
Larval development of the hermit crab Clibanarius albidigitus (Crustacea: Anomura: Diogenidae)...
11
Marine Biology 116, 603-613 (1993) Marine ............... Biology (~ Springer-Verlag 1993 Larval development of the hermit crab Clibanarius albidigitus (Crustacea: Anomura: Diogenidae) reared under laboratory conditions E A. Siddiqui 1, p. A. McLaughlin 2, j. A. Crain 2 1 Marine Reference Collection Centre, University of Karachi, Karachi 75270, Pakistan 2 Shannon Point Marine Center, 1900 Shannon Point Road, Anacortes, Washington 98221-4042, USA Received: 9 March 1993 / Accepted: 7 April 1993 Abstract. The larval development of Clibanarius albidigi- tus Nobili is described and illustrated from laboratory- reared specimens. At 30 ~ this species passes through four zoeal stages before molting to the megalopa. Of the 120 individual larvae reared, survival was high, with 88 % reaching the megalopal stage. Zoeal stage durations varied from 3 to 18 d. Rearing was terminated after 45 d, and at that time no megalopae had molted to the first- crab stage. Among known larvae of CIibanarius species, C. albidigitus is immediately distinguished by the pres- ence of dorsomedial and dorsolateral carapace keels. Introduction The usefulness of decapod larval characters in assessing phylogenetic relationships has received considerably sup- port from carcinologists (e.g. Williamson 1976, Rice 1980, 1981, Gore and Scotto 1983, Martin 1984, Gore 1985). In the Brachyura, larval characters have provided the basis for hypotheses of phylogenetic relationships at the generic and familial levels (e.g. Rice and Ingle 1975, Scotto and Gore 1981, Pohle 1989). In contrast to the wealth of information available on brachyuran larvae, complete descriptions of developmental sequences in her- mit crabs are available for relatively few species in most genera. Clibanarius is one of the most specious genera of the Diogenidae, with more than 58 species described (cf. Fize and Ser6ne 1955), and from the standpoint of our knowledge of diogenid larval deVelopment, also the best known. The description of the larval development of C. albidigitus Nobili brings to 11 the number of Clibanarius species for which larval sequences have been described. It is also the first of the eastern Pacific species of the genus to be reared in the laboratory. Brossi-Garcia (1987) summarized the principal mor- phological characters of eight species of Clibanarius, Correspondence to: P. A. McLaughlin omitting only C. longitarsus (DeHaan) and C. olivaceus Henderson described by Ajmal Khan and Natarajan (1981 a, b). A perusal of her data shows a generally con- servative developmental pattern within the genus, al- though Siddiqui et al. (1991) have reported appreciable intraspecific variations among populations of the western Atlantic C. antillensis Stimpson. Only in the development of carapace keels do the zoeae of C. albidigitus differ significantly from other known CIibanarius larvae. Materials and methods Five ovigerous females of Clibanarius albidigitus Nobili were col- lected intertidally on 21 December 1990 at Culebra Island on the Pacific coast of Panam~ by one of us (JAC). The collection site consisted of rocks, cobble and areas of sand. In addition to C. albidigitus, the area was populated by Calcinus explorator Boone and an unidentified species of Paguristes. The semiterrestrial Coeno- bita compressus H. Milne Edwards was found occasionally among the rocks. The five females of Cilbanariusalbidigitus were placed in aerated local sea water and returned to the laboratory at the Smithsonian Tropical Research Institute (STRI). Sea water temperatures at STRI ranged from 29 to 31 ~ On 24 December, the crabs were transferred to plastic "Whirl-Pak ''| bags, each containing ~ 150 ml of local sea water, enclosed in second "Ziploc''| plastic bags, placed in an insulated styrofoam container, and hand-carried via air to Tampa, Florida. Upon arrival in Florida, the water in the "Whirl- Pak" bags was again aerated; the crabs were then shipped by over- night express to the Shannon Point Marine Center on 26 December. Upon their arrival, the crabs, while still in the "Whirl-Pak" bags, were placed in a glass culture dish of sea water until they acclimated to the culture temperature of 30 ~ On 28 December, the crabs were transferred to glass, culture dishes, each with ~ 500 ml filtered sea water (29%o S). The first larvae hatched or 3 January 1991; the majority hatched on 4 January and the remainder on 6 January. Twenty-fourlarvae from each female were reared in individual com- partments in 24-compartmented plastic trays each with ~ 50 ml of natural, filtered sea water (29 to 31%o S); the remainder of the larvae were maintained in mass culture (~ 25 larvae per 500 ml filtered sea water of the same salinity). The trays and mass culture bowls were kept in a constant-temperature unit at 30 ~ 0.5 C~ without light, during the entire rearing period. Water was changed daily and the larvae were provided with a diet of rotifers (Brachionus plicatilis
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Larval development of the hermit crab Clibanarius albidigitus (Crustacea: Anomura: Diogenidae)...

Marine Biology 116, 603-613 (1993)
Marine . . . . . . . . . . . . . . . Biology
(~ Springer-Verlag 1993
Larval development of the hermit crab Clibanarius albidigitus (Crustacea: Anomura: Diogenidae) reared under laboratory conditions E A. Siddiqui 1, p. A. McLaughlin 2, j. A. Crain 2
1 Marine Reference Collection Centre, University of Karachi, Karachi 75270, Pakistan 2 Shannon Point Marine Center, 1900 Shannon Point Road, Anacortes, Washington 98221-4042, USA
Received: 9 March 1993 / Accepted: 7 April 1993
Abstract. The larval development of Clibanarius albidigi- tus Nobili is described and illustrated from laboratory- reared specimens. At 30 ~ this species passes through four zoeal stages before molting to the megalopa. Of the 120 individual larvae reared, survival was high, with 88 % reaching the megalopal stage. Zoeal stage durations varied from 3 to 18 d. Rearing was terminated after 45 d, and at that time no megalopae had molted to the first- crab stage. Among known larvae of CIibanarius species, C. albidigitus is immediately distinguished by the pres- ence of dorsomedial and dorsolateral carapace keels.
