
Oxygen availability causes morphological changes and a different VEGF/FIk‐1/HIF‐2 expression...
10
This article was downloaded by: [91.72.147.254] On: 22 March 2014, At: 05:18 Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK Italian Journal of Zoology Publication details, including instructions for authors and subscription information: http://www.tandfonline.com/loi/tizo20 Oxygen availability causes morphological changes and a different VEGF/FIk‐1/HIF‐2 expression pattern in sea bass gills Liliana Rinaldi a , Patrizia Basso a , Gianluca Tettamanti a , Annalisa Grimaldi a , Genciana Terova a , Marco Saroglia a & Magda de Eguileor a a Department of Structural and Functional Biology , University of Insubria , Via J. H. Dunant 3, Varese, I‐1100, Italy Published online: 28 Jan 2009. To cite this article: Liliana Rinaldi , Patrizia Basso , Gianluca Tettamanti , Annalisa Grimaldi , Genciana Terova , Marco Saroglia & Magda de Eguileor (2005) Oxygen availability causes morphological changes and a different VEGF/FIk‐1/ HIF‐2 expression pattern in sea bass gills, Italian Journal of Zoology, 72:2, 103-111, DOI: 10.1080/11250000509356660 To link to this article: http://dx.doi.org/10.1080/11250000509356660 PLEASE SCROLL DOWN FOR ARTICLE Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) contained in the publications on our platform. However, Taylor & Francis, our agents, and our licensors make no representations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of the Content. Any opinions and views expressed in this publication are the opinions and views of the authors, and are not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon and should be independently verified with primary sources of information. Taylor and Francis shall not be liable for any losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoever or howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use of the Content. This article may be used for research, teaching, and private study purposes. Any substantial or systematic reproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any form to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http:// www.tandfonline.com/page/terms-and-conditions
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Oxygen availability causes morphological changes and a different VEGF/FIk‐1/HIF‐2 expression...

This article was downloaded by: [91.72.147.254]On: 22 March 2014, At: 05:18Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: MortimerHouse, 37-41 Mortimer Street, London W1T 3JH, UK
Italian Journal of ZoologyPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/tizo20
Oxygen availability causes morphological changesand a different VEGF/FIk‐1/HIF‐2 expressionpattern in sea bass gillsLiliana Rinaldi a , Patrizia Basso a , Gianluca Tettamanti a , Annalisa Grimaldi a ,Genciana Terova a , Marco Saroglia a & Magda de Eguileor aa Department of Structural and Functional Biology , University of Insubria , Via J. H.Dunant 3, Varese, I‐1100, ItalyPublished online: 28 Jan 2009.
To cite this article: Liliana Rinaldi , Patrizia Basso , Gianluca Tettamanti , Annalisa Grimaldi , Genciana Terova , MarcoSaroglia & Magda de Eguileor (2005) Oxygen availability causes morphological changes and a different VEGF/FIk‐1/HIF‐2 expression pattern in sea bass gills, Italian Journal of Zoology, 72:2, 103-111, DOI: 10.1080/11250000509356660
To link to this article: http://dx.doi.org/10.1080/11250000509356660
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”)contained in the publications on our platform. However, Taylor & Francis, our agents, and our licensorsmake no representations or warranties whatsoever as to the accuracy, completeness, or suitability for anypurpose of the Content. Any opinions and views expressed in this publication are the opinions and viewsof the authors, and are not the views of or endorsed by Taylor & Francis. The accuracy of the Contentshould not be relied upon and should be independently verified with primary sources of information.Taylor and Francis shall not be liable for any losses, actions, claims, proceedings, demands, costs,expenses, damages, and other liabilities whatsoever or howsoever caused arising directly or indirectly inconnection with, in relation to or arising out of the use of the Content.
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in anyform to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions

Ital. J. Zool., 72: 103-111 (2005)
Oxygen availability causes morphologi-cal changes and a different VEGF/FIk-1/HIF-2 expression pattern in sea bassgills
LILIANA RINALDIPATRIZIA BASSOGIANLUCA TETTAMANTIANNALISA GRIMALDIGENCIANA TEROVAMARCO SAROGLIAMAGDA DE EGUILEORDepartment of Structural and Functional Biology,University of Insubria,Via J. H. Dunant 3, I-1100 Varese (Italy)
ABSTRACT
The sea bass, Dicentrarchus labrax, is abundantly reared in in-tensive aquaculture and oxygen is one of the most important pa-rameters. The gills of D. labrax exposed for three months to threedifferent oxygen partial pressure conditions were investigated:normoxia, mild hyperoxia and mild hypoxia (respectively, 90--100%, 120-130% and 60-70% of the saturation value). Modifica-tions to Po2 levels were studied with morphological and immuno-histochemical techniques. In normoxia and mild hyperoxia condi-tions, the gills had the typical structure with rows of parallel fila-ments; nevertheless, hyperoxia treatment altered the pillar cellstructure, leading to an enlargement of the vascular lumen. In hy-poxia condition, gills were disorganized, the vascular lumen wasreduced and the lamellae showed apical blebs and outer epithe-lial detachment. In vertebrates, the vascular endothelial growthfactor and its main receptor Flk-1 are two important oxygen de-pendent molecules whose expression can be regulated by the hy-poxia inducible factor 2. In D. labrax, high expression of thesefactors was found in normoxia and mild hypoxia, while in mildhyperoxia they were very weak. Our data characterize the mor-phological changes occurring in D. labrax gills and the differentexpression pattern of the above-mentioned factors after exposureto different oxygen partial pressure conditions.
