4.0----------T.clavicauda Seinhorst, 1968 (syn. Clavicaudatus Seinhorst, 1963)
Tail shape cylindrical to sub cylindrical, lip region with 5-7 annules, T/ABW= 2.0-3.2-----------------------------------------------------------------------21
21. T/ABW = 2.0, spicule 30 µ long, lip region with 5 annules----- T. huesingiPaetzold, 1958
T/ABW = 3.0-3.2, spicule 12-24 µ long, lip region with 7 annules-------------------------22
23. Tail with 40-58 annules----------------------------------------24 Tail with 8-40 annules------------------------------------------2624. Tail with 40-45 annules, stylet 23µ long, tail cylindrical, tail terminus
Tail with 45-58 annules, stylet 28-39 µ long, tail sub-cylindrical or conoid, tail terminus sub-hemispherical-----------------------------------------------25
17. Tail with 66 annules, anterior surface of stylet knobs inclined anteriorly--------------------------------------------------------T. canalis Thorne and Malek, 1968
long--------------------------------------------------------T. bryobius Sturhan, 1966
Tail cylindrical, tail terminus hemispherical, body 0.98-1.40 mm long-------------------------------------------------------------------- -T. maximus Allen, 1955
22. Tail sub cylindrical, tail terminus sub-hemispherical, tail with 46-48annules----- T. dubius (Bütschli, 1873) Filipjev, 1936
hemispherical--------------------------------T. robustus Thorne and Malek, 1968
Tail cylindrical, tail terminus hemispherical, tail with 35-43 annules------------ T. parvus, Allen, 1955

33
25. Tail with 45-47 annules, stylet 31-39 µ long, T/ABW = 2.2--- T. galeatusLitvinova, 1946
Tail with 51-58 annules, stylet 28-31 µ long, T/ABW = 3.3-4.8------T.kegenicus Litvinova, 1946
26. Stylet 23-27 µ long---------------------------------------------27 Stylet 11-23 µ long---------------------------------------------2927. Tail with 17-23 annules, lip region continuous, tail cylindrical, tail terminus
Tail with 14-16 annules, lip region offset, tail sub-cylindrical to conoid, tailterminus sub-hemispherical to bluntly pointed----------------------28
29. Lip region with only 2 annules-------------------------------30 Lip region with 3 or more annules---------------------------3130. Tail conoid, tail terminus hemispherical, T/ABW = 2.8, tail with 18-20
31. Tail with 8-15 annules------------------------------------32 Tail with 15-40 annules-----------------------------------3432. Tail cylindrical, tail terminus hemispherical, tail with 8-10 annules-----------
33. Lip region offset, T/ABW = 2.2-------------------------T. latus Allen, 1955 Lip region continuous, T/ABW = 2.6--------T. clarus Allen, 1955 (syn. T.
tener Erzhanova, 196434. Lip region with 3-4 annules-------------------------------35 Lip region with 5-7 annules-------------------------------40
hemispherical----------------------------------------- T. silvaticus Ferris, 1963
28. Tail conoid, tail terminus bluntly pointed, body 0.65-0.99 mm long, V% =54-65-------------------------------------------------------T. cylindricus Cobb, 1913
Tail sub-cylindrical, tail terminus sub-hemispherical, body 0.50-0.62 mmlong, V%=52-54------------------------------------T. tarjani Andrássy, 1969
annules---------------------------------------------------- T. nudus Allen, 1955 Tail sub-cylindrical, tail terminus sub-hemispherical, T/ABW = 4.0, tail with
29 annules---------------------------------------T. delhiensis Chawla et al., 1968
Tail conoid, tail terminus bluntly pointed, tail with 10-15annules-------------33
-----------------------------------------------------T. georgiensis Eliashvili, 1971

34
35. Stylet 11-14 µ long, tail with 28-32 annules-------------T. ventrosignatusTobar-J.,1969
Stylet 16-23 µ long, tail with 15-33 annules-------------3636. Lip region well setoff, tail conoid, tail terminus bluntly pointed---------------
T. brassicae Siddiqi, 1961 Lip region continuous or only weakly setoff, tail cylindrical to sub-
cylindrical, tail terminus hemispherical to sub-hemispherical------------3737. Spermatheca and males not observed----------------------T. martini Fielding,
1956 Spermatheca and males present-----------------------------3838. Stylet 20-23µ long, spicule 22-25µ long, anterior surface of stylet knobs
39. Stylet 16-19µ long, tail terminus sub-hemispherical to hemispherical---------T. mashhoodi Siddiqi and Basir, 1959
Stylet 18-20 µ long, tail terminus sub-hemispherical-----------------T. ewingiHopper, 1959
40. Lip region well offset, stylet 13-15 µ long----------------------------T. goffartiSturhan, 1966
Lip region continuous or weakly set off, stylet 15-22 µ long---------------------------------41
41. Tail with 35-38 annules, T/ABW = 3.3-----------------------T. manubriatusLitvinova, 1946
42. Stylet 21-22 µ long, cephalic framework heavily sclerotized----------T.ebriensis Seinhorst, 1963
Stylet 15-18 µ long, cephalic framework lightly to moderately sclerotized---------------43
inclined laterally--------------------------------------------------T. agri Ferris, 1963
Stylet 16-20 µ long, spicule 18-22 µ long, anterior surface of stylet knobsinclined posteriorly-----------------------------------------------------39
Tail with 16-27 annules, T/ABW = 2.0-2.8------------------------------------42
43. Lateral field areolated, lip region set off, cephalic framework moderatelysclerotized--------------------------------------------- T. areolatus Tobar-J., 1970
Lateral field not areolated, lip region continuous, cephalic frameworklightly sclerotized----------------------------------------44

35
Tylenchorhynchus
1. Stylet 10–31 m long; tail terminus smooth----------------------------- 21a. Stylet 13–30 m long; tail terminus annulated ------------------------ 802(1). Stylet 10–15 m long --------------------------------------------------- 32a. Stylet 15–23 m long ----------------------------------------------------- 132b. Stylet 23–31 m long------------------------------------------------------ 743(2). Lip region 0–4 annules ------------------------------------------------- 43a. Lip region 4–7 annules ---------------------------------------------------- 94(3). Lip region continuous with 0–4 annules------------------------------- 54a. Lip region set off or conoid with 1–4 annules -------------------------- 75(4). Lip region without any annules; tail clavate with hemispherical terminus
8(7). Lip region with 4 annules; stylet 11–14 m long; tail sub-cylindrical with abluntly pointed terminus bearing 28–32 annules; wave-like structures
Body 0.42-0.63 mm long, proximal end of gubernaculums only slightlycurved anteriad ---------------------------------------- T. contractus Loof, 1964
44. Body 0.58-0.72 mm long, proximal end of gubernaculum curved anteriadalmost 90º-------------------------------------------------- T. striatus Allen, 1955
bearing 20–26 annules ------------------------------------------------ T. cynodoni
5a. Lip region with 2–4 annules; tail conoid to cylinidrical with a bluntly pointed terminus bearing 14–27 annules---------------------------------------------- 6
6(5a). Stylet 11–13 m long; tail conoid with a bluntly pointed terminus bearing14–19 annules --------------------------------------------- -----------T. nordiensis
6a. Stylet 14–15 m long; tail cylindrical with bluntly pointed terminus bearing24–27 annules -------- --------------------------------------------------T. punensis
7(4a). Lip region set off with 2–4 annules; tail cylindrical to sub-cylindrical witha bluntly rounded to bluntly pointed terminus bearing 15–32 annules------ 8
7a. Lip region conoid with 1–2 annules; tail sub-cylindrical with a bluntlyrounded terminus bearing 13 annules---------------- ----------------- T. pachys

36
present near vulva on ventral side of body of mature females------------------------------------------------------ T. ventrosignatus
9a. Stylet 12–15 m long ---------------------------------------------------- 1010(9a). Lip region 4–5 annules --------------------------------------------- 1110a. Lip region 5–7 annules ------------------------------------------------- 1211(10). Tail conoid with a bluntly pointed terminus bearing 38 annules -----------
------- T. oleraceae11a. Tail sub-cylindrical with a conoid terminus bearing 21–32 annules ----------
------- T. zambiensis12(10a). Lip region sunken, dome shaped with 5–6 annules; tail cylindrical with
12a. Lip region set off, not sunken or dome-shaped, with 6–7 annules; tail sub-cylindrical with a bluntly pointed terminus; lateral field not areolated -------------T. goffarti
8a. Lip region with 2–3 annules; stylet 12–15 m long; tail cylindrical with abluntly pointed terminus bearing15–23 annules; no such wave-likestructures present ---- T. tritici
9(3a). Stylet 10–11 m long --------------------------------------------- T. depressus
a conoid terminus; lateral field areolated ---------------------- -------T. quaidi
bluntly pointed or conoid terminus ----------------------------------- 19
body) ------------- 4414(13). Lip region 0–3 annules ------------------------------------------- 1514a. Lip region 3–6 annules ----------------------------------------------- 3615(14). Stylet 15–18 m long --------------------------------------------- 1615a. Stylet 18–23 m long ------------------------------------------------- 2416(15). Tail 14–20 annules ------------------------------------------------ 1716a. Tail 20–36 annules ---------------------------------------------------- 2317(16). Tail clavate to sub-clavate with a hemispherical to conoid terminus ------
13(2a). Lip region continuous -------------------------------------- 1413a. Lip region set off (except T. paratriversus sometimes continuous with
-------------- 817a. Tail sub-cylindrical to conoid or bluntly rounded with a hemispherical to

37
18a. Lip region with 2 annules; tail sub-clavate with a conoid terminus bearing14–17 annules T. rosei
19a. Tail conoid to bluntlyrounded with a hemispherical to bluntly pointedterminus ----------21
20(19). Tail sub-cylindrical with a hemispherical terminus bearing 17–21annules; spicule and gubernaculum longer, measuring 23–27 m and 12–14
m, respectively ----------- T. bohrrensis20a. Tail sub-cylindrical with a bluntly pointed terminus bearing 15–17 annules;
spicule and gubernaculum shorter, measuring 22 m and 10 m, respectively--------------------- T. penniseti
21(19a). Tail conoid with a bluntly pointed or hemispherical terminus; malesabsent ------- 22
21a. Tail bluntly rounded with a hemispherical terminus; males present -------------------- T. gossypii
22(21). Lip region with 2 annules; stylet 17–18 m long with anteriorly directedknobs; tail conoid with a bluntly pointed terminus bearing 14–15 annules -------------------------- T. variacaudatus
22a. Lip region with 3 annules; stylet 16–18 m long with posteriorly directedknobs; tail conoid with a hemispherical terminus bearing 14–20 annules -------------- T. haki
23(16a). Lip region with 2–3 annules; tail sub-cylindrical with a hemisphericalterminus bearing 20–36 annules; vulva at 55–59%; males present-------------------- T. goldeni
23a. Lip region with 2 annules; tail sub-cylindrical with a bluntly pointedterminus bearing 29 annules; vulva at 47–48%; males absent------------------------- T. delhiensis
24(15a). Lip region 0–2 annules ---------------------------------------- 2524a. Lip region 2–3 annules --------------------------------------------- 3225(24). Body length less than 1.00 mm; stylet 18–23 m long; tail with 16–35
18(17). Lip region with 3 annules; tail clavate with a hemispherical terminusbearing 18–22 annules --------------------------------------------------T. sacchari
19(17a). Tail sub-cylindrical with a hemispherical to bluntly pointed terminus ---------------- 20
annules ------------ 26

38
25a. Body length about 1.00 mm or longer; stylet 23 m long; tail with 40–45annules ---T. robustus
26a. Tail sub-clavate with a rounded terminus bearing 25–35 annules--------------- T. microcephalus
27(26). Tail conoid with a hemispherical terminus bearing 18–20 annules -------------------- T. nudus
28(27a). Lip region 0 annules (except T. leviterminalis sometimes with 1annule)------------------ 29
28a. Lip region 1–2 annules -------------------------------------------- 3129(28). Stylet 12–22 m long with anteriorly directed knobs; males absent -------
------------------ 3029a. Stylet 17–19 m long with posteriorly directed knobs; males present --------
-- T. leviterminalis30(29). Stylet 19–20 m long; tail sub-cylindrical with a subhemispherical
31(28a). Lip region with 2 annules; stylet 17–20 m long with posteriorlydirected knobs; tail sub-cylindrical with a bluntly pointed terminus bearing19 annules ----------------------- T. coffeae
31a. Lip region with 1–2 annules; stylet 18–19 m long with anteriorly directedknobs; tail sub-cylindrical with a bluntly pointed terminus bearing 16–28annules ------------------- T. musae
32(24a). Tail clavate with a hemispherical to rounded terminus bearing 15–50annules----------- 33
32a. Tail conical or sub-cylindrical with a bluntly pointed terminus bearing 15–25 annules------ 35
26(25). Tail sub-cylindrical or conoid with hemispherical to sub-hemisphericalor bluntly pointed terminus bearing 12–28 annules ------------------ ------- 27
27a. Tail sub-cylindrical with a hemispherical to subhemispherical or bluntlypointed terminus bearing 12–28 annules ----------------------------------- --- 28
terminus bearing 16–20 annules ------------------------------- --------- T. amgi
30a. Stylet 20–22 m long; tail sub-cylindrical with a hemispherical terminusbearing 12–19 annules -------------------------------------- --------- T. tuberosus
33(32). Tail 15–21 annules ------------------------------------- ----- T. kegasawai