Introduction
The usefulness of decapod larval characters in assessing phylogenetic relationships has received considerably sup- port from carcinologists (e.g. Williamson 1976, Rice 1980, 1981, Gore and Scotto 1983, Martin 1984, Gore 1985). In the Brachyura, larval characters have provided the basis for hypotheses of phylogenetic relationships at the generic and familial levels (e.g. Rice and Ingle 1975, Scotto and Gore 1981, Pohle 1989). In contrast to the wealth of information available on brachyuran larvae, complete descriptions of developmental sequences in her- mit crabs are available for relatively few species in most genera. Clibanarius is one of the most specious genera of the Diogenidae, with more than 58 species described (cf. Fize and Ser6ne 1955), and from the standpoint of our knowledge of diogenid larval deVelopment, also the best known. The description of the larval development of C. albidigitus Nobili brings to 11 the number of Clibanarius species for which larval sequences have been described. It is also the first of the eastern Pacific species of the genus to be reared in the laboratory.
Brossi-Garcia (1987) summarized the principal mor- phological characters of eight species of Clibanarius,
Correspondence to: P. A. McLaughlin
omitting only C. longitarsus (DeHaan) and C. olivaceus Henderson described by Ajmal Khan and Natarajan (1981 a, b). A perusal of her data shows a generally con- servative developmental pattern within the genus, al- though Siddiqui et al. (1991) have reported appreciable intraspecific variations among populations of the western Atlantic C. antillensis Stimpson. Only in the development of carapace keels do the zoeae of C. albidigitus differ significantly from other known CIibanarius larvae.
Materials and methods
Five ovigerous females of Clibanarius albidigitus Nobili were col- lected intertidally on 21 December 1990 at Culebra Island on the Pacific coast of Panam~ by one of us (JAC). The collection site consisted of rocks, cobble and areas of sand. In addition to C. albidigitus, the area was populated by Calcinus explorator Boone and an unidentified species of Paguristes. The semiterrestrial Coeno- bita compressus H. Milne Edwards was found occasionally among the rocks.
The five females of Cilbanarius albidigitus were placed in aerated local sea water and returned to the laboratory at the Smithsonian Tropical Research Institute (STRI). Sea water temperatures at STRI ranged from 29 to 31 ~ On 24 December, the crabs were transferred to plastic "Whirl-Pak ''| bags, each containing ~ 150 ml of local sea water, enclosed in second "Ziploc ''| plastic bags, placed in an insulated styrofoam container, and hand-carried via air to Tampa, Florida. Upon arrival in Florida, the water in the "Whirl- Pak" bags was again aerated; the crabs were then shipped by over- night express to the Shannon Point Marine Center on 26 December. Upon their arrival, the crabs, while still in the "Whirl-Pak" bags, were placed in a glass culture dish of sea water until they acclimated to the culture temperature of 30 ~ On 28 December, the crabs were transferred to glass, culture dishes, each with ~ 500 ml filtered sea water (29%o S). The first larvae hatched or 3 January 1991; the majority hatched on 4 January and the remainder on 6 January. Twenty-four larvae from each female were reared in individual com- partments in 24-compartmented plastic trays each with ~ 50 ml of natural, filtered sea water (29 to 31%o S); the remainder of the larvae were maintained in mass culture (~ 25 larvae per 500 ml filtered sea water of the same salinity). The trays and mass culture bowls were kept in a constant-temperature unit at 30 ~ 0.5 C ~ without light, during the entire rearing period. Water was changed daily and the larvae were provided with a diet of rotifers (Brachionus plicatilis

604 E A. Siddiqui et al.: Development in Clibanarh4s albidigitus
I IA I
\
C ~--JK, D I ~ E
I B, C
Fig. 1. Clibanarius albidigitus. (A) Zoea I; (B) Zoea II; (C) Zoea III; (D) Zoea IV; (E) Megalopa, Scale bars = 0.25 mm
I
Miiller) and algae (Isochrisis galbana Parke). On the second day after hatching, the larval diet was supplemented with freshly hatched Artemia sp. nauplii. Methodology as well as terminology for the descriptions follow those of McLaughlin and Gore (1988)- Descriptions are based on molts and on preserved and living stages observed throughout development. N refers to the number of isolat- ed individuals reported for each stage that survived through that stage and successfully molted to the next; n refers to the mmaber of individuals measured and/or dissected. One measurement, CL, re- ferring to carapace length, was taken with an ocular micrometer. This length was measured from the rostral tip to the posterior midpoint of the carapace. Voucher specimens have been deposited in the National Museum of Natural History, Smithsonian Institu- tion.
Results - Description of Clibanarius albidigitus larvae
Zoea I
Size. C L = 0 . 6 3 to 0.76 ram; n=21.
Duration. 4 to 9 d; N = 119.
Carapace (Fig. 1A). Rostrum broadly triangular, mod- erately elongate. Carapace with short, weak median keel and pair of longer dorsolateral ridges; posterolateral margins without spines; eyes sessile.
Abdomen (Fig. 1A). Five somites and telson; pos- terodorsal margins and posterolateral angles unarmed; no pleopod buds.
Telson (Figs. 1 A, 2 A ). Elongate, fan-shaped posteriorly and with deep median cleft; posterior margin with 7 + 7 processes (1, ii, 3 -7) , outermost articulated simple pro- cess (1), second process an anomuran hair (ii), third through seventh articulated processes, plumose setae (3 to 7); no anal spine.