KEY WORDS: Hyperoxia - Hypoxia - Hypoxia inducible factor -Vascular endothelial growth factor.
ACKNOWLEDGMENTS
We are grateful to Dr Roberto Ferrarese for graphical assistance.This work was supported in part by "V Piano Triennale déliaPesca ed Acquicoltura, legge 41/82, Direzione Generale dellaPesca, Ministero per le politiche Agricole e Forestali".
(Received 6 May 2004 - Accepted 21 October 2004)
INTRODUCTION
In teleost fishes, gills are the principal organ responsi-ble for gas transfer and the lamellae are the primarysites of oxygen uptake. The lamella is a vascular sheetmade up of an endothelial and two epithelial layers,whose shape can be maintained by pillars consisting ofspecialized cells, called pillar cells (Farrell et al., 1980;Laurent, 1984). The internal volume of the lamellae canvary due to a passive distension generated by increasedblood pressure. This distension is responsible for im-portant adjustments of functional respiratory surfaces(Soivio & Hughes, 1978; Booth & Amer, 1979; Soivio &Tuurala, 1981; Farrel et al., 1980).
Sea bass Dicentrarchus labrax (Linneus, 1758) is oneof the most reared species in Mediterranean aquacultureand its important commercial role has prompted re-search into modern techniques aimed at improving fishperformances within artificial rearing environments. Inaquaculture, oxygen partial pressure (Po2) is a very im-portant and critical parameter regulating the complicat-ed relationships between structure and function. Pro-longed and intensive hypoxia and hyperoxia cause irre-versible structural and functional damages (Barthélémyet al., 1981; Leknes, 1985).
A morphometric study of sea bass gills at differentdissolved Po2 was previously performed by Saroglia etal. (2000, 2002). In those works, the Authors evaluatedthe effects of Po2 (normoxia, mild hyperoxia and mildhypoxia conditions, respectively, 90%, 140% and 60% ofthe saturation value) on gill Gas Diffusion Distance(GDD) and Respiratory Surface Area (RSA), and theydescribed an increase of GDD and a reduction of RSAas a consequence of high Po2 availability. In addition,we previously reported preliminary data about morpho-logical changes of sea bass D. labrax gills to these dif-ferent Po2, showing that mild hypoxic and hyperoxicstress conditions lead to morphological changes of gillfilaments (Rinaldi et al., 2003).
In mammals, hypoxia is a triggering stimulus forblood vessel remodelling, altered cellular permeabilityand angiogenesis (Marti & Risau, 1998). All these effectsare related to induction of the Vascular EndothelialGrowth Factor (VEGF) by hypoxia. VEGF is an endo-thelial cell specific mitogen and is important not onlyfor vasculogenesis and angiogenesis, but also for main-tenance of existing blood vessels (Horowitz et al.,1997). Its biological effects are mediated by three spe-cific cell surface-expressed receptors, VEGFR-1 (Flt-1),VEGFR-2 (Flk-1), and VEGFR-3 (Flt-4), which belong tothe platelet derived growth factor receptor subfamily oftyrosine-kinase receptors (Neufeld et al., 1999). TheVEGF-A family comprises five closely related isoformswhich are expressed in different tissues: VEGFl65 is themost abundant and most biologically active of the fiveforms in vivo, and is commonly used in studies investi-gating the biological effects of VEGF. In addition, it hasbeen demonstrated that, under hypoxic conditions,
Dow
nloa
ded
by [
91.7
2.14
7.25
4] a
t 05:
18 2
2 M
arch
201
4

104 L. RINALDI, P. BASSO, G. TETTAMANTI, A. GRIMALDI, G. TEROVA, M. SAROGLIA, M. DE EGUILEOR
VEGF and VEGF receptor-2 synthesis is induced throughbinding of protein factors, Hypoxia-inducible factor 1(HIF-1) and factor 2 (HIF-2), to the hypoxia response el-ement (HRE) localized upstream of the VEGF and VEGFreceptor-2 gene (Ema et al., 1997; Tian et al, 1997). Un-like the ubiquitously expressed HIF-1, HIF-2 is essentiallypresent in endothelial cells (Wiesener et al., 2003).
The present study was carried out with the aims of cor-relating the effects of different Po2 conditions on lamellamorphology and on induction of VEGF and its receptors.