39
T. neoclavicaudatus35(32a). Stylet 18–20 m long; tail sub-cylindrical with a bluntly pointed
36(14a). Stylet 15–20 m long --------------------------------------- 3736a. Stylet 20–22 m long -------------------------------------------- 4337(36). Lip region 3–4 annules -------------------------------------- 3837a. Lip region 5–6 annules ------------------------------------------ 4038(37). Tail conoid with a bluntly pointed terminus bearing 21–34 annules ------
---- T. thermophilus38a. Tail cylindrical to subcylindrical with a bluntly pointed to bluntly rounded
39(38a). Stylet 16–20 m long; tail cylindrical with a bluntly pointed terminusbearing 14–29 annules ---------------------------------------------------------------------- T. mashhoodi
39a. Stylet 15–18 m long; tail sub-cylindrical with a bluntly rounded terminusbearing 22–23 annules ----------------------------------------------------------- T.elegans
40(37a). Tail conoid with a bluntly pointed terminus bearing 10–23 annules -------------------------- 41
40a. Tail sub-cylindrical with a bluntly pointed or hemispherical terminusbearing 20–38 annules -- 42
34a. Stylet 20–23 m long; tail with 32–50 annules --------------------------------
33a. Tail 31–50 annules ---------------------------------------------- 34 34(33a). Stylet 18–19 m long; tailwith 31 annules ------
terminus bearing 15–19 annules - ---------------------------------------T. ewingi
35a. Stylet 19–21 m long; tail conical bearing 25annules--------------------------T. alami
terminus bearing 14–29 annules------------------------------------------------39
41a. Lip region with 5–6 annules; stylet 16–18 m long with laterally directedknobs; tail with 20–23 annules --------------------------------------T. contractus
41(40). Lip region with 5 annules; stylet 15–18 m long with anteriorly directedknobs; tail with 10–20 annules ----------------------------------------- T. clarus

40
42(40a). Stylet 16–17 m long with anteriorly directed knobs; tail sub-cylindrical with a bluntly pointed terminus bearing 20–27 annules ------------------- T. striatus
43(36a). Lip region with 5 annules; stylet 21–22 m long with posteriorlydirected knobs; tail conoid with a bluntly pointed terminus bearing 25annules ------- T. ebriensis
44(13a). Lip region 0–3 annules ------------------------------------ 4544a. Lip region 3–6 annules ------------------------------------------ 5044b. Lip region 6–10 annules ---------------------------------------- 6945(44). Lip region smooth, 0 annules ------------------------------- 4645a. Lip region 2–3 annules ------------------------------------------ 4746(45). Stylet 18–21 m long; tail clavate with a hemispherical terminus bearing
47(45a). Tail 11–17 annules ------------------------------------------- 4847a. Tail 17–24 annules ------------------------------------------------ 4948(47). Lip region with 3 annules; tail conoid with a bluntly pointed terminus
49(47a). Stylet 17–21 m long with laterally directed knobs; tail sub-cylindricalwith a hemispherical
42a. Stylet 18 m long with posteriorly directed knobs; tail sub-cylindrical witha hemispherical terminus bearing 35–38 annules ------------- T. manubriatus
43a. Lip region with 4 annules; stylet 20–22 m long with anteriorly directedknobs; tail conoid with a conoid terminus bearing 13 annules---T. projectus
14–25 annules --------------------------------------------------------T. paranudus
46a. Stylet 23 m long; tail cylindrical with a deeply indented terminus bearing26–27 annules ------------------------------------------------------ T. bicaudatus
bearing 11–12 annules ---------------------------------------------- T. aspericutis
48a. Lip region with 2 annules; tail conoid with a bluntly rounded terminusbearing 17 annule ----------------------------------------------T.vishwanathensis
terminus bearing 18–24 annules -----------T. annulatus

41
---------------------- 5951(50). Stylet 15–18 m long ---------------------------------------- 5251a. Stylet 18–23 m long -------------------------------------------- 5552(51). Tail 10–18 annules -------------------------------------------- 5352a. Tail 18–49 annules ------------------------------------------------ 5453(52). Tail conoid with a bluntly pointed terminus bearing 10–15 annules;
55(51a). Tail conoid with a bluntly rounded to hemispherical or bluntly pointedterminus----------- 56
49a. Stylet 17–19 m long with anteriorly directed knobs; tail conoid with abluntly pointed terminus bearing 17–22 annules --------------- T.badliensis
50(44a). Lip region 3–4 annules ------------------------------------- 5150a. Lip region 4–6 annules (except T. paratriversus sometimes with 3 annules)-
males absent ------------------------------------------------- -----T. microconus
53a. Tail sub-cylindrical with a bluntly pointed terminus bearing 12–18 annules;males present ------------------------------------- ------------------ T. spinaceae
54(52a). Tail with 18–33 annules; sinuous canals present in the region ofintestine, extending back to the tail terminus ------ ------------ T. paracanalis
54a. Tail with 49 annules; no such sinuous canals present ----------T. brassicae
55a. Tail sub-cylindrical to conical with a hemispherical terminus------------------ 58
56(55). Body with 29 longitudinal striae at mid-body; stylet 18–21 m long; tailconoid with a bluntly rounded terminus bearing 9–20 annules-------------- ----T. claytoni
56a. Body without longitudinal striae ------------------------------ --- 5757(56a). Stylet 19–21 m long with posteriorly directed knobs; tail conoid with
a bluntly rounded terminus bearing 18–22 annules ------------ T. mexicanus
57a. Stylet 18–20 m long with anteriorly directed knobs; tail conoid with ahemispherical terminus bearing 9–19 annules ------------- T. ancorastyletus

42
58a. Stylet 19–21 m; tail conical with 21 annules; c = 3–5; males absent ------------------ T. kamlae
forward; tail conoid with a bluntly rounded terminus bearing 14–15 annules----- T. latus
66(59a). Tail 8–38 annules --------------------------------------------- 6766a. Tail 42–46 annules ------------------------------------------------- T. hordei
58(55a). Stylet 20–23 m long; tail sub-cylindrical with a hemisphericalterminus bearing 18–26 annules; c = 2.6; males present ----------- T. agri
62a. Lip region with 4–5 annules; stylet knobs rounded; tail conoid with a bluntterminus bearing 21 annules -------------------- --------------------- T. crotoni
63(61a). Stylet 16–17 m long; tail conoid with a bluntly pointed terminusbearing 34 annules--------------------------------------------------- ---T. solani
63a. Stylet 15–16 m long; tail conoid with a narrow terminus bearing 34–40annules ---------------------------------------------------------------T. usmanensis
64(60a). Tail 16–22 annules --------------------------------------------- 6564a. Tail 32–49 annules--------------------------- -- T.
65(64). Stylet 15–17 m long with posteriorly directed knobs; tailcylindrical with a bluntly pointed terminus ------------- --- T. aerolatus
------------------- 6160a. Tail sub-cylindrical with a hemispherical or bluntly pointed terminus----
61(60). Tail 14–21 annules; males absent---------------------------- 6261a. Tail 34–40 annules; males present ------------------------------ 6362(61). Lip region with 6 annules; stylet knobs with anterior margins directed
59(50a). Stylet 15–18 m long --------------------------------------- 6059a. Stylet 18–23 m long -------------------------------------------- 6660(59). Tail conoid with a bluntly rounded to blunt or narrow terminus -
65a. Stylet 17–18 m long with anteriorly directed knobs; tail sub-cylindricalwith a hemispherical terminus--------------------------------- T. graciliformis

43
67a. Tail cylindrical with a hemispherical terminus bearing 8–10 annules ---------------- T. georgiensis
68(67). Stylet 18–20 m long with posteriorly directed knobs; tail conoid with abluntly pointed terminus bearing 19–27 annules; bursa reaching male tailend -------------------------- T. aduncus
68a. Stylet 21–23 m long with anteriorly directed more or less anchor-shapedknobs; tail conical to almost funnel shaped with variously rounded terminusbearing 27–38 annules; bursa not reaching male tail end------------------------------------------------T. paratriversus
69(44b). Stylet 15–17 m long--------------------------------------------- 7069a. Stylet 17–23 m long -------------------------------------------------- 7170(69). Tail sub-cylindrical with a bluntly pointed terminus bearing 35–42
annules --------T. vulgaris70a. Tail sub-cylindrical with a hemispherical terminus bearing 34–47 annules --
--------------- T. allii71(69a). Lip region with 8–10 annules; tail sub-cylindrical with a hemispherical
71a. Lip region with 6–9 annules; tail conoid, sub-cylindrical or narrow with arounded to bluntly rounded terminus bearing 27–66 annules; males present --------- 72
72(71a). Lip region with 6–7 annules; stylet 19–23 m long; tail conoid with abluntly rounded terminus bearing 28–66 annules; gubernaculum peculiarlyshaped like a hammer----T. estherae
72a. Lip region with 7–9 annules; stylet 11–23 m long; tail sub-cylindrical ornarrow with a rounded terminus bearing 27–51 annules ---------------------------------------73
73(72a). Lip region with 7–8 annules; stylet 17–23 m long; tail narrow with arounded terminus bearing 39–51 annules ----------------------------------------T. novenus
67(66). Tail conoid or conical with a bluntly pointed to pointed or variouslyrounded terminus bearing 19–38 annules------------------------------------- 68
68b. Stylet 21–22 m long with anteriorly directed knobs; tail conical to conoidwith a pointed to finely rounded terminus bearing 27 annules; bursareaching male tail end ------- --- T. paulettae
terminus bearing 50 annules; males absent ----------------- ----- T. tobari

44
73a. Lip region with 6–9 annules; stylet 16–20 m long; tail sub-cylindrical witha rounded terminus bearing 27–51 annules ----------------------------------------T. namibiensis
74(2b). Lip region continuous -------------------------------------- 7574a. Lip region set off------------------------------------------------ 7875(74). Lip region 3–4 annules ------------------------------------- 7675a. Lip region 6–10 annules --------------------------------------- 7776(75). Body length longer (L =0.80–1.00 mm); stylet 23–26 m long; tail
cylindrical with a hemispherical terminus bering 17–23 annules; spicule andgubernaculums longer, measuring 28–32 m and 16–19 m, respectively --------------------- T. silvaticus
77(75a). Lip region with 6–7 annules; stylet 25–26 m long; tail conoid with abluntly pointed terminus bearing 18–35 annules; males absent----------------------------------------- T. eroshenkoi
77a. Lip region with 7–10 annules; stylet 28–31 m long; tail sub-cylindricalwith a bluntly pointed terminus bearing 51–58 annules; males present ----------- T. kegenicus
80(1a). Stylet 13–21 m long ---------------------------------------- 81
76a. Body length shorter (L = 0.74–0.79 mm); stylet 24–27 m long; tailelongateclavate bearing 12–19 annules; spicule and gubernaculum shorter, measuring 21–25 m and 11–14 m, respectively ------------- --- T. clavus
78(74a). Lip region with 4–5 annules; stylet 24–29 m long; tail conoid to sub-cylindrical with a bluntly pointed terminus bearing 14–20 annules ------ 79
78a. Lip region with 8 annules; stylet 23–26 m long; tail cylindrical with anacutely pointed terminus bearing 38–65 annules ----------- T. tenuicaudatus
79a. Body length shorter (L =0.50–0.62 mm); stylet 24–25 m long withposteriorly directed knobs; vulva at 52–54%; tail sub-cylindrical with abluntly pointed terminus bearing 14–15 annules --------- ---- T. tarjani
79(78). Body length longer (L =0.65–1.17 mm); stylet 24–29 m long withanteriorly directed knobs; vulva at 51–65%; tail conoid with a bluntlypointed terminus bearing 15–20 annules -------------- ---T. cylindricus

45
80a. Stylet 21–30 m long --------------------------------------------10081(80). Stylet 13–15 m long ----------------------------------------- 8281a. Stylet 16–21 m long --------------------------------------------- 8382(81). Stylet knobs inclined posteriorly; vulva at 52–56%; tail cylindrical with
83(81a). Tail with 13–21 annules ------------------------------------ 8483a. Tail with 23–66 annules ----------------------------------------- 8784(83). Lip region continuous with 3 annules; stylet 20 m long; tail clavate
85(84a). Stylet 17–18 m long; vulva at 55–56%; tail with 18–21 annules ---------------- T. leucaenus
87(83a). Lip region continuous with 4–7 annules------------------------ 8887a. Lip region set off with 2–8 annules ---------------------------------- 9188(87). Lip region continuous with 4–5 annules------------------------- 8988a. Lip region continuous with 6–7 annules----------------------------- 9089(88). Stylet 18–19 m long with posteriorly directed knobs; tail cylindrical
a bluntly pointed terminus --------------------------- ----------- T. swarupi
82a. Stylet knobs inclined anteriorly; vulva at 53–60%; tail cylindrical with ahemispherical terminus ---------------- ------------------ T. cuticaudatus
with a hemispherical terminus ---------------- ----------- T. crassicaudatus
84a. Lip region set off with 3–5 annules; stylet 17–21 m long; tail sub-cylindrical with a narrow to bluntly pointed terminus --------------------- 85
85a. Stylet 17–21 m long; vulva at 54–64%; tail with 13–17 annules ---- - 86
86(85a). Lip region set off with 3 annules; tail sub-cylindrical with a bluntlypointed terminus; vulva at 62–64%; males present ---- T. kashmirensis
86a. Lip region set off with 4–5 annules; tail sub-cylindrical with a narrowterminus; vulva at 54–61%; males absent -------------------------- T. ismailii
with a hemispherical terminus bearing 32–36 annules ------------ T. huesingi

51
direction, making sure that they were resting on the surface of glass slide and not floating on the surface of drop. Microscopic cover glass of 18mm was placed over the specimen and sealed by paraffin wax-ring method (de Maeseneer and d’Herde, 1963).
3.2 Observations on morphological and morphometric variations of Tylenchorhynchus Cobb, 1913 associated with groundnut
Important morphological and morphometric features of taxonomic significance have been studied in detail for each population of Tylenchorhynchus obtained from different groundnut growing areas of Hooghly district. Several populations of Tylenchorhynchus were examined for morphometric characterization for the generation of information on inter-specific and intra-specific variations. Forty nine females, 12 juveniles and 10 males of Tylenchorhynchus were examined for the purpose. The following set of parameters used to characterize nematode species were developed initially by De Man in 1876 and 1880 and have been added to, modified and amended by Cobb (1914), Thorne (1949) and others. They are often known as the De Man Formula or the De Man Indices.
n = number of specimens on which measurements are taken
L = total body length
Tail = portion of body from anus or cloaca to posterior terminus
Ratios
a = body length / greatest body width
b = body length / distance from anterior (head) to oesophago-intestinal valve (cardia)
c = body length / tail length
c' = tail length / anal body width (abw)
Percentage m%= (length of conus/length of stylet) × 100 o %= (distance of dorsal esophageal gland opening from stylet knobs/ stylet length) × 100

