
Identification of new subgroup of HSP70 in Bythograeidae (hydrothermal crabs) and Xanthidae
9
Identification of new subgroup of HSP70 in Bythograeidae (hydrothermal crabs) and Xanthidae Vincent Leignel ⁎ , Marie Cibois, Brigitte Moreau, Benoît Chénais Laboratoire de Biologie et Génétique Evolutive (EA3265), Université du Maine, 72085, Le Mans, France Received 13 November 2006; received in revised form 9 February 2007; accepted 21 February 2007 Received by M. Di Giulio Available online 15 March 2007 Abstract Crabs of the Bythograeidae family (Crustacea: Brachyura: Bythogreoidea) are the only endemic crab family living in hydrothermal fields. The hydrothermal environment is characterized by unique ecological parameters, such as the high temperature gradient around the hydrothermal chimney (2–350 °C), a fluid environment containing high levels of metals and numerous gases. The 70-kDa Heat Shock Protein (HSP70) group is the most-studied HSP, because it is ubiquitous, and a strong positive correlation has been found between the amounts of HSP70 produced in response to stress, and the ability of the organism to withstand stressful conditions. The 70-kDa heat shock protein genes from Bythograeids (species analyzed: Bythograea thermydron, Cyanagraea praedator and Segonzacia mesatlantica) were characterized. Our results revealed that Bythograeidae possess genes which are similar with those present in Xanthids (coastal crabs). The deduced protein sequences displayed motifs distinct from those in the other crustacean HSC70/HSP70s available in the databases. Phylogenetic analysis showed that these members of HSP70 family identified in Bythograeidae and Xanthidae constitute a new subgroup within this family. © 2007 Elsevier B.V. All rights reserved. Keywords: Heat shock protein; HSP70; Characterization; Hydrothermal crab; Bythograeidae 1. Introduction Crabs of the Bythograeidae family (Eubrachyura, Decapoda) are the only endemic crab family living in hydrothermal fields (Martin and Haney, 2005; Desbruyères et al., 2006). This family includes the Allograea (1 species: Allograea tomentosa), Aus- tinograea (4 species), Bythograea (6 species), Cyanagraea (1 species: Cyanagraea praedator) genera present in the Pacific Ocean (except for Austinograea rodriguezensis, reported from the Indian ocean) and the Segonzacia (1 species: Segonzacia mesatlantica) genus which has been collected only in the Atlantic Ocean (Williams, 1980; Hessler and Martin, 1989; Tsuchida and Hashimoto, 2002; Guinot and Hurtado, 2003; Martin and Haney, 2005). The hydrothermal ecosystem depends on chemical interactions between the magma source and the surrounding seawater, which is released as superheated water (at about 300–350 °C) with various chemical compositions (Sarradin et al., 1999; Van Dover, 2000). The environmental conditions of the hydrothermal fields are thought to be rather unique, because they display high pressure, high levels of potentially toxic compounds such as hydrogen sulfide and heavy metals, and a low pH (Van Dover, 2000). The synthesis of the Heat Shock Proteins (HSP) is induced under stressful conditions (Feder, 1999; Grover, 2002). Environmental stresses, such as exposure to high temperature (Buckler et al., 2001; Spees et al., 2002), heavy metals (Pedersen and Lundebye, 1996), radiation (Nepple and Bachofen, 1997), changes in hydrostatic pressure (Kaarniranta et al., 1998), salinity (Gonza- lez and Bradley, 1994), polluting chemical compounds (hydrogen peroxide, organo-phosphate pesticides, sulhydryl reagents) (Levinson et al., 1980; Love et al., 1986; Werner and Nagel, 1997) and hypoxia/reoxygenation (Sanders and Martin, Gene 396 (2007) 84 – 92 www.elsevier.com/locate/gene Abbreviations: HSP, heat shock proteins. ⁎ Corresponding author. Laboratoire de Biologie et Génétique Evolutive, Université du Maine, UFR Sciences et Techniques, Avenue Olivier Messiaen, F-72085 Le Mans cedex 9, France. Tel.: +33 2 43833904; fax: +33 2 43833917. E-mail address: [email protected] (V. Leignel). 0378-1119/$ - see front matter © 2007 Elsevier B.V. All rights reserved. doi:10.1016/j.gene.2007.02.025
-
Upload
univ-lemans -
Category
Documents
-
view
0 -
download
0
Transcript of Identification of new subgroup of HSP70 in Bythograeidae (hydrothermal crabs) and Xanthidae

) 84–92www.elsevier.com/locate/gene
Gene 396 (2007
Identification of new subgroup of HSP70 in Bythograeidae(hydrothermal crabs) and Xanthidae
Vincent Leignel ⁎, Marie Cibois, Brigitte Moreau, Benoît Chénais
Laboratoire de Biologie et Génétique Evolutive (EA3265), Université du Maine, 72085, Le Mans, France
Received 13 November 2006; received in revised form 9 February 2007; accepted 21 February 2007
Available onlin
Received by M. Di Giulio
e 15 March 2007
Abstract
Crabs of the Bythograeidae family (Crustacea: Brachyura: Bythogreoidea) are the only endemic crab family living in hydrothermal fields. Thehydrothermal environment is characterized by unique ecological parameters, such as the high temperature gradient around the hydrothermalchimney (2–350 °C), a fluid environment containing high levels of metals and numerous gases. The 70-kDa Heat Shock Protein (HSP70) group isthe most-studied HSP, because it is ubiquitous, and a strong positive correlation has been found between the amounts of HSP70 produced inresponse to stress, and the ability of the organism to withstand stressful conditions. The 70-kDa heat shock protein genes from Bythograeids(species analyzed: Bythograea thermydron, Cyanagraea praedator and Segonzacia mesatlantica) were characterized. Our results revealed thatBythograeidae possess genes which are similar with those present in Xanthids (coastal crabs). The deduced protein sequences displayed motifsdistinct from those in the other crustacean HSC70/HSP70s available in the databases. Phylogenetic analysis showed that these members of HSP70family identified in Bythograeidae and Xanthidae constitute a new subgroup within this family.© 2007 Elsevier B.V. All rights reserved.
