
Modulation of ion uptake across posterior gills of the crab Chasmagnathus granulatus by dopamine and...
7
Modulation of ion uptake across posterior gills of the crab Chasmagnathus granulatus by dopamine and cAMP J. Halperin a,b, * , G. Genovese a,b , M. Tresguerres a , C.M. Luquet a,b a Departamento de Biodiversidad y Biologı ´a Experimental, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, Pabello ´n II, Ciudad Universitaria (C1428EHA) Buenos Aires, Argentina b CONICET (Consejo Nacional de Investigaciones Cientı ´ficas y Te ´cnicas) Rivadavia 1917 (C1033AAJ) Buenos Aires, Argentina Received 12 December 2003; received in revised form 27 May 2004; accepted 20 July 2004 Abstract Cyclic AMP (cAMP) and dopamine modulate ion uptake across isolated and perfused posterior gills of Chasmagnathus granulatus acclimated to 10x salinity. Addition of cAMP agonists, such as cp-cAMP, forskolin, and IBMX, produced a significant increase in the transepithelial potential difference (Vte ), which reflects ion transport activity. Dopamine (DA) also had a stimulatory effect on ion uptake, increasing Vte and Na + influx, although this effect was transient, since both variables remained elevated for less than 30 min. In addition, the dose–response curve for DA concentration-Vte was biphasic, and the maximum stimulation was obtained with 10 Amol l 1 . When the effects of forskolin and DA on the Na + /K + -ATPase activity were tested, they correlated well with the Vte and Na + influx experiments; the enzyme activity increased significantly after preincubation of gill fragments for 10 min with forskolin or DA (51 and 64%, respectively), but there was no effect after pre-incubation with DA for 20 min. Finally, KT5720, a specific inhibitor of cAMP-dependent protein kinase (PKA), completely abolished the stimulatory effect of DA on Vte , suggesting the involvement of PKA in this mechanism. D 2004 Elsevier Inc. All rights reserved. Keywords: cAMP; Chasmagnathus granulatus ; Crab; Dopamine; Gills; Ion transport; Na + /K + -ATPase; PKA; Transepithelial potential differences 1. Introduction Hyper-regulating crabs compensate for diffusive salt loss by actively absorbing NaCl across the posterior gills (Mantel, 1985). These organs are lined with a specialized epithelium composed of thick cells with great numbers of mitochondria that supply the energy for the active transport of ions against their diffusional gradients (Taylor and Taylor, 1992; Pe ´queux, 1995). Located at the basolateral membrane of these cells, the enzyme Na + /K + -ATPase plays a principal role in ion uptake from dilute media, driving sodium uptake (and, in some cases, also chloride) across the gill epithelium of crabs experiencing hypo-osmotic stress (Siebers et al., 1985; Towle and Kays, 1986; Onken and Riestenpatt, 1998; Onken et al., 2003). Na + /K + -ATPase shows both a long-term and a rapid regulation in hypo-osmotic conditions. The former typically takes 1–2 weeks after transferring crabs from full-strength to dilute seawater, and it implies an increase in the enzyme activity measured in posterior gill homogenates (see Lucu and Towle, 2003, for a review). Recently, this response was linked to increased expression of Na + /K + -ATPase a-subunit mRNA in the blue crab Callinectes sapidus (Lucu and Towle, 2003), suggesting that the long-term response depends on the synthesis of more units of this enzyme. On the other hand, rapid activation of Na + /K + -ATPase can be achieved in several different ways, the most studied being neurohormonal action. Injections of hemolymph from crabs acclimated to low-salinity (Savage and Robinson, 1983), dopamine (DA), and pericardial organ extract (Sommer and Mantel, 1988) have all been reported to 1095-6433/$ - see front matter D 2004 Elsevier Inc. All rights reserved. doi:10.1016/j.cbpb.2004.07.001 * Corresponding author. Department of Physiology and Biophysics, University of Illinois at Chicago, 835 S. Wolcott St., M/C 901, Chicago, IL 60612, USA. Tel.: +1 312 996 7688; fax: +1 312 996 1414. E-mail address: [email protected] (J. Halperin). Comparative Biochemistry and Physiology, Part A 139 (2004) 103– 109 www.elsevier.com/locate/cbpa
Transcript of Modulation of ion uptake across posterior gills of the crab Chasmagnathus granulatus by dopamine and...

