
Miocene squat lobsters (Decapoda, Anomura, Galatheoidea) of the Central Paratethys – a review,...
28
Miocene squat lobsters (Decapoda, Anomura, Galatheoidea) of the Central Paratethys – a review, with description of a new species of Munidopsis M. Hyžný, R. Gašparič, C.M. Robins & J. Schlögl Hyžný, M., Gašparič, R., Robins, C.M. & Schlögl, J. Miocene squat lobsters (Decapoda, Anomura, Gala- theoidea) of the Central Paratethys – a review, with description of a new species of Munidopsis. In: Fraaije, R.H.B., Hyžný, M. Jagt, J.W.M., Krobicki, M. & Van Bakel, B.W.M. (eds.), Proceedings of the 5 th Symposi- um on Mesozoic and Cenozoic Decapod Crustaceans, Krakow, Poland, 2013: A tribute to Pál Mihály Müller. Scripta Geologica, 147: 241-267, 7 figs., 2 tables, 6 pls., Leiden, October 2014. Matúš Hyžný, Naturhistorisches Museum Wien, Burgring 7, 1010 Vienna, Austria, and Department of Geology and Palaeontology, Faculty of Natural Sciences, Comenius University, Mlynská dolina G1, 842 15 Bratislava, Slovakia ([email protected]); Rok Gašparič, Ljubljanska cesta 4j, 1241 Kamnik, Slovenia ([email protected]); Cristina M. Robins, Florida Museum of Natural History, University of Florida, Dickinson Hall, 1659 Museum Road, Gainesville, FL 32608, USA; Ján Schlögl, Department of Geology and Palaeontology, Faculty of Natural Sciences, Comenius University, Mlynská dolina G1,842 15 Bratis- lava, Slovakia. Key words – Neogene, Austria, Slovakia, revision, new species. All squat lobsters of the families Galatheidae, Munididae and Munidopsidae from the Miocene of the Central Paratethys are reviewed taxonomically. Based on additional observations emended diagnoses are provided for Agononida cerovensis and Galathea weinfurteri, from the Lower and Middle Miocene, respectively. Munidopsis is represented by two species in the study area; additional data for M. lieskoven- sis from the Lower Miocene of Slovakia are presented and a new species, M. palmuelleri, from the Middle Miocene of Slovenia is erected. Implications for palaeoenvironmental reconstructions are briefly dis- cussed for each taxon. Contents Introduction ............................................................................................................................................................ 241 Material and methods ...................................................................................................................................... 242 Systematic palaeontology .............................................................................................................................. 243 Notes on palaeoenvironment ....................................................................................................................... 253 Acknowledgements ........................................................................................................................................... 255 References ................................................................................................................................................................ 255 Introduction The superfamily Galatheoidea currently comprises five families; Galatheidae Samou- elle, 1819, Munididae Ahyong, Baba, Macpherson & Poore, 2010, Munidopsidae Whiteaves, 1874, Porcellanidae Haworth, 1825 and Retrorsichelidae Feldmann, Tshudy & Thomson, 1993. The last-named is known exclusively from the fossil record. Except for the Retrorsichelidae, all galatheoid families have previously been recorded from the Mio- cene of the Central Paratethys (see e.g., Bachmayer, 1950; Müller, 1984a; Hyžný & Schlögl, 2011; see Table 1). The present contribution is a review of galatheid, munidid and muni- dopsid squat lobsters from the Miocene of the Central Paratethys; the porcelain crabs (Porcellanidae) will be revised in a forthcoming paper.
Transcript of Miocene squat lobsters (Decapoda, Anomura, Galatheoidea) of the Central Paratethys – a review,...

Miocene squat lobsters (Decapoda, Anomura, Galatheoidea) of the Central Paratethys – a review, with description
of a new species of Munidopsis
M. Hyžný, R. Gašparič, C.M. Robins & J. Schlögl
Hyžný, M., Gašparič, R., Robins, C.M. & Schlögl, J. Miocene squat lobsters (Decapoda, Anomura, Gala-theoidea) of the Central Paratethys – a review, with description of a new species of Munidopsis. In: Fraaije, R.H.B., Hyžný, M. Jagt, J.W.M., Krobicki, M. & Van Bakel, B.W.M. (eds.), Proceedings of the 5th Symposi-um on Mesozoic and Cenozoic Decapod Crustaceans, Krakow, Poland, 2013: A tribute to Pál Mihály Müller. Scripta Geologica, 147: 241-267, 7 figs., 2 tables, 6 pls., Leiden, October 2014. Matúš Hyžný, Naturhistorisches Museum Wien, Burgring 7, 1010 Vienna, Austria, and Department of Geology and Palaeontology, Faculty of Natural Sciences, Comenius University, Mlynská dolina G1, 842 15 Bratislava, Slovakia ([email protected]); Rok Gašparič, Ljubljanska cesta 4j, 1241 Kamnik, Slovenia ([email protected]); Cristina M. Robins, Florida Museum of Natural History, University of Florida, Dickinson Hall, 1659 Museum Road, Gainesville, FL 32608, USA; Ján Schlögl, Department of Geology and Palaeontology, Faculty of Natural Sciences, Comenius University, Mlynská dolina G1,842 15 Bratis-lava, Slovakia.
Key words – Neogene, Austria, Slovakia, revision, new species.All squat lobsters of the families Galatheidae, Munididae and Munidopsidae from the Miocene of the Central Paratethys are reviewed taxonomically. Based on additional observations emended diagnoses are provided for Agononida cerovensis and Galathea weinfurteri, from the Lower and Middle Miocene, respectively. Munidopsis is represented by two species in the study area; additional data for M. lieskoven-sis from the Lower Miocene of Slovakia are presented and a new species, M. palmuelleri, from the Middle Miocene of Slovenia is erected. Implications for palaeoenvironmental reconstructions are briefly dis-cussed for each taxon.
Contents
Introduction ............................................................................................................................................................ 241Material and methods ...................................................................................................................................... 242Systematic palaeontology .............................................................................................................................. 243Notes on palaeoenvironment ....................................................................................................................... 253Acknowledgements ........................................................................................................................................... 255References ................................................................................................................................................................ 255
Introduction
The superfamily Galatheoidea currently comprises five families; Galatheidae Samou-elle, 1819, Munididae Ahyong, Baba, Macpherson & Poore, 2010, Munidopsidae Whiteaves, 1874, Porcellanidae Haworth, 1825 and Retrorsichelidae Feldmann, Tshudy & Thomson, 1993. The last-named is known exclusively from the fossil record. Except for the Retrorsichelidae, all galatheoid families have previously been recorded from the Mio-cene of the Central Paratethys (see e.g., Bachmayer, 1950; Müller, 1984a; Hyžný & Schlögl, 2011; see Table 1). The present contribution is a review of galatheid, munidid and muni-dopsid squat lobsters from the Miocene of the Central Paratethys; the porcelain crabs (Porcellanidae) will be revised in a forthcoming paper.

242 Hyžný et al. Miocene squat lobsters of the Central Paratethys. Scripta Geol., 147 (2014)
The Paratethys was an extensive network of inland seas intermittently connected to the Mediterranean and Indo-West Pacific (Rögl & Steininger, 1983; Rögl, 1998, 1999; Popov et al., 2004; Harzhauser & Piller, 2007; Harzhauser et al., 2007). Changing sea-ways and landbridges led to internal differentiation of the northern domain into a Western, Central and Eastern Paratethys. The Central Paratethys covered the area from present-day Austria to Poland, Ukraine and Romania; this includes the eastern Alpine-Carpathian Foreland basins (from Lower Austria to Moldavia) and the Pannonian Ba-sin System (see Harzhauser et al., 2003, fig. 10). Summaries of the palaeogeography and stratigraphy of the Central Paratethys include those by Rögl (1998, 1999), Popov et al. (2004), Harzhauser & Piller (2007) and Piller et al. (2007), to which reference is made.
Extinct galatheoids have recently been discussed and/or listed by Schweitzer & Feldmann (2000), De Angeli & Garassino (2002), Schweitzer et al. (2010) and Ahyong et al. (2011). Recently, numerous Jurassic taxa have been reassessed, and a range of new taxa have been erected (Robins et al., 2012a, 2013).
Material and methods
The material studied often is preserved in different ways, being dependent of the sedimentary facies. In carbonates usually isolated carapaces, of which often the rostra are broken, are found; (semi-)articulated specimens have yet to be discovered from such facies. This is the case for a widely distributed species, Galathea weinfurteri (see below). In fine siliciclastic sediments, specimens may retain articulated appendages, but usually only dorsal carapaces are preserved. In addition, the rostrum may be bro-ken (e.g., Agononida cerovensis), but in many cases it is retained, as in both species of Munidopsis described below. Our terminology follows Klompmaker et al. (2012) and Robins et al. (2013). The material studied was obtained either from institutional repositories (e.g., G. wein furteri) or recent collections made by three of us (MH, RG and JS). Specimens were prepared with needles and, where deemed necessary, by using a vibrating pneumatic needle. Where only imprints were preserved, a cast was produced (Stomaflex® Putty). Specimens were photographed dry, either uncoated or coated with ammonium chloride.
Measurements – The following parameters were measured: LR – total length of cara-pace, including rostrum; L – length of carapace, excluding rostrum; GH – length of
Table 1. Galatheoid taxa recorded from the Miocene of the Central Paratethys (CP).
Family Taxon Stratigraphic range in CPGalatheidae Galathea weinfurteri Bachmayer, 1950 ‘Badenian’Munididae Agononida cerovensis Hyžný & Schlögl, 2011 ‘Karpatian’Munidopsidae Munidopsis lieskovensis Hyžný & Schlögl, 2011 ‘Karpatian’Munidopsidae Munidopsis palmuelleri n. sp. ‘Badenian’Porcellanidae Petrolisthes haydni Müller, 1984a ‘Badenian’Porcellanidae Petrolisthes magnus Müller, 1984a ‘Badenian’Porcellanidae Pisidia kokayi (Müller, 1974a) ‘Badenian’Porcellanidae Pisidia aff. viai Müller, 1984b ‘Badenian’

