
Paleomagnetism in northwestern Bulgaria: geological implications of widespread remagnetization

Available online at www.sciencedirect.com
alaeoecology 255 (2007) 342–360www.elsevier.com/locate/palaeo
Palaeogeography, Palaeoclimatology, P
Late Oligocene and Miocene climate and vegetation in the EasternParatethys area (northeast Bulgaria), based on pollen data
Dimiter A. Ivanov a,⁎, Abdul R. Ashraf b, Volker Mosbrugger c
a Bulgarian Academy of Sciences, Institute of Botany, Acad. G. Bonchev Street 23, BG-1113 Sofia, Bulgariab University of Tübingen, Institute of Geosciences, Sigwartstr. 10, D-72076 Tübingen, Germany
c Senckenberg Research Institute and Natural History Museum, Senckenberganlage 25, D-60325 Frankfurt, Germany
Received 3 November 2006; received in revised form 13 July 2007; accepted 29 August 2007
Abstract
The results of palynological studies of the Oligo-Miocene in the western part of the Euxinian Basin (Eastern Paratethys) arebased on the analysis of 57 samples from core C-136A drilled near the town of Balchik, Varna district. Some samples were barrenor contained very little pollen and were not included in the analysis. Quantitative data are thus confined to 26 polleniferoussamples. We present results for the Upper Oligocene as well as the Tarkhanian, Karaganian, Bessarabian, and Chersonian stages ofthe middle to upper Miocene.
Mixed deciduous forests were widespread during the time period studied. Swamp forests are also recorded, with distributionmaxima observed in the Oligocene and in the Bessarabian. The widest extent of herbaceous vegetation is recorded in the lateBessarabian and Chersonian, when it occupied open landscapes after a marine regressional phase.
The climatic data reconstructed by the Coexistence Approach indicate mean annual temperatures (MATs) of ca. 15–17 °C andmean annual precipitation (MAPs) of 800–1300 mm for the Upper Oligocene. The narrowest coexistence intervals of theTarkhanian and Karaganian display MATs of 14–17 °C and MAPs of 800–1300 mm. The most substantial data are for theBessarabian, in which the lower part is characterized by MATs of 15.6–16.6 °C and MAPs from 800–1200 (1300) mm. The resultsfor the upper part of the Bessarabian are more variable, especially with respect to precipitation that is generally lower (700–800 mm). The same is true for the Chersonian, where besides lower precipitation, lower annual temperature and temperature ofcoldest month are observed. These results correlate with previous vegetation and palaeoclimate data from the Forecarpathian Basin(NW Bulgaria).© 2007 Elsevier B.V. All rights reserved.
Keywords: Miocene; Oligocene, Palynology; Palaeoclimate; Vegetation; Bulgaria; Southeastern Europe
1. Introduction
Palaeoclimate reconstructions are essential for analysisof the response of the climate system to internal andexternal forcing factors, such as geographic reorganiza-
⁎ Corresponding author. Tel.: +35929793773; fax: +3592719032.E-mail address: [email protected] (D.A. Ivanov).
0031-0182/$ - see front matter © 2007 Elsevier B.V. All rights reserved.doi:10.1016/j.palaeo.2007.08.003
tion, orogenic activities, changing proportions of green-house gases, volcanic activity, solar energy, etc. Thedistribution pattern of vegetation strongly depends onclimatic conditions, and thus vegetation reconstructionshelp to understand past climates. In recent years,numerous climate reconstructions with different quanti-tative techniques (e.g. Coexistence Approach: Mosbrug-ger and Utescher, 1997; Climate Amplitude Method:
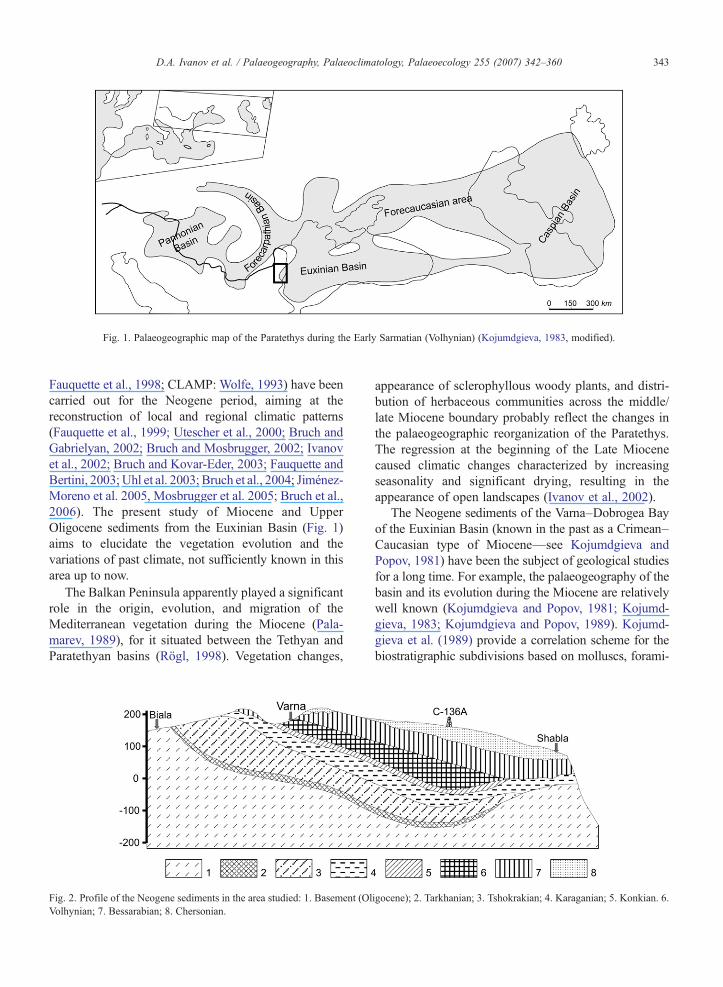
Fig. 1. Palaeogeographic map of the Paratethys during the Early Sarmatian (Volhynian) (Kojumdgieva, 1983, modified).
343D.A. Ivanov et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 255 (2007) 342–360
Fauquette et al., 1998; CLAMP: Wolfe, 1993) have beencarried out for the Neogene period, aiming at thereconstruction of local and regional climatic patterns(Fauquette et al., 1999; Utescher et al., 2000; Bruch andGabrielyan, 2002; Bruch and Mosbrugger, 2002; Ivanovet al., 2002; Bruch and Kovar-Eder, 2003; Fauquette andBertini, 2003; Uhl et al. 2003; Bruch et al., 2004; Jiménez-Moreno et al. 2005, Mosbrugger et al. 2005; Bruch et al.,2006). The present study of Miocene and UpperOligocene sediments from the Euxinian Basin (Fig. 1)aims to elucidate the vegetation evolution and thevariations of past climate, not sufficiently known in thisarea up to now.
The Balkan Peninsula apparently played a significantrole in the origin, evolution, and migration of theMediterranean vegetation during the Miocene (Pala-marev, 1989), for it situated between the Tethyan andParatethyan basins (Rögl, 1998). Vegetation changes,
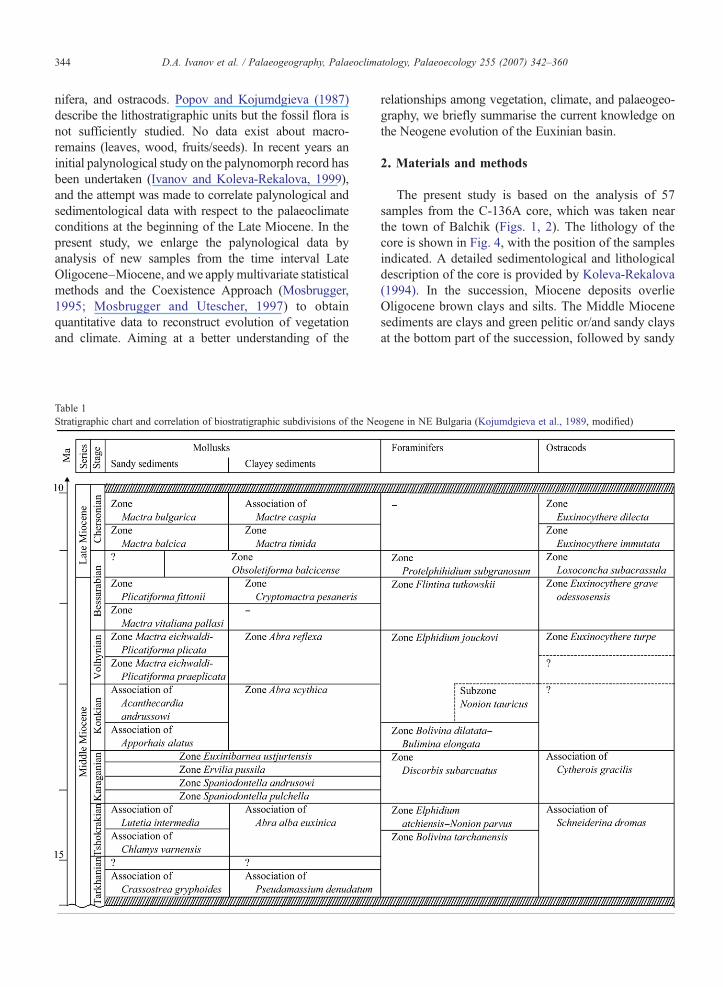
Fig. 2. Profile of the Neogene sediments in the area studied: 1. Basement (OlVolhynian; 7. Bessarabian; 8. Chersonian.
appearance of sclerophyllous woody plants, and distri-bution of herbaceous communities across the middle/late Miocene boundary probably reflect the changes inthe palaeogeographic reorganization of the Paratethys.The regression at the beginning of the Late Miocenecaused climatic changes characterized by increasingseasonality and significant drying, resulting in theappearance of open landscapes (Ivanov et al., 2002).
The Neogene sediments of the Varna–Dobrogea Bayof the Euxinian Basin (known in the past as a Crimean–Caucasian type of Miocene—see Kojumdgieva andPopov, 1981) have been the subject of geological studiesfor a long time. For example, the palaeogeography of thebasin and its evolution during the Miocene are relativelywell known (Kojumdgieva and Popov, 1981; Kojumd-gieva, 1983; Kojumdgieva and Popov, 1989). Kojumd-gieva et al. (1989) provide a correlation scheme for thebiostratigraphic subdivisions based on molluscs, forami-
igocene); 2. Tarkhanian; 3. Tshokrakian; 4. Karaganian; 5. Konkian. 6.
344 D.A. Ivanov et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 255 (2007) 342–360
nifera, and ostracods. Popov and Kojumdgieva (1987)describe the lithostratigraphic units but the fossil flora isnot sufficiently studied. No data exist about macro-remains (leaves, wood, fruits/seeds). In recent years aninitial palynological study on the palynomorph record hasbeen undertaken (Ivanov and Koleva-Rekalova, 1999),and the attempt was made to correlate palynological andsedimentological data with respect to the palaeoclimateconditions at the beginning of the Late Miocene. In thepresent study, we enlarge the palynological data byanalysis of new samples from the time interval LateOligocene–Miocene, and we apply multivariate statisticalmethods and the Coexistence Approach (Mosbrugger,1995; Mosbrugger and Utescher, 1997) to obtainquantitative data to reconstruct evolution of vegetationand climate. Aiming at a better understanding of the
Table 1Stratigraphic chart and correlation of biostratigraphic subdivisions of the Ne
relationships among vegetation, climate, and palaeogeo-graphy, we briefly summarise the current knowledge onthe Neogene evolution of the Euxinian basin.
2. Materials and methods
The present study is based on the analysis of 57samples from the C-136A core, which was taken nearthe town of Balchik (Figs. 1, 2). The lithology of thecore is shown in Fig. 4, with the position of the samplesindicated. A detailed sedimentological and lithologicaldescription of the core is provided by Koleva-Rekalova(1994). In the succession, Miocene deposits overlieOligocene brown clays and silts. The Middle Miocenesediments are clays and green pelitic or/and sandy claysat the bottom part of the succession, followed by sandy
ogene in NE Bulgaria (Kojumdgieva et al., 1989, modified)