51
direction, making sure that they were resting on the surface of glass slide and not floating on the surface of drop. Microscopic cover glass of 18mm was placed over the specimen and sealed by paraffin wax-ring method (de Maeseneer and d’Herde, 1963).
3.2 Observations on morphological and morphometric variations of Tylenchorhynchus Cobb, 1913 associated with groundnut
Important morphological and morphometric features of taxonomic significance have been studied in detail for each population of Tylenchorhynchus obtained from different groundnut growing areas of Hooghly district. Several populations of Tylenchorhynchus were examined for morphometric characterization for the generation of information on inter-specific and intra-specific variations. Forty nine females, 12 juveniles and 10 males of Tylenchorhynchus were examined for the purpose. The following set of parameters used to characterize nematode species were developed initially by De Man in 1876 and 1880 and have been added to, modified and amended by Cobb (1914), Thorne (1949) and others. They are often known as the De Man Formula or the De Man Indices.
n = number of specimens on which measurements are taken
L = total body length
Tail = portion of body from anus or cloaca to posterior terminus
Ratios
a = body length / greatest body width
b = body length / distance from anterior (head) to oesophago-intestinal valve (cardia)
c = body length / tail length
c' = tail length / anal body width (abw)
Percentage m%= (length of conus/length of stylet) × 100 o %= (distance of dorsal esophageal gland opening from stylet knobs/ stylet length) × 100

51
direction, making sure that they were resting on the surface of glass slide and not floating on the surface of drop. Microscopic cover glass of 18mm was placed over the specimen and sealed by paraffin wax-ring method (de Maeseneer and d’Herde, 1963).
3.2 Observations on morphological and morphometric variations of Tylenchorhynchus Cobb, 1913 associated with groundnut
Important morphological and morphometric features of taxonomic significance have been studied in detail for each population of Tylenchorhynchus obtained from different groundnut growing areas of Hooghly district. Several populations of Tylenchorhynchus were examined for morphometric characterization for the generation of information on inter-specific and intra-specific variations. Forty nine females, 12 juveniles and 10 males of Tylenchorhynchus were examined for the purpose. The following set of parameters used to characterize nematode species were developed initially by De Man in 1876 and 1880 and have been added to, modified and amended by Cobb (1914), Thorne (1949) and others. They are often known as the De Man Formula or the De Man Indices.
n = number of specimens on which measurements are taken
L = total body length
Tail = portion of body from anus or cloaca to posterior terminus
Ratios
a = body length / greatest body width
b = body length / distance from anterior (head) to oesophago-intestinal valve (cardia)
c = body length / tail length
c' = tail length / anal body width (abw)
Percentage m%= (length of conus/length of stylet) × 100 o %= (distance of dorsal esophageal gland opening from stylet knobs/ stylet length) × 100

46
90(88a). Stylet 17–18 m long with rounded laterally directed knobs; tailcylindrical with a hemispherical terminus bearing 35–43 annules ------------------- T. parvus
90a. Stylet 19–21 m long with posteriorly directed knobs; tail cylindrical toslightly clavate with a hemisphericalterminus bearing 26–34 annules -------------------- T. malinus
94(93). Body length 1 mm or longer; stylet 20 m long; tail with 66 annules ---------------- T. canalis
94a. Body length less than 1 mm long; stylet 16–19 m long; tail with 22–66annules ------- 95
96(93a). Stylet 17–20 m long; tail clavate with a bluntly pointed terminus ----------------- T. iphilus
89a. Stylet 19–20 m long with anteriorly directed knobs; tail conoid with abluntly pointed terminus bearing 27–29 annules ---------- T. eremicolus
bluntly pointed terminus bearing 30–31 annules --------------- T. sanwali
92a. Stylet 19–21 m long with posteriorly directed knobs; tail conoid with abluntly pointed terminus bearing 20–26 annules ---------- - T. irregularis
93(91a). Stylet 16–20 m long; tail sub-cylindrical with a bluntly pointed orhemispherical terminus -------------------------------------------------------- 94
93a. Stylet 17–21 m long; tail clavate with bluntly pointed terminus orcylindrical to conoid with a hemispherical to rounded terminus ---------- 96
91(87a). Lip region set off with 2–4 annules ----------------------------- - 9291a. Lip region set off with 5–8 annules ---------------------------------- -- 9392(91). Stylet 20 m long with anteriorly directed knobs; tail conoid with a
95(94a). Lip region with 6–8 annules; stylet 18–19 m long; tail sub-cylindricalwith a bluntly pointed terminus bearing 36–66 annules-------------------------T. dubius
95a. Lip region with 4–5 annules; stylet 16–17 m long; tail sub-cylindrical witha hemispherical terminus bearing 22–30 annules --------- ---------T. queirozi

55
High population of Tylenchorhynchus mashhoodi (906/200cc of soil) and Meloidogyne spp. (173.5/200cc of soil) were also recorded from this same locality of Pursurah block. Rotylenchulus reniformis was also recorded in huge number (2642/200cc of soil) from Samanta Road areas of Pursurah block. Presence of Hirschmanniella in the soil may be due to the existence of rice based cropping system in the surveyed areas of the district. Saprozoic nematodes were also recorded in huge number from all the soil samples collected from the district. 4.2 Community analysis of rhizospheric nematodes associated with
groundnut in Hooghly district of West Bengal
Community analysis of major soil inhabiting plant parasitic nematodes associated with groundnut in Hooghly district of West Bengal is presented in the table-2. Six major plant parasitic nematode genera were identified from the rhizosphere of peanut. These were Criconemoides Taylor, 1936, Rotylenchulus Linford and Oliviera, 1940 Tylenchorhynchus Cobb, 1913, Meloidogyne Goeldi, 1892, Hirschmanniella Luc and Goodey, 1964 and Pratylenchus Filipjev, 1936. Criconemoides sp. Taylor, 1936 has been identified as most predominant plant parasitic nematode in groundnut. Rotylenchulus reniformis Linford and Oliviera, 1940 and Tylenchorhynchus mashhoodi Siddiqi and Basir, 1959 were identified as prevalent nematode species under genera Rotylenchulus and Tylenchorhynchus, respectively. Among root-knot nematodes, two species viz. Meloidogyne incognita (Kofoid and White, 1919) Chitwood, 1949 and Meloidogyne javanica (Treub, 1885) Chitwood, 1949 were identified.
Predominance of plant parasitic nematodes associated with peanut in Hooghly district of West Bengal is presented in descending order of their relative prominence value (Table 2). In reference to the density (absolute & relative) and prominence value Criconemoides sp. ranked first followed by Rotylenchulus reniformis, Tylenchorhynchus mashhoodi and Meloidogyne incognita. Considering frequency (absolute & relative) of occurrence Tylenchorhynchus mashhoodi ranked second after Criconemoides sp. Therefore, it is pertinent to workout prominence value for determining the parasitic importance of nematodes in a diverse community (Beals, 1960). The prominence value calculated did not seem to clearly reflect the pathogenic significance of the members of the community.

56
Table 2: Community analysis of rhizospheric nematodes associated with
groundnut in Hooghly district of West Bengal (Based on 43 samples)
Nematode species Absolute frequency
Relative frequency
Absolute density
Relative density
PV RPV
Criconemoides spp. 97.7 19.1 547.0 20.5 540.6 22.3
Rotylenchulus reniformis 51.2 10.0 674.2 25.3 482.3 19.9
Tylenchorhynchus mashhoodi 93.0 18.2 309.9 11.6 298.9 12.3
Meloidogyne incognita 67.4 13.2 61.0 2.3 50.1 2.1
Hirschmanniella spp. 48.8 9.5 37.5 1.4 26.2 1.1
Pratylenchus sp. 20.9 4.1 12.4 0.5 5.7 0.2
Meloidogyne javanica 20.9 4.1 11.3 0.4 5.2 0.2
Mononchids 11.6 2.3 4.4 0.2 1.5 0.1
Saprozoic nematodes 100.0 19.5 1011.8 37.9 1011.8 41.8
Note: PV=Prominence Value; PV=Absolute densityAbsolute Frequency,
RPV=Relative Prominence value
It was reported that Rotylenchulus reniformis is a most frequently occurring nematode parasite of crops in West Bengal at present (Mukhopadhyay and Haque, 1974; Mukherjee and Dasgupta, 1983; Mukhopadhyay and Roy, 2006; Roy et al., 2007; Roy and Mukhopadhyay; 2011). Root lesion nematode, Pratylenchus sp. was also recorded as a parasitic nematode in groundnut. Frequency of Hirschmanniella spp. was recorded as 48.8%. Farmers of the surveyed area used to follow rice-potato-groundnut cropping sequence. Hirschmanniella had been reported to be associated with rice, cabbage, sugarcane, vegetables, weeds and many other crops (Jairajpuri and Baqri, 1991). This finding too corroborates the present observation regarding close association of Hirschmanniella spp. with groundnut in West Bengal. Beside phytonematodes, mononchids (especially Mylonchulus sp.) the predatory nematodes were also found in a frequency of 11.6%. However, saprozoic nematodes comprising of free living dorylaimids and rhabditids were also observed as very prominent member among soil nematode communities of groundnut.

55
High population of Tylenchorhynchus mashhoodi (906/200cc of soil) and Meloidogyne spp. (173.5/200cc of soil) were also recorded from this same locality of Pursurah block. Rotylenchulus reniformis was also recorded in huge number (2642/200cc of soil) from Samanta Road areas of Pursurah block. Presence of Hirschmanniella in the soil may be due to the existence of rice based cropping system in the surveyed areas of the district. Saprozoic nematodes were also recorded in huge number from all the soil samples collected from the district. 4.2 Community analysis of rhizospheric nematodes associated with
groundnut in Hooghly district of West Bengal
Community analysis of major soil inhabiting plant parasitic nematodes associated with groundnut in Hooghly district of West Bengal is presented in the table-2. Six major plant parasitic nematode genera were identified from the rhizosphere of peanut. These were Criconemoides Taylor, 1936, Rotylenchulus Linford and Oliviera, 1940 Tylenchorhynchus Cobb, 1913, Meloidogyne Goeldi, 1892, Hirschmanniella Luc and Goodey, 1964 and Pratylenchus Filipjev, 1936. Criconemoides sp. Taylor, 1936 has been identified as most predominant plant parasitic nematode in groundnut. Rotylenchulus reniformis Linford and Oliviera, 1940 and Tylenchorhynchus mashhoodi Siddiqi and Basir, 1959 were identified as prevalent nematode species under genera Rotylenchulus and Tylenchorhynchus, respectively. Among root-knot nematodes, two species viz. Meloidogyne incognita (Kofoid and White, 1919) Chitwood, 1949 and Meloidogyne javanica (Treub, 1885) Chitwood, 1949 were identified.
Predominance of plant parasitic nematodes associated with peanut in Hooghly district of West Bengal is presented in descending order of their relative prominence value (Table 2). In reference to the density (absolute & relative) and prominence value Criconemoides sp. ranked first followed by Rotylenchulus reniformis, Tylenchorhynchus mashhoodi and Meloidogyne incognita. Considering frequency (absolute & relative) of occurrence Tylenchorhynchus mashhoodi ranked second after Criconemoides sp. Therefore, it is pertinent to workout prominence value for determining the parasitic importance of nematodes in a diverse community (Beals, 1960). The prominence value calculated did not seem to clearly reflect the pathogenic significance of the members of the community.

56
Table 2: Community analysis of rhizospheric nematodes associated with
groundnut in Hooghly district of West Bengal (Based on 43 samples)
Nematode species Absolute frequency
Relative frequency
Absolute density
Relative density
PV RPV
Criconemoides spp. 97.7 19.1 547.0 20.5 540.6 22.3
Rotylenchulus reniformis 51.2 10.0 674.2 25.3 482.3 19.9
Tylenchorhynchus mashhoodi 93.0 18.2 309.9 11.6 298.9 12.3
Meloidogyne incognita 67.4 13.2 61.0 2.3 50.1 2.1
Hirschmanniella spp. 48.8 9.5 37.5 1.4 26.2 1.1
Pratylenchus sp. 20.9 4.1 12.4 0.5 5.7 0.2
Meloidogyne javanica 20.9 4.1 11.3 0.4 5.2 0.2
Mononchids 11.6 2.3 4.4 0.2 1.5 0.1
Saprozoic nematodes 100.0 19.5 1011.8 37.9 1011.8 41.8
Note: PV=Prominence Value; PV=Absolute densityAbsolute Frequency,
RPV=Relative Prominence value
It was reported that Rotylenchulus reniformis is a most frequently occurring nematode parasite of crops in West Bengal at present (Mukhopadhyay and Haque, 1974; Mukherjee and Dasgupta, 1983; Mukhopadhyay and Roy, 2006; Roy et al., 2007; Roy and Mukhopadhyay; 2011). Root lesion nematode, Pratylenchus sp. was also recorded as a parasitic nematode in groundnut. Frequency of Hirschmanniella spp. was recorded as 48.8%. Farmers of the surveyed area used to follow rice-potato-groundnut cropping sequence. Hirschmanniella had been reported to be associated with rice, cabbage, sugarcane, vegetables, weeds and many other crops (Jairajpuri and Baqri, 1991). This finding too corroborates the present observation regarding close association of Hirschmanniella spp. with groundnut in West Bengal. Beside phytonematodes, mononchids (especially Mylonchulus sp.) the predatory nematodes were also found in a frequency of 11.6%. However, saprozoic nematodes comprising of free living dorylaimids and rhabditids were also observed as very prominent member among soil nematode communities of groundnut.