Antennule (Fig. 2B). Unsegmented; 2 to 4 aesthetascs and 2 or 3 simple setae terminally, 1 long plumose termi- nal seta at site of future endopodal bud.
Antenna (Fig. 2 C). Endopod unsegmented, with 2 or 3 terminal plumose setae; scaphocerite bluntly blade-
E A. Siddiqui et al.: Development in Clibanarius albidigitus
, /
////
605
% f
I IB.
I I d
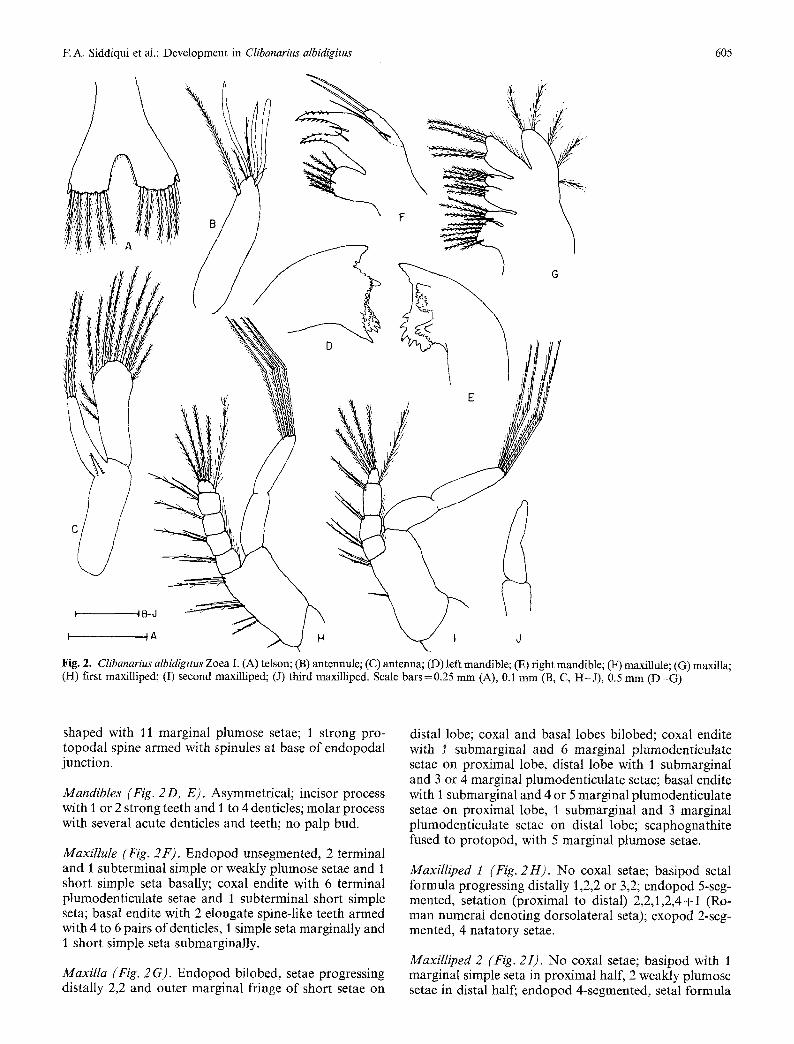
Fig. 2. Clibanarius albidigitus Zoea I. (A) tels0n; (B) antennule; (C) antenna; (D) left mandible; (E) right mandible; (F) maxillule; (G) maxilla; (H) first maxiUiped; (I) second maxilliped; (J) third maxilliped. Scale bars=0.25 mm (A), 0.1 mm (B, C, H-J) , 0.5 mm (D-G)
shaped with l l marginal plumose setae; 1 strong pro- topodal spine armed with spinules at base of endopodal junction.
Mandibles (Fig. 2 D, E). Asymmetrical; incisor process with I or 2 strong teeth and 1 to 4 denticles; molar process with several acute denticles and teeth; no palp bud.
Maxillule (Fig. 2F). Endopod unsegmented, 2 terminal and I subterminal simple or weakly plumose setae and 1 short simple seta basally; coxal endite with 6 terminal plumodenticulate setae and I subterminal short simple seta; basal endite with 2 elongate spine-like teeth armed with 4 to 6 pairs ofdenticles, 1 simple seta marginally and 1 short simple seta submarginally.
Maxilla (Fig. 2 G). Endopod bilobed, setae progressing distally 2,2 and outer marginal fringe of short setae on
distal lobe; coxal and basal lobes bilobed; coxal endite with I submarginal and 6 marginal plumodenticulate setae on proximal lobe, distal lobe with 1 submarginal and 3 or 4 marginal plumodenticulate setae; basal endite with 1 submarginal and 4 or 5 marginal plumodenticulate setae on proximal lobe, I submarginal and 3 marginal plumodenticulate setae on distal lobe; scaphognathite fused to protopod, with 5 marginal plumose setae.
MaxiIliped 1 (Fig. 2H). No coxal setae; basipod setal formula progressing distally 1,2,2 or 3,2; endopod 5-seg- mented, setation (proximal to distal) 2,2,1,2,4+I (Ro- man numeral denoting dorsolateral seta); exopod 2-seg- mented, 4 natatory setae.
Maxitliped 2 (Fig. 2I). No coxal setae; basipod with 1 marginal simple seta in proximal half, 2 weakly plumose setae in distal half; endopod 4-segmented, setal formula

EA. Siddiqui et al.: Development in Clibanarius albidigitus
a .