MATERIALS AND METHODS
Fish and experimental conditions
Sea bass, 150 ± 20 g body weight, were divided into three sep-arate experimental groups (10 animals/group). Each group waskept for 10 days in 200-1 tanks, supplied with a recirculationbrackish water system (water salinity 10%o) at a constant tempera-ture of 20 ± 0.5 CC. The total gas pressure, measured with a No-vatec 3000 tensionometer, ranged from 98-102% of the expectedsaturation value, while dissolved oxygen concentration, moni-tored with a Bob-Cat 9000® oxygen probe-data-logger and con-trolled with addition of pure oxygen, was normally maintained at90-100% of the saturation value (131.9-146.6 torr). Carbon dioxidenever exceeded 5-0 torr and pH was controlled at 7.4-7.5 with afeed-back system automatically adding NaHCO3 according toneed. In each tank, all the above-mentioned parameters werecontinuously monitored and controlled. After 10 days, Po2 wasmodified in two tanks, thus creating three different oxygenregime ranges:
Tank 1. Normoxia group (i.e., control group): 90-100% (131.9--146.6 torr).Tank 2. Mild hypoxia group: 60-70% saturation (87.9-102.6 torr).Tank 3. Mild hyperoxia group: 120-130% saturation (175.9-190.6torr).The main oxygen supply was reduced and pure oxygen was
added according to the regime, utilizing a microbubble generatordevice in all the tanks, apart from the mild hypoxia group. Freeammonia concentration was < 30 ug I"1 as N-NHj, pH ranged from7.4 to 7.5. After 3 months of acclimatization to the different condi-tions, all animals (there was 100% survival) were anaesthetizedwith 0.1% MS-222 (Sandoz®) and sacrificed. The surgical removalswere performed at the first and the second left gill arches.
Samples of twenty gill filaments were examined. For morpho-logical and immunohistochemical analysis, sections were obtainedas shown in Figure 1. Within each treatment group, three animalswere used for morphological study, three for immunohistochemi-cal analysis and four for biochemical procedures.
Light and transmission electron microscopy
The gills were fixed for 4 h at room temperature in a solution of2% glutaraldehyde in 0.1 M Na-cacodylate buffer pH 7.2, thenwashed in the same buffer and post-fixed for 1 h with 1% osmicacid in 0.1 M Na-cacodylate buffer (pH 7.2) at room temperature. Af-ter following the standard procedure of serial ethanol dehydration,the specimens were embedded in an Epon-Araldite 812 mixture.
Semi-thin sections (750 nm) were stained using conventionalmethods (crystal violet and basic fuchsin) (Moore et al., I960) andobserved with a BH2 light microscope (Olympus, Tokyo, Japan).Thin sections (80 nm) were observed with a Jeol 1010 EX elec-tron microscope <Jeo'. Tokyo, Japan).
Immunocytochemistry
Paraffin embeddingThe samples, fixed in 4% paraformaldehyde in 0.2 M Phosphate
Fig. 1 - Schematic representation of a gill arch portion: the sec-tioning plane is indicated.
Buffered Saline (PBS) pH 7.2 for 6 h at room temperature, werewashed five times in PBS, dehydrated in an ethanol series, treatedin Bioclear (Bio-Optica, Milan, Italy) and embedded in paraffin.Sections (6 urn) were cut with a Jung Multicut 2045 microtome(Leica, Nussolch, Germany) and placed on 3-aminoproryltri-ethoxy-silane (AES)-coated slides.
ImmunofluorescenceDeparaffinized sections were rehydrated in an ethanol series,
washed in distilled water, and treated for 15 min with an 0.05 Msolution of NH4CI to reduce autofluorescence (De la Lande & Wa-terson, 1968). After washing with PBS, sections were incubated ina dark moist chamber for 1 h with the following primary antibod-ies/antisera: a) anti-human VEGFj65 polyclonal antibody (Neo-Markers, Fremont, U.S.A.; diluted 1:20); b) anti-human Flt-1/mono-clonal antibody (Sigma, Milan, Italy; diluted 1:20); c) anti-humanFlk-1 monoclonal antibody (Sigma; diluted 1:50); d) anti-humanHIF-2 polyclonal antibody (Santa Cruz Biotechnology, CA, U.S.A.;diluted 1:50); e) anti-mouse Myosin Light Chain monoclonal anti-body (Sigma; diluted 1:50). After incubation with primary antibod-ies, the specimens were washed and incubated in a dark moistchamber with appropriate FITC-conjugated or Cy3-conjugated sec-ondary antibodies (Jackson, Immuno Research Laboratories, WestGrove, PA, U.S.A.; diluted 1:100).
The PBS buffer used for the washing steps and antibody dilu-tions contained 2% bovine serum albumin (BSA). In the controlsamples, primary antibodies were omitted and sections weretreated with BSA-containing PBS.
Coverslips were mounted in Vectashield mounting medium forfluorescence (Vector Laboratories, Burlingame, CA, U.SA) and theslides were examined with a fluorescence microscope (Olympus).
Alcian blue pH 2.5The specific technique was used to detect acid mucopolysac-
charides (Daniel & Zanin, 1997). Paraffin sections were rehydrat-ed in an ethanol series; after being washed in distilled water, thesections were incubated for 30 min with an Alcian blue solution(0.05 g in 15 ml of a 3% acetic acid solution in distilled water) pH2.5. The coverslips were mounted with Eukitt after being repeat-edly washed with 3% acetic acid and distilled water, and observedwith a light microscope. Data are presented as means ± SE. Statis-tical differences were estimated by Student's t-test. Differenceswere considered significant at P < 0.01.
Biochemical procedures
Specimens of D. labrax gills were destroyed in liquid nitrogen
Dow
nloa
ded
by [
91.7
2.14
7.25
4] a
t 05:
18 2
2 M
arch
201
4

EFFECTS OF DIFFERENT OXYGEN LEVELS IN SEA BASS GILLS 105
with a mortar. For SDS-polyacrylamide gel electrophoresis (SDS--PAGE), homogenates were suspended in extraction buffer(Laemmli's Buffer 2x) in the presence of commercial protease in-hibitor cocktail (water-soluble protease inhibitor specific for ser-ine, cysteine, aspartate and metalloproteases) (Sigma); paniculatematerial was removed by centrifugation at 13,000 rpm for 10 minat 4 °C in a refrigerated Eppendorf Minispin microcentrifuge. Su-pernatants were denatured at 100 °C for 10 min.
