
The IκB kinase inhibitor sulfasalazine impairs long-term memory in the crab Chasmagnathus
12
THE IUB KINASE INHIBITOR SULFASALAZINE IMPAIRS LONG-TERM MEMORY IN THE CRAB CHASMAGNATHUS E. MERLO, R. FREUDENTHAL and A. ROMANO Laboratorio de Neurobiolog| ¤a de la Memoria, Facultad de Ciencias Exactas y Naturales, Departamento de Ciencias Biolo ¤ gicas, Pabello ¤ n II, Universidad de Buenos Aires, (1428) Buenos Aires, Argentina AbstractɆEvidence for the participation of Rel/NF-UB transcription factors in long-term memory has recently been reported in the context-signal learning paradigm of the crab Chasmagnathus, in which a high correlation between long-term memory formation and NF-UB activation was observed. Two components of the NF-UB pathway in the crab brain have now been identi¢ed by cross-immunoreactivity using mammalian antibodies for IUB-K and IUB kinase K. Furthermore, IUB kinase-like phosphotransferase activity, which was inhibited by the IUB kinase inhibitor sulfasala- zine, was detected in brain extracts. We have evaluated the e¡ect of sulfasalazine administration on long-term memory tested at 48 h. Amnesia was found when sulfasalazine was administered pre-training and 5 h after training but not at 0 or 24 h after training. Thus, two periods for sulfasalazine-induced amnesia were found in coincidence with the two phases of NF-UB activation previously described (immediately and 6 h after training). The cyclooxygenase inhibitor indomethacin did not induce amnesia when administered pre-training. Thus, the possibility that sulfasalazine induces amnesia by means of cyclooxygenase inhibition is unlikely to be tenable. In vivo sulfasalazine inhibition of basal NF-UB activity was found between 30 and 45 min after injection, as assessed by electrophoretic mobility shift assay. On the other hand, in vivo sulfasalazine administration 6 h after training inhibited the second phase of training-induced NF-UB activation, providing evidence that the sulfasalazine e¡ect on memory is due to a direct e¡ect of the drug on the NF-UB pathway. These results provide the ¢rst evidence that IUB kinase and NF-UB activation are necessary for memory formation. ȣ 2002 IBRO. Published by Elsevier Science Ltd. All rights reserved. Key words: long-term memory, IUB kinase, Rel/NF-UB, neural plasticity, crab. An almost universal feature of long-lasting changes in neural plasticity and in memory consolidation is the reg- ulation of transcription and translation processes (Agrano¡, 1972; Barondes, 1975; Davis and Squire, 1984; Crow and Forrester, 1990; Pedreira et al., 1995). Some pathways by which signals, originating from syn- aptic activity, reach the nucleus have been proposed (e.g., Alberini et al., 1994; Meberg et al., 1996; Deisseroth et al., 1996; Wellmann et al., 2001). All of them involve the activation of constitutive transcription factors that rapidly activate transcription in response to external signals. It has been demonstrated that the cAMP response element binding protein family of tran- scription factors is implicated in memory and neuronal plasticity in both vertebrates and invertebrates (Kaang et al., 1993; Yin et al., 1995; Bernabeu et al., 1997; Bourtchuladze et al., 1998). More recently, results obtained with the context-signal learning paradigm of the crab Chasmagnathus have implicated the participa- tion of the Rel/nuclear factor-UB (NF-UB) family in long-term memory (LTM) formation (Freudenthal et al., 1998; Freudenthal and Romano, 2000). Spaced train- ing yields a protein synthesis-dependent LTM which cor- relates with NF-UB activation in the nucleus of the crab central brain. Conversely, NF-UB was not activated after massed training which in turn induces a protein synthe- sis-independent intermediate-term memory (ITM). NF-UB was also activated in synaptosomal extract from central brain of animals receiving spaced training (Freudenthal and Romano, 2000), supporting the hypothesis of a local activation of NF-UB as part of a synapse-to-nucleus signaling system (Kaltschmidt et al., 1993; Meberg et al., 1996). Two phases of Rel/NF-UB activation were found in crabs that received spaced train- ing. The ¢rst phase, found after 10 or more trials, is coincident with the number of trials required for memory formation. Such activation decayed to basal level 3 h after training. The second phase was found 6 h after 161 *Corresponding author. Tel. : +54-11-45763348 ; fax : +54-11-45763384. E-mail address : [email protected] (A. Romano). Abbreviations : ANOVA, analysis of variance ; COX, cyclooxyge- nase; CT, control group; DMSO, dimethylsulfoxide; EDTA, eth- ylenediaminetetra-acetate ; EMSA, electrophoretic mobility shift assay ; GST, glutathione S-transferase ; HEPES, N-(2-hydroxye- thyl)piperazine-NP(2-ethanesulfonic acid); IUB, NF-UB inhibitor ; IKK, IUB kinase; IND, indomethacin group; ITI, intertrial inter- val ; ITM, intermediate-term memory ; LTM, long-term memory ; NF-UB, nuclear factor UB; NMDA, N-methyl-D-aspartate ; NV, naive ; PAGE, polyacrylamide gel electrophoresis ; PBS, phos- phate-bu¡ered saline ; PT, pre-training test ; ROD, relative optical density; SAL, saline group; SDS, sodium dodecylsulfate; SSZ, sulfasalazine group; TNF, tumor necrosis factor; TR, trained group ; VEH, vehicle group. NSC 5503 22-5-02 www.neuroscience-ibro.com Neuroscience Vol. 112, No. 1, pp. 161^172, 2002 ȣ 2002 IBRO. Published by Elsevier Science Ltd All rights reserved. Printed in Great Britain PII:S0306-4522(02)00049-0 0306-4522 / 02 $22.00+0.00
Transcript of The IκB kinase inhibitor sulfasalazine impairs long-term memory in the crab Chasmagnathus

THE IUB KINASE INHIBITOR SULFASALAZINE IMPAIRS LONG-TERMMEMORY IN THE CRAB CHASMAGNATHUS
E. MERLO, R. FREUDENTHAL and A. ROMANO�
Laboratorio de Neurobiolog|¤a de la Memoria, Facultad de Ciencias Exactas y Naturales, Departamento de Ciencias Biolo¤gicas,Pabello¤n II, Universidad de Buenos Aires, (1428) Buenos Aires, Argentina
Abstract4Evidence for the participation of Rel/NF-UB transcription factors in long-term memory has recently beenreported in the context-signal learning paradigm of the crab Chasmagnathus, in which a high correlation betweenlong-term memory formation and NF-UB activation was observed. Two components of the NF-UB pathway in thecrab brain have now been identi¢ed by cross-immunoreactivity using mammalian antibodies for IUB-K and IUB kinaseK. Furthermore, IUB kinase-like phosphotransferase activity, which was inhibited by the IUB kinase inhibitor sulfasala-zine, was detected in brain extracts. We have evaluated the e¡ect of sulfasalazine administration on long-term memorytested at 48 h. Amnesia was found when sulfasalazine was administered pre-training and 5 h after training but not at 0 or24 h after training. Thus, two periods for sulfasalazine-induced amnesia were found in coincidence with the two phases ofNF-UB activation previously described (immediately and 6 h after training). The cyclooxygenase inhibitor indomethacindid not induce amnesia when administered pre-training. Thus, the possibility that sulfasalazine induces amnesia by meansof cyclooxygenase inhibition is unlikely to be tenable. In vivo sulfasalazine inhibition of basal NF-UB activity was foundbetween 30 and 45 min after injection, as assessed by electrophoretic mobility shift assay. On the other hand, in vivosulfasalazine administration 6 h after training inhibited the second phase of training-induced NF-UB activation, providingevidence that the sulfasalazine e¡ect on memory is due to a direct e¡ect of the drug on the NF-UB pathway.
These results provide the ¢rst evidence that IUB kinase and NF-UB activation are necessary for memoryformation. C 2002 IBRO. Published by Elsevier Science Ltd. All rights reserved.
Key words: long-term memory, IUB kinase, Rel/NF-UB, neural plasticity, crab.
