
Journal of Natural History The sentinel crabs of the genus Chaenostoma (Stimpson, 1858) (Crustacea:...
15
This article was downloaded by: [University of Tehran] On: 07 October 2013, At: 05:36 Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK Journal of Natural History Publication details, including instructions for authors and subscription information: http://www.tandfonline.com/loi/tnah20 The sentinel crabs of the genus Chaenostoma (Stimpson, 1858) (Crustacea: Brachyura: Macrophthalmidae), with description of a new species and new records Reza Naderloo a a Forschungsinstitut Senckenberg , Frankfurt am Main , Germany Published online: 05 Sep 2013. To cite this article: Reza Naderloo , Journal of Natural History (2013): The sentinel crabs of the genus Chaenostoma (Stimpson, 1858) (Crustacea: Brachyura: Macrophthalmidae), with description of a new species and new records, Journal of Natural History, DOI: 10.1080/00222933.2013.807948 To link to this article: http://dx.doi.org/10.1080/00222933.2013.807948 PLEASE SCROLL DOWN FOR ARTICLE Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) contained in the publications on our platform. However, Taylor & Francis, our agents, and our licensors make no representations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of the Content. Any opinions and views expressed in this publication are the opinions and views of the authors, and are not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon and should be independently verified with primary sources of information. Taylor and Francis shall not be liable for any losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoever or howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use of the Content. This article may be used for research, teaching, and private study purposes. Any substantial or systematic reproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any form to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http://www.tandfonline.com/page/terms- and-conditions
Transcript of Journal of Natural History The sentinel crabs of the genus Chaenostoma (Stimpson, 1858) (Crustacea:...

This article was downloaded by: [University of Tehran]On: 07 October 2013, At: 05:36Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registeredoffice: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
Journal of Natural HistoryPublication details, including instructions for authors andsubscription information:http://www.tandfonline.com/loi/tnah20
The sentinel crabs of thegenus Chaenostoma (Stimpson,1858) (Crustacea: Brachyura:Macrophthalmidae), with description ofa new species and new recordsReza Naderloo aa Forschungsinstitut Senckenberg , Frankfurt am Main , GermanyPublished online: 05 Sep 2013.
To cite this article: Reza Naderloo , Journal of Natural History (2013): The sentinel crabs of thegenus Chaenostoma (Stimpson, 1858) (Crustacea: Brachyura: Macrophthalmidae), with descriptionof a new species and new records, Journal of Natural History, DOI: 10.1080/00222933.2013.807948
To link to this article: http://dx.doi.org/10.1080/00222933.2013.807948
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the“Content”) contained in the publications on our platform. However, Taylor & Francis,our agents, and our licensors make no representations or warranties whatsoever as tothe accuracy, completeness, or suitability for any purpose of the Content. Any opinionsand views expressed in this publication are the opinions and views of the authors,and are not the views of or endorsed by Taylor & Francis. The accuracy of the Contentshould not be relied upon and should be independently verified with primary sourcesof information. Taylor and Francis shall not be liable for any losses, actions, claims,proceedings, demands, costs, expenses, damages, and other liabilities whatsoever orhowsoever caused arising directly or indirectly in connection with, in relation to or arisingout of the use of the Content.
This article may be used for research, teaching, and private study purposes. Anysubstantial or systematic reproduction, redistribution, reselling, loan, sub-licensing,systematic supply, or distribution in any form to anyone is expressly forbidden. Terms &Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions

Journal of Natural History, 2013http://dx.doi.org/10.1080/00222933.2013.807948
The sentinel crabs of the genus Chaenostoma (Stimpson, 1858)(Crustacea: Brachyura: Macrophthalmidae), with description of a newspecies and new records
Reza Naderloo*†
Forschungsinstitut Senckenberg, Frankfurt am Main, Germany
(Received 30 September 2012; final version received 20 May 2013)
The genus Chaenostoma (Stimpson, 1858) so far includes four distinct species, i.e.C. boscii (Audouin, 1826), C. lisae (Poupin and Bouchard, 2010), C. punctula-tus Miers, 1884, and C. sinuspersici (Naderloo and Türkay, 2011). Chaenostomajava n. sp. is here described as a fifth species from Java (Indonesia) easily dis-tinguished from its congeners by the features of the male first pleopod, thepenultimate segment of the male abdomen, and the unique female gonopore. Thenew species is most similar to C. lisae, but they differ in the morphology of the malechelipeds and female gonopores. Chaenostoma lisae is redescribed and recordedfrom Madagascar, Mayotte I. and New Caledonia. An identification key for thegenus is provided.
Keywords: Crustacea; Decapoda; Brachyura; Macrophthalmidae; Chaenostomajava n. sp.; new species
Introduction
The classification of the genus Macrophthalmus Desmarest, 1823 is problematic, a sit-uation that has been discussed in several studies (Barnes 2010; Kitaura et al. 2010;McLay et al. 2010; Naderloo & Türkay 2011; Naderloo et al. 2011). Subdivision ofthe genus Macrophthalmus was first proposed by Barnes (1967), who divided the genusinto six different subgenera, a classification followed by subsequent authors. Ng et al.(2008) added the two subgenera Euplax H. Milne Edwards and Paramareotis Komai,Goshima and Murai, 1995, and listed eight subgenera under this genus. They remarkedthat the subgeneric classification within the genus is not satisfactory, and added thatmost of the subgenera should be raised to generic level. This idea was supported byDavie (2009) and Naderloo et al. (2011).