G
606
Fig. 3. Ctibanarius albidigitus Zoea II. (A) telson; (B) antennule; (C) antenna; (D) left mandible; (E) right mandible; (F) maxillule; (G) maxilla; (H) first maxilliped; (I) second maxilliped; (J) third maxilliped. Scale bars = 0.25 mm (A), 0.1 mm (B-E, H-J), 0.5 mm (F-G)
progressing distally 2,2,2,4+I; exopod 2-segmented, 4 terminal natatory setae.
Maxilliped 3 (Fig. 2J). Exopod unsegmented, simple, with pointed or blunt tip.
CoIour. Zoea transparent but with overall reddish-yellow tint; red chromatophores as follows: on rostrum basally, at bases of antennules, antennae, mandibles and first and second maxillipeds, on lateral and posterior carapace; Yellow chromatophores dorsally over gut. Corneae black.
Zoea II
Size. CL=0.85 to 1.20 ram; n=13.
Duration. 3 to 9 d; N=118.
Carapace (Fig. 1 B). Larger; median keel extending onto rostrum at least in proximal half, dorsolateral ridges con- tinuous with proximal rostral margins. Rostrum still broadly triangular, but now with slender tip; eyes stalked.
Abdomen (Fig. 1 B). Essentially unchanged from previ- ous stage.
Telson (Figs. 1B, 3 A). Posterior marginal formula now 8 + 8 (l,ii, 3-8) with the addition of pair of median setae; median cleft still present but not as deep.
Antennule (Fig. 3 B). Exopod bud clearly delineated, still with 3 or 4 terminal aesthetascs and 3 or 4 simple or weakly plumose terminal setae; i long plumose seta on faintly developing endopod; 1 or 2 short simple setae and 2 long plumose setae at protopodal margin.

E A. Siddiqui et al.: Development in Clibanarius albidigitus
\
B
607
f H
~ G " ~ " ~ ~ / ~ I I B- H F J ~A
Fig. 4. Clibanarius albidigitus Zoea III. (A) telson; (B) antennule; (C) antenna; (D) maxillule; (E) maxilla; (F) first maxilliped; (G) second maxilliped; (H) third maxilliped. Scale bars=0.25 mm (A); 0.1 mm (B, C, F-H); 0.5 mm (D, E)
Antenna (Fig. 3 C). Endopod lengthened, with 3 terminal plumose setae; scaphocerite still with 11 or 12 plumose marginal setae; protopodal spine unchanged.
Mandibles (Fig. 3 D, E). Larger; dentition stronger but essentially unchanged from previous stage.
Maxillule (Fig. 3F). Endopod and coxal endite un- changed from previous stage; basal endite now with 4 spine-like teeth armed with marginal spinules, still with 2 short simple marginal or submarginal setae.
Maxilla (Fig. 3 G). Endopod, coxal and basal endites un- changed from previous stage; scaphognathite now with 7 marginal plumose setae.
Maxilliped 1 (Fig. 3 H). Basipod unchanged; endopod with setal formula now 1 + I, 2 + I, 1 + I, 2,4 + I; exopod incompletely 2-segmented, with 6 natatory setae.
Maxilliped 2 (Fig. 3I). Basipod unchanged; endopod with setal formula now 2,2 + 1,2 + 1,4 + I; exopod with 6 natatory setae.
Maxilliped 3 (Fig. 3 J). Exopod 2-segmented, with 5 or 6 natatory setae.
Colour. Generally as in Stage I, but with additional pair of yellow chromatophores on second abdominal somite.

608 E A. Siddiqui et al.: Development in Clibanarius albidigitus
Zoea III
Size. C L = I . 2 to 1.40; n=13.
Colour. Additional red-orange chromatophore on ros- trum distally, and on protopod of third maxilliped, dis- tinctly red chromatophore on sixth somite and on telson.
Duration. 4 to 12 d; N=116.
Carapace (Fig. 1 C). Larger than in preceding stage; dor- somedial keel now extending to near posterior carapace margin.
Abdomen (Fig. 1 C). Sixth somite clearly delineated; uropods with endopodal bud not delineated from pro- topod; exopod with 6 to 8 marginal plumose setae.
Telson (Figs. 1 C, 4 A). Posterior marginal formula now 8 + 1 + 8 (1,ii,3,IV,5- 8 + 1) with addition of single, articu- lated median process; first process often slightly reduced in size, fourth process transformed to strong fused spine; median cleft absent.
Antennule (Fig. 4B). Exopod bud still with 2 to 4 termi- nal aesthetascs, now with 2 short and 3 long setae on protopod at exopodal junction and 1 submarginally; en- dopod delineated from protopod, naked.
Zoea IV
Size. C L = 1.5 to 1.7 mm; n = 12.
Duration. 5 to 18 d; N = 111.
Carapace (Fig. 1D). Larger but otherwise unchanged; dorsomedial carapace keel extending to posterior mar- gin.
Abdomen (Fig. 1 D). Somites unchanged from previous stage; uropods clearly biramous, endopods no longer fused to protopods; exopod with 8 to 10 marginal plumose setae and 2 or 3 short simple setae on dorsal surface, endopod with 4 to 6 marginal plumose setae; pleopod buds present.
Telson (Figs. 1 D, 5 A ). Except for continued reduction of first process, unchanged from previous stage.
Antenna (Fig. 4 C). Scaphocerite spatulate, now with 12 marginal plumose setae; protopod still with prominent plumodenticulate spine; endopod still unsegmented, with 1 simple or plumose terminal seta; protopodal spine still well developed.
Mandible. Teeth of incisor process stronger, but configu- ration unchanged.
Maxillule (Fig. 4D). Endopod 2-segmented, 2 terminal plumose setae on ultimate segment, 1 long plumose seta distally and 1 short simple seta proximally on basal seg- ment; coxal and basal endites unchanged, or 1 submar- ginal seta of basal endite occasionally denticulate.
