
Transsexual limb transplants in fiddler crabs and expression of novel sensory capabilities
Upload
independentCategory
view
1download
0
NEW SPECIES AND NEW RECORDS OF ENDEMIC FRESHWATER CRABS FROM THE ATLANTIC
FOREST IN SOUTHERN BRAZIL (ANOMURA: AEGLIDAE)
Georgina Bond-Buckup, Carlos G. Jara, Ludwig Buckup, Marcos Perez-Losada, Alessandra A.P. Bueno,Keith A. Crandall, and Sandro Santos
(GBB, LB) Departamento de Zoologia, Instituto Biociencias, Universidade Federal do Rio Grande do Sul, Av.Bento Goncalves, 9500,
90501-970, Porto Alegre, RS, Brazil;
(CGJ) Instituto de Zoologia, Casilla 567, Universidad Austral de Chile, Valdivia, Chile;
(MPL) CIBIO, Centro de Investigacao em Biodiversidade e Recursos Geneticos, Universidade do Porto, Campus Agrario de Vairao,
4485-661, Vairao, Portugal;
(AAPB) Universidade Federal de Lavras, Departamento de Biologia, Campus Universitario, 37200-000 Lavras, MG, Brazil;
(KAC) Department of Biology, Brigham Young University, Provo UT 84602-5255, USA;
(SS, correspondence, [email protected]) Departamento de Biologia, Universidade Federal de Santa Maria, 97105-900,
Santa Maria, RS, Brazil
A B S T R A C T
Two new species of freshwater anomurans, Aegla pomerana and Aegla muelleri (Decapoda: Anomura: Aeglidae), are described from the
Itajaı River basin, the major basin of the Atlantic range, located in the northeastern part of the state of Santa Catarina, Brazil. The new
taxa can be distinguished from their congeners based on both morphological and molecular evidence (the nuclear gene 28S, and the
mitochondrial genes 12S, 16S, COI, and COII). Based on molecular data, A. pomerana has a phylogenetic relationship with A.
leptodactyla, but morphologically these two species differ in several characters. Aegla muelleri is a member of the same clade as A.
leptochela, but several morphological characters distinguish the two species. New records of occurrence of A. jarai and A. odebrechtii,
which occur in the same hydrographic basin, are provided.
KEY WORDS: Aegla, Anomura, biogeography, Brazil, Itajaı basin, molecular systematics
DOI: 10.1651/09-3186.1
INTRODUCTION
The Atlantic Forest is considered one of the regions withthe highest biodiversity in the world, a hotspot area(Mittermeier et al., 2004). The forest originally extendedalong almost the entire coast of Brazil, from the states ofPiauı to Rio Grande do Sul. It covers an area of1,300,000 Km2, which corresponds to 15% of the territoryof the country, of which only 7% remains preserved.
Even today the rivers and lakes of this forest shelters richaquatic ecosystems, many of them threatened by thedestruction of the gallery forests and consequent siltationof the water sources, by water pollution, and by theconstruction of reservoirs without due care for theenvironment. This intricate mosaic of basins is formed byrivers of national and regional importance, such as the RioItajaı in the state of Santa Catarina. The basin of the RioItajaı is the largest basin of the Atlantic drainage in the stateof Santa Catarina, covering approximately 15,500 km2,equivalent to 16.5% of the area of the state (Fig. 1). Theregion has 53 municipalities occupying its territory, ofwhich 47 sit within the basin, where about a millioninhabitants live. In the Itajaı Valley, it is still possible tofind remnants of the Atlantic Forest and watercourses in agood state of conservation, as is the case for the Serra doItajaı National Park, in the municipality of Indaial.
Living in rivers, creeks, and lakes, Aeglidae areconsidered good indicators of environmental quality, beingfound only in localities with clean, well-oxygenated water
(Bond-Buckup, 2003). The first species of this familydescribed from southern Brazilian continental waters wasAegla odebrechtii Muller 1876, recorded in streams thatdescend the Serra do Mar and enter the Rio Itajaı (Bond-Buckup and Buckup, 1994). Presently, six additionalspecies of Aegla are recorded from the state: A. franciscanaBuckup and Rossi 1977, A. platensis Schmitt1942, A.rossiana Bond-Buckup and Buckup 1994, A. spinosa Bond-Buckup and Buckup 1994, A. jarai Bond-Buckup andBuckup 1994 and A. parva Bond-Buckup and Buckup1994. These last two have already been recorded in thebasin of the Rio Itajaı (Bond-Buckup, 2003; Boos et al.,2006).