Among the unsolved taxonomic issues in Macrophthalmus is the Macrophthalmus(Chaenostoma) boscii complex, which is thought to include several different species(Ng et al. 2008; Naderloo & Türkay 2011). The type material of C. boscii (Audouin,1826) was destroyed and not re-examined after its original description (Naderloo &Türkay 2011). The unavailability of the type material resulted in huge taxonomicuncertainties and the assumption that this species is widely distributed from the RedSea throughout the Indian Ocean to Australia and New Caledonia. Naderloo andTürkay (2011) redescribed C. boscii based on material from the north Red Sea (type
*Email: [email protected].†Present address: School of Biology and Center of Excellence in Phylogeny of Living Organisms,College of Science, University of Tehran, Tehran, Iran
© 2013 Taylor & Francis
Dow
nloa
ded
by [
Uni
vers
ity o
f T
ehra
n] a
t 05:
36 0
7 O
ctob
er 2
013

2 R. Naderloo
locality), designated a neotype, and established that C. boscii is restricted to the RedSea. Consequently, all non-Red Sea records of the species examined by them wereplaced in a second species, Chaenostoma sinuspersici (Naderloo and Türkay, 2011).Chaenostoma lisae (Poupin and Bouchard, 2010) is an additional species describedfrom the island of Mayotte in the western Indian Ocean. Poupin and Bouchard (2010)provided detailed description and drawings for C. lisae. But, some small differencescan be seen between the present specimens and drawings (e.g. female gonopore, maleabdomen) given by Poupin and Bouchard (2010) for the type specimens from the islandof Mayotte. Therefore, a detailed description and drawings are provided for C. lisae aswell. Barnes (1967) included Macrophthalmus punctulatus Miers, 1884, together withM. boscii, in the subgenus Mopsocarcinus Barnes, 1967. The subgenus was later con-sidered a subjective synonym of Chaenostoma Stimpson, 1858 (see Barnes 2010). Herewe describe a fifth species of Chaenostoma from Java, Indonesia.
Material and methods
The specimens of Chaenostoma java n. sp. were in Geneva Museum and were orig-inally identified as Macrophthalmus boscii (Audouin, 1826). I examined them in theSenckenberg Museum where the specimens were on loan to Michael Türkay. Thespecimens of Chaenostoma lisae were collected from Europa I., Western Indian Oceanduring Biodiversite, Ressources et Conservation des Recifs Coralliens des Iles Eparses(BioReCIE) Expedition in 2011 and from New Caledonia by Joseph Poupin andMadagascar by Alain Crosnier in 1962. These specimens have also been identified asM. boscii. Drawings were made using a camera lucida (Leica MZ8). Abbreviationsused are as follows: CL, carapace length; CB, carapace breadth; G1, first male gono-pod (pleopod); MHNG, Muséum d’Histoire naturelle de Genève; MNHN, Muséumnational d’Histoire Naturelle, Paris, France.
Key to male specimens of the species of Chaenostoma Stimpson, 1858
1.Distinct chitinous plectrum near lower margin of inner surface of merus ofmale chelipeds . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Chaenostoma sinuspersici
– Without plectrum on the upper surface of merus of male chelipeds . . . . . . . . 2
2.Movable finger of male cheliped without differentiated tooth; inner surface ofpalm with dense setal cover, not extending to inner surface of fingers . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Chaenostoma lisae
– Movable finger of male cheliped with distinct differentiated tooth, innersurface of palm with dense setal cover extending along inner surface of fingers. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3
3.Exorbital angle acute; posterior surface of carapace uneven, mostly granulate;first male gonopod with apical process plate-shaped . . . . . Chaenostoma boscii
– Exorbital angle relatively obtuse; posterior surface of carapace not extremelyuneven, not granulate . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4
4.Posterior surface of carapace with scattered small granules; apical process offirst male gonopod short, rounded, directed laterally at nearly 60◦ . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Chaenostoma java n. sp.
– Posterior surface of carapace without granules; first male gonopod withelongate apical process . . . . . . . . . . . . . . . . . . . . . . . . . . . Chaenostoma punctulatus
Dow
nloa
ded
by [
Uni
vers
ity o
f T
ehra
n] a
t 05:
36 0
7 O
ctob
er 2
013

Journal of Natural History 3
Systematic account
Subsection THORACOTREMATA Guinot, 1977Family MACROPHTHALMIDAE Dana, 1851
Chaenostoma (Stimpson, 1858)
Type species: Macrophthalmus boscii Audouin, 1826
Diagnosis
Small-sized (CL < 15 mm) species; carapace slightly broader than long (CB/CL < 1.3),posterior surface with scattered small granules, without granular ridges; lateral marginwith three teeth, first one largest, last one smaller. Front broad, larger than 0.2 as broadas carapace in basal part of front, slightly constricted medially; eyestalks short, stout,not extending beyond exorbital teeth. Chelipeds equal; palm moderately long, about1.5 times as long as high in its distal portion or slightly shorter, outer surface withlongitudinal granular ridge parallel to lower margin, extending from tip of immovablefinger backward near to articulation with carpus, inner surface with patch of densecover of setae, setal patch may continue to inner surface of fingers; immovable fingernot deflexed or very slightly deflexed; movable finger with more or less distinct dif-ferentiated proximal tooth. Walking legs medium-sized, scarcely covered with setae,without any spine, second and third ones larger, nearly of same length. Third max-illipeds relatively long, with gap between them, ischium about 1.5 times as long asquadrate-shaped merus, with circular pattern of long setae on proximal portion ofouter surface. Posteromedian margin of epistome straight, median part of anteriorbuccal cavity smooth, without any ridge. Male G1 with small apical process and shortmesial palp. Female cheliped with outer surface of palm finely granular, with distinctgranular ridge, running subparallel to lower margin; cutting edge of fingers smooth;female gonopore with small operculum directing outwards.