Fig. 3. Sketch map showing the structural/palaeogeographic areas innortheast Bulgaria during the Neogene as well as the location of thedrilling C-136A (redrawn from Popov and Kojumdgieva, 1987).Legend: 1. Land; 2. South-Dobrodgea strait; 3. Marginal area ofVarna–Balchik depression; 4. Varna–Balchik depression: IIIa Varnapart; IIIb Balchik part; 5. Boundaries of palaeogeographic structures;6. Core.
345D.A. Ivanov et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 255 (2007) 342–360
biogenic limestones, in rare cases interbedded with thinclay laminae (clayey-limestone formation, 76.1 m thick,Fig. 4). The overlying Euxinograd Formation in itslower part consists of diatomaceous clays interbeddedwith sandy clays and clayey limestone (28.8 m thick).The next sediment succession consists of white shellyand sandy limestone, calcareous sandstone, and sands(49.3 m thick). The sediments of the upper part of theEuxinograd Formation consist of diatomaceous clayswith layers of biogenic clays and micritic limestone.Unconsolidated aragonite sediments predominate in theTopola Formation, but clays, and micritic limestoneinterbeds are also identified. In the upper part of theformation, the sediment is different, and laminatedaragonite sediments alternate with laminae of aragoniteclays and micritic limestone. The rest of the profile(Fig. 4: Karvuna Formation) consists of an alternation ofshelly and micritic limestone. Pleistocene sandy clayscover the Miocene succession disconformably at the topof the core.
As indicated by data obtained from the marine fauna(foraminifera, molluscs, and ostracods) the sedimentsequence covers the stratigraphic range from the LateOligocene to the lower part of the Late Miocene. TheEarly Miocene is missing (Figs. 2, 4, Table 1).
Only 26 out of the 57 samples contain enoughpalynomorphs to provide reliable data (full taxonomiccomposition and distribution of the taxa in all studiedsamples are presented in Appendix A). Unfortunately,21 samples are entirely devoid of pollen, while tenothers contain pollen in low concentrations (with lessthan 100 grains counted). To minimise statistical errorsand to avoid wrong interpretations, we do not considerthese 10 pollen spectra in our interpretation ofvegetation and palaeoclimate. The 26 pollen spectraare used to reconstruct the climate evolution with theCoexistence Approach (Mosbrugger, 1995, Mosbruggerand Utescher, 1997). Thereby the following climaticparameters are considered: MAT—mean annual tem-perature (°C); TCM—mean temperature of the coldestmonth (°C); TWM—mean temperature of the warmestmonth (°C); and MAP—mean annual precipitation(mm).
In addition, a pollen diagram is constructed from thepalynological data, and the results are analysed andinterpreted. The percentage of each identified taxon inthe pollen spectra was calculated with respect to thepollen sum AP+NAP=100%. The proportions ofspores were calculated with respect to the total sumAP+NAP+L=100%. Multivariate statistical methodswere used to show the succession of palynomorphassociations. Minimum variance clustering (Ward's
method) based on squared Euclidian distance is applied.The data matrix for the cluster analysis includes theproportion of each spore/pollen taxon in the samplerelative to the total pollen sum counted for that sample(Fig. 8). In addition, relative abundances of the taxawere correlated using Pearson's correlation. As a result,a data matrix with correlation coefficients between taxawas obtained and then used in cluster analysis in order todetect similarities among individual taxa, as well as todetermine the groups of plants with simultaneouslychanged distribution patterns (Fig. 9).
3. Palaeogeography of the basin
The Euxinian Basin represents the western part of theEastern Paratethys (Fig. 1). It played an important rolein connecting Miocene basins, providing seawaysbetween the eastern and central parts of the Paratethysthroughout the Black Sea Basin (Rögl, 1998). The briefdescription of the geology and palaeogeographicevolution of the area presented here is based on thedata published by Kojumdgieva and Popov (1981,1989), Kojumdgieva (1983), Popov et al. (1986), Popov

Fig. 4. Lithologic column of the profile studied. Legend: 1. Clays; 2. Sandy clays; 3. Limestone; 4. Sandy limestone; 5. Aragonitic sediments;6. Clayey aragonitic sediments; 7. Shelly limestone; 8. Diatom-spongolitic clays; 9. Sands; 10. Calcareous clays; 11. Siltstones (silts); 12. Calcareousdolomites; 3. Biodetritic clay; 14. Transgressive surface; 15. Terra rossa.
346 D.A. Ivanov et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 255 (2007) 342–360