In a recent survey of the Aeglidae of South America, theRio Itajaı Basin was investigated in a most detailed manner,and the results are presented here. Two new species aredescribed, and the distributions of A. jarai and A.odebrechtii are reviewed and extended.
MATERIAL AND METHODS
The specimens were collected in watercourses of the Rio Itajaı-Acu Basin,during several sampling campaigns and were deposited in the Collection ofCrustaceans of the Department of Zoology, Institute of Biosciences,Federal University of Rio Grande do Sul, Porto Alegre, Brazil (collectioncode: UFRGS) and in Museu de Zoologia da Universidade de Sao Paulo(MZUSP). The descriptions of the new species were based on examinationof the type-series. Measurements were made according to the method ofBond-Buckup and Buckup (1994), with the following abbreviations: ‘‘m’’5 males, ‘‘f’’ 5 females, ‘‘f ov’’ 5 ovate females, ‘‘j’’ 5 juveniles, CL 5
JOURNAL OF CRUSTACEAN BIOLOGY, 30(3): 495-502, 2010
495

total cephalothorax length (between the tip of rostrum and the midpoint ofthe posterior margin of the carapace), PCW 5 precervical width (betweenthe left and right epibranchial margins), FW 5 frontal width (between thetips of the spines of the anterolateral angles of the carapace), AL 5 areolalength, AW 5 areola width, RL 5 rostrum length (between the tip of therostrum and the midpoint of the orbital margin.
Morphometric Analysis
For the description of the new species, the following body dimensionswere recorded for all animals of the type series, and also for the non-paratype animals: TCL, CL, PCW, FW, AL, AW and RL. Themorphometric ratios TCL/RL, CL/RL, PCW/FW and AL/AW werecalculated for the set of animals from each locality.
Molecular Analysis
The molecular data and phylogenetic analyses presented in this study wereobtained from Perez-Losada et al. (2009). However, at the time of thatpublication, the taxonomic status of two species described here wasunknown. Examination of the lots collected in the Rio Itajaı-Acuhydrographic basin allowed us to verify two new species and to addnew localities of occurrence, with the respective geographical coordinates,of two species of previously described aeglids.
SYSTEMATICS
Aeglidae Dana, 1852Aegla pomerana n. sp. Bond-Buckup and Buckup
(Fig. 2)
Type-material.—Holotype male, Brazil, state of SantaCatarina, Pomerode, Sıtio Mundo Antigo tributary in theRio Itajaı-Acu Basin, 26u469220S, 49u119260W; 276 ma.s.l., 18.viii.1999, G. Bond-Buckup and L. Buckup col.(MZUSP 20463).
Paratypes: 2 m, 1 f with juveniles, 2 j, same data asholotype (UFRGS 2679P); 1 m, same data as holotype,02.i.2001, F. Faraco col. (UFRGS 2993); 4 m, 1 f, CampoAlegre, Joinvile, i.2002 (UFRGS 3262).
Diagnosis.—Antero-lateral spine of carapace extendingbeyond base of cornea; protogastric lobes present; extra-orbital sinus absent, rostrum triangular, slightly ligulate,slightly recurved distally, lacking carena on distal third, outerproximal margin of movable finger of cheliped with lobe;palmar crest of cheliped subrectangular; anterior angle ofventral margin of epimeron 2 unarmed; inner margin ofventral surface of ischium of cheliped with a distal tubercle.
Description.—Carapace moderately compressed dorsoven-trally, area of gastric region more elevated, dorsal surfacescabrous, covered with punctae. Front wide; LPC/LF ratioof holotype male 1.87.
Rostrum triangular, slightly ligulate, elevated in medianportion, slightly recurved distally, without carena on distalthird. Sub-rostral process developed; in profile, rostrumwith ventral portion deeper than dorsal. Rostral carenabegins between protogastric lobes, with two parallel rowsof scales that do not reach apex; excavated in medianportion. Lateral margins of rostrum with small scales.
Orbits wide, deep, without orbital spine. Orbital marginwith small sparse scales. Extraorbital sinus lacking.
Antero-lateral angle of carapace projecting anteriorly ina spine, which extends beyond base of cornea. Outer andinner margins of antero-lateral lobe with sparse scales. Firsthepatic lobe delimited anteriorly by a distinct fissure;lateral margin with scales; second and third hepatic lobesnot delimited, with only a slight indication; lateral marginswith scales.