Maxilla (Fig. 4 E). Endopod unchanged from previous stage; coxal endite now with 5 submarginal and 3 margin- al plumose or plumodenticulate setae on proximal lobe, distal lobe with 1 marginal and 3 submarginal plumose or plumodenticulate setae; basal endite unchanged; scaphognathite now with 9 to 11 marginal plumose setae, posterior lobe usually not delineated.
Maxilliped 1 (Fig. 4F). Basipodal setal formula now usually 1,2,3,3; endopodal setation unchanged; exopod still with 6 natatory setae.
MaxilIiped 2 (Fig. 4G). Basipod and endopod un- changed or, rarely, antipenultimate segment with 1 + I setae; exopod still with 6 natatory setae.
Maxilliped 3 (Fig. 4 H). Endopod bud present; exopod with 5 or 6 natatory setae.
Pereopods. Developing beneath carapace posteriorly.
Antennule (Fig. 5 B). Exopodal bud incompletely 3-seg- mented, with 3 terminal aesthetascs and 3 or 4 simple or plumose setae, pair of aesthetascs on penultimate and antipenultimate segments; endopodal bud slightly larger but otherwise unchanged; protopod with 3 long marginal and 1 submarginal plumose setae and 3 or 4 short simple setae.
Antenna (Fig. 5 C). Scaphocerite still spatulate, now with 11 to 12 marginal, plumose setae; endopod considerably overreaching scaphocerite, weakly 4- or 5-segmented and with I or occasionally 2 terminal setae.
Mandibles (Fig. 5D, E). Enlarged and with more pro- nounced dentition; palp bud now present, well developed.
MaxilluIe (Fig. 5F). Endopod unchanged from previous stage; coxal endite now with 1 or 2 submarginal and 7 or 8 marginal setae; basal endite unchanged from previous stage.
Maxilla (Fig. 5 G). Endopod unchanged; coxal endite now with 3 or 4 marginal and 7 submarginal setae on proximal lobe, 3 or 4 marginal and 1 submarginal setae on distal lobe; basal endite with 3 marginal and 3 or 4 submarginal setae on proximal lobe, 4 marginal and 1 submarginal setae on distal lobe; scaphognathite with 12 to 16 marginal plumose setae on distal lobe, 0 to 2 mar- ginal plumose setae on now distinct proximal lobe.
Maxilliped 1 (Fig. 5 H). Endopod unchanged; exopod with 6 or 7 natatory setae.
Maxilliped 2 (Fig. 5I). Endopod unchanged; exopod with 7 or 8 natatory setae.

F. A, Siddiqui et al.: Development in Clibanarius albidigitus 609
\
k.,
B
,/
J J J ~
I tB-J
t I A I \ J
f
E
Fig. 5. Clibanarius albidigitus Zoea IV. (A) telson; (B) antennule; (C) antenna; (D) left mandible; (E) right mandible; (F) maxilluIe; (G) maxilla; (H) first maxilliped; (I) second maxilliped; (J) third maxilliped. Scale bars =0.25 mm (A); 0.1 mm (B-E, H-J); 0.5 mm (F-G)
Maxilliped 3 (Fig. 5 J). Endopod bud now incompletely 3- to 5-segmented; exopod unchanged.
Pereopods. Chelipeds with chelae enlarged, dactyls dis- tinct; ambulatory legs moderately well developed.
Colour. Carapace and appendages with overall yellow hue, but with margins bluish; abdominal somites bluish- yellow ventrally and yellow dorsally; additional red chro- matophores on ventral carapace anteriorly beneath eyes.
Megalopa
Size. CL=0.8 to 1.0 ram; n=12.
Duration. At least 30 d. None survived to moult to Crab Stage 1.
Carapace (Fig. 1 E). Shield slightly broader than long to longer than broad, branchiostegites with few to numer- ous marginal plumose setae; rostrum broad and bluntly

610 E A. Siddiqui et al.: Development in Clibanarius albidigitus
F
...... E
I tA,B
Fig. 6. Clibanarius albidigitus Megalopa. (A) antennule; (B) antenna; (C) mandible; (D) maxillule; (E) maxilla; (F) first maxilliped; (G) second maxilliped; (H) third maxilliped. Scale bars=0.25 mm (A, B); 0.1 mm (C-H)
triangular. Ocular peduncles stout, distal margin of corneae often overreaching ultimate antennular segment; ocular acicles only weakly developed.
Antennule (Fig. 6 A). Biramous; peduncle 3-segmented; basal segment with 5 or 6 short setae dorsally, 1 or 2 setae on distal margin ventrally and 1 or 2 on ventral margin; penultimate and ultimate segments each with 3 to 5 short setae; exopod (upper ramus) 5- or more frequently 6-seg- mented, aesthetascs (shown in parentheses) and setae (no parentheses) progressing distally (0),(6),(4 or 5),(3),3 or 4 (5th segment), and 3 on terminal segment; endopod (lower ramus) 3-segmented, with 2 to 4, 2 or 3 and 4 or 5 setae, respectively.
Antenna (Fig. 6B). Supernumerary segment not dearly delineated; second segment with acute dorsomesial distal angle; acicle weakly serrate, with 3 or 4 simple marginal
setae and bifid spinose tip, occasionally also small spine on lateral margin; flagellum with 10 segments, basal seg- ment naked, remaining articles each with I to several short setae, 2 or 3 longer terminal setae.
Mandible (Fig. 6 C). Reduced, simple; palp 2- or indis- tinctly 3-segmented, with several marginal or submar- ginal plumodenticulate bristles.