www.elsevier.com/locate/cbpa
Comparative Biochemistry and Physiol
Modulation of ion uptake across posterior gills of the crab
Chasmagnathus granulatus by dopamine and cAMP
J. Halperina,b,*, G. Genovesea,b, M. Tresguerresa, C.M. Luqueta,b
aDepartamento de Biodiversidad y Biologıa Experimental, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, Pabellon II,
Ciudad Universitaria (C1428EHA) Buenos Aires, ArgentinabCONICET (Consejo Nacional de Investigaciones Cientıficas y Tecnicas) Rivadavia 1917 (C1033AAJ) Buenos Aires, Argentina
Received 12 December 2003; received in revised form 27 May 2004; accepted 20 July 2004
Abstract
Cyclic AMP (cAMP) and dopamine modulate ion uptake across isolated and perfused posterior gills of Chasmagnathus granulatus
acclimated to 10x salinity. Addition of cAMP agonists, such as cp-cAMP, forskolin, and IBMX, produced a significant increase in the
transepithelial potential difference (Vte), which reflects ion transport activity. Dopamine (DA) also had a stimulatory effect on ion uptake,
increasing Vte and Na+ influx, although this effect was transient, since both variables remained elevated for less than 30 min. In addition, the
dose–response curve for DA concentration-Vte was biphasic, and the maximum stimulation was obtained with 10 Amol l�1. When the effects
of forskolin and DA on the Na+/K+-ATPase activity were tested, they correlated well with the Vte and Na+ influx experiments; the enzyme
activity increased significantly after preincubation of gill fragments for 10 min with forskolin or DA (51 and 64%, respectively), but there
was no effect after pre-incubation with DA for 20 min. Finally, KT5720, a specific inhibitor of cAMP-dependent protein kinase (PKA),
completely abolished the stimulatory effect of DA on Vte, suggesting the involvement of PKA in this mechanism.
D 2004 Elsevier Inc. All rights reserved.
Keywords: cAMP; Chasmagnathus granulatus; Crab; Dopamine; Gills; Ion transport; Na+/K+-ATPase; PKA; Transepithelial potential differences
1. Introduction
Hyper-regulating crabs compensate for diffusive salt loss
by actively absorbing NaCl across the posterior gills
(Mantel, 1985). These organs are lined with a specialized
epithelium composed of thick cells with great numbers of
mitochondria that supply the energy for the active transport
of ions against their diffusional gradients (Taylor and
Taylor, 1992; Pequeux, 1995). Located at the basolateral
membrane of these cells, the enzyme Na+/K+-ATPase plays
a principal role in ion uptake from dilute media, driving
sodium uptake (and, in some cases, also chloride) across the
gill epithelium of crabs experiencing hypo-osmotic stress
1095-6433/$ - see front matter D 2004 Elsevier Inc. All rights reserved.
doi:10.1016/j.cbpb.2004.07.001
* Corresponding author. Department of Physiology and Biophysics,
University of Illinois at Chicago, 835 S. Wolcott St., M/C 901, Chicago, IL
60612, USA. Tel.: +1 312 996 7688; fax: +1 312 996 1414.
E-mail address: [email protected] (J. Halperin).
(Siebers et al., 1985; Towle and Kays, 1986; Onken and
Riestenpatt, 1998; Onken et al., 2003).
Na+/K+-ATPase shows both a long-term and a rapid
regulation in hypo-osmotic conditions. The former typically
takes 1–2 weeks after transferring crabs from full-strength to
dilute seawater, and it implies an increase in the enzyme
activity measured in posterior gill homogenates (see Lucu
and Towle, 2003, for a review). Recently, this response was
linked to increased expression of Na+/K+-ATPase a-subunit
mRNA in the blue crab Callinectes sapidus (Lucu and
Towle, 2003), suggesting that the long-term response
depends on the synthesis of more units of this enzyme.
On the other hand, rapid activation of Na+/K+-ATPase
can be achieved in several different ways, the most studied
being neurohormonal action. Injections of hemolymph from
crabs acclimated to low-salinity (Savage and Robinson,
1983), dopamine (DA), and pericardial organ extract
(Sommer and Mantel, 1988) have all been reported to
ogy, Part A 139 (2004) 103–109

J. Halperin et al. / Comparative Biochemistry and Physiology, Part A 139 (2004) 103–109104
produce a short-term activation of the Na+/K+-ATPase in
aquatic crabs. In addition, Trausch et al. (1989) have shown
an increase of protein phosphorylation through DA and
serotonin (5-HT) receptors in the gills of the Chinese mitten
crab Eriocheir sinensis and a positive influence of this
phosphorylation on Na+/K+-ATPase activity. The intra-
cellular second messenger likely involved in the response
to these neurohormones is 3V–5-Vcyclic adenosine mono-
phosphate (cAMP), since DA, 5-HT, and octopamine
elevate its intracellular content in the gills of C. sapidus
and E. sinensis (Kamemoto and Oyama, 1985; Lohrmann
and Kamemoto, 1987; Bianchini and Gilles, 1990; Mo et al.,
1998). However, in E. sinensis, there are contradictory
reports about the relationship between Na+/K+-ATPase and
cAMP. While incubation of isolated gills with dibutyryl
(db)-cAMP produced an activation of the enzyme (Mo et al.,
1998), Riestenpatt et al. (1994) working on split gill
lamellae mounted in an Ussing chamber found that the
principal effect of db-cAMP on Na+ transport was to
increase the current and conductivity of this ion at constant
electromotive force. These authors concluded that db-cAMP
increased the transcellular Na+ conductivity through apical
Na+ channels without direct stimulation of the Na+/K+-
ATPase or that, at least, this stimulation was unimportant.
Despite these conflicting results on the Chinese crab, the
association between hypo-osmotic stress–DA–cAMP–Na+/
K+–ATPase and a fast activation of ion uptake in aquatic
hyper-regulating crabs is well documented and generally
accepted (see Morris, 2001 for a review).