Fig. 5. Sketch map showing outlines of the basin during Tarkhanian–Konkian: (compiled after Popov and Kojumdgieva, 1987): Legend:1. Tarkhanian; 2. Tshokrakian; 3. Karaganian; 4. Konkian.
Fig. 6. Sketch map showing outlines of the basin during Sarmatian:(compiled after Popov andKojumdgieva, 1987): Legend: 1. Volhynian;2. The beginning of Bessarabian; 3. The end of Bessarabian;4. Chersonian.
347D.A. Ivanov et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 255 (2007) 342–360
and Kojumdgieva (1987), and Kojumdgieva et al.(1989).
Three main structural-palaeogeographic areas wereintroduced for the area by Kojumdgieva and Popov(1981) (see Fig. 3): I. South-Dobrogea strait—slightlyuplifted area, flooded by the sea during two shortperiods: first at the beginning of the Middle Miocene,and then during the Bessarabian, forming a sea channelto the Forecarpathian (Dacic) Basin. II. Marginal areaof the Varna–Balchik depression—comparatively stableand uplifted belt surrounding the depression to the westand north. This area was the shelf of the Euxinian Sea.III. Varna–Balchik depression, which appeared at thebeginning of the Middle Miocene and was active up tothe Chersonian. Probably it was only the westernmostpart of a larger depression, the main part of which wassituated in the Black Sea. The depression was dividedinto two parts: The Varna and the Balchik part (IIIA andIIIB on Fig. 2).
During the Early Miocene, northeast Bulgaria was anupland, and no sediments are known from that time. Atectonics-controlled reorganization in the latest Early toearliest Middle Miocene in the Eastern Paratethysresulted in the emergence of the Greater Caucasian
archipelago (Meulenkamp and Sissingh, 2003), fol-lowed by a transgression in the territories west of theBlack sea. The first transgression in the Varna–Balchikarea took place in the Tarkhanian (Fig. 5), when only afew meters of clays, rich in marine foraminifers, weredeposited, as well as some detritic limestones in theSouth-Dobrogea strait. These sediments disconformablyoverlie the Oligocene basement. In the TshokrakianBasin a regressive trend existed, but the sedimentationwas still intensive in the Varna–Balchik depression.During that time, the Ponto-Caspian area was sealed offagain from the open oceans (Rögl, 2001), thus leading tothe appearance of brackish environments. The salinity ofthe basin was low, as evident from the endemiceurihaline fauna (Kostadinov and Kojumdgieva,1964). Diatom analysis indicates changes in watersalinity from 8–10‰ at the beginning of the Tshokra-kian to 12–15‰ at the end of the period (Temniskova-Topalova, 1994).
A slight transgression took place in the Karaganian,when marginal areas were flooded by seawater, andsandy and calcareous sediments were deposited in
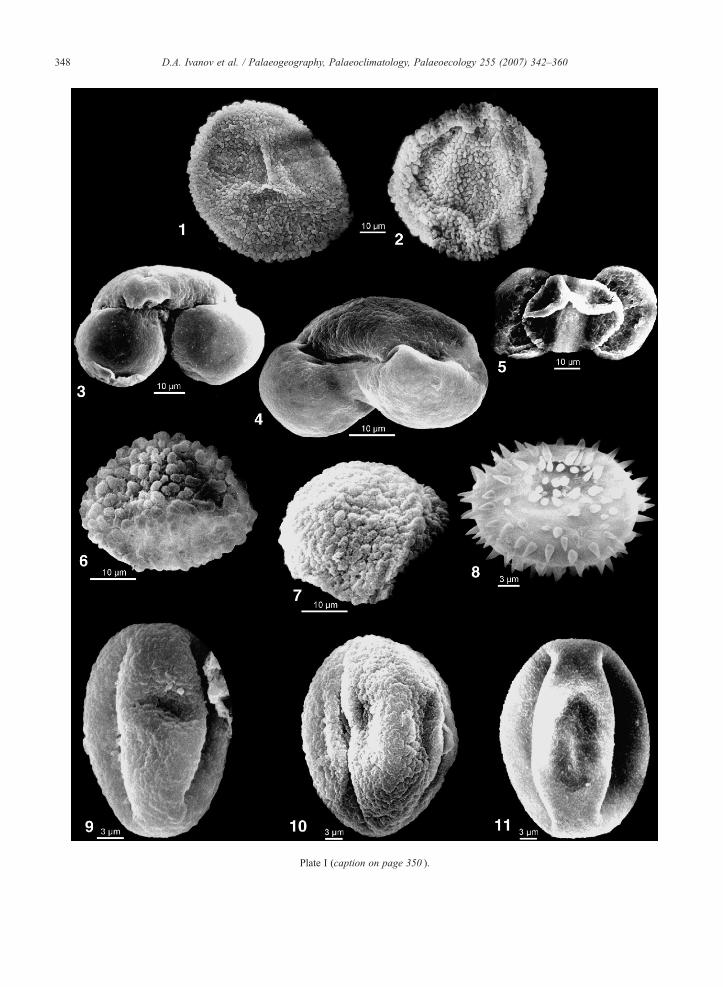
Plate I (caption on page 350 ).
348 D.A. Ivanov et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 255 (2007) 342–360
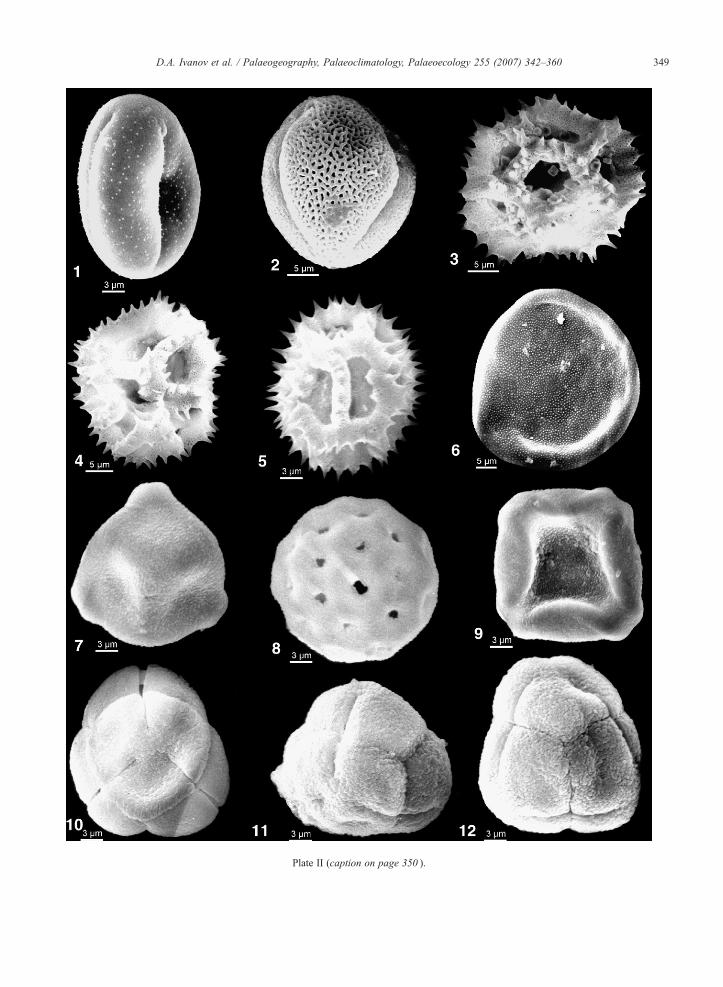
Plate II (caption on page 350 ).
349D.A. Ivanov et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 255 (2007) 342–360

