
Trophic relationships of juvenile blue crabs (Callinectes sapidus) in estuarine habitats
12
Primary Research Paper Trophic relationships of juvenile blue crabs (Callinectes sapidus) in estuarine habitats Ana I. Dittel 1, *, Charles E. Epifanio 1 & Marilyn L. Fogel 2 1 College of Marine Studies, University of Delaware, Lewes, DE 19958, USA 2 Carnegie Institution of Washington, Washington, DC 20015, USA (*Author for correspondence: E-mail: [email protected]) Received 12 September 2005; in revised form 27 January 2006; accepted 24 March 2006; published online 29 May 2006 Key words: stable isotopes, carbon and nitrogen sources, food webs, blue crab juveniles Abstract Salt marshes and shallow-water macroalgal beds are known to provide nursery habitat for many species of fish and invertebrates. The role of these habitats as refuge from predation is well established, but the degree to which indigenous primary production within the nursery provides food for growth and development of estuarine species remains unresolved. In this study, we tested the hypothesis that juvenile blue crabs depend on indigenous primary production, directly or indirectly, during their entire stay within the nursery. To test this hypothesis, we conducted isotopic studies and stomach content analyses of juveniles from habitats near the mouth of Delaware Bay and from an adjacent lagoonal estuary (ca. 39.5° N, 75.1° W). Primary producers, marsh detritus, various life-history stages of blue crabs and potential prey species were sampled in the main estuary and in an adjacent marsh during the summer and early fall of two consecutive years. Newly settled juveniles (<15 mm carapace width) from the marsh were about 1.8& lighter in carbon ()17.2&) relative to larger juveniles from the marsh (15–30 mm carapace width) and appeared to have retained a carbon isotopic signature indicative of the phytoplankton-based food web associated with larval stages. However, the signature of juveniles changed as a function of size. Large juveniles and crabs >60 mm were enriched in d 13 C()14.7 ± 0.1&) compared to small crabs, suggesting a gradual shift in diet from a planktonic to a detritus-based food web with increasing size. As with crabs from Delaware Bay, the d 13 C signature of juvenile crabs sampled from macroalgal beds in the lagoonal estuary (Rehoboth Bay) changed as a function of size. Also, d 13 C ratios of crabs varied among the various species of macroalgae. The d 15 N composition of primary producers in the marsh and main estuary also was reflected in the d 15 N values of crabs and other benthic consumers in the respective habitats. Results of stomach-content analysis in this study were consistent with isotope data. Observed changes in prey preferences were related to changes in size of juvenile crabs and also differed among habitats. Gut content analyses of the three size classes of juveniles in macroalgal beds from Rehoboth Bay indicated that the crabs depend heavily on various amphipod species that occur on the seaweeds. These amphipods graze directly on the macroalgae and are among the most abundant invertebrates in the macroalgal beds. This implies a direct trophic relationship between the juvenile crabs and the macroalgae. In summary, our study provides strong evi- dence that the value of nursery areas such as salt marshes and macroalgal beds goes beyond that of providing refuge from predation, and that species using these nurseries (e.g. juvenile blue crabs) are ultimately dependent on primary production originating in benthic plants indigenous to the nursery. Hydrobiologia (2006) 568:379–390 Ó Springer 2006 DOI 10.1007/s10750-006-0204-2
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Trophic relationships of juvenile blue crabs (Callinectes sapidus) in estuarine habitats

Primary Research Paper
Trophic relationships of juvenile blue crabs (Callinectes sapidus) in estuarine
habitats
Ana I. Dittel1,*, Charles E. Epifanio1 & Marilyn L. Fogel21College of Marine Studies, University of Delaware, Lewes, DE 19958, USA2Carnegie Institution of Washington, Washington, DC 20015, USA(*Author for correspondence: E-mail: [email protected])
Received 12 September 2005; in revised form 27 January 2006; accepted 24 March 2006; published online 29 May 2006
Key words: stable isotopes, carbon and nitrogen sources, food webs, blue crab juveniles
Abstract
Salt marshes and shallow-water macroalgal beds are known to provide nursery habitat for many species offish and invertebrates. The role of these habitats as refuge from predation is well established, but the degreeto which indigenous primary production within the nursery provides food for growth and development ofestuarine species remains unresolved. In this study, we tested the hypothesis that juvenile blue crabs dependon indigenous primary production, directly or indirectly, during their entire stay within the nursery. To testthis hypothesis, we conducted isotopic studies and stomach content analyses of juveniles from habitats nearthe mouth of Delaware Bay and from an adjacent lagoonal estuary (ca. 39.5� N, 75.1� W). Primaryproducers, marsh detritus, various life-history stages of blue crabs and potential prey species were sampledin the main estuary and in an adjacent marsh during the summer and early fall of two consecutive years.Newly settled juveniles (<15 mm carapace width) from the marsh were about 1.8& lighter in carbon()17.2&) relative to larger juveniles from the marsh (15–30 mm carapace width) and appeared to haveretained a carbon isotopic signature indicative of the phytoplankton-based food web associated with larvalstages. However, the signature of juveniles changed as a function of size. Large juveniles and crabs>60 mm were enriched in d13C ()14.7±0.1&) compared to small crabs, suggesting a gradual shift in dietfrom a planktonic to a detritus-based food web with increasing size. As with crabs from Delaware Bay, thed13C signature of juvenile crabs sampled from macroalgal beds in the lagoonal estuary (Rehoboth Bay)changed as a function of size. Also, d13C ratios of crabs varied among the various species of macroalgae.The d15N composition of primary producers in the marsh and main estuary also was reflected in the d15Nvalues of crabs and other benthic consumers in the respective habitats. Results of stomach-content analysisin this study were consistent with isotope data. Observed changes in prey preferences were related tochanges in size of juvenile crabs and also differed among habitats. Gut content analyses of the three sizeclasses of juveniles in macroalgal beds from Rehoboth Bay indicated that the crabs depend heavily onvarious amphipod species that occur on the seaweeds. These amphipods graze directly on the macroalgaeand are among the most abundant invertebrates in the macroalgal beds. This implies a direct trophicrelationship between the juvenile crabs and the macroalgae. In summary, our study provides strong evi-dence that the value of nursery areas such as salt marshes and macroalgal beds goes beyond that ofproviding refuge from predation, and that species using these nurseries (e.g. juvenile blue crabs) areultimately dependent on primary production originating in benthic plants indigenous to the nursery.