Epigastric prominences little pronounced, surface irreg-ular, of indefinite shape, elongated toward base of firsthepatic lobe, with sparse scales. Protogastric lobesmoderately elevated, anterior margin marked by row ofscales.
Transverse dorsal line sinuous. Areola subquadrate tosubrectangular. CA/LA ratio of holotype male 1.5.
Epibranchial area triangular, with an apical tuberclefollowed by scales. Lateral margins of anterior andposterior branchial areas with subequal scales.
Anterior angle of ventral margin of epimeron 2 unarmed,with only small scales; ventrolateral margin slightlyconvex; posterior angle of ventral margin obtuse, unarmed.Epimera of third to sixth segments projecting; on third andfourth the lateral projection ornamented with a small apicalscale.
Telson divided by longitudinal suture.Anterior extremity of third sternite truncate, projected
between the coxae of the exopods of the third maxillipeds.Fourth thoracic sternite elevated in median region, withoutornament, lateral margins slightly recurved.
Chelipeds subequal, hand subrectangular. Smaller cheladelicate in appearance, elongated, covered by scales.Larger cheliped with more globose appearance, palmslightly more inflated in posterolateral region, coveredwith corneal scales. Palmar crest subrectangular, withmargin ornamented with scaliform tubercles, indicatinglobes. Pre-dactylar lobe forming a small angle with anteriormargin of propodus, ornamented with scales. Fingersslender, covered by setae, scales, and scaliform tubercles.Proximal outer margin of movable finger with distinct lobetipped with scales. Prehensile margins of fingers withscaliform denticles on their entire length, and withpronounced fitted opposed lobular teeth. Dorsal surfaceof carpus scabrous, with scales; inner margin with fourspines, with distalmost spine being most developed; thesespines with sparse scales on the lateral margins; small spinebetween the distal spine and the inner antero-lateral angleof the carpus; inner antero-lateral angle sub-obtuse, withapical spine; anterodorsal margin with scales. Distal part of
Fig. 1. Drainage basin of the Rio Itajaı, in the state of Santa Catarina. 15 Municipality of Pomerode; 2 5 Municipality of Indaial; 3 5Municipality of Blumenau.
496 JOURNAL OF CRUSTACEAN BIOLOGY, VOL. 30, NO. 3, 2010

Fig. 2. Aegla pomerana, n. sp. a: male holotype (MZUSP 20463), dorsal view; b: precervical portion of carapace, lateral view; c: ischium of cheliped,ventral view; d: third and fourth sterna, ventral view; e: second pleomere epimeron.
BOND-BUCKUP ET AL.: NEW SPECIES OF AEGLIDAE 497

dorsal surface with depression parallel to distal margin.Carpal crest pronounced, more elevated in proximal region,with clumps of three to five scales on elevations of crest;outer ventral angle of carpus with scales; ventral surfacewith one conical spine and tufts of setae. Dorsal margin ofmerus of cheliped with one spine, remainder of margin withscaliform tubercles; antero-dorsal margin with scales.Lateral surfaces with sparse punctae. Inner ventral marginof merus with one distal spine followed by scaliformtubercles; outer ventral margin with pronounced distaltubercle followed by scales. Dorsal margin of ischium withone spine; inner margin of ventral surface with small distalspine, followed by several spines along margin. Dorsalmargin of dactylus, propodus, and carpus of second, third,and fourth pereiopods with rows of setae, scales, andscaliform tubercles arranged in longitudinal series, lendingthe surface a pubescent appearance.
Variations.—The rostrum is more excavated in the largerparatypes, and the larger chela is more globose.
Morphometry.—Holotype male with LC 16.89 mm. Para-types (n 5 5) with mean LC 13.60 mm. A small-sizedspecies. LPC/LF ratios of paratypes (n 5 3) ranging from1.50 to 1.90.
CA/LA ratios of paratypes varying little, from 1.7 to 1.8.
Distribution.—Brazil: State of Santa Catarina, Rio Itajaı-Acu Basin.