Hyžný et al. Miocene squat lobsters of the Central Paratethys. Scripta Geol., 147 (2014) 243
gastric region; TW – total width of anterior margin; MW – maximum carapace width. All measurements are in millimetres.
Repositories – Specimens studied are housed in the following institutions: IPUW, Institut für Paläontologie, Universität Wien, Vienna (Austria); KGP-MH, Department of Geology and Palaeontology, Comenius University, Bratislava (Slovakia); NHMW, Naturhistorisches Museum Wien, Vienna (Austria); RGA/SMNH, Slovenian Museum of Natural History, Ljubljana (Slovenia); SNM, Natural History Museum of the Slovak National Museum, Bratislava.
Systematic palaeontology
Order Decapoda Latreille, 1802Infraorder Anomura MacLeay, 1838
Superfamily Galatheoidea Samouelle, 1819Family Galatheidae Samouelle, 1819
Genus Galathea Fabricius, 1793
Type species – Cancer strigosus Linnaeus, 1761, by subsequent designation of Latreille (1810).
Diagnosis – See Baba (2005, p. 74).
Remarks – In the fossil record, the present genus is represented by twelve species (see Garassino et al., 2008; Schweitzer et al., 2010), the oldest being G. sahariana Garas-sino, De Angeli & Pasini, 2008 from the Upper Cretaceous of Morocco. However, it should be noted that the chelae of G. sahariana possess proportionally long palms with rather short fingers (Garassino et al., 2008, figs. 11, 12), while typical Galathea chelae have relatively short and spinose palms (Baba, 2005).
Galathea weinfurteri Bachmayer, 1950Figs. 1, 7A; Pls. 1, 2.
1928 Galathea sp.; Glaessner, pp. 164, 206, pl. 3, fig. 1.1950 Galathea weinfurteri Bachmayer, p. 135, pl. 1, figs. 2-4.1952 Galathea weinfurteri; Papp, p. 124.1953 Galathea weinfurteri; Bachmayer, pp. 242, 243, pl. 5, figs. 3, 4, 6.1953 Galathea spec. ind.; Bachmayer & Tollmann, p. 312.1974b Galathea weinfurteri; Müller, p. 276, pl. 1, fig. 4.1976 Galathea weinfurteri; Müller, p. 516.1979 Galathea weinfurteri; Müller, p. 274.1984a Galathea weinfurteri; Müller, p. 60, pl. 21, figs. 4, 5, pl. 22, figs. 1-5.1996 Galathea weinfurteri; Müller, p. 8.1998 Galathea weinfurteri; Müller, p. 14.2002 Galathea weinfurteri; De Angeli & Garassino: p. 26. 2002 Galathea weinfurteri; Górka, p. 528.2006 Galathea weinfurteri; Radwański et al., pp. 96, 97, pl. 2, fig. 1.2010 Galathea weinfurteri; Gatt & De Angeli, p. 1326, pl. 2, fig. 4.

244 Hyžný et al. Miocene squat lobsters of the Central Paratethys. Scripta Geol., 147 (2014)
2010 Galathea sp.; Gatt & De Angeli, p. 1326, text-fig. 3, pl. 2, fig. 3.2010 Galathea weinfurteri; Schweitzer et al., p. 49.2011 Galathea weinfurteri; Ossó & Stalennuy, figs. 3.7, 3.13.
Emended diagnosis – Carapace subrectangular (L/MW 1-1.1); slightly vaulted trans-versely. Rostrum subtriangular with three pairs of lateral spines, one pair of supraorbi-tal spines; no median carina. Rostrum ornamented with small, irregularly spaced tu-bercles. Cervical groove deep; anterior and posterior branch equally incisive. Carapace ornamented with transverse striae; lateral margins with seven spines; carapace surface ornamented with transverse striae; two postfrontal striae, curved in median part and interrupted by short groove.
Type material – The type lot originated from the locality of Furth in Lower Austria (Bachmayer, 1950). That author mentioned that the type material was held in the ‘Wein-furter Sammlung’ at IPUW. However, even after repeated searches by one of us (MH), this material could not be traced. We here consider the holotype (carapace without rostrum) and two paratypes (fragmentary carapace with left part of the rostrum, and left propodus) lost. For comparison, we here reproduce (Pl. 1) the original illustrations of the types.
Fig. 1. Distribution of squat lobsters in the Central Paratethys during the Miocene, projected onto the Re-cent Carpathian-Pannonian Basin complex (after Harzhauser et al., 2003). White areas represent respective Neogene basins. Legend: ‘star’ = Galathea weinfurteri Bachmayer, 1950; ‘circle’ = Agononida cerovensis Hyžný & Schlögl, 2011 and Munidopsis lieskovensis Hyžný & Schlögl, 2011; ‘pentagon’ = Munidopsis palmuelleri n. sp.; 1 - Furth; 2 - Fenk quarry at Gross Höflein; 3 - Bad Deutsch-Altenburg; 4 – Budapest area; 5 - Visegrád and Nagymaros; 6 - Grobie; 7 - Węglinek; 8 - Maksymivka; 9 - Cerová-Lieskové; 10 - Košaki.

Hyžný et al. Miocene squat lobsters of the Central Paratethys. Scripta Geol., 147 (2014) 245
Other material – From the Austrian part of the Vienna Basin: six specimens (NHMW 2011/0167/0132-0137), Fenk quarry (Gross Höflein, Lower Austria; for details see Bach-mayer & Tollmann, 1953; Riegl & Piller, 2000; Wiedl et al., 2013); 20 specimens (NHMW 2013/0580/0001-0020), quarry in the Hundsheim Hills (Deutsch-Altenburg, Lower Aus-tria). From the Budapest area, Hungary (for details see Müller, 1984a): numerous speci-mens from the Tétényi Plateau (M.86.21, M.86.191, M.86.507, M.86.831), Diósd (M.89.64), Kerepesi út (M.86.417), keresztúri út (M.86.231), Örs vezér tere (M.86.73) and Rákos (M.86.493). From the Hungarian part of the Danube Basin (for details see Müller, 1984a): several specimens from Visegrád (M.86.342) and Nagymaros-Törökmező (M.86.103).
Measurements – Measurements (only of sufficiently well-preserved specimens) are as follows:NHMW 2011/0167/0134: L=11.8; GH=6.4; TW=9.0; MW=11.8; L/MW=1.0NHMW 2013/0580/0005: L=11.0; GH=5.5; TW=8.0; MW=10.5; L/MW=1.05NHMW 2013/0580/0014: LR=4.9; L=3.6; GH=2.0; TW=2.9; MW=3.6; L/MW=1.0NHMW 2013/0580/0012: LR=5.3; L=4.0; GH=2.2; TW=3.0; MW=3.6; L/MW=1.1NHMW 2013/0580/0016: L=5.3 (estimated value); GH=3.0; TW=4.4; MW=5.4; L/MW=0.99
Description – Carapace subrectangular, slightly longer than wide, excluding rostrum, lateral margins convex, widest in posterior-median part. Rostrum projects slightly downwards, concave at midline, subtriangular, with three pairs of lateral spines, with one pair of supraorbital spines. Rostral surface ornamented with small, irregularly spaced tubercles, no median carina. Orbits with concave margins and one small, ante-riorly directed outer-orbital spine. Lateral margins slightly convex, with seven spines. Posterior margin slightly concave, rimmed. Cervical groove deep; postcervical groove weakly indicated. Carapace surface ornamented with transverse, sinuous and parallel striae. Striae occasionally interrupted. On rostral base two postfrontal raised striae, curved in the median part and interrupted by a short groove. Postfrontal striae orna-mented with several spines pointing forwards. Epigastric regions ornamented with up to four pairs of small spines. Gastric regions with three continuous transverse striae and several interspersed and interrupted striae. Posterior area of the carapace undivided by grooves, ornamented with three or four continuous transverse striae and several inter-rupted ones. Cardiac region weakly marked by interruptions in groove pattern. Sternum slightly longer than broad, maximum width at sternites 6 and 7; sternite 3 narrowest; sternites 4 and 5 narrowing anteriorly; sternites 6 and 7 approximately 2.5 times wider than sternite 3; sternum surface depressed in midline. Abdominal tergites smooth, unarmed.
Remarks – Bachmayer (1950, p. 136) provided a rather vague diagnosis, which is here emended. In view of the fact that the type material is here considered lost, our emendation is based both on the illustrations of the original material (see Pl. 1) and on additional material from Deutsch-Altenburg which Bachmayer himself considered to be conspecific (see Bachmayer, 1953). However, there do appear to be some differences between Bachmayer’s original description and the material examined herein. Bachmay-er (1950) noted that in the type material the carapace was widest anteriorly. Interest-ingly, in all other specimens known, the carapace is widest posteriorly. This discrepancy

