
Nucleotide utilization requirements that render ClpB active as a chaperone
Upload
cumhuriyetCategory
view
0download
0
10.1128/EC.4.2.289-297.2005.
2005, 4(2):289. DOI:Eukaryotic Cell Eisenberg, Lois E. Greene and Daniel C. MasisonYoutao Song, Yue-xuan Wu, Giman Jung, Yusuf Tutar, Evan Replication
Prion SeedSaccharomyces cerevisiaeRole for Hsp70 Chaperone in
http://ec.asm.org/content/4/2/289Updated information and services can be found at:
These include:
REFERENCEShttp://ec.asm.org/content/4/2/289#ref-list-1at:
This article cites 53 articles, 28 of which can be accessed free
CONTENT ALERTS more»articles cite this article),
Receive: RSS Feeds, eTOCs, free email alerts (when new
http://journals.asm.org/site/misc/reprints.xhtmlInformation about commercial reprint orders: http://journals.asm.org/site/subscriptions/To subscribe to to another ASM Journal go to:
on August 5, 2014 by guest
http://ec.asm.org/
Dow
nloaded from
on August 5, 2014 by guest
http://ec.asm.org/
Dow
nloaded from

EUKARYOTIC CELL, Feb. 2005, p. 289–297 Vol. 4, No. 21535-9778/05/$08.00�0 doi:10.1128/EC.4.2.289–297.2005
Role for Hsp70 Chaperone in Saccharomyces cerevisiaePrion Seed Replication
Youtao Song,1 Yue-xuan Wu,2 Giman Jung,1† Yusuf Tutar,1 Evan Eisenberg,2Lois E. Greene,2 and Daniel C. Masison1*
Laboratory of Biochemistry and Genetics, National Institute of Diabetes Digestive and Kidney Diseases,1
and Laboratory of Cell Biology, National Heart, Lung, and Blood Institute,2
National Institutes of Health, Bethesda, Maryland
Received 26 October 2004/Accepted 14 December 2004
The Saccharomyces cerevisiae [PSI�] prion is a misfolded form of Sup35p that propagates as self-replicatingcytoplasmic aggregates. Replication is believed to occur through breakage of transmissible [PSI�] prion par-ticles, or seeds, into more numerous pieces. In [PSI�] cells, large Sup35p aggregates are formed by coalescenceof smaller sodium dodecyl sulfate-insoluble polymers. It is uncertain if polymers or higher-order aggregates orboth act as prion seeds. A mutant Hsp70 chaperone, Ssa1-21p, reduces the number of transmissible [PSI�]seeds per cell by 10-fold but the overall amount of aggregated Sup35p by only two- to threefold. This discrep-ancy could be explained if, in SSA1-21 cells, [PSI�] seeds are larger or more of the aggregated Sup35pdoes not function as a seed. To visualize differences in aggregate size, we constructed a Sup35-green fluorescentprotein (GFP) fusion (NGMC) that has normal Sup35p function and can propagate like [PSI�]. Unlike GFPfusions lacking Sup35p’s essential C-terminal domain, NGMC did not form fluorescent foci in log-phase[PSI�] cells. However, using fluorescence recovery after photobleaching and size fractionation techniques, wefind evidence that NGMC is aggregated in these cells. Furthermore, the aggregates were larger in SSA1-21 cells,but the size of NGMC polymers was unchanged. Possibly, NGMC aggregates are bigger in SSA1-21 cellsbecause they contain more polymers. Our data suggest that Ssa1-21p interferes with disruption of large Sup35paggregates, which lack or have limited capacity to function as seed, into polymers that function more efficientlyas [PSI�] seeds.
The Saccharomyces cerevisiae [PSI�] prion is a self-repli-cating altered form of the essential translation release factorSup35p that is cytoplasmically transmissible (infectious) (37,42, 50, 54). Purified Sup35p spontaneously assembles into amy-loid, which is a highly ordered fibrous aggregate that grows byrecruiting protein into the fibers, after a lag time thought to berequired for formation of amyloid nuclei. Adding fragmentsresulting from breakage of mature fibers eliminates the lag inamyloid formation by providing preformed nuclei. Althoughthe precise physical properties of infectious forms of Sup35pare uncertain, there is ample evidence suggesting that suchentities, or [PSI�] “seeds,” are an amyloid form of Sup35p(6, 9, 17, 25, 26, 28, 41, 43, 44, 51). In particular, [PSI�] can beinduced to appear in yeast cells by infection with Sup35p amy-loid produced in vitro (25, 43).
In order for [PSI�] to be maintained in a rapidly dividingyeast population, the number of [PSI�] seeds must at leastdouble in number in the time that a cell divides. Hsp104, aprotein chaperone that protects cells from exposure to stressby resolubilizing denatured protein from aggregates, is thoughtto facilitate [PSI�] seed generation by breaking Sup35p aggre-
gates into more numerous pieces (5, 27, 35, 37, 39). In vitro,protein disaggregation by Hsp104 requires the assistance ofHsp70 (18). Hsp70s are conserved protein chaperones thatassist protein folding in general and prevent aggregation ofpartially folded proteins. They are important for many cellularprocesses, such as folding of nascent polypeptides, disassem-bling clathrin, and transporting protein across membranes, andthey protect cells from exposure to environmental stresses (4,16, 32).
S. cerevisiae has four cytosolic Hsp70s of the Ssa subfamily,and expression of at least one of them is essential for growth(49). Ssa1p is one of two constitutively expressed Ssa Hsp70s.We earlier identified a dominant mutant of Ssa1p (Ssa1-21p)that impairs propagation of the yeast [PSI�] prion, weakeningthe phenotype caused by [PSI�] and causing frequent mitoticloss of [PSI�] (23). This [PSI�] instability is due to a reductionin the number of [PSI�] seeds per cell, which suggests thatSsa1p is involved in [PSI�] seed generation.
Kryndushkin et al. showed that high-molecular-weight ma-terial containing Sup35p from [PSI�] cells consists of largeaggregates and smaller sodium dodecyl sulfate (SDS)-insoluble“polymers” of Sup35p (27). In the absence of SDS the poly-mers fractionate with the larger aggregates, which are up to30-fold larger than the polymers. Under denaturing conditionsthe large aggregates fall apart, but SDS-resistant polymersof a size equivalent to 8 to 50 Sup35p monomers remain. Theauthors propose that the large aggregates are composed ofindividual Sup35p polymers that have self-associated and thatHsp104 generates new [PSI�] seeds by fragmenting polymers
* Corresponding author. Mailing address: Laboratory of Biochem-istry and Genetics, National Institute of Diabetes Digestive and Kid-ney Diseases, National Institutes of Health, Bethesda, MD 20892.Phone: (301) 594-1316. Fax: (301) 496-9431. E-mail: [email protected].