In contrast, the role of dopamine in the regulation of ion
uptake in terrestrial and semi-terrestrial crabs might be
different, due to the rapidly changing conditions that these
crabs must face regularly. The animals must cope with
sudden changes in the osmotic conditions due to tide action,
exposure to air with the consequent evaporation of gill
chamber water, or entrance into rain pools. In the semi-
terrestrial purple shore crab Leptograpsus variegatus, DA or
cAMP increases the branchial Na+/K+-ATPase activity when
injected into intact animals (Morris and Edwards, 1995). It
was concluded that DA enhanced Na+/K+-ATPase activity
via cAMP, however, direct evidence for this relationship
was not provided.
Chasmagnathus granulatus (Dana, 1851) is a grapsoid
crab that inhabits estuaries in Brazil, Uruguay and Argentina
(Boschi, 1964). Adults of this species actively leave water
and are able to regulate acid–base balance after several
hours of air exposure (Luquet et al., 1998). C. granulatus is
well adapted to cope with extreme tidal and seasonal
fluctuations in salinity (D’Incao et al., 1992; Anger et al.,
1994; Spivak et al., 1994), thanks to its great osmoregula-
tory capacity both in oligohaline and hyperhaline media
(Charmantier et al., 2002). This capacity is based on active
absorption and excretion of ions through the epithelium of
the posterior gills, using the Na+/K+-ATPase as the main
driving force (Luquet et al., 2002; Onken et al., 2003). The
aim of this work was to investigate the role of dopamine and
cAMP in the regulation of ion uptake and Na+/K+-ATPase
activity in a species that experiences both rapid and long-
term salinity variations.
2. Materials and methods
2.1. Animals
Adult male crabs in intermoult stage C (Drach and
Tchernigovtzeff, 1967) of C. granulatus species were
collected at Punta Rasa Beach, San Clemente del Tuyu,
Buenos Aires, Argentina. Animals were kept in plastic
containers with aerated artificial brackish water of 10xsalinity and fed twice a week with pellets of rabbit food.
Temperature was kept a 20F1 8C with a 12L:12D photo-
period. Animals with an average carapace width of
29.7F0.4 mm were chosen for the study.
2.2. Gill perfusion
Crabs were sacrificed by destroying the nervous system
with a spike and scissors. After removing the dorsal
carapace, gills number 6 (the largest among posterior gills)
were gently excised, and prepared according to Siebers et
al., 1985. The afferent and efferent vessels were connected
by fine polyethylene tubing (a=0.4 mm) to a peristaltic
pump (afferent) and to a collecting tube (efferent). The
tubing was held in position by an acrylic clamp covered
with smooth neoprene to minimize gill damage. The
preparation was bathed in a beaker containing approx-
imately 25 ml of aerated saline and was perfused at a rate of
0.1 ml min�1. The perfusate was collected in a second
beaker. Under these conditions, the preparation remains
viable for several hours; gills not showing stable Vte after
the usual stabilization period were discarded.
2.3. Transepithelial potential difference (Vte)
Ag/AgCl electrodes were connected via agar bridges to
the external bath and to the beaker collecting the perfusate
(internal side). Electrical potential differences were meas-
ured with reference to the external medium and recorded
with a chart recorder.
2.4. Sodium flux
Unidirectional sodium influx was measured in perfused
gills by applying 22Na in the bathing solution according to
Luquet et al. (2002) at a final concentration of 0.25 ACiml�1. After Vte stabilization, at least three samples were
collected from the perfusate leaving the gills at 10-min
intervals. Finally, dopamine was added to the perfusate at a
final concentration of 30 Amol l�1 and additional six
samples were collected in the same way. Radioactivity
was measured with a Canberra Series 35 plus gamma
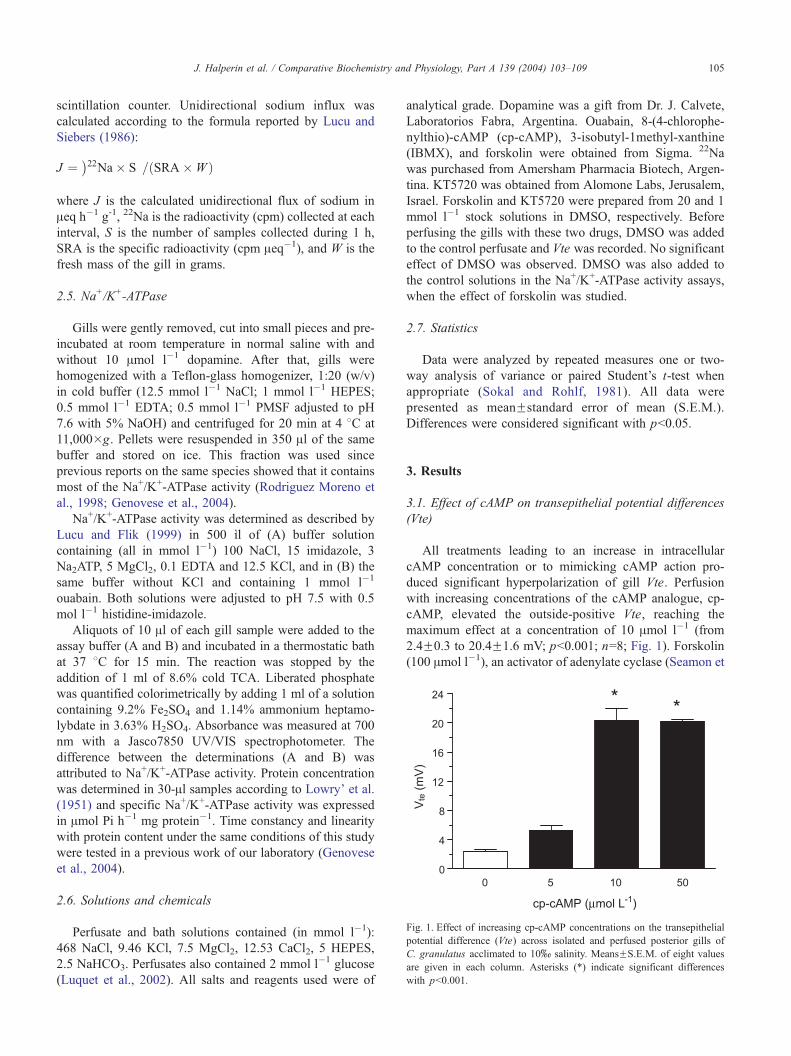
Fig. 1. Effect of increasing cp-cAMP concentrations on the transepithelial
potential difference (Vte) across isolated and perfused posterior gills of
C. granulatus acclimated to 10x salinity. MeansFS.E.M. of eight values
are given in each column. Asterisks (*) indicate significant differences
with pb0.001.