Hydrobiologia (2006) 568:379–390 � Springer 2006DOI 10.1007/s10750-006-0204-2

Introduction
In estuaries, food for invertebrates and fish isderived from various sources of primary produc-tion. These include benthic micro- and macroal-gae, phytoplankton, and both submersed andemergent vascular plants. The relative importanceof these different sources varies among species andhabitats, and our understanding of this variation isstill limited (Deegan & Garritt, 1997; Peterson,1999). Of particular interest are species such as theblue crab (Callinectes sapidus), which use macro-algal beds, seagrass meadows, oyster reefs, shallowbare sediments and marshes as nursery habitat(Heck & Thoman, 1984; Dittel et al., 1995; Par-dieck et al., 1999; Epifanio et al., 2003). Blue crabsare important predators in estuaries from NewJersey to Argentina (Virnstein, 1977, 1979; Hol-land et al., 1980; Williams, 1984), and the speciesexploits both estuarine and coastal ecosystemsduring its complex life cycle (Dittel & Epifanio,1982; Epifanio, 1995; Jones & Epifanio, 1995;Garvine et al., 1997). Maximum settlement ofblue crab larvae in nursery habitat occurs in latesummer and autumn. For example, in DelawareBay juveniles remain in the nursery until thefollowing summer when they eventually emigrateto adjacent open-water habitat (Epifanio et al.,2003). While the role of these habitats as refugefrom predation is well established (Heck & Tho-man, 1984; Hines et al., 1987; Orth & vanMontfrans, 1987) the degree to which indigenousprimary production within the nursery supportsthe growth and development of blue crabsand other species using the nursery remainsunresolved.
Wetland food webs were traditionally thoughtto be fueled via detritus-based food webs, throughthe decomposition of vascular marsh plants.However, more recent investigations have come tovarying conclusions about the relative importanceof vascular plant detritus and algal carbon (e.g.Peterson & Howarth, 1987; Sullivan & Montcreiff,1990; Peterson, 1999; Wainright et al., 2000; Hart& Lovvron, 2002) in supporting the nutrition ofinvertebrates and fish. Until now, there was noconsensus in the literature regarding the contri-bution of Spartina detritus in supporting thegrowth and development of species that use themarsh.
It is well documented that isotopic data areuseful for identifying animals that switch diets andfor tracing their migratory routes and habitats(Fry, 1981; Bardonnet & Riera, 2005; Norris et al.,2005). However, additional information fromstomach content analyses and spatial sampling isvaluable in the interpretation of stable isotopedistributions (Creach et al., 1997; Peterson, 1999;Hart & Lovvorn, 2002). Studies based solely onstomach-content analyses have shown that juve-nile blue crabs are omnivores and feed on a varietyof prey including small invertebrates and plantdetritus. Nonetheless, little information is avail-able concerning trophic linkages at different pointsin the life history of the species. Moreover, mostresearch combining isotopic and gut-contentanalysis has focused on adult organisms and fewstudies have analyzed the isotopic signatures ofjuvenile stages of invertebrates or fish in any sys-tematic way (Laughlin, 1982; Stoner & Buchanan,1990; Wainright et al., 1996). In this paper, weexpand our field and laboratory experiments todiscern whether the sources of carbon for juvenilecrab growth and development are habitat related(e.g. marshes, macroalgae beds).
In earlier work, we showed that blue crabsswitched from a phytoplankton-based food web toa detritus-based food web after settling in marshhabitats in the estuary (Fantle et al., 1999). Inaddition, laboratory experiments showed thatnewly metamorphosed juveniles fed natural dietsfrom potential settlement sites had isotopic signa-tures reflecting those of their respective diets andthat these signatures were consistent with expectedtrophic fractionation (Dittel et al., 2000).
In the present study, we hypothesized thatjuvenile blue crabs depend on locally producedsources of organic matter during their entire stay innursery habitats such as salt marshes or shallow-water macroalgal beds. To test our hypothesis, wesampled juvenile blue crabs from various estuarinehabitats with different types of algal and macro-phyte communities. We used stable isotopes of Cand N to assess the trophic relationships of earlyjuvenile crabs entering and settling in three differ-ent estuarine habitats. We complemented isotopicstudies with stomach content analyses in order toassess ontogenetic and habitat-related changes inthe diet of the crabs and to discern trophic rela-tionships within the respective habitats.
380

Materials and methods
Study area
The study was conducted at two locations near themouth of Delaware Bay (Fig. 1) and in an adja-cent lagoonal estuary (Rehoboth Bay). We chosethe different sites based on the occurrence ofjuvenile crabs and the presence of variousprimary producers (e.g. Spartina, macroalgae,phytoplankton). The two habitats in DelawareBay were the Great Marsh and Cape HenlopenFlats. The Great Marsh is part of a larger systemof salt marshes that extends almost continuouslyalong the 150 km western shoreline of the bay. Thedominant emergent vegetation at the study site ismarsh cord grass Spartina alterniflora. CapeHenlopen Flats are located approximately 7 kmseaward of the marsh. Sediments in the intertidalzone consist of mostly well sorted medium sand
(Miller et al., 1992). Eroded peat from old marshesis prevalent in surface sediments (personal obser-vation). Benthic macrofauna is dominated bycrustaceans, gastropods (Ilyanassa) and severalpolychaete worms such as Nereis and Marezilleriaviridis, (Bock & Miller, 1995; Miller et al., 2002).Subtidal areas consist of a mixture of well sortedmedium sand. Submersed vegetation is limited totiny patches of macroalgae (Enteromorpha spp.,Ulva lactuca, and several species of red algae)associated with large shells or occasional sub-merged objects (Miller et al., 2002). Rehoboth Bayis a lagoonal estuary located approximately 15 kmsouth of Delaware Bay and connected to the largerestuary by a canal. Benthic habitat in RehobothBay is dominated by large beds of macroalgaeconsisting primarily of the green Ulva lactuca andthe red algae Agardhiella tenera and Gracillariaspp (Timmons & Price, 1996). An earlier investi-gation by our group has shown that these macro-
Longitude-75.4 -75.2 -75.0 -74.8 -74.6
Latit
ude
38.2
38.4
38.6
38.8
39.0
39.2
Broadkill River
Great Marsh
Rehoboth Bay
Delaware Bay
Cape Henlopen
Figure 1. Location of the study area in the Delaware Bay.