SDS-PAGEAnalytical SDS-PAGE using 5%, 7.5% and 15% acrylamide
minigels was performed at 200 V for 1 h. Molecular weights weredetermined by concurrently running broad and high range stan-dards from Bio-Rad (Bio-Rad, Richmond, MA, U.S.A.) or New Eng-land Biolabs (Hertfordshire, U.K.). Gels were stained with 2.5%Coomassie blue (Bio-Rad) in methanol-acetic acid-water (5:1:5).
Western blotProteins separated by SDS-PAGE were transferred onto Bio-Rad
nitrocellulose filters. Before immunostaining, membranes were sat-urated with 2% bovine serum albumin, 0.01% Tween 20 in TrisBuffered Saline (TBS, 20 mM Tris-HCl buffer, 500 mM NaCl pH
7.5) at room temperature for 2 h. Nitrocellulose membranes wereincubated overnight at 4 °C with different primary polyclonal anti-bodies: a) anti-human VEGF (diluted 1:250); b) anti-human Flk-1monoclonal antibody (diluted 1:250); c) anti-human HIF-2 poly-clonal antibody (diluted 1:250). After three washes of the mem-brane with TBS, antigens were revealed with appropriate sec-ondary antibodies diluted 1:500 if conjugated with alkaline phos-phatase (Sigma), or diluted 1:5000 if conjugated with peroxidase(Jackson). Immunoreactivity was detected with SIGMA FASTBCIP/NBT (Sigma) or with the SuperSignal West Pico Chemilumi-nescent Substrate (Pierce Biotechnology, Rockford, U.S.A.).
RESULTS
Normoxic condition
The gills of sea bass reared under normoxic condi-tions showed a typical regular structure (Fig. 2) charac-terized by protruding lamellae at each side of the fila-
Figs 2-6 - 2-3 - Light and electron micrographs of a gill filament in normoxia condition. Note the regular distribution of lamellae (L) andthe shape of pillar cells (P) with a large nucleus and cytoplasmic projections (arrowheads). E: erythrocytes. Bars: 50 urn, 5 pm. 4 -Myosin immunostainings in gill lamellae: the signal is localized in the peripheral cytoplasm of pillar cells. Bar: 10 urn. 5-6 - Electron mi-crographs of mucous (Fig. 5) and chloride cells (Fig. 6). Bars: 5 pm.
Dow
nloa
ded
by [
91.7
2.14
7.25
4] a
t 05:
18 2
2 M
arch
201
4

106 L. RINALDI, P. BASSO, G. TETTAMANTI, A. GRIMALDI, G. TEROVA, M. SAROGLIA, M. DE EGUILEOR
ment. Flattened lamellae, showing ordered and parallelarrangement, were formed by a coaxial system made upof wrapping epithelium and inner endothelium support-ed by pillar cells (also referred to as Bietrix cells) withtypical "hourglass" shape. The central portion of pillarcells was occupied by a large nucleus, while flattenedcytoplasmic projections were laterally expanded (Fig.3). Pillar and endothelial cells lined the vascular lumenwhere red blood cells freely circulated. Pillar cells, spe-cialized endothelial cells, were stained by the myosinantibody, which was localized in the peripheral cyto-plasm (Fig. 4). Mucous and chloride cells (Figs 5-6)were distributed along the gill filament structure.
Mild hyperoxic condition
The gills of sea bass reared under mild hyperoxic con-ditions showed a regular structure (Fig. 7) with flattenedlamellae ordered and parallel disposed. Nevertheless,pillar cells were frequently disjoined from the basal lami-na (Fig. 8) and this process allowed the appearance ofmyosin filament bundles in the cytoplasm (Fig. 9); de-tached membranes lined small peripheral microenviron-ments (Fig. 8) characterized by laminar folds (Fig. 10).
Mild hypoxia condition
The gills of sea bass reared under mild hypoxia condi-tions lost all regular structure (Fig. 11): lamellae were dis-organized, twisted and showed frequent apical blebs(Fig. 12). In particular, there were many lacunae betweenthe epithelial and endothelial layers, which caused themassive deformation of the lamellae. The pillar cellsseemed to be contracted, the vascular lumen was re-duced and the red blood cells were deformed (Fig. 13).
Immunohistochemistry
Immunohistochemical analysis (Figs 14-25) revealedthe presence of VEGF and its receptor Flk-1 in the en-dothelial cells of lamellae of fishes reared under nor-moxic (Figs 14 and 17) and mild hypoxic conditions(Figs 16 and 19). On the contrary, a lower signal wasdetected in hyperoxic samples (Figs 15 and 18). HIF-2was markedly expressed in the epithelial layer undernormoxic and mild hypoxic conditions (Figs 20 and 22),but it was absent in mild hyperoxia (Fig. 21).
Unlike the three factors mentioned above, Flt-1 wasnot found to be expressed under the three differentconditions analysed (Figs 23-25).
ÍO3J •.