J. Halperin et al. / Comparative Biochemistry and Physiology, Part A 139 (2004) 103–109 105
scintillation counter. Unidirectional sodium influx was
calculated according to the formula reported by Lucu and
Siebers (1986):
J ¼ 22Na� S� �
= SRA�Wð Þ
where J is the calculated unidirectional flux of sodium in
Aeq h�1 g-1, 22Na is the radioactivity (cpm) collected at each
interval, S is the number of samples collected during 1 h,
SRA is the specific radioactivity (cpm Aeq�1), and W is the
fresh mass of the gill in grams.
2.5. Na+/K+-ATPase
Gills were gently removed, cut into small pieces and pre-
incubated at room temperature in normal saline with and
without 10 Amol l�1 dopamine. After that, gills were
homogenized with a Teflon-glass homogenizer, 1:20 (w/v)
in cold buffer (12.5 mmol l�1 NaCl; 1 mmol l�1 HEPES;
0.5 mmol l�1 EDTA; 0.5 mmol l�1 PMSF adjusted to pH
7.6 with 5% NaOH) and centrifuged for 20 min at 4 8C at
11,000�g. Pellets were resuspended in 350 Al of the same
buffer and stored on ice. This fraction was used since
previous reports on the same species showed that it contains
most of the Na+/K+-ATPase activity (Rodriguez Moreno et
al., 1998; Genovese et al., 2004).
Na+/K+-ATPase activity was determined as described by
Lucu and Flik (1999) in 500 ıl of (A) buffer solution
containing (all in mmol l�1) 100 NaCl, 15 imidazole, 3
Na2ATP, 5 MgCl2, 0.1 EDTA and 12.5 KCl, and in (B) the
same buffer without KCl and containing 1 mmol l�1
ouabain. Both solutions were adjusted to pH 7.5 with 0.5
mol l�1 histidine-imidazole.
Aliquots of 10 Al of each gill sample were added to the
assay buffer (A and B) and incubated in a thermostatic bath
at 37 8C for 15 min. The reaction was stopped by the
addition of 1 ml of 8.6% cold TCA. Liberated phosphate
was quantified colorimetrically by adding 1 ml of a solution
containing 9.2% Fe2SO4 and 1.14% ammonium heptamo-
lybdate in 3.63% H2SO4. Absorbance was measured at 700
nm with a Jasco7850 UV/VIS spectrophotometer. The
difference between the determinations (A and B) was
attributed to Na+/K+-ATPase activity. Protein concentration
was determined in 30-Al samples according to Lowry’ et al.
(1951) and specific Na+/K+-ATPase activity was expressed
in Amol Pi h�1 mg protein�1. Time constancy and linearity
with protein content under the same conditions of this study
were tested in a previous work of our laboratory (Genovese
et al., 2004).
2.6. Solutions and chemicals
Perfusate and bath solutions contained (in mmol l�1):
468 NaCl, 9.46 KCl, 7.5 MgCl2, 12.53 CaCl2, 5 HEPES,
2.5 NaHCO3. Perfusates also contained 2 mmol l�1 glucose
(Luquet et al., 2002). All salts and reagents used were of
analytical grade. Dopamine was a gift from Dr. J. Calvete,
Laboratorios Fabra, Argentina. Ouabain, 8-(4-chlorophe-
nylthio)-cAMP (cp-cAMP), 3-isobutyl-1methyl-xanthine
(IBMX), and forskolin were obtained from Sigma. 22Na
was purchased from Amersham Pharmacia Biotech, Argen-
tina. KT5720 was obtained from Alomone Labs, Jerusalem,
Israel. Forskolin and KT5720 were prepared from 20 and 1
mmol l�1 stock solutions in DMSO, respectively. Before
perfusing the gills with these two drugs, DMSO was added
to the control perfusate and Vte was recorded. No significant
effect of DMSO was observed. DMSO was also added to
the control solutions in the Na+/K+-ATPase activity assays,
when the effect of forskolin was studied.