Maxillule (Fig. 6 D). Endopod with internal lobe moder- ately well developed, 1 terminal bristle, external lobe well developed, recurved; coxal endite with 2 rows of short bristles; basal endite with many small teeth and 2 or 3 long plumose setae.
Maxilla (Fig. 6 E). Endopod narrowing distally, slightly twisted, naked or occasionally with short seta subtermi- nally; proximal and distal lobes of coxal and basal endites

E A. Siddiqui et al.: Development in Clibanarius albidigitus 611
\
f
A
r
i - r
I
Fig. 7. Clibanarius albidigitus Megalopa. (A) left cheliped; (B) right second pereopod; (C) right third pereopod; (D) right fourth pereopod; (E) left fifth pereopod; (F-H), pleopods of abdominal somites 2, 3, and 5; (I) tailfan. Scale bars=0.25 mm
each with numerous setae, in double rows on coxal lobes and single rows on basal lobes; scaphognathite elongate, with 45 to 60 marginal plumose setae on proximal and distal lobes collectively.
Maxilliped ! (Fig. 6F). Exopod 1-segmented, with 9 or 10 marginal setae; endopod unsegmented, simple; coxal endite with several short plumodenticulate setae; basal endite with numerous short plumodenticulate setae and 1 or 2 longer plumose setae distally.
Maxilliped 2 (Fig. 6 G). Exopod 2-segmented, with 6 or 7 terminal setae and 1 or 2 short setae on inner margin; endopod 4-segmented, ultimate segment with numerous plumose or plumodenticulate setae, remaining segments each with 1 to 3 setae.
Maxilliped 3 (Fig. 6H). Protopod occasionally with 2 to 4 blunt denticles on inner margin. Exopod 2-segmented, with 6 terminal setae; endopod 5-segmented, ultimate and penultimate segments with numerous setae, many
barbed; remaining segments with setae as shown, crista dentata with 4 or 5 denticles.
Gills. Moderately well developed, lobular arthrobranchs on chelipeds and Pereopods 2 to 4; developing pleuro- branch on body wall above Pereopod 4.
Pereopods. Chelipeds (Fig. 7 A) generally equal, unarmed but with scattered setae. Ambulatory legs (Fig. 7 B, C) moderately long; dactyls each with 3 or 4 corneous spin- ules on ventral margins; all segments with scattered setae. Fourth pereopods (Fig. 7 D) with 2 incomplete rows of corneous scales comprising propodal rasp; dactyl with 1 long plumose seta distally. Fifth pereopods (Fig. 7E) with few corneous scales on propodi and dactyls, 3 long plumose setae on distal margin of propodus.
Pleopods (Fig. 7 F, G, H). Paired pleopods developed on Abdominal Somites 2 to 5, decreasing in size posteriorly; exopods with 9 plumose setae; endopods unsegmented, each with appendix interna of 2 small apical hooks.
612
Abdomen (Fig. 1 E). Somites 2 to 5 with posterolateral angles rounded, each with 1 or 2 short setae; sixth somite with 2 pairs of setae on posterior margin, 3 short setae at posterolateral angles dorsally, 3 or 4 short setae on lateral margins and 2 or 3 pairs of setae on dorsal surface.
Tailfan (Figs. 1 E, 7I). Uropods with small spine and 1 or 2 short setae on protopodal distal angle; exopods with 16 to 19 marginal setae and 11 to 13 corneous scales; en- dopods with 1 or 2 simple setae on dorsal surface, 9 to 13 marginal setae and 8 to 10 corneous scales. Telson with terminal margin rounded, 9 setae on posterior margin, shorter setae on lateral margins and on dorsal surface (as illustrated).
Colour. Entire body and appendages with yellowish-red hue; shield with horizontal red band anteriorly, bran- chiostegites and posterior carapace margin red. Mandibles and bases of appendages also red. Corneae black.
E A. Siddiqui et al.: Development in Clibanarius albidigitus ,00 8 0 ~ 1
t
2~ ' 0 ~ I ~ , , ~ ~ , , , ~ , , ~ b ~ h L , h
5 9 13 17 21 25 29 33 Days
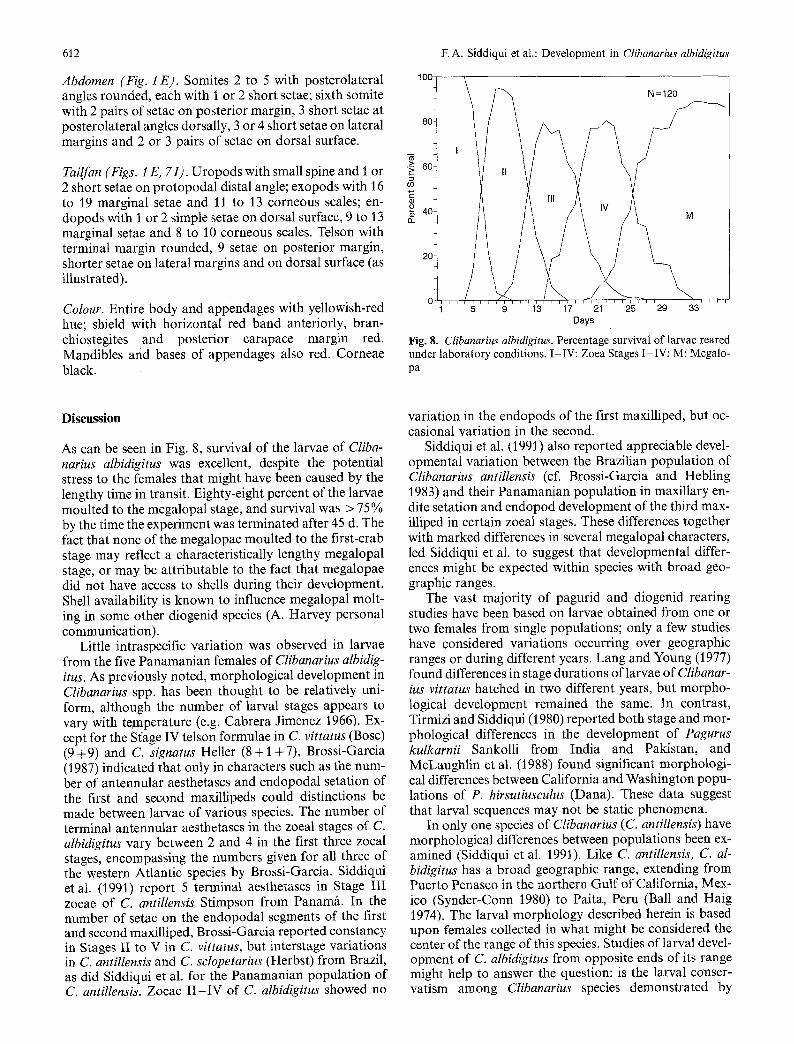
Fig. 8. Clibanarius albidigitus. Percentagelsurvival of larvae reared under laboratory conditions. I-IV: Zoea Stages I-IV; M: Megalo- pa
Discussion
As can be seen in Fig. 8, survival of the larvae of Cliba- narius albidigitus was excellent, despite the potential stress to the females that might have been caused by the lengthy time in transit. Eighty-eight percent of the larvae moulted to the megalopal stage, and survival was > 75% by the time the experiment was terminated after 45 d. The fact that none of the megalopae moulted to the first-crab stage may reflect a characteristically lengthy megalopal stage, or may be attributable to the fact that megalopae did not have access to shells during their development. Shell availability is known to influence megalopal molt- ing in some other diogenid species (A. Harvey personal communication).