Chaenostoma java n. sp.(Figures 1A–F, 2A–D)
Material examined
Holotype. 1♂ (CL 9. 60 mm, CB 12.09 mm) (MHNG), Rosoeki, Java, July 1897, leg.Zehntner.
Paratypes. 1♂ (CL 8.97 mm, CB 11.24 mm), 4♀ ovig. (CL 7.94–5.07 mm, CB10.07–6.89 mm) (MHNG), locality same as holotype.
Description
Carapace sub-rectangular (Figure 1A), about 1.2–1.3 times as broad as long, maxi-mum width on posterior part of posterolateral margin; posterior surface with smallgranules scarcely on lateral regions, invisible with naked eyes; long setae on lateralregions, continuous to lateral margin. Regions weekly defined; furrows of gastricregion distinct on posterior portion; cardiac region slightly elevated, intestinal region
Dow
nloa
ded
by [
Uni
vers
ity o
f T
ehra
n] a
t 05:
36 0
7 O
ctob
er 2
013
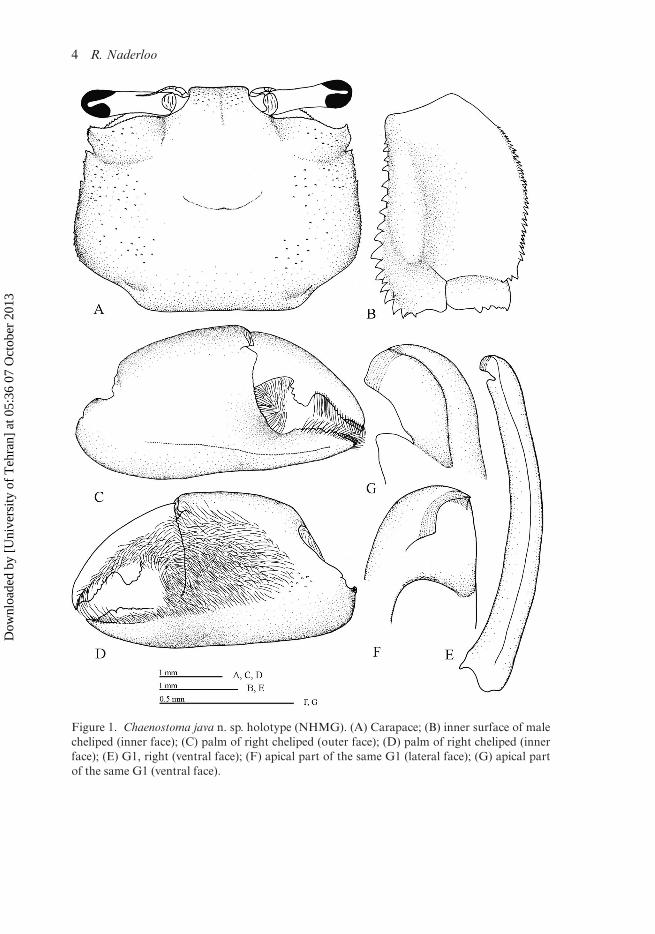
4 R. Naderloo
Figure 1. Chaenostoma java n. sp. holotype (NHMG). (A) Carapace; (B) inner surface of malecheliped (inner face); (C) palm of right cheliped (outer face); (D) palm of right cheliped (innerface); (E) G1, right (ventral face); (F) apical part of the same G1 (lateral face); (G) apical partof the same G1 (ventral face).
Dow
nloa
ded
by [
Uni
vers
ity o
f T
ehra
n] a
t 05:
36 0
7 O
ctob
er 2
013
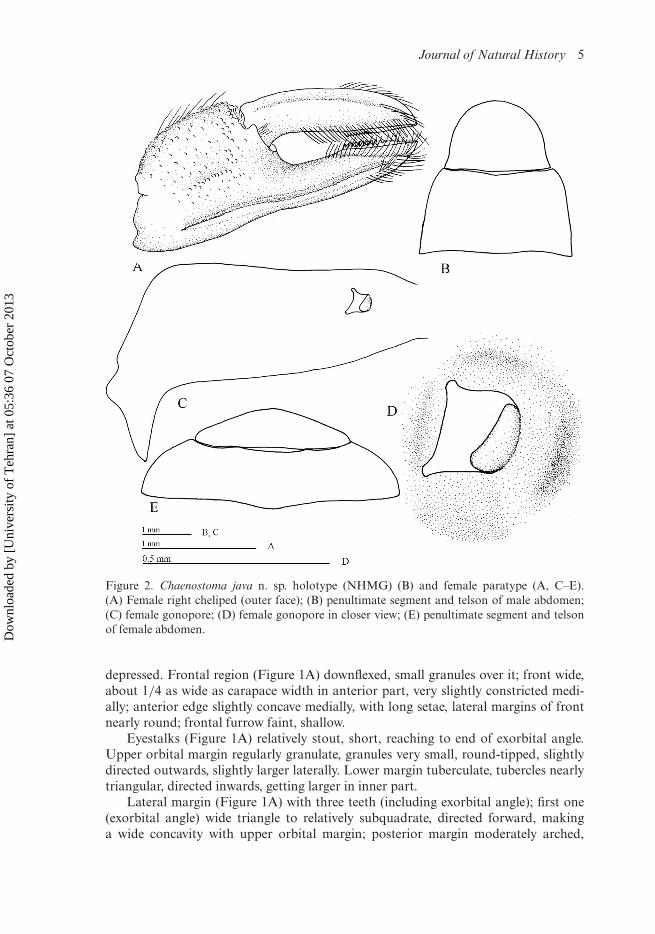
Journal of Natural History 5
Figure 2. Chaenostoma java n. sp. holotype (NHMG) (B) and female paratype (A, C–E).(A) Female right cheliped (outer face); (B) penultimate segment and telson of male abdomen;(C) female gonopore; (D) female gonopore in closer view; (E) penultimate segment and telsonof female abdomen.
depressed. Frontal region (Figure 1A) downflexed, small granules over it; front wide,about 1/4 as wide as carapace width in anterior part, very slightly constricted medi-ally; anterior edge slightly concave medially, with long setae, lateral margins of frontnearly round; frontal furrow faint, shallow.