An almost universal feature of long-lasting changes inneural plasticity and in memory consolidation is the reg-ulation of transcription and translation processes(Agrano¡, 1972; Barondes, 1975; Davis and Squire,1984; Crow and Forrester, 1990; Pedreira et al., 1995).Some pathways by which signals, originating from syn-aptic activity, reach the nucleus have been proposed(e.g., Alberini et al., 1994; Meberg et al., 1996;Deisseroth et al., 1996; Wellmann et al., 2001). All ofthem involve the activation of constitutive transcription
factors that rapidly activate transcription in response toexternal signals. It has been demonstrated that thecAMP response element binding protein family of tran-scription factors is implicated in memory and neuronalplasticity in both vertebrates and invertebrates (Kaanget al., 1993; Yin et al., 1995; Bernabeu et al., 1997;Bourtchuladze et al., 1998). More recently, resultsobtained with the context-signal learning paradigm ofthe crab Chasmagnathus have implicated the participa-tion of the Rel/nuclear factor-UB (NF-UB) family inlong-term memory (LTM) formation (Freudenthal etal., 1998; Freudenthal and Romano, 2000). Spaced train-ing yields a protein synthesis-dependent LTM which cor-relates with NF-UB activation in the nucleus of the crabcentral brain. Conversely, NF-UB was not activated aftermassed training which in turn induces a protein synthe-sis-independent intermediate-term memory (ITM).NF-UB was also activated in synaptosomal extractfrom central brain of animals receiving spaced training(Freudenthal and Romano, 2000), supporting thehypothesis of a local activation of NF-UB as part of asynapse-to-nucleus signaling system (Kaltschmidt et al.,1993; Meberg et al., 1996). Two phases of Rel/NF-UBactivation were found in crabs that received spaced train-ing. The ¢rst phase, found after 10 or more trials, iscoincident with the number of trials required for memoryformation. Such activation decayed to basal level 3 hafter training. The second phase was found 6 h after
161
*Corresponding author. Tel. : +54-11-45763348;fax: +54-11-45763384.E-mail address: [email protected] (A. Romano).Abbreviations: ANOVA, analysis of variance; COX, cyclooxyge-
nase; CT, control group; DMSO, dimethylsulfoxide; EDTA, eth-ylenediaminetetra-acetate ; EMSA, electrophoretic mobility shiftassay; GST, glutathione S-transferase ; HEPES, N-(2-hydroxye-thyl)piperazine-NP(2-ethanesulfonic acid); IUB, NF-UB inhibitor ;IKK, IUB kinase; IND, indomethacin group; ITI, intertrial inter-val ; ITM, intermediate-term memory; LTM, long-term memory;NF-UB, nuclear factor UB; NMDA, N-methyl-D-aspartate ; NV,naive; PAGE, polyacrylamide gel electrophoresis ; PBS, phos-phate-bu¡ered saline; PT, pre-training test ; ROD, relative opticaldensity; SAL, saline group; SDS, sodium dodecylsulfate; SSZ,sulfasalazine group; TNF, tumor necrosis factor; TR, trainedgroup; VEH, vehicle group.
NSC 5503 22-5-02
www.neuroscience-ibro.com
Neuroscience Vol. 112, No. 1, pp. 161^172, 2002C 2002 IBRO. Published by Elsevier Science Ltd
All rights reserved. Printed in Great BritainPII: S 0 3 0 6 - 4 5 2 2 ( 0 2 ) 0 0 0 4 9 - 0 0306-4522 / 02 $22.00+0.00

training, waning to basal level between 12 and 24 h(Freudenthal and Romano, 2000).
In mammals the Rel/NF-UB family of transcriptionfactors comprises ¢ve members which share the Relhomology domain and form homo- and hetero-dimers.In Drosophila two members of the Rel subfamily, Dorsaland DIF, and a partial homologue of the NF-UB sub-family, Relish, were found. The dimers are normallyfound in cytoplasm associated with members of theNF-UB inhibitor (IUB) family (Cactus in Drosophila).The IUB proteins inhibit the transcription factor byoccluding its nuclear location sequence. In response tospeci¢c stimuli, Rel/NF-UB translocates to the nucleus asa result of the proteolytic degradation of IUB proteins.The signal for the degradation of IUB is the phosphory-lation of two serine residues followed by polyubiquitina-tion and association with the 26S proteasome (DiDonatoet al., 1996). IUB is phosphorylated by a speci¢c serinekinase multimeric complex recently identi¢ed, namedIUB kinase (IKK) complex, formed by two kinase sub-units (IKKK and IKKL) (DiDonato et al., 1997;Mercurio et al., 1997; Zandi et al., 1997) and a structuralcomponent (NEMO/IKKQ/IKKAP) (Rothwarf et al.,1998; Yamoaka et al., 1998). The mechanism by whichdistinct signals are transduced to IKK activation remainsto be elucidated (Israel, 2000). An IKK-like protein wasisolated in oyster, which shares structural and functionalproperties with its mammalian homologues, suggestingthat this component of the Rel/NF-UB pathway is evo-lutionarily conserved (Escoubas et al., 1999).
The e¡ects of non-steroidal anti-in£ammatory drugssuch as acetylsalicylic acid and sodium salicylate havebeen classically attributed to their inhibitory action onthe enzyme cyclooxygenase (COX) and the consequentdecrease in prostaglandin levels. However, an inhibitorye¡ect of acetylsalicylic acid and sodium salicylate on NF-UB was recently found (Koop and Ghosh, 1994; Grilli etal., 1996). This inhibition is due to interference of thedrug with IKKK (Yin et al., 1998), which is normallyactivated in response to immune and in£ammatory sig-nals. The drug sulfasalazine was synthesized to combinean antibiotic, sulfapyridine, and an anti-in£ammatoryagent, 5-aminosalicylic acid, and is used in chronicin£ammatory diseases. Its action was initially attributedto COX inhibition, but the e¡ect on prostaglandin levelswas controversial and, in general, sulfasalazine and itsmetabolite 5-aminosalicylic acid seem to be weak orvery weak inhibitors of COX (Hoult, 1986). Threeyears ago, Wahl et al. (1998) found that sulfasalazine,but not its metabolites, is a potent and speci¢c inhibitorof NF-UB. More recently, the same group reported directinhibition of IKKK and IKKL due to interference of thedrug with the ATP binding site (Weber et al., 2000). Thisdrug was previously used to inhibit NF-UB in the ner-vous system. The administration of sulfasalazine in theretina of rats impeded injury-induce NF-UB activationand its anti-apoptotic action (Choi et al., 2000).
In the context-signal learning paradigm of Chasmag-nathus, the repeated presentation of a danger stimulus(an opaque ¢gure passing overhead) provokes the fadingof the escape response that was initially elicited (Lozada
et al., 1990). Non-associative habituation learning isinduced by massed training (e.g., 300 trials of stimuluspresentations, with a 4-s intertrial interval (ITI)). Withthis training protocol an ITM is generated, so-called sig-nal memory, that lasts no more than 3 or 4 days and thatdoes not show up at the ¢rst testing trial but only duringthe following trials (retraining phase) of the testing ses-sion. On the other hand, spaced training (e.g., 15 trials,171-s ITI) induces an association between the iteratedstimulus and contextual features (Tomsic et al., 1998)(context-signal associative learning), which is expressedmainly at the ¢rst testing trial but is also evident at theretraining phase (Pedreira et al., 1998). A LTM is formed(context-signal memory) that lasts for at least a week andentails a di¡erent mechanism than that underlying ITM(Pedreira et al., 1998; Hermitte et al., 1999).
The aims of the present report were (1) to initiate thestudy of the mechanisms involved in NF-UB activation inthe context of LTM formation and (2) to test whetherthe interference of IKK and NF-UB activation impairsmemory consolidation.
EXPERIMENTAL PROCEDURES
Animals
Adult male Chasmagnathus granulatus intertidal crabs, 2.6^2.9cm across the carapace, weighing 17 Q 0.2 g (n= 60), were col-lected from water less than 1 m deep in the estuarine coasts ofSan Clemente del Tuyu¤, Argentina. They were transported tothe laboratory, where they were lodged in plastic tanks(30U45U20 cm) ¢lled to 0.5 cm depth with diluted (12%, pH7.4^7.6) marine water (prepared from Cristalsea Marinemixsalts, USA), to a density of 20 crabs per tank. The holdingroom was maintained on a 12-h light^dark cycle (light on07.00^19.00 h). Animals were fed rabbit pellets (NutrientesS.A., Argentina) every 3 days and water was changed afterfeeding. The temperature of both holding and experimentalrooms was maintained within a range of 22^24‡C. Experimentswere carried out within the ¢rst week after the animal’s arrival.Each crab was used in only one experiment.
Training-testing apparatus
The experimental unit, the ‘actometer’, was constructed in thelaboratory and is described in detail elsewhere (Romano et al.,1990). In brief, it consists in a bowl-shaped plastic containerwhere the crab is lodged and an opaque rectangular screenwhich moves horizontally over the animal. Screen displacementsprovoke a crab’s running response and, as a consequence, con-tainer oscillations which induce electrical signals through a pie-zoelectric transducer. Signals recorded during a trial weretranslated into numerical units ranging from 0 to 10 000. Acomputer was employed to program trial sequences, trial dura-tion and ITIs, as well as to monitor experimental events.