† Present address: New Frontiers Research Laboratories, Toray In-dustries, Inc., Kanagawa, Japan.
289
on August 5, 2014 by guest
http://ec.asm.org/
Dow
nloaded from

into more numerous pieces and by dismantling Sup35p aggre-gates into individual polymers.
Although SSA1-21 mutants have a large reduction in [PSI�]seeds per cell, fractionation of Sup35p by centrifugation showsthat there is only a slight reduction in the amount of aggre-gated Sup35p in SSA1-21 cells (23). This discrepancy could beexplained if the [PSI�] seeds in SSA1-21 cells are larger thannormal or if more of the aggregated Sup35p in SSA1-21 cellsdoes not function as [PSI�] seeds.
To determine visually if Sup35p aggregated to a greaterextent in live SSA1-21 cells, we replaced Sup35p with a Sup35-green fluorescent protein (GFP) fusion that functions normallyin translation termination and in [PSI�] propagation. In con-trast to what is seen with widely used GFP fusions lacking theessential C-terminal domain Sup35p, visual differences in GFPfluorescence between [PSI�] and [psi–] were not obvious inlog-phase cells. Using more sensitive techniques, however, wefind evidence of Sup35p aggregates in the [PSI�] cells and thatthese aggregates are larger in SSA1-21 cells. We further findthat SSA1-21 has little effect on the size of Sup35p polymers in[PSI�] cells, suggesting that Sup35p aggregates in SSA1-21cells are larger because they contain more polymers. We pre-sent a new technique for analyzing aggregation of prion pro-teins in S. cerevisiae and provide insight into the role of Hsp70in yeast prion propagation.
MATERIALS AND METHODS
Strains, media, and growth conditions. The yeast strains are listed in Table 1.Strain 779-6A is [PSI�], which is the origin of [PSI�] for all strains derived from779-6A. Strain 74D-694 and its sup35::TRP1 derivative (designated 74-D694s)were gifts from Y. Nakamura (19). Where strains lacking [PSI�] were used,[PSI�] was eliminated by growth on medium containing 3 mM guanidine hydro-chloride. The SSA1-21 allele of SSA1 encodes a tryptophan in place of leucine atcodon 483 (23). Gene replacements at the chromosomal SSA1 and SSA2 locihave been described (21).
Strain J780 is a [PSI�] diploid formed by a cross of strains 1001 and 1013, inwhich one SUP35 allele was replaced with KanMX by transformation with DNAamplified by PCR (2, 47). To obtain strains with wild-type and GFP-taggedSUP35 (designated NGMC), strain J780 was first transformed by pJ501 (seebelow), which is a HIS3-based plasmid carrying wild-type SUP35. J780/pJ501meiotic segregants with pJ501 that are sup35::KanMX and either wild type (780-1D) or SSA1-21 (780-1C) were selected. To replace SUP35 with NGMC, 780-1Cand 780-1D were first transformed with pJ510 (see below), which is a LEU2-based plasmid with NGMC. Sup35p is essential for growth, so transformantsmust maintain either pJ501 or pJ510. Moreover, when cells have either pJ501 orpJ510, the plasmid is maintained without selection (i.e., on rich medium). Aftergrowing transformants having both plasmids on rich medium, clones were iden-tified that had lost pJ501 and retained pJ510. All such clones retained a typical[PSI�] phenotype, indicating that the resident [PSI�] formed of the untaggedSup35p (expressed from pJ501) induced conversion of NGMC to a stable [PSI�]form (see below). A similar plasmid shuffle was used to replace Sup35p withNGMC in strain 74-D694s.
1/2YPD medium contains 0.5% yeast extract, 2% peptone, and 2% dextrose.YPAD is similar but contains 1% yeast extract and 400 mg of adenine/liter.1/2YPD/G3 is 1/2YPD containing 3 mM guanidine. Solid media contained 2%agar. Growth conditions were as described (23) or as indicated. Cytoplasmictransmission of prions between strains (cytoduction) was done as described (23).
Estimation of [PSI�] seed numbers. Quantitative assays were a slight modi-fication of a previously described method (7). Cells grown on selection plateslacking adenine at 25°C were streaked onto 1/2YPD plates and grown for 1, 2,and 4 days at 30°C. Cells were then restreaked onto 1/2YPD/G3 and grown for40 h at 30°C. Entire single colonies were picked up and diluted in 200 �l of water,spread onto selection plates lacking adenine, and grown for 2 weeks at 25°C. Theresulting colonies were replica plated onto 1/2YPD and 1/2YPD/G3 plates,which were then incubated for 2 days at 30°C followed by 3 or more days at 25°C.
Colonies that were white or pink on 1/2YPD and red on 1/2YPD/G3 were scoredas [PSI�].
Plasmids. Plasmid pJ501 is single-copy HIS3 plasmid pRS313 (40) with SUP35controlled by its own promoter on an XhoI-XbaI fragment PCR amplified fromstrain 628-3A (23). Plasmid pJ509 is pJ501 with GFP between the N and Mregions of SUP35 (NGMC, see Results). It was built by first adding in-frameBamHI and EcoRI sites between codons 123 and 124 of SUP35 in pJ501, creatingpJ508. GFP from plasmid pFA6a-GFP(S65T)-TRP1 (30) was then amplifiedwith primers with in-frame flanking BamHI and EcoRI sites, digested withBamHI and EcoRI, and inserted into pJ508 cut with the same enzymes. In ourhands, in-frame fusions of GFP at the N or C terminus or between M and C didnot support growth when expressed in place of Sup35p. Plasmid pJ510 is pRS315with the NGMC allele on an ApaI-NotI fragment from pJ509. Plasmid pH317/NMG is multicopy LEU2 plasmid pH317 (12) with SUP35NM-GFP on aBamHI-PvuII DNA fragment from p316CUP1-3SGFPSG (29), under control ofthe Gal1-10 promoter.