í -
Figs 7-10 - Light and electron micrographs of a gill filament in mild hyperoxia. Pillar cell (P) detaches from the basal lamina through acontraction of the myosin filaments (9, arrow) leading to the consequent formation of enclosed vesicles aligned on the periphery of thecell (8, arrowheads). These microenvironments are lined by laminar cytoplasmic expansions (10). E: erythrocytes. Bars: 50 urn, 2 pm, 1um, 0.3 um.
Dow
nloa
ded
by [
91.7
2.14
7.25
4] a
t 05:
18 2
2 M
arch
201
4
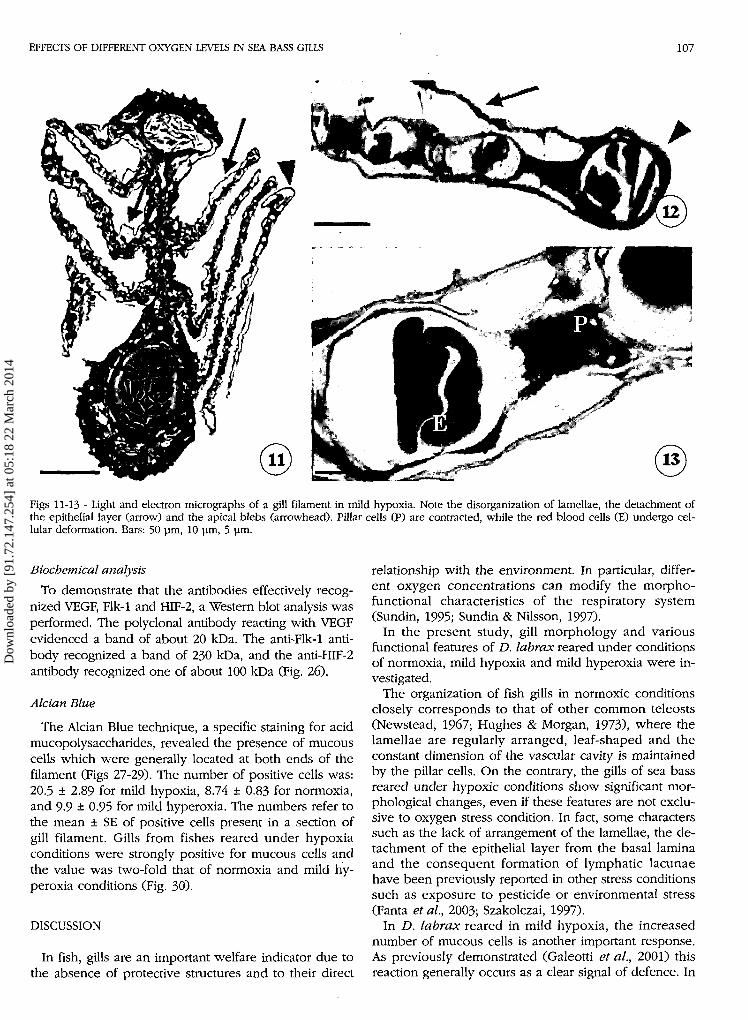
EFFECTS OF DIFFERENT OXYGEN LEVELS IN SEA BASS GILLS 107
Figs 11-13 - Light and electron micrographs of a gill filament in mild hypoxia. Note the disorganization of lamellae, the detachment ofthe epithelial layer (arrow) and the apical blebs (arrowhead). Pillar cells (P) are contracted, while the red blood cells (E) undergo cel-lular deformation. Bars: 50 pm, 10 pm, 5 pm.
Biochemical analysisTo demonstrate that the antibodies effectively recog-
nized VEGF, Flk-1 and HIF-2, a Western blot analysis wasperformed. The polyclonal antibody reacting with VEGFevidenced a band of about 20 kDa. The anti-Flk-1 anti-body recognized a band of 230 kDa, and the anti-HIF-2antibody recognized one of about 100 kDa (Fig. 26).
Aldan Blue
The Alcian Blue technique, a specific staining for acidmucopolysaccharides, revealed the presence of mucouscells which were generally located at both ends of thefilament (Figs 27-29). The number of positive cells was:20.5 ± 2.89 for mild hypoxia, 8.74 ± 0.83 for normoxia,and 9-9 ± 0.95 for mild hyperoxia. The numbers refer tothe mean ± SE of positive cells present in a section ofgill filament. Gills from fishes reared under hypoxiaconditions were strongly positive for mucous cells andthe value was two-fold that of normoxia and mild hy-peroxia conditions (Fig. 30).
DISCUSSION
In fish, gills are an important welfare indicator due tothe absence of protective structures and to their direct
relationship with the environment. In particular, differ-ent oxygen concentrations can modify the morpho-functional characteristics of the respiratory system(Sundin, 1995; Sundin & Nilsson, 1997).
In the present study, gill morphology and variousfunctional features of D. labrax reared under conditionsof normoxia, mild hypoxia and mild hyperoxia were in-vestigated.
The organization of fish gills in normoxic conditionsclosely corresponds to that of other common teleosts(Newstead, 1967; Hughes & Morgan, 1973), where thelamellae are regularly arranged, leaf-shaped and theconstant dimension of the vascular cavity is maintainedby the pillar cells. On the contrary, the gills of sea bassreared under hypoxic conditions show significant mor-phological changes, even if these features are not exclu-sive to oxygen stress condition. In fact, some characterssuch as the lack of arrangement of the lamellae, the de-tachment of the epithelial layer from the basal laminaand the consequent formation of lymphatic lacunaehave been previously reported in other stress conditionssuch as exposure to pesticide or environmental stress(Fanta et al, 2003; Szakolczai, 1997).