350 D.A. Ivanov et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 255 (2007) 342–360
shallow water (Fig. 5). Terrigenous sedimentationpredominated in the area of Varna, while in the Balchikpart of the depression mainly clays and limestones weredeposited, succeeded during late Karaganian by lami-nated calcareous clays and diatomites (EuxinogradFormation, lower part—Fig. 4). The character of thediatom flora suggests a water salinity of ca. 18–20‰(Temniskova-Topalova, 1994).
In the Konkian Basin a regressive trend is observed,and only Balchik and the northern part of the Varnadepressions were flooded (Fig. 5). There mainlycalcareous and diatomaceous clays were deposited.Despite their small thickness (less than 10 m) they aredivided into two zones by the contained fauna (Table 1):the lower one with a fauna living at normal marinesalinity (25–30‰ according to diatom data presented byTemniskova-Topalova, 1994), and the upper one with aeuryhaline fauna, typical for an isolated basin (Popovand Kojumdgieva, 1987). Rögl (1998, 2001) suggestsan open connection during the early Konkian in easternAnatolia that linked the Indian Ocean to the Paratethys,and full marine conditions returned in the Euxinianbasin with stenohaline molluscs and foraminifers.Regarding the late Konkian, Ilyina et al. (2004) presumereduced water salinity (about 20‰), based on changesin the composition of the mollusc fauna.
Plate I. (see page 348)
1. Osmunda sp. (Osmundacidites primarius (Wolff) Nagy); Proxim2. Osmunda sp. (Osmundacidites primarius (Wolff) Nagy); Distal3. Pinus sylvestris type (Pityosporites labdacus (Pot.) Raatz); C-134. Keteleeria sp. (Keteleeriapollenites komloënsis Nagy); C-136A,5. cf. Podocarpus (Podocarpidites); C-136A, 133.0 m, Bessarabia6. Sciadopitys sp. (Sciadopityspollenites serratus (Pot. & Ven.) Ra7. Sciadopitys sp. (Sciadopityspollenites serratus (Pot. & Ven.) Ra8. Nuphar sp. (Nupharipollis echinatus Krutzsch); C-136A, 146.59. Quercus sp. 1. (Quercoidites henrici (Pot.) Pot. Th. & Thierg.);10. Quercus sp. 2. (Quercoidites granulatus (Nagy) Slodkowska); C11. Quercus sp. 3. (Quercoidites sp.); C-136A, 133.0 m, Bessarabia
Plate II. (see page 349)
1. Ranunculus sp.; C-136A, 141.0 m, Bessarabian;2. Tricolporopollenites sp.; C-136A, 146.5 m, Bessarabian;3. Cichorioideae (Cichoreacidites gracilis (Nagy) Nagy); C-136A,4. Cichorioideae (Cichoreacidites gracilis (Nagy) Nagy); C-136A,5. Cichorioideae (Cichoreacidites gracilis (Nagy) Nagy); C-136A,6. Carya sp. (Caryapollenites simplex (Pot.) Raatz); C-136A, 146.7. Betula sp. (Betulaepollenites betuloides (Pfl.) Nagy); C-136A, 18. Chenopodiaceae (Chenopodipollis multiplex (Weyl. & Pfl.) Kru9. Alnus sp. (Alnipollenites verus Potonié); C-136A, 133.0 m, Bes10. Ericaceae (Ericipites ericius (Pot.) Krutzsch); C-136A, 214.8 m11. Ericaceae (Ericipites callidus (Pot.) Krutzsch); C-136A, 214.8 m12. Ericaceae (Ericipites callidus (Pot.) Krutzsch); C-136A, 214.8 m
The Euxinian Sea transgressed onto large land areasduring the Volhynian–Chersonian, especially at thenorthern and eastern margins (Paramonova et al., 2004).In the beginning of the Volhynian, a slight probablyeustatic transgression took place also in our study area,and mainly diatomaceous clays with thin detritic layerswere deposited. Later, in Bessarabian the sea flooded theSouth-Dobrogea strait (Fig. 6), where shallow lime-stones of the Odartsi Formation were deposited.Chemical sedimentation started in the Balchik depres-sion (zone Cerastoderma balcicense), and aragoniticsediments accumulated (Koleva-Rekalova, 1994). Dur-ing the Chersonian, a small regression occurred, andbefore the end of this stage the sea abandoned northeastBulgaria. The Chersonian is represented mainly bylaminated sediments: aragonitic laminae alternate withlaminae rich in clay minerals (Koleva-Rekalova, 1994;Ivanov and Koleva-Rekalova, 1999). The basin wasisolated from the ocean, and the salinity was lower,probably between 10–15‰ during this period (Temnis-kova-Topalova, 1994).
4. Vegetation
The samples studied from the Euxinian Basinprovide in total 85 different spore and pollen taxa
al view, C-136A, 146.5 m, Bessarabian;view, C-136A, 133.0 m, Bessarabian;6A, 301.1 m, Tarkhanian;141.0 m, Bessarabian;n;atz); C-136A, 141.0 m, Bessarabian;atz); C-136A, 301.1 m, Tarkhanian;m, Bessarabian;C-136A, 146.5 m, Bessarabian;-136A, 141.0 m, Bessarabian;n;
141.0 m, Bessarabian;141.0 m, Bessarabian;133.0 m, Bessarabian;5 m, Bessarabian;46.5 m, Bessarabian;tzsch); C-136A, 141.0 m, Bessarabian;sarabian;, Karaganian;, Karaganian;, Karaganian;

Fig. 7. Pollen diagram of core C-136A. Shaded area: 20 times exaggerated. Legend: 1. Herbaceous communities (cluster A1— see Fig. 8); 2. Mixed mesophytic forests (cluster A2); 3. Swamp forests(cluster B); 4. Pinus-dominated pollen spectra (cluster C).
351D.A.Ivanov
etal.
/Palaeogeography,
Palaeoclim
atology,Palaeoecology
255(2007)
342–360

352 D.A. Ivanov et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 255 (2007) 342–360
(Appendix A; Plates I, II) belonging to pteridophytes,gymnosperms, and angiosperms. The angiospermsshow the highest diversity, with a total of 62 fossiltaxa. The gymnosperms are represented by 14 taxa, andthe pteridophytes by 9 taxa. Spore plants are poorlyrepresented in the pollen spectra, both in speciesrichness and abundance. The highest amount of sporesis found in samples from the bottom part of the core, insediments of Oligocene and Tarkhanian age. In generalthe palynoflora is composed of a mix of taxa, whosenearest living relatives have temperate and subtropicaldistributions.
The palaeoecological analysis of the fossil microfloraand the abundance of individual fossil taxa (Fig. 7)provide evidence about the past plant communities.During the whole period mixed mesophytic forestsappear to have been widespread in the lowlands andmid-altitude uplands surrounding the basin, thusrepresenting the zonal vegetation. Species of Quercusand Ulmus dominated in these forests, accompanied byCastanea, Carya, Pterocarya, Juglans, Ilex, Eucom-mia, Betula, Carpinus, Corylus etc. Thermophilousplants like Platycarya, Engelhardia, Symplocos, Ree-vesia, Sapotaceae, Araliaceae, and Arecaceae are alsopresent in the pollen spectra, with frequencies varyingwith respect to age and facies of the strata. These plantsare more abundant in the pollen spectra from theOligocene, partly Tarkhanian and the lower part of theBessarabian (Fig. 7).
The presence of pollen of Pinus, Tsuga, Abies, Ke-teleeria, and Picea suggests the existence of mixedconiferous–broadleaved forests on elevated areas, ormesophytic forests with a mixture of coniferous species.In some of the samples the high abundance of Pinus-pollen, with its ease in air and water transport (Suc andDrivaliari 1991), may reflect long distance transportfrom neighbouring mountains, namely the eastern partof Palaeo-Balkan Mts., located to the southwest, and/orthe Palaeo-Carpathians to the northwest.
Swamp forests, as azonal vegetation, grew on wetterrain and swampy places around the water basin,where the soil was under the water level during theentire vegetation season or the greater part of it.Taxodiaceae dominated in these habitats, along withspecies of Alnus, Myrica, Nyssa, Sciadopitys, Cyper-aceae, and Osmunda. On more elevated places (or inriver valleys), Salix, Populus, Platanus, and Liquidam-bar were accessory in these forests. The maximumdistribution of these plant communities was recorded forthe Oligocene and partly for the lower part of theBessarabian sediments. These palaeofloristic data cor-respond to our data from Northwest Bulgaria, where the
basal parts of the Bessarabian are characterized by avegetation rich in palaeotropical elements, but at the endof this stage their abundance tends to decrease(Palamarev and Ivanov, 2001; Ivanov et al., 2002).
Herbaceous plants formed important communitiescomposed of species belonging to the following generaand families/subfamilies: Apiaceae, Lamiaceae, Poa-ceae, Asteroideae, Cichorioideae, Persicaria, Artemi-sia, and Chenopodiaceae. The maximum distribution ofherbaceous vegetation is recorded in the upper part ofBessarabian and Chersonian (i.e. the beginning of theLate Miocene) with higher proportions of its compo-nents. These higher proportions are combined withsubxerophitic/xerophitic plants, which became increas-ingly important—species of Celtis, Pistacia, Oleaceae,Ephedra. For other areas of Bulgaria, in addition tothese plants other subxerophytes were identified asmacrofossils: Quercus (ex. sect. ilex), Robinia, Arbu-tus, Berberis, Paliurus, Celastrus, and Cesalpinites(Palamarev, 1989, 1990, 1991, Palamarev and Ivanov,1998, 2001). Simultaneously, the distribution area ofswamp forests was reduced. These palaeobotanical datasuggest an appearance of open landscapes and thedevelopment of xerophytic plant communities, inaccordance with the suggestion of Jacobs et al. (1999)about the steady spread of steppe vegetation and theexistence of a more arid interior in southeastern Europe,Ukraine, and Turkey at about 10 Ma. These initial openlandscapes were probably of the so-called parklandtype, where a mosaic of open landscapes and smaller orlarger stands of trees exist.
The palynofloras studied were grouped by clusteranalysis into four clusters, coded as A1, A2, B, and C onFig. 8. The major distances observed among the casesare between clusters A–B and C. The C-clustercombines palynofloras from Tarkhanian, Karaganian,and some from Bessarabian sediments with highestabundance of Pinus-pollen. We suggest that thesepalynofloras reflect changes in the basin configurationand correspond to transgressive periods and themaximum extension of the basin (see previous chapter),when the core has a maximum distance from the coastline. The B-cluster includes only two floras (one fromOligocene and one from Bessarabian sediments), wherethe widest distribution of swamp forests was recorded.We suggest that they also reflect changes in thepalaeogeography, i.e. periods linked with the beginningof regressions or eustatic sea-level changes, whenswampy areas developed along the margin of the basin.
The A1-cluster corresponds to the maximum occur-rence of herbaceous vegetation and xerophytes, whereasthe A2-cluster reflects the widest extension of mixed