Keywords: Heat shock protein; HSP70; Characterization; Hydrothermal crab; Bythograeidae
1. Introduction
Crabs of the Bythograeidae family (Eubrachyura, Decapoda)are the only endemic crab family living in hydrothermal fields(Martin and Haney, 2005; Desbruyères et al., 2006). This familyincludes the Allograea (1 species: Allograea tomentosa), Aus-tinograea (4 species), Bythograea (6 species), Cyanagraea (1species: Cyanagraea praedator) genera present in the PacificOcean (except for Austinograea rodriguezensis, reported fromthe Indian ocean) and the Segonzacia (1 species: Segonzaciamesatlantica) genus which has been collected only in theAtlantic Ocean (Williams, 1980; Hessler and Martin, 1989;Tsuchida and Hashimoto, 2002; Guinot and Hurtado, 2003;
Abbreviations: HSP, heat shock proteins.⁎ Corresponding author. Laboratoire de Biologie et Génétique Evolutive,
Université du Maine, UFR Sciences et Techniques, Avenue Olivier Messiaen,F-72085 Le Mans cedex 9, France. Tel.: +33 2 43833904; fax: +33 2 43833917.
E-mail address: [email protected] (V. Leignel).
0378-1119/$ - see front matter © 2007 Elsevier B.V. All rights reserved.doi:10.1016/j.gene.2007.02.025
Martin and Haney, 2005). The hydrothermal ecosystem dependson chemical interactions between the magma source and thesurrounding seawater, which is released as superheated water (atabout 300–350 °C) with various chemical compositions(Sarradin et al., 1999; Van Dover, 2000). The environmentalconditions of the hydrothermal fields are thought to be ratherunique, because they display high pressure, high levels ofpotentially toxic compounds such as hydrogen sulfide andheavy metals, and a low pH (Van Dover, 2000). The synthesis ofthe Heat Shock Proteins (HSP) is induced under stressfulconditions (Feder, 1999; Grover, 2002). Environmental stresses,such as exposure to high temperature (Buckler et al., 2001;Spees et al., 2002), heavy metals (Pedersen and Lundebye,1996), radiation (Nepple and Bachofen, 1997), changes inhydrostatic pressure (Kaarniranta et al., 1998), salinity (Gonza-lez and Bradley, 1994), polluting chemical compounds(hydrogen peroxide, organo-phosphate pesticides, sulhydrylreagents) (Levinson et al., 1980; Love et al., 1986; Werner andNagel, 1997) and hypoxia/reoxygenation (Sanders and Martin,

85V. Leignel et al. / Gene 396 (2007) 84–92
1993), are all stimuli for the production of HSP. Within the cells,the HSP70s act by protecting the newly synthesized polypep-tides from misfolding, which aids their transportation acrossmembranes by preventing inappropriate interactions with othermolecules, the formation of multimeric complexes, and thestructural conformation of denatured proteins (Gullo and Teoh,2004; Mayer and Bukau, 2005). The HSP70s seem to be themost highly conserved and abundant group within the family of
Fig. 1. Example of the nucleotide sequence and deduced amino acid sequence of theaccession no. AM410073). The 189-bp intron in the open reading frame is shown i
stress proteins (Lindquist and Craig, 1988). The HSP70s havetwo active domains: an NH2-terminal, ATPase domain(44 kDa), and a COOH-terminal region of 25 kDa dividedinto a substrate-binding domain (∼15 kDa), and a COOH-terminal domain (10 kDa with a conserved EEVD terminalsequence) (James et al., 1997; Bukau and Horwich, 1998; Kiangand Tsokos, 1998). The HSP70 is approximately 650 aminoacids long. Generally, two closed cytosolic forms of HSP70
heat shock protein 70 kDa isolated from Bythograeidae (Cyanagraea praedator,n bold letters.

86 V. Leignel et al. / Gene 396 (2007) 84–92
coexist inside Eukaryota: constitutive HSP70, denoted asHSC70, and inducible HSP70, denoted as HSP70.
The aim of our investigation was to identify the 70-kDa heatshock protein genes fromhydrothermal crabs that live in particularenvironmental conditions. Three Bythograeids were studiedBythograea thermydron, C. praedator and S. mesatlantica. Amolecular comparison was carried out between the genes ofBythograeids and those of some others invertebrates (Arthropoda,Cnidaria, Echiura, Echnodermata, Mollusca, Nematoda, Plathel-minthes, and Porifera phyla).We also isolated the members of theHSP70 family from Portunidae and Xanthidae families, whichhave been suggested as being close to Bythograeidae.
2. Materials and methods
2.1. Biological materials
Hydrothermal crabs (Bythograeidae) were obtained duringthe oceanographic missionsMARVEL (1997) andHOPE (1999)using the submersible Nautile operating on the Mid-AtlanticRidge (Lucky Strike, 37°18′N, 1700 m depth), and East PacificRise (13°N, 2620 m depth). The hydrothermal crabs collectedwere identified as B. thermydron Williams, 1980, C. praedatorDe Saint Laurent, 1984, S. mesatlantica (Williams, 1988). B.thermydron and C. praedatorwere collected from the 13°N site,whereas the S. mesatlantica was found at the Lucky Strike site.Coastal crabs (Portunidae and Xanthidae) were also collected.Demania scaberrima (Walker, 1887) (Xanthidae) and Portu-nus sanguinolentus (Herbst, 1783) (Portunidae) were caught offthe Vietnamese coast (Nha Trang, 12.16N, 109.10E), whereasXantho incisus Leach, 1814 (Xanthidae), was found along theFrench coast (Noirmoutier, 46.58N 02.10W). The morpholog-ical identification of the coastal species was carried out
Fig. 2. Intron sequences of Bythograeidae (1) and Xanthidae (2) Hsp70 membersnucleotides. The positions of the introns are indicated in parenthesis.
according to Apel and Spiridonov (1998), Falciai and Minervini(1992) and Stephenson and Campbell (1959). All the organismscollected were preserved and conserved at −70 °C or in liquidnitrogen. The tissues were crushed using a sterile pestle andmortar cooled with liquid nitrogen. Then DNA was extractedusing standard procedure.