55
High population of Tylenchorhynchus mashhoodi (906/200cc of soil) and Meloidogyne spp. (173.5/200cc of soil) were also recorded from this same locality of Pursurah block. Rotylenchulus reniformis was also recorded in huge number (2642/200cc of soil) from Samanta Road areas of Pursurah block. Presence of Hirschmanniella in the soil may be due to the existence of rice based cropping system in the surveyed areas of the district. Saprozoic nematodes were also recorded in huge number from all the soil samples collected from the district. 4.2 Community analysis of rhizospheric nematodes associated with
groundnut in Hooghly district of West Bengal
Community analysis of major soil inhabiting plant parasitic nematodes associated with groundnut in Hooghly district of West Bengal is presented in the table-2. Six major plant parasitic nematode genera were identified from the rhizosphere of peanut. These were Criconemoides Taylor, 1936, Rotylenchulus Linford and Oliviera, 1940 Tylenchorhynchus Cobb, 1913, Meloidogyne Goeldi, 1892, Hirschmanniella Luc and Goodey, 1964 and Pratylenchus Filipjev, 1936. Criconemoides sp. Taylor, 1936 has been identified as most predominant plant parasitic nematode in groundnut. Rotylenchulus reniformis Linford and Oliviera, 1940 and Tylenchorhynchus mashhoodi Siddiqi and Basir, 1959 were identified as prevalent nematode species under genera Rotylenchulus and Tylenchorhynchus, respectively. Among root-knot nematodes, two species viz. Meloidogyne incognita (Kofoid and White, 1919) Chitwood, 1949 and Meloidogyne javanica (Treub, 1885) Chitwood, 1949 were identified.
Predominance of plant parasitic nematodes associated with peanut in Hooghly district of West Bengal is presented in descending order of their relative prominence value (Table 2). In reference to the density (absolute & relative) and prominence value Criconemoides sp. ranked first followed by Rotylenchulus reniformis, Tylenchorhynchus mashhoodi and Meloidogyne incognita. Considering frequency (absolute & relative) of occurrence Tylenchorhynchus mashhoodi ranked second after Criconemoides sp. Therefore, it is pertinent to workout prominence value for determining the parasitic importance of nematodes in a diverse community (Beals, 1960). The prominence value calculated did not seem to clearly reflect the pathogenic significance of the members of the community.
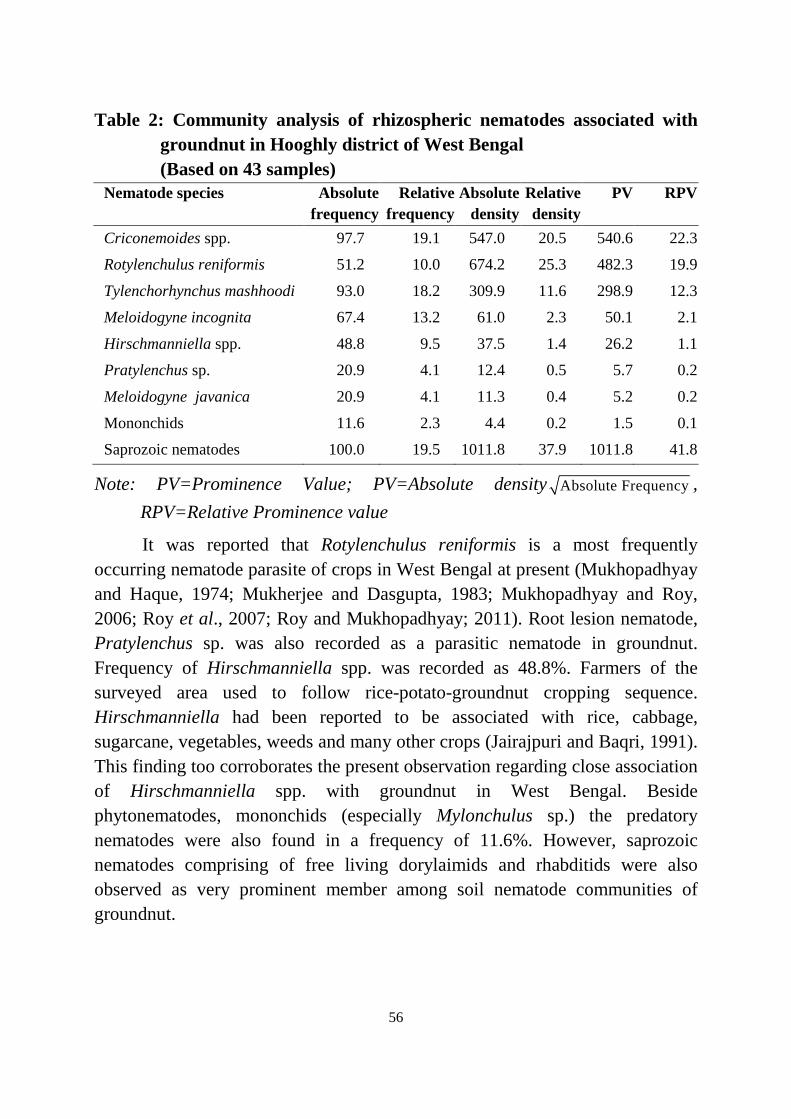
56
Table 2: Community analysis of rhizospheric nematodes associated with
groundnut in Hooghly district of West Bengal (Based on 43 samples)
Nematode species Absolute frequency
Relative frequency
Absolute density
Relative density
PV RPV
Criconemoides spp. 97.7 19.1 547.0 20.5 540.6 22.3
Rotylenchulus reniformis 51.2 10.0 674.2 25.3 482.3 19.9
Tylenchorhynchus mashhoodi 93.0 18.2 309.9 11.6 298.9 12.3
Meloidogyne incognita 67.4 13.2 61.0 2.3 50.1 2.1
Hirschmanniella spp. 48.8 9.5 37.5 1.4 26.2 1.1
Pratylenchus sp. 20.9 4.1 12.4 0.5 5.7 0.2
Meloidogyne javanica 20.9 4.1 11.3 0.4 5.2 0.2
Mononchids 11.6 2.3 4.4 0.2 1.5 0.1
Saprozoic nematodes 100.0 19.5 1011.8 37.9 1011.8 41.8
Note: PV=Prominence Value; PV=Absolute densityAbsolute Frequency,
RPV=Relative Prominence value
It was reported that Rotylenchulus reniformis is a most frequently occurring nematode parasite of crops in West Bengal at present (Mukhopadhyay and Haque, 1974; Mukherjee and Dasgupta, 1983; Mukhopadhyay and Roy, 2006; Roy et al., 2007; Roy and Mukhopadhyay; 2011). Root lesion nematode, Pratylenchus sp. was also recorded as a parasitic nematode in groundnut. Frequency of Hirschmanniella spp. was recorded as 48.8%. Farmers of the surveyed area used to follow rice-potato-groundnut cropping sequence. Hirschmanniella had been reported to be associated with rice, cabbage, sugarcane, vegetables, weeds and many other crops (Jairajpuri and Baqri, 1991). This finding too corroborates the present observation regarding close association of Hirschmanniella spp. with groundnut in West Bengal. Beside phytonematodes, mononchids (especially Mylonchulus sp.) the predatory nematodes were also found in a frequency of 11.6%. However, saprozoic nematodes comprising of free living dorylaimids and rhabditids were also observed as very prominent member among soil nematode communities of groundnut.

47
97a. Tail 41–66 annules ------------------------------------------------------- 9898(97a). Body length more than 1mm; vulva at 49–50%; tail conoid with a
98a. Body length less than 1 mm; vulva at 51–57%; tail cylindrical with ahemispherical terminus - 99
99(98a). Stylet 19–21 m long; males absent; tail with 52–66 annules ---------------------- T. natalensis
99a. Stylet 17–18 m long; males present; tail with 44–57 annules --------------------------------- T. teeni
100(80a). Body length greater than 1mm------------------------------101100a. Body length less than 1 mm--------------------------------------105 101 (100). Stylet 21–24 m long (except T. ibericus with a range of 21–26 m) --
105(100a). Stylet 21–24 m long (except T. velatus with a range of 22–25 m) -----------------------106
96a. Stylet 17–21 m long; tail conoid to cylindrical with a bluntly pointed orrounded to hemispherical terminus ------------------------------------------ 97
97(96a). Tail 23–25 annules ----------------------------- ----- T. cristatus
rounded terminus------------------------------------------------------- T. dewaeli
---------------------102101a. Stylet 24–28 m long --------------------------------- ------------104102 (101). Lip region continuous with 6–9 annules; males present --103102a. Lip region set off with 6 annules; males absent ------------- ----T. ibericus103 (102). Lip region continuous with 7 annules; tail cylindrical with a
hemispherical terminus bearing 38–41 annules--------------------- T. maximus
103a. Lip region continuous with 7–9 annules; tail cylindrical with a roundedterminus bearing 23–29 annules ------------------------------------- T. kidwaii
104 (101a). Lip region continuous with 6 annules; tail conoid with a bluntlyrounded terminus bearing 43 annules --------------------- ----- T. lamilliferus
104a. Lip region set off with 6 annules; tail cylindrical with a hemisphericalterminus bearing 29–34 annules ------------------------------------- T. wilskii

48
107a. Lip region set off with 6–8 annules; stylet 24–30 m long with anteriorlydirected knobs; tail cylindrical with a rounded terminus bearing 16–30annules ------------------------------ T. siccus
105a. Stylet 24–30 m long --------------------------------------- -------107106 (105). Lip region conoid, set off with 5–6 annules; tail subcylindrical with a
bluntly pointed terminus bearing 35–45 annules ------------------ T. bryobius
106a. Lip region set off with 5–6 annules; tail cylindrical with a hemisphericalterminus bearing 25 annules ----------------------------------- ---- T. velatus
107(105a). Lip region set off with 5–7 annules; stylet 24–26 m long withposteriorly directed knobs; tail subcylindrical with a hemispherical terminusbearing 39 annules ----------- -- T. antarcticus

49
This chapter is dealing with survey for collection of nematode populations, counting of the population and preparation of permanent slides for identificationand observation on morphological features of Tylenchorhynchus spp.
To study the nematode population, forty three soil samples (200cc) werecollected from the ground nut fields of Hooghly district of West Bengal, Indiaduring May, 2012. Survey was conducted in four major groundnut growingblocks (Haripal, Pursurah, Khanakul-I and Jangipara) of the district. GPS(Geographical Positioning System) information for the each area under surveywas also recorded during sampling. Soil samples from the mentioned crop fieldwere collected by making a ‘V’ shaped core to a depth of 20cm with the help ofkhurpi from the rhizosphere of the crop. Each composite sample was obtainedfrom ten such random cores at each site. The samples were kept in a polythenebag, labelled and brought to the laboratory for storing in refrigerator at 5 C forfurther extraction and processing of the nematode specimens.

50
The nematodes were extracted from 200cc of composite soil sample byCobb’s decanting and sieving method (Cobb, 1918) followed by modifiedBaermann technique (Christie and Perry, 1951).
The nematode specimens were killed by hot-water-bath method. Theconcentrated nematode suspension was plunged into a beaker containing hot
water at about 60-65oC and shaken the suspension continuously inside thebeaker for about 2-3 minutes. Then the nematodes were killed and suspension ofnematode was kept out of hot water to bring the suspension at room temperatureas well as to allow the nematode for settling at the bottom of suspension. Toconcentrate the nematode suspension, excess amount of water was removedgently from the top of suspension by means of dropper without disturbing thewhole. Killed nematodes were then fixed in 4:1 formalin glacial acetic acid andkept separately in the labelled vial for further study.
Vial containing fixed nematode suspension was taken in a measuringcylinder to measure the total volume of nematode suspension obtained from200cc of soil. Nematodes remained in the fixed suspension were counted understereoscopic binocular microscope (Carl Zeiss-Stemi 2000C) using multi-chambered counting disc. The nematode suspension from each location wasobserved and mean of three aliquot was taken for calculating the populationdensity per 200cc of soil.
For morphological and morphometric studies the required number of fixednematode specimens from each population were processed by glycerol-ethanolmethod (Seinhorst, 1959).
The processed specimens were permanently mounted in pure anhydrousglycerol. A small drop of glycerol was placed on the centre of clean glass slidemeasuring 3 ×1 . About 7-8 specimens of processed nematodes were picked upand placed in the centre of glycerol drop so that their head pointing at same

52
Tylenchorhynchus
µµ
1. n = number of specimen2. L =Total body length i.e., head to tail end 3. Body width 4. Esophagus length (up to cardia)5. Stylet length6. Length of anterior (conical) parts of spear (conus)7. Length of shaft of spear8. Stylet knob width 9. Inclination of stylet knob 10. Body width at base of spear/stylet knob11. Depth of annule at esophageal end, mid body and at tail region12. Depth of annule at mid body region13. Depth of annule at tail region14. Centre of Median bulb from the base of stylet knob15. Median bulb width 16. Position of hemizonid (anterior or posterior to excretory pore)17. Excretory pore from anterior end 18. Number of lateral lines19. Width of lateral field20. Distance of anus from anterior end21. Distance of phasmid from anus (+ = anterior to anus, - = posterior to anus)22. Distance of phasmid from tail terminus
P %= (distance of phasmid from anus / tail length) × 100V% = (distance of vulva from anterior/ total body length) × 100T %= (distance from cloaca to anterior part of testis/total body length) × 100The measurements of all the specimens were taken under compound microscope(Olympus BX-51) with the help of ocular micrometer as well as with the imageanalyzing device ProgRes CT5 of Jenoptic Version 2.80. All the drawings weremade with the help of drawing tube of the compound binocular microscope(Olympus BX-51).

53
23. Number of annules from tail terminus to phasmid24. Spicule length 25. Gubernaculum length26. Lip region (continuous/offset/sunken) 27. Lip height 28. Lip annules 29. Tail annules30. Tail tip (smooth/annulated)31. Tail shape32. Shape of tail terminus
a , b, c, c´
m, o, P, V, T
For community study, absolute and relative frequency, absolute andrelative density, and prominence value (absolute and relative) of majorrhizospheric nematodes of groundnut were determined according to theformulae given by Norton (1978). For comparing the morphometric data of thespecies under genus Tylenchorhynchus Cobb, 1913 arithmetic mean, standarddeviation (SD), and co-efficient of variation (CV) were determined. ConsideringCV value the taxonomic characters were rated as least variable (CV<6%), moderately variable (CV 6-8%) and highly variable (CV>8%) for the female, male and juveniles of Tylenchorhynchus.