Fig. 8. Dendrogram for pollen spectra based on squared Euclidian distance. The data matrix for the cluster analysis includes the relative proportion ofeach spore/pollen taxon in the sample relative to the total pollen sum counted for that sample.
353D.A. Ivanov et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 255 (2007) 342–360
mesophytic forests. The differentiation of the palyno-floras into these two groups could be related to climaticchanges, especially to drying of the climate in someparts of the Bessarabian and Chersonian. This assump-tion agrees with palaeoclimate data obtained from theCoexistence Approach (see below).
To obtain additional information about the structureof the vegetation and changes of different vegetationtypes we made a correlation analysis with relativefrequencies of palynomorph taxa. The taxa are groupedin several more or less well-defined clusters (Fig. 9). Forexample, the mesophytic species, preferring wet habitats(Engelhardia, Pterocarya, Platanus, Alnus, Myrica,with admixture of some herbs Apiaceae, Thalictrum)are groped together, but the presence of Pistacia(growing in dry habitats) in this cluster cannot beexplained. Also the group of herbaceous species is well-defined (Poaceae, Asteroideae, Cichorioideae, Caryo-
phyllaceae, and Chenopodiaceae), together with Celtis.Ulmus, and Fagus join this group of plants, andapparently their increasing abundance in the fossilflora reflects climatic change towards drier conditions,when the herbaceous communities became morewidespread. These plants replaced some of the speciesrequiring more humid environments in mixed meso-phytic forests. Another well-defined group combinesTaxodiaceae, Nyssa, Sequoia, and Liquidambar (plantsforming swamp vegetation), probably restricted to smallareas adjacent to rivers. The group of Pinus, Abies,Tsuga, Sciadopitys combined with Tilia and Ilex form aseparate cluster. This cluster could confirm the presenceof mixed coniferous–broadleaved forests in somehabitats surrounding the basin. It is not clear whyEphedra, a genus that nowadays grows in rocky and dryplaces, is grouped in this cluster. One possibleexplanation for the unexpected presence of taxa in

Fig. 9. Dendrogram for correlations between taxa. The data matrix for the cluster analysis was obtained by correlation analysis of the taxa in thesection studied.
354 D.A. Ivanov et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 255 (2007) 342–360
single groups is the wide stratigraphic range of thestudied profile. The correlation analysis works wellwhen we trace the dynamics within the phytocoenoses.In our case we observe not only dynamics within theplant communities, but also the appearance of newcommunities (herbaceous vegetation, groups of hemi-xerophytic and sclerophyllous taxa). On the other hand,we have a lot of barren samples (see above), and in some
cases bisaccate pollen is over-represented, leading togaps in the reconstruction of the vegetation evolution.Finally we have to keep in mind that ecological require-ments of Neogene plants could significantly differ fromthose of their recent relatives.
The relative abundance of the clusters (Fig. 10) in theprofile studied shows simultaneous decrease during theLate Bessarabian and Chersonian in the percentage of

355D.A. Ivanov et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 255 (2007) 342–360
the clusters 1 and 2 including mainly hygrophilous taxaand the increase in the abundance of clusters 3 and 4,combining herbaceous taxa and temperate deciduousplants. Components of swamp forests (grouped incluster 5) also decrease. All these changes coincidewith precipitation change (Fig. 11) and support ourassumption about climate-driven changes in the vege-tation structure.
5. Palaeoclimate
The palaeoclimate records obtained from the appli-cation of the Coexistence Approach to the palynoflorasof the C-136A core are shown in Fig. 11. The present-day climate of northeast Bulgaria is characterized byMAT 12.0 °C, TCM 1.9 °C, TWM 22.0 °C and MAP480 mm at the meteorological station of Varna and byMAT 11.2 °C, TCM −0.4 °C, TWM 22.8 °C and MAP592 mm at the meteorological station near Shumen,80 km to the west of the Black Sea (Velev, 1997).
Fig. 10. Relative abundance of the clusters in the profile studied: 1. PterocaryCorylus, Alnus, Betula, Engelhardia, Engelhardieae; 3. Celtis, Ulmus, Caryoph4. Quercus, Caprifoliaceae, Ericaceae, Carya, Salix, Keteleeria; 5. TaxodiaceaCyperaceae, Ephedra, Pinus haploxylon type, Pinus sylvestris type.
The data obtained from the Upper Oligocene sedi-ments display a relatively warm and humid subtropicalclimate. The MAT coexistence intervals range between15.6 and 17.2 °C, but wider intervals also occur; TCM isbetween 5.0 and 7.0 °C, and the TWM coexistenceinterval is constantly 24.7–27.8 °C. The lower boundariesof the MAP coexistence intervals are 740 and 823 mm,respectively, and the upper boundary is 1322 mm. Bruch(1998) and Bruch and Mosbrugger (2002) calculatedcomparable coexistence intervals for the Oligocene floraof Slovenia—mean annual temperatures of 16–19 °C,temperatures of coldest and warmest months of 6–9 °Cand 25–28 °C, and precipitation between 1100 and1300 mm. Similar results were reported for the Oligocenein northwest Germany by Utescher et al. (2000), e.g. forthe Enspel flora annual temperatures range between15.7 °C and 16.8 °C, summer (around 25 °C), and winter(5–6 °C) temperatures, and the mean annual precipitation(970–1200 mm), values very close to our data. Thus allthe data evidence the relative homogeneity of climate
a, Thalictrums, Apiaceae, Sparganium, Platanus, Carpinus; 2.Myrica,yllaceae, Chenopodiaceae, Cichorioideae, Asteroideae, Poaceae, Fagus,e, Nyssa, Liquidambar, Sequoia; 6. Abies, Sciadopitys, Tilia, Tsuga, Ilex,

Fig. 11. Coexistence intervals derived from samples of core C-136A. MAT—mean annual temperature; TCM—mean temperature of the coldest month; TWM— mean temperature of the warmestmonth; MAP — mean annual precipitation. (Note: The calculations for sample at 16.0 m without single grains are shown with hollow bars next to the previous ones. — see the explanation in thechapter about palaeoclimate.)
356D.A.Ivanov
etal.
/Palaeogeography,
Palaeoclim
atology,Palaeoecology
255(2007)
342–360