Microscopy. Microscopic analysis of the NGMC and NM-GFP fusion proteinswas done with a Nikon E-800 microscope with cells grown as indicated on YPADplates or in YPAD liquid medium. Fluorescent images were captured with IPlabsoftware and processed with Adobe Photoshop software.
Confocal microscopy and fluorescence recovery after photobleaching. Gain israised to the maximum of the linear range prior to the photobleach. A definedregion was repeatedly photobleached at 100% laser power, resulting in 50 to 80%reduction in fluorescence intensity. Fluorescence recovery was monitored byscanning at 2% laser power. In order to get time points in the subsecond timerange when measuring the fluorescence recovery of photobleached highly dif-fusible protein, the scanning distance along the ordinate axis was markedlyreduced. When data sets were compared, the cells were photobleached andmonitored identically including the number of bleaches, the area of the photo-bleach region, and the time course of imaging at low laser power. This methodenables us to detect quantitative differences in mobility of the GFP proteinswhich are diffusing freely in the cell.
Preparation of S. cerevisiae cell lysates. Cell lysates were prepared as described(27) from cultures grown in YPAD to an optical density at 600 nm of 1.5. Celldebris was removed by centrifugation at 5,000 � g for 8 min. Protein concen-trations were determined according to the protein assay kit (Bio-Rad).
Column fractionation. Twenty milligrams of protein from cell lysates wasloaded on a Hiprep Sephacryl 500 (16 by 88 cm) column (Amersham PharmaciaBiotech) with the AKTA-fast protein liquid chromatography system. The run-ning buffer contains 50 mM Tris-HCl (pH 7.4) and 200 mM NaCl, and 2-mlfractions were collected. Samples (40 �l) were added to 10 �l of SDS samplebuffer (five times) and boiled for 10 min, and proteins were separated by SDS–7.5% polyacrylamide gel electrophoresis (Bio-Rad). Western analysis was per-formed as described (23). In separate experiments, 3 mg of protein from lysatesin the same running buffer was separated on Hiprep Sephacryl 300 (16 by 60column) with the AKTA-FPLC system. Very similar results were obtained (datanot shown).
Agarose gel electrophoresis. We used a modification of a previously describedmethod for agarose gel electrophoresis (27). Cell lysates were incubated insample buffer (Tris-borate-EDTA, 0.5% SDS, 5% glycerol, and 0.05% bromo-phenol blue) for 10 min at 42°C, and then proteins were separated by electro-phoresis in horizontal 1.5% agarose gels (20 by 25 cm) in Tris-borate-EDTAbuffer with 0.1% SDS. Proteins were electrophoretically transferred to Immo-
TABLE 1. Yeast strains
Straina Relevant genotype
779-6A MATa kar1-1 SUQ5 ade2-1 his3�202 leu2�1 trp1�63 ura3-52780-1D MAT� sup35::KanMX SSA1/pJ501 or pJ510 or pH317/NMG780-1C MAT� sup35::KanMX SSA1-21/pJ501 or pJ510 or pH317/NMGJ104x MATa hsp104::KanMX1001 MAT� SSA11013 MATa SSA1-211002 MAT� SSA1 ssa2::HIS31014 MATa SSA1-21 ssa2::HIS31011 MATa ssa1::KanMX74-D694s MATa ade1-14 his3 leu2 trp1 ura3 sup35::TRP1/pJ501 or pJ510
or pH317/NMG
a All strains except 74-D694s originated in our laboratory and are isogenic to779-6A.
290 SONG ET AL. EUKARYOT. CELL
on August 5, 2014 by guest
http://ec.asm.org/
Dow
nloaded from

bilon polyvinylidene difluoride sheets (Millipore) at 100 V for 60 min at 4°C, andWestern analysis was performed as described (23).
Luciferase reactivation assays. Luciferase reactivation assays were performedas described (24).
RESULTS
Functional Sup35-GFP fusion protein. Sup35p is a transla-tion termination factor that catalyzes release of nascent pep-tides when ribosomes encounter stop codons. In [PSI�] cells,much Sup35p is in an aggregated form, which reduces trans-lation termination efficiency and causes nonsense suppression.We monitored the presence of [PSI�] by its ability to suppressthe ade2-1 nonsense allele when the SUQ5 tRNA is present(8). Nonsuppressed ade2-1 mutants require adenine and arered due to accumulation of a substrate of Ade2p. [PSI�] re-stores adenine prototrophy and white colony color.
Sup35p can be subdivided into three regions: a dispensableamino-terminal domain (N) rich in polar amino acids that isnecessary for Sup35p prion formation, a highly charged middledomain (M) of unknown function that is also dispensable, anda C-terminal domain (C) that is necessary and sufficient forSup35p release factor function (45, 52). Fusion proteins withGFP appended to the N or N and M domains of Sup35p havebeen used widely to observe Sup35p aggregation in [PSI�] cells(3, 10, 36, 53). They are useful for diagnosing the presence orabsence of [PSI�] because fluorescence of the fusion proteinsis diffuse in [psi–] cells but coalesces into foci in [PSI�] cells,apparently because the fusions incorporate into existing Sup35paggregates through N domain interactions. Such reporters pro-vide only indirect information on the state of endogenousSup35p, however, and the foci might also represent self-aggre-gation of the fusion protein alone that is “seeded” by Sup35pprion aggregates. Moreover, these fusion proteins can induce[PSI�] to appear when expressed in [psi–] cells. This inductionlikely reflects an efficient ability of the fusions to drive aggre-gation, complicating interpretations regarding properties ofthe intact Sup35p.
Our earlier observations that SSA1-21 mutants have manyfewer transmissible [PSI�] particles or seeds per cell but onlya slight decrease in the amount of aggregated Sup35p could beexplained if more aggregated Sup35p does not function asseeds or if the seeds are larger than normal (23). In an attemptto visualize differences in Sup35p aggregation directly in vivoand avoid the complications of truncated Sup35-GFP fusions,we created a functional full-length Sup35-GFP fusion protein.The fusion protein has GFP inserted between the N and Mdomains and is designated NGMC to distinguish it fromSup35p, which we refer to as NMC (Fig. 1A). Cells lackingchromosomal SUP35 and expressing NMC or NGMC from theSUP35 promoter on a single-copy plasmid grew at similar rates(106 and 108 min/cell division, respectively; also see Fig. 1B forrate of colony formation). Therefore, NGMC performs as wellas NMC with respect to essential Sup35p function.