381

algal beds are the primary nursery habitat for bluecrabs in Rehoboth Bay (Epifanio et al., 2003).
Field collections
Field samples were collected throughout the sum-mer and early fall of 1997 and 1998. Juvenile bluecrabs of different sizes (carapace width, CW) werecollected with a seine or dip net and immediatelyplaced in a cooler with ice to minimize furtherdigestion of stomach contents. Primary producersand common estuarine fish and invertebrate spe-cies were collected simultaneously with crabs at alllocations. Potential prey items included fiddlercrabs, mussels, snails, grass shrimp and polychae-tes. Particulate organic matter (POM) was col-lected by filtering one liter of seawater through apre-combusted 0.7 lm glass fiber filter. Phyto-plankton was collected by filtering seawaterthrough a 3 lm glass fiber filter (Fantle et al.,1999). Benthic microalgae were collected at lowtide by scraping the marsh surface carefully toexclude as much non-algal material as possible(Fantle et al., 1999; Dittel et al., 2000). Macroal-gae were collected by hand in the various habitats.Upon return to the laboratory, all samples wererinsed in distilled water, placed in glass vials andstored at )20 �C until analyses were performed.Prior to isotope analyses, individual samples werefreeze-dried, ground, and placed in a desiccatoruntil analyses were done.
Stomach content analyses
In order to assess whether dietary changes occurredsoon after crabs entered the estuary and settled, weclassified juveniles into four main size categories asin Dittel et al. (2000): (1) Newly settled juveniles(<15 mm CW), (2) Small juveniles (16–30 mmCW), (3) Intermediate juveniles (31.5–45 mm CW)and (4) Large juveniles (45.1–60 mm CW). (5)Crabs >60 mm include juveniles and adults.
Thawed crabs were measured to the nearestmm CW and dissected using standard microscopictechniques (e.g. Dittel, 1993). Individual gut-con-tent analysis was performed on a total of 164crabs. Once an individual stomach had been re-moved, the percent fullness was estimated and theproportion of each food category was determined.A weighted average of each food category was
then calculated for each of the various size classesof juvenile crabs following the methods of Hineset al. (1987). Tissue from individual crabs wasobtained and prepared for isotope analyses.
Isotopic analyses
Samples were dried at 50 �C in an oven that wasflushed with N2 gas. Dried samples were thenground to a fine powder and sealed in a glassdesiccator with concentrated HC1 vapors to re-move carbonates. Samples were analyzed isotopi-cally by a modified Dumas combustion thatconverts organic carbon and organic nitrogen toCO2 and N2 gas for mass spectral analysis(Macko, 1981). The samples were placed in quartztubes with Cu and CuO and sealed off undervacuum (Macko et al., 1982). The quartz tubeswere heated to 90 �C for one hour, and cooled toroom temperature at a rate of 60 �C h)1. The slowcooling cycle ensured that any oxides of nitrogenwere decomposed to N2. CO2 gas was separatedfrom N2 gas by cryogenic distillation and analyzedby mass spectrometry. A Nier-Johnson type dou-ble-focusing mass spectrometer was used to runthe nitrogen samples whereas the isotopic com-position of carbon dioxide was measured on aFinnigan MAT-252 mass spectrometer. Stableisotope ratios are presented in standard notation:
dZ¼ðRsample=Rstandard�1Þ � 1000&
where Z denotes the heavier isotope (either 13C or15N), and R represents the ratio (either 13C:12C or15N:14N). The standard for nitrogen was atmo-spheric N2 and the standard for carbon was Pee-Dee Belemnite. For d15N and d13C the precisionwas ±0.2& and ±0.02&, respectively. Carbonand nitrogen concentrations were measured on aCarlo Erba EA-1108 Elemental Analyzer.
Data analysis
Mean values for stable isotope ratios werecompared among ontogenetic stages by one-wayANOVA followed by Tukey’s pairwise compari-son test for each of the study sites. In addition, wetested the effect of macroalgae type and size classof juvenile crabs from Rehoboth Bay using a
382

General Linear Model with size class and substratetype as factors and isotope ratio (d13C, d15N) asthe response variable. Preliminary analysis withLevene’s Test for Equality of Variance obviatedthe need for transformation of data.
Results
We measured the isotopic composition of primaryproducers and key members of the estuarine foodweb from Delaware and Rehoboth Bays in orderto assess the types of primary producers thatsupport growth of early life stages of blue crabsand discern trophic linkages in these habitats.Phytoplankton from Delaware Bay and particu-late organic matter (POM) isolated from marshwaters were depleted in d13C (� )22&) relative toprimary producers in the marsh such as Spartinadetritus, Ulva and benthic microalgae. Red mac-roalgae (e.g. Agardhiella) collected at HenlopenFlats and Rehoboth Bay were depleted in d13C()19.4& and )20.7&, respectively) compared togreen algae like Ulva and Enteromorpha ()11.2and )11.9&, respectively) from the various habi-tats (Table 1).
Delaware Bay consumers such as suspensionfeeders (e.g. mussels and razor clams) and zoo-plankton had the lightest d13C composition ()16.8to )20.1&). Blue crab megalopae had d13C values()19.5&) similar to other zooplankton species andconsistent with a phytoplankton-derived diet. Incontrast, most other consumers (d13C: )12.9 to)15.7&) in the marsh and bay areas had d13Cvalues closer to Spartina detritus and benthic algae.
We found a significant effect of size on d13Cisotopic ratios among the various crab stages. Thed13C composition of newly settled juveniles col-lected in the marsh was significantly higher(ANOVA, F4,51 = 65.5; p<0.001; Tukey’s mul-tiple comparison test) than that of all other juve-niles and adults. The signature of juvenile crabschanged as a function of size (Fig. 2a). Newlysettled juveniles were about 1.8& lighter in carbon()17.2&) relative to small juveniles (15–30 mm).Large juvenile stages and adults were enriched ind13C ()14.7±0.1&) compared to early crabstages and small juveniles, suggesting a gradualshift in diet from a planktonic to a detritus foodweb with increasing size (Fig. 2a). Figure 3 shows
the isotope values of different size juveniles fromthe marsh. Mean d13C values of intermediate andlarge juveniles resemble those of Spartina detritus(Table 1). Statistical analyses showed differencesbetween the two size categories collected at Henl-open Flats (ANOVA, F1,15 = 8.3; p = 0.01).Newly settled juveniles at Henlopen Flats wereabout 2.5& heavier in d13C ()15.9±0.5&) rela-tive to small juveniles (Table 1). Again, it ap-peared that crabs were switching diets and feedingon prey that relied on heavier C sources such asSpartina.