Eyestalks (Figure 1A) relatively stout, short, reaching to end of exorbital angle.Upper orbital margin regularly granulate, granules very small, round-tipped, slightlydirected outwards, slightly larger laterally. Lower margin tuberculate, tubercles nearlytriangular, directed inwards, getting larger in inner part.
Lateral margin (Figure 1A) with three teeth (including exorbital angle); first one(exorbital angle) wide triangle to relatively subquadrate, directed forward, makinga wide concavity with upper orbital margin; posterior margin moderately arched,
Dow
nloa
ded
by [
Uni
vers
ity o
f T
ehra
n] a
t 05:
36 0
7 O
ctob
er 2
013

6 R. Naderloo
granulate; second longer than first one, lower than it, outer margin nearly straight,granulate; third very small, usually distinguishable, indicated by large granules; pos-terolateral margin relatively straight, diverging posteriorly, with small round granules,wide-based, long setae along it.
Third maxillipeds with large gap between. Ischium long, about 1.8 times as longas wide, slightly shorter than two times as long as merus, inner margin distally convex,with long setae along it, outer margin nearly straight, with short setae; circular patternof long setae on proximal portion of outer surface, long setae irregularly along distalmargin, short setae scarcely on outer surface. Merus quadrate, inner margin with longsetae; outer margin nearly straight, arched proximally, long setae along distal part,outer surface widely depressed near inner margin, narrow depression near along outermargin.
Posteromedian margin of epistome straight, median part of anterior buccal cavitysmooth, without any ridge.
Chelipeds (Figure 1C, D) nearly equal. Ischium denticulate on anterior margin,with long setae. Merus (Figure 1B) upper surface smooth, slightly elevated near innermargin, without chitinous plectrum; outer surface with transverse ridges of very smallgranules, sparsely covered with short setae; inner surface smooth, densely covered withlong setae on distal two third: upper inner margin with small denticles, getting slightlylarger distally, densely covered with long setae, setae continuous to anterior surface;inner lower margin with large triangular tubercles, larger distally, covered with scarcesetae; outer margin denticulate, long setae scarcely along it. Carpus finely granulateon outer surface; margins with small denticles, some large denticles medially on innermargin, long setae along outer, inner, lower distal margins. Palm (Figure 1C) rela-tively swollen, about 1.2 times as long as high; outer surface (Figure 1C) smooth, withmicroscopic granules, slightly larger proximally, longitudinal granular ridge on lowerportion, running from near tip of immovable finger backward, subparallel to lowermargin; lower margin nearly straight, minutely granular, not deflexed near immovablefinger; upper margin slightly arched, finely granular; inner surface (Figure 1D) withsmall granules on proximal portion, mostly covered with long setae, setal patch contin-uous along inner surface of fingers. Movable finger curved downward, curved inwarddistally, cutting edge with subproximal differentiated tooth, large, small denticles distalto large one. Immovable finger short, smooth proximally, without large differentiatedtooth, teeth along proximal two-thirds, getting larger distally; large round gap betweenfingers, tip of fingers spooned, with long setae.
Female chelipeds (Figure 2A) small; palm nearly flat, inferior ridge prominent, par-allel to lower margin, gently curving medially, granular, granules small distally; shortlongitudinal row of granules on middle portion, depressed above this ridge; upper mar-gin granular, with small granules; inner surface of palm smooth, row of long setae nearupper margin. Fingers relatively stout, movable finger distinctly longer than palm,long setae along inner surface, longitudinal ridge along outer surface, cutting edgesof fingers smooth, chitinous ridge comb-shaped proximally.
Walking legs medium-sized, second, third legs largest, nearly in same length, fourthleg smallest, long setae scarcely along anterior margin of segments, except dactylus.Merus posterior surface smooth; anterior, posterior margins serrate, merus of secondand third legs with subdistal tooth on anterior margin, merus of third leg about threetimes as long as wide. Carpus serrate on anterior and posterior margins; carpus of
Dow
nloa
ded
by [
Uni
vers
ity o
f T
ehra
n] a
t 05:
36 0
7 O
ctob
er 2
013

Journal of Natural History 7
third leg slightly shorter than propodus. Propodus finely serrate on anterior and pos-terior margins, slightly longer than dactylus, that of last leg about as long as dactylus.Dactylus with scarce short setae on proximal portion, with six longitudinal smoothridges; dense setal patch on anterior, posterior margins of daclytus and propodus offirst leg; setal patch just on proximal parts of dactylus and propodus of second andthird legs; fourth leg without setal patch.
Male abdomen long; segments 3, 4 nearly in same length, segment 5 distinctlylonger than segments 3, 4, nearly as long as segment 6; segment 6 (Figure 2B) withlateral margins arched, very slightly swollen proximally, converging distally; telsonslightly shorter than segment 6, margins arched, apically rounded.
G1 gently curved medially (Figure 1E); apical chitinous process short, directed lat-erally at nearly 60◦ (Figure 1F, G), lateral surface of apical process distinctly depressed,concave on apical surface; long setae around apical process, long feather-shaped setaealong ventral margin, short feather-shaped setae along mesial margin.