To obtain the NGMC strains, we replaced NMC with NGMCby a plasmid shuffle technique (see Materials and Methods).The resulting wild-type and SSA1-21 strains, which expressNGMC as the only source of Sup35p, retained typical [PSI�]phenotypes, indicating that NGMC could propagate as a prion.The letter G is added to the prion designation of NGMC cells([GPSI�]) to distinguish it from [PSI�]. [GPSI�] therefore
arose from conversion of NGMC by [PSI�] prions composedof NMC while the two proteins were both present in the samecell. Thus, the [PSI�] and [GPSI�] prions are compatible andare of the same “strain” origin. The [PSI�] and [psi–] states arenormally very stable: spontaneous appearance or mitotic lossof [PSI�] occurs very rarely. We did not quantify such eventsbut observed no difference in spontaneous appearance or lossof [PSI�] or [GPSI�] during routine handling and storage ofcultures. Thus, NGMC was as stable as NMC in both the prionand nonprion states.
Compared to [PSI�] cells, [GPSI�] cells had a very faintpink color when grown at 30°C (Fig. 1B), which is indicative ofa slight reduction in nonsense suppression. [PSI�] and [GPSI�]phenotypes were otherwise indistinguishable. [GPSI�] wassimilarly inhibited by SSA1-21, displaying comparable reduc-tions in nonsense suppression and mitotic stability (Fig. 1B).[PSI�] and [GPSI�] also were the same regarding curability bydeletion and overexpression of Hsp104 or by including milli-molar amounts of guanidine hydrochloride in the growth me-diu, which inactivates Hsp104 (data not shown) (5, 15, 24,46). Finally, [PSI�] and [GPSI�] could both be transmittedcytoplasmically (by cytoduction, see above) with equal effi-ciency to both NGMC and NMC strains (data not shown).These results demonstrate that the GFP insertion did not sig-nificantly affect prion interactions of the N domains of NGMC
FIG. 1. Sup35-GFP fusion protein functions normally. (A) (Top)NMC: Sup35p domains N, codons 1 to 123; M, codons 124 to 253; andC, codons 254 to 685 are indicated. (Bottom) NGMC: the Sup35-GFPfusion has GFP inserted in-frame between codons 123 and 124. (B)SSA1-21 (top) and SSA1 (bottom) cells expressing NMC (left) orNGMC (right) in place of endogenous Sup35p were grown on 1/2YPDfor 2 days at 30°C followed by 3 days at 22°C. For NMC and NGMC,the rate of colony formation and [PSI�] phenotype, including impair-ment of [PSI�] by SSA1-21, are essentially indistinguishable.
VOL. 4, 2005 Hsp70 ROLE IN YEAST PRION SEED REPLICATION 291
on August 5, 2014 by guest
http://ec.asm.org/
Dow
nloaded from

or of those between NGMC and NMC. Strains discussed here-after, unless indicated otherwise, express NGMC in place ofNMC.
NGMC forms fluorescent foci only in [GPSI�] cells of agingcultures. GFP fluorescence in [Gpsi–] wild-type and SSA1-21cells was diffuse throughout the cytoplasm in both log-phaseand stationary-phase cultures (Fig. 2). Unexpectedly, for bothwild-type and SSA1-21 strains, GFP fluorescence in log-phase[GPSI�] cells was uniformly diffuse throughout the cytoplasm,similar to that of [Gpsi–] cells. Thus, aggregation of NGMCinto particles of a size detectable as fluorescent foci was notrequired for propagation of [GPSI�], suggesting that [PSI�]similarly does not require such aggregation of NMC.
Numerous fluorescent foci appeared throughout the cyto-plasm of both wild-type and SSA1-21 cells from aging [GPSI�]cultures (Fig. 2). In focusing through the cells, we estimatedthe number of foci, which streamed throughout the cytoplasm,to be several hundred to a thousand. There was no consistentdifference in the size or number of aggregates between age-matched wild-type and SSA1-21 cells. No such foci appeared in[Gpsi–] cells in cultures of any age. Thus, foci were restricted to[GPSI�] cells and appeared only in stationary-phase cells.
The diffuse fluorescence in [GPSI�] log-phase cells was un-expected because other widely used GFP fusions, which lackthe Sup35p C terminus, show clear focus formation after ex-pressing the fusions for a relatively short time. Since our GFPfusion and strain background both differed from those usedpreviously to visualize Sup35p in vivo, we tested both our GFPfusion in another strain background, and the commonly used
NM-GFP fusion in our strains. When NMC was replaced withNGMC in another strain (74-D694) commonly used to moni-tor [PSI�], we again saw the appearance of [GPSI�] foci onlyin stationary-phase cells (Fig. 3A). Additionally, in our wild-type and SSA1-21 log-phase [PSI�] cells, fluorescent foci wereclearly present after expressing NM-GFP for only 2 h (Fig. 3B).NM-GFP did not form foci in [psi–] variants of the same strains
FIG. 2. NGMC fluorescence in SSA1 and SSA1-21 cells forms focionly in aging [GPSI�] cultures. Cells expressing NGMC in place ofendogenous Sup35p were grown on YPAD plates at 30° for 1, 2, and4 days (1d, 2d, and 4d, respectively). One-day cells are in log-phase,while most 2-day and 4-day cells are in or approaching stationaryphase. NGMC fluorescence in [GPSI�] cells from 1-day colonies ofboth strains is indistinguishable from that in [Gpsi–] cells. Foci ofNGMC fluorescence are apparent only in [GPSI�] cells from oldercolonies of the same cultures.
FIG. 3. NGMC and NM-GFP fluorescence patterns are indepen-dent of strain background. (A) NGMC fluorescence was assayed as inFig. 2 in an unrelated strain (74-D694) commonly used to monitorSup35NM-GFP aggregation. As in Fig. 2, fluorescence in log-phasecultures of [GPSI�] cells is similar to that of [Gpsi–] cells, and foci ap-pear only in [GPSI�] cells from older colonies of the same cultures. (B)Fluorescence in log-phase cells 2 h after galactose-induced expressionof Sup35NM-GFP. Fluorescence remains diffuse in [psi–] cells (top),while foci appear in both wild-type and SSA1-21 [PSI�] cells (bottom).