54
Forty three soil samples from groundnut fields of Hooghly district werecollected to study the occurrence of associated nematode species. Survey wasconducted in four major groundnut growing blocks (Haripal, Pursurah, Khanakul-I and Jangipara) of the district. Six genera of plant parasiticnematodes (Criconemoides, Rotylenchulus, Tylenchorhynchus, Meloidogyne, Hirschmanniella and Pratylenchus) were encountered from the soil samples. Ofwhich, Criconemoides, Rotylenchulus, Tylenchorhynchus and Meloidogyne weremore frequent over other genera of plant parasitic nematodes (Table 2). Criconemoides were recorded in the range of 31-2016/200cc of soil, the highestbeing from Chiladangi area of Pursurah block of Hooghly district (Table 1).
Mi Mj Tyl Roty Crico Praty Hirsch
Nalikul (2) 186.3 20.7 89.0 0.0 0.0 0.0 59.0 29.6 1184.0
Belechonga (2) 0.0 0.0 116.5 0.0 474.0 71.5 43.0 0.0 647.5
Samanta Road (3) 0.0 0.0 126.7 2642.0 31.0 0.0 10.3 0.0 578.7
Balarampur (8) 63.6 20.6 408.4 256.1 449.1 33.8 69.9 0.0 1030.4
Chiladangi (5) 173.5 48.9 906.0 6.4 2016.0 11.0 77.2 0.0 672.6
Balipur (10) 50.3 0.0 201.9 1014.7 402.1 3.2 26.0 9.9 1452.6
Purba Radhanagar (7) 39.9 0.0 75.9 0.0 307.6 4.1 19.3 3.9 881.6
Arunda (4) 47.1 11.2 280.3 1929.3 323.3 0.0 0.0 6.1 747.8
Radhanagar (2) 21.0 15.0 477.0 53.0 470.0 0.0 35.0 0.0 1273.0
Note: Mi- Meloidogyne incognita, Mj- M. javanica, Tyl – Tylenchorhynchus mashhoodi, Roty- Rotylenchulusreniformis, Crico- Criconemoides sp., Praty- Pratylenchus sp., Hirsch – Hirschmanniella spp., Mon-Mononchids, Sapro – Saprozoic nematode including free living dorylaimids and rhabditids

57
Tylenchorhynchus
All the populations of Tylenchorhynchus sp. studied were identified asTylenchorhynchus mashhoodi Siddiqi & Basir, 1959. The comparativemorpphometric diagnostic data of females of T. mashhoodi Siddiqi & Basir, 1959 is presented in the table- 3. The lip region was continuous, stylet knob wasround and inclined posteriorly, tail cylindrical to sub-cylindrical with smoothand sub-hemispherical terminus (Fig. 2, 5, 9, 10). Body length (0.44-0.84 mm), c' (1.7-5.0), lip annules (3), stylet length (15-23 µm) and number of tail annules(15-28) are found similar to the findings of Handoo (2000). Presentmorphometric observations confirmed the studied population ofTylenchorhynchus as Tylenchorhynchus mashhoodi Siddiqi & Basir, 1959. T.mashhoodi has been reported as one of the common Indian species under genusTylenchorhynchus (Bajaj et al., 2010). Baqri and Jairajpuri (1970) have reportedT. mashhoodi and T. annulatus from rice growing areas of Burdwan district ofWest Bengal. Mukerjee and Dasgupta (1983) recorded T. coffeae Siddiqi &Basir, 1959 and T. crassicaudatus Williams, 1960 from banana growing areas ofHooghly district of West Bengal. In the present investigation, Tylenchorhynchusmashhoodi was detected as the only dominant species under the genusTylenchorhynchus. It was recorded in abundance from the rhizospheric soil ofgroundnut crop in the surveyed areas of Hooghly district of West Bengal whererice-potato-groundnut is a major adopted cropping sequence among the farmingcommunity.
Tylenchorhynchusmashhoodi
Length (mm) 0.66 (0.44-0.84) 0.49-0.76Lip region Continuous ContinuousLip annules 3 3-4Stylet length (µm) 19.1 (15-23) 16-20Stylet knob inclination Posterior PosteriorTail annules 23 (15-28) 14-29Tail shape Cylindrical to sub-cylindrical CylindricalTail terminus Sub-hemispherical Bluntly pointedTail tip annulation Smooth Smoothc' 3.9 (1.7-5.0) 2.5-4.0

58
Tylenchorhynchus mashhoodi
Data on morphometric variability of Tylenchorhynchus mashhoodi hasbeen presented in the table-4. The body length of T. mashhoodi varied from 440-840µm in females and 570-680µm in males. Siddiqi and Basir (1959) observedthat, the body length of holotype female ranged from 490-760µm. Later, Handoo (2000) studied the paratype females of T. mashhoodi population fromUSDA Nematode Collection at Beltsville, Maryland and mentioned similarrange of body length.
Baqri and Jairajpuri (1970) documented 540-720µm body length for T.mashhoodi. The present population of T. mashhoodi revealed highly variable(CV>8) body length in females than in males (Table 4). Number of lip annulesand lateral lines were found very consistent (CV<6) among the population of T.mashhoodi. Therefore, these could be considered as stable characters foridentification and differentiation of T. mashhoodi from other species. The styletlength (15-23µm) showed least variability among juveniles, moderate variabilityamong males and high variability among females. Handoo (2000) presentedstylet length of T. mashhoodi female ranging from 16-20µm, while 16-19µmhad been cited by earlier workers (Siddiqi and Basir,1959; Baqri andJairajpuri,1970). The mean stylet length of the females of T. mashhoodi in thepresent investigation was recorded as 19.1 µm. This observation was in paritywith the findings made by earlier workers.
Ratios like, a, b, c, c´ were found highly variable in females; while malesshowed least to moderate variability with regard to a and b ratios (Table 4). V%in females, distance of median valve from stylet base and excretory pore fromanterior end in males are found least variable (CV<6). Observation of thepresent author with regard to V% and T% were in agreement with Siddiqi andBasir (1959). Percentage like, m, o, p and T were found highly variable in thepopulation of T. mashhoodi recovered from the rhizosphere of groundnut. Inmale length of spicule was found very consistent while gubernaculum lengthwas highly variable (Table 4). Both these variables were found at par with theobservations of Siddiqi and Basir (1959); Siddiqi (1961) and Baqri andJairajpuri (1970). Distance of phasmid from anus and number of tail annules inthe present population of T. mashhoodi were at par with the observations ofSiddiqi and Basir (1959) and Handoo (2000).

59
Tylenchorhynchus mashhoodi
µµ
L 657.0 ± 99.4(440-840)
15.1 636.3 ± 50.2 (570-680)
7.9 552.5 ± 61.3(420-640)
11.1
a 23.5 ± 3.6(16-33.8)
15.4 22.5 ± 1.0(21.1-23.1)
4.2 22.4 ± 2.6(19.1-26.9)
11.7
b 4.5 ± 0.5(3.4-5.3)
11.0 4.8 ± 0.3(4.4-5.0)
6.2 4.0 ± 0.4(3.4-4.6)
9.5
c 13.9 ± 2.5(10.7-26.9)
18.2 15.3 ± 1.5(13.6-16.8)
9.5 12.4 ± 1.2(10.3-14.1)
9.9
c´ 3.9 ± 0.7(1.7-5.0)
17.0 3.2 ± 0.3(2.9-3.6)
9.0 3.9 ± 0.5(2.7-4.5)
13.0
m % 52.2 ± 7.1(31.6-63.2)
13.6 56.6 ± 7.3(50.0-66.7)
12.9 56.7 ± 6.9(44.4-66.7)
12.1
o % 16.3 ± 3.8(10.0-26.3)
23.4 12.8 ± 2.7(10.5-16.7)
21.2 14.6 ± 3.2(10.5-20.0)
22.1
Body width 28.2 ± 3.0(20-30)
10.6 28.3 ± 1.5(27.0-30.0)
5.3 24.8 ± 3.0(20.0-29.0)
12.2
Lip region Continuous - Continuous - Continuous -Lip height 4.1 ± 0.7
(3.0-6.0)16.2 4 ± 0 0.0 4.0 ± 0.5
(2.5-5.0)13.7
Lip annules 3 ± 0.0 0.0 3 ± 0 0.0 3.0 ± 0 0.0Stylet length 19.1 ± 1.6
(15-23)8.4 17.6 ± 1.3
(16.0-19.0)7.1 18.5 ± 1.1
(17.0-20.0)5.8
Esophagus length 117.3 ± 8.6(100-132.5)
7.3 107.5 ± 7.1 (97.5-112.5)
6.6 112.5 ± 11.9(85.0-125.0)
10.6
Length of conus ofspear
9.9 ± 1.7(6-13)
17.2 10 ± 1.6(8.0-12.0)
16.3 10.5 ± 1.3(8.0-12.0)
12.8
Length of shaft ofspear
7.4 ± 1.0(5.0-9.0)
14.0 6.1 ± 1.2(4.5-7.0)
19.3 6.3 ± 1.3(4.0-8.0)
20.5
Stylet knob width 4.7 ± 0.8(3.0-6.0)
16.2 4 ± 0.8(3.0-5.0)
20.4 4.6 ± 0.5(3.8-5.0)
11.2
Inclination of styletknob
Posterior - Posterior - Posterior -
Body width at base ofstylet knob
11.9 ± 1.2(9.0-15.0)
10.1 10.6 ± 0.8(10.0-11.5)
7.1 11.1 ± 1.1(10.0-13.5)
9.6
Depth of annule atesophageal end
1.0 ± 0.0(1.0-1.25)
3.6 1 ± 0 0.0 0.94 ± 0.2(0.5-1.3)
23.1
Depth of annule atmid body region
1.0 ± 0.1(1-2)
14.0 1 ± 0 0.0 1.0 ± 0.1(1.0-1.3)
7.1
Depth of annule at tailregion
1.3 ± 0.4(1-2)
31.8 1 ± 0 0.0 1.2 ± 0.3(1.0-2.0)
30.1
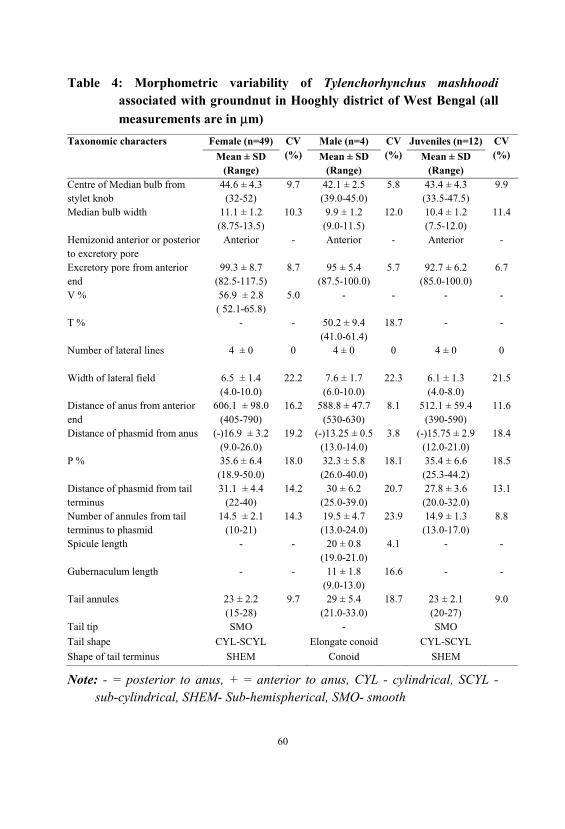
60
Tylenchorhynchus mashhoodi
µµ
Centre of Median bulb fromstylet knob
44.6 ± 4.3(32-52)
9.7 42.1 ± 2.5(39.0-45.0)
5.8 43.4 ± 4.3(33.5-47.5)
9.9
Median bulb width 11.1 ± 1.2(8.75-13.5)
10.3 9.9 ± 1.2(9.0-11.5)
12.0 10.4 ± 1.2(7.5-12.0)
11.4
Hemizonid anterior or posteriorto excretory pore
Anterior - Anterior - Anterior -
Excretory pore from anteriorend
99.3 ± 8.7(82.5-117.5)
8.7 95 ± 5.4(87.5-100.0)
5.7 92.7 ± 6.2(85.0-100.0)
6.7
V % 56.9 ± 2.8( 52.1-65.8)
5.0 - - - -
T % - - 50.2 ± 9.4(41.0-61.4)
18.7 - -
Number of lateral lines 4 ± 0 0 4 ± 0 0 4 ± 0 0
Width of lateral field 6.5 ± 1.4(4.0-10.0)
22.2 7.6 ± 1.7(6.0-10.0)
22.3 6.1 ± 1.3(4.0-8.0)
21.5
Distance of anus from anteriorend
606.1 ± 98.0 (405-790)
16.2 588.8 ± 47.7 (530-630)
8.1 512.1 ± 59.4 (390-590)
11.6
Distance of phasmid from anus (-)16.9 ± 3.2 (9.0-26.0)
19.2 (-)13.25 ± 0.5 (13.0-14.0)
3.8 (-)15.75 ± 2.9 (12.0-21.0)
18.4
P % 35.6 ± 6.4(18.9-50.0)
18.0 32.3 ± 5.8(26.0-40.0)
18.1 35.4 ± 6.6(25.3-44.2)
18.5
Distance of phasmid from tailterminus
31.1 ± 4.4(22-40)
14.2 30 ± 6.2(25.0-39.0)
20.7 27.8 ± 3.6(20.0-32.0)
13.1
Number of annules from tailterminus to phasmid
14.5 ± 2.1(10-21)
14.3 19.5 ± 4.7(13.0-24.0)
23.9 14.9 ± 1.3(13.0-17.0)
8.8
Spicule length - - 20 ± 0.8(19.0-21.0)
4.1 - -
Gubernaculum length - - 11 ± 1.8(9.0-13.0)
16.6 - -
Tail annules 23 ± 2.2(15-28)
9.7 29 ± 5.4(21.0-33.0)
18.7 23 ± 2.1(20-27)
9.0
Tail tip SMO - SMO
Tail shape CYL-SCYL Elongate conoid CYL-SCYL
Shape of tail terminus SHEM Conoid SHEM
Note: - = posterior to anus, + = anterior to anus, CYL - cylindrical, SCYL -sub-cylindrical, SHEM- Sub-hemispherical, SMO- smooth