246 Hyžný et al. Miocene squat lobsters of the Central Paratethys. Scripta Geol., 147 (2014)
may be explained by diverging styles of preservation; the type material came from marls, while all other material is from carbonates. In the original sketch (Bachmayer, 1950, pl. 1, fig. 2a) the cardiac region is relatively well marked, which is not true for other material. In fact, an indistinct cardiac region is typical of Galathea (see Baba, 2005). However, this feature is less well visible in the photograph (Bachmayer, 1950, pl. 1, fig. 1). In the interest of taxonomic stability we attribute these differences (if they do exist) to intraspecific variation, until the issue can be studied in more detail on the basis of topotypical material, or in case the type lot is traced. Müller (1984a, p. 60) noted that there had already been some doubts on the unity of this particular taxon. It should be noted, however, that carapace ornament remains reasonably constant in all specimens studied. Bachmayer (1953) provided a drawing of the sternum, but failed to add a pho-tograph of the same. Re-examination of this specimen (NHMW 2013/0580/0010) has now shown that Bachmayer’s illustration is slightly inaccurate, in that sternites 5-7 were shown to be more or less parallel (Bachmayer, 1953, pl. 5, fig. 6), while in actual fact they are convergent (Pl. 2, fig. 5). The present species is fairly similar to the extant G. squamifera Leach, 1814 and G. strigosa. However, the former has less prominent postfrontal striae, while these are al-ways well developed in G. weinfurteri. Galathea strigosa has more discontinuous trans-verse striae than G. weinfurteri. Gatt & De Angeli (2010) recorded Galathea sp. from the Upper Miocene of Malta, together with G. weinfurteri. Of the former only a single fragmentary specimen is known; this differs G. weinfurteri in showing a larger number of sinuous secondary striae inter-spersed with primary ones and in having some spines on the hepatic and epibranchial regions (Gatt & De Angeli, 2010, p. 1326). It should be noted that this particular speci-men is significantly larger than co-occurring G. weinfurteri, which suggests that these differences may be ontogenetic. Similarly, at Deutsch-Altenburg larger-sized specimens of G. weinfurteri exhibit more secondary striae than smaller ones (Pl. 2). Another point is that secondary striae are often better visible when lit from different angles. Gatt & De Angeli (2010) also noted the presence of small spines on the hepatic and epibranchial regions in what they named Galathea sp. Although spine position is considered taxo-nomically significant (Poore, 2004), this may vary to a certain degree (e.g., Baba, 2005). In fact, Bachmayer (1950) described G. weinfurteri with more than seven spines in the area of ‘the upper margin of the mesogastriac region’. These may correspond to the spines recorded in Galathea sp. by Gatt & De Angeli (2010). Similarly, Müller (1984a, p. 60) pointed out that, ‘behind the rostrum there is a spiny ridge but the spines are very small or even rudimentary on some specimens’. Bachmayer (1950) and Müller (1984a) also described isolated propodi which they at-tributed to G. weinfurteri. These are stout and striated as is typical of members of the ge-nus. However, it should be noted that they have never been found directly associated with carapaces, although the occur at localities where G. weinfurteri is the sole species of squat lobster on record. Specimens with ‘bopyriform’ swellings have also been recorded; Müller (1984, pl. 22, fig. 5) mentioned a specimen with a swelling on the left branchial area, whereas the one illustrated herein (Pl. 2, fig. 3) has a swelling on the right-hand side.
Occurrence – This species is known from the Middle Miocene (‘Badenian’; Lang-hian) of Austria (Bachmayer, 1950, 1953), Hungary (Müller, 1984a), Poland (Müller,

Hyžný et al. Miocene squat lobsters of the Central Paratethys. Scripta Geol., 147 (2014) 247
1996; Górka, 2002), Ukraine (Radwański et al., 2006; Ossó & Stalennuy, 2011) and the Upper Miocene of Malta (Gatt & De Angeli, 2010).
Family Munididae Ahyong, Baba, Macpherson & Poore, 2010Genus Agononida Baba & de Saint Laurent, 1996
Type species – Munida incerta Henderson, 1888, by original designation.
Diagnosis – See Baba & de Saint Laurent (1996, p. 441).
Remarks – To date, there is only a single fossil record of this genus (Hyžný & Schlögl, 2011). Agononida is closely similar to Munida Leach, 1820, but differs in the constant pos-session of pairs of epigastric and postcervical spines (Baba & de Saint Laurent, 1996; Poore, 2004). It should be noted that spines are very delicate structures and in the sole fossil member known, Agononida cerovensis Hyžný & Schlögl, 2011, they were identified only on the basis of the counterpart of the fossil, whereas the latter shows only large swellings or tubercles (Hyžný & Schlögl, 2011, text-fig. 6A). Fossil crustaceans often oc-cur in fragments, which holds true in particular for material preserved in carbonates; this might, in part, explain the paucity of the genus in the fossil record. In contrast, ex-tinct members of a close relative, Munida, are well known from different parts of the world (Schweitzer & Feldmann, 2000; Robins et al., 2012b).
Agononida cerovensis Hyžný & Schlögl, 2011Figs. 2, 7B; Pl. 3.
2011 Agononida cerovensis Hyžný & Schlögl, p. 332, text-figs. 6, 7.2011 Agononida cerovensis; Ahyong et al., p. 96.
Emended diagnosis – Carapace cordate, nearly as long as wide, with convexly round-ed lateral margins; transverse carapace ridges (striae) distinct, anterior of ridges orna-mented with granules, ridges subparallel, becoming less so in anteriormost and poste-riormost portions of carapace; anterolateral margin with three spines; cervical and postcervical grooves deeply incised; 4 epigastric spines, 6 postcervical spines and spines on the posterior rim form two distinct rows oriented longitudinally.
Holotype – Carapace without rostrum (SNM Z-24.864); L = 6.8; GH = 3.6; TW = 6.0; MW = 7.0; L/MW = 0.97.
Remarks – Hyžný & Schlögl (2011) interpreted the four tubercles in the anterior por-tion of the carapace to be bases of two supraorbital and two epigastric spines. A re-ex-amination of the holotype has now shown that all of them are in fact bases of epigastric spines. In the illustration (see Fig. 6B), the rostrum and supraorbital spines are conjec-tural, being based on comparisons with extant members of the genus. This species is based upon a single individual; in spite of numerous visits to the type locality during the last two years, no additional material has so far come to light.
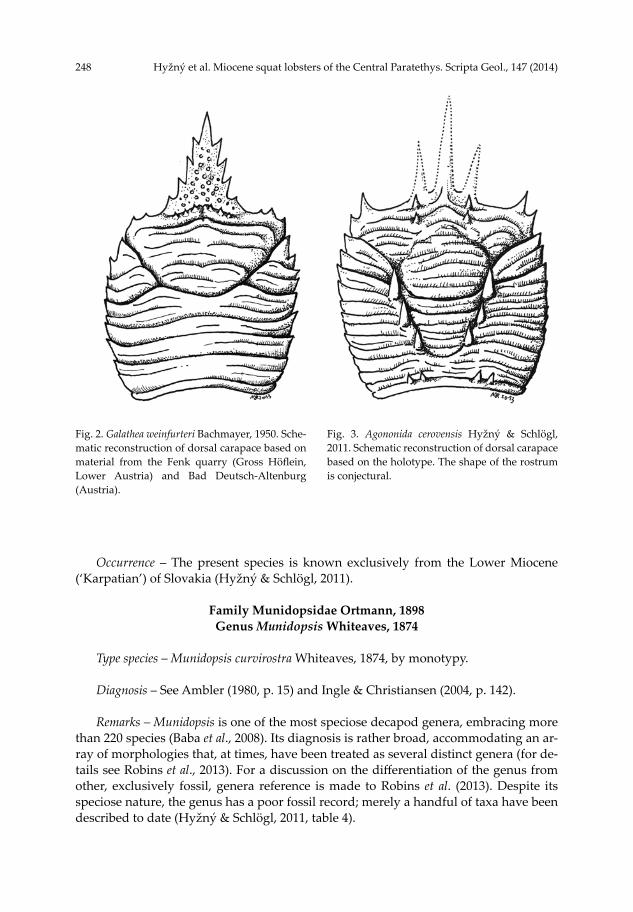
248 Hyžný et al. Miocene squat lobsters of the Central Paratethys. Scripta Geol., 147 (2014)
Occurrence – The present species is known exclusively from the Lower Miocene (‘Karpatian’) of Slovakia (Hyžný & Schlögl, 2011).
Family Munidopsidae Ortmann, 1898Genus Munidopsis Whiteaves, 1874
Type species – Munidopsis curvirostra Whiteaves, 1874, by monotypy.
Diagnosis – See Ambler (1980, p. 15) and Ingle & Christiansen (2004, p. 142).
Remarks – Munidopsis is one of the most speciose decapod genera, embracing more than 220 species (Baba et al., 2008). Its diagnosis is rather broad, accommodating an ar-ray of morphologies that, at times, have been treated as several distinct genera (for de-tails see Robins et al., 2013). For a discussion on the differentiation of the genus from other, exclusively fossil, genera reference is made to Robins et al. (2013). Despite its speciose nature, the genus has a poor fossil record; merely a handful of taxa have been described to date (Hyžný & Schlögl, 2011, table 4).
Fig. 2. Galathea weinfurteri Bachmayer, 1950. Sche-matic reconstruction of dorsal carapace based on material from the Fenk quarry (Gross Höflein, Lower Austria) and Bad Deutsch-Altenburg (Austria).
Fig. 3. Agononida cerovensis Hyžný & Schlögl, 2011. Schematic reconstruction of dorsal carapace based on the holotype. The shape of the rostrum is conjectural.