2.7. Statistics
Data were analyzed by repeated measures one or two-
way analysis of variance or paired Student’s t-test when
appropriate (Sokal and Rohlf, 1981). All data were
presented as meanFstandard error of mean (S.E.M.).
Differences were considered significant with pb0.05.
3. Results
3.1. Effect of cAMP on transepithelial potential differences
(Vte)
All treatments leading to an increase in intracellular
cAMP concentration or to mimicking cAMP action pro-
duced significant hyperpolarization of gill Vte. Perfusion
with increasing concentrations of the cAMP analogue, cp-
cAMP, elevated the outside-positive Vte, reaching the
maximum effect at a concentration of 10 Amol l�1 (from
2.4F0.3 to 20.4F1.6 mV; pb0.001; n=8; Fig. 1). Forskolin
(100 Amol l�1), an activator of adenylate cyclase (Seamon et

J. Halperin et al. / Comparative Biochemistry and Physiology, Part A 139 (2004) 103–109106
al., 1981), also augmented Vte dramatically (from 2.5F0.7
to 15.0F1.5 mV; pb0.001; n=5). Phosphodiesterase is the
enzyme that catalyzes the degradation of cAMP. Inhibition
of this enzyme with 500 Amol l�1 IBMX (Chasin and
Harris, 1976) caused a much less pronounced effect on Vte
than cp-cAMP but a significant stimulation was still
observed (from 2F0.2 to 2.7F0.1 mV; pb0.05; n=6). All
Fig. 2. Representative time courses of transepithelial potential differences
(Vte) across isolated and perfused posterior gills of C. granulatus
acclimated to 10x salinity. Arrows indicate addition of cp-cAMP 10 Amol
mol l�1 (a), forskolin 100 Amol l�1 (b) and IBMX 500 Amol l�1 (c).
Fig. 3. Representative time courses of the effect of dopamine on the
transepithelial potential difference (Vte) across isolated and perfused
posterior gills. The arrow indicates the addition of different concentrations
of dopamine (DA) to the perfusate. 10 Amol l�1 of DA produced the
maximum stimulation, producing a more than twofold increase in the
control Vte ( pb0.05, n=7).
these effects were sustained for at least 1 h. Representative
time courses of these experiments are shown in Fig. 2.
3.2. Effect of dopamine (DA) on Vte
Representative time courses of Vte stimulation by
different concentrations of DA are shown in Fig. 3. The
minimum concentration that produced significant hyper-
polarization was 5 Amol l�1, while the highest response was
obtained with 10 Amol l�1 (from 3.2F0.4 to 8.3F0.5 mV;
pb0.05, N=7). At higher concentrations (50 and 500 Amol
l�1), the effect was still significant but less pronounced.
Hence, the dose–response curve of Vte of gills vs. DA
concentration showed a biphasic profile (Fig. 4). All the DA
Fig. 4. Effect of different dopamine concentrations on the transepithelial
potential difference (Vte) across isolated and perfused posterior gills of
C. granulatus acclimated to 10x salinity. (a) Time course and (b)
DVte=gain in Vte (treated�control). DVte increased significantly from
the corresponding controls (*pb0.05; n=4–10) with all concentrations
tested except for 1 Amol l�1 ( pN0.05, n=6).

Fig. 6. Effect of KT5720 on the transepithelial potential differences (Vte)
across isolated and perfused posterior gills treated with dopamine. (a)
Representative time course (filled circles: dopamine alone; empty circles:
dopamine plus KT5720). (b) MeanFS.E.M of five values for each
treatment. Asterisks indicate statistical differences after dopamine treatment
( pb0.05). Dopamine (DA) was added at a concentration of 10 Amol l�1.
KT5720 (a specific PKA inhibitor) at 250 nmol l�1.
J. Halperin et al. / Comparative Biochemistry and Physiology, Part A 139 (2004) 103–109 107
concentrations tested produced transient effects; Vte
remained stimulated for less than 30 min.
3.3. Effect of DA on sodium influx
DA at 10 Amol l�1 did not produce consistent effects in
this experiment. A higher concentration, 30 Amol l�1,
produced a significant but transient stimulation of sodium
influx, similar to the effect of DA on Vte. Maximum
augmentation of 34% (from 398F73 to 534F119 Aeq h�1
g�1; pb0.05; n=5) was recorded 30 min after the addition of
dopamine. Sodium influx declined to control values 30 min
later (60 min after the addition of DA, see Fig. 5).
3.4. Linkage between DA action and cAMP-dependent
protein kinase
In order to assess whether dopamine stimulates ion
transport through a pathway involving the cAMP-dependent
protein kinase (PKA), KT5720, a specific cell permeable
inhibitor of this enzyme (Kase et al., 1987) was added to the
perfusate before and during perfusion with dopamine. As
shown in Fig. 6a, 250 nmol l�1 KT5720 almost completely
abolished the stimulation of Vte caused by dopamine. Fig.