Female abdomen round, segment 6 (Figure 2E) distinctly longer than telson, witharched lateral margins, telson distinctly wider than half as time as segment 6, slightlymore than four times as wide as long.
Female gonopore (Figure 2C) small, with outer margin nearly straight, nearlyparallel to median line of sternum; operculum small (Figure 2D), directed forwardslaterally; margins of opening slightly elevated.
Habitat
The specimens were collected from the rocky intertidal, under small boulders.
Distribution
Presently only known from its type locality, Java (Indonesia) in the eastern IndianOcean.
Etymology
The species is named after the type locality, Java in Indonesia.
Remarks
Chaenostoma java n. sp. is mainly characterized by a blunt exorbital angle, non-deflexed male cheliped, a dense setal patch covering the inner surface of the palmand fingers of the male chelipeds, a G1 with a short rounded apical process, and afemale gonopore with a small operculum directed forwards. The non-deflexed malechelipeds and the morphology of the male G1 distinguish the new species fromC. boscii (Audouin, 1826) and the widely distributed species C. sinuspersici (Naderlooand Türkay, 2011). Male chelipeds of the last two species are more or less deflexeddownwards near the base of the immovable finger. The G1 of M. sinuspersici hasthe apical process of G1 distinctly mesiolaterally inclined, strongly emarginated, butthe apical process of G1 of M. boscii is plate-shaped, directed obliquely in respect
Dow
nloa
ded
by [
Uni
vers
ity o
f T
ehra
n] a
t 05:
36 0
7 O
ctob
er 2
013

8 R. Naderloo
to stem (see Naderloo & Türkay 2011, figure 3). The relatively smooth dorsal sur-face of the carapace of C. java n. sp. is rather similar to that of C. lisae (Poupin andBouchard, 2010) and C. punctulatus (Miers, 1884). The latter has been described ashaving a completely smooth dorsal carapace surface (see Miers 1884; Barnes 1967,p. 229), whereas C. java n. sp. and C. lisae have few scattered granules on the frontaland the lateral regions of the carapace. It should be noted that Miers (1884, pl. XXV,figure A) described C. punctulatus based on a single female from Port Jackson, NewSouth Wales, Australia. Miers mentioned a large differentiated tooth on the cuttingedge of the immovable finger and a smaller one on the cutting edge of the movablefinger, which are clearly seen in his drawings (Miers 1884, pl. XXV, figure A). Barnes(1967, p. 229, figure 10d) examined a large number of specimens from eastern andsouthern Australia and mentioned that only the movable finger has the differentiatedtooth.
Two characters examined here, the male G1 and female gonopore, are proposedto be important diagnostic characters among the species of the group (Naderloo& Türkay 2011). Chaenostoma java n. sp. is rather similar to C. lisae in these twocharacters and these two species are not easily distinguishable from each other.Chaenostoma java n. sp. differs from C. lisae by having a setal patch on the innersurface of the male cheliped palm extending to the fingers (see Figure 1D), whereasin C. lisae the setal patch is present only on the inner surface of the palm and thefingers are glabrous. Furthermore, in the new species a proximal large gap is rec-ognized between the male cheliped fingers with a distinctly differentiated tooth onthe cutting edge of the movable finger, whereas in C. lisae there is no gap betweenthe fingers and the tooth of the movable finger is low and not very differentiatedas in other species of the group (see Figure 3C, D; Poupin & Bouchard 2010,figure 2F, G).
Chaenostoma lisae (Poupin and Bouchard, 2010)(Figures 3A–H, 4A–D)
Macrophthalmus (Chaenostoma) lisae Poupin and Bouchard, 2010, 62, figures 1A–C,2A–O.
Material examined
Indian Ocean. 2♂ (CL 4.14–4.39 mm, CB 5.26–5.59 mm), 1♀ ovig. (CL 5.04 mm, CB6.34 mm) (MNHN-B10717), Nosy Be, Madagascar, February 1962, A. Crosnier; 2♂(CL 4.66–4.05 mm, CB 3.67–3.17 mm) (MNHN); Mozambique Channel, Europa I.BioReCIE Expedition, St. 4, outer reef, high intertidal, 8 November 2011, J. Poupin.
Pacific. 1♂ (CL 4.50 mm, CB 5.69 mm), 1♀ ovig. (CL 5.16 mm, CB 6.79 mm)(MNHN), Pindai, New Caledonia.