2.2. DNA amplifications, cloning and sequencing
The Hsp70 were amplified by PCR, using the primers HS1(forward), 5′-GGCGGATCCAGTAATTGGC-3′, and HS2 (re-verse), 5′- TAATCGACTTCCTCGATGGTKG-3′, designedfrom multiple alignment using Genedoc, version 2.6 (Nicholasand Nicholas, 1997), and included the Hsp70 cDNA fromcrustaceans available in Genbank™ database. The amplifica-tion mixture used was as follows: 100 ng of DNA added to 20 μlof a reaction mixture consisting of 1.5 mMMgCl2, 50 mMKCl,1 U Taq polymerase® (Promega), 1.25 μM of each primer, and0.2 mM dNTPs. The amplification conditions required were:35 cycles consisting of the initial denaturing (94 °C, 1 min),annealing (61 °C, 1 min) and extension (72 °C, 2 min 30 s)steps. The PCR products were integrated into the PCRII-TOPOplasmid (TA cloning®, Invitrogen). The recombined vector wasintegrated into TOP10 competent cells® (Invitrogen). Positivecolonies were identified by white/blue selection. The sequenc-ing reactions were carried out by Genome Express (Meylan,France, http://www.gexbyweb.com) using SP6 and T7-promo-tors, and successive internal primers.
2.3. Sequence analysis
The sequences obtained were identified using the FASTA andBLAST available at the NCBI GenBank biocomputing site
. The (.) symbol denotes the gap positions and the (-) indicate the conserved

Fig. 3. Multiple alignment of protein sequences deduced in silico, from HSP70 family members isolated from Decapoda. The HSP70 signature motifs are boxed. The (⁎) symbole denotes the gap positions. The speciesare numbered as follows: 1 (Pachygrapsus marmoratus, crab), 2 (Callinectes sapidus, crab), 3 (Portunus sanguinolentus, crab), 4 (Ferropenaeus chinensis, prawn), 5 (Homarus americanus, lobster), 6 (Macrobrachiumrosenbergii, shrimp), 7 (Cyanagraea praedator, crab), 8 (Segonzacia mesatlantica, crab), 9 (Demania scaberrima, crab), 10 (Xantho incisus, crab), 11–12 (Bythograea thermydron, crab).
87V.
Leignel
etal.
/Gene
396(2007)
84–92
88 V. Leignel et al. / Gene 396 (2007) 84–92
(Altschul et al., 1990; Pearson, 1990). The deduced amino acidsequences were obtained using Translate tool software, availableat the ExPASy (Expert Protein Analysis System) server of theSwiss Institute of Bioinformatics (http://www.expasy.org/tools/dna.htm). The multiple alignment of the sequenced fragmentswas carried out using ClustalW, version 1.83 (Thompson et al.,1994) and visualized using Genedoc, version 2.6 (Nicholas andNicholas, 1997). To eliminate any artifactual alignment errors,
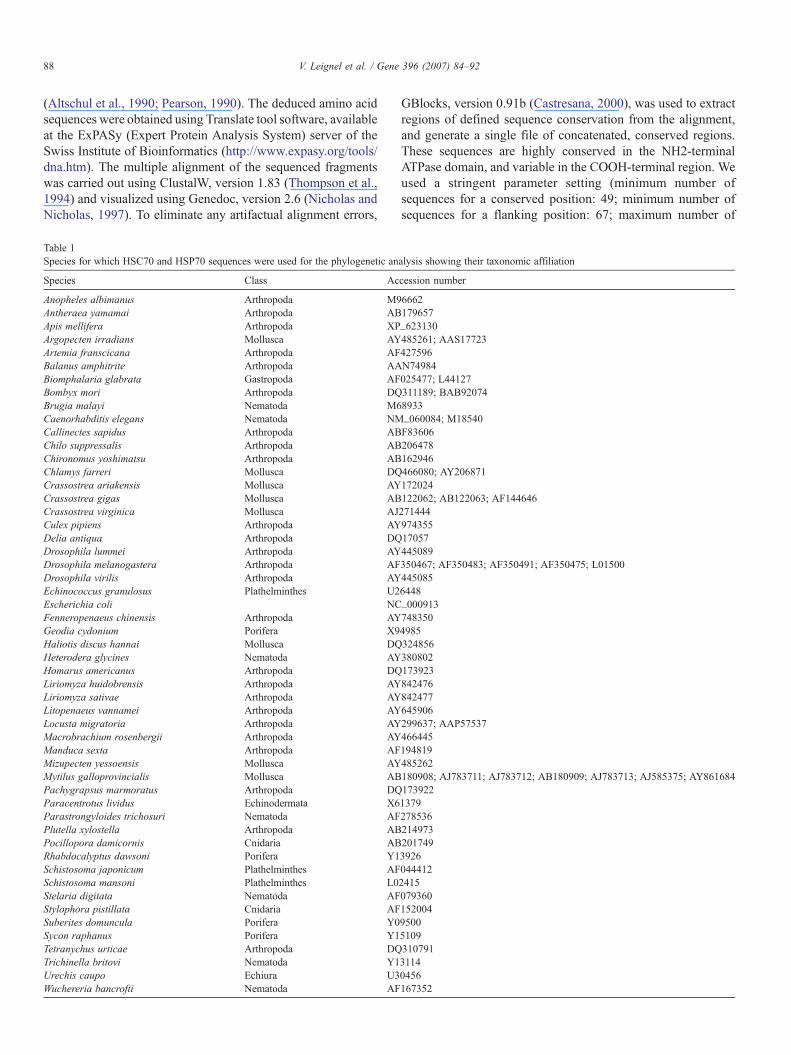
Table 1Species for which HSC70 and HSP70 sequences were used for the phylogenetic an
Species Class Ac
Anopheles albimanus Arthropoda M9Antheraea yamamai Arthropoda ABApis mellifera Arthropoda XPArgopecten irradians Mollusca AYArtemia franscicana Arthropoda AFBalanus amphitrite Arthropoda AABiomphalaria glabrata Gastropoda AFBombyx mori Arthropoda DQBrugia malayi Nematoda M6Caenorhabditis elegans Nematoda NMCallinectes sapidus Arthropoda ABChilo suppressalis Arthropoda ABChironomus yoshimatsu Arthropoda ABChlamys farreri Mollusca DQCrassostrea ariakensis Mollusca AYCrassostrea gigas Mollusca ABCrassostrea virginica Mollusca AJCulex pipiens Arthropoda AYDelia antiqua Arthropoda DQDrosophila lummei Arthropoda AYDrosophila melanogastera Arthropoda AFDrosophila virilis Arthropoda AYEchinococcus granulosus Plathelminthes U2Escherichia coli NCFenneropenaeus chinensis Arthropoda AYGeodia cydonium Porifera