Little intraspecific variation was observed in larvae from the five Panamanian females of Clibanarius albidig- itus. As previously noted, morphological development in Clibanarius spp. has been thought to be relatively uni- form, although the number of larval stages appears to vary with temperature (e.g. Cabrera Jimenez 1966). Ex- cept for the S~/age IV telson formulae in C. vittatus (Bosc) (9 +9) and C. signatus Heller (8 + 1 + 7), Brossi-Garcia (1987) indicated that only in characters such as the num- ber of antennular aesthetascs and endopodal setation of the first and second maxillipeds could distinctions be made between larvae of various species. The number of terminal antennular aesthetascs in the zoeal stages of C. albidigitus vary between 2 and 4 in the first three zoeal stages, encompassing the numbers given for all three of the western Atlantic species by Brossi-Garcia. Siddiqui et al. (1991) report 5 terminal aesthetascs in Stage III zoeae of C. antiIlensis Stimpson from Panamg. In the number of setae on the endopodal segments of the first and second maxilliped, Brossi-Garcia reported constancy in Stages II to V in C. vittatus, but interstage variations in C. antillensis and C. selopetarius (Herbst) from Brazil, as did Siddiqui et al. for the Panamanian population of C. antillensis. Zoeae I I - IV of C. albidigitus showed no
variation in the endopods of the first maxilliped, but oc- casional variation in the second.
Siddiqui et al. (1991) also reported appreciable devel- opmental variation between the Brazilian population of Clibanarius antillensis (cf. Brossi-Garcia and Hebling 1983) and their Panamanian population in maxillary en- dite setation and endopod development of the third max- illiped in certain zoeat stages. These differences together with marked differences in several megalopal characters, led Siddiqui et al. to suggest that developmental differ- ences might be expected within species with broad geo- graphic ranges.
The vast majority of pagurid and diogenid rearing studies have been based on larvae obtained from one or two females from single populations; only a few studies have considered variations occurring over geographic ranges or during different years. Lang and Young (1977) found differences in stage durations of larvae of Clibanar- ius vittatus hatched in two different years, but morpho- logical development remained the same. In contrast, Tirmizi and Siddiqui (1980) reported both stage and mor- phological differences in the development of Pagurus kulkarnii Sankolli from India and Pakistan, and McLaughlin et al. (1988) found significant morphologi- cal differences between California and Washington popu- lations of P. hirsutiusculus (Dana). These data suggest that larval sequences may not be static phenomena.
In only one species of Ctibanarius (C. antillensis) have morphological differences between populations been ex- amined (Siddiqui et al. 1991). Like C. antillensis, C. al- bidigitus has a broad geographic range, extending from Puerto Penasco in the northern Gulf of California, Mex- ico (Synder-Conn 1980) to Paita, Peru (Ball and Haig 1974). The larval morphology described herein is based upon females collected in what might be considered the center of the range of this species. Studies of larval devel- opment of C. albidigitus from opposite ends of its range might help to answer the question: is the larval conser- vatism among Clibanarius species demonstrated by

E A. Siddiqui et al.: Development in Clibanarius albidigitus
Bross i -Ga rc i a (1987) an a r t i fac t o f res t r ic ted rear ing events o r is the in t raspeci f ic va r i a t i on r epo r t ed for C. antillensis a un ique p h e n o m e n o n for this genus?
Acknowledgements. The authors acknowledge, with thanks, the co- operation extended by Dr. J. Christie and the staff of the Smithso- nian Tropical Research Institute in the collection of the females, and the assistance provided by the staff of the Shannon Point Marine Center in the larval rearing. This research was supported, in part, by National Science Foundation Grant BSR-906574, and is a scien- tific contribution from the Shannon Point Marine Center.