357D.A. Ivanov et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 255 (2007) 342–360
conditions during this period. Utescher et al. (2000)suggest a warming trend toward the end of the Oligocene,which our present results do not confirm. This might bedue to imprecise dating and restricted stratigraphicresolution in the time-interval mentioned.
The Early Miocene is not present in our study area,and our record continues in the Middle Miocene. Des-pite the high number of samples from the sequenceTarkhanian–Volhynian, only a few pollen samples pro-vide palaeoclimate results. Some data are obtained forTarkhanian and Karaganian horizons. There wide rangesof coexistence intervals are characteristic (Fig. 11.) dueto the low taxonomic diversity and insufficient numberof taxa in coexistence in these palynofloras. The nar-rowest CA-interval for the MAT is 13.6–16.6 (17.2)°C,for TCM 2.4–7.0 °C, TWM 24.7–27.8 °C, and for theMAP 823–1206 mm. Comparison of these data withquantitative climatic results from corresponding sedi-ments (of the Badenian age) in other areas of Bulgaria(Ivanov et al., 2002; Palamarev and Ivanov, 2004)suggests that they are less precise for taphonomic rea-sons. Comparing the data from different regions inEurope, Bruch et al. (2004) demonstrate that the MiddleMiocene temperature parameters do not indicate anyspatial pattern or latitudinal gradient, and that the cli-mate was more or less homogenous within Europe.
Most extensive data were obtained for the Bessar-abian and Chersonian sediments. The lower part of theBessarabian is characterized by relatively stable climaticconditions (Fig. 11), and the most common intervals forclimate parameters are as follows: mean annual tem-perature of 15.6–16.6 °C (and even higher in somecases), temperatures of the warmest month 24.7–27.3 °C, temperatures of the coldest month ca. 5.0–7.0 °C, and mean annual precipitation 823–1206 (1308)mm. Towards the end of Bessarabian and duringChersonian changes in the climate parameters occurred,and probably climate became more variable and drier. Insome cases we observe a decline of the lower limit oftemperature intervals, but most significant are lowervalues obtained for the coexisting intervals of meanannual precipitation—(559 mm) 740–760 mm. Theseresults coincide with sedimentological data (Koleva-Rekalova, 1994; Ivanov and Koleva-Rekalova, 1999)for the Bessarabian and Chersonian, when massive andlaminated aragonitic sediments were deposited. Koleva-Rekalova (1994) suggests an inorganic origin foraragonite–chemical precipitation at high temperature,shallow-water conditions, and limited inflow of terrig-enous components. Nowadays, aragonitic muds aredeposited in tropical and subtropical arid to semiaridareas, e.g. in the Persian Gulf and the Red Sea.
Regarding the Chersonian laminated aragonitic sedi-ments with varved structure, deposition in seasonalclimate can be assumed (Ivanov and Koleva-Rekalova,1999). During the hot dry period of the year aragonitelaminae were precipitated, while the clay laminas weredeposited in the wet season. The existence of drierclimatic conditions is also in accordance with thevegetation changes discussed above: the increasing roleof herbaceous plant communities and increasing pollenproportions of the subxerophytes.
One of the most important questions is the following:Are the observed vegetation changes climaticallydriven, or are they induced by sea-level changes? TheEuxinian Sea had its widest extent during earlyBessarabian, while in the late Bessarabian and Cherso-nian a regression took place (see Fig. 6). The continentalareas abandoned by the sea were suitable for halophytes(Chenopodiaceae/Amaranthaceae), which appeared aspioneer vegetation. They should be rapidly replaced bytree vegetation under wet climate conditions. Such atype of succession must have existed for only fewdecades (as compare to the rate of recent plantsuccessions, e.g. Ohmann and Spies 1998; Foster andTilman 2000; Zhang 2005, etc.), while in our case werecorded a long-lasting presence of this vegetation type.So the long-term existence of herbaceous communitiescannot be explained only with the presence of salt soils.Apparently the drier climate with well expressedseasonality favored the distribution of this vegetationtype during late Bessarabian and Chersonian. Thepresent-day vegetational distribution pattern displays astrong climatic dependence. For example, deciduousforests of the temperate zone are confined to climaticregions of an oceanic nature, where there are no sharpextremes of temperature and where the rainfall is moreor less evenly distributed throughout the year, usuallywith a summer maximum (Walter, 1973). Steppesoccupy regions with a continental type of climate,where the annual temperature amplitude is greater, thewinters are colder, and the summers are hotter and morearid.
As emphasized above, the upper part of the Bes-sarabian and Chersonian has to be characterized ascomparatively dry and warm, possibly with some trendsto slightly lower annual temperatures and increasedseasonality. Only the data derived from the palynoflora at16.0 m (see Fig. 11) do not coincide with the data fromunderlying palynofloras: higher intervals for all temper-ature parameters, and high precipitation rates—1187–1206mm. These intervals were derived from the climaticrequirements of Reevesia and Engelhardia (presented inthe pollen spectrum by single pollen grains—0.2%).

358 D.A. Ivanov et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 255 (2007) 342–360
When excluding single specimens from the calculationsthe results do not indicate any climatic change. Probablythere was no significant climatic change, because of thelack of other evidences, e.g., there is neither a significantchange in the type of sedimentation, nor are changes inthe proportion of other pollen taxa observed in thisspectrum. When we exclude the taxa mentioned abovefrom our climatic calculations, similar values for allclimatic parameters as for the rest of the samples of thelater Bessarabian and Chersonian were obtained (Fig. 11:the calculations without single grains, for sample of16.0 m, are shown with hollow bars next to the previousones). Regretfully, the studied samples from the intervalat 81.0–16.0 m have not yielded any pollen grains, andwe cannot trace the vegetation succession for this period.Also, the upper part of the Chersonian is not documen-ted, because the sea retreated from the territory ofnortheast Bulgaria.
The climatic changes in the time period from the lateBessarabian to the Chersonian (the beginning of theLate Miocene) discussed above are in good accordancewith quantitative palaeoclimate data for the Forecar-pathian (Dacian) Basin (Central Paratethys) in north-west Bulgaria (Ivanov et al., 2002), where a slightdecrease in temperatures and a significant drying issimilarly observed: CA intervals for MATs 13.3 (14.4)–17.2 °C and CA intervals for MAPs 652 (740)–752 mm.In the area of the Euxinian Basin, however, this drierperiod started somewhat earlier, as is evidenced byvegetation changes and sedimentology. Moreover, thesedimentological data (absence of massive aragonitesediments in the Forecarpathian Basin) suggest drierconditions in the Euxinian Basin (Ivanov and Koleva-Rekalova, 1999). These data support a previous hypoth-esis about declining precipitation in the Paratethyanrealm (Ivanov and Koleva-Rekalova, 1999, Stuchliket al., 1999, Ivanov et al., 2002). Palynological datatestified to the increasingly xerophytic nature of thevegetation at the end of the Bessarabian and Chersonianin southern Ukraine (Syabryay and Stekina, 1983),proved also by Belokris (1988) on the basis of thepresence of dolomites, which form under very dryconditions. These data are in agreement with the dataobtained from small mammals (van Dam, 2006), whichalso indicate an aridification trend at the beginning ofthe Late Miocene. The ecomorphological analyses oflarge Tortonian mammals (Brayard et al., 2004) andcomputed ecomorphologic parameter for Europeanlarge mammals localities also support the existence ofaridity gradient by Late Miocene time, with more openand arid environments in southern Europe. The analysisof the hypsodonty-based maps of precipitation distribu-
tion (Fortelius et al., 2006) suggests that since ca.11.1 Ma (MN 9 zone) the climate of Europe wasdominated by a humidity gradient from continental tomore marine conditions from east to west.
The regression of the Paratethys at the beginning ofthe Late Miocene caused a greater seasonality of climateand significant drying, due to shrinking water basins andthe appearance of open landscapes. As evident fromvegetation data (Stuchlik et al., 1999), the climate didnot change equally in the northern and southern areas:To the North it was cooler and less dry. Kováč et al.(2006) also observed that vegetation with subtropicalfloral elements was replaced by vegetation of thetemperate zone in the area of the Pannonian Basin.The authors interpreted these changes as an appearanceof seasonality with lowered humidity. Also drierconditions can be assumed for the eastern areas incomparison to the western ones. Ivanov and Koleva-Rekalova (1999) suggest that a dry climate beltestablished north or northeast of the Carpathians,which gradually spread to the south to the westernparts of the Euxinian Basin, later occupying the southernparts of the Forecarpathian Basin and the PannonianBasin to the west. The palaeoclimatic reconstructions ofUtescher et al. (2000) for western Germany inferred thatwet climate existed during the whole Miocene. Probablyto the west the climate was somewhat softened by theinfluence of the Atlantic Ocean and predominance ofwesterly winds that brought humid air over westernEurope.
6. Conclusions
By analysing palynofloras from the western part ofthe Euxinian Basin we reconstruct vegetation andpresent quantitative palaeoclimate data for the followingregional stages of the Eastern Paratethys: UpperOligocene, Tarkhanian, Karaganian, Bessarabian, andChersonian. Mixed deciduous forests were widespreadduring most of the time period studied. Swamp forestswere also present, with two distribution maximaobserved in the Oligocene and in the Bessarabian, asconfirmed by cluster analysis. The widest distribution ofherbaceous vegetation is recorded for the upper part ofthe Bessarabian and the Chersonian, when it probablyoccupied the open landscapes after regression of the seaand was also favoured by climatic changes.
The data obtained for the Late Oligocene suggestmean annual temperatures (MATs) of ca. 15–17 °C andmean annual precipitation rates (MAPs) of 800–1300 mm. The narrowest coexistence intervals derivedfor the Tarkhanian and Karaganian display MATs of 14–