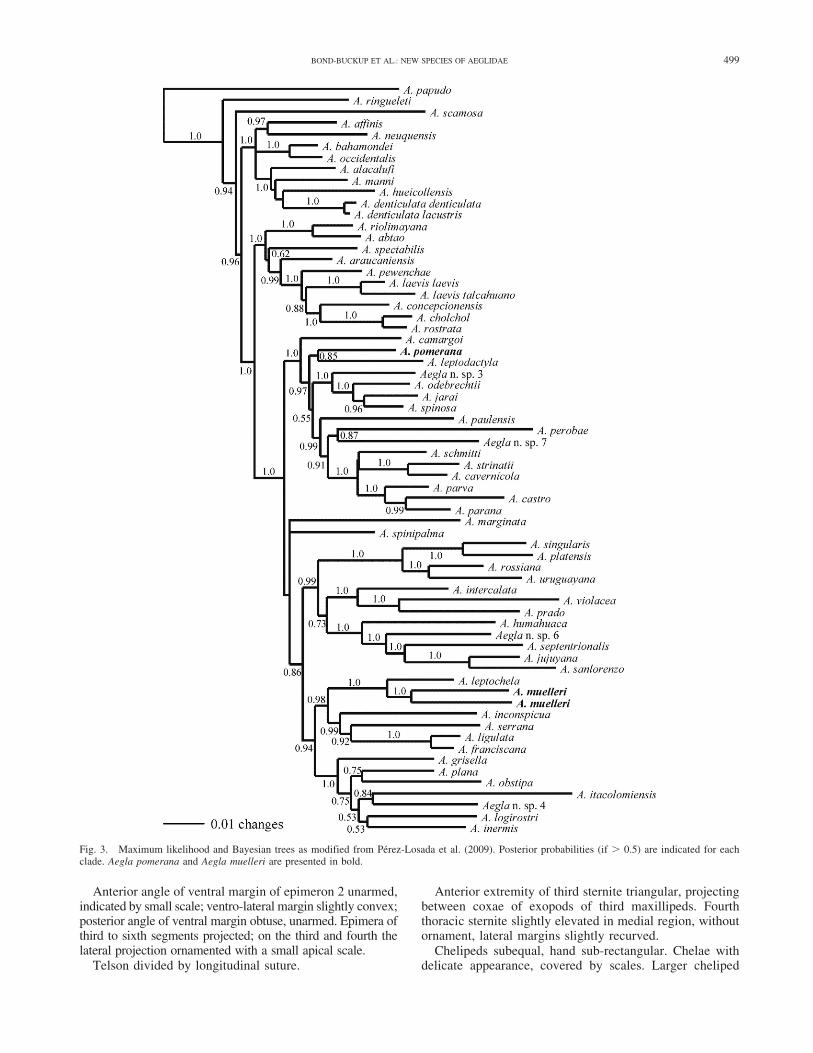
Remarks.—The Bayesian phylogenetic analysis of themolecular data showed that A. pomerana belongs to CladeC as defined in Perez-Losada et al. (2004). With regard tothe relationship to other species, this was considered asister-species of A. leptodactyla (Fig. 3), which occurs inthe Rio Uruguay basin on the high plateau of Rio Grandedo Sul and Santa Catarina. Nevertheless, there are severalmorphological differences between these two species. A.pomerana does not bear an extra-orbital sinus, whereas A.leptodactyla possesses this character. The rostrum of A.pomerana is only slightly ligulate, recurved, lacks a carinaon the distal third, and ends without a spine on the tip;whereas this structure in A. leptodactyla is of mediumlength, and its distal end terminates in a spine. A. pomeranahas wide chelipeds; its sister species has narrow chelipeds.
The relationship of A. pomerana with species found inthe Rio Uruguay basin, which at present has no directconnection to the Rio Itajaı basin, can be attributed to acommon geological origin of the two basins, which is relatedto the formation of the Serra Geral in southern Brazil(Ribeiro, 2006). The uplift of the Serra Geral separated theUruguay and Itajaı-Acu rivers creating the opportunity fornew species to arise through allopatric speciation.
Morphologically, A. pomerana resembles A. marginataBond-Buckup and Buckup, 1994, in some aspects,particularly in the slightly arched antero-lateral border ofthe carapace. However, A. pomerana has a subrectangularpalmar crest, and A. marginata has no palmar crest.
Biology.—Unknown.
Etymology.—Named in honor of the inhabitants ofPomerode, the type-locality of the species.