Diagnosis
Carapace sub-rectangular (Figure 3A), about 1.26 times as broad as long, maxi-mum width between second lateral teeth; posterior surface with very small granules,
Dow
nloa
ded
by [
Uni
vers
ity o
f T
ehra
n] a
t 05:
36 0
7 O
ctob
er 2
013
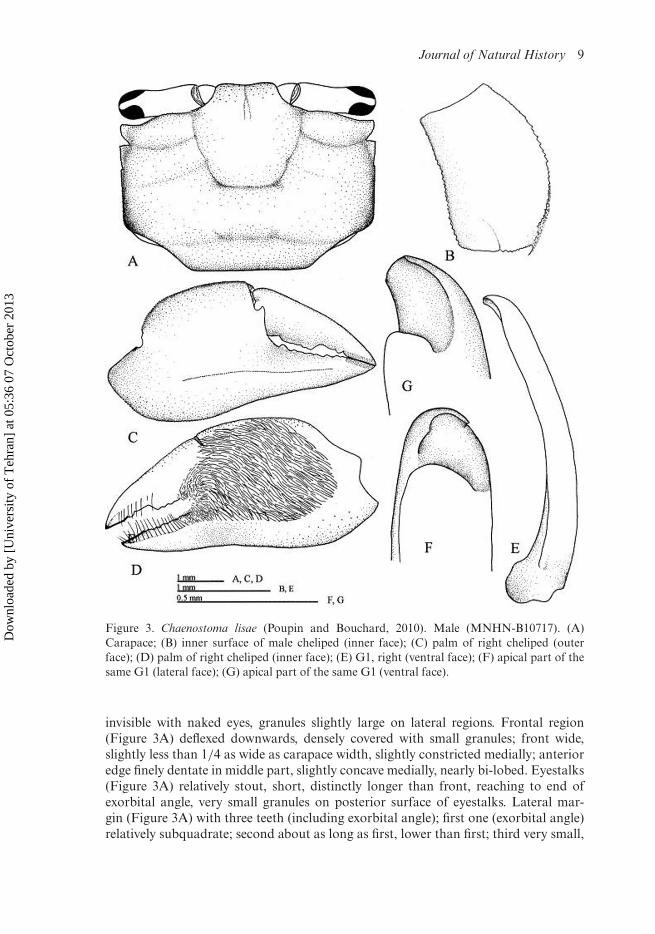
Journal of Natural History 9
Figure 3. Chaenostoma lisae (Poupin and Bouchard, 2010). Male (MNHN-B10717). (A)Carapace; (B) inner surface of male cheliped (inner face); (C) palm of right cheliped (outerface); (D) palm of right cheliped (inner face); (E) G1, right (ventral face); (F) apical part of thesame G1 (lateral face); (G) apical part of the same G1 (ventral face).
invisible with naked eyes, granules slightly large on lateral regions. Frontal region(Figure 3A) deflexed downwards, densely covered with small granules; front wide,slightly less than 1/4 as wide as carapace width, slightly constricted medially; anterioredge finely dentate in middle part, slightly concave medially, nearly bi-lobed. Eyestalks(Figure 3A) relatively stout, short, distinctly longer than front, reaching to end ofexorbital angle, very small granules on posterior surface of eyestalks. Lateral mar-gin (Figure 3A) with three teeth (including exorbital angle); first one (exorbital angle)relatively subquadrate; second about as long as first, lower than first; third very small,
Dow
nloa
ded
by [
Uni
vers
ity o
f T
ehra
n] a
t 05:
36 0
7 O
ctob
er 2
013
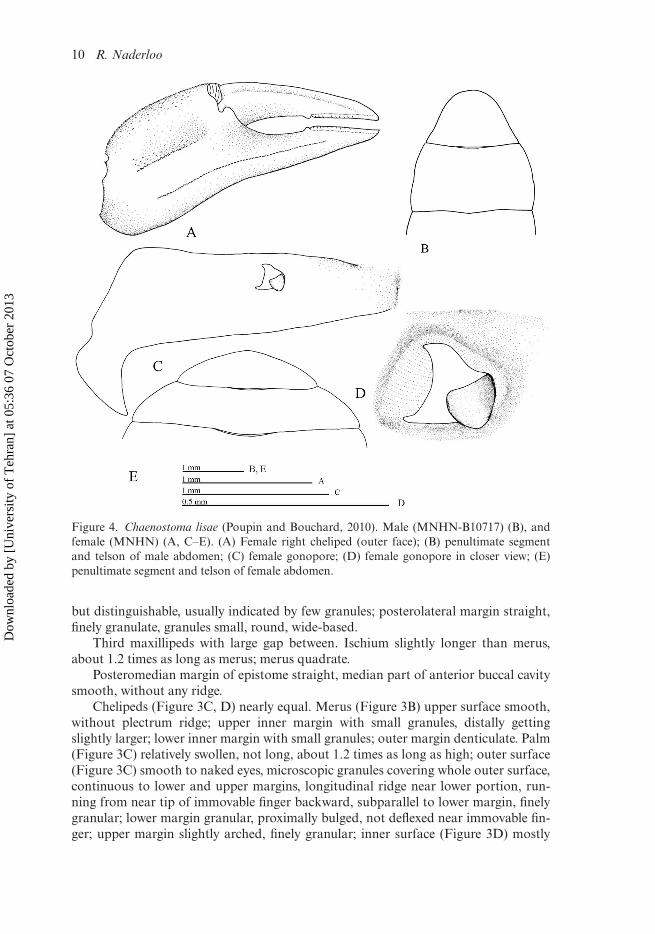
10 R. Naderloo
Figure 4. Chaenostoma lisae (Poupin and Bouchard, 2010). Male (MNHN-B10717) (B), andfemale (MNHN) (A, C–E). (A) Female right cheliped (outer face); (B) penultimate segmentand telson of male abdomen; (C) female gonopore; (D) female gonopore in closer view; (E)penultimate segment and telson of female abdomen.
but distinguishable, usually indicated by few granules; posterolateral margin straight,finely granulate, granules small, round, wide-based.
Third maxillipeds with large gap between. Ischium slightly longer than merus,about 1.2 times as long as merus; merus quadrate.
Posteromedian margin of epistome straight, median part of anterior buccal cavitysmooth, without any ridge.