Drugs and injection protocol
Sulfasalazine [2,4-hydroxy((4-((2-pyridinylamino)sulfonyl)-phenyl)azo) benzoic acid] (Sigma, St. Louis, MO, USA) solutionwas freshly dissolved in crustacean saline (Hoeger and Florey,1989) with 10 mM HEPES pH 7.6 plus 15% dimethylsulfoxide(DMSO), ¢nal pH 7.6. Indomethacin [1-(4-chlorobenzoyl)-5-methoxy-2-methyl-1H-indole-3-acetic acid] was dissolved at aconcentration of 1 mM or less with crustacean saline with 10mM HEPES pH 7.6 plus 20% DMSO. Fifty microliters ofvehicle or drug solution were given through the right side ofthe dorsal cephalothoracic^abdominal membrane by means of
NSC 5503 22-5-02
E. Merlo et al.162

a syringe ¢tted with a sleeve to control depth of penetration to4 mm, thus ensuring that the injected solution was releasedroughly at the center of the pericardial sac. The total volumeof hemolymph was estimated at 5 ml (30% of the body weight)(Gleeson and Zubko¡, 1977) resulting in an approximate 100-fold dilution of the drug in hemolymph.
Memory evaluation experiments
Each trial lasted 9 s and consisted of two cycles of presenta-tion of the screen over the actometer. Each cycle lasted 2.5 swith a 2-s interval between cycles. The crab’s activity wasrecorded during the entire trial time.
Before the training session the animals underwent a pre-train-ing test (PT), which consisted of one trial. Animals were rankedaccording to their PT score and distributed in the experimentalgroups, ensuring similar mean response values between groups.
Training and testing sessions were separated by a 48-h inter-session interval. Crabs were individually housed during theinter-session interval in plastic containers, covered to a depthof 0.5 cm with water and kept inside dimly lighted drawers. Thetraining session consisted of 15 trials with an ITI of 171 s andthe testing session consisted of one trial. Both sessions werepreceded by 15 min of adaptation in the actometer.
One group of crabs was injected with vehicle (VEH group)and another was given sulfasalazine (SSZ group). Half of theanimals in each group were trained (TR group) while the otherhalf were located in the actometers but remained untrained(control group, CT group). Thus, each experiment consisted offour groups (n= 40 for each one) termed CT-VEH, TR-VEH,CT-SSZ and TR-SSZ. A similar design was used in the twoexperiments in which indomethacin was administered at dosesof 100 WM and 500 WM. In the ¢rst one, CT-VEH and TR-VEHgroups were run together with the trained groups administeredindomethacin (TR-IND100 WM and TR-IND500 WM). In thesecond, CT-VEH group were run together with CT-IND100 WMand CT-IND500 WM groups.
Data analysis and evaluation of amnestic e¡ect
Retention of learning acquired during training was consideredwhen a statistically lower level of response in the testing sessionwas found for the trained group relative to the control groupinjected with the same solution (drug or saline). A comparisonbetween control groups injected with drug or vehicle was neces-sary in order to determine any drug side e¡ects a¡ecting theresponse level at testing in a way not related to training experi-ence.
The statistical analysis of testing data included a set of three apriori planned comparisons, namely CT-VEH vs. TR-VEH, CT-SSZ vs. TR-SSZ and CT-VEH vs. CT-SSZ, using a weightedmeans analysis of variance (ANOVA) with K (per comparisonerror rate) = 0.05 (Howell, 1987; Rosenthal and Rosnow, 1985).In the ¢rst comparison a di¡erence between CT-VEH and TR-VEH groups was expected due to the reduction in response levelinduced by training in the latter group. In contrast, in the sec-ond comparison, if the drug produced an amnestic e¡ect, nodi¡erence was expected between CT-SSZ and TR-SSZ. Finally,as long as the drug does not a¡ect the level of response at test-ing, no di¡erence was expected in the comparison between con-trol groups. In the experiments with indomethacinadministration, CT-VEH was compared with the rest of thegroups in each experiment.
Western blot assay
The animals were anesthetized by immersion in an ice^watermix for 2 min. For each sample 10 central brains (supraesopha-geal ganglion) were dissected, pooled in 1 ml bu¡ered crab sa-line solution (pH 7.6) and then centrifuged for 20 s at 1000Ug,4‡C. For total protein extracts the ganglia were homogenized inRIPA bu¡er (phosphate-bu¡ered saline, PBS, pH 7.4; 1% igepalCA-630; 0.5% sodium deoxycholate; 0.1% sodium dode-
cylsulfate (SDS) and protease inhibitors: pepstatin A, 1 Wg/ml;leupeptin, 10 Wg/ml; aprotinin, 10 Wg/ml and phenylmethylsul-fonyl £uoride, 0.5 mM) with eight strokes of Te£on^glasshomogenizer. The sample was centrifuged at 1000Ug for 5 minat 4‡C and the supernatant stored at 320‡C. Nuclear proteinextracts were obtained as described previously (Freudenthal andRomano, 2000). Thirty micrograms of proteins were electropho-resed in 10% SDS^polyacrylamide gel electrophoresis (PAGE)at 150 V for 1 h and then electroblotted to nitrocellulose mem-brane (Gibco BRL, Grand Island, NY, USA) at 150 V for 1 h.Immunodetection was performed for a polyclonal full-lengthmouse IUB-K antibody (Santa Cruz Biotechnology, SantaCruz, CA, USA; sc-847) and for a polyclonal mouse IKKK
antibody (Santa Cruz Biotechnology sc-7182) at 1/500 dilutionfollowed by anti-rabbit IgG-horseradish peroxidase secondaryantibody (Santa Cruz Biotechnology) at 1/10 000 dilution.Detection was made with a Luminol chemiluminescence kit(Santa Cruz Biotechnology) as described by the manufacturer,exposing the membranes to XAR-5 ¢lms (Kodak). Proteinquantities in the samples were determined by Lowry method.
In vitro IKK-like kinase activity
Ten ganglia of naive or trained animals extracted immediatelyafter training were homogenized in IKK extraction bu¡er(HEPES 10 mM, L-glycerol phosphate 20 mM, p-nitrophenyl-phosphate 10 mM, Na3VO4 100 WM, EDTA 5 mM with pro-tease inhibitors) with eight strokes of a Te£on^glasshomogenizer. The sample was centrifuged at 1000Ug for 5 minat 4‡C and the supernatant stored at 380‡C. The phosphotrans-ferase activity assay was performed according to DiDonato etal. (1997), using the recombinant glutathione S-transferase(GST)-human IUB-K peptide (Santa Cruz Biotechnology) as spe-ci¢c substrate for IKK. Twenty to 25 Wg of extracts were addedto IKK reaction bu¡er (HEPES 10 mM, L-glycerolphosphate 20mM, p-nitrophenylphosphate 10 mM, Na3VO4 100 WM andNaCl 200 mM, pH 7.5), ATP mix (ATP 20 WM, MgCl2 10mM, aprotinin 10 Wg/ml and [Q-32P]ATP, pH 7.5) and 1 Wg ofGST-IUB-K peptide. Fifteen percent of DMSO with or withoutsulfasalazine (5 mM ¢nal concentration) was added to the reac-tion, performed at 30‡C for 30 min. The kinase reaction mix-tures were resolved by SDS^PAGE and the phosphorylation ofGST-IUB-K substrate was detected by autoradiography. The rel-ative optical density (ROD) was measured using the NIH SionImage Beta (4.0.2 version) software. The performance of alltrained groups during the training session was similar to thatdescribed in Fig. 2A. The individual performance was analyzedbefore dissection. Animals with an initial level of response below1000 were not considered.
Immunoprecipitation
Total protein extracts obtained from naive animals (500 Wg)were added to 1 Wg of rabbit IgG (Santa Cruz Biotechnology)and to 20 Wl of protein A-agarose conjugate (Santa Cruz Bio-technology) and then stirred for 30 min at 4‡C. Samples werecentrifuged at 2500 r.p.m. for 5 min (at 4‡C). Five microliters ofIKK-K antibody were added to the supernatant and incubatedfor 1 h at 4‡C. Protein A-agarose was then added (20 Wl) andincubated with stirring on. Samples were then centrifuged (2500r.p.m., 5 min, 4‡C) and pellets were washed four times with 1 mlPBS. The IKK kinase activity was measured as described above.
Electrophoretic mobility shift assay (EMSA)
At di¡erent times after treatment (see Results section) animalswere anesthetized by immersion in ice-cold water for 2 min. Thecentral brain (supraesophageal ganglion) was then dissected.Twenty ganglia per sample were pooled in 1 ml bu¡ered crabsaline solution (pH 7.6). Nuclear extracts were obtained asdescribed previously (Freudenthal and Romano, 2000). Toassess NF-UB activity, double-stranded oligonucleotide DNAcontaining the NF-UB binding site (5P-AGTTGAGGG-
NSC 5503 22-5-02
E¡ect of IUB kinase inhibitor on long-term memory 163

GACTTTCCCAGGC-3P, binding site in bold) (Promega, Mad-ison, WI, USA) was used as probe. The ROD of the bandobtained with this probe was estimated using NIH ScionImage software. All measures were made with exposures withinthe linear range of the ¢lm (Kodak XAR5). Images were digi-tized by means of a scanner for negatives. Protein contents ofthe extracts were measured in triplicate by the Bradford methodand checked for quality and quantity by comparing patternintensities in SDS^PAGE.