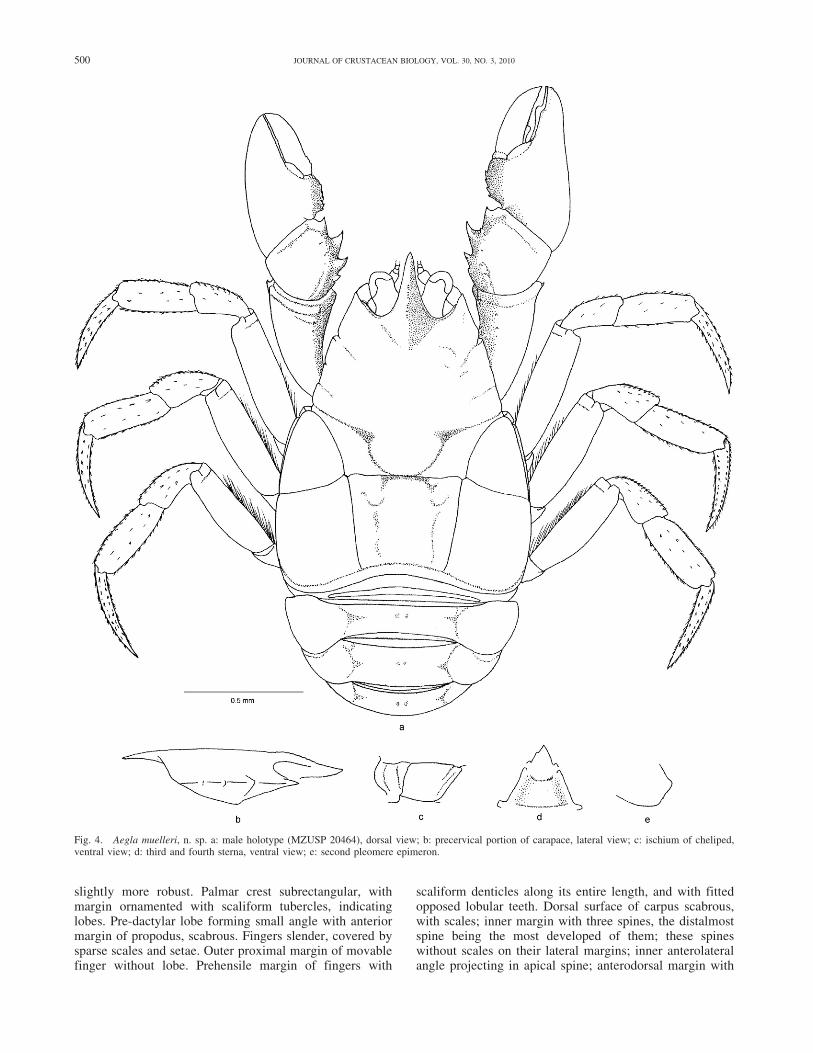
Aegla muelleri n. sp. Bond-Buckup and Buckup(Fig. 4)
Type-material.—Holotype male, Brazil, state of SantaCatarina, Indaial, Serra do Itajaı National Park, Rio Itajaı-Acu basin, 27u019240S, 49u099060W; ii.2006, H. Boos col.(MZUSP 20464).
Paratypes: 1 m, 6 f, same data as holotype (UFRGS4117P); 2 m, 2 f, same data as holotype, 11.v.2001, H. Boosand G. Bond-Buckup col. (UFRGS 2982); id., 2 m,21.ix.2002 (UFRGS 3570); id., juvenile (UFRGS 3571);1 m, Cascata do Rio Passa Quatro, Monte Castelo, RioItajaı-Acu basin, 26o289140S, 50u119590W; 24.x.2000,A.A.P. Bueno, G. Bond-Buckup, C. Jara and M. Perez-Losada col. (UFRGS 2979); id., ib., ib., 11 m , 11 f , 1 f ov,5 j (UFRGS 2980).
Material Examined.—Brazil, Santa Catarina, EspingardaCreek, Serra do Itajaı National Park, Rio Itajaı-Acu basin,27u01924.10S, 49u0996.70W; 20.vi.2002, H. Boos and K.Schacht col., 4 dissected specimens, (UFRGS 3385).
Diagnosis.—Antero-lateral spine of carapace reaching baseof cornea; protogastric and epigastric lobes absent; extra-orbital sinus absent, rostrum triangular, tapered, slightlyrecurved distally, carinate, proximal outer margin ofmovable finger of cheliped without lobe; palmar crest ofcheliped small, subrectangular; anterior angle of ventralmargin of epimeron 2 with small scale; inner margin ofventral surface of ischium of cheliped with distal scaliformtubercle.
Description.—Carapace strongly compressed dorsoventral-ly, dorsal surface scabrous, covered by punctations andscales; anterior dorsal region delimiting base of rostrum byV-shaped depression.