Analysis of Variance showed a significant effectof size and macroalgae type on d13C isotopic ratiosof crabs from Rehoboth Bay. Interaction effectswere not significant. Early crab stages from beds ofgreen algae had a d13C value ()14.0&±0.3) thatresembled those of U. lactuca ()14.1&) whereascrabs collected among red algal taxa()17.3&±0.2) were enriched relative to red algae(Table 1). Carbon composition of newly settledjuveniles (<15 mm) from beds of green algae wassignificantly higher (ANOVA, F1,24= 45.4;p<0.001) than for those from beds of red algae.
In Delaware Bay, there was a difference in Ncomposition between primary producers in themarsh and the bay (Table 1). Except for live dunegrass and peat, primary producers in the bay suchas phytoplankton had the lightest 15N values(7.1&) whereas macro algae from Henlopen Flatshad the heaviest (e.g. Enteromorpha and Ulva:�12&). In the marsh, Spartina detritus and ben-thic microalgae had 15N values of 5.8&. The d 15Nvalue for U. lactuca (7.3&) in Rehoboth Bay wasabout 2& lighter than that for A. tenera.
There was variation in the nitrogen composi-tion of juvenile crabs from the different habitats.The average d15N values of early stages from themarsh were considerably lighter than those inHenlopen Flats (5.9±0.5 and 9.4±0.3&,respectively). Crabs from Rehoboth Bay hadaverage d15N values of 5.4&±0.7 and6.9&±0.4 in red and green algal beds, respec-tively. In general, the 15N composition of the pri-mary producers was reflected in that of the crabsand other benthic consumers.
As with carbon isotope ratios, there was a sig-nificant difference in the 15N isotopic values amongthe various crab stages collected in the marsh(ANOVA, F4, 51 = 10.5; p<0.001; Tukey’s
383

Table 1. Isotopic composition (&) of primary producers and organisms associated with blue crab juveniles in Delaware and Rehoboth
Bay study sites
Sample type d13C n d15N n Site
A. Primary producers
Phytoplankton )20.8±0.2 3 7.1±0.3 5 Roosevelt Inlet
POM )22.8±0.6 5 5.4±0.8 4 Great Marsh
Benthic algae )17.6±0.1 5 5.8 2 Great Marsh
Spartina detritus )15.7±0.4 8 5.8±0.5 6 Great Marsh
Ulva )11.2±0.7 6 11.7±0.3 6 Henlopen Flats
Enteromorpha )11.9 1 12.0 1 Henlopen Flats
Agardhiella )19.4 1 11.0 1 Henlopen Flats
Live dune grass )24.8 1 0.6 1 Henlopen Flats
Peat )15.6±1.1 5 3.1±2.3 5 Henlopen Flats
Agardhiella )20.7 1 9.3 1 Rehoboth Bay
Gracillaria spp. )17.6 1 – Rehoboth Bay
Ulva )14.1 1 7.3 1 Rehoboth Bay
B. Consumers
Meiofauna )16.6 1 3.0 1 Great Marsh
Littoraria littorea )14.6±0.5 3 10.3 2 Great Marsh
Uca pugnax )15.3±0.4 7 7.1±0.2 7 Great Marsh
Ilyanassa obsolete )13.0±0.2 3 9.7 1 Great Marsh
Geukensia demissa )18.4±0.3 6 7.7±0.4 5 Great Marsh
Paleomonetes )15.3 2 8.1 2 Great Marsh
Cyprinidon variegates )14.5 1 7.6 1 Great Marsh
Fundulus sp. )13.5 1 10.2 1 Great Marsh
Naked goby )15.6 1 10.8 1 Great Marsh
Silverside )16.7 1 12.3 1 Great Marsh
Zooplankton (>500 lm) )16.8 2 10.1 2 Roosevelt Inlet
Copepods (253–500 lm) )18.2 2 11.2 1 Roosevelt Inlet
Crab larvae (>500 lm) )17.1±0.4 3 10.0 1 Roosevelt Inlet
Zooplankton (253–500 lm) )17.5 1 8.7 1 Broadkill River
Mytilus edulis )20.1±0.3 3 9.1±0.7 3 Broadkill River
Illyanasa obsolete )12.5±0.2 10 11.4±0.3 10 Henlopen Flats
Ensis )17.0 1 9.1 1 Henlopen Flats
Nereis )14.0±0.2 6 12.9±0.9 6 Henlopen Flats
Merenzellaria viridis )15.6±0.6 4 10.8±0.3 4 Henlopen Flats
Unidentifiable Hermit crab )14.6 1 9.8 1 Henlopen Flats
Unidentifiable sipunculid )13.2 1 9.1 1 Henlopen Flats
Crabs
C. sapidus (<15 mm) )17.4±0.1 13 6.0±0.6 13 Great Marsh
C. sapidus (15–30 mm) )15.5±0.3 6 6.3±0.7 6 Great Marsh