RESULTS
IKK and IUB-like immunoreactivity in Chasmagnathuscentral brain extracts
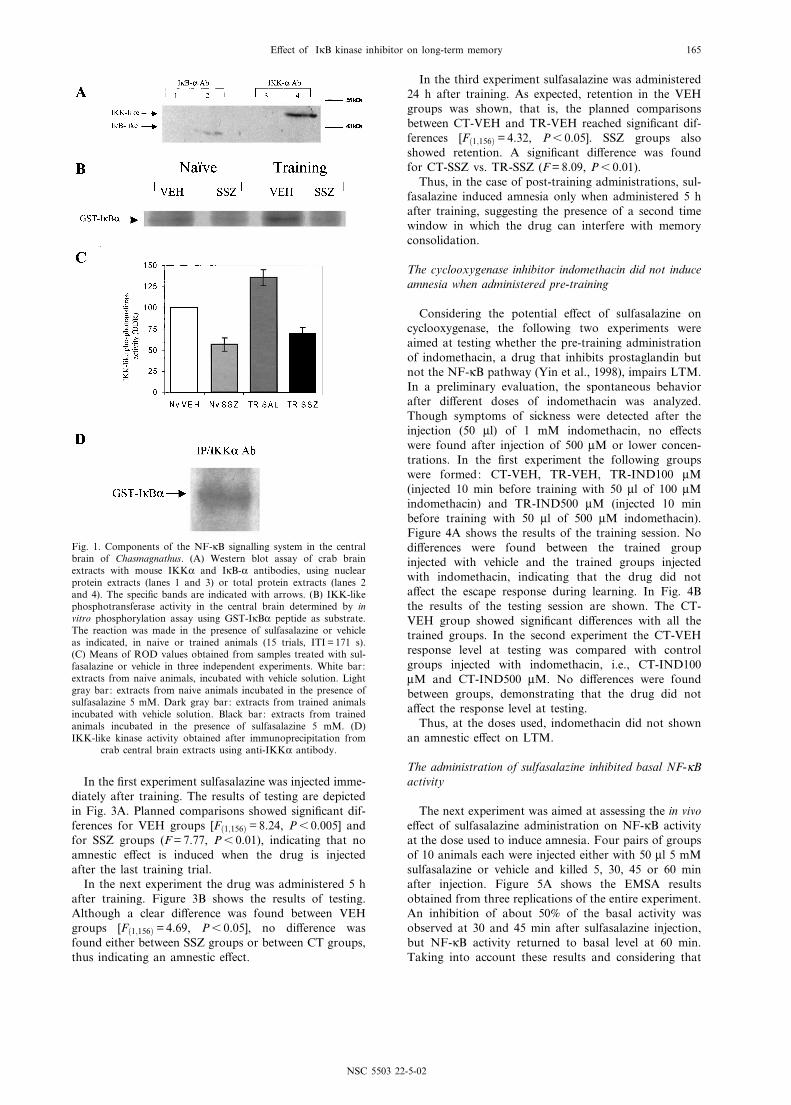
Previous evidence for the presence of NF-UB familymembers in the nervous system of the crab was obtainedwith speci¢c DNA binding activity assays (Freudenthalet al., 1998; Freudenthal and Romano, 2000), westernblot analysis with human p65 antibody and UV-cross-linking analysis of the complex found in nucleus (R.Freudenthal and A. Romano, unpublished data). Theaim of this experiment was to detect components of theNF-UB signalling system in the central brain, i.e., thearea of the CNS in which changes in NF-UB activityare correlated with memory formation. For this purpose,mouse IKKK and IUB-K antibodies were used in westernblots in order to determine cross-reactivity in crab brainextracts. IKK and IUB are normally present in the cyto-plasm but not in the nucleus. For this reason nuclear andtotal protein extracts were compared. Figure 1A shows arepresentative result of the western blot assay performedwith the IKKK and IUB-K antibodies. The antibodiesshowed immunoreactivity in total protein extracts butnot in nuclear extracts. The proteins detected had amolecular weight of 41 kDa for the IUB-like proteinand 48 kDa for IKK-like protein.
IKK-like kinase activity in crab brain is inhibited in vitroby sulfasalazine
The presence of IKK-like immunoreactivity suggestedthe existence of an IKK complex homologue in Chas-magnathus. To obtain further evidence for the functionof this complex we performed in vitro phosphorylationassays using central brain protein extracts incubated witha speci¢c IKK substrate, the GST-IUBK peptide. Figure1B shows a representative autoradiogram in which thepeptide was phosphorylated and the IKK inhibitor sul-fasalazine markedly reduced such phosphorylation inextracts from both naive (NV) and trained (TR) animalsimmediately after training. As presented in Fig. 1C, theROD values for samples treated with (SSZ) or without(VEH) sulfasalazine in four independent replications ofthe experiment showed higher values for the TR-VEHgroup than the NV-VEH group. Besides, sulfasalazineinduced an inhibitory e¡ect of 43.8% for NV and of48.4% for TR groups. Ongoing experiments with appro-priate controls and temporal course analysis are aimed atevaluating the IKK activation during memory consolida-tion. To further con¢rm that IUB-K substrate is phos-phorylated by the crab IKK homologue, the complex
was puri¢ed by immunoprecipitation with the IKKK
antibody. The precipitate conserved IKK-like phospho-transferase activity as determined by IKK kinase assay insamples obtained from naive animals (Fig. 1D).
Pre-training administration of sulfasalazine inducesamnesia
A remarkable correlation between NF-UB activity andLTM formation was previously found in Chasmagnathus(Freudenthal et al., 1998; Freudenthal and Romano,2000). The aim of the following series of experimentswas to evaluate whether the activation of NF-UB is nec-essary for memory consolidation. For this purpose thee¡ect of in vivo administration of the IKK and NF-UBinhibitor sulfasalazine on memory was evaluated.
In preliminary experiments the e¡ect of an injection of50 Wl sulfasalazine at concentrations ranging from 1 mMto 25 mM on spontaneous behavior was analyzed. Re-versible stress symptoms and/or muscular sti¡ness wasfound over 10 mM administration.
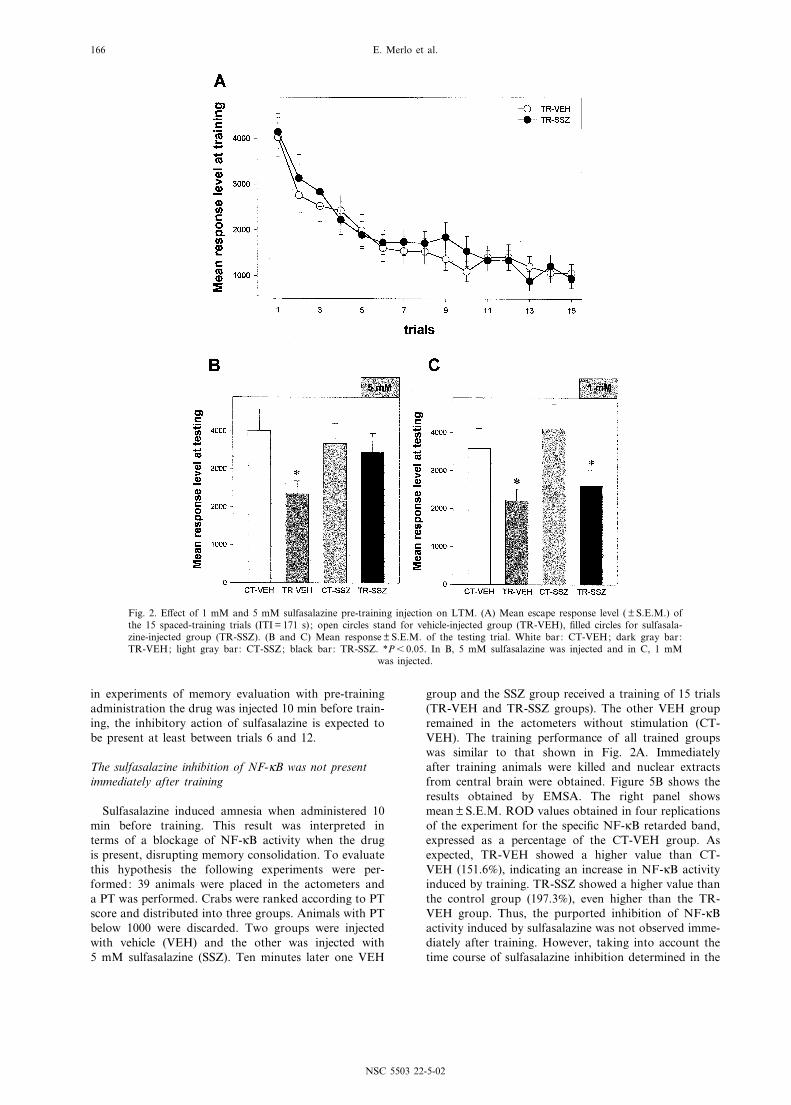
The next two experiments were performed to evalu-ate the pre-training e¡ect of sulfasalazine on long-termretention. Two doses, 1.17 Wg/g (50 Wl of 1 mM) and5.75 Wg/g (50 Wl of 5 mM), were administered 10 minbefore training. No e¡ect on training performance wasfound either for the lower dose (data not shown) or forthe highest dose (Fig. 2A), indicating that the drug didnot a¡ect the escape response of the animals and sug-gesting that it did not a¡ect learning performance. Arepeated measures ANOVA applied on these data yiel-ded no signi¢cant di¡erences.