Hyžný et al. Miocene squat lobsters of the Central Paratethys. Scripta Geol., 147 (2014) 249
Munidopsis lieskovensis Hyžný & Schlögl, 2011Figs. 3, 7C; Pl. 4.
2011 Munidopsis lieskovensis Hyžný & Schlögl, p. 334, text-figs. 8, 9.2011 Munidopsis lieskovensis; Ahyong et al., p. 96.2013 Munidopsis lieskovensis; Robins et al., table 9.
Diagnosis – Carapace subsquare; moderately convex transversely. Rostrum styli-form, keeled, unidentate. Anterolateral margin with one spine at anterior of hepatic region. Circumgastric groove strong; V-shaped in posteriormost part. Regions slightly inflated. Epigastric regions circular in outline. Urogastric region extending across width of carapace; twice as wide as cardiac region. Cardiac region triangular, apex directed posteriorly. Carapace uniformly ornamented with short, transverse, squamous ridges.
Type material – The holotype is a near-complete dorsal carapace with the rostrum (SNM Z-24.865); both paratypes are near-complete dorsal carapaces, lacking the ros-trum (SNM Z-24.866 and SNM Z-24.867).
Other material – Six near-complete dorsal carapaces (KGP-MH CL-002-007) and a single sternum (KGP-MH CL-008).
Measurements – Measurements were taken only from sufficiently well-preserved specimens:SNM Z-24.865 (holotype): GH=2.6; TW=5.1; MW=5.7SNM Z-24.866 (paratype): GH=2.3; TW=4.8; MW=5.0SNM Z-24.867 (paratype): GH=2.6; TW=4.7; MW=5.1KGP-MH CL-002: LR=6.1; L=4.3; GH=2.0; TW=3.8; MW=4.3; L/MW=1KGP-MH CL-003: LR=5.1; L=3.7; GH=1.8; TW=3.6 (reconstructed); MW=3.6; L/MW=1KGP-MH CL-004: GH=1.9; TW=3.8; MW=4.1KGP-MH CL-005: GH=1.6; TW=2.9; MW=3.4KGP-MH CL-006: L=3.4; GH=1.7; TW=2.8 (estimated); MW=3.4; L/MW=1KGP-MH CL-007: LR=6.6 (estimated); L=4.4; GH=2.1; TW=3.4; MW=4 (estimated); L/MW=1.1
Emended description – Carapace subsquare (L/MW 1-1.1); moderately convex trans-versely. Rostrum styliform, keeled, unidentate, bearing pustular ornamentation. Ante-rior margin sinuous. Lateral margins slightly convex, with two spines. Circumgastric groove strong; V-shaped in posteriormost part. Branchiocardiac groove strongest ap-proaching lateral margins. Epigastric, hepatic, epibranchial, urogastric and cardiac re-gions well defined. Regions slightly inflated. Hepatic region quadrate with one spine at its anterior; epigastric regions circular and inflated. Urogastric region extending across width of carapace; twice as wide as cardiac region. Cardiac region triangular, apex di-rected posteriorly. Carapace uniformly ornamented with short, transverse, squamous ridges. Posterior margin inflected concave posterior, strongly rimmed. Ornamentation does not continue onto rim. Sternum: sternites 4 and 5 narrowing anteriorly; sternite 5 approximately 1.5 times wider than sternite 4; sternum surface depressed in midline.

250 Hyžný et al. Miocene squat lobsters of the Central Paratethys. Scripta Geol., 147 (2014)
Remarks – The type lot comprises three incomplete specimens. Recently, new mate-rial has been recovered from the type locality, Cerová-Lieskové, including numerous complete specimens (Pl. 4) that allows the original description to be emended in terms of rostral length and details of posterior margin and sternum.
Occurrence – The species is known exclusively from the Lower Miocene (‘Karpa-tian’) of Slovakia (Hyžný & Schlögl, 2011).
Munidopsis palmuelleri n. sp.Figs. 5, 7D; Pl. 5.
Diagnosis – Carapace with well-defined regions; rostrum styliform, unkeeled, uni-dentate, flat in cross section; carapace surface covered with short, parallel striae, striae forming longer ridges in mesogastric region and posterior part of carapace.
Etymology – The specific name honours Pál Müller, who greatly improved our knowledge of Miocene decapod crustaceans of the Central Paratethys.
Holotype – The holotype (RGA/SMNH 0351) is an imprint of a complete carapace; measurements: LR=8.9; L=6.6; GH=3.5; TW=5.3; MW=5.6; L/MW=1.18.
Description – Carapace subrectangu-lar, slightly longer than wide, L/W 1.18; moderately convex transversely. Ros-trum styliform, unkeeled, unidentate, flat in cross section. Anterolateral mar-gin with small, anteriorly directed spine. Lateral margins subparallel. Circumgas-tric groove strong; V-shaped in posteri-ormost part. Branchiocardiac groove strongest approaching lateral margins. Hepatic, epibranchial, urogastric and cardiac regions well defined. Epigastric regions poorly defined. Urogastric re-gion extending across middle half of carapace; wider than cardiac region. Cardiac region strongly defined anteri-orly, less defined posteriorly; triangular in shape with apex directed posteriorly. Carapace ornamented with narrow, short, sinuous transverse ridges anteri-orly; ridges broaden posteriorly. Posteri-or margin inflected concave posterior, weakly rimmed. Ornamentation not continuing onto rim. Ventral surface and appendages not preserved.
Fig. 4. Munidopsis lieskovensis Hyžný & Schlögl, 2011. Schematic reconstruction of dorsal carapace based on newly collected material (presented here-in).