C. sapidus (31–45 mm) )14.9±0.2 5 9.3±0.8 5 Great Marsh
C. sapidus (46–60 mm) )14.7±0.1 7 8.7±0.3 7 Great Marsh
C. sapidus (>60 mm) )14.7±0.1 21 8.8±0.2 21 Great Marsh
C. sapidus (<15 mm) )15.9±0.5 12 9.4±0.3 12 Henlopen Flats
C. sapidus (15–30 mm) )14.2±1.0 4 9.8±0.6 4 Henlopen Flats
C. sapidus (<15 mm) )17.3±0.2 7 5.4±0.7 9 Rehoboth Bay (Red algae)
C. sapidus (15–30 mm) )15.9±0.6 6 5.7±1.2 7 Rehoboth Bay (Red algae)
Continued on next page
384
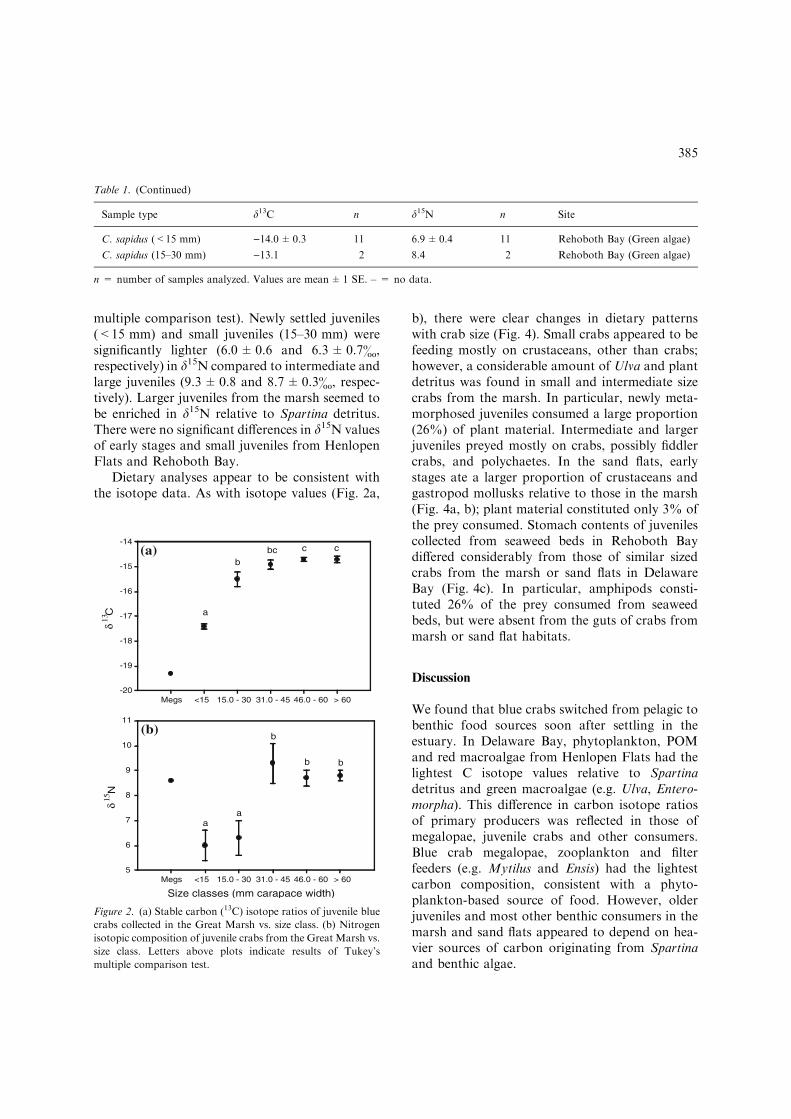
multiple comparison test). Newly settled juveniles(<15 mm) and small juveniles (15–30 mm) weresignificantly lighter (6.0±0.6 and 6.3±0.7&,respectively) in d15N compared to intermediate andlarge juveniles (9.3±0.8 and 8.7±0.3&, respec-tively). Larger juveniles from the marsh seemed tobe enriched in d15N relative to Spartina detritus.There were no significant differences in d15N valuesof early stages and small juveniles from HenlopenFlats and Rehoboth Bay.
Dietary analyses appear to be consistent withthe isotope data. As with isotope values (Fig. 2a,
b), there were clear changes in dietary patternswith crab size (Fig. 4). Small crabs appeared to befeeding mostly on crustaceans, other than crabs;however, a considerable amount of Ulva and plantdetritus was found in small and intermediate sizecrabs from the marsh. In particular, newly meta-morphosed juveniles consumed a large proportion(26%) of plant material. Intermediate and largerjuveniles preyed mostly on crabs, possibly fiddlercrabs, and polychaetes. In the sand flats, earlystages ate a larger proportion of crustaceans andgastropod mollusks relative to those in the marsh(Fig. 4a, b); plant material constituted only 3% ofthe prey consumed. Stomach contents of juvenilescollected from seaweed beds in Rehoboth Baydiffered considerably from those of similar sizedcrabs from the marsh or sand flats in DelawareBay (Fig. 4c). In particular, amphipods consti-tuted 26% of the prey consumed from seaweedbeds, but were absent from the guts of crabs frommarsh or sand flat habitats.
Discussion
We found that blue crabs switched from pelagic tobenthic food sources soon after settling in theestuary. In Delaware Bay, phytoplankton, POMand red macroalgae from Henlopen Flats had thelightest C isotope values relative to Spartinadetritus and green macroalgae (e.g. Ulva, Entero-morpha). This difference in carbon isotope ratiosof primary producers was reflected in those ofmegalopae, juvenile crabs and other consumers.Blue crab megalopae, zooplankton and filterfeeders (e.g. Mytilus and Ensis) had the lightestcarbon composition, consistent with a phyto-plankton-based source of food. However, olderjuveniles and most other benthic consumers in themarsh and sand flats appeared to depend on hea-vier sources of carbon originating from Spartinaand benthic algae.
Table 1. (Continued)
Sample type d13C n d15N n Site
C. sapidus (<15 mm) )14.0±0.3 11 6.9±0.4 11 Rehoboth Bay (Green algae)
C. sapidus (15–30 mm) )13.1 2 8.4 2 Rehoboth Bay (Green algae)
n = number of samples analyzed. Values are mean±1 SE. – = no data.
δ 15
N
5
6
7
8
9
10
11
Size classes (mm carapace width)
aa
b
b b
Megs <15 46.0 - 60
δ 13
C
-20
-19
-18
-17
-16
-15
-14
a
bbc c
> 6031.0 - 4515.0 - 30
Megs <15 46.0 - 60 > 6031.0 - 4515.0 - 30
c
(b)
(a)
Figure 2. (a) Stable carbon (13C) isotope ratios of juvenile blue
crabs collected in the Great Marsh vs. size class. (b) Nitrogen
isotopic composition of juvenile crabs from the Great Marsh vs.
size class. Letters above plots indicate results of Tukey’s
multiple comparison test.