In D. labrax reared in mild hypoxia, the increasednumber of mucous cells is another important response.As previously demonstrated (Galeotti et al., 2001) thisreaction generally occurs as a clear signal of defence. In
Dow
nloa
ded
by [
91.7
2.14
7.25
4] a
t 05:
18 2
2 M
arch
201
4

108 L. RINALDI, P. BASSO, G. TETTAMANTI, A. GRIMALDI, G. TEROVA, M. SAROGLIA, M. DE EGUILEOR
normoxia hyperoxia hypoxia
Figs 14-25 - 14-16 - Immunodetections of VEGF in gill endothelium under normoxia, mild hyperoxia and mild hypoxia conditions. Bars:30 um. 17-22 - Immunodetections of Flk-1 (green) and HIF-2 (red) respectively in gill endothelium and epithelium under normoxia,mild hyperoxia and mild hypoxia conditions. Bars: 20 pm, 10 urn, 10 um, 5 um, 10 um, 10 um. 23-25 - Immunodetections of Flt-1shows no signal in any conditions. Bars: 30 pm.
Dow
nloa
ded
by [
91.7
2.14
7.25
4] a
t 05:
18 2
2 M
arch
201
4

EFFECTS OF DIFFERENT OXYGEN LEVELS IN SEA BASS GILLS 109
66-
2 6 - I
20 - f
200-
-20kDa
116-
-230 kDa
115-93
49-
35-
29-
21-
-100 kDa
VEGF Flk-1 HIF-2
Fig. 26 - Western blot analysis of D. labrax gills. The anti-VEGF antibody recognizes a band of about 20 kDa, the anti-Flk-1 antibodyrecognizes one of 230 kDa, and the anti-HIF-2 antibody one of about 100 kDa.
addition, Khunyakari et al. (2001) reported that fishesexposed to heavy metal pollution develop an increasein mucous cells on the gill epithelium. In the case of in-sufficient oxygen supply, we can hypothesize a doublerole for the increased mucous secretion: first for de-fence, since mucous contains lysozyme that could avoid
possible infections, and second for increasing oxygensolubility. The number of chloride cells did not showevident changes in the three oxygen conditions.
Two specific morpho-functional changes related tomild hypoxia are represented by the apical blebs of thelamellae and the reduced vascular space: the restricted
5.M « v
% ..rmm -
1«- if .-/•%• '
Figs 27-29 -Alcian Blue technique. Gills under mild hypoxia (27), normoxia (28), mild hyperoxia (29) conditions. Mucous cells arestained in blue and are arranged at both ends of the filaments. In hypoxia (27) the number of mucous cells is twice that of those pre-sent in normoxia and hyperoxia (28-29). Bars: 50 urn.
Dow
nloa
ded
by [
91.7
2.14
7.25
4] a
t 05:
18 2
2 M
arch
201
4

110 L. RINALDI, P. BASSO, G. TETTAMANTI, A. GRIMALDI, G. TEROVA, M. SAROGLIA, M. DE EGUILEOR
lumen of the vessel could force erythrocytes to moveinto a reduced space thus giving rise to cellular defor-mation. Also in D. pabrax as proposed by Nilsson et al.(1995) in rainbow trout and in roach, this deformationcould be an adaptation to enhance oxygen uptake,since it can counteract the formation of a diffusionboundary layer around the erythrocytes and can causemixing of intracellular haemoglobin molecules. Howev-er, due to the narrow passage, cells have to pass slowlythrough the lamella thus favouring oxygen uptake andthe local increase of intralamellar haematocrit.
The gills of sea bass reared under mild hyperoxic con-ditions appear similar to those subjected to normoxicones, maintaining an ordered distribution of the lamellaeand the primary sites of oxygen uptake, except for thepillar cells which have undergone structural changes. Infact, these cells show a peculiar general organizationand shape, probably due to the stretching of their cyto-plasmic acto-myosin system responsible for the detach-ment of the cell membrane from the basal lamina. Con-sequently, the detached and overlapped membranesgive rise to a series of aligned microenvironments linedby the laminar cytoplasmic process: the morpho-func-tional modification of the pillar cells leads to the en-largement of the vascular space where the erythrocytesmove freely, thus assuring optimal gas exchange.
Although the gross morphology of teleostean gills hasbeen extensively studied, little is known about in-ducibility of VEGF (a molecule fundamental not onlyfor angiogenesis but also for endothelial permeability),VEGF receptors and hypoxia-inducible factors (HIF-1,HIF-2) in relation to different Po2 in gills.
The immunohistochemical analysis revealed a highexpression of VEGF and Flk-1 in the lamellae of D.labrax reared under mild hypoxic and normoxic condi-tions with only small differences between the two,while VEGF and its receptor were practically unde-tectable in mild hyperoxic condition.
15
* *
T
*T
* *T
Hypoxia Normoxia Hyperoxia
Fig. 30 - Number of mucous cells in cross-sectioned gill filaments.Each value represents the mean of 20 cross-sectioned gills fromfive different fishes. *,**: the number of mucous cells in hypoxiacondition is significantly different from those in normoxia and hy-peroxia conditions (P < 0.01).