Hyžný et al. Miocene squat lobsters of the Central Paratethys. Scripta Geol., 147 (2014) 251
Remarks – Although the species is based on a single specimen, it differs significantly from all other extinct congeners. It can be differentiated from M. scabrosa Feldmann & Wilson, 1988 and M. foersteri Feldmann, Tshudy & Thompson, 1993 by the shape of the circumgastric groove. The base of the circumgastric groove of M. palmuelleri n. sp. is V-shaped, while it is rounded in the other two. Moreover, M. foersteri has a pyriform cara-pace and the rostrum is nearly as long as the carapace (Feldmann et al., 1993), quite unlike M. palmuelleri n. sp. Munidopsis scabrosa has a subsquare carapace with forward-ly directed scabrose ornamentation and a keeled rostrum (Feldmann & Wilson, 1988). The closest morphological match with M. palmuelleri n. sp. is M. lieskovensis (see above), but the former can be differentiated on ornamentation; that of M. palmuelleri n. sp. is much more ridge like than the squamous sculpture in M. lieskovensis. The rostrum in
Fig. 5. The Košaki section, Slo-venia; type locality of Munidop-sis palmuelleri n. sp.; A. Schemat-ic lithological section; B. Geo-graphical position.

252 Hyžný et al. Miocene squat lobsters of the Central Paratethys. Scripta Geol., 147 (2014)
the latter also has a different shape, while the lateral carapace margins are near parallel at the base, whereas in M. palmuelleri n. sp. they are strongly con-vergent, yielding a more triangular cara-pace shape. Moreover, unlike M. liesko-vensis, the new species has a rostrum without a keel and the epigastric regions are poorly defined.
Occurrence – The holotype and sole specimen known to date (RGA/SMNH 0351) was recovered from monotonous grey marls at a construction site in the now abandoned Košaki brickyard. The locality is situated approximately 2 km northeast of the city of Maribor in north-eastern Slovenia (GPS co-ordinates: 46°35’27.62”N, 15°39’56.25”E; see Fig. 5). The Košaki section consists of a rather uniform succession of poorly fossilifer-ous (macrofossil) grey clayey marl beds of the Slovenske Gorice Formation which is the lithological equivalent of the Kreuzkrumpel Formation (Steirischer Schlier) exposed in the Austrian part of the Styrian Basin (Gross et al., 2007). Near the top of the 15-m-thick section the carbonate content of the marl in-creases, the marl becoming much harder and lamellate. The sequence is here interrupted by several thin (up to 10 cm) sandstone interbeds. The carbonate fraction of ‘lower Badenian’ marls of the Slovenske Gorice Formation consists mainly of nannofossils, whereas clastic components are predominantly clay and silt (Bartol, 2009). The marls usually contain only rare macrofossils, but are rich in calcareous nannoplankton. Apart of the present specimen, we are not aware of any ad-ditional decapod remains from this locality. An enrichment of thin-shelled pectinid bi-valves (Amusium cristatum) was observed in the same layer, along with rare schizasterid echinoids (RG, pers. obs.). Bartol (2009) recorded a nannoplankton assemblage from a several sampling sites near the Košaki section within the same lithological unit; this defines the interval zone MuN5a sensu Martini (1971). This interval was correlated with the lower part of NN5 sensu Martini (1971) and probably corresponds to the transition between the lower and middle ‘Badenian’ (Rögl et al., 2002). Thus, Munidopsis palmuelleri n. sp. is of Middle Miocene age.
Fig. 6. Munidopsis palmuelleri n. sp. Schematic recon-struction of dorsal carapace based on the holotype.

Hyžný et al. Miocene squat lobsters of the Central Paratethys. Scripta Geol., 147 (2014) 253
Galatheoidea genus and species indeterminatePl. 6.
Material – A single specimen (NHMW 2013/0607/0001), consisting of four articulat-ed pereiopods, without dorsal carapace, sternum or pleon.
Description – Pereiopod 1 (cheliped), with propodus twice as long as dactylus, car-pus short, merus approximately 2.5 times length of carpus; all elements ornamented with rows of prominent spines (Pl. 6, fig. 2); occlusal surface of fingers armed with tiny teeth (Pl. 6, fig. 3). Pereiopods 2 and 3(?) long and slender, no details discernible.
Remarks – Unfortunately, a family-level identification of this material is impossible in the absence of a dorsal carapace. The preservation of appendages of galatheoids in the fossil record is rare, which also means that comparative material is scant. Modern galatheoids are quite variable in appendage size and structure, even within genera. Munidids tend to have long chelae (Ahyong et al., 2010), which would suggest that the present specimen is not a member of this family; however, this is inconclusive.
Occurrence – Kummer quarry at Sankt Margarethen, Burgenland, Austria (Eisen-stadt-Sopron Basin). A detailed sedimentological description and facies analysis of the strata exposed here was provided by Schmid et al. (2001). The age is based on calcareous nannoplankton, which indicates zone NN5b sensu Martini (1971), i.e., Middle Miocene, around the Langhian/Serravallian boundary (late ‘Badenian’).
Notes on palaeoenvironment
Squat lobsters inhabit a wide range of habitats, ranging from the intertidal zone to hydrothermal vents, and are typically members of benthic communities (Baba, 2005; Baba et al., 2008; Lowrich & Thiel, 2011). Occasionally, they occur in dense aggrega-tions, which is especially true for taxa that live on the continental shelf and often are
Fig. 7. Miocene squat lobsters of the Central Paratethys; reconstructions (not to scale) of dorsal cara-paces of Galathea weinfurteri (A), Agononida cerovensis (B), Munidopsis lieskovensis (C) and M. palmuelleri n. sp. (D). The shape of the rostrum in A. cerovensis is based on extant congeners.

254 Hyžný et al. Miocene squat lobsters of the Central Paratethys. Scripta Geol., 147 (2014)
associated with other species of animal (for a review see Lowrich & Thiel, 2011; refer-ences therein). Squat lobsters commonly inhabit coral reefs, rocky or muddy bottoms (Baba et al., 2008; Lowrich & Thiel, 2011); our fossil material from the Central Paratethys is no exception in that respect (Table 2).
Galathea weinfurteri – This is the most widely distributed squat lobster in the Mio-cene of the Central Paratethys known to date. Extant species of Galathea usually inhabit shallow waters (Baba et al., 2008); indeed, most material of G. weinfurteri comes from shallow-marine reefal facies (e.g., Bachmayer & Tollmann, 1953; Müller, 1984a, 1996; Radwański et al., 2006) and specimens from Deutsch-Altenburg are confined to bryo-zoan marls and calcarenites with typical reefal decapods (Bachmayer, 1953). Interest-ingly, the type collection came from a different facies (marls; see above), and differs slightly from other occurrences of this species. Some squat lobster species may exhibit rather broad ecological preferences, and assuming that all reported occurrences of G. weinfurteri belong to the same species, this indicates that G. weinfurteri followed such a pattern. An alternative interpretation would be that the type specimens of G. weinfurteri are not conspecific with other specimens assigned to that species, as discussed above. Studies by Nicol (1932) and Pike (1947) demonstrated that the mouth parts of Galathea are ill-adapted for life on muddy substrates. For instance, G. squamifera and G. strigosa, extant relatives of G. weinfurteri, are restricted to rocky areas (De Grave & Turner, 1997).
Agononida cerovensis – The species is known from a single locality, the sedimentary setting of which has been interpreted to represent bathyal settings of depths between 240 and 330 m (Schlögl et al., 2011). Today, the genus Agononida is considered to be a typical deep-water inhabitant (Poore, 2004). Baba et al. (2008) stated that it inhabited most typically environments shallower than 300 m. Because A. cerovensis is the sole fos-sil member of the genus known, its ecological preferences cannot be compared with any extinct relative.
Munidopsis lieskovensis – The species has been recorded only from its type locality (Cerová Lieskové), but, unlike A. cerovensis from the same setting, it is known from numerous specimens. Munidopsis is generally considered a deep-water taxon, com-monly inhabiting the continental slope, often below 500 m (Ambler, 1980; Baba et al., 2008; Osawa et al., 2008; Macpherson, 2011). The palaeobathymetry at Cerová-Liesko-vé has been estimated to have been 240-330 m, thus confirming the bathymetric pref-erence of the genus also in Miocene times (see also Takeda et al., 1986). Late Creta-ceous (Feldmann et al., 1993) and Eocene (Feldmann & Wilson, 1988) occurrences of Munidopsis, however, are known from shallow-marine settings, implying different
Table 2. Palaeoenvironment of Miocene squat lobsters from the Central Paratethys.
Taxon Substrate PalaeoenvironmentGalathea weinfurteri carbonate, siliciclastic reefal, non-reefal (shallow-water)Agononida cerovensis Siliciclastic muddy bottom (deep-water)Munidopsis lieskovensis Siliciclastic muddy bottom (deep-water)Munidopsis palmuelleri n. sp. Siliciclastic muddy bottom (deep-water)