6b depicts the effects of dopamine on control and KT5720-
treated gills. KT5720 alone had no significant effect.
3.5. Na+/K+-ATPase
The stimulatory effects of cAMP and DA are likely due
to an activation of one or several ion-transporting proteins,
thus producing an increase in ion uptake that is reflected in
the Vte. Since the Na+/K+-ATPase generates the main
driving force for ion uptake in this epithelium (Luquet et
al., 2002; Onken et al., 2003), we tested the effect of
forskolin and DA on the activity of this enzyme. Pre-
incubation of gill fragments with 100 Amol l�1 forskolin for
Fig. 5. Representative time course of the effect of dopamine on sodium
influx across an isolated and perfused posterior gill of C. granulatus
acclimated to 10x salinity. Arrow indicates the addition of 30 Amol l�1
dopamine (DA).
10 min produced a significant increase of Na+/K+-ATPase
activity (from 20.5F2.5 to 30.9F2.8 Amol Pi h�1 mg�1;
n=5). After 10 min of pre-incubation with 10 Amol l�1
dopamine, a significant increase in Na+/K+-ATPase activity,
from 24F3.1 to 39.3F2.8 Amol Pi h�1 mg�1, was also
recorded (N=13). When the gills were incubated with
Fig. 7. Na+/K+-ATPase activity in posterior gills pre-incubated with
forskolin (100 Amol l�1, 10 min, n=6) or dopamine (10 Amol l�1, 10 or
20 min, n=13 for each time). The data are given as percentages (FS.E.M.)
of control value (Na+/K+-ATPase activity of the paired, untreated gill).
Asterisks indicate statistical differences between treated and control gills,
with pb0.05.

J. Halperin et al. / Comparative Biochemistry and Physiology, Part A 139 (2004) 103–109108
dopamine for 20 min, no significant increase from control
values was observed. These results correlate well with the
Vte and sodium influx experiments. Fig. 7 illustrates the
percentage increase in Na+/K+-ATPase activity after pre-
incubation with forskolin and dopamine.
4. Discussion
There is a general consensus within the literature about
the role of dopamine as a neuroendocrine factor involved in
the response of euryhaline aquatic crabs to dilution of the
ambient salinity. The prevailing concept is that DA
enhances ion uptake by activating the Na+/K+-ATPase,
and that the second messenger involved in this regulation
seems to be cAMP. Activation of this signaling pathway has
frequently been observed to result in increased sodium
influxes that compensate for salt lost during hypo-osmotic
stress (see Kamemoto, 1991; Morris, 2001 for reviews).
In this study, we showed that DA enhances trans-
epithelial potential difference (Vte), sodium uptake and
Na+/K+-ATPase activity in posterior gills of C. granulatus.
Activators of the PKA transduction pathway, cp-cAMP,
forskolin and IBMX, produced comparable stimulation of
Vte and Na+/K+-ATPase activity. The connection between
DA and the cAMP-PKA transductional pathway was
studied by adding a specific inhibitor of PKA (KT5720)
to perfused gills before the addition of DA, while monitor-
ing the Vte. The complete abolition of the DA effect by
KT5720 is direct evidence showing that the dopamine
stimulation of ion transport across C. granulatus gills
depends on PKA. Trausch et al. (1989) have demonstrated
dopamine and serotonin-dependant phosphorylation of
proteins in a microsomal fraction containing Na+/K+-
ATPase of E. sinensis gill homogenates. Moreover, they
have been able to inhibit the serotonin-induced phosphor-
ylation with ouabain, a specific inhibitor of Na+/K+-ATPase.
Recently, Towle et al. (2001) have shown a putative site for
phosphorylation by PKA in the sequence of C. sapidus gill
Na+/K+-ATPase. All these findings together lead to the
conclusion that dopamine binds to specific receptors at the
gill ionocyte basolateral membrane and increases cAMP
levels, leading to PKA-mediated phosphorylation of Na+/
K+-ATPase.
So far, the action of DA in the gills of C. granulatus
follows the model proposed for aquatic crabs. Novel results
are obtained when the time course of the response is taken
into account. In all of our experiments, the effect of DA is
transient, lasting no more than 30 min, in contrast to the
sustained stimulation caused by all the agonists of the PKA
pathway tested. Vte and sodium influx show the same
response after the addition of DA: an instant but transient
increase followed by a return to control levels. Modulation
of the Na+/K+-ATPase activity by DA follows the same
temporal pattern, indicating that it is one of the ion-
transporting proteins involved in this mechanism.
Perfusion experiments performed on gills of aquatic
hyper-regulating crabs have shown sustained effects of
dopamine, positively linked to cAMP (Kamemoto and
Oyama, 1985; Mo et al., 1998). The transient effects of
DA in C. granulatus may indicate a more complex
regulatory system, probably related to the amphibious
lifestyle of this species, which experiences sudden changes
from full to diluted or even to concentrated seawater. In
another amphibious crab, L. variegatus, DA has been
reported to increase Na+/K+-ATPase activity, presumably
by increasing cAMP levels (Morris and Edwards, 1995).