Our results on VEGF expression are similar to previ-ous data about mice from Marti and Risau (1998) whoshowed that endothelial cell maintenance-related VEGFexpression is rather unresponsive to hypoxia, whereascell angiogenesis-related VEGF gene expression appearsto be tightly regulated by oxygen: we hypothesize thatin sea bass the Flk-1 receptor is subjected to the sameregulation. Our hypothesis is supported by the patternthat we found for HIF-2, a hypoxia-inducible transcrip-tion factor, which usually induces many genes such asVEGF and Flk-1. HIF-2 expression is normally abundantin fish gills and therefore also in D. labrax the HIF-2/VEGF/Flk-1 signalling pathway may be an importantregulating mechanism for vessel maintenance.
In mild hyperoxic conditions, lamellae show weak pos-itivity for VEGF and Flk-1 antibodies. These results are inaccordance with Klekamp's previous data which showedthat exposure to high levels of oxygen causes a down-regulation of VEGF and Flk-1 in adult rat lungs (Klekampet al., 1999) and with the above mentioned hypothesisabout the HIF-2 /VEGF/Flk-1 pathway, since the gills inmild hyperoxia showed no HIF-2 expression, supportinga possible down-regulation also for this factor.
We did not find any positivity for Flt-1 in sea bass,and this is in accordance with the results of Liao et al.(1997) in zebrafish. In fact those Authors characterizedthe presence of only one tyrosine kinase receptor,whose sequence has several regions of marked homolo-gy with human and mouse Flk-1 and Flt-1: thus, theyhypothesized that this sequence, which they called Flk-1,represents an ancestral gene which, only in higher ver-tebrates, diversified into two different molecules (Flk-1and Flt-1).
Summarizing, our results allow an evaluation of bothhypoxic and hyperoxic fish conditions: in particular,mild hypoxia is easily recognizable due to changes inlamellae morphology and high expression of factors in-volved in the regulation of endothelial cells, i.e. VEGF,Flk-1 and HIF-2; conversely, mild hyperoxia can be dis-tinguished from normoxia due to lamellae morphologyand low expression of VEGF, Flk-1 and HIF-2.
Sea bass D. labrax is one of the most reared speciesin Mediterranean aquaculture and a preventive recogni-tion of respiratory stress conditions is important to en-sure fish welfare. In particular, high and prolonged hy-poxia is responsible for the appearence of debilitatingconditions that lead to death; on the contrary, mild hy-poxia and hyperoxia usually determine transient effectson fish: thus, while mild hypoxia is dangerous as it de-termines reversible sufferance symptoms, mild hyperox-ia could be considered a useful parameter for the im-provement of fish health in aquaculture systems.
In addition, since in aquaculture Po2 is a critical para-meter (Scapigliati et al., 1999) which allows fish to bereared at high densities with significant physiologicaladvantages, it is interesting to evaluate any possiblemarker that can give information about fish perfor-mances within artificial rearing environments.
Dow
nloa
ded
by [
91.7
2.14
7.25
4] a
t 05:
18 2
2 M
arch
201
4

EFFECTS OF DIFFERENT OXYGEN LEVELS IN SEA BASS GILLS 111
REFERENCES
Barthelemy L., Beland A., Chastel C., 1981 - A comparative studyof oxygen toxicity in vertebrates. Respir. Physiol., 44: 261-268.
Booth J. A., Amer S., 1979 - Circulation in trout gills: the relation-ship between branchial perfusion and the width of the lamel-lar blood space. Can. J. Zool., 57: 2183-2185.
Daniel S., Zanin T., 1997 - Manuale di tecnica cito-istologica. Do-cumentazione Scientifica Editrice, Bologna.
De la Lande I. S., Waterson J. G., 1968 - Modification of autofluo-rescence in the formaldehyde-treated rabbit ear artery by Evansblue. J. Histochem. Cytochem., 16: 281-282.
Ema M., Taya S., Yokotani N., Sogawa K., Matsuda Y., Fujii--Kuriyama Y., 1997 - A novel bHLH-PAS factor with close se-quence similarity to hypoxia-inducible factor l a regulates theVEGF expression and is potentially involved in lung and vascu-lar development. Proc. Natl. Acad. Sci. U.S.A., 94: 4273-4278.
Fanta E., Rios F. S., Romao S., Vianna A. C., Freiberger S., 2003 -Histopathology of the fish Corydoras paleatus contaminatedwith sublethal levels of organophosphorus in water and food.Ecotoxicol. Environ. Saf., 54: 119-130.
Farrell A. P., Sobin S. S., Randall D. J., Crosby S., 1980 - In-tralamellar blood flow patterns in fish gills. Am. J. Physiol., 263:R741-R746.
Galeotti M., Volpatti D., Berton D., Beraldo P., Contessi B., Gus-mani, L, 2001 - Patologie in acquacoltura. In: S. Cataudella & P.Bronzi (eds), Acquacoltura responsabile. Unimar-Uniprom, Ro-ma, pp. 200-201.
Horowitz J. R., Rivard A., van der Zee R., Hariawala M., Sheriff D.D., Esakof D. D., Chaudhry G. M., Symes J. F., Isner J. M., 1997- Vascular endothelial growth factor/vascular permeability factorproduces nitric oxide-dependent hypotension. Evidence for amaintenance role in quiescent adult endothelium. Arterio.Thromb. Vasc. Biol., 17: 2793-2800.