61
Tylenchorhynchus mashhoodi
Morphometric data of T. mashhoodi associated with groundnut inHooghly district of West Bengal has been presented in the table -4.
: Length 440-840 µm (mean 657.0 µm, standard deviation [SD]99.4 µm); body width = 20-30 µm (28.2, 3.0); a = 16-33.8 (23.5, 3.6); b = 3.4-5.3 (4.5, 0.5); c = 10.7-26.9 (13.9, 2.5); c´ = 1.7-5.0 (3.9, 0.7); m % = 31.6-63.2(52.2, 7.1); o % = 10.0-26.3 (16.3, 3.8); P % = 18.9-50.0 (35.6, 6.4); V % = 52.1-65.8 (56.9, 2.8); lip height = 3.0-6.0(4.1,0.7); lip annules =3; stylet length = 15-23 µm (19.1, 1.6); esophagus length= 100-132.5 µm (117.3, 8.6); length of conus of spear = 6-13 µm (9.9, 1.7);length of shaft of spear = 5.0-9.0 µm (7.4, 1.0); stylet knob width = 3.0-6.0 µm(4.7, 0.8); body width at the base of stylet knob = 9.0-15.0 µm (11.9, 1.2); depthof annule at esophageal end = 1.0-1.25 µm (1.0, 0.0); depth of annule at midbody region = 1-2 µm (1.0, 0.1); depth of annule at tail region = 1-2 µm(1.3,0.4); distance from stylet knob to centre of median bulb = 32-52 µm(44.6, 4.3); median bulb width = 8.75-13.5 µm (11.1, 1.2); distance of excretory porefrom anterior end = 82.5-117.5 µm (99.3, 8.7); number of lateral lines = 4; widthof lateral field = 4.0-10.0 µm(6.5, 1.4); distance of anus from anterior end =405-790 µm (606.1, 98.0); distance of phasmid from anus = 9.0-26.0 µm (16.9, 3.2); distance of phasmid from tail terminus = 22-40 µm (31.1, 4.4); number ofannules from tail terminus to phasmid = 10-21 (14.5, 2.1); number of tailannules = 15-28 (23, 2.2).
Body slightly arcuate or curved ventrally in posterior half of its lengthwhen heat relaxed, tapering slightly towards both extremities. Cuticle finelystriated. Annules were 1-2µm wide on mid body and tail. Lateral field was 4-10
(6.5) µm wide with four incisures, which were not areolated. Longitudinalstriations were absent. Lip region was round, continuous with the body contourand bearing 3 annules; cephalic framework was lightly sclerotized. Stylet waswell developed, 15-23 (19.1) µm long, conus was longer than shaft, basal knobwas rounded and slopping backward (posteriorly directed or inclined). Esophagus was typical of the genus. Median esophageal bulb was spheroid with8.75-13.5 µm wide and prominent refractive valvular apparatus in the centre.

62
Median valve was located at 32-52 (44.6) µm away from stylet knob. Orifice ofdorsal esophageal gland was 3.1µm behind stylet base. Nerve ring wasencircling esophagus slightly anterior or middle of isthmus. Hemizonid was oneto two body annule long, located at one annule anterior to excretory pore at thebase of isthmus. Basal esophageal bulb was pyriform, cardia small andhemispherical.Vulva was like a transverse slit, lacking epiptygma or lateralmembranes; vagina at right angle to the body axis. Ovaries were amphidelphic, outstretched. Spermatheca was round, filled with sperm. Tail was 3.9 anal bodywidth (abw) long, marked with 15-28 annules ventrally, cylindrical to sub-cylindrical in shape, slightly narrowing down after anus. Tail terminus was sub-hemispherical in shape and smooth (unstriated). Phasmidwas located in anterior half (18.9-50%) of tail.

63

64

65

66

67

68

69

70
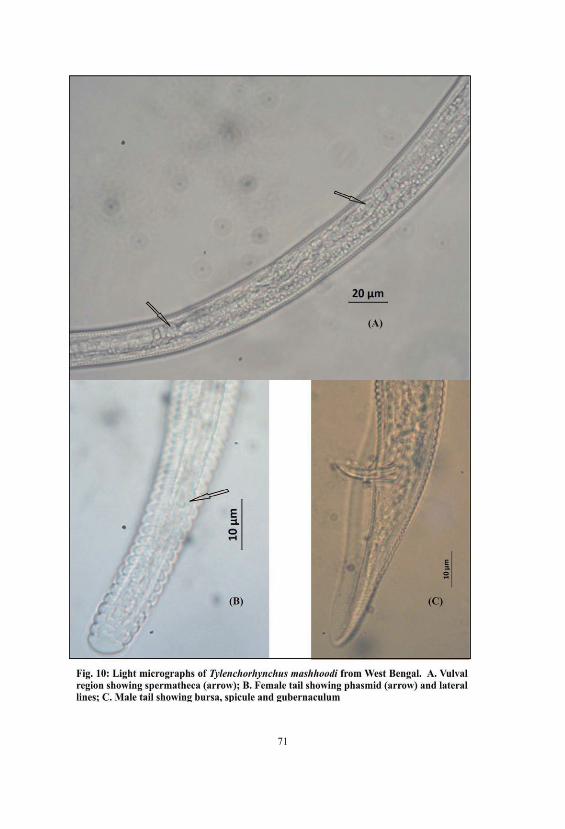
71

72
: Length 570-680 µm (mean 636.3 µm, standard deviation [SD] 50.2µm); body width = 27.0-30.0 µm (28.3, 1.5); a = 21.1-23.1 µm (22.5, 1.0); b =4.4-5.0 (4.8,0.3); c = 13.6-16.8 (15.3,1.5); c´= 2.9-3.6 (3.2, 0.3); m % = 50.0-66.7(56.6, 7.3); o % = 10.5-16.7 (12.8, 2.7); P % = 26.0-40.0 (32.3, 5.8); T % =41.0-61.4 (50.2, 9.4); lip height = 4µm; lip annules = 3; stylet length = 16.0-19.0µm (7.6, 1.3); esophagus length = 97.5-112.5µm (107.5, 7.1); length of conus ofspear = 8.0-12.0 µm (10, 1.6); length of shaft of spear = 4.5-7.0µm (6.1, 1.2);stylet knob width = 3.0-5.0 µm (4,0.8); body width at the base of stylet knob =10.0-11.5 µm (10.6-0.8); depth of annule at esophageal end, mid body regionand tail region = 1.0µm; distance from stylet knob to centre of median bulb =39-45 µm(42.1, 2.5); median bulb width = 9.0-11.5µm(9.9, 1.2); excretory porefrom anterior end = 87.5-100.0µm (95, 5.4); number of lateral lines = 4.0-5.0 (4, 0.5); width of lateral field = 6.0-10.0 µm(7.6, 1.7); distance of anus fromanterior end = 530-630 µm (588.8, 47.7); distance of phasmid from anus =13.0-14.0 µm (13.25, 0.5); distance of phasmid from tail terminus = 25.0-39.0µm (30.0, 6.2); spicule length = 19.0-21.0µm (20.0, 0.8); gubernaculum length =9.0-13.0 µm (11, 1.8); number of annules from tail terminus to phasmid = 13.0-24.0 (19.5, 4.7); number of tail annules = 21-33 (29, 5.4).
Males were similar to female in general morphology, but slightly smaller thanfemale. Testis single, outstretched. Spicule tylenchid, arcuate, pointed at distalend, 19-21 (20)µm long medially. Gubernaculum 9-13 (11)µm long. Bursatylenchoid, well developed with transverse cuticular striae, completelyenveloping tail (peloderan). Tail was elongate conoid with acute or sub-acuteterminus, 2.9-3.6 anal body width long, bearing 21-33 annules (transverse striae)ventrally. Phasmid located in anterior half (26-40%) of tail.
: Length 420-640 µm (mean 552.5, µm, standard deviation [SD]61.3 µm); body width 20.0-29.0 µm (24.8, 3.0); a = 19.1-26.9 µm (22.4, 2.6); b= 3.4-4.6 (4.0, 0.4); c = 10.3-14.1 (12.4, 1.2); c´ = 2.7-4.5 (3.9, 0.5); m % =44.4-66.7 (56.7, 6.9); o % = 10.5-20.2 (14.6, 3.2); P % = 25.3-44.2 (35.4, 6.6);lip height = 2.5-5.0 µm (4.0, 0.5); lip annules = 3; stylet length = 17.0-20.0 µm(18.5, 1.1); esophagus length = 88.0-125.0 µm (112.5, 11.9); length of conus ofspear = 8.0-12.0 µm (10.5-1.3); length of shaft of spear = 4.0-8.0 µm (6.3, 1.3);stylet knob width = 3.8-5.0 µm (4.6, 0.5); body width at the base of stylet knob= 10.0-13.5 µm (11.1, 1.1); depth of annule at esophageal end = 0.5-1.3 µm

73
(0.94,0.2); depth of annule at mid body region = 1.0-1.2 µm (1.0, 0.1); depth ofannule at tail region = 1.0-2.0 µm (1.2, 0.3); distance from stylet knob to thecentre of median bulb = 33.5-47.5 µm (43.4, 4.3); median bulb width = 7.5-12.0µm (10.4, 1.2); excretory pore from anterior end = 85.0-100.0 µm (92.7, 6.2);number of lateral lines = 4; width of lateral field = 4.0-8.0 µm (6.1-1.3); distanceof anus from anterior end = 390-590 µm (512.1, 59.4); distance of phasmidfrom anus = 12-21 µm (15.75, 2.9); distance of phasmid from tail terminus =20.0-30.0 µm (27.8, 3.6); number of annules from tail terminus to phasmid = 13-17 (14.9, 1.3); number of tail annules = 20-27 (23, 2.1).
Twelve juveniles of unknown development stage were examined, and allshowed the characteristic continuous head with posteriorly inclined stylet knobas described in adults.
Tylenchorhynchus mashhoodi
Data pertaining to the comparative morphometric variability ofTylenchorhynchus mashhoodi has been presented in the table -5. Most of theTylenchorhynchus species encountered in the experiment was found to beTylenchorhynchus mashhoodi when taxonomic characters observed in thepresent experiment and compared with the findings of earlier observation byHandoo (2000), Siddiqi & Basir (1959), Siddiqi (1961) and Baqri & Jairajpuri(1970). Slight variations on morphometric characters in the groundnutpopulation of Tylenchorhynchus mashhoodi have been observed and arecompared in the table- 5.

74
µµ Tylenchorhynchusmashhoodi
L 657.0 (440-840) 490-760 540-720Lip region Continuous Continuous ContinuousLip annule 3 3-4 -Stylet length 19.1 (15-23) 16-19 16-19Stylet knobshape/inclination
Posteriorly directed Posteriorlydirected
Round, flattened
Spermatheca Functional - FunctionalLateral lines 4 4 -Spicule length 20 (19.0-21.0) 18-22 18-22Gubernaculum length 11 (9.0-13.0) 10-15 10-15Post anal extension Absent Absent -Tail shape CYL-SCYL CYL CYL-SCYLTail terminus SHEM & SMO SHEM & SMO RND-BLR& SMOTail annule 23 (15-28) 15-30 14-29a 23.5 (16-33.8) 25.8-37.0 28-37b 4.5 (3.4-5.3) 4.4-5.8 4.4-5.8c 13.9 (10.7-26.9) 13.0-19.4 13-18c´ 3.9 (1.7-5.0) 2.5-4.0 2.5-4.0V % 56.9 ( 52.1-65.8) 50-59 52-58Note: CYL - cylindrical, SCYL - sub-cylindrical, SHEM- Sub-hemispherical, SMO- smooth, RND- round, BLR-bluntly rounded

75
A roving survey was conducted in four major groundnut growing blocks(Haripal, Pursurah, Khanakul-I and Jangipara) of the Hooghly district of WestBengal, India during May, 2012. To study the occurrence and distribution ofrhizospheric nematode population forty three soil samples (200cc) werecollected. Community study of major soil inhabiting plant parasitic nematodesrevealed presence of six nematode genera in the rhizosphere of peanut. Thesewere Criconemoides Taylor, 1936, Rotylenchulus Linford and Oliviera, 1940Tylenchorhynchus Cobb, 1913, Meloidogyne Goeldi, 1892, HirschmanniellaLuc and Goodey, 1964 and Pratylenchus Filipjev, 1936. Among these, Criconemoides, Rotylenchulus, Tylenchorhynchus and Meloidogyne were foundmore frequent over other genera of plant parasitic nematodes. Criconemoides sp. has been identified as most predominant plant parasitic nematode in groundnutfollowed by Rotylenchulus and Tylenchorhynchus. Criconemoides (2016/200ccsoil) and Tylenchorhynchus mashhoodi (906/200cc soil) were recorded highestfrom Chiladangi area of Pursurah block of Hooghly district. Meloidogyneincognita was recorded maximum (186.5/200cc soil) from Haripal block. Rotylenchulus reniformis was also recorded in huge number (2642/200cc of soil)from Samanta Road areas of Pursurah block. Those areas could be treated ashot spot for the respective nematodes. Beside phytonematodes, mononchids(especially Mylonchulus sp.) the predatory nematodes were also found in afrequency of 11.6%. However, saprozoic nematodes comprising of free livingdorylaimids and rhabditids were also observed as very prominent memberamong soil nematode communities of groundnut crop. Rotylenchulus reniformis Linford and Oliviera, 1940 and Tylenchorhynchusmashhoodi Siddiqi and Basir, 1959 were identified as prevalent nematodespecies under genera Rotylenchulus and Tylenchorhynchus, respectively. Amongroot-knot nematodes, two species viz., Meloidogyne incognita (Kofoid andWhite, 1919) Chitwood, 1949 and Meloidogyne javanica (Treub, 1885)Chitwood, 1949 were identified. The present findings on the morphometric characters of the populations ofTylenchorhynchus from groundnut growing areas of Hooghly district confirmedthe species as Tylenchorhynchus mashhoodi Siddiqi and Basir, 1959. The body length of T. mashhoodi varied from 440-840µm in females and 570-680µm in males. Present population of T. mashhoodi revealed highly variable