Chelipeds (Figure 3C, D) nearly equal. Merus (Figure 3B) upper surface smooth,without plectrum ridge; upper inner margin with small granules, distally gettingslightly larger; lower inner margin with small granules; outer margin denticulate. Palm(Figure 3C) relatively swollen, not long, about 1.2 times as long as high; outer surface(Figure 3C) smooth to naked eyes, microscopic granules covering whole outer surface,continuous to lower and upper margins, longitudinal ridge near lower portion, run-ning from near tip of immovable finger backward, subparallel to lower margin, finelygranular; lower margin granular, proximally bulged, not deflexed near immovable fin-ger; upper margin slightly arched, finely granular; inner surface (Figure 3D) mostly
Dow
nloa
ded
by [
Uni
vers
ity o
f T
ehra
n] a
t 05:
36 0
7 O
ctob
er 2
013

Journal of Natural History 11
covered with long setae except on marginal portions, setal patch continuous to proxi-mal of fingers. Movable finger gently curved downward, curved inward distally, uppermargin granular along proximal two third; cutting edge with subproximal differen-tiated tooth, long, low, two or three small denticles distal to large tooth. Immovablefinger short, nearly straight, without differentiated tooth, tuberculate teeth along entireit, getting larger distally.
Chelipeds of female (Figure 4A) small; palm nearly flat, inferior ridge prominent,subparallel to lower margin, gently curving medially, granulate, granules small dis-tally; short longitudinal row of granules on middle portion, depressed above this ridge;upper margin with two rows of small granules; inner surface of palm smooth, row oflong setae near upper margin. Fingers narrow, movable finger about as long as palm,long setae along inner surface, longitudinal ridge along outer surface, cutting edge offingers smooth, distinct round teeth on middle part of both fingers, close to chitinouspart.
Walking legs medium-sized, third leg largest, fourth leg smallest, long setaescarcely along anterior margin of segments, except dactylus. Merus posterior surfacesmooth; anterior, posterior margins serrate, merus of second and third legs with verysmall subdistal tooth on anterior margin, merus of third leg slightly more than twice aslong as wide. Carpus serrate on anterior, posterior margins; carpus of third leg slightlyshorter than propodus. Propodus finely serrate on anterior, posterior margins, slightlylonger than dactylus, that of last leg about as long as dactylus. Dactylus with scarceshort setae on proximal portion, with six longitudinal smooth ridges.
Male abdomen long; segments 3, 4 nearly in same length, segment 5 distinctlylonger than segments 3, 4, nearly as long as segment 6; segment 6 (Figure 4B)with lateral margins arched, very slightly swollen proximally, converging distally; tel-son slightly shorter than segment 6, margins nearly sharply converging proximally,rounded apically.
G1 gently curved medially (Figure 3E); apical chitinous process short, directedlaterally at 60◦ (Figure 3F, G), lateral surface of apical process distinctly depressed,concave on apical surface, with distal margin slightly concave; long setae around apicalprocess, long feather-shaped setae along ventral margin, short feather-shaped setaealong mesial margin.
Female gonopore (Figure 4C) small, with outer margin arched, nearly parallel tomedian line of sternum; operculum small (Figure 4D), directed forwards; margins ofopening slightly elevated.
Colour
“Dorsal surface of carapace and appendages are white-cream scattered with brownishpatches” (Poupin & Bouchard 2010, p. 65), which makes the species hardly dis-tinguishable in its habitat. The preserved specimens examined here all are evenlycream-coloured.
Habitat
Poupin and Bouchard (2010) provided detailed information on the habitat of thespecies in its type locality. Chaenostoma lisae, unlike its congeners that are mainlyfound on rocky shores, digs burrows in the upper intertidal zone in sandy-mud habitats.
Dow
nloa
ded
by [
Uni
vers
ity o
f T
ehra
n] a
t 05:
36 0
7 O
ctob
er 2
013

12 R. Naderloo
Distribution
Mayotte (Comoro Islands), Madagascar, Japan, New Caledonia.
Remarks
Chaenostoma lisae (Poupin and Bouchard, 2010) is characterized by its blunt firstanterolateral teeth of the carapace, a setal patch on the inner surface of the malecheliped palm not extending to cover inner surface of fingers, a rounded short apicalprocess of the male G1, and a female gonopore parallel to the median line of ster-num with a small operculum laterally directing forward. Regarding the blunt form ofthe first anterolateral teeth of the carapace, this species is readily distinguished fromthe type species of the genus, C. boscii (Audouin, 1926) and is similar to other con-geners. Features of the male G1 of C. lisae make it completely distinct from C. boscii,C. sinuspersici (Naderloo and Türkay, 2011) and C. punctulatus (Miers, 1884), whereasit is very close to C. java n. sp. The apical process of the G1 in C. lisae and C. java n. sp.is short, directed in anterior–posterior direction, convex on mesial surface, while thatof M. sinuspersici is strongly emarginated and distinctly concave on the mesial sur-face (see Naderloo & Türkay 2011; Naderloo et al. 2011). The apical margin of G1 inC. sinuspersici is distinctly two-horned, while in C. lisae it is simply slightly concave.The apical process of G1 in C. boscii is unique by having a plate-shaped form, andbeing directed obliquely ventroposteriorly (see Naderloo & Türkay 2011); therefore itis not comparable with that of other species.
Male chelipeds of C. lisae are not long, about 1.2 times as long as high, while thoseof C. boscii and C. sinuspersici are relatively long with length/height ratio about 1.5(see Naderloo & Türkay 2011). The inner surface of the chelipeds of C. lisae has a setalpatch covering the whole inner surface, but just extending to the proximal parts of fin-gers, but in all other congeners the setal patch extends to the end of the fingers. Thefemale gonopore is an additional important character allowing distinction of C. lisaefrom both C. boscii and C. sinuspersici, the latter have female gonopores which areoblique to the central line of the sternum, and the opercula are directed posterolat-erally. But in C. lisae and the new species the outer margin of the genital opening isnearly parallel to the central line of the sternum and the small opercula are directedfrontolaterally.