Hughes G. M., Morgan M., 1973 - The structure of fish gills in re-lation to their respiratory function. Biol. Rev., 48: 419-475.
Khunyakari R. P., Tare V., Sharma R. N., 2001 - Effects of sometrace heavy metals on Poecilia reticulata (Peters). J. Environ.Biol., 22: 141-144.
Klekamp J. G., Jarzecka K., Perkett E. A., 1999 - Exposure to hy-peroxia decreases the expression of vascular endothelialgrowth factor and its receptors in adult rat lungs. Am. J. Pathol.,154: 823-831.
Laurent P., 1984 - Gill internal morphology. In: W. S. Hoar & D. J.Randall (eds), Fish physiology. Academic Press, Orlando, pp.73-183.
Leknes I. L., 1985 - Ultrastructural and ultrahistochemical studiesof post-mortem changes and effects of hypoxia in the bony fishheart. J. Submicroscopic. Cytol, 17: 223-228.
Liao W., Bisgrove B. W., Sawyer H., Hug B., Bell B., Peters K. G.,Grunwald D. J., Stainier D. Y. R., 1997 - The zebrafish genecloche acts upstream of a flk-1 homologue to regulate endothe-lial cell differentiation. Development, 124: 381-389.
Marti H. H., Risau W., 1998 - Systemic hypoxia changes the organ--specific distribution of vascular endothelial growth factor andits receptors. Proc. Natl. Acad. Sci. U.S.A., 95: 15809-15814.
Moore R. D., Mumaw V., Shonberg M. D., I960 - Optical microscopyof ultrathin tissue sections. J. Ultrastruct. Res., 4: 113-116.
Neufeld G., Cohen T., Gengrinovitch S., Poltorak Z., 1999 - Vascu-lar endothelial growth factor (VEGF) and its receptors. FASEBJ., 13: 9-21.
Newstead J. D., 1967 - Fine structure of the respiratory lamellae ofteleostean gills. Z. Zellforsch. Mikrosk Anat., 79: 396-428.
Nilsson G. E., Lofman C. O., Block M., 1995 - Extensive erythro-cyte deformation in fish gills observed by in vivo microscopy:apparent adaptations for enhancing oxygen uptake. J. Exp. Bi-ol., 198: 1151-1156.
Rinaldi L., Grimaldi A., Tettamanti G., Terova G., Saroglia M., Val-vassori R., de Eguileor M., 2003 - Oxygen level influences thestructural organization of gills in Dicentrarchus labrax. Biol.Mar. Mediterr, 10: 1133-1135.
Saroglia M., Cecchini S., Terova G., Caputo A., De Stradis A., 2000- Influence of environmental temperature and water oxygenconcentration on gas diffusion distance in sea bass (Dicentrar-chus labrax L.). Fish Physiol. Biochem., 23: 55-58.
Saroglia M., Terova G., De Stradis A., Caputo A., 2002 - Morpho-metric adaptations of sea bass gills to different dissolved oxy-gen partial pressure. J. Fish Biol., 60: 1423-1430.
Scapigliati G., Scalia D., Marras A., Meloni S., Mazzini M., 1999 -Immunoglobulin levels in the teleost sea bass Dicentrarchuslabrax (L.) in relation to age, season, and water oxygenation.Aquaculture, 174: 207-212.
Soivio A., Hughes G. M., 1978 - Circulatory changes in secondarylamellae of Salmo gairdneri gills in hypoxia and anesthesia.Ann. Zool. Fenn., 15: 221-225.
Soivio A., Tuurala H., 1981 - Structural and circulatory responsesto hypoxia in the secondary lamellae of Salmo gairdneri gills attwo temperatures. J. Comp. Physiol., 45: 37-43.
Sundin L., 1995 - Responses of the branchial circulation to hypox-ia in the Atlantic cod, Gadus morhua. Am. J. Physiol. Regul. In-tegr. Comp. Physiol., 268: R771-R778.
Sundin L., Nilsson G. E., 1997 - Neurochemical mechanisms be-hind gill microcirculatory responses to hypoxia in trout: in vivomicroscopy study. Am. J. Physiol. Regul. Integr. Comp. Physiol.,272: R576-R585.
Szakolczai J., 1997 - Histopathological changes induced by envi-ronmental stress in common carp, Japanese coloured carp, Eu-ropean eel and African catfish. Acta Vet. Hung., 45: 1-10.
Tian H., McKnight S. L, Russel D. W., 1997 - Endothelial PAS do-main protein 1 (EPAS 1), a transcription factor selectively ex-pressed in endothelial cells. Genes Dev., 11: 72-82.
Wiesener M. S., Jurgensen J. S., Rosenberg C., Scholze C.,Horstrup J. H., Warnecke C., Mandriota S., Bechmann I., Frei U.A., Pugh C. W., Ratcliffe P. J., Bachmann S., Maxwell P. H.,Eckardt K. U., 2003 - Widespread, hypoxia-inducible expressionof HIF-2α in distinct cell populations of different organs. FASEBJ., 17: 271-273.
Dow
nloa
ded
by [
91.7
2.14
7.25
4] a
t 05:
18 2
2 M
arch
201
4




