Hyžný et al. Miocene squat lobsters of the Central Paratethys. Scripta Geol., 147 (2014) 255
environmental preferences. A scant fossil record does not allow further speculations on this issue.
Munidopsis palmuelleri n. sp. – The type locality of this new species is in the Mari-bor subbasin of the Mura Depression. The time of deposition of the Slovenske Gorice marls was characterised by the ‘early Badenian’ transgression from the Mediterrane-an through the Slovenian corridor (Pavšič & Horvat, 2009). The extensional tectonics at that time were intense and subsidence very rapid, which resulted in basins exceed-ing 500 m in depth (Mioč & Žnidarčič, 1989). The highly diverse nannoplankton as-semblage in the marls with M. palmuelleri n. sp. is consistent with an offshore deposi-tional environment (Bartol, 2009); foraminifera from the same strata (Rijavec, 1976) and planktonic molluscs such as Clio pedemontana (Mayer, 1868) from similar layers of the nearby locality of Polički Vrh (Mikuž et al., 2012) corroborate the bathymetrical setting inferred.
Acknowledgements
We wish to thank Gerhard Wanzenböck (Bad Vöslau, Austria) for providing an in-determinate squat lobster specimen for the present study. Reinhardt Roetzel (Geologis-che Bundesanstalt, Wien, Austria) helped with data on the locality Furth. Michał Kro-bicki (AGH University of Science and Technology, Kraków, Poland) and Karen E. Schna bel (National Institute of Water and Atmospheric Research, Wellington, New Zealand) are thanked for careful reviews. MH’s research has been supported by Aus-trian Science Fund (FWF; Lise Meitner Program M 1544-B25). This work was supported by the Slovak Research and Development Agency under contracts no. APVV-0640-10 and APVV-0436-12.
References
Ahyong, S.H., Baba, K., Macpherson, E. & Poore, G.C. 2010. A new classification of the Galatheoidea (Crustacea: Decapoda: Anomura). Zootaxa, 2676: 57-68.
Ahyong, S.H., Schnabel, K.E. & Macpherson, E. 2011. Phylogeny and fossil record of marine squat lob-sters. In: Poore, G.C.B., Ahyong, S.T. & Taylor, J. (eds.), The biology of squat lobsters: 73-104. CSIRO Publishing, Melbourne.
Ambler, J.W. 1980. Species of Munidopsis (Crustacea, Galatheidae) occurring off Oregon and in adjacent waters. Fishery Bulletin, 78: 13-34.
Baba, K. 2005. Deep-sea chirostylid and galatheid crustaceans (Decapoda: Anomura) from the Indo-Pacific, with a list of species. Galathea Report, 20: 1-317.
Baba, K., Macpherson, E., Poore, G.C.B., Ahyong, S.T., Bermudez, A., Cabezas, P. & Lin, Ch.-W. 2008. Catalogue of squat lobsters of the world (Crustacea: Decapoda: Anomura – families Chirostylidae, Galatheidae and Kiwaidae). Zootaxa, 1905: 1-220.
Baba, K. & Saint Laurent, M. de. 1996. Crustacea Decapoda: Revision of the genus Bathymunida Balss, 1914, and description of six new related genera (Galatheidae). In: Crosnier, A. (ed.), Résultats des Campagnes MUSORSTOM, Volume 15. Mémoires du Muséum national d’Histoire naturelle, 168: 433-502.
Bachmayer, F. 1950. Neue Dekapoden aus dem österreichischen Tertiär. Annalen des Naturhistorischen Museums in Wien, 57: 133-140.
Bachmayer, F. 1953. Die Dekapodenfauna des tortonischen Leithakalkes von Deutsch-Altenburg (Nied-erösterreich). Mitteilungen der Geologischen Gesellschaft in Wien, 44 (for 1951): 237-262.

256 Hyžný et al. Miocene squat lobsters of the Central Paratethys. Scripta Geol., 147 (2014)
Bachmayer, F. & Tollmann A. 1953. Die Crustaceen-Fauna aus dem tortonischen Leithakalk (Steinbrüche der Firma Fenk) bei Groß-Höflein im Burgenland. Kober-Festschrift 1953, Skizzen zum Antlitz der Erde. Geologische Arbeiten, herausgegeben aus Anlaβ des 70. Geburtstages von Prof. Dr. L. Kober, Univer-sität Wien: 308-314. Gesellschaftsbuchdruckerei Brüder Hollinek, Wien.
Bartol, M. 2009. Middle Miocene calcareous nannoplankton of NE Slovenia (Western Central Paratethys). Paleontološki institute Ivana Rakovca ZRC SAZU: 1-136.
De Angeli, A. & Garassino, A. 2002. Galatheid, chirostylid and porcellanid decapods (Crustacea, De-capoda, Anomura) from the Eocene and Oligocene of Vicenza (N Italy). Memorie della Società italiana di Scienze naturali e del Museo civico di Storia naturale di Milano, 30: 3-31.
De Grave, S. & Turner, J.R. 1997. Activity rhythms of the squat lobsters, Galathea squamifera and G. strigosa (Crustacea: Decapoda: Anomura) in south-west Ireland. Journal of the Marine Biological As-sociation of the United Kingdom, 77: 273-276.
Feldmann, R.M., Tshudy, D.M. & Thomson, M.R.A. 1993. Late Cretaceous and Paleocene decapod crus-taceans from James Ross Basin, Antarctic Peninsula. The Paleontological Society Memoir, 28: 1-41.
Feldmann, R.M. & Wilson, M.T. 1988. Eocene decapod crustaceans from Antarctica. In: Feldmann, R.M. & Woodburne, M.O. (eds.), Geology and paleontology of Seymour Island, Antarctic Peninsula. Geo-logical Society of America Memoir, 169: 465-488.
Fabricius, J.C. 1793. Entomologia systematica emendata et aucta. Secundum classes, ordines, genera, species adjectis synonymis, locis, observationibus, descriptionibus, 2. Proft et Storch, Hafniae: 1-519.
Garassino, A., De Angeli, A. & Pasini, G. 2008. New decapod assemblage from the Upper Cretaceous (Cenomanian) of southeastern Morocco. Atti della Società italiana di Scienze naturali e del Museo civico di Storia naturale in Milano, 149: 37-67.
Gatt, M. & De Angeli, A. 2010. A new coral-associated decapod assemblage from the Upper Miocene (Messinian) upper Coralline Limestone of Malta (Central Mediterranean). Palaeontology, 53: 1315-1348.
Glaessner, M.F. 1928. Die Dekapodenfauna des österreichischen Jungtertiärs. Jahrbuch der Geologischen Bundesanstalt Wien, 78: 161-219.
Górka, M. 2002. The Lower Badenian (Middle Miocene) coral patch reef at Grobie (southern slopes of the Holy Cross Mountains, Central Poland), its origin, development and demise. Acta Geologica Polonica, 53: 201-234.
Gross, M., Fritz, I., Piller, W.E., Soliman, A., Harzhauser, M., Hubmann, B., Moser, B., Scholger, R., Suttner, T.J. & Bojar, H.-P. 2007. The Neogene of the Styrian Basin - guide to excursions. Joannea, Geologie und Paläontologie, 9: 117-193.
Harzhauser, M. & Piller, W.E. 2007. Benchmark data of a changing sea – palaeogeography, palaeobioge-ography and events in the Central Paratethys during the Miocene. Palaeogeography, Palaeoclimatolo-gy, Palaeoecology, 253: 8-31.
Harzhauser, M., Mandic, O. & Zuschin, M. 2003. Changes in Paratethyan marine molluscs at the Early/Middle Miocene transition: diversity, palaeogeography and palaeoclimate. Acta Geologica Polonica, 53: 323-339.
Harzhauser, M., Kroh, A., Mandic, O., Piller, W.E., Göhlich, U., Reuter, M. & Berning, B. 2007. Biogeo-graphic responses to geodynamics: a key study all around the Oligo-Miocene Tethyan Seaway. Zo-ologischer Anzeiger, 246: 241-256.
Haworth, A.H. 1825. A new binary arrangement of the macrurous Crustacea. The Philosophical Magazine and Journal, 65: 183-184.
Henderson, J.R. 1888. Report on the Anomura collected by H.M.S Challenger during the years 1873-1876. In: Murray, J. (ed.), Zoology. Report on the scientific results of the voyage of H.M.S. Challenger dur-ing the years 1873-76 under the command of Captain George S. Nares, R.N., F.R.S. and the late Captain Frank Tourle Thomson, R.N. Wyville Thomson, C. and J. Murray (series eds.), Vol. 27. Neill and Company, Edinburgh: xi+221 pp.
Hyžný, M. & Schlögl, J. 2011. An early Miocene deep-water decapod crustacean faunule from the Vienna Basin (Western Carpathians, Slovakia). Palaeontology, 54: 323-349.
Ingle, R.W. & Christiansen, M.E. 2004. Lobsters, mud shrimps and anomuran crabs. Keys and notes for the identification of species. In: Crothers, J.H. & Hayword, P.J. (eds.), Synopses of the British Fauna,