292 SONG ET AL. EUKARYOT. CELL
on August 5, 2014 by guest
http://ec.asm.org/
Dow
nloaded from
under the same conditions. Therefore, focus formation or lackthereof was a characteristic of the GFP fusion rather than thestrain background.
Fluorescent foci do not directly represent [GPSI�] seeds. Instationary-phase cells, the number and appearance of NGMCaggregates that have attained a size visible by GFP fluores-cence were very similar in wild-type and SSA1-21 cells. SinceSSA1-21 mutants have fewer seeds per cell, we inferred thatthese foci do not represent prion seeds. To test this hypothesis,and to determine if stationary-phase cells had different num-bers of seeds than log-phase cells, we used a recently describedtechnique (7) based on guanidine inactivation of Hsp104 (15,24) to estimate seed number per cell. Log-phase and station-ary-phase cells, grown like those used for the fluorescenceimaging, were spread onto rich medium containing 3 mM gua-nidine and allowed to form colonies. Inhibition of Hsp104 byguanidine blocks replication of [PSI�] seeds (33). As a cellgrows into a colony, its limited number of seeds become dis-tributed to only a portion of its progeny in the colony. Thenumber of [PSI�] cells in the resulting colony, determined byplating the cells of the colony onto medium selecting for[PSI�], provides an estimate of the number of [PSI�] seedsthat were present in the cell that began the colony.
In agreement with our earlier estimates of a 10-fold differ-ence (23), we found about 80 seeds/cell for wild-type cells andabout 10/cell for SSA1-21 cells (Table 2). These seed numbersare much lower than the number of fluorescent foci. Moreover,this difference does not correlate with a difference in the num-ber of foci between wild-type and SSA1-21 cells. Together withthe observation that [GPSI�] seeds propagate in the absenceof foci in log-phase cells, these data suggest that the foci do notdirectly represent [GPSI�] seeds. Although these results donot rule out the possibility that aggregates seen as foci havesome ability to act as [GPSI�] seeds, they imply that suchability is limited. On average, the seed numbers of stationary-phase cells were lower than those of log-phase cells for bothwild-type and SSA1-21 strains. Apparently, in stationary-phase[GPSI�] cells, aggregation of Sup35p continues but seed rep-lication does not.
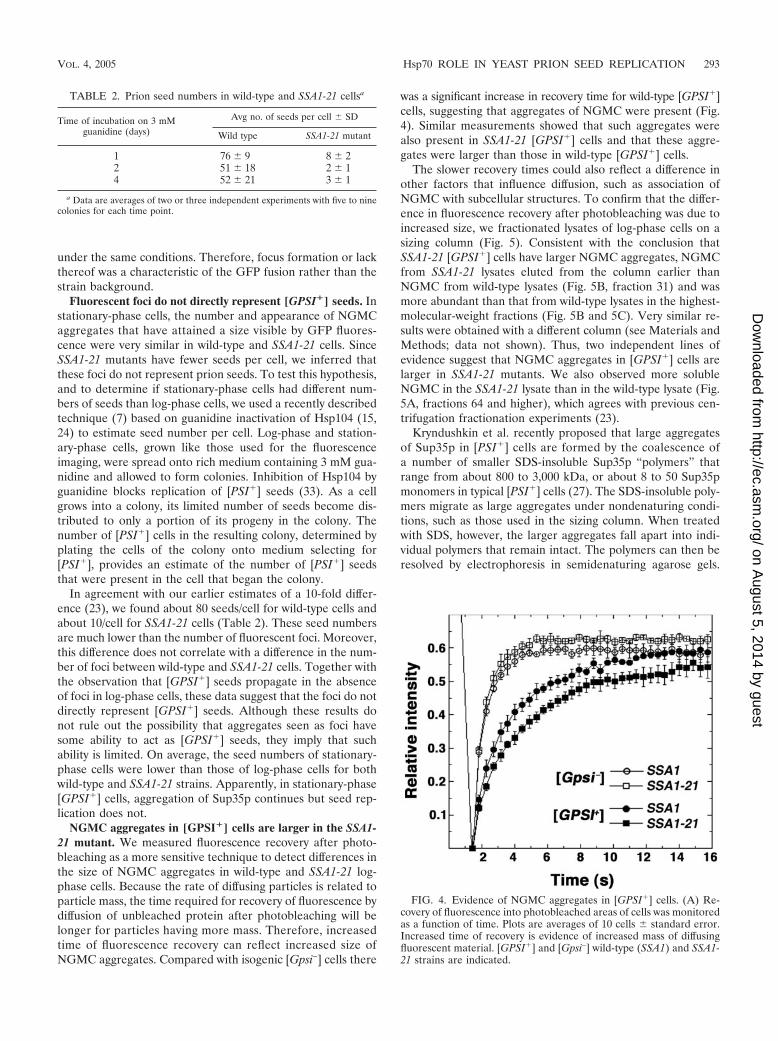
NGMC aggregates in [GPSI�] cells are larger in the SSA1-21 mutant. We measured fluorescence recovery after photo-bleaching as a more sensitive technique to detect differences inthe size of NGMC aggregates in wild-type and SSA1-21 log-phase cells. Because the rate of diffusing particles is related toparticle mass, the time required for recovery of fluorescence bydiffusion of unbleached protein after photobleaching will belonger for particles having more mass. Therefore, increasedtime of fluorescence recovery can reflect increased size ofNGMC aggregates. Compared with isogenic [Gpsi–] cells there
was a significant increase in recovery time for wild-type [GPSI�]cells, suggesting that aggregates of NGMC were present (Fig.4). Similar measurements showed that such aggregates werealso present in SSA1-21 [GPSI�] cells and that these aggre-gates were larger than those in wild-type [GPSI�] cells.