X9Haliotis discus hannai Mollusca DQHeterodera glycines Nematoda AYHomarus americanus Arthropoda DQLiriomyza huidobrensis Arthropoda AYLiriomyza sativae Arthropoda AYLitopenaeus vannamei Arthropoda AYLocusta migratoria Arthropoda AYMacrobrachium rosenbergii Arthropoda AYManduca sexta Arthropoda AFMizupecten yessoensis Mollusca AYMytilus galloprovincialis Mollusca ABPachygrapsus marmoratus Arthropoda DQParacentrotus lividus Echinodermata X6Parastrongyloides trichosuri Nematoda AFPlutella xylostella Arthropoda ABPocillopora damicornis Cnidaria ABRhabdocalyptus dawsoni Porifera Y1Schistosoma japonicum Plathelminthes AFSchistosoma mansoni Plathelminthes L0Stelaria digitata Nematoda AFStylophora pistillata Cnidaria AFSuberites domuncula Porifera Y0Sycon raphanus Porifera Y1Tetranychus urticae Arthropoda DQTrichinella britovi Nematoda Y1Urechis caupo Echiura U3Wuchereria bancrofti Nematoda AF
GBlocks, version 0.91b (Castresana, 2000), was used to extractregions of defined sequence conservation from the alignment,and generate a single file of concatenated, conserved regions.These sequences are highly conserved in the NH2-terminalATPase domain, and variable in the COOH-terminal region. Weused a stringent parameter setting (minimum number ofsequences for a conserved position: 49; minimum number ofsequences for a flanking position: 67; maximum number of
alysis showing their taxonomic affiliation
cession number
6662179657_623130485261; AAS17723427596N74984025477; L44127311189; BAB920748933_060084; M18540F83606206478162946466080; AY206871172024122062; AB122063; AF14464627144497435517057445089350467; AF350483; AF350491; AF350475; L015004450856448_0009137483504985324856380802173923842476842477645906299637; AAP57537466445194819485262180908; AJ783711; AJ783712; AB180909; AJ783713; AJ585375; AY8616841739221379278536214973201749392604441224150793601520049500510931079131140456167352
89V. Leignel et al. / Gene 396 (2007) 84–92
contiguous non-conserved positions: 8; minimum length of ablock: 10) to extract the conserved regions that contained morereliable phylogenetic signals. The phylogenetic analysis wasperformed using the maximum parsimony (MP) method, with
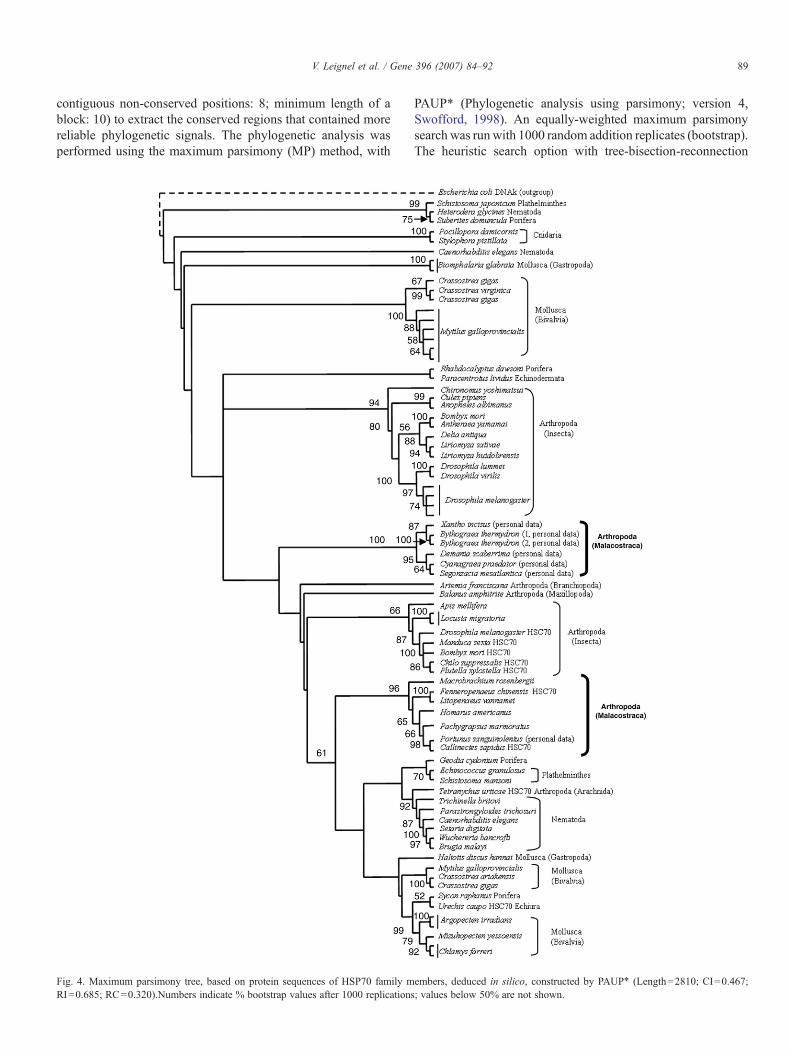
Fig. 4. Maximum parsimony tree, based on protein sequences of HSP70 family mRI=0.685; RC=0.320).Numbers indicate % bootstrap values after 1000 replications
PAUP⁎ (Phylogenetic analysis using parsimony; version 4,Swofford, 1998). An equally-weighted maximum parsimonysearch was runwith 1000 random addition replicates (bootstrap).The heuristic search option with tree-bisection-reconnection
embers, deduced in silico, constructed by PAUP⁎ (Length=2810; CI=0.467;; values below 50% are not shown.

90 V. Leignel et al. / Gene 396 (2007) 84–92
(TBR) branch swapping was used to find the best tree(s) for theprotein data set. The best trees were visualized by TreeView,version 1.6.6. (Page, 1996).
3. Results
3.1. Analysis of genes identified in coastal and hydrothermalcrabs