Front wide; LPC/LF ratio of holotype male 1.73.Rostrum long, triangular, tapering, slightly recurved
distally, carinate, distal part of rostrum slightly recurved.Sub-rostral process well developed; in profile, ventralportion of rostrum deeper than dorsal portion. Rostralcarina begins at height of orbits, with two juxtaposed rowsof scales that reach to apex; carina elevated in middleportion and very low in distal third. Lateral margins ofrostrum scabrous.
Orbits wide and deep, orbital spine indicated by anincision. Orbital margin scabrous. Extra-orbital sinusabsent.
Antero-lateral angle of carapace projecting anteriorly ina spine, extending beyond base of cornea. Outer and innermargins of antero-lateral lobe with sparse scales. Firsthepatic lobe delimited anteriorly by small incision; lateralmargin smooth; 2nd hepatic lobe with small indication, and3rd not delimited; lateral margins scabrous. Epigastricprominences lacking, surface scabrous. Protogastric lobeslacking, without indication.
Transverse dorsal line sinuous. Areole quadrate. CA/LAratio of holotype male 1.34.
Epibranchial area triangular, with an apical tuberclefollowed by scales. Lateral margins of anterior andposterior branchial area with subequal scales.
498 JOURNAL OF CRUSTACEAN BIOLOGY, VOL. 30, NO. 3, 2010
Anterior angle of ventral margin of epimeron 2 unarmed,indicated by small scale; ventro-lateral margin slightly convex;posterior angle of ventral margin obtuse, unarmed. Epimera ofthird to sixth segments projected; on the third and fourth thelateral projection ornamented with a small apical scale.
Telson divided by longitudinal suture.
Anterior extremity of third sternite triangular, projectingbetween coxae of exopods of third maxillipeds. Fourththoracic sternite slightly elevated in medial region, withoutornament, lateral margins slightly recurved.
Chelipeds subequal, hand sub-rectangular. Chelae withdelicate appearance, covered by scales. Larger cheliped
Fig. 3. Maximum likelihood and Bayesian trees as modified from Perez-Losada et al. (2009). Posterior probabilities (if . 0.5) are indicated for eachclade. Aegla pomerana and Aegla muelleri are presented in bold.
BOND-BUCKUP ET AL.: NEW SPECIES OF AEGLIDAE 499
slightly more robust. Palmar crest subrectangular, withmargin ornamented with scaliform tubercles, indicatinglobes. Pre-dactylar lobe forming small angle with anteriormargin of propodus, scabrous. Fingers slender, covered bysparse scales and setae. Outer proximal margin of movablefinger without lobe. Prehensile margin of fingers with
scaliform denticles along its entire length, and with fittedopposed lobular teeth. Dorsal surface of carpus scabrous,with scales; inner margin with three spines, the distalmostspine being the most developed of them; these spineswithout scales on their lateral margins; inner anterolateralangle projecting in apical spine; anterodorsal margin with
Fig. 4. Aegla muelleri, n. sp. a: male holotype (MZUSP 20464), dorsal view; b: precervical portion of carapace, lateral view; c: ischium of cheliped,ventral view; d: third and fourth sterna, ventral view; e: second pleomere epimeron.
500 JOURNAL OF CRUSTACEAN BIOLOGY, VOL. 30, NO. 3, 2010

scales. Dorsal surface distally with small depressionparallel to distal margin. Carpal crest distinct, moreelevated in proximal region; medial and distal portionswith scales clumped in groups of three to five on elevationsof crest; outer ventral angle of carpus obtuse, unarmed;ventral surface with small conical spine. Dorsal margin ofmerus of cheliped with elevations tipped with scales, whichdecrease in size proximally, followed by long setae; antero-dorsal margin with sparse scales. Lateral surfaces withpunctae and scales. Inner ventral margin of merus with adistal spine followed by scaliform tubercles and scales;prominent distal tubercle followed by scales on outerventral margin. Dorsal margin of ischium with conicaltubercle and long setae; inner margin of ventral surface ofischium elevation tipped with scale.
Dorsal margins of dactylus, propodus, and carpus ofsecond, third, and fourth pereiopods with rows of setae andscales arranged in longitudinal rows. Dorsal margins ofsecond, third, and fourth pereiopods covered with longsetae, lending the surface a pubescent appearance.
Variations.—In some paratypes, the protogastric lobes aredelimited anteriorly by sparse scales, although these arelacking in other specimens. Proximal outer margin ofmovable finger, in some specimens, bears scales suggestinga lobe. Other paratypes do not possess fitted opposedlobular teeth on the prehensile margin of the fingers, butonly scaliform denticles on their entire length. The lotUFRGS 2979 has more-developed tubercles on thepereiopods, and the palmar crest is slightly more distinct.This is a small-sized species, comparable to A. violaceaBond-Buckup and Buckup, 1994 and A. leptodactyla.