Figure 2B shows the results of the testing session per-formed 48 h after training when the higher dose wasused. As expected, the TR-VEH group showed a signi¢-cantly lower response compared with the respectivecontrol group, CT-VEH [Fð1;156Þ = 5.7, P6 0.05], dem-onstrating retention at testing. In contrast, a high lev-el of response was found for the TR-SSZ group, notdi¡erent from that observed in the CT-SSZ group.Both CT groups also showed a similar level of response,indicating that the drug treatment alone did not a¡ectthe level of response at testing. In contrast, when thelower dose was used, both CT group vs. TR group com-parisons were signi¢cantly di¡erent [Fð1;156Þ = 4.0,P6 0.05 for VEH group comparison and F= 4.78,P6 0.05 for SSZ group comparison] (Fig. 2C).
Thus, an amnestic e¡ect of sulfasalazine was foundwhen used at 5 mM but not at 1 mM concentration,suggesting that the drug a¡ects retention in a dose-de-pendent manner.
Time course of the amnestic e¡ect of sulfasalazine whenadministered after training
The next three experiments were aimed at determiningwhether the amnestic e¡ect of the higher dose of sulfa-salazine is present when administered immediately aftertraining or after a delay between training and injectiontime with the purpose of assessing the e¡ect of the drugat di¡erent times during LTM consolidation.
NSC 5503 22-5-02
E. Merlo et al.164
In the ¢rst experiment sulfasalazine was injected imme-diately after training. The results of testing are depictedin Fig. 3A. Planned comparisons showed signi¢cant dif-ferences for VEH groups [Fð1;156Þ = 8.24, P6 0.005] andfor SSZ groups (F= 7.77, P6 0.01), indicating that noamnestic e¡ect is induced when the drug is injectedafter the last training trial.
In the next experiment the drug was administered 5 hafter training. Figure 3B shows the results of testing.Although a clear di¡erence was found between VEHgroups [Fð1;156Þ = 4.69, P6 0.05], no di¡erence wasfound either between SSZ groups or between CT groups,thus indicating an amnestic e¡ect.
In the third experiment sulfasalazine was administered24 h after training. As expected, retention in the VEHgroups was shown, that is, the planned comparisonsbetween CT-VEH and TR-VEH reached signi¢cant dif-ferences [Fð1;156Þ = 4.32, P6 0.05]. SSZ groups alsoshowed retention. A signi¢cant di¡erence was foundfor CT-SSZ vs. TR-SSZ (F= 8.09, P6 0.01).
Thus, in the case of post-training administrations, sul-fasalazine induced amnesia only when administered 5 hafter training, suggesting the presence of a second timewindow in which the drug can interfere with memoryconsolidation.
The cyclooxygenase inhibitor indomethacin did not induceamnesia when administered pre-training
Considering the potential e¡ect of sulfasalazine oncyclooxygenase, the following two experiments wereaimed at testing whether the pre-training administrationof indomethacin, a drug that inhibits prostaglandin butnot the NF-UB pathway (Yin et al., 1998), impairs LTM.In a preliminary evaluation, the spontaneous behaviorafter di¡erent doses of indomethacin was analyzed.Though symptoms of sickness were detected after theinjection (50 Wl) of 1 mM indomethacin, no e¡ectswere found after injection of 500 WM or lower concen-trations. In the ¢rst experiment the following groupswere formed: CT-VEH, TR-VEH, TR-IND100 WM(injected 10 min before training with 50 Wl of 100 WMindomethacin) and TR-IND500 WM (injected 10 minbefore training with 50 Wl of 500 WM indomethacin).Figure 4A shows the results of the training session. Nodi¡erences were found between the trained groupinjected with vehicle and the trained groups injectedwith indomethacin, indicating that the drug did nota¡ect the escape response during learning. In Fig. 4Bthe results of the testing session are shown. The CT-VEH group showed signi¢cant di¡erences with all thetrained groups. In the second experiment the CT-VEHresponse level at testing was compared with controlgroups injected with indomethacin, i.e., CT-IND100WM and CT-IND500 WM. No di¡erences were foundbetween groups, demonstrating that the drug did nota¡ect the response level at testing.
Thus, at the doses used, indomethacin did not shownan amnestic e¡ect on LTM.
The administration of sulfasalazine inhibited basal NF-UBactivity
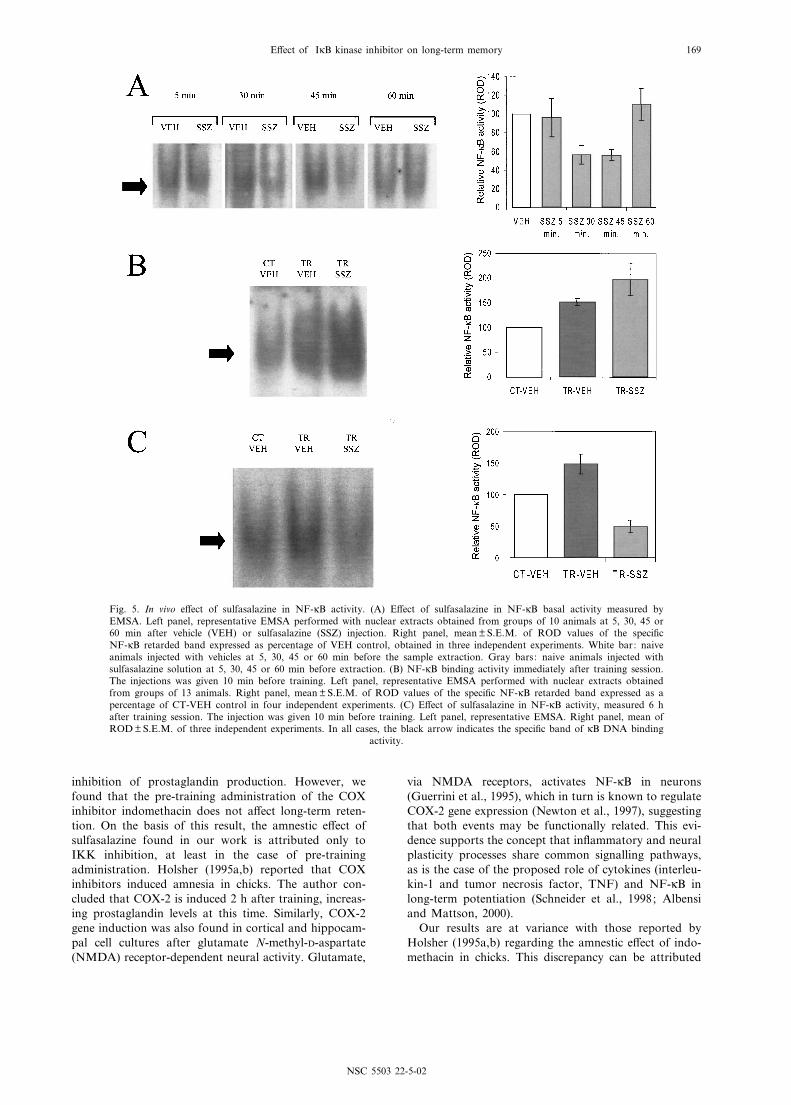
The next experiment was aimed at assessing the in vivoe¡ect of sulfasalazine administration on NF-UB activityat the dose used to induce amnesia. Four pairs of groupsof 10 animals each were injected either with 50 Wl 5 mMsulfasalazine or vehicle and killed 5, 30, 45 or 60 minafter injection. Figure 5A shows the EMSA resultsobtained from three replications of the entire experiment.An inhibition of about 50% of the basal activity wasobserved at 30 and 45 min after sulfasalazine injection,but NF-UB activity returned to basal level at 60 min.Taking into account these results and considering that
Fig. 1. Components of the NF-UB signalling system in the centralbrain of Chasmagnathus. (A) Western blot assay of crab brainextracts with mouse IKKK and IUB-K antibodies, using nuclearprotein extracts (lanes 1 and 3) or total protein extracts (lanes 2and 4). The speci¢c bands are indicated with arrows. (B) IKK-likephosphotransferase activity in the central brain determined by invitro phosphorylation assay using GST-IUBK peptide as substrate.The reaction was made in the presence of sulfasalazine or vehicleas indicated, in naive or trained animals (15 trials, ITI = 171 s).(C) Means of ROD values obtained from samples treated with sul-fasalazine or vehicle in three independent experiments. White bar:extracts from naive animals, incubated with vehicle solution. Lightgray bar: extracts from naive animals incubated in the presence ofsulfasalazine 5 mM. Dark gray bar: extracts from trained animalsincubated with vehicle solution. Black bar: extracts from trainedanimals incubated in the presence of sulfasalazine 5 mM. (D)IKK-like kinase activity obtained after immunoprecipitation from
crab central brain extracts using anti-IKKK antibody.