Hyžný et al. Miocene squat lobsters of the Central Paratethys. Scripta Geol., 147 (2014) 257
new series, 55: 1-271. Field Studies Council for Linnaean Society of London Estuarine and Coastal Sciences Association, Shrewsbury.
Klompmaker, A.A., Feldmann, R.M., Robins, C.M. & Schweitzer, C.E. 2012. Peak diversity of Cretaceous galatheoids (Crustacea, Decapoda) from northern Spain. Cretaceous Research, 36: 125-145.
Latreille, P.A. 1802. Histoire naturelle, générale et particulière des Crustacés et des Insectes. Ouvrage faisant suite aux Œuvres de Leclerc de Buffon, et partie du Cours complet d’Histoire naturelle rédigé par C.S. Son-nini, membre de plusieurs Sociétés savantes. Familles naturelles des genres, 3: 1-467. F. Dufart, Paris.
Latreille, P.A. 1810. Considerations générales sur l’ordre naturel des animaux composant les classes des Crus-tacés, des Arachnidés, et des Insectes; avec un tableau méthodique de leur genres, disposés en familles. F. Dufart, Paris: 444 pp.
Leach, W.E. 1814. Crustaceology. Brewster’s Edinburgh Encyclopedia, 7: 383-437.Leach, W.E. 1820. Galatéadées, Galateadæ. (Crust.). In: Cuvier, F. (ed.), Dictionnaire des Sciences Naturelles,
dans lequel on trait Méthodiquement des Différens étres de la Nature, considérés soit en eux-mêmes, d’après l’état actuel de nos connoissances, soit relativement a l’utilité qu’en peuvent retirer la Médecine, l’Agriculture, le Commerce et les Arts. Suivi d’une biographie des plus Célèbres Naturalistes. Ouvrage destiné aux mé-decins, aux agriculteurs, aux commerçans, aux artistes, aux manufacturiers, et à tous ceux qui ont intérêt à connoître les productions de la nature, leurs caractères génériques et spécifiques, leur lieu natal, leurs propié-tés et leurs usages, 18: 49-56. F.G. Levrault/ Le Normant, Strasbourg/Paris.
Linnaeus, C. 1761. Fauna Suecica sistens Animalia Sueciae Regni: Mammalia, Aves, Amphibia, Pisces, Insecta, Vermes. Distributa per Classes, Ordines, Genera, Species, cum Differentiis Specierum, Synonymis Aucto-rum, Nominibus Incolarum, Locis Natalium, Descriptionibus Insectorum. Laurentius Salvius, Stockhol-miae: 578 pp.
Lowrich, G.A. & Thiel, M. 2011. Ecology, physiology, feeding and trophic role of squat lobsters. In: Poore, G.C.B., Ahyong, S.T. & Taylor, J. (eds.), The biology of squat lobsters: 183-222. CSIRO Publish-ing, Melbourne.
MacLeay, W.S. 1838. On the brachyurous decapod Crustacea brought of the Annulosa of South Africa. In: Smith, A., Illustrations of the Annulosa of South Africa: being a portion of the objects of natural history chiefly collected during an expedition into the interior of South Africa, under the direction of Dr. Andrew Smith in the years 1834, 1835, and 1836; fitted out by “The Cape of Good Hope Association for Exploring Central Africa”: 53-71 Smith, Elder and Co., London.
Macpherson, E. 2011. A new squat lobster of the genus Munidopsis (Crustacea: Decapoda: Munidopsi-dae) from the Mediterranean Sea. Scientia Marina, 75: 525-532.
Martini, E. 1971. Standard Tertiary and Quaternary calcareous nanoplankton zonation. In: Farinacci, A. (ed.), Proceedings of the Second Plankton Conference, Rome, 1970: 739-785. Tecnoscienza, Roma.
Mayer, C. 1868. Description de coquilles fossiles des terrains tertiaires supérieurs (suite). Journal de Con-chyliologie (3)8(16): 102-112.
Mikuž, V., Gašparič, R., Bartol, M., Horvat, A. & Pavšič, J. 2012. Miocene pteropods from Polički Vrh in Slovenske gorice, northeast Slovenia. Geologija, 55: 67-76. [in Slovenian, with English abstract and summary].
Mioč, P. & Žnidarčič, M. 1989. Tolmač za lista Maribor in Leibnitz. Osnovna geološka karta SFRJ 1:100.000. Zvezni geološki zavod Beograd, Beograd: 60 pp.
Müller, P. 1974a. Decapoda (Crustacea) fauna a budapesti miocénből (1). Földtani Közlöny, 102: 119-132.Müller, P. 1974b. Decapoda (Crustacea) fauna a budapesti miocénből (2). Földtani Közlöny, 102: 275-287.Müller, P. 1976. Decapoda (Crustacea) fauna a budapesti miocénből (3). Földtani Közlöny, 105: 506-517.Müller, P. 1979. Decapoda (Crustacea) fauna a budapesti miocénből (5). Földtani Közlöny, 108: 272-312.Müller, P. 1984a. Decapod Crustacea of the Badenian. Geologica Hungarica, Series Palaeontologica, 42:
3-317.Müller, P. 1984b. Messinian and older decapods from the Mediterranean with description of two new
species. Annales Géologiques des Pays Helléniques, 32: 25-34.Müller, P. 1996. Middle Miocene decapod Crustacea from southern Poland. Prace Muzeum Ziemi, 43:
3-16.

258 Hyžný et al. Miocene squat lobsters of the Central Paratethys. Scripta Geol., 147 (2014)
Müller, P. 1998. Crustacea Decapoda. In: Flügel, H.W. (ed.), Catalogus Fossilium Austriae: 1-48. Verlag der Österreichischen Akademie der Wissenschaften, Wien.
Nicol, E.A.T. 1932. The feeding habits of the Galatheidea. Journal of the Marine Biological Association of the United Kingdom, 18: 87-106.
Ortmann, A.E. 1898. Gliedlerfüssler: Arthropoda. In: Gerstäcker, A. & Ortmann, A.E. (eds.), Dr. H.G. Bronn’s Klassen und Ordnungen des Thier-Reichs, wissenschaftlich dargestellt in Wort und Bild, Bd. V, Abt. 2 (Crustacea), 2 Hälfte (Malacostraca): 1057-1168.
Osawa, M., Lin, C.W. & Chan, T.Y. 2008. Species of Galacantha and Munidopsis (Crustacea: Decapoda: Anomura: Galatheidae) from the deep-waters off Taiwan, with the description of two new species. Scientia Marina, 72: 35-57.
Ossó, A. & Stalennuy, O. 2011. Description of the first fossil species of Bathynectes (Brachyura, Polybii-dae) in the Badenian (middle Miocene) of the Medobory Hills (Ukraine, Central Paratethys), with remarks on its habitat ecology. Treballs Museu Geologia Barcelona, 18: 37-46.
Papp, A. 1952. Zur Kenntnis des Jungtertiärs in der Umgebung von Krems a.d. Donau (N.-Ö). Verhand-lungen der geologischen Bundesanstalt Wien, 1952: 49-53.
Pavšič, J. & Horvat, A. 2009. Eocene, Oligocene and Miocene in Central and Eastern Slovenia. In: Pleničar, M. (ed.), The geology of Slovenia: 373-426, Geološki zavod Slovenije.
Pike, R.B. 1947. L.M.B.C. Memoirs No. 34. Galathea. Proceedings and Transactions of the Liverpool Biological Society, 55: 1-179.
Piller, W.E., Harzhauser, M. & Mandic, O. 2007. Miocene Central Paratethys stratigraphy – current sta-tus and future directions. Stratigraphy, 4: 71-88.
Poore, G.C.B. 2004. Marine decapod Crustacea of southern Australia. A guide to identification with chapter on Stomatopoda by Shane Ahyong. CSIRO Publishing, Melbourne: 574 pp.
Popov, S.V., Rögl, F., Rozanov, A.Y., Steininger, F.F., Shcherba, I.G. & Kováč, M. (eds.) 2004. Lithological-paleogeographic maps of Paratethys. Courier Forschungsinstitut Senckenberg, 250: 1-46.
Radwański, A., Górka, M. & Wysocka, A. 2006. Middle Miocene coralgal facies at Maksymivka near Ternopil (Ukraine): A preliminary account. Acta Geologica Polonica, 56: 89-103.
Riegl, B. & Piller, W.E. 2000. Biostromal coral facies – a Miocene example from the Leitha Limestone (Austria) and its actualistic interpretation. Palaios, 15: 399-413.
Rijavec, L. 1976. Biostratigrafija miocena v Slovenskih goricah. Geologija, 19: 53-79.Robins, C.M., Feldmann, R.M. & Schweitzer, C.E. 2012a. The oldest Munididae (Decapoda: Anomura:
Galatheoidea) from Ernstbrunn, Austria (Tithonian). Annalen des Naturhistorischen Museums in Wien, 114A: 289-300.
Robins, C.M., Feldmann, R.M. & Schweitzer, C.E. 2012b. Clarification of the South American genus Austromunida Schweitzer & Feldmann, 2000 (Crustacea: Decapoda: Anomura), with the establish-ment of a new species. Annalen des Naturhistorischen Museums in Wien, 114A: 301-306.
Robins, C.M., Feldmann, R.M. & Schweitzer, C.E. 2013. Nine new genera and 24 new species of the Munidopsidae (Decapoda: Anomura: Galatheoidea) from the Jurassic Ernstbrunn Limestone of Austria, and notes on fossil munidopsid classification. Annalen des Naturhistorischen Museums in Wien, 115A: 167-251.
Rögl, F. 1998. Palaeogeographic considerations for Mediterranean and Paratethys seaways (Oligocene to Miocene). Annalen des Naturhistorischen Museums in Wien, 99A: 279-310.
Rögl, F. 1999. Mediterranean and Paratethys. Facts and hypotheses of an Oligocene to Miocene paleoge-ography (short overview). Geologica Carpathica, 50: 339-349.
Rögl, F., Spezzaferri, S. & Ćorić, S. 2002. Micropaleontology and biostratigraphy of the Karpatian-Bade-nian transition (Early-Middle Miocene boundary) in Austria (Central Paratethys). Courier Forschun-gsinstitut Senckenberg, 237: 47-67.
Rögl, F. & Steininger, F. 1983. Vom Zerfall der Tethys zu Mediterran und Paratethys. Die Neogene Paläo-geographie und Palinspastik des zirkum-mediterranen Raumes. Annalen des Naturhistorischen Mu-seums in Wien, 85A: 135-163.
Samouelle, G. 1819. The entomologist’s useful compedium; or an introduction to the knowledge of British in-sects, comprising the best means of obtaining and preserving them, and a description of the apparatus gener-ally used; together with the genera of Linné, and the modern method of arranging the classes Crustacea,