Morphometry.—Holotype male with LC 12.20 mm. Theovigerous female measured 13.29 mm LC. The mean LC of25 paratypes was 11.52 mm, ranging from 8.0 to 15.33 mm.
LPC/LF ratio of paratypes (n 5 10) ranging from 1.71 to1.91, i.e., characterizing a wide front. CA/LA ratio ofparatypes (n 5 11) ranging from 1.14 to 1.32.
Etymology.—Named in honor of Fritz Muller, the firstscientist to describe an aeglid from Brazilian waters.
Biology.—Unknown.
Distribution.—Brazil: state of Santa Catarina, Rio Itajaı-Acu Basin.
Remarks.—The new species A. muelleri resembles A. planaBuckup and Rossi, 1977 in the dorsoventrally compressedcephalothorax and the absence of the epigastric andprotogastric lobes. However it is distinguished from A.plana by the shape of the chelipeds, which are wider in A.plana; by the configuration of the spines of the carpus; andby the shape of the palmar crest, which is more developedin A. plana. This latter species also possesses an extra-orbital spine, which is lacking in A. muelleri.
The total length of the cephalothorax of the paratypes (n5 7) reached only 13.43 mm, showing it to be a small-sizedspecies.
The species occurs in sympatry with A. jarai in theRibeirao Garcia in the Parque das Nascentes, an environ-
mental preservation area that is among the few localitiesthat shelter species in Atlantic Forest.
The Bayesian phylogenetic analysis of the moleculardata showed that A. muelleri belongs to Clade E as definedin Perez-Losada et al. (2004). Aegla muelleri has a sisterrelationship to A. leptochela (Fig. 3), with which it sharesthe delicate form of the chelipeds and the convexity of thecarapace. However, the two species differ in relation to theantero-lateral spine of the carapace, which in A. muellerireaches the base of the cornea, and in A. leptochela doesnot. In A. muelleri, the extra-orbital sinus and the lobe ofthe movable finger are lacking, and the palmar crest is sub-rectangular; whereas in A. leptochela the extra-orbital sinusas well as the lobe of the movable finger are present, andthe palmar crest is lacking.
Aegla jarai Bond-Buckup and Buckup, 1994
Diagnosis.—Antero-lateral spine of carapace extendingbeyond middle of cornea; protogastric lobes subtle; rostrumvery long in adult males, styliform, carinate along its entirelength; anterior angle of ventral margin of epimeron 2unarmed; outer proximal margin of movable finger ofcheliped with lobe tipped with tubercle; palmar crest ofcheliped disc-shaped, strongly excavated; inner margin ofventral surface of ischium of cheliped with one conicaldistal spine and up to three tubercles; dorsal margin ofmerus of second pereiopod with spine, followed bytubercles; ventral margin of merus of second pereiopodwith scaliform tubercles.
Formerly Known Distribution.—Brazil: Central and south-ern part of state of Santa Catarina, and northern part of stateof Rio Grande do Sul (Bond-Buckup, 2003).
New Records.—Brazil: State of Santa Catarina: Rio Itajaıbasin: Rio Itajaı-Acu: Ribeirao Encoro, Indaial, 13.v.2000,H. Boos col., 2 m, 5 f (UFRGS 2948); Ribeirao Espingarda,Indaial, 06.x.2000, H. Boos col., 2 m, 2 f (UFRGS 2991);Ribeirao Caete, Cascata Ferdinand Schaback, Blumenau,03.iii.2001, H. Boos col., 2 f, 1 j (UFRGS 3203). Theoccurrence of A. jarai in the sub-basin of the Rio Itajaı-Acubroadens its known geographical distribution to northeast-ern Santa Catarina.
Biology.—Aspects of the growth of this species werecharacterized by Boos et al. (2006).
Remarks.—The phylogeny of the aeglids shows that A.jarai belongs to Clade C, with an estimated divergencetime of 33.2 6 1.8 MYA Perez-Losada et al. (2004). Theseveral populations of A. jarai form a nonmonophyleticgroup, according to these authors, indicating that newsystematic studies on this species are needed.