Literature cited
Ajmal Khan, S., Natarajan, R. (1981a). Laboratory rearing of larval stages of the estuarine hermit crab Clibanarius longitarus (De Haan) (Decapoda: Anomura). Indian J. mar. Sci. 10:74-81
Ajmal Khan, S., Natarajan, R. (1981 b). Metamorphosis of an estu- arine hermit crab Clibanarius olivaceus Henderson in the labora- tory (Crustacea: Decapoda: Anomura). Mahasagar (Bull. natn. Inst. Oceanogr., India) 14:265-276
Ball, E. E., Haig, J. (1974). Hermit crabs from the tropical eastern Pacific. I. Distribution, color, and natural history of some com- mon shallow-water species. Bull. Sth. Calif. Acad. Sci. 73: 95- 144
Brossi-Garcia, A. L. (1987). Morphology of the larval stages of Clibanarius scIopetarius (Herbst, 1796) (Decapoda, Diogenidae) reared in the laboratory. Crustaceana 52:251-275
Brossi-Garcia, A. L., Hebling, N. J. (1983). Desenvolvimento p6sembrionario de Clibanarius antillensis Stimpson, 1859 (Crus- tacea, Diogenidae), em laborat6rio. Bolm Zool., Univ. S Paulo 6:89-111
Cabrera Jimenez, J. A. (1966). Le metamorfosis larvaria de Cliba- narius vittatus (Bosc): descripcion de los primeros estadios y efecto de la temperatura en la supervivencia. Thesis. Universi- dad Nacional Aut6mona de Mexico City
Fize, A., Ser+ne R (1955). Les pagures du Vietnam. Notes Inst. oc6anogr., Nhatrang 45:1-228
Gore, R. H. (1985). Molting and growth in decapod larvae. In: Wenner, A. M. (ed.) Crustacean growth: larval growth. Crus- tacean issues. VoI. 2. A. A. Balkema, Rotterdam, p. 1-65
Gore, R. H., Scotto, L. E. (1983). Studies on decapod Crustacea from the Indian River region of Florida. XXVII. Phimochirus holthuisi (Provenzano, 1961) (Anomura: Paguridae): the com- plete larval development under laboratory conditions, and the systematic relationships of its larvae. J. Crustacean Biol. 3: 93- 116
Lang, W H., Young, A. M. (1977). The larval development of Clibanarius vittatus (Bosc) (Crustacea: Decapoda; Diogenidae)
613
reared in the laboratory. Biol. Bull. mar. biol. Lab., Woods Hole 152:84-t04
Martin, J. W. (1984). Notes and bibliography of the larvae of xan- thid crabs, with a key to the known xanthid zoeae of the western Atlantic and Gulf of Mexico. Bull. mar. Sci. 34:220-239
McLaughlin, P. A., Gore, R. H. (1988). Studies on the provenzanoi and other pagurid groups. I. The larval stages of Pagurus maclaughlinae Garcia-Gdmez, 1982 (Decapoda: Anomura: Paguridae) reared under laboratory conditions, J. Crustacean Biol. 8:247-277
McLaughlin, P. A., Gore, R. H., Crain, J. A. (1988). Studies on the provenzanoi and other pagurid groups. II. A reexamination of the larval stages of Pagurus hirsutiusculus hirsutiusculus (Dana) (Decapoda: Anomura: Paguridae) reared in the laboratory. J. Crustacean Biol. 8:430-450
Pohle, G. (1989). Development and morphology of laboratory- reared larvae of Dissodactylus nitidus Smith, 1840 (Decapoda: Brachyura), with a discussion of phylogenetic aspects in the Pinnotheridae. J. Crustacean Biol. 9:278-296
Rice, A. L. (1980). Crab zoeal morphology and its bearing on the classification of the Brachyura. Trans. zool. Soc. Lond. 35: 1 461
Rice, A. L. (1981). Crab zoeae and brachyuran classification: a reappraisal. Bull Br. Mus. nat. Hist. (D: Zool.) 40:287-296
Rice, A. L., Ingle, R. W (1975). A comparative study of the larval morphology of the British portunid crabs Macropipus puber (L.) and M. hoIsatus (Fabricius), with a discussion of generic and sub-familial larval characters within the Portunidae. Bull. Br. Mus. nat. Hist. (D: Zool). 28:123-151
Scotto, L. E., Gore, R. H. (1981). Studies on decapod Crustacea from the Indian River region of Florida. XXIII. The laboratory cultured zoeal stages of the coral gall-forming crab Troglocarci- nus corallicola Verrill, 1908 (Brachyura: Hapalocarcinidae) and its familial position. J. Crustacean Biol. 1:486-505
Siddiqui, E A., McLaughlin, P. A., Crain, J. A. (1991). Larval development of Clibanarius antillensis Stimpson, 1859 (Crus- tacea: Anomura: Diogenidae) reared under laboratory condi- tions: a comparison between Panamanian and Brazilian popula- tions. J. nat. Hist. 25:917-932
Snyder-Conn, E. (1980). Arthropoda: Crustacea Paguroidea and Coenobitoidea (hermit crabs). In: Brusca, R. C. (ed.) Common intertidal invertebrates of the Gulf of California. University of Arizona Press, Tucson, p. 275-285
Tirmizi, N. M., Siddiqui, E A. (1980). Notes on the laboratory reared larvae of Pagurus kulkarnii Sankolli (Decapoda, Paguridae). Crustaceana 38:155-168
Williamson, D. I. (1976). Larval characters and the origin of crabs (Crustacea, Decapoda, Brachyura). Thalassia jugosl. 10: 401- 414
C o m m u n i c a t e d by M. G. Hadf ie ld , H o n o l u l u




