359D.A. Ivanov et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 255 (2007) 342–360
17 °C and MAPs of 800–1300 mm. The most extensivedata are for the Bessarabian. The lower part of this stageis characterized by MATs between 15.6–16.6 °C andMAPs from 800 to 1200 (1300) mm. The results for theupper part are more variable, especially with respect toprecipitation—with declining CA intervals to levels of700–800 mm. The same is true to the Chersonian, wherebesides lower precipitation values a decrease in meanannual temperature and temperature of the coldestmonth is observed. These results coincide withsedimentological and vegetational data. The Paratethyanregression at the beginning of the Late Miocene alongwith shrinking water basins induced a higher seasonalityof climate and significant drying. These results agreewell with the quantitative palaeoclimatic estimates forthe Forecarpathian Basin in northwest Bulgaria (Ivanovet al. 2002), where the same trend is observed, thusconfirming our previous suggestions.
Acknowledgments
We are grateful to H.E. Wright (University ofMinnesota, USA) for revising the English. T. Utescher(Bonn, Germany) is thanked for critical reading of themanuscript. Our sincere thanks go to C.A.E. Strömberg(Stockholm, Sweden) and A. Bertini (Firenze, Italy)who greatly improved an earlier version of themanuscript by their comments and suggestions. Manythanks to P. Kersaw (Australia) and D. Uhl (Germany)for their final discussions and corrections. This work is acontribution to the international research program“Neogene Climate Evolution in Eurasia—NECLIME”and project B-1525 (NSF, Bulgaria).
Appendix A. Supplementary data
Supplementary data associated with this articlecan be found, in the online version, at doi:10.1016/j.palaeo.2007.08.003.
References
Belokris, L., 1988. Climate zonation and its change during theMiocene(Climatic-stratigraphic aspects). Geol. J. 1, 82–90 (in Russian).
Bruch, A.A., 1998. Palynologische Untersuchungen im OligozänSloweniens—Paläo-Umvelt und Paläoklima im Ostalpenraum.Tueb. mikropalaeontol. Mitt. 18, 193 S.
Bruch, A.A., Gabrielyan, I., 2002. Quantitative data of the NeogeneClimatic Development in Armenia and Nakhichevan. ActaCarolinae 46 (4), 41–48.
Bruch, A.A., Kovar-Eder, J., 2003. Climatic evaluation of the florafrom Oberdorf (Styria, Austria, Early Miocene) based onCoexistence Approach. Phytol. Balc. 9, 175–185.
Bruch, A.A., Mosbrugger, V., 2002. Palaeoclimate versus vegetationreconstruction—palynological investigations on the Oligocenesequence of the Sava Basin, Slovenia. Rev. Palaeobot. Palynol.198, 279–301.
Bruch, A.A., Utescher, T., Olivares, C.A., Dolakova, N., Ivanov, D.,Mosbrugger, V., 2004. Middle and Late Miocene spatialtemperature patterns and gradients in Europe—preliminary resultsbased on palaeobotanical climate reconstructions. Cour. Forsch.inst. Senckenb. 249, 15–27.
Bruch, A.A., Utescher, T., Mosbrugger, V., Gabrielyan, L., Ivanov, D.A.,2006. Late Miocene climate in the circum-Alpine realm—aquantitative analysis of terrestrial palaeofloras. Palaeogeogr. Palaeo-climatol. Palaeoecol. 238, 270–280.
Brayard, A., Héran, M.-A., Costeur, L., Escarguel, G., 2004. Triassic andCenozoic Palaeobiogeography: two case studies in quantitativemodel-ling using IDLÒ. Palaeontol. Electronica 7 (2) Art. 6A:22p, 1MB;http://palaeo-electronica.org/paleo/2004_2/triassic/issue2_04.htm.
Fauquette, S., Bertini, A., 2003. Quantification of the northern ItalyPliocene climate from pollen data—evidence for a very peculiarclimate pattern. Boreas 32, 361–369.
Fauquette, S., Guiot, J., Suc, J.-P., 1998. A method for climaticreconstruction of the Mediterranean Pliocene using pollen data.Palaeogeogr. Palaeoclimatol. Palaeoecol. 144, 183–201.
Fauquette, S., Suc, J.-P., Guiot, J., Diniz, F., Feddi, N., Zheng, Z.,Bessais, E., Drivaliari, A., 1999. Climate and biomes in the WestMediterranean area during the Pliocene. Palaeogeogr. Palaeocli-matol. Palaeoecol. 152, 15–36.
Fortelius, M., Eronen, J., Liu, L., Pushkina, D., Tesakov, A.,Vislobokova, I., Zhang, Z., 2006. Late Miocene and Pliocenelarge land mammals and climatic changes in Eurasia. Palaeogeogr.Palaeoclimatol. Palaeoecol. 238, 219–227.
Foster, B.L., Tilman, D., 2000. Dynamic and static views ofsuccession: testing the descriptive power of the chronosequenceapproach. Plant Ecol. 146, 1–10.
Ilyina, L.B., Shcherba, I.G., Khondkarian, S.O., Goncharova, I.A.,2004. Map 6: mid Middle Miocene (Middle Serravalian, LateBadenian, Konkian). In: Popov, S.V., Rögl, F., Rozanov, A.Y.,Steininger, F.F., Shcherba, I.G., Kovac, M. (Eds.), Lithological-Palaeogeographic maps of Paratethys. Cour. Forsch.-Inst.Senckenberg, vol. 250, pp. 23–25.
Ivanov, D., Koleva-Rekalova, E., 1999. Palynological and sedimen-tological data about late Sarmatian paleoclimatic changes in theForecarpathian and Euxinian basins (Northern Bulgaria). ActaPalaeobot. (Suppl. 2), 307–313.
Ivanov, D., Ashraf, A.R., Mosbrugger, V., Palamarev, E., 2002.Palynological evidence for Miocene climate change in theForecarpathian Basin (Central Paratethys, NW Bulgaria). Palaeo-geogr. Palaeoclimatol. Palaeoecol. 178, 19–37.
Jacobs, B.F., Kingston, J.D., Jacobs, L.L., 1999. The origin ofgrassdominated ecosystems. Ann. Mo. Bot. Gard. 86 (2), 590–643.
Jiménez-Moreno, G., Rodríguez-Tovar, F.J., Pardo-Igúzquiza, E.,Fauquette, S., Suc, J.-P., Müller, P., 2005. High-resolutionpalynological analysis in late early-middle Miocene core fromthe Pannonian Basin, Hungary: climatic changes, astronomicalforcing and eustatic fluctuations in the Central Paratethys.Palaeogeogr. Palaeoclimatol. Palaeoecol. 216 (1–2), 73–97.
Kojumdgieva, E., 1983. Palaeogeographic environment during thedessication of the Black Sea. Palaeogeogr. Palaeoclimatol.Palaeoecol. 43, 195–204.
Kojumdgieva, E., Popov, N., 1981. Regions structuro-paléogéogra-phiques en Bulgarie du Nord-Est pendant le Néogène. C.R. Acad.Bulg. Sci. 34, 1273–1275.