385

Changes in the isotope composition of the var-ious crab stages further indicate that early lifestages switch diets soon after settling and meta-morphosis of megalopae occur. While newlymetamorphosed juveniles collected in the marshappeared to retain a pelagic isotope signal()17.5&), the carbon isotope composition changedas a function of size. Small juveniles showed a trendtowards d13C enrichment; by the time crabsreached sizes of 30 mm or more, average d13Cvalues ()14.8±0.4&) were closer to those ofSpartina detritus. These findings are consistentwith our previous work, which suggested thatheavy marsh carbon is an important dietary com-ponent of juveniles living in the marsh (Fantleet al., 1999; Dittel et al., 2000). In that study, newlymetamorphosed crabs were fed different diets un-der controlled laboratory conditions. After21 days, crabs had changed their initial isotopiccomposition (d13C: )17.1&; d15N: 10.0&) and re-flected that of the diet. For example, crabs fed Ucapugnax, a detritivore crab (Haines & Montague,1979; Montague, 1980; Currin et al., 1995), weresignificantly enriched in d13C ()15.8&), consistentwith this species eating heavy marsh carbon. Incontrast, crabs fed a mixed diet of meiofauna plusdetritus had isotope values of )19.5& whereasthe average d13C of the diet was )16.1&. This
difference in isotopic signatures between the crabsand the diet suggested that juveniles were feedingselectively on prey that depended on an isotopicallylighter C source (e.g. benthic algae) rather thanmarsh carbon. However, in the current study, mostintermediate and large size juveniles isotopicallyresembled Spartina detritus. These results indicatethat juvenile crabs remain in the marsh long en-ough to acquire a distinct isotopic signature, fur-ther demonstrating the importance of marshcarbon in supporting growth and development ofblue crab juveniles in these habitats.
There was a wide variation in carbon isotopesignatures of newly settled juveniles in the sandflats. Although d13C isotope ratios of most earlystage crabs were between )16.0 and )17.8&, sev-eral individuals were significantly heavier andisotopically resembled macroalgae such as Ulvaand Enteromorpha collected in the flats (Table 1).Again, it appears that early life stages switch dietssoon after settlement and metamorphosis of meg-alopae occur. Moreover, it appears that benthicfood webs in the marsh and sand flats are drivenby heavier sources of carbon, rather than byphytoplankton-based food chains. In our previousstudy, we concluded that juveniles in the mainestuary depended on phytoplankton-derived or-ganic matter as the main source of carbon (Fantle
-20 -18 -17 -16 -15 -145
6
7
8
9
10
11δ 15
Ν
δ13C
<15
15.0 - 30
31.0 - 45
46.0 - 60
> 60Megs
-19
Figure 3. Stable carbon (13C) isotope vs. Nitrogen stable (d15N) of various size classes of juvenile crabs from the Great Marsh. Values
above plots indicate size of crabs (mm CW).
386

et al., 1999; Dittel et al., 2000). However, thatinvestigation was based on isotope analyses of alimited number of small crabs. In the presentstudy, a detailed analysis of crabs and other con-sumers in the sand flats revealed that carbonsources other than phytoplankton might beimportant components of benthic food webs in the
main estuary. Large clumps of eroded marshsubstrate (vegetation plus sediment) are carried bylittoral transport to the tip of Cape Henlopen anddeposited on the tidal flats (Kraft et al., 1976). Thesampling area is a ‘‘null area’’ where sedimentsfrom Delaware Bay and Cape Henlopen converge(Kraft et al., 1976). Moreover, recent modelingstudies indicate that marsh detritus is potentiallytransported downstream from the Great Marsh,through Roosevelt Inlet or from Broadkill Beachand accumulated in this area (Petrone et al., 2005).While juvenile blue crabs, for example, do notnecessarily assimilate marsh or peat detritus di-rectly, these juveniles possibly derive their nutri-tion from microbial assemblages associated withthe detritus (Dittel et al., 2000).
Earlier work by our group indicates that mac-roalgal beds are also important settlement sites forblue crab megalopae and appear to be sources ofprimary production for juvenile crabs (Epifanioet al., 2003). Results of our study corroboratethose previous findings. Carbon isotope ratios ofred and green algae from Rehoboth Bay weresimilar to those reported previously for these spe-cies (Fry et al., 1982; Maberly et al., 1992; Currinet al., 1995), and higher than typical values forestuarine phytoplankton (Fogel et al., 1992; Dittelet al., 2000). In turn, the isotope ratios of crabscollected among the various species of macroalgaereflected those of the dominant macroalgae (redvs. green) from where juveniles were collected.These differences imply that juvenile crabs weredependent on macroalgal food webs originatingwithin the beds and that individual crabs remainwithin particular seaweed beds for periods longenough to assume distinct isotopic compositions.These results are consistent with previous studiesthat have found that consumers use locally pro-duced sources of organic matter (Deegan & Gar-ritt, 1997; Wainright et al., 2000; Guest &Connolly, 2004; Guest et al., 2004).
The nitrogen isotope composition of primaryproducers in the marsh and main estuary werereflected in the d15N values of crabs and otherbenthic consumers in the respective habitats. Forexample, d15N values of small crabs in the flatswere at least 3& heavier than those in the marsh.Other consumers like polychaetes and zooplank-ton had nitrogen isotope compositions thatreflected the heavier nitrogen values of the various
0
20
40
60
80Ave
rag
e p
erce
nta
ge
of
Eac
h F
oo
d C
ateg
ory 0
20
40
60
80 <15 mm 315-30 mm31-60 mm>60 mm
0
20
40
60
80
Great Marsh n=51
Sand Flats n=27<15 mm 15-30 mm 31-60 mm
<15 mm 15-30 mm31-60 mm
Rehoboth Bay n=86
Oth
er C
rust
acea
ns
Cra
bsA
mph
ipod
s
Biv
alve
sO
ther
mol
lusk
sP
olyc
haet
es
Ulv
aP
lant
det
ritu
sFo
ram
inife
ra
San
d
Un
id. t
issu
e
(a)
(b)
(c)
Figure 4. Average percentage of the various food categories in
the diet of different size classes of blue crabs collected in Del-
aware and Rehoboth Bays. n = number of crabs analyzed at
each study site.
387

primary producers in the main estuary. These re-sults are consistent with those of our previousstudy, which showed that nitrogen isotope com-position of primary producers was reflected inthose of experimental crabs fed various diets.Furthermore, isotope analyses also suggested thatprimary producers in the marsh and main estuaryderived their nitrogen from different sources(Dittel et al., 2000). Several studies in the Dela-ware Bay have attributed the high d15N values ofphytoplankton to anthropogenic inputs of dis-solved organic nitrogen derived from sewage oragricultural run-off (Cifuentes et al., 1988; Cifu-entes et al., 1989). In our study, several fish speciesin the marsh had considerably higher d15N ratiosrelative to other consumers in that habitat. Oneexplanation is that fish are occupying higher tro-phic levels. Alternatively, fish may be feeding inthe main estuary on a d15N nitrogen-enriched foodweb based on anthropogenic nitrogen.