Hyžný et al. Miocene squat lobsters of the Central Paratethys. Scripta Geol., 147 (2014) 259
Myriapoda, Spiders, Mites and Insects, from their affinities and structure, according to the views of Dr. Leach. Also an explanation of the terms used in entomology; a calendar of the times of appearance and usual situations of near 3,000 species of British insects; with instructions for collecting and fitting up objects for the microscope. Thomas Boys, London: 1-496.
Schmid, H.P., Harzhauser, M. & Kroh, A. 2001. Hypoxic events on a Middle Miocene carbonate platform of the Central Paratethys (Austria, Badenian, 14 Ma) with contributions by Ćorić, S., Rögl, F. & Schultz, O. Annalen des Naturhistorischen Museums in Wien, 102A: 1-50.
Schweitzer, C.E. & Feldmann, R.M. 2000. First notice of the Chirostylidae (Decapoda) in the fossil record and new Tertiary Galatheidae (Decapoda) from the Americas. Bulletin of the Mizunami Fossil Muse-um, 27: 147-165.
Schweitzer, C.E., Feldmann, R.M., Garassino, A., Karasawa, H. & Schweigert, G. 2010. Systematic list of fossil decapod crustacean species. Crustaceana Monographs, 10: 1-222.
Takeda, M., Mizuno, Y. & Yamaoka, M. 1986. Some fossil crustaceans from the Miocene Morozaki Group in the Chita peninsula, Central Japan. Kaseki no Tomo (Publication of the Tokai Fossil Society), 28: 12-22 [in Japanese].
Whiteaves, J.F. 1874. On recent deep-sea dredging operations in the Gulf of St. Lawrence. American Journal of Science, (3)7: 210-219.
Wiedl, T., Harzhauser, M., Kroh, A., Ćorić, S. & Piller, W.E. 2013. Ecospace variability along a carbonate platform at the northen boundary of the Miocene reef belt (Upper Langhian, Austria). Palaeogeogra-phy, Palaeoclimatology, Palaeoecology, 370: 232-246.

260 Hyžný et al. Miocene squat lobsters of the Central Paratethys. Scripta Geol., 147 (2014)
Plate 1
Galathea weinfurteri Bachmayer, 1950; type collection (considered lost), reillustrated from Bachmayer (1950, pl. 1, figs. 2-4). All figures are to scale; scale bar equals 5 mm.
Figs. 1-3. Carapace without rostrum (holotype), in dorsal (1, 2) and lateral (3) views.Figs. 4, 5. Carapace with fragmentary rostrum (detail).Figs. 6, 7. Left propodus.

Hyžný et al. Miocene squat lobsters of the Central Paratethys. Scripta Geol., 147 (2014) 261

262 Hyžný et al. Miocene squat lobsters of the Central Paratethys. Scripta Geol., 147 (2014)
Plate 2
Galathea weinfurteri Bachmayer, 1950
Fig. 1. Carapace without rostrum (NHMW 2011/0167/0134).Fig. 2. Carapace without rostrum (NHMW 2013/0580/0005).Fig. 3. Carapace with ‘bopyriform’ swelling (NHMW 2011/0167/0137).Fig. 4. Sternum (NHMW 2013/0580/0010).Fig. 5. Abdominal somite (NHMW 2011/0167/0133).Fig. 6. Carapace with rostrum (NHMW 2013/0580/0014).Fig. 7. Carapace with rostrum (NHMW 2013/0580/0012).Fig. 8. Carapace with rostrum (NHMW 2013/0580/0016).Specimens of Figs. 1, 3 and 4 are from the Fenk quarry (Austria), those of Figs. 2 and 5-8 are from Bad Deutsch-Altenburg (Austria). All specimens, except that of Fig. 8, were covered with ammonium chlo-ride prior to photography. Scale bars equal 1 mm (Figs. 4-7) and 5 mm (Figs. 1-3, 8).
Plate 3Figs. 1-3. Agononida cerovensis Hyžný & Schlögl, 2011, holotype (SNM Z-24.864); part (Figs. 2, 3) and counterpart (Fig. 1), photographed under different light conditions. All photos are to scale; scale bar equals 5 mm.

Hyžný et al. Miocene squat lobsters of the Central Paratethys. Scripta Geol., 147 (2014) 263

264 Hyžný et al. Miocene squat lobsters of the Central Paratethys. Scripta Geol., 147 (2014)
Plate 4
Munidopsis lieskovensis Hyžný & Schlögl, 2011
Figs. 1, 2. Carapace, and counterpart of the same specimen (KGP-MH CL-002).Fig. 3. Imprint of carapace (KGP-MH CL-003).Fig. 4. Sternum (KGP-MH CL-008).Fig. 5. Carapace (KGP-MH CL-004).Fig. 6. Carapace (KGP-MH CL-005)Fig. 7. Carapace (KGP-MH CL-006).Fig. 8. Carapace (KGP-MH CL-007).
Scale bar equals 1 mm.
Plate 5
Munidopsis palmuelleri n. sp.
Figs. 1, 2. Imprint of holotype (RGA/SMNH 0351) photographed uncoated and coated with ammonium chloride, respectively.Fig. 3. Stomaflex® Putty cast of same specimen, coated with ammonium chloride prior to photography.
Scale bars equal 5 mm.

Hyžný et al. Miocene squat lobsters of the Central Paratethys. Scripta Geol., 147 (2014) 265

266 Hyžný et al. Miocene squat lobsters of the Central Paratethys. Scripta Geol., 147 (2014)
Plate 6
Figs. 1-3. Galatheoidea, genus and species indeterminate. Associated pereiopods without carapace or venter. Note spines arranged in rows on P1 merus (arrows in Fig. 2) and tiny teeth on the occlusal sur-face of P1 fingers (arrows in Fig. 3).
Scale bar equals 5 mm.

Hyžný et al. Miocene squat lobsters of the Central Paratethys. Scripta Geol., 147 (2014) 267





