360 D.A. Ivanov et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 255 (2007) 342–360
Kojumdgieva, E., Popov, N., 1989. Paléogéographic et evolutiongéodinamique de la Bulgarie Septentrionale au Néogène. Geolo-gica Balc. 19, 73–92.
Kojumdgieva, E., Popov, N., Stancheva, M., Darakchieva, S., 1989.Correlation of the biostratigraphic subdivisions of the Neogene inBulgaria after molluscs, foraminifers and ostracodes. Geol. Balc.16, 9–22 Sofia (in Russian, with English abstract).
Koleva-Rekalova, E., 1994. Sarmatian aragonite sediments in North-eastern Bulgaria—origin and diagenesis. Geol. Balc. 24, 47–64.
Kostadinov, V., Kojumdgieva, E., 1964. Upper Helvetian in northeastBulgaria. Rev. Bulg. Geol. Soc. 25, 186–190 (in Bulgarian).
Kováč, M., Baráth, I., Fordinál, K., Grigorovich, A.S., Halásová, E.,Hudáčková, N., Joniak, P., Sabol, M., Slamková, M., Sliva, Ľ.,Vojtko, R., 2006. Late Miocene to Early Pliocene sedimentaryenvironments and climatic changes in the Alpine–Carpathian–Pannonian junction area: a case study from the Danube Basinnorthern margin (Slovakia). Palaeogeogr. Palaeoclimatol. Palaeoe-col. 238, 32–52.
Meulenkamp, J.E., Sissingh, W., 2003. Teriary palaeogeography andtectonostratigraphic evolution op the Northern and Southern Peri-Tethys platforms and the intermediate domains of theAfrican–Eurasianconvergent plate boundary zone. Palaeogeogr. Palaeoclimatol. Palaeoe-col. 196, 209–228.
Mosbrugger, V., 1995. New methods and approaches in Tertiarypalaeoenvironmental research. Abhandl. Staatl. Mus. Mineralog.Geol. Dresden 41, 41–52.
Mosbrugger, V., Utescher, T., 1997. The coexistence approach—amethod for quantitative reconstruction of Tertiary terrestrialpalaeoclimate using plant fossils. Palaeogeogr. Palaeoclimatol.Palaeoecol. 134, 61–86.
Mosbrugger, V., Utescher, T., Dilcher, D.L., 2005. Cenozoic continentalclimatic evolution of Central Europe. PNAS 102, 14964–14969.
Ohmann, J.L., Spies, T.A., 1998. Regional gradient analysis andspatial pattern of woody plant communities of Oregon forests.Ecol. Monogr. 68, 151–182.
Palamarev, E., 1989. Palaeobotanical evidences of the Tertiary historyand origin of the Mediterranean sclerophyll dendroflora. PlantSyst. Evol. 162, 93–107.
Palamarev, E., 1990. Grundzuge der paläofloristischen Paläosukzes-sionen im Spätmiozän (Sarmatien-Pontien) Bulgariens. In:Knobloch, E., Kvačec, Z. (Eds.), Paleofloristic and paleoclimaticchanges in the Cretaceous and Tertiary, Prague, pp. 257–263.
Palamarev, E. 1991. Composition, structure and main stages in theevolution of Miocene paleoflora in Bulgaria. Autoref. DSc thesis,BAS, Sofia, 1–60, (in Bulgarian).
Palamarev, E., Ivanov, D., 1998. über einige Besonderheiten der tertiärenFloren in Bulgarien und ihre Bedeutung für die Entwicklungs-geschichte der Pflanzenwelt in Europa. Acta Palaeobot. 38, 147–165.
Palamarev, E., Ivanov, D., 2001. Charakterzüge der Vegetation derSarmatien (Mittel-bis Obermiozän) im südlichen Teil des DazischenBeckens (Südost Europa). Palaeontographica, B 259, 209–220.
Palamarev, E., Ivanov, D., 2004. Badenian vegetation of Bulgaria:biodiversity, palaeoecology and palaeoclimate. Cour. Forsch.inst.Senckenb. 249, 63–69.
Paramonova, N.P., Shcherba, I.G., Khondkarian, S.O., 2004. Map 7: lateMiddleMiocene (Late Serravallian, Sarmatian s.s.,Middle Sarmatians.l. In: Popov, S.V., Rögl, F., Rozanov, A.Y., Steininger, F.F.,Shcherba, I.G., Kovac, M. (Eds.), Lithological-Palaeogeographicmaps of Paratethys. Cour. Forsch.-Inst. Senckenberg, vol. 250,pp. 27–29.
Popov, N., Kojumdgieva, E., 1987. The Miocene in NortheasternBulgaria (lithostratigraphic subdivision and geological evolution).Rev. Bulg. Geol. Soc. 48, 15–33 (in Bulgarian).
Popov, N., Stancheva, M., Darakchieva, S., 1986. Reference sectionsof the Neogene from Northeastern Bulgaria. Palaeontol., Stratigr.Lithol. 23, 25–45 (in Bulgarian).
Rögl, F., 1998. Palaeogeographic considerations for Mediterraneanand Paratethys seaways (Oligocene to Miocene). Ann. Naturhist .Mus. Wien 99A, 279–310.
Rögl, F., 2001. Mid-Miocene Circum-Mediterranean palaeogeogra-phy. Ber. Inst. Geol. Palaeontol. Karl-Franzens Univ. Graz Oesterr.4, 49–59.
Stuchlik, L., Ivanov, D., Palamarev, E., 1999. Middle and LateMiocene floristic changes in the Northern and Southern parts of theCentral Paratethys. Acta Palaeobot. (Suppl. 2), 391–397.
Suc, J.-P., Drivaliari, A., 1991. Transport of bisaccate coniferous fossilpollen grains to coastal sediments: An example from the earliestPliocene Orb ria (Languedoc, southern France). Rev. Palaeobot.Palynol. 70, 247–253.
Syabryay, S.V., Stekina, N.A., 1983. History of the development of thevegetation of Ukraine during the Miocene. Naukovaya Dumka,Kiev. 166 pp. (in Russian).
Temniskova-Topalova, D. 1994. Miocene diatom floras in Bulgaria—composition, structure, palaeoecology, and biostratigraphy. Auto-ref. DSc Thesis, Sofia University, 68 p. Sofia (in Bulgarian).
Uhl, D., Mosbrugger, V., Bruch, A.A., Utescher, T., 2003. Recon-structing palaeotemperatures using leaf floras—case studies for acomparison of leaf margin analysis and the coexistence approach.Rev. Palaeobot. Palynol. 126, 49–64.
Utescher, T., Mosbrugger, V., Ashraf, A.R., 2000. Terrestrial climateevolution in Northwest Germany over the last 25 million years.Palaios 15, 430–449.
van Dam, J., 2006. Geographic and temporal patterns in the lateNeogene (12–3 Ma) aridification of Europe. The use of smallmammals as paleoprecipitation proxies. Palaeogeogr. Palaeocli-matol. Palaeoecol. 238, 190–218.
Velev, S., 1997. Contemporary air temperature and precipitationfluctuations in Bulgaria. In: Jordanova, M., Donchev, D. (Eds.),Geography of Bulgaria. Acad. Publ. House, Sofia, pp. 145–150(In Bulgarian).
Walter, H., 1973. Vegetation of the Earth on relation to climate andecophysiological conditions. Heidelberg Science Library, vol. 15.English Universities Press Ltd., London.
Wolfe, J.A., 1993. A method of obtaining climatic parameters fromleaf assemblages. U. S. Geol. Surv. Bull. 2040, 1–71.
Zhang, J.-T., 2005. Succession analysis of plant communities inabandoned croplands in the eastern Loess Plateau of China. J. AridEnviron. 63, 458–474.
Copyright © 2022 FDOKUMEN




