76
body length in females than in males. Number of lip annules and lateral lineswere found very consistent among the population of T. mashhoodi. The styletlength (15-23µm) showed least variability among juveniles, moderate variabilityamong males and high variability among females. The mean stylet length of thefemales of T. mashhoodi in the present investigation was recorded as 19.1 µm. Ratios like, a, b, c, c´ were found highly variable in females; while malesshowed least to moderate variability with regard to a and b ratios. V% infemales, distance of median valve from stylet base and excretory pore fromanterior end in males are found least variable. Percentage like, m, o, p and Twere found highly variable in the population of T. mashhoodi recovered fromthe rhizosphere of groundnut. In male, length of spicule was found veryconsistent while gubernaculum length was highly variable. Morphometric data of T. mashhoodi associated with groundnut in Hooghlydistrict of West Bengal can be summarized as followsL 440-840 µm; body width = 20-30 µm a = 16-33.8; b = 3.4-5.3; c = 10.7-26.9;c´ = 1.7-5.0; m% = 31.6-63.2; o% = 10.0-26.3; P% = 18.9-50.0; V% = 52.1-65.8; T % = 41.0-61.4; lip annules =3; stylet length = 15-23 µm; esophaguslength = 100-132.5 µm; stylet knob width = 3.0-6.0 µm; depth of annule at midbody region = 1-2 µm; number of lateral lines = 4; distance of phasmid fromanus = 9.0-26.0 µm (16.9, 3.2); number of tail annules = 15-28, length of spicule19-21µm medially; gubernaculum 9-13 µm long. Body slightly arcuate or curved ventrally in posterior half of its length whenheat relaxed, tapering slightly towards both extremities. Cuticle finely striated. Annules were 1-2µm wide on mid body and tail. Lateral field was 6.5µm widewith four incisures. Longitudinal striations were absent. Lip region was round, continuous with the body contour and bearing 3 annules; cephalic frameworkwas lightly sclerotized. Stylet was well developed, 15-23 (19.1) µm long, conuswas longer than shaft, basal knob was rounded and slopping backward(posteriorly directed or inclined). Esophagus was typical of the genus. Medianesophageal bulb was spheroid with 8.75-13.5 µm wide and prominent refractivevalvular apparatus in the centre. Median valve was located at 32-52 (44.6) µm
away from stylet knob. Orifice of dorsal esophageal gland was 3.1µm behindstylet base. Nerve ring was encircling esophagus slightly anterior or middle ofisthmus. Hemizonid was one to two body annule long, located at one annuleanterior to excretory pore at the base of isthmus. Basal esophageal bulb waspyriform, cardia small and hemispherical. Vulva was like a transverse slit,

77
lacking epiptygma or lateral membranes; vagina at right angle to the body axis. Ovaries were amphidelphic, outstretched. Spermatheca was round, filled withsperm. Tail was 3.9 anal body width (abw) long, marked with 15-28 annulesventrally, cylindrical to sub-cylindrical in shape, slightly narrowing down afteranus. Tail terminus was sub-hemispherical in shape and smooth (unstriated). Phasmid was located in anterior half of tail. Males were similar to female in general morphology, but slightly smaller thanfemale. Testis single, outstretched. Spicule arcuate, pointed at distal end. Gubernaculum rod shaped. Bursa well developed with transverse cuticularstriae, completely enveloping tail. Tail was elongate conoid. Twelve juveniles of unknown development stage were examined, and allshowed the characteristic continuous head with posteriorly inclined stylet knobas described in adults.

78
In the present experiment community of the rhizospheric nematodesassociated with groundnut crop, their identification and detailed morphometricvariability of Tylenchorhynchus mashhoodi had been studied. All the groundnutgrowing areas of the state need to be surveyed to get a detail idea about theextent of nematode infestation on the crop. Other than Tylenchorhynchusmashhoodi, many plant parasitic nematode species like Rotylenchulusreniformis, Meloidogyne incognita, Meloidogyne javanica, Criconemoides sp. and Pratylenchus sp. were found associated with the crop. Species levelidentification and morphometric variability (if any) of those parasitic nematodespecies need to be carried out in future. Importance value, index of similarity, richness of the species and diversity index among nematode species of differentlocality need to be worked out in future to have an idea about the interactionamong nematode community of groundnut crop. Observation on thepathogenicity of the parasitic nematodes species associated with groundnut cropand their eco-friendly management strategy to be developed in future.

79
Alexander, P. M. (1963). Stylet-bearing nematodes associated with variousplants in South Carolina. Plant Disease Reporter, : 978–982.
Allen, M. W. (1955). A review of the nematode genus Tylenchorhynchus. University of California Publications in Zoology, :129–166.
Allen, M. W. and Sher, S. A. (1967). Taxonomic problems concerning thephytoparasitic nematodes. Ann. Rev. Phytopath., :247-264.
Anderson, R. V. and Potter, J. W. (1991). Stunt nematodes: Tylenchorhynchus, Merlinius, and related genera. In. Manual of Agricultural Nematology(Ed. Nickle, W. R.), New York: Marcel Dekker, pp. 529–586.
Andrássy, I. (1976). Evolution as a basis for the Systematization of Nematodes. London, UK: Pitman Publishing, pp. 288.
Anonymous (1973). Plant Nematology. In. Secretary for Agriculture Report forthe Period 10 october 1971 to 30 September 1972.Salisbury, Rhodesia.
Anonymous (2010). Final Estimates of Groundnut in West Bengal during 2009-10. Evaluation Wing, Directorate of Agriculture, West Bengal.
Bajaj, H. K., Walia, R. K. and Walia, K. K. (2010). Identification of ImportantPlant Parasitic Nematodes. Department of Nematology, CCS HaryanaAgricultural University, Hisar, pp.30.
Baqri, Q. H. and Jairajpuri, M. S. (1970). On the intra-specific variations ofTylenchorhynchus mashoodi Siddiqi and Basir, 1959 and an emended keyto species of Tylenchorhynchus Cobb, 1913 (Nematoda). Revista BrasiiaBiologica, :61–68.
Barker, K. R., Schmitt, D. P. and Campos, V. P. (1982). Response of peanut, corn, tobacco, and soybean to Criconemella ornata. Journal ofNematology. : 576–581.
Basu, M. S. and Ghosh, P. K. (1995). The status of technologies used to achievehigh groundnut yields in India. In Achieving high Groundnut Yields. Patancheru, India: ICRISAT.
Beals, E. (1960). Forest bird communities in the Apostle Island of WisconsinWilson Bulletin, :156-181.
Bertioli, D. J., Seijo, G., Freitas, F. O., Valls, J. F. M., Leal-Bertioli, S. C. M. and Moretzsohn, M. C. (2011). An overview of peanut and its wildrelatives. Plant Genetic Resources: Characterization and Utilization,
(1): 134-149.

80
Boswell, T. E. (1968). Pathogenicity of Pratylenchus brachyurus to Spanishpeanut. Ph. D. Dissertation, Texas A&M University, College Station, Texas.
Boyle, L.W. (1950). Several species of parasitic nematodes on peanuts inGeorgia. Plant Disease Reporter, : 61–62.
Brzeski, M. W., and Dolinski, C. M. (1998). Compendium of the genusTylenchorhynchus Cobb, 1913 sensulato (Nematoda: Belonolaimidae). Russian Journal of Nematology, :189–199.
Carneiro, R. M. D. G., Carneiro, R. G., Das Neves, D. I. and Almeida, M. R. A. (2003). Nova raça de Meloidogyne javanica detectada em Arachispintoi no Estado do Paraná. Nematologia Brasileira, : 219–221.
Cetintas, R., Lima, R. D., Mendes, M. L., Brito, J. A. and Dickson, D. W. (2003). Meloidogyne javanica on peanut in Florida. Journal ofNematology, : 433–436.
Chabra, H. K. and Mahajan, R. (1976). Pratylenchus coffeae , the root-lesionnematode in groundnut and its control by granular nematicides. NematologiaMediterranea, : 241–242.
Cheng, Y. H. and Tu, C. C. (1980). Identification of Meloidogyne arenaria ofpeanut in Taiwan. Journal of Agricultural Research of China, : 47–53.
Cheng, Y. H., Lo, W. C. and Tu, C. C. (1981). Studies on host rangeof Meloidogyne arenaria and its chemical control. Research Bulletin ofTainan Dais, : 55–64.
Chitwood, B. G. (1950). Genearl structures of nematodes. In. An Introduction toNematology (Eds. Chitwood, B. G. and Chitwood, M. B.), Section 1, Anatomy. Baltimore, USA:Monumental Printing Co., pp. 7-27.
Choi, Y. E. (1981). The root-knot nematodes, Meloidogyne spp., in Korea. Proceedings of the 3rd Research Planning Conference on Root-knotNematodes , Meloidogyne spp., Region VI ,Jakarta, Indonesia.
Christie, J. R. and Perry, V. G. (1951). Removing nematode from soil. Proc. Helminth. Soc. Washington, : 106-108.
Cobb, N. A. (1913). New nematode genera found inhabiting fresh-water andnon-brackish soils. Journal of the Washington Academy of Sciences,
:432–445. Cobb, N. A. (1918). Estimating the nema population of the soil. Agricultural
Technology Circular, Bureau of Plant Industry, United State Departmentof Agriculture, No. :48.

81
Cobb, N. A. 1914. Antarctic marine free-living nematodes of the Shakeltonexpedition. Contributions to a Science of Nematology, pp. 3-33.
Colbran, R.C. (1958). Studies of plant and soil nematodes. 2. Queensland hostrecords of root-knot nematodes (Meloidogyne spp.). Queensland Journalof Agricultural Science, : 101–136.
Colbran, R. C. (1968). Nematodes – important pests of peanuts. Division ofPlant Industry, Department of Primary Industries. Queensland, Australia, Advisory Leaflet No. 955.
De Maeseneer, J. and D’Herde, J. (1963). Methodes utilisees pourl’etude desanguillules libres du sol. Revue Agric. Brux., :441-447.
De Man, J. G. 1876. Onderzoekingen over vrij in de aarde levende nematoden. Tijdschr. Ned. Dierk. Ver., : 78-196.
De Man, J. G. 1880. Die Eingheimischen, frei in der reinen erde und im süssenwasser lebends Nematoden. Tijdschr. Ned. Dierk. Ver., : 1-104.
De Waele, D., Jones, B. L., Bolton, C. and Van den Berg, E. (1988). Ditylenchusdestructor in hulls and seeds of peanut. Journal of Nematology, : 10–15.
Dhurj, I. and Vaishnav, M. U. (1981). Pathogenicity of root-knot nematode ongroundnut. Indian Journal of Nematology, : 217–218.
Dickson, D. W. and Struble, F. B. (1965). A sieving–staining technique forextraction of egg masses of Meloidogyne incognita from soil.Phytopathology, :497.
Dickson, D. W. (1998). Peanut. In. Plant and Nematode Interactions (Eds. Barker, K.R., Pederson, G. A. and Windham, G. L.), American Society ofAgronomy, Crop Science Society of America, and Soil Science Societyof America, Madison, Wisconsin, pp. 523–566.
Eisenback, J. D. and Triantaphyllou, H. H. (1991). Root-knot nematodes:Meloidogyne species and races. In. Manual of Agricultural Nematology(Ed. Nickle, W.R.), Marcel Dekker, New York, pp. 191–274.
Eisenback, J. D., Bernard, E. C., Starr, J. L., Lee, T. A. and Tomaszewski, E. K. (2003). Meloidogyne haplanaria n.sp. (Nematoda: Meloidogynidae), aroot-knot nematode parasitizing peanut in Texas. Journal of Nematology, : 395–403.
Eliava, I. Y. (1964). The position of the genus Tylenchorhynchus within theTylenchoidea (Nematoda:Tylenchida). Soobshch. Akad. Nauk. Gruz. SSR., :669-673 (in Russian).

82
Esser, R. P. (1991). A computer-ready checklist of the genera and species ofphytoparasitic nematodes, including a list of mnemonically coded subjectcategories. Bulletin 13. Florida Department of Agriculture and ConsumerServices, Division of Plant Industry, Contribution No. 387, Gainesville.
FAO (2006). FAO Production Yearbook, Vol.60, Rome, Italy, 37. Fortuner, R. and Luc, M. (1987). A reappraisal of Tylenchina (Nemata). 6. The
family Belonolaimidae Whitehead, 1960. Revue de Ne´matologie, :183–202.
Germani, G. and Dhéry, M. (1973). Observations et expérimentationsconcernant le role des nématodes dansdeux affections de l’arachide enHaute Volta: la ‘chlorose’ et le ‘clump’. Oléagineux, : 235–242.
Golden, A. M., Maqbool, M. A. and Handoo, Z. A. 1987. Description of twonew species of Tylenchorhynchus Cobb, 1913 (Nematoda: Tylenchida), with details of morphology and variation of T. claytoni. Journal ofNematology , :58–68.
Good, J. M., Boyle, L.W. and Hammons, R. D. (1958). Studies of Pratylenchusbrachyurus on peanuts. Phytopathology, : 530–535.
Handoo, Z. A. (2000). A key and diagnostic compendium to the species of thegenus Tylenchorhynchus Cobb, 1913 (Nematoda: Belonolaimidae). Journal of Nematology, :20–34.
Handoo, Z. A., Khan, A. and Islam, S. (2007). A key and diagnosticcompendium to the species of the genus Merlinius Siddiqi, 1970(Nematoda: Tylenchida) with description of Merlinius khuzdarensis n.sp. associated with date palm. Nematology, :251–260.
Hooper, D. J. (1978). The Tylenchorhynchidae. The identification of stuntnematodes (Tylenchorhynchinae, Merlininae and Trophurinae), especiallythose in western Europe. In. Spiral and Stunt Nematodes. Manualprepared for workshop sponsored by the Nematology Group of theAssociation of Applied Biologists, Rothamsted Experiment Station, England, pp. 1–21.
Ibrahim, I. K. A. and El-Saedy, M. A. (1976a) Plant parasitic nematodesassociated with peanuts in Egypt. Egyptian Journal of Phytopathology, :31–35.
Ibrahim, I. K. A. and El-Saedy, M. A. (1976b) Development and pathogenesisof Meloidogyne javanica in peanut roots. Nematologica Mediterranea, :231–234.