NSC 5503 22-5-02
E¡ect of IUB kinase inhibitor on long-term memory 165
in experiments of memory evaluation with pre-trainingadministration the drug was injected 10 min before train-ing, the inhibitory action of sulfasalazine is expected tobe present at least between trials 6 and 12.
The sulfasalazine inhibition of NF-UB was not presentimmediately after training
Sulfasalazine induced amnesia when administered 10min before training. This result was interpreted interms of a blockage of NF-UB activity when the drugis present, disrupting memory consolidation. To evaluatethis hypothesis the following experiments were per-formed: 39 animals were placed in the actometers anda PT was performed. Crabs were ranked according to PTscore and distributed into three groups. Animals with PTbelow 1000 were discarded. Two groups were injectedwith vehicle (VEH) and the other was injected with5 mM sulfasalazine (SSZ). Ten minutes later one VEH
group and the SSZ group received a training of 15 trials(TR-VEH and TR-SSZ groups). The other VEH groupremained in the actometers without stimulation (CT-VEH). The training performance of all trained groupswas similar to that shown in Fig. 2A. Immediatelyafter training animals were killed and nuclear extractsfrom central brain were obtained. Figure 5B shows theresults obtained by EMSA. The right panel showsmean Q S.E.M. ROD values obtained in four replicationsof the experiment for the speci¢c NF-UB retarded band,expressed as a percentage of the CT-VEH group. Asexpected, TR-VEH showed a higher value than CT-VEH (151.6%), indicating an increase in NF-UB activityinduced by training. TR-SSZ showed a higher value thanthe control group (197.3%), even higher than the TR-VEH group. Thus, the purported inhibition of NF-UBactivity induced by sulfasalazine was not observed imme-diately after training. However, taking into account thetime course of sulfasalazine inhibition determined in the
Fig. 2. E¡ect of 1 mM and 5 mM sulfasalazine pre-training injection on LTM. (A) Mean escape response level ( Q S.E.M.) ofthe 15 spaced-training trials (ITI = 171 s); open circles stand for vehicle-injected group (TR-VEH), ¢lled circles for sulfasala-zine-injected group (TR-SSZ). (B and C) Mean response Q S.E.M. of the testing trial. White bar: CT-VEH; dark gray bar:TR-VEH; light gray bar: CT-SSZ; black bar: TR-SSZ. *P6 0.05. In B, 5 mM sulfasalazine was injected and in C, 1 mM
was injected.
NSC 5503 22-5-02
E. Merlo et al.166

previous section, the e¡ect of the drug is expected todisappear at the end of training. This fact could explainthe lack of e¡ect of sulfasalazine in this experiment inwhich a rebound after inhibition could be present con-sidering that IKK upstream signals could be still acti-vated.
Pre-training administration of sulfasalazine inhibitedNF-UB 6 h after training
In a previous report we found two phases of NF-UBactivation after training. The ¢rst one takes place after10 or more trials but drops to basal level 3 h after train-ing. A second wave of Rel/NF-UB activity, as high as the¢rst, is observed about 6 h after training. This secondwave of activity decreased gradually reaching basal level24 h after training. In order to evaluate if the pre-train-ing administration of sulfasalazine interferes with NF-UBactivation during this second phase we performed thefollowing experiment. The design was equal to the pre-vious experiment but animals were individually housedafter training and nuclear extracts from brain wereobtained 6 h later. Results are depicted in Fig. 5C. Inthe right panel the mean Q S.E.M. ROD values obtainedin three replications of the experiment for the speci¢cNF-UB retarded band are shown, expressed as a percent-age of the CT-VEH group. The TR-saline (SAL) groupshowed a higher value than the CT-VEH group(148.3%), according to the second phase of training-de-pendent NF-UB activation. In contrast, TR-SSZ showeda lower value relative to the TR-SAL group and evenlower than CT-VEH (48.8%). Thus, an inhibitory e¡ectof sulfasalazine on NF-UB activity was found at 6 h aftertraining.
DISCUSSION
On the basis of previous results we predicted thatNF-UB activation was necessary for LTM formation in
the context-signal learning paradigm of Chasmagnathus.In this work, experimental data supporting this predic-tion are reported.
The ¢rst series of experiments indicates that compo-nents of the NF-UB pathway are present in crustaceans.IUB- and IKK-like immunoreactivity, as well as IKK-likekinase activity were detected in the central brain. Thisactivity was inhibited by sulfasalazine, a drug previouslyfound to inhibit IKK and NF-UB in mammals. TheIKK-like kinase activity in trained animals was higherthan in naive controls.
In the second group of experiments, two periods werefound in which sulfasalazine was e¡ective at inducingamnesia, the ¢rst during training and the second 5 hafter training. These results are interpreted consideringthe existence of two periods in which IKK and NF-UBactivation are necessary for memory consolidation.Accordingly, two phases of NF-UB activity induced byspaced training were previously found in this paradigm(Freudenthal and Romano, 2000). The time courses oftraining-induced NF-UB activity and of sulfasalazine-induced amnesia are compared in Fig. 6. The periodsin which the drug is e¡ective at producing amnesia arecoincident with the time windows when NF-UB activityin nuclei is upsurging, namely, during training andbetween 3 and 6 h after training. Such coincidence isexpected considering that IKK activation should be pre-vious to NF-UB nuclear translocation. On the otherhand, the drug is not e¡ective when NF-UB is alreadyactivated (0 h) or when it returns to basal activity (24 h).
The third series of experiments were aimed at con¢rm-ing this hypothesis, analyzing the in vivo e¡ect of thedrug on NF-UB activity in the brain. The temporalcourse of the e¡ect of sulfasalazine administration(Fig. 5A) indicates that the drug e¡ectively inhibitbasal NF-UB activity. The inhibition was present at 45min but vanished 60 min after injection. However, imme-diately after 15 trials (55 min after injection) NF-UBactivity was even higher in animals injected with sulfasa-lazine than in trained animals injected with vehicle. This
Fig. 3. E¡ect of 5 mM sulfasalazine injected at di¡erent times after training on LTM. Mean response level Q S.E.M. at test-ing. Sulfasalazine was injected immediately (A), 5 h (B) or 24 h (C) after the last training trial. Symbols as in Fig. 2.
*P6 0.05; **P6 0.01.
NSC 5503 22-5-02
E¡ect of IUB kinase inhibitor on long-term memory 167

increment, which was not found when the system wasinactive, may be interpreted as a rebound after an arrestof the NF-UB activation mechanism. Anyway, theamnestic e¡ect of the drug could be a consequence ofan interference of this signalling pathway either by alack of NF-UB pathway activation during training, anunspeci¢c over-activation after training or both e¡ects.A clear inhibition of NF-UB by sulfasalazine was foundwhen the second phase of activation was analyzed, sug-gesting that this activation, several hours after training,plays a key role in LTM consolidation. A two-phaseinduction of Rel/NF-UB activity was reported in othersystems. The ¢rst transient phase is attributed to a rapidIKK activation and IUB phosphorylation in response totransducing signals that allows translocation of the pre-formed NF-UB to the nucleus. The second long-lastingphase is considered to be dependent on NF-UB geneinduction. An alternative interpretation for the secondphase is that activated NF-UB forms initially present insynaptic terminals reach the nucleus several hours later.
Such synaptic NF-UB activation was found in Chasmag-nathus after training (Freudenthal and Romano, 2000).However, the amnestic e¡ect of sulfasalazine at 5 h aftertraining suggests that IKK activity during this period isrequired for memory consolidation. Previous studies inother memory models showed two time windows forprotein kinase activation and protein synthesis, oneimmediately and other from 4 to 6 h after training(Bernabeu et al., 1997; Bourtchuladze et al., 1998;Freeman et al., 1995). In Chasmagnathus, cAMP-depen-dent kinase activity is enhanced 6 h after training(F. Locatelli, H. Maldonado and A. Romano, unpub-lished data) and is required for LTM formation(Locatelli et al., 2001). The memory consolidation pro-cess appears to have two periods in which mechanisms ofneural plasticity are required.