The coding sequence, from the start codon (ATG) to the stopcodon (TAA), ranged in length from 1950 and 2280 bp(Portunidae, Xanthidae and Bythograeidae families). Forexample C. praedator displayed an 70-kDa heat shock proteingene of 2097 bp (Fig. 1), and the predicted ORF encodes for635 amino acids (69.74 kDa). The nucleic sequences obtainedhave been submitted to the EMBL database: AM410071 toAM410077. The members of HSP70 family isolated from theBythograeids (B. thermydron, C. praedator and S. mesatlan-tica) and Xanthidae (D. scaberrima, X. incisus) displayed anintron (Fig. 2), whereas those from P. sanguinolentus(Portunidae) had no intron. The intron lengths ranged from127 (D. scaberrima, X. incisus) to 372 bp (S. mesatlantica:153 bp; C. praedator: 189 bp; B. thermydron (two individuals):from 370 to 372 bp). The intron was located at the same positionin the genes isolated from both families, and displayed similarsplicing signal (5′GTgAGTGtYTYGCCGC—TTCTCCCTCa-CAG 3′) and similar internal flanking regions (Fig. 2). Theseintrons also contained one or two microsatellite(s) (CA)n was/were observed in intron. The number of (CA) replicates rangedfrom 16 (D. scaberrima and X. incisus) to 45 (S. mesatlantica:33; C. praedator: 45). B. thermydron (2 individuals) had theparticular characteristic of possessing two microsatellites (CA)n, flanking a large insertion (189 bp) (Fig. 2). The length of thefirst microsatellite (CA)n of the two B. thermydron analysedwas (CA)22 and (CA)24; whereas the second microsatellite wassimilar ((CA)15). This difference in the first (CA)n could becaused by a sequencing error. The large insertion observed in B.thermydron and flanked by the microsatellites (CA)n was notidentified using the BLAST and FASTA programs. The proteinsencoded by the genes isolated from Bythograeidae, Portunidaeand Xanthidae displayed the conserved motifs characteristic ofthe HSP70 family: GIDLGTTY, DLGGGTFD, IVLVGG-STRIPKIQL, KRKYKKDPSENKRALRRLRTACERAKRTL(nuclear localization signal, NLS), and the final motif: EEVD(Liu et al., 2004; Demand et al., 1998; Rensing and Maier,1994). The comparison at both nucleic and protein levelsrevealed that the 70-kDa heat shock proteins from Bythograei-dae and Xanthidae constituted a distinct group. Theirs proteinsshowed some distinct amino acid residues notably in the NLS(Fig. 3). The protein divergence between Bythograeidae/Xanthidae group and the others crustacean HSC70/HSP70sequences was estimated to be 20.2%, using Genedoc.
3.2. Phylogenetic analysis
To determine the relationship between the members of theHSP70 family isolated from invertebrates, we carried out a
phylogenetic analysis (maximum parsimony, PAUP⁎) including70 HSC70/HSP70 sequences from Arthropoda, Cnidaria,Echiura, Echnodermata, Mollusca, Nematoda, Plathelminthesand Porifera phyla extracted from GenBank™ database. Theaccession number of these HSC70/HSP70 sequences is shownin Table 1. The phylogenetic trees were based on the GBlocksalignment with 519 selected positions out of 678 available afterClustalW alignment. All characters had equal weight. Out of atotal of 519 characters, 103 were constant, 109 were variable butparsimony uninformative, and 307 were informative. Parsimo-ny analysis (heuristic search option with tree-bisection-reconnection branch swapping) identified the 3 shortest trees(Length=2810; CI=0.467; RI=0.685; RC=0.320). Eachphylum (Arthropoda, Mollusca, Nematoda, Platyhelminthes,Porifera) displayed two HSP70 groups (Fig. 4). The proteinsfrom Bythograeidae and Xanthidae, appeared to be distinctfrom the HSC70/HSP70s of others crustaceans (barnacle, crab,prawn, shrimp, lobster). In spite of the nucleic difference(intron), indicated in Fig. 2, the phylogenetic relationship basedon protein sequences did not distinguish between members ofthe Bythograeidae and Xanthidae. D. scaberrima was close toC. praedator and S. mesatlantica, whereas X. incisus wasgrouped with B. thermydron.