83
Inglis, W. E. G. (1983). An outline classification of the phylum Nematoda. Australian Journal of Zoology, :243-255.
Ingram, E. G. and Rodríguez-Kábana, R. (1980). Nematodes parasitic onpeanuts in Alabama and evaluationof methods for detection and study ofpopulation dynamics. Nematropica, : 21–30.
Jackson, E.E. and Sturgeon, R.V. Jr. (1973). Effects of nematicides upon root lesion nematode populations. Journal of the American Peanut Research andEducation Association, : 178–181.
Jain, R. K., Mathur, K. N. and Singh, R. V. (2007). Estimation of losses due toplant parasitic nematodes on different crops in India. Indian Journal ofNematology, (2):219-221.
Jairajpuri, M. S. and Baqri, Q. H. (1991). Nematode Pests of Rice. Oxford &IBH Publishing Co. Pvt. Ltd., pp. 24.
John, K., Vasanthi, R. P. and Venkateswarlu, O. (2006). Variability andheritability studies in groundnut (Arachis hypogaea L.). Legume Research, (3): 225-227.
Jones, B. L. and De Waele, D. (1988). First report of Ditylenchus destructor inpods and seeds of peanut. Plant Disease, : 453.
Kalaiarasan, P., Rajendran, G. and Lakshmanan, P. (2006). Pathogenic potentialof root-knot nematode, Meloidogyne arenaria on groundnut (Arachishypogaea L.). Indian Journal of Nematology, (2): 278-279.
Kepenekci, I. and Ozturk, G. (2002). Plant parasitic nematodes of Tylenchidae(nematoda) associated with groundnut (Arachis hypogaea) fields in theMediterranean region of Turkey. Phytoparasitica, (3): 288-289.
Khan, F. A. and Misari, S. M. (1992). Parasitic nematodes associated withgroundnut crop in four ecological zones of Nigeria. Journal of AfricanZoology, (3): 263-271.
Loof, P. A. A. (1964). Free-living and plant-parasitic nematodes fromVenezuela. Nematologica, : 201–300.
Lordello, A. I. L. and Gerin, M. (1981). Nematóide javanes parasitando raízes enódules de amendoim. Revista de Agricultura Piracicaba, : 278.
Machmer, J. H. (1951). Root-knot of peanut. I. Distribution. Plant DiseaseReporter,
: 364–366. Machmer, J. H. (1953). Criconemoides sp., a ring nematode associated with
peanut ‘yellows’. Plant Disease Reporter, : 156.

84
Mani, A. and Kumar, G. R. (1992). Plant parasitic nematodes associated withgroundnut in Andhra Pradesh. Indian Journal of Nematology, (1): 44-48.
Martin, G. C. (1958). Root-knot nematodes (Meloidogyne spp.) in theFederation of Rhodesia and Nyasaland. Nematologica, : 332–349.
Martin, G. C. (1961). Plant species attacked by root-knot nematodes(Meloidogyne spp.) in the Federation of Rhodesia and Nyasaland. Nematologica, : 130–134.
Mc Donald, A. H., Loots, G. C., Fourie, H. and De Waele, D. (2005). Amicroplot study on Ditylenchus africanus population densities anddamage symptoms on groundnut in relation to commercial yields. Nematology, (5): 647-653.
Mehan, V. K., Reddy, D. D. R. and Mc Donald, D. (1993). Resistance ingroundnut genotypes to “Kalahasti” malady caused by the stuntnematode, Tylenchorhynchus brevilineatus. International Journal of Pest Management, (2): 201-203.
Merny, G., Fortuner, R. and Luc, M. (1974) Plant parasitic nematodes inGambia. L’Agronomie Tropicale, Nogent, :702–707.
Miller, L. I. and Duke, G. B. (1961). Peanut nematode disease control. VirginiaAgricultural Experiment Station Bulletin. 520.
Minton, N. A. and Bell, D. K. (1969). Criconemoides ornatus parasitic onpeanuts. Journal of Nematology, : 349–351.
Minton, N. A. Hammons, R. O. and Parham, S. A. (1970). Infection of shelland peg tissues of six peanut cultivars by Pratylenchus brachyurus. Phytopathology, : 472–474.
Minton, N. A., Mc Gill, J. F. and Golden, A. M. (1969). Meloidogyne javanicaattacks peanuts in Georgia. Plant Disease Reporter, : 668.
Minz, G. (1956). The root-knot nematode, Meloidogyne spp., in Israel. PlantDisease Reporter, : 798–801.
Mitsui, Y., Yoshida, T., Okamoto, K. and Ishii, R. (1976). Relationship between nematode-trapping fungi and Meloidogyne hapla inpeanut field. Japanese Journal of Nematology, : 47–55.
Motsinger, R. E; Crawford, J. L and Thompson, S. S. (1976). Nematodesurvey of peanuts and cotton in south-west Georgia. Peanut Science, :72–74.

85
Mukherjee, B. and Dasgupta, M. K. (1983). Community analysis of nematodesassociated with banana plantations in the Hooghly district, West Bengal, India. Nematologia Mediterranea, : 43-48.
Mukhopadhyay, A. K. and Roy, K. (2006). Community analysis of major plantparasitic nematodes associated with vegetable crops in Eastern andNortheastern India. International Journal of Nematology, (2): 194-199.
Mukhopadhyay, M. C. and Haque, S. (1974). Nematodes associated with fieldcrops, fruit and fodder crops in West Bengal, India. Indian Journal ofNematology, : 104-107.
Naidu, P. H. and Moses, G. J. (2000). Identification and evaluation of sources ofresistance to Kalahasti malady in groundnut. Indian Phytopathology,
(4): 395-398. Norton, D. C. (1978). Some simple methods of community analysis. (In)
Ecology of Plant Parasitic Nematodes. John Willey & Sons, New York. p. 66-68.
Orion, D. and Cohn, E. (1975). Crop rotation for control of certain nematodes. Phytoparasitica, : 69 (Abstract).
Oteifa, B. A. (1962). Species of root-lesion nematodes commonly associatedwith economic crops in the Delta of the U.A.R. Plant DiseaseReporter, : 572–575.
Patel, S. K. and Patel, D. J. (1999). Determination of threshold level ofMeloidogyne javanica pt. 2 attacking groundnut cv. GG 20. Proceedingsof national symposium on rational approaches in nematode managementfor sustainable agriculture, Anand, India, 82-83.
Patel, S. K. Patel, D. J. and Talati (1999). Biochemical alterations induced byMeloidogyne javanica pt. 2 infection in groundnut cv. GG 20. Proceedings of national symposium on rational approaches in nematodemanagement for sustainable agriculture, Anand, India, 76-78.
Powers, T. O. (1983). Systematic analysis of Merlininae Siddiqi, 1971(Nematoda: Dolichodoridae). Ph. D. Thesis. University of California, Riverside.
Powers, T. O., Baldwin, J. G. and Bell, A. H. (1983). Taxonomic limits of thegenus Nagelus (Thorne and Malek, 1968) Siddiqi, 1979 with a descriptionof Nagelus borealis n. sp. from Alaska. Journal of Nematology, : 582–593.

86
Reddy, B. M. R., Sharma, S. B. and Krishnappa, K. (1993). New record on theoccurrence of lesion nematode, Pratylenchus brachyurus on groundnut inKerala. Indian Journal of Nematology, (1): 91.
Reddy, D. D. R., Subrahmanian, P., Sankara Reddy, G. H., Raja Reddy, C. andSiva Ras, D. V. (1984). A nematode disease of peanut caused byTylenchorhynchus brevilineatus. Plant Disease, : 526–529.
Rich, J. R. and Kinloch, R. A. (2009). Peanut Nematode Management. Entomology & Nematology Department, Florida Cooperative ExtensionService, Institute of Food and Agricultural Sciences, University ofFlorida.
Roman, J. and Hirschmann, H. (1969). Morphology and morphometrics of sixspecies of Pratylenchus. Journal of Nematology, : 363–386.
Roy, K. and Mukhopadhyay, A. K. (2011). Community study of soil nematodesin the rhizosphere of solanaceous vegetable crops in West Bengal, India. Journal of Crop and Weed, (2):200-201.
Roy, K., Mukhopadhyay, A. K. and Pramanik, A. (2007). Occurrence, distribution and community analysis of plant parasitic nematodesassociated with leguminous vegetable crops in West Bengal. IndianJournal of Nematology, (1): 58-62.
Rudolphi, C. A. (1808). Entozoorum sive vermium Intestinalium HistoriaNaturalis. Amsterdum: :525.
Saint Smith, J.H., McCarthy, G. J. P., Rawson, J. E., Langford, S. and Colbran, R. C. (1972). Peanut growing. Queensland Agricultural Journal, : 639–644.
Sakhuja, P. K. and Sethi, C. L. (1985a). Growth of groundnut as influenced bydifferent inocula of Meloidogyne javanica. Indian Journal ofNematology, : 135–137
Sakhuja, P. K. and Sethi, C. L. (1985b). Frequency of occurrence of variousplant parasitic nematodes and root-rot fungi on groundnut in Punjab. Indian Journal of Nematology, : 191–194
Sasser, J. N. and Freckman, W. D. (1987). A world perspective onNematology: the role of the society. Vistas on Nematology (Veech, J.A. &Dickson, D.W. eds.), Society of Nematologists, Hyattsville, Maryland, USA, pp. 7-14.

87
Sasser, J. N. (1954). Identification and host–parasite relationships of certainroot-knot nematodes (Meloidogyne spp.). University ofMaryland, Agricultural Experiment Station, Technical Bulletin A-77.
Sasser, J. N. (1977). Worldwide dissemination and importance of the root-knotnematodes, Meloidogyne spp. Journal of Nematology, : 26–29.
Sehgal, M. and Gaur, H. S. (1999). Important Nematode Problems of India. Tech. Bull. NCIPM, New Delhi, India, pp. 16.
Seinhorst, J. W. (1959). A rapid method for the transfer of nematodes fromfixative to anhydrous glycerine. Nematologica, : 67-69.
Seinhorst, J. W. (1959). A rapid method for the transfer of nematodes fromfixative to anhydrous glycerine. Nematologica, :67-69.
Senthamizh, K. and Sivakumar, M. (2004). Community analysis of thenematodes associated with groundnut (Arachis hypogaea L.) in NamakkalDistrict of Tamil Nadu, India. Current Nematology, : 27-28.
Sharma, N. K., Sharma, S. K. and Midha, S. K. (1978) Root-knot disease ofgroundnut in Punjab. Indian Farming, : 19–21.
Sharma, S. B., Siddiqi, M. R., Van, N. V., Hong, N. X. (1994). Plant-parasiticnematodes associated with groundnut in North Vietnam. Afro AsianJournal of Nematology, (2): 185-189.
Sharma, S. B. and Mc Donald, D. (1992). Nematode-caused diseases ofgroundnut. International Journal of Tropical Plant Diseases, (1): 21-30.
Siddiqi, M. R. (1961). Studies on Tylenchorhynchus spp. (Nematoda:Tylenchida) from India. Z. F. Parasitenkunde, :46–64.
Siddiqi, M. R. (1986). Tylenchida Parasites of Plants and Insects. Commonwealth Agricultural Bureaux, Slough, London, United Kingdom, pp. 1-615.
Siddiqi, M. R. and Basir, M. A. (1959). On some plant parasitic nematodesoccurring in South India, with the description of two new species of thegenus Tylenchorhynchus Cobb, 1913. Proceedings of the 46th meetingIndian Science Congress, :35.
Singh, R. V. and Khera, S. (1978). Culturing and the life history studies ofRotylenchulus reniformis Linford and Oliveira on brinjal (Nematoda). Bull. Zool. Surv. India, (2): 115-128.

88
Sreenivasulu, B., Naidu, P. H., Sitaramaiah, K., Rao, S. V. R. and Rao, V. K. (2001). Reaction of groundnut varieties/genotypes to Tylenchorhynchusbrevilineatus. Indian Journal of Nematology, (2): 161-163.
Steenkamp, S., Jordaan, A., McDonald, A. H. and De Waele, D. (2011). Differential responses of resistant and susceptible groundnut genotypes atcellular level to Ditylenchus africanus. Nematology, (2): 177-183.
Steiner, G. (1949). Plant nematodes the grower should know. Proceedings of theSoil Science Society of Florida, 1942, -B: 72–117.
Stigter, K. (2006). Agrometeorological services in various parts of the world, under conditions of a changing climate. Austin Bourke Memorial Lecturepresented in the Royal Irish Academy, Dublin, in the evening of 2 March. Extended Abstract available on the INSAM website(www.agrometeorology.org) under “Accounts of OperationalAgrometeorology”.
Tarjan, A. C. (1964). A compendium of the genus Tylenchorhynchus(Tylenchidae: Nematoda). Proceedings of the Helminthological Society ofWashington, :270–280.
Tarjan, A. C. (1973). A synopsis of the genera and species in theTylenchorhynchinae (Tylenchoidea, Nematoda). Proceedings of theHelminthological Society of Washington, :123–144.
Thies, J., Merrill, S. B. and Corley, E. L. (2002). Red food coloring stain: new, safer procedures for staining nematodes in roots and egg masses on rootsurfaces. Journal of Nematology, : 179–181.
Thorne, G. (1949). On the classification of the Tylenchida, new order(Nematoda, Phasmida). Proceedings of the Helminthological Society ofWashington, :37-73.
Thorne, G. 1949. On the classification of the Tylenchida, new order (Nematode, Phasmidia). Proc. Helmithol. Soc. Washington, :37-73.
Tomaszewski, E. K., Khalil, M. A. M., El-Deeb, A. A., Powers, T. O. and Starr, J. L. (1994). Meloidogyne javanica parasitic on peanut. Journal ofNematology, : 436–441.
Van der Linde, J.W. (1956). The Meloidogyne problem in South Africa. Nematologica. : 177–183.
Weiss, E. A. (2000). Oilseed Crops. London: Blackwell Science. Wheeler, T. A. and Starr, J. L. (1987). Incidence and economic importance of
plant-parasitic nematodes on peanut in Texas. Peanut Science. :94–96.

89
Yang, B. (1984). The identification of 15 root-knot nematode populations. ActaPhytopathologica Sinica, :107–112.
Yin, K. C. and Feng, Z. X. (1981). A preliminary survey on the parasiticnematodes of agricultural crops. Phytopathologica, : 111–126.
Zhang, Y. (1985). Occurrence and control of peanut root-knot disease in non-irrigated sloping fields of Zhangjiang District. Agricultural Science ofGuangdong Province, : 48–49.
List of peanut diseases, accessed on 26.3.13 at http://en.wikipedia.org




