Although controversial, some evidence suggests ane¡ect of sulfasalazine on COX activity in vertebrates.Therefore, the amnestic e¡ect of sulfasalazine found inthis report could be attributed, at least in part, to the
Fig. 4. E¡ect of the COX inhibitor indomethacin on LTM. (A) Mean escape response level ( Q S.E.M.) of the 15 spacedtraining trials (ITI = 171 s); ¢lled circles stand for vehicle-injected group (TR-VEH), open circles for the group injected withindomethacin 100 WM (TR-IND100 WM), ¢lled triangles for the group injected with indomethacin 500 WM (TR-IND500 WM).
(B and C) Mean response Q S.E.M. of the testing trial. *P6 0.05; **P6 0.01.
NSC 5503 22-5-02
E. Merlo et al.168
inhibition of prostaglandin production. However, wefound that the pre-training administration of the COXinhibitor indomethacin does not a¡ect long-term reten-tion. On the basis of this result, the amnestic e¡ect ofsulfasalazine found in our work is attributed only toIKK inhibition, at least in the case of pre-trainingadministration. Holsher (1995a,b) reported that COXinhibitors induced amnesia in chicks. The author con-cluded that COX-2 is induced 2 h after training, increas-ing prostaglandin levels at this time. Similarly, COX-2gene induction was also found in cortical and hippocam-pal cell cultures after glutamate N-methyl-D-aspartate(NMDA) receptor-dependent neural activity. Glutamate,
via NMDA receptors, activates NF-UB in neurons(Guerrini et al., 1995), which in turn is known to regulateCOX-2 gene expression (Newton et al., 1997), suggestingthat both events may be functionally related. This evi-dence supports the concept that in£ammatory and neuralplasticity processes share common signalling pathways,as is the case of the proposed role of cytokines (interleu-kin-1 and tumor necrosis factor, TNF) and NF-UB inlong-term potentiation (Schneider et al., 1998; Albensiand Mattson, 2000).
Our results are at variance with those reported byHolsher (1995a,b) regarding the amnestic e¡ect of indo-methacin in chicks. This discrepancy can be attributed
Fig. 5. In vivo e¡ect of sulfasalazine in NF-UB activity. (A) E¡ect of sulfasalazine in NF-UB basal activity measured byEMSA. Left panel, representative EMSA performed with nuclear extracts obtained from groups of 10 animals at 5, 30, 45 or60 min after vehicle (VEH) or sulfasalazine (SSZ) injection. Right panel, mean Q S.E.M. of ROD values of the speci¢cNF-UB retarded band expressed as percentage of VEH control, obtained in three independent experiments. White bar: naiveanimals injected with vehicles at 5, 30, 45 or 60 min before the sample extraction. Gray bars: naive animals injected withsulfasalazine solution at 5, 30, 45 or 60 min before extraction. (B) NF-UB binding activity immediately after training session.The injections was given 10 min before training. Left panel, representative EMSA performed with nuclear extracts obtainedfrom groups of 13 animals. Right panel, mean Q S.E.M. of ROD values of the speci¢c NF-UB retarded band expressed as apercentage of CT-VEH control in four independent experiments. (C) E¡ect of sulfasalazine in NF-UB activity, measured 6 hafter training session. The injection was given 10 min before training. Left panel, representative EMSA. Right panel, mean ofROD Q S.E.M. of three independent experiments. In all cases, the black arrow indicates the speci¢c band of UB DNA binding
activity.
NSC 5503 22-5-02
E¡ect of IUB kinase inhibitor on long-term memory 169

either to the fact that di¡erent animals were used, to thedi¡erent method of drug administration used (systemicvs. intracranial injections) or to the di¡erent time of test-ing (48 h vs. 2 h after training) employed. Holsher foundamnesia, in coincidence with an inhibition of prostaglan-dins, not before 2 h after training. Experiments in prog-ress in our laboratory are aimed at evaluating whetherindomethacin has an amnestic e¡ect when administeredat di¡erent times after training and when the training^testing interval is shortened.
Sulfasalazine was e¡ective when administered beforebut not immediately after training. This fact does notimply, however, that the drug a¡ected learning perfor-mance. Data shown in Fig. 2A support that the drug, infact, did not interfere with learning.
The ¢rst IKK activity period seems to be only neces-sary for memory consolidation in a narrow time window
con¢ned to training session because post-training admin-istration was not e¡ective. A similar result was obtainedwith the protein kinase A inhibitor, Rp-8-Cl-cAMPS,which induced amnesia when administered pre-trainingor after a shortened training of 12 trials, but not after theusual 15 trials training (Locatelli et al., 2001). In the crabmodel a turning point between 10 and 15 trials wasfound. Ten trials are not enough to induce long-termretention, while 15 trials yield a good retention (Pedreiraet al., 1998). Accordingly, NF-UB is activated after 15 ormore trials but not after 10 or fewer trials (Freudenthaland Romano, 2000).
This work, together with previous reports, supportsthe participation of the UB-dependent transcriptionalregulation system in memory. Two main questionsremain to be clari¢ed: (1) which are the memory-relatedsignals that activate this pathway? and (2) which are the
Fig. 6. Comparison between the time courses of training-induced NF-UB activity and sulfasalazine-induced amnesia. (A) NF-UB activity for training groups during training and at di¡erent times after training determined in a previous work (resultsredrawn from Freudenthal and Romano, 2000). UB DNA binding activity was determined by EMSA. Mean ROD values ofthe speci¢c retarded band from spaced training groups relative to the values obtained from the corresponding control group.Dotted line: UB binding activity in the control groups. (B) E¡ect of sulfasalazine on LTM. The abscissa represents the timeof injection with respect to the end of training. Values were normalized to the respective control group (CT-SSZ). Opencircles : mean Q S.E.M. relative response level at testing of CT-SSZ groups. Gray bars: mean Q S.E.M. relative response level
at testing of TR-SSZ groups. Hatched rectangle: training session. *P6 0.05. **P6 0.01.
NSC 5503 22-5-02
E. Merlo et al.170

memory-related genes to be regulated by this pathway?Among neurotransmitters that are known to activateNF-UB, glutamate through NMDA receptors and angio-tensin II play key roles in memory and particularly in theChasmagnathus model (Delorenzi and Maldonado, 1999;Troncoso and Maldonado, personal communication).Other signals like nerve growth factor, TNF and neu-ral cell adhesion molecule have been proposed to beinvolved in memory and are also known to activatethis pathway. Among several genes that are regulatedby the UB consensus sequences, angiotensinogen, proen-
kephalin and cytokine genes are particularly interestingcandidates to participate in the processes that subservememory formation.
Acknowledgements1We thank Dr. H. Maldonado for his sup-port and critical reading of the manuscript and Dr. L. Orelli andDr. G. Hermitte for their assistance. This work was supportedby the following grants: Third World Academy of Science 98-241 RG/BIO/LA, Fundacio¤n Antorchas A-13622/1 and AgenciaNacional de Promocio¤n Cient|¤¢ca y Tecnolo¤gica ^ ArgentinaPICT 1-6041.
REFERENCES
Agrano¡, B.W., 1972. The Chemistry of Mood Motivation and Memory. Plenum, New York.Albensi, B.C., Mattson, M.P., 2000. Evidence for the involvement of TNF and NF-kappaB in hippocampal synaptic plasticity. Synapse 35,
151^159.Alberini, C.M., Ghirardi, M., Metz, R., Kandel, E.R., 1994. C/EBP is an immediate-early gene required for the consolidation of long-term
facilitation in Aplysia. Cell 76, 1099^1114.Barondes, S.H., 1975. Protein synthesis-dependent and protein synthesis-independent memory storage processes. In: Deutsch, D., Deutsch, J.A.
(Eds.), Short-Term Memory. Academic, New York.Bernabeu, R., Bevilaqua, L., Ardenghi, P., Bromberg, E., Schmitz, P., Bianchin, M., Izquierdo, I., Medina, J., 1997. Involvement of hippocampal
cAMP/cAMP-dependent protein kinase signaling pathways in a late memory consolidation phase of aversively motivated learning in rats. Proc.Natl. Acad. Sci. USA 94, 7041^7046.
Bourtchuladze, R., Abel, T., Berman, N., Gordon, R., Lapidus, K., Kandel, E.R., 1998. Di¡erent training procedures recruit either one or twocritical periods for contextual memory consolidation, each of which requires protein synthesis and PKA. Learn. Mem. 5, 365^374.
Choi, J.-S., Kim, J., Kim, D.-H., Chun, M.-H., Gwag, B.-J., Yoon, S.K., Joo, C.-K., 2000. Failure to activate NF-kappaB promotes apoptosis ofretinal ganglion cells following optic nerve transection. Brain Res. 883, 60^68.
Crow, T., Forrester, J., 1990. Inhibition of protein synthesis blocks long-term enhancement of generator potentials produced by one-trial in vivoconditioning in Hermissenda. Proc. Natl. Acad. Sci. USA 87, 4490^4494.