4. Discussion
The members of HSP70 family identified in Bythograeidae(hydrothermal crabs) and Xanthidae (coastal crabs) displaymultiple similarities and seem to constitute a new subgroup inthe HSP70 family. Their genes present an intron including one ortwo microsatellite (CA)n and similar splicing signals. Thepresence of the microsatellite (CA)n within the intron sequencessuggests that this intron has evolved rapidly by increases in the(CA) repeat. The 189-bp insertion, localized in the intron,observed in B. thermydron may be explained by geneticrecombination. The proteins deduced in silico present multiplespecific amino acid residues by comparison with othercrustacean HSC70/HSP70s. These specific amino acid residueswere noted along the sequence, and notably in the nuclearlocalization signal which play a function in the translocation ofthe heat shock proteins into the nucleus (Knowlton and Salfity,1996). To differentiate between the HSC70 and HSP70 forms,some authors have indicated thatHsp70 do not possess an intron(Gunther and Walter, 1994; Boutet et al., 2003a,b). IntronlessHsp70 (that encode inducible HSP70) may respond morerapidly to stress conditions than intron-containing Hsc70(encoding constitutive HSP70), because RNA splicing isunnecessary. Indeed many marine invertebrate Hsp70 do notpossess any intron such as in Crassostrea gigas (Boutet et al.,2003b), Litopenaeus vannamei (AY645906, not published);Macrobrachium rosenbergii (Liu et al., 2004); Mytilus gallo-provincialis (Kourtidis and Scouras, 2005) and Ostrea edulis(Boutet et al., 2003a). Nevertheless, some introns were observedin the coding region of several Hsp70 in Blastocladiellaemersonii (Stephani and Gomes, 1995),Caenorhabditis elegans(Snutch et al., 1988), Locusta migratoria (Qin et al., 2003),Neurospora crassa (Kapoor et al., 1995) and Paracentrotus

91V. Leignel et al. / Gene 396 (2007) 84–92
lividus (Sconzo et al., 1992). So, the location of the intron isspecific for each organism. This means that the presence/absenceof the intron cannot be used to distinguish between Hsc70 andHsp70. The comparison of protein sequences, deduced in silico,showed that members of HSP70 family isolated in Bythograei-dae and Xanthidae display a shorter COOH-terminal region andcytosolic motifs, such as EEVD, but did not display any GAP orGGMPmotifs. None of the structural characteristics can be usedto differentiate between HSC70 and HSP70. Consequently, thebest criteria to use to distinguish between Hsc70 and Hsp70 stillseem to be gene expression studies.
The presence of similar members of HSP70 family identifiedin Bythograeidae and Xanthidae could be considered as acommon ancestral character? Recently, some studies havesuggested that Bythograeidae could be derived from anancestral form close to the Potamoidea, Portunoidea orXanthoidea superfamilies. Delamare-Deboutteville and Guinot(1981) found marked morphological similarity between Bytho-graeids and Typhlopseudothelphusa mocinoi (Pseudothelphusi-dae, Potamoidea), both of which display a loss of carapacepigmentation and a small ocular system. Nevertheless, theseauthors concluded that deducing any relationship between thesecrabs seems to be hazardous. Guinot (1988) suggested that theorigin and evolution of hydrothermal crabs (after studying theBythograea and Cyanagraea genera) may not be derivedfrom bathyal ancestral crabs. After an ultrastructural investiga-tion of their spermatozoal morphology, Tudge et al. (1998)suggested a similarity between Bythograeidae (Bythograea,Austinograea and Segonzacia genera analyzed) and Calocar-cinus africanus (Xanthoidea: Trapeziidae). In the conclusion oftheir analysis, Tudge et al. (1998) proposed that C. africanusmay be included in Bythograeids sensu lato. These authorssuggested that the origin of Bythograeids may be a Calocarci-nus-like, deep-water xanthoid form, which may have colonizedthe hydrothermal system in/after the Eocene period. Inagreement with Tudge et al. (1998), Sternberg et al. (1999)established a morphology-based cladistic analysis from multi-ple crab families that suggested that there is a close relationshipbetween the Bythograeids and Xanthoids. In contrast, Sternbergand Cumberlidge (2003), by their examination of the thoracicendophragmal system, have shown that Bythograeids havesternite patterns organized in a similar way to those ofTrichodactylids (Portunoidea). Our results indicate a closerelationship between the 70-kDa heat shock protein genes fromthe Bythograeids and Xanthids and match closely the conclu-sions of Guinot (1988), Tudge et al. (1998) and Sternberg et al.(1999). In this case, these genes could derive from an inheritedgene from a common ancestral crab (perhaps an ancestral formlike Calocarcinus, according to Tudge et al., 1998) living in ahydrothermal environment.
Acknowledgements
We would like to thank Drs. D. Desbruyères, F. Lallier, chiefscientists of the HERO-1997 and HOPE-99 missions and theNautile submersible pilots of the crew of Atalante. We alsoacknowledged Drs. D. Desbruyères, D. Jollivet and S. Hourdez
who identified the hydrothermal crabs. The authors are gratefulto Monika Ghosh for revising the English text.
References
Altschul, S.F., Gish, W., Miller, W., Myers, E.W., Lipman, D.J., 1990. Basiclocal alignment search tool. J. Mol. Biol. 215, 403.