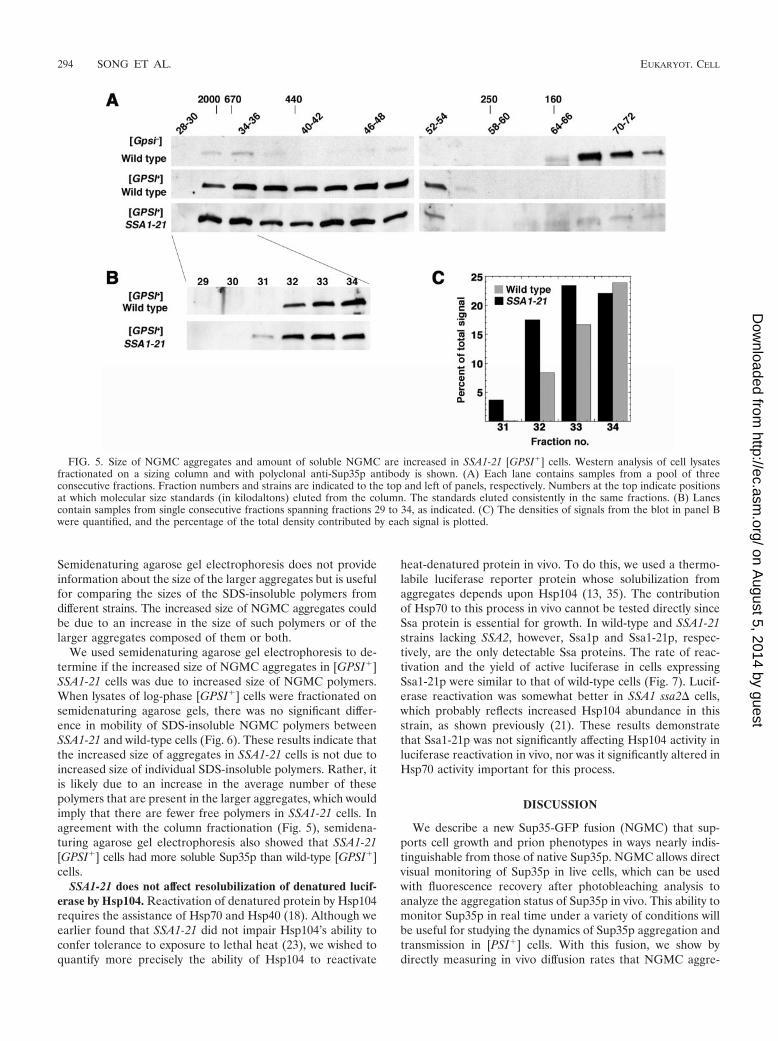
The slower recovery times could also reflect a difference inother factors that influence diffusion, such as association ofNGMC with subcellular structures. To confirm that the differ-ence in fluorescence recovery after photobleaching was due toincreased size, we fractionated lysates of log-phase cells on asizing column (Fig. 5). Consistent with the conclusion thatSSA1-21 [GPSI�] cells have larger NGMC aggregates, NGMCfrom SSA1-21 lysates eluted from the column earlier thanNGMC from wild-type lysates (Fig. 5B, fraction 31) and wasmore abundant than that from wild-type lysates in the highest-molecular-weight fractions (Fig. 5B and 5C). Very similar re-sults were obtained with a different column (see Materials andMethods; data not shown). Thus, two independent lines ofevidence suggest that NGMC aggregates in [GPSI�] cells arelarger in SSA1-21 mutants. We also observed more solubleNGMC in the SSA1-21 lysate than in the wild-type lysate (Fig.5A, fractions 64 and higher), which agrees with previous cen-trifugation fractionation experiments (23).
Kryndushkin et al. recently proposed that large aggregatesof Sup35p in [PSI�] cells are formed by the coalescence ofa number of smaller SDS-insoluble Sup35p “polymers” thatrange from about 800 to 3,000 kDa, or about 8 to 50 Sup35pmonomers in typical [PSI�] cells (27). The SDS-insoluble poly-mers migrate as large aggregates under nondenaturing condi-tions, such as those used in the sizing column. When treatedwith SDS, however, the larger aggregates fall apart into indi-vidual polymers that remain intact. The polymers can then beresolved by electrophoresis in semidenaturing agarose gels.
FIG. 4. Evidence of NGMC aggregates in [GPSI�] cells. (A) Re-covery of fluorescence into photobleached areas of cells was monitoredas a function of time. Plots are averages of 10 cells � standard error.Increased time of recovery is evidence of increased mass of diffusingfluorescent material. [GPSI�] and [Gpsi–] wild-type (SSA1) and SSA1-21 strains are indicated.
TABLE 2. Prion seed numbers in wild-type and SSA1-21 cellsa
Time of incubation on 3 mMguanidine (days)
Avg no. of seeds per cell � SD
Wild type SSA1-21 mutant
1 76 � 9 8 � 22 51 � 18 2 � 14 52 � 21 3 � 1
a Data are averages of two or three independent experiments with five to ninecolonies for each time point.
VOL. 4, 2005 Hsp70 ROLE IN YEAST PRION SEED REPLICATION 293
on August 5, 2014 by guest
http://ec.asm.org/
Dow
nloaded from
Semidenaturing agarose gel electrophoresis does not provideinformation about the size of the larger aggregates but is usefulfor comparing the sizes of the SDS-insoluble polymers fromdifferent strains. The increased size of NGMC aggregates couldbe due to an increase in the size of such polymers or of thelarger aggregates composed of them or both.