Regarding the presence and morphology of the differentiated teeth on the cuttingedge of the male cheliped fingers, C. lisae is easily separated from its congeners byhaving a long and low tooth on the cutting edge of the movable finger, and by theabsence of a gap between the fingers.
Conclusion
With the present description of a new species, the genus Chaenostoma now contains fivespecies including the new species herein described. The five species are unevenly dis-tributed throughout the Indo-West Pacific region. Chaenostoma sinuspersici (Naderlooand Türkay, 2011) has the widest range, from East Africa, Gulf of Aden, PersianGulf through the Indian Ocean to Japan, Australia and as far east as New Caledonia.Chaenostoma lisae (Poupin and Bouchard, 2010) had previously been recorded exclu-sively from its type locality in Mayotte, Western Indian Ocean (Poupin & Bouchard,
Dow
nloa
ded
by [
Uni
vers
ity o
f T
ehra
n] a
t 05:
36 0
7 O
ctob
er 2
013

Journal of Natural History 13
2010), but the present record from New Caledonia documents a wide Indo-West Pacificdistribution. The remaining three species are regionally restricted, which could be theresult of local speciation: Chaenostoma punctulatus (Miers, 1884) seems restricted toeastern and south-eastern Australia (see Davie 2002), whereas C. boscii (Audouin,1826) is only known from the Red Sea (see Naderloo & Türkay 2011). It is difficultto suggest an origin for the species using zoogeographical scenarios, but molecu-lar studies, which are currently being undertaken by this author and C. Schubart(Regensburg University, Germany) should reveal the phylogeography of this groupwithin the paraphyletic genus Macrophthlamus.
Acknowledgements
I am very grateful to Regis Cleva (formerly at MNHN), Joseph Poupin (Institut de Recherchede l’Ecole Navale, IRENav), Paul Clark (Natural History Museum, London) and PeterSchwendinger (Muséum d’histoire naturelle de Genève) for their hospitality during my stays inParis and London and for providing me with material. I am indebted to Peter Castro (CaliforniaState Polytechnic University, Pomona) for reviewing and editing the manuscript. My specialthanks to Michael Türkay (Forschungsinstitut Senckenberg of Frankfurt, Germany) for his kindsupport as supervisor.
References
Barnes RSK. 1967. The Macrophthalminae of Australasia; with a review of the evolutionand morphological diversity of the type genus Macrophthalmus. Trans Zool Soc Lond.31:195–262.
Barnes RSK. 2010. A review of the sentinel and allied crabs (Crustacea: Brachyura:Macrophthalmidae), with particular references to the genus Macrophthalmus. Bull RafflesMus. 58:31–49.
Crosnier A. 1965. Crustacés Décapodes, Grapsidae et Ocypodidae. Faune de Madagascar.18:1–143.
Davie PJF. 2002. Crustacea: Malacostraca: Eucarida (Part 2): Decapoda, Anomura, Brachyura.In: Wells, A. & Houston, WWK. editors. Zoological catalogue of Australia. Vol. 19.3B.Melbourne: CSIRO Publishing.
Davie PJF. 2009. A new genus and species of Macrophthalmidae (Crustacea: Decapoda:Brachyura: Ocypodoidea) from the Northern Territory, Australia. Crustaceana.82:815–827.
Kitaura J, Wada J, Fukui Y, McLay C. 2010. Molecular phylogenetic position of the N Zealandsentinel crab, Macrophthalmus (Hemiplax) hirtipes (Jacquinot, in Homborn & Jacquinot,1846) (Decapoda, Brachyura, Macrophthalmidae). Crustaceana. 83:1315–1326.
McLay CL, Kitaura J, Wada K. 2010. Behavioural and molecular evidence for the systematcposition of Macrophthalmus (Hemiplax) hirtipes Hombron & Jacquinot, 1846, with com-ments on Macrophtahlminae subgenera (Decapoda, Brachyura, Macrophthalmidae). In:Fransen C, de Grave S, Ng P. editors. Studies on Malacostraca: Lipke Bijdeley HolthuisMemorial Volume. Crustaceana Monographs, 14. Leiden: BRILL. p. 483–503.
Miers EJ. 1884. Crustacea. In: Report on the zoological collections made in the Indo-Pacific Ocean during the voyage of H.M.S. “Alert” 1881-82. Part I. The collections fromMelanesia. London; p. 178–322, pls. 18–32.
Naderloo R, Türkay M. 2011. A new species of the Macrophthalmus boscii-group (Decapoda:Brachyura: Macrophthalmidae) from the Persian Gulf with designation a neotype forM. boscii Audouin (1826). Marine Biodiversity. 41:503–515.
Dow
nloa
ded
by [
Uni
vers
ity o
f T
ehra
n] a
t 05:
36 0
7 O
ctob
er 2
013

14 R. Naderloo
Naderloo R, Türkay M, Apel M. 2011. Brachyuran crabs of the family MacrophthalmidaeDana, 1851 (Decapoda: Brachyura: Macrophthalmidae) of the Persian Gulf. Zootaxa.2911:1–42.
Ng PKL, Guinot D, Davie PJF. 2008. Systema brachyurorum: Part I. An annotated checklist ofextant brachyuran crabs of the world. Bull Raffles Mus. Suppl. 17:pp. 286.
Poupin J, Bouchard JM. 2010. A new dwarf sentinel crab from Mayotte Island, western IndianOcean (Decapoda: Brachyura: Macrophthalmidae). Zootaxa. 2501:61–67.
Dow
nloa
ded
by [
Uni
vers
ity o
f T
ehra
n] a
t 05:
36 0
7 O
ctob
er 2
013




