Apel, M., Spiridonov, V.A., 1998. Taxonomy and zoogeography of the portunidcrabs (Crustacea: Decapoda: Brachyura: Portunidae) of the Arabian Gulfand adjacent waters. Fauna Arab. 17, 159–331.
Boutet, I., Tanguy, A., Moraga, D., 2003a. Organization and nucleotide sequenceof the European flat oyster Ostrea edulis heat shock cognate 70 (hsc70) andheat shock protein 70 (hsp70) genes. Aquat. Toxicol. 65, 221–225.
Boutet, I., Tanguy, A., Rousseau, S., Auffret, M., Moraga, D., 2003b. Molecularidentification and expression of heat shock cognate 70 (hsc70) and heatshock protein 70 (hsp70) genes in the Pacific oyster Crassostrea gigas. CellStress Chaperones 8 (1), 76–85.
Buckler, B.A., Owen, M.E., Hofmann, G.E., 2001. Adjusting the thermostat: thethreshold induction temperature for the heat-shock response in intertidalmussels (genus Mytilus) changes as a function of thermal history. J. Exp.Biol. 204, 3571–3579.
Bukau, B., Horwich, A.L., 1998. The Hsp70 and Hsp60 chaperone machines.Cell 92, 351–366.
Castresana, J., 2000. Selection of conserved blocks from multiple alignments fortheir use in phylogenetic analysis. Mol. Biol. Evol. 17, 540–552.
Delamare-Deboutteville, C., Guinot, D., 1981. Considérations sur lesBythograeoidea Williams, nouvelle superfamille de crabes de la dorsalePacifique Est, conférence présentée à la VIIe Réunion des Carcinologistes delangue française, Banyuls-Sur-Mer, France, 1–6 juin 1981.
Demand, J., Lüders, J., Höhfeld, J., 1998. The carboxy-terminal domain ofHSC70 provides binding sites for a distinct set of chaperone cofactors. Mol.Cell. Biol. 18, 2023–2028.
Desbruyères, D., Segonzac, M., Bright, M., 2006. Handbook of Deep-Seahydrothermal vent fauna. Denisia 18, 1–544.
Falciai, L., Minervini, R., 1992. In: Muzzio, F., Editore, C. (Eds.), Guida deiCrostacei Decapodi d'Europa, pp. 1–287. Padova, Italia.
Feder, M.E., 1999. Heat-shock proteins, molecular chaperones, and the stressresponse: Evolutionary and Ecological Physiology. Annu. Rev. Physiol. 61,243–282.
Gonzalez, C.R.M., Bradley, B.P., 1994. Are there salinity stress proteins ? Mar.Enivron. Res. 35, 79–83.
Grover, A., 2002. Meeting review: molecular biology of stress responses. CellStress Chaperones 7, 1–5.
Guinot, D., 1988. Les crabes des sources hydrothermales de la dorsale duPacifique oriental (campagne Biocyarise, 1984). Oceanol. Acta 8 (SpecialIssue), 109–118.
Guinot, D., Hurtado, L.A., 2003. Two new species of hydrothermal vent crabs of thegenus Bythograea from the southern East Pacific Rise and from the GalapagosRift (CrustaceaDecapodaBrachyuraBythograeidae). C.R., Biol. 326, 423–439.
Gullo, C.A., Teoh, G., 2004. Minireview: heat shock proteins: to present or not,that is the question. Immunol. Lett. 94, 1–10.
Gunther, E., Walter, L., 1994. Genetic aspects of the hsp70 multigene family invertebrates. Experientia 50, 987–1001.
Hessler, R.R., Martin, J.W., 1989. Austinograea williamsi, new genus, newspecies, a hydrothermal vent crab (Decapoda: Bythograeidae) from theMariana Back-Arc Basin, western Pacific. J. Crustac. Biol. 9 (4), 645–661.
James, P., Pfund, C., Craig, E.A., 1997. Functional specificity among Hsp70molecular chaperones. Science 275, 387–389.
Kaarniranta, K., et al., 1998. Hsp70 accumulation in chondrocytic cells exposedto high continous hydrostatic pressure coincides with mRNA stabilizationrather than transcriptional activation. Proc. Natl. Acad. Sci. U. S. A. 95,2319–2324.
Kapoor, M., Curle, C.A., Runham, C., 1995. The hsp70 family of Neurosporacrassa: cloning, sequence analysis, expression, and genetic mapping of themajor stress-inducible member. J. Bacteriol. 177 (1), 212–221.
Kiang, J.G., Tsokos, G.C., 1998. Heat Shock Protein 70 kDa: molecular biology,biochemistry, and physiology. Pharmacol. Ther. 80, 183–201.

92 V. Leignel et al. / Gene 396 (2007) 84–92
Knowlton, A.A., Salfity, M., 1996. Nuclear localisation signal and the heatshock proteins. J. Biosci. 21, 123–132.
Kourtidis, A., Scouras, Z.G., 2005. Analysis and characterization of thetranscriptional unit of a new Mytilus galloprovincialis (Mollusca: Bivalvia)hsp70 gene. DNA Seq. 16 (1), 36–43.
Levinson, W., Oppermann, H., Jackson, J., 1980. Transition series metals andsulhydryl reagents induce the synthesis of four proteins in eukaryotic cells.Biochem. Biophys. Acta 606, 170–180.
Lindquist, S., Craig, E.A., 1988. The heat shock proteins. Annu. Rev. Genet. 22,631–637.
Liu, J., Yang, W.J., Zhu, X.J., Karouna-Renier, N.K., Rao, R.K., 2004.Molecular cloning and expression of two HSP70 genes in the prawn, Ma-crobrachium rosenbergii. Cell Stress Chaperones 9 (3), 313–323.