Davis, H.P., Squire, L.R., 1984. Protein synthesis and memory: a review. Psychol. Bull. 96, 518^559.Deisseroth, K., Bito, H., Tsien, R.W., 1996. Signaling from synapse to nucleus: postsynaptic CREB phosphorylation during multiple forms of
hippocampal synaptic plasticity. Neuron 16, 89^101.Delorenzi, A., Maldonado, H., 1999. Memory enhancement by the angiotensinergic system in the crab Chasmagnathus is mediated by endogenous
angiotensin II. Neurosci. Lett. 266, 1^4.DiDonato, J.A., Hayakawa, M., Rothwarf, D.M., Zandi, E., Karin, M., 1997. A cytokine-responsive IUB kinase that activates the transcription
factor NF-UB. Nature 388, 548^554.DiDonato, J.A., Mercurio, F., Rosette, C., Wu-li, J., Suyang, H., Ghosh, S., Karin, M., 1996. Mapping of the inducible IUB phosphorylation sites
that signal its ubiquitination and degradation. Mol. Cell. Biol. 16, 1295^1304.Escoubas, J.M., Briant, L., Montagnani, C., Hez, S., Devaux, C., Roch, P., 1999. Oyster IKK-like protein shares structural and functional
properties with its mammalian homologues. FEBS Lett. 453, 293^298.Freeman, F.M., Rose, S.P.R., Scholey, A.B., 1995. Two time windows of anisomycin-induced amnesia for passive avoidance training in the day-
old chick. Neurobiol. Learn. Mem. 63, 291^295.Freudenthal, R., Locatelli, F., Hermitte, G., Maldonado, H., Delorenzi, A., Romano, A., 1998. Kappa-B like DNA binding activity is enhanced
after spaced training that induces long-term memory in the crab Chasmagnathus. Neurosci. Lett. 242, 143^146.Freudenthal, R., Romano, A., 2000. Participation of Rel/NF-UB transcription factor in long-term memory in the crab Chasmagnathus. Brain Res.
855, 274^281.Gleeson, R.A., Zubko¡, P.L., 1977. The determination of hemolymph volume in the blue crab, Callinectes sapidus, utilizing 14C-thiocyanate.
Comp. Biochem. Physiol. 56A, 411^413.Grilli, M., Pizzi, M., Memo, M., Spano, P., 1996. Neuroprotection by aspirin and sodium salicylate through blockade of NF-kappaB activation.
Science 274, 1383^1385.Guerrini, L., Blasi, F., Denis-Donini, S., 1995. Synaptic activation of NF-kappa B by glutamate in cerebellar granule neurons in vitro. Proc. Natl.
Acad. Sci. USA 92, 9077^9081.Hermitte, G., Pedreira, M.E., Tomsic, D., Maldonado, H., 1999. Context shift and protein synthesis inhibition disrupt long-term habituation after
spaced, but not massed, training in the crab Chasmagnathus. Neurobiol. Learn. Mem. 71, 34^49.Hoeger, U., Florey, E., 1989. Cathecholamine degradation in the hemolimph of the Chinese crab Eriocher sinensis. Comp. Biochem. Physiol. 92,
323^327.Holsher, C., 1995a. Inhibitors of cyclooxigenases produce amnesia for a passive avoidance task in the chick. Eur. J. Neurosci. 7, 1360^1365.Holsher, C., 1995b. Prostaglandins play a role in memory consolidation in the chick. Eur. J. Pharmacol. 294, 253^259.Hoult, J.R., 1986. Pharmacological and biochemical actions of sulphasalazine. Drugs 32, 18^26.Howell, D.C., 1987. Statisticals Methods of Psychology, 2nd edn. Duxbury, Boston, MA.Israel, A., 2000. The IKK complex: an integrator of all signals that activate NF-kappaB? Trends Cell Biol. 10, 129^133.Kaang, B.K., Kandel, E., Grant, S.G.N., 1993. Activation of cAMP-responsive genes by stimuli that produce long-term facilitation in Aplysia
sensory neurons. Neuron 10, 427^435.Kaltschmidt, C., Kaltschmidt, B., Bauerle, P.A., 1993. Brain synapses contain inducible forms of the transcription factor NF-kappa B. Mech. Dev.
43, 135^147.Koop, E.B., Ghosh, S., 1994. Inhibition of NF-kappa B by sodium salicylate and aspirin. Science 265, 956^959.Locatelli, F., Maldonado, H., Romano, A., 2002. Two critical periods for cAMP-dependent protein kinase activity during long-term memory
consolidation in the crab Chasmagnathus. Neurobiol. Learn. Mem. 77, 234^249.
NSC 5503 22-5-02
E¡ect of IUB kinase inhibitor on long-term memory 171

Lozada, M., Romano, A., Maldonado, H., 1990. Long-term habituation to a danger stimulus in the crab Chasmagnathus. Physiol. Behav. 47, 35^41.
Meberg, P.J., Kinney, W.R., Valcourt, E.G., Routtenberg, A., 1996. Gene expression of the transcription factor NF-B in hippocampus: regulationby synaptic activity. Mol. Brain Res. 38, 179^190.
Mercurio, F., Zhu, H., Murray, B.W., Shevchenko, A., Bennet, B.L., Li, J., Young, D.B., Barbosa, M., Mann, M., Manning, A., Rao, A., 1997.IKK-1 and IKK-2-cytokine-activated IUB kinases essential for NF-UB activation. Science 278, 860^866.
Newton, R., Kuitert, L.M.E., Bergman, M., Adcock, I.M., Barnes, P., 1997. Evidence for the involvement of NF-kappaB in the transcriptionalcontrol of COX-2 gene expression by IL-1beta. Biochem. Biophys. Res. Commun. 237, 28^32.
Pedreira, M.E., Dimant, B., Tomsic, D., Quesada-Allue, L.A., Maldonado, H., 1995. Cycloheximide inhibits long-term habituation and contextmemory in the crab Chasmagnathus. Pharmacol. Biochem. Behav. 52, 385^395.
Pedreira, M.E., Romano, A., Tomsic, D., Lozada, M., Maldonado, H., 1998. Massed and spaced training build up di¡erent components of long-term habituation in the crab Chasmagnathus. Animal Learn. Behav. 26, 34^45.
Romano, A., Lozada, M., Maldonado, H., 1990. E¡ects of naloxone pretreatment on habituation in the crab Chasmagnathus granulatus. Behav.Neur. Biol. 53, 113^122.
Rosenthal, R., Rosnow, R.L., 1985. Contrast Analysis Focused Comparisons in the Analysis of Variance. Cambridge University Press, Cam-bridge.
Rothwarf, D.M., Zandi, E., Natoli, G., Karin, M., 1998. IKK-Q is an essential regulatory subunit of the IUB kinase complex. Nature 395, 297^300.Schneider, H., Pitossi, F., Balshun, D., Wagner, A., Del Rey, A., Besedovsky, H.O., 1998. A neuromodulatory role of interleukin-1beta in the
hippocampus. Proc. Natl. Acad. Sci. USA 95, 7778^7783.Tomsic, D., Pedreira, E.M., Romano, A., Hermitte, G., Maldonado, H., 1998. Association to context as a determinant of long-term habituation in
the crab Chasmagnathus. Animal Learn. Behav. 26, 196^209.Wahl, C., Liptay, S., Adler, G., Schmid, R., 1998. Sulfasalazine: a potent and speci¢c inhibitor of nuclear factor kappa B. J. Clin. Invest. 101,
1163^1174.Weber, C.K., Liptay, S., Wirth, T., Adler, G., Schmid, R.M., 2000. Suppression of NF-kappaB activity by sulfasalazine is mediated by direct
inhibition of IkappaB kinases alpha and beta. Gastroenterology 119, 1209^1218.Wellmann, H., Kaltschmidt, B., Kaltschmidt, C., 2000. Retrograde transport of transcription factor NF-kappaB in living neurons. J. Biol. Chem.
276, 11821^11829.Yamoaka, S., Courtois, G., Bessia, C., Whiteside, S.T., Weil, R., Agou, F., Kirk, H.E., Kay, R.J., Israel, A., 1998. Complementation cloning of
NEMO, a component of the IUB kinase complex essential for NF-UB activation. Cell 93, 1231^1240.Yin, J.C.P., Del Vecchio, M., Zhou, H., Tully, T., 1995. CREB as a memory modulator: induced expression of a dCREB2 activator isoform
enhances long-term memory in Drosophila. Cell 81, 107^115.Yin, M.-J., Yamamoto, Y., Gaynor, R.B., 1998. The anti-in£ammatory agents aspirin and salicylate inhibit the activity of IkappaB kinase-beta.
Nature 396, 77^80.Zandi, E., Rothwarf, D.M., Delhase, M., Hayakawa, M., Karin, M., 1997. The IUB kinase complex (IKK) contains two kinase subunits, IKK-K
and IKK-L, necessary for IUB phosphorylation and NF-UB activation. Cell 91, 243^252.
(Accepted 21 December 2001)
NSC 5503 22-5-02
E. Merlo et al.172




