Dietary analyses appear to be consistent withthe isotope data. There were changes in feedingbehavior with crab size as well as differences in theprey ingested by crabs collected in the varioushabitats. Early stages and small crabs fed on agreater variety of prey types than larger juveniles.These patterns in feeding behavior seemed to bereflected in the wider variation in isotopic signa-tures observed in newly settled juveniles comparedto those in larger crabs. We found clear differencesin the diets of same size crabs collected in thevarious habitats. For example, amphipods com-prised the single most important food type in thestomachs of blue crabs collected from seaweedbeds in Rehoboth Bay, but are completely absentfrom the diets of juveniles using alternative nurs-ery areas such as salt marshes or sand flats(Epifanio et al., 2003). Our results appear toindicate that the trophic linkage between juvenilecrabs and macroalgae depends heavily on amphi-pods species common in seaweeds. Various speciesof seaweed-grazing amphipods are among themost abundant macrofauna found in seaweed bedsin Rehoboth Bay (Timmons & Price, 1996).
In conclusion, our data clearly indicate thatblue crabs switch from a phytoplankton-basedfood chain to benthic sources of organic mattersoon after settling in nursery habitat. In particular,juveniles collected in the marsh showed a clearpattern towards carbon isotope enrichment as
crabs grew in size. While early crab stages retainedisotopic signatures indicative of a phytoplankton-based food web, intermediate and large juvenilesappeared to be supported by a food chain basedon heavy marsh carbon. Previous studies docu-mented the refuge value of habitats like marshesand seaweed beds for species of invertebrates andfish. Our study now provides strong evidence thatlocal carbon sources like marsh detritus and macroalgal production play an important role insupporting growth and development of early lifestages of species such as the blue crab.
Acknowledgements
This study was supported in part by a grant fromthe Marsh Estuarine Research Program (MERP)(no. G98-04A) and NOAA Sea Grant Program.We thank Ray A. Rodriguez for his help withfield sampling. Sandy Schwalm and Matt Fantlehelped with the isotope analyses. We also thankBill Ullman and Doug Miller (SG RME 31) fortheir collaboration on work done at CapeHenlopen.
References
Bardonnet, A. & P. Riera, 2005. Feeding of glass eels (Anguilla
anguilla) in the course of their estuarine migration: new in-
sights from stable isotope analysis. Estuarine, Coastal, and
Shelf Science 63: 201–209.
Bock, M. J. & D. C. Miller, 1995. Storm effects on particulate
food resources on an intertidal sandflat. Journal of Experi-
mental Marine Biology and Ecology 187: 81–101.
Cifuentes, L. A., M. L. Fogel, J. R. Pennock & J. H. Sharp,
1989. Biogeochemical factors that influence the stable
nitrogen isotope ratio of dissolved ammonium in the Dela-
ware Estuary. Geochimica et Cosmochimica Acta 53: 2713–
2721.
Cifuentes, L. A., J. H. Sharp & M. L. Fogel, 1988. Stable
carbon and nitrogen isotope biogeochemistry in the Dela-
ware estuary. Limnology and Oceanography 33: 1102–1115.
Creach, V., M. T. Schricke, G. Bertru & A. Mariotti, 1997.
Stable isotopes and gut analyses to determine feeding
relationships in saltmarsh macroconsumers. Estuarine,
Coastal, and Shelf Science 44: 599–611.
Currin, C. A., S. Y. Newell & H. W. Paerl, 1995. The role of
standing dead Spartina alterniflora and benthic microalgae in
salt marsh food webs: considerations based on multiple
stable isotope analysis. Marine Ecology Progress Series 121:
99–116.
388

Deegan, L. A. & R. H. Garritt, 1997. Evidence for spatial
variability in estuarine food webs. Marine Ecology Progress
Series 147: 31–47.
Dittel, A. I., 1993. Changes in food-habits of Callinectes arcu-
atus (Ordway) (Crustacea – Decapoda) in the Gulf of Ni-
coya, Costa-Rica. Revista de Biologia Tropical 41: 639–646.
Dittel, A., A. Hines, G. Ruiz & K. Ruffin, 1995. Effects of
shallow water refuge on behavior and density-dependent
mortality of juvenile blue crabs in Chesapeake Bay. Bulletin
of Marine Science 57: 902–916.
Dittel, A. I., C. E. Epifanio, S. M. Schwalm, M. S. Fantle & M.
L. Fogel, 2000. Carbon and nitrogen sources for juvenile
blue crabs Callinectes sapidus in coastal wetlands. Marine
Ecology Progress Series 194: 103–112.
Dittel, A. & C. E. Epifanio, 1982. Seasonal abundance and
vertical distribution of crab larvae in Delaware Bay. Estu-
aries 5: 197–202.
Epifanio, C. E., 1995. Transport of blue crab (Callinectes sa-
pidus) larvae in the waters off Mid-Atlantic states. Bulletin of
Marine Science 57: 713–725.
Epifanio, C. E., A. I. Dittel, R. A. Rodriguez & T. E. Targett,
2003. The role of macroalgal beds as nursery habitat for
juvenile blue crabs, Callinectes sapidus. Journal of Shellfish
Research 22: 881–886.
Fantle, M. S., A. I. Dittel, S. M. Schwalm, C. E. Epifanio & M.
L. Fogel, 1999. A food web analysis of the juvenile blue crab,
Callinectes sapidus, using stable isotopes in whole animals
and individual amino acids. Oecologia 120: 416–426.
Fogel, M. L., L. A. Cifuentes, D. J. Velinsky & J. H. Sharp,
1992. Relationship of carbon availability in estuarine phy-
toplankton to isotopic composition. Marine Ecology Pro-
gress Series 82: 291–300.
Fry, B., 1981. Natural stable isotope tag traces Texas shrimp
migrations. Fishery Bulletin 79: 337–345.
Fry, B., R. Lutes, M. Northam, P. L. Parker & J. Ogden, 1982.
A 13C/12C comparison of food webs in Caribbean seagrass
meadows and coral reefs. Aquatic Botany 14: 389–398.
Garvine, R. W., C. E. Epifanio, C. C. Epifanio & K. C. Wong,
1997. Transport and recruitment of blue crab larvae: a model
with advection and mortality. Estuarine Coastal and Shelf
Science 45: 99–111.
Guest, M. A. & R. M. Connolly, 2004. Fine-scale movement
and assimilation of carbon in saltmarsh and mangrove
habitat by resident animals. Aquatic Ecology 38: 599–609.
Guest, M. A., R. M. Connolly & N. R. Loneragan, 2004.