Love, J., Vivino, A.A., Minton, K.W., 1986. Hydrogen peroxide toxicity may beenhanced by heat shock gene induction in Drosophila. J. Cell. Physiol. 126,60–68.
Martin, J.W., Haney, T., 2005. Decapod crustaceans from hydrothermal ventsand cold sepps: a review through 2005. Zool. J. Linn. Soc. 145, 445–522.
Mayer, M.P., Bukau, B., 2005. Hsp70 chaperones: cellular functions andmolecular mechanism. Cell. Mol. Life Sci. 62, 670–684.
Nepple, B.B., Bachofen, R., 1997. Induction of stress proteins in thephototrophic bacterium Rhodobacter sphaeroides. FEMS Microbiol. Lett.153, 173–180.
Nicholas, K.B., Nicholas, H.B., 1997.GENEDOC aTool for Editing andAnnotingMultiple Sequence Alignments. http://www.psc.edu/biomed/genedoc.
Page, R.D., 1996. TreeView: an application to display phylogenetic trees onpersonal computers. Comput. Appl. Biosci. 12, 357–358.
Pearson, W.R., 1990. Rapid and sensitive sequence comparison with FASTP andFASTA. Methods Enzymol. 183, 63–98.
Pedersen, S.N., Lundebye, A.K., 1996. Metallothionein and stress protein levelsin shore crabs (Carcinus maenas) along a trace metal gradient in the FalEstuary System (UK). Mar. Environ. Res. 42, 241–246.
Qin, W., Tyshenko, M.G., Wu, B.S., Walker, V.K., Robertson, R.M., 2003.Cloning and characterization of a member of the hsp70 gene family fromLocusta migratoria, a highly thermotolerant insect. Cell Stress Chaperones2, 144–152.
Rensing, S.A., Maier, U.G., 1994. Phylogenetic analysis of the stress-70 proteinfamily. J. Mol. Evol. 39, 80–86.
Sanders, B.M., Martin, L.S., 1993. Stress proteins as biomarkers of contaminantexposure in archived environmental samples. Sci. Total Enivron. 1, 459–470.
Sarradin, P.M., Caprais, J.C., Riso, R., Kerouel, R., Aminot, A., 1999. Chemicalenvironment of the hydrothermal mussel communities in the Lucky Strike andMenez Gwen vent fields, Mid Atlantic Ridge. Cah. Biol. Mar. 40, 93–104.
Sconzo, G., Scardina, G., Ferraro, M.G., 1992. Characterization of a newmember of the sea urchin Paracentrotus lividus hsp70 gene family and itsexpression. Gene 121 (2), 353–358.
Snutch, T.P., Heschl, M.F., Baillie, D.L., 1988. The Caenorhabditis eleganshsp70 gene family: a molecular geneticx characterization. Gene 64,241–255.
Spees, J.L., Chang, S.A., Snyder, M.J., Chang, E.S., 2002. Thermal acclimationand stress in the amercian lobster, Homarus americanus: equivalenttemperature shifts elicit unique gene expression patterns for molecularchaperones and polyunbiquitin. Cell Stress Chaperones 7 (1), 97–106.
Stephani, R.M.P., Gomes, S.L., 1995. A unique intron-containing hsp70 geneinduced by heat shock and during sporulation in the aquatic fungus Blas-tocladiella emersonii. Gene 152, 19–26.
Stephenson, W., Campbell, B., 1959. The australian portunids (Crustacea:Portunidae). III. The genus Portunus. Aust. J. Mar. Freshw. Res. 10 (1),84–124.
Sternberg, R.N.V., Cumberlidge, N., 2003. Autapomorphies of the enphragmalsystem in Trichodactylid freshwater crabs. J. Morphol. 256, 23–28.
Sternberg, R.N.V., Cumberlidge, N., Rodriguez, G., 1999. On the marine sistergroups of the freshwater crabs (Crustacea: Decapoda: Brachyura). J. Zoolog.Syst. Evol. Res. 37, 19–38.
Swofford, D.L., 1998. PAUP⁎. Phylogenetic Analysis Using Parsimony (⁎andOthers Methods). Version 4. Sinauer Associates. ©Smithsonian Institution,Sunderland, Massachusetts.
Thompson, J.D., Higgins, D.G., Gibson, T.J., 1994. CLUSTALW improving thesensitivity of progressive multiple sequence alignment through sequenceweighting, positions specific gap penalties and weight matrix choice.Nucleic Acids Res. 22, 4673–4680.
Tsuchida, S., Hashimoto, J., 2002. A new species of Bythograeid crab, Austi-nograea rodriguezensis (Decapoda, Brachyura), associated with activehydrothermal vents from the Indian ocean. J. Crustac. Biol. 22 (3), 642–650.
Tudge, C.C., Jamieson, B.G.M., Segonzac, M., Guinot, D., 1998. Spermatozoalultrastruture in three species of hydrothermal vent crab, in the generaBythograea, Austinograea and Segonzacia (Decapoda, Brachyura, Bytho-graeidae). Invertebr. Reprod. Dev. 34 (1), 13–23.
Van Dover, C.L., 2000. The ecology of deep-sea hydrothermal vents. Princetonuniversity press, Princeton, New Jersey. 424 pp.
Werner, I., Nagel, R., 1997. Stress proteins HSP60 and HSP70 in three speciesof amphipods exposed to cadmium, diazinon, dieldrin, and fluoranthene.Environ. Toxicol. Chem. 16, 2393–2403.
Williams, A.B., 1980. A new crab family from the vicinity of submarine thermalvents on the Galapagos Rift (Crustacea: Decapoda: Brachyura). Proc. Biol.Soc. Wash. 93, 443–472.




