However, nothing is known about the time course of this
response, and, interestingly, it was only obtained in studies
of intact animals; no effect was seen on isolated gills. The
few fully terrestrial crabs studied show very different
regulatory patterns, including a response to serotonin but
not to DA or cAMP in Gecarcoidea natalis and inhibiting
effects of dopamine and cAMP in Birgus latro (see Morris,
2001 for a review).
Although the activation of the PKA pathway by DA is
evident from our results, the transient effect of this bioamine
in C. granulatus suggests the presence of either a second
signaling pathway leading to the total reversion of the initial
changes or a rapid deactivation of the receptors or signaling
intermediates. This complex response could also be related
to the existence of different DA receptors in the gills of this
species. Trausch et al. (1989) have prevented the effects of
DA on the Vte of isolated gills of E. sinensis by perfusing
with the D2 antagonist, chlorpromazine. More recently, Mo
et al. (2002) have characterized D1 dopamine receptors in
the gills of the same species by binding of the D1
antagonist, SCH23390. The temporal pattern of the response
to dopamine as well as the presence and interaction of D1
and D2 receptors in ion-regulating crab gills still require
extensive investigation.
References
Anger, K., Spivak, E., Bas, C., Ismael, D., Luppi, T., 1994. Hatching
rhythms and dispersion of decapod crustacean larvae in a brackish
coastal lagoon in Argentina. Helgol. Meeresunters. 48, 445–466.
Bianchini, A., Gilles, R., 1990. Cyclic AMP as a modulator of NaCl
transport in gills of the euryhaline chinese crab Eriocheir sinensis. Mar.
Biol. 104, 191–195.
Boschi, E.E., 1964. Los crustaceos decapodos brachyura del litoral
bonaerense. Bol. Inst. Biol. Mar. 164, 1–34.
Charmantier, G., Gimenez, L., Charmantier-Daures, M., Anger, K., 2002.
Ontogeny of osmoregulation, physiological plasticity and larval export
strategy in the grapsid crab Chasmagnathus granulata (Crustacea,
Decapoda). Mar. Ecol., Prog. Ser. 229, 185–194.
Chasin, M., Harris, D.N., 1976. Inhibitors and activators of cyclic
nucleotide phosphodiesterase. Adv. Cyclic Nucleotide 7, 225–264.
D’Incao, F., Ruffino, M.L., Silva, K.G., Braga, A.C., 1992. Response of
Chasmagnathus granulata Dana (Decapoda, Grapsidae) to salt-marsh
environmental variations. J. Exp. Mar. Biol. Ecol. 161, 179–188.
Drach, P., Tchernigovtzeff, C., 1967. Sur la methode de determination des
stades d’intermue et son application generale aux crustaces. Vie Milieu
18, 597–607.

J. Halperin et al. / Comparative Biochemistry and Physiology, Part A 139 (2004) 103–109 109
Genovese, G., Luchetti, C.G., Luquet, C.M., 2004. Na+/K+-ATPase activity
and gill ultrastructure in the hyper-hypo-regulating crab Chasmagna-
thus granulatus acclimated to dilute, normal and concentrated seawater.
Mar. Biol. 144, 111–118.
Kamemoto, F.I., 1991. Neuroendocrinology of osmoregulaton in crabs.
Zool. Sci. 8, 827–833.
Kamemoto, F.I., Oyama, S.N., 1985. Neuroendocrine influence on effector
tissues of hydromineral balance in crustaceans. In: Lofts, B., Holmes,
W.N. (Eds.), Current Trends in Comparative Endocrinology. Hong
Kong Univ. Press, Hong Kong, pp. 883–886.
Kase, H., Iwahashi, K., Nakanishi, S., Matsuda, Y., Yamada, K., Takahashi,
M., Murakata, C., Sato, A., Kaneka, M., 1987. K252 compounds: novel
and potent inhibitors of protein kinase C and cyclic-nucleotide-
dependent kinases. Biochem. Biophys. Res. Commun. 142, 436–440.
Lohrmann, D.M., Kamemoto, F.I., 1987. The effect of dibutyryl cAMP on
sodium uptake by isolated perfused gills of Callinictes sapidus. Gen.
Comp. Endocrinol. 65, 300–305.
Lowry, O.H., Rosebrough, N.J., Farr, A.L., Randall, R.J., 1951. Protein
measurement with the Folin phenol reagent. J. Biol. Chem. 193, 265–275.
Lucu, C., Flik, G., 1999. Na+-K+-ATPase and Na+/Ca+ exchange activities
in gills of hyperegulating Carcinus maenas. Am. J. Physiol. 276,
R490–R499.
Lucu, C., Siebers, D., 1986. Amilorid-sensitive sodium flux and potentials
in perfused Carcinus gill preparations. J. Exp. Biol. 122, 25–35.
Lucu, C., Towle, D.W., 2003. Na++K+-ATPase in gills of aquatic crustacea.
Comp. Biochem. Physiol. C 135, 195–214.
Luquet, C.M., Cervino, C.O., Ansaldo, M., Pereyra Carrera, V., Kocmur, S.,
Dezi, R.E., 1998. Physiological response to emersion in the amphibious
crab Chasmagnathus granulata Dana (Decapoda Grapsidae):
biochemical and ventilatory adaptations. Comp. Biochem. Physiol., A
121, 385–393.