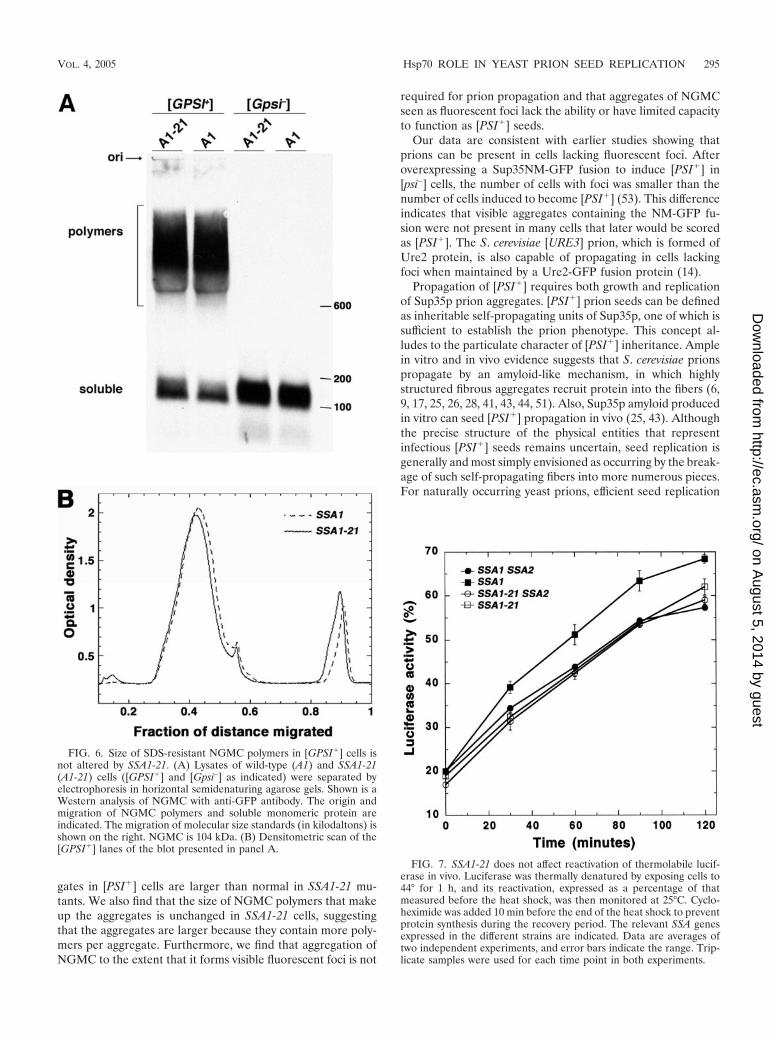
We used semidenaturing agarose gel electrophoresis to de-termine if the increased size of NGMC aggregates in [GPSI�]SSA1-21 cells was due to increased size of NGMC polymers.When lysates of log-phase [GPSI�] cells were fractionated onsemidenaturing agarose gels, there was no significant differ-ence in mobility of SDS-insoluble NGMC polymers betweenSSA1-21 and wild-type cells (Fig. 6). These results indicate thatthe increased size of aggregates in SSA1-21 cells is not due toincreased size of individual SDS-insoluble polymers. Rather, itis likely due to an increase in the average number of thesepolymers that are present in the larger aggregates, which wouldimply that there are fewer free polymers in SSA1-21 cells. Inagreement with the column fractionation (Fig. 5), semidena-turing agarose gel electrophoresis also showed that SSA1-21[GPSI�] cells had more soluble Sup35p than wild-type [GPSI�]cells.
SSA1-21 does not affect resolubilization of denatured lucif-erase by Hsp104. Reactivation of denatured protein by Hsp104requires the assistance of Hsp70 and Hsp40 (18). Although weearlier found that SSA1-21 did not impair Hsp104’s ability toconfer tolerance to exposure to lethal heat (23), we wished toquantify more precisely the ability of Hsp104 to reactivate
heat-denatured protein in vivo. To do this, we used a thermo-labile luciferase reporter protein whose solubilization fromaggregates depends upon Hsp104 (13, 35). The contributionof Hsp70 to this process in vivo cannot be tested directly sinceSsa protein is essential for growth. In wild-type and SSA1-21strains lacking SSA2, however, Ssa1p and Ssa1-21p, respec-tively, are the only detectable Ssa proteins. The rate of reac-tivation and the yield of active luciferase in cells expressingSsa1-21p were similar to that of wild-type cells (Fig. 7). Lucif-erase reactivation was somewhat better in SSA1 ssa2� cells,which probably reflects increased Hsp104 abundance in thisstrain, as shown previously (21). These results demonstratethat Ssa1-21p was not significantly affecting Hsp104 activity inluciferase reactivation in vivo, nor was it significantly altered inHsp70 activity important for this process.
DISCUSSION
We describe a new Sup35-GFP fusion (NGMC) that sup-ports cell growth and prion phenotypes in ways nearly indis-tinguishable from those of native Sup35p. NGMC allows directvisual monitoring of Sup35p in live cells, which can be usedwith fluorescence recovery after photobleaching analysis toanalyze the aggregation status of Sup35p in vivo. This ability tomonitor Sup35p in real time under a variety of conditions willbe useful for studying the dynamics of Sup35p aggregation andtransmission in [PSI�] cells. With this fusion, we show bydirectly measuring in vivo diffusion rates that NGMC aggre-
FIG. 5. Size of NGMC aggregates and amount of soluble NGMC are increased in SSA1-21 [GPSI�] cells. Western analysis of cell lysatesfractionated on a sizing column and with polyclonal anti-Sup35p antibody is shown. (A) Each lane contains samples from a pool of threeconsecutive fractions. Fraction numbers and strains are indicated to the top and left of panels, respectively. Numbers at the top indicate positionsat which molecular size standards (in kilodaltons) eluted from the column. The standards eluted consistently in the same fractions. (B) Lanescontain samples from single consecutive fractions spanning fractions 29 to 34, as indicated. (C) The densities of signals from the blot in panel Bwere quantified, and the percentage of the total density contributed by each signal is plotted.
294 SONG ET AL. EUKARYOT. CELL
on August 5, 2014 by guest
http://ec.asm.org/
Dow
nloaded from
gates in [PSI�] cells are larger than normal in SSA1-21 mu-tants. We also find that the size of NGMC polymers that makeup the aggregates is unchanged in SSA1-21 cells, suggestingthat the aggregates are larger because they contain more poly-mers per aggregate. Furthermore, we find that aggregation ofNGMC to the extent that it forms visible fluorescent foci is not
required for prion propagation and that aggregates of NGMCseen as fluorescent foci lack the ability or have limited capacityto function as [PSI�] seeds.
Our data are consistent with earlier studies showing thatprions can be present in cells lacking fluorescent foci. Afteroverexpressing a Sup35NM-GFP fusion to induce [PSI�] in[psi–] cells, the number of cells with foci was smaller than thenumber of cells induced to become [PSI�] (53). This differenceindicates that visible aggregates containing the NM-GFP fu-sion were not present in many cells that later would be scoredas [PSI�]. The S. cerevisiae [URE3] prion, which is formed ofUre2 protein, is also capable of propagating in cells lackingfoci when maintained by a Ure2-GFP fusion protein (14).