Aegla odebrechtii Muller, 1876
Diagnosis.—Antero-lateral spine of carapace reachingmiddle of cornea; protogastric lobes obsolete; rostrum ofmedium length in adult males, deflected and slightlyrecurved distally, without carina on distal third; anterior
BOND-BUCKUP ET AL.: NEW SPECIES OF AEGLIDAE 501

angle of ventral margin of epimeron 2 unarmed; proximalouter margin of movable finger of cheliped with lobe tippedwith tubercles; palmar crest of cheliped subdisciform,excavated; inner margin of ventral surface of ischium ofcheliped with one distal conical spine, one proximalsmaller spine, and up to three tubercles between them.
Formerly Known Distribution.—Brazil: The majority ofrecords refer to the central region of the state of SantaCatarina, and the northern part of Rio Grande do Sul. In theRio Itajaı basin, there is only the record of the holotypedescribed from the Rio Itajaı-Acu, municipality ofBlumenau.
New Records.—Brazil: Santa Catarina: Rio Itajaı basin, atributary of the Rio Itajaı do Sul (Fig. 1), Atalanta,16.viii.1999, G. Bond-Buckup and L. Buckup col., 7 m, 2f, 3 j (UFRGS 2676); Ribeirao Belchior, a tributary of RioItajaı-Acu, Gaspar, 14.vi. 1986, A da Silva col., 3 m, 2 f(UFRGS 2390).
Biology.—Unknown.
Remarks.—The phylogeny of the group, presented byPerez-Losada et al. (2004), showed that A. odebrechtiibelongs to clade C, with A. jarai as the sister species, and adivergence time estimated at 33.2 6 1.8 MYA.
ACKNOWLEDGEMENTS
We thank our fellow biologists who participated in the samplingcampaigns, especially Harry Boos Jr., Karin Schacht, and Fabio Faraco.We also thank CNPq for a productivity grant to GBB and SS, the USNational Science Foundation (OISE-0530267, DEB-0075600) and theNational Geographic Society for partial funding of this work.
REFERENCES
Bond-Buckup, G. 2003. Famılia Aeglidae, pp. 21-116. In, G.A.S. Melo(ed.), Manual de identificacao dos Crustacea Decapoda, de agua doce doBrasil. Editora Loyola, Sao Paulo.
———, and L. Buckup. 1994. A famılia Aeglidae (Crustacea, Decapoda,Anomura). Arquivos de Zoologia 32: 159-347.
Buckup, L., and A. Rossi. 1977. O genero Aegla no Rio Grande do Sul,Brasil (Crustacea, Decapoda, Anomura, Aeglidae). Revista Brasiliero deBiologia 37: 879-892.
Boos Jr., H., D. Silva-Castiglioni, K. Schacht, L. Buckup, and G. Bond-Buckup. 2006. Crescimento de Aegla jarai Bond-Buckup and Buckup(Crustacea, Anomura, Aeglidae). Revista Brasileira de Zoologia 23:490-496.
Mittermeier, R.A., P.R. Gil, M. Hoffmann, J. Pilgrim, T. Brooks, C.G.Mittermeier, J. Lamoreux, and G.A.B. Fonseca. 2004. HotspotsRevisited: Earth’s Biologically Richest and Most Endangered TerrestrialEcoregions. CEMEX/Agrupacion Sierra Madre. 432 pp.
Muller, F. 1876. Aegla odebrechtii n. sp. Jenaische Zeitschrift furNaturwissenschaft, n.s., Jena 10(3): 13-24.
Perez-Losada, M., G. Bond-Buckup, C.G. Jara, and K.A. Crandall. 2004.Molecular systematics and biogeography of the southern SouthAmerican fresh-water ‘‘crabs’’ Aegla (Decapoda: Anomura: Aeglidae)using multiple heuristic tree search approaches. Systematic Biology 53:767-780.
———, ———, ———, and ———. 2009. Conservation Assessment ofSouthern South American Freshwater Ecoregions on the Basis of theDistribution and Genetic Diversity of Crabs from the Genus Aegla.Conservation Biology, 23: 692-702.
Ribeiro, A. C. 2006. Tectonic history and the biogeography of thefreshwater fishes from the coastal drainages of eastern Brazil: anexample of faunal evolution associated with a divergent continentalmargin. Neotropical Ichthyology 4: 225-246.
Schmitt, W.L. 1942. The species of Aegla, endemic South Americancrustaceans. Proceedings of the United States National Museum 91:431-524.
RECEIVED: 9 June 2009.ACCEPTED: 18 November 2009.
502 JOURNAL OF CRUSTACEAN BIOLOGY, VOL. 30, NO. 3, 2010
Copyright © 2022 FDOKUMEN




