Luquet, C.M., Postel, U., Halperin, J., Urcola, M.R., Marques, R., Siebers,
D., 2002. Transepithelial potential differences and Na+ flux in isolated
perfused gills of the crab Chasmagnathus granulatus (Grapsidae)
acclimated to hyper- and hypo-salinity. J. Exp. Biol. 205, 71–77.
Mantel, L.H., 1985. Neurohormonal integration of osmotic and ionic
regulation. Am. Zool. 25, 253–263.
Mo, J.L., Devos, P., Trausch, G., 1998. Dopamine as a modulator of ionic
transport and Na+/K+-ATPase activity in the gills of the Chinese crab
Eriocheir sinensis. J. Crustac. Biol. 18, 442–448.
Mo, J.L., Devos, P., Trausch, G., 2002. Dopamine D1 receptors in the gills
of Chinese crab Eriocheir sinensis. Comp. Biochem. Physiol. C 131,
433–438.
Morris, S., 2001. Neuroendocrine regulation of osmoregulation and the
evolution of air-breathing in decapod crustaceans. J. Exp. Biol. 204,
979–989.
Morris, S., Edwards, T., 1995. Control of osmoregulation via regulation of
Na+/K+-ATPase activity in the amphibious purple shore crab Leptog-
rapsus variegatus. Comp. Biochem. Physiol. C 112, 129–136.
Onken, H., Riestenpatt, S., 1998. NaCl absorption across split gill lamellae
of hyperregulating crabs: transport mechanisms and their regulation.
Comp. Biochem. Physiol., A 119, 883–893.
Onken, H., Tresguerres, M., Luquet, C.M., 2003. Active NaCl absorption
across posterior gills of hyperosmoregulating Chasmagnathus gran-
ulatus. J. Exp. Biol. 206, 1017–1023.
Pequeux, A., 1995. Osmotic regulation in crustaceans. J. Crustac. Biol. 15,
1–60.
Riestenpatt, S., Zeiske, W., Onken, H., 1994. Cyclic AMP stimulation of
electrogenic uptake of Na+ and Cl� across the gill epithelium of the
Chinese crab Eriocheir sinensis. J. Exp. Biol. 188, 159–174.
Rodriguez Moreno, P.A., Schwarzbaum, P.J., Rodriguez, E.M., 1998.
Effects of cadmium on gill Na,K-ATPase of the estuarine crab
Chasmagnathus granulata (Decapoda, Grapsidae) during postmolt: in
vivo and in vitro studies. Bull. Environ. Contam. Toxicol. 61, 629–636.
Savage, J.P., Robinson, G.D., 1983. Inducement of increased gill Na+/K+-
ATPase activity by a hemolymph factor in hyperosmoregulating
Callinectes sapidus. Comp. Biochem. Physiol., A 75, 65–69.
Seamon, K.B., Padgett, W., Daly, J.W., 1981. Forskolin: unique deterpene
activator of adenylate cyclase in membranes and in intact cells. Proc.
Natl. Acad. Sci. U. S. A. 78, 3363–3367.
Siebers, D., Winkler, A., Lucu, C., Thedens, G., Weichar, tD., 1985. Na-K-
ATPase generates an active transport potential in the gills of the
hyperregulating shore crab Carcinus maenas. Mar. Biol. 87, 185–192.
Sokal, R.R., Rohlf, F.J., 1981. Biometry. W.H. Freeman, New York.
Sommer, M.J., Mantel, L.H., 1988. Effect of dopamine, cyclic AMP, and
pericardial organs on sodium uptake and Na/K-ATPase activity in gills
of the green crab Carcinus maenas (L). J. Exp. Zool. 248, 272–277.
Spivak, E., Anger, K., Luppi, T., Bas, C., Ismael, D., 1994. Distribution
and habitat preferences of two grapsid crab species in Mar Chiquita
Lagoon (Province of Buenos Aires, Argentina). Helgol. Meeresunters.
48, 59–78.
Taylor, H.H., Taylor, E.W., 1992. Gills and lungs: the exchange of gases
and ions. In: Harrison, F.W., Humes, A.G. (Eds.), Microscopic
Anatomy of Invertebrates. Wiley-Liss, New York, pp. 203–293.
Towle, D.W., Kays, W.T., 1986. Basolateral localization of Na K -ATPase
in gill epithelium of two osmoregulating crabs, Callinectes sapidus and
Carcinus maenas. J. Exp. Zool. 239, 311–318.
Towle, D.W., Paulsen, R.S., Weihrauch, D., Kordylewski, M., Salvador, C.,
Lignot, J.H., Spanings-Pierrot, C., 2001. Na++K+-ATPase in gills of the
blue crab Callinectes sapidus: cDNA sequencing and salinity-related
expression of a-subunit mRNA and protein. J. Exp. Biol. 204,
4005–4012.
Trausch, G., Forge, tM.-C., Devos, P., 1989. Bioamines-stimulated
phosphorylation and (Na+, K+)-ATPase in the gills of the chinese crab,
Eriocheir sinensis. Comp. Biochem. Physiol., B 94, 487–492.




