Carbon movement and assimilation by invertebrates in
estuarine habitats at a scale of metres. Marine Ecology
Progress Series 278: 27–34.
Haines, E. B. & C. L. Montague, 1979. Food sources of estu-
arine invertebrates analyzed using 13C/12C ratios. Ecology
60: 48–56.
Hart, E. A. & J. R. Lovvorn, 2002. Interpreting stable isotopes
from macroinvertebrate foodwebs in saline wetlands. Lim-
nology and Oceanography 47: 580–584.
Heck, K. L. & T. A. Thoman, 1984. The nursery role of sea-
grass meadows in the upper and lower reaches of the Ches-
apeake Bay. Estuaries 7: 70–92.
Hines, A. H., R. N. Lipcius & A. M. Haddon, 1987. Population
dynamics and habitat partitioning by size, sex and molt stage
of blue crabs Callinectes sapidus in a subestuary of central
Chesapeake Bay. Marine Ecology Progress Series 36: 55–64.
Holland, A. F., N. K. Mountford, K. R. Kaumeyer & J. A.
Mihursky, 1980. Influence of predation on infaunal abun-
dance in upper Chesapeake Bay. Marine Biology 57: 221–
235.
Jones, M. B. & C. E. Epifanio, 1995. Settlement of brachyuran
megalopae in Delaware Bay: an analysis of time series data.
Marine Ecology Progress Series 125: 67–76.
Kraft, J. C., E. A. Allen, D. F. Belknap, C. J. John & E. M.
Maurmeyer, 1976. Delaware’s Changing Shoreline. Techni-
cal Report Number 1, Delaware Coastal Zone Management
Program, May 1976.
Laughlin, R. A., 1982. Feeding habits of the blue crab, Calli-
nectes sapidus Rathbun, in the Apalachicola estuary, Flor-
ida. Bulletin of Marine Science 32: 807–822.
Maberly, S. C., J. A. Raven & A. M. Johnston, 1992. Dis-
crimination between 12C and 13C by marine plants. Oeco-
logia 91: 481–492.
Macko, S. A., 1981. Stable nitrogen isotope ratios as tracers of
organic geochemical processes. Dissertation, University of
Texas, Austin.
Macko, S. A., W. Y. Lee & P. L. Parker, 1982. Nitrogen and
carbon isotope fractionation by two species of marine am-
phipods: laboratory and field studies. Journal of Experi-
mental Marine Biology and Ecology 63: 145–149.
Miller, D. C., M. J. Bock & E. J. Turner, 1992. Deposit and
suspension feeding in oscillatory flows and sediment fluxes.
Journal of Marine Research 50: 489–520.
Miller, D. C., C. L. Muir & O. A. Hauser, 2002. Detrimental
effects of sedimentation on marine benthos: what can be
learned from natural processes and rates?. Ecological Engi-
neering 19: 211–232.
Montague, C. L., 1980. A natural-history of temperate western
Atlantic fiddler crabs (genus Uca) with reference to their
impact on the salt-marsh. Contributions in Marine Science
23: 25–55.
Norris, D. R., P. P. Marra, T. K. Kyser & L. M. Ratcliffe, 2005.
Tracking habitat use of a long-distance migratory bird, the
American redstart Setophaga ruticilla, using stable-carbon
isotopes in cellular blood. Journal of Avian Biology 36: 164–
170.
Orth, R. J. & J. van Montfrans, 1987. Utilization of seagrass
meadow and tidal marsh creek by blue crabs (Callinectes
sapidus). I. Seasonal and annual variations in abundance
with emphasis on post-settlement juveniles. Marine Ecology
Progress Series 41: 283–294.
Pardieck, R. A., R. J. Orth, R. J. Diaz & R. N. Lipcius, 1999.
Ontogenetic changes in habitat use by postlarvae and young
juveniles of the blue crab. Marine Ecology Progress Series
186: 227–238.
Peterson, B. J., 1999. Stable isotopes as tracers of organic
matter input and transfer in benthic food webs: a review.
Acta Oecologica-International Journal of Ecology 20: 479–
487.
Peterson, B. J. & R. W. Howarth, 1987. Sulfur, carbon and
nitrogen isotopes used to trace organic matter flow in the
salt-marsh estuaries of Sapelo Island, Georgia. Limnology
and Oceanography 32: 1195–1213.
389

Petrone, C. J., L. B. Jancaitis, M. B. Jones, C. C. Natunewicz,
C. E. Tilburg & C. E. Epifanio, 2005. Dynamics of larval
patches: spatial distribution of fiddler crab larvae in Dela-
ware Bay and adjacent waters. Marine Ecology Progress
Series 293: 177–190.
Stoner, A. W. & B. A. Buchanan, 1990. Ontogeny and overlap
in the diets of four tropical Callinectes sp. Bulletin of Marine
Science 46: 3–12.
Sullivan, M. J. & C. A. Moncreiff, 1990. Edaphic algae are an
important component of salt marsh food-webs: evidence
from multiple stable isotope analyses. Marine Ecology Pro-
gress Series 62: 149–159.
Timmons, M. & K. S. Price, 1996. The macroalgae and asso-
ciated fauna of Rehoboth and Indian River Bays, Delaware.
Botanica Marina 39: 231–238.
Virnstein, R. W., 1977. The importance of predation by crabs
and fishes on benthic infauna in Chesapeake Bay. Ecology
58: 1199–1217.
Virnstein, R. W., 1979. Predation on estuarine infauna: re-
sponse patterns of component species. Estuaries 2: 69–86.
Wainright, S. C., C. M. Fuller, R. H. Michener & R. A.
Richards, 1996. Spatial variation of trophic position and
growth rate of juvenile striped bass (Morone saxatilis) in the
Delaware River. Canadian Journal of Fisheries and Aquatic
Sciences 53: 685–692.
Wainright, S. C., M. P. Weinstein, K. W. Able & C. A. Currin,
2000. Relative importance of benthic microalgae, phyto-
plankton and the detritus of smooth cordgrass Spartina
alterniflora and the common reed Phragmites australis to
brackish-marsh food webs. Marine Ecology Progress Series
200: 77–91.
Williams, A. B., 1984. Shrimps, Lobsters, and Crabs of the
Atlantic Coast of the EasternUnited States,Maine toFlorida.
Smithsonian Institution Press, Washington DC 550 pp.
390




