Propagation of [PSI�] requires both growth and replicationof Sup35p prion aggregates. [PSI�] prion seeds can be definedas inheritable self-propagating units of Sup35p, one of which issufficient to establish the prion phenotype. This concept al-ludes to the particulate character of [PSI�] inheritance. Amplein vitro and in vivo evidence suggests that S. cerevisiae prionspropagate by an amyloid-like mechanism, in which highlystructured fibrous aggregates recruit protein into the fibers (6,9, 17, 25, 26, 28, 41, 43, 44, 51). Also, Sup35p amyloid producedin vitro can seed [PSI�] propagation in vivo (25, 43). Althoughthe precise structure of the physical entities that representinfectious [PSI�] seeds remains uncertain, seed replication isgenerally and most simply envisioned as occurring by the break-age of such self-propagating fibers into more numerous pieces.For naturally occurring yeast prions, efficient seed replication
FIG. 6. Size of SDS-resistant NGMC polymers in [GPSI�] cells isnot altered by SSA1-21. (A) Lysates of wild-type (A1) and SSA1-21(A1-21) cells ([GPSI�] and [Gpsi–] as indicated) were separated byelectrophoresis in horizontal semidenaturing agarose gels. Shown is aWestern analysis of NGMC with anti-GFP antibody. The origin andmigration of NGMC polymers and soluble monomeric protein areindicated. The migration of molecular size standards (in kilodaltons) isshown on the right. NGMC is 104 kDa. (B) Densitometric scan of the[GPSI�] lanes of the blot presented in panel A.
FIG. 7. SSA1-21 does not affect reactivation of thermolabile lucif-erase in vivo. Luciferase was thermally denatured by exposing cells to44° for 1 h, and its reactivation, expressed as a percentage of thatmeasured before the heat shock, was then monitored at 25°C. Cyclo-heximide was added 10 min before the end of the heat shock to preventprotein synthesis during the recovery period. The relevant SSA genesexpressed in the different strains are indicated. Data are averages oftwo independent experiments, and error bars indicate the range. Trip-licate samples were used for each time point in both experiments.
VOL. 4, 2005 Hsp70 ROLE IN YEAST PRION SEED REPLICATION 295
on August 5, 2014 by guest
http://ec.asm.org/
Dow
nloaded from

requires the disaggregating activity of Hsp104 (5, 11, 31, 37–39).
Kryndushkin et al. showed that aggregated Sup35p in [PSI�]cells is found as individual SDS-insoluble polymers and aslarge aggregates formed by the coalescence of a number ofsuch polymers (27). They propose that Hsp104 generates new[PSI�] seeds by severing the polymers and by dismantlingthe large Sup35p aggregates into individual polymers, both ofwhich increase the number of free polymers. Additionally, itwas suggested that Sup35p polymers within the aggregatescould be severed without generating new seeds if the resultingpolymer fragments remained associated with the aggregate.The implications of this work are that the Sup35p polymers actefficiently as [PSI�] seeds and that higher-order aggregatescomposed of them, while less efficient, can act as [PSI�] seedsby being a source of Sup35p polymers that are liberated by theaction of cellular chaperones. Although this study providesvery useful insight into the physical basis of [PSI�] seeds, itshould be emphasized that the precise properties of seeds isuncertain, as they are quantified genetically as the transmissi-ble entities that perpetuate the [PSI�] state.
The interpretation that individual Sup35p polymers act mostefficiently as [PSI�] seeds also provides a simple explanationfor our observations. Our conclusion that NGMC aggregatesare larger in SSA1-21 cells because there are more NGMCpolymers per aggregate would explain the concomitant de-crease in [GPSI�] seeds. If a larger proportion of NGMCpolymers were associated with large aggregates, then therewould be fewer free NGMC polymers. Moreover, since the ef-ficiency with which Sup35p is recruited into prion aggregatesdepends upon the number of seeds (33), the increase inSup35p solubility in SSA1-21 cells can be explained by thereduced number of free polymers capable of recruiting andtherefore depleting Sup35p. A similar conclusion relating freepolymers to Sup35p solubility has been reported by others (1,27).
An explanation for the effects of SSA1-21 is that Ssa1p(Hsp70) normally acts in [PSI�] seed propagation by disman-tling polymers from Sup35p aggregates. Our earlier workshowed that Ssa1-21p has enhanced substrate binding com-pared with Ssa1p (20, 21). By binding more avidly to Sup35paggregates, Ssa1-21p might be impaired in an Hsp70 activitythat promotes the disassembly of aggregates into polymers,either directly or through interaction with Hsp104. Althoughan inhibitory effect on Hsp104 would be expected to causean increase in size of Sup35p aggregates, those that inhibitHsp104 alone, such as guanidine inactivation, increase the sizeof Sup35p polymers (22, 27). The effect of Ssa1-21p on [PSI�]could be due to altered interaction of Ssa1-21p with Hsp104 inthe dismantling process or to steric hindrance of Hsp104 accessto Sup35p aggregates by increased amounts of bound Ssa1-21p. Any of these direct or indirect effects would also explainthe ability of Ssa1-21p to impair [PSI�] in the presence of thefunctionally redundant Ssa2p. Since cells expressing Ssa1-21pas the only Ssa protein have no apparent defect in thermotol-erance or reactivation of denatured luciferase in vivo, the pro-cess of breaking highly ordered Sup35p aggregates to generatenew prion seeds should be different than disaggregation ofamorphous aggregates of thermally denatured protein. Alter-natively, replication of prion aggregates might be much more
sensitive to disruptions of Hsp70 or Hsp104 machinery func-tion.
An alternative explanation also consistent with the dominanteffects of Ssa1-21p is that it is hyperactive in an Hsp70 functionthat promotes aggregation of the Sup35p polymers into higher-order aggregates. Work showing that excess wild-type Ssa1causes fluorescent Sup35NM-GFP aggregates in [PSI�] cells toincrease in size led to the proposal that Ssa1p helps assembleSup35p into larger aggregates that are less sensitive to the dis-aggregating activity of Hsp104 (3). In contrast to Ssa1-21p, how-ever, excess Ssa1p increases nonsense suppression and doesnot cause [PSI�] instability (34). Moreover, unlike Ssa1-21,excess Ssa1p increases the size of Sup35p polymers in [PSI�]cells (1). Although Ssa1p might help Sup35p polymerize invivo, the effects of Ssa1-21p on [PSI�] are not consistent withits being more efficient at a wild-type Hsp70 function.
Fluorescent foci in [PSI�] cells expressing Sup35NM-GFPfusions are interpreted as reflecting the extent of aggregationof endogenous Sup35p. Depletion of Hsp104 causes such focito increase in size while decreasing in number (48). Similarly,foci in cells harboring variants of [PSI�] that appear partiallyresistant to Hsp104 are larger and fewer (3). These differencescorrelate with reduced [PSI�] stability, which is consistent withthe notion that Hsp104 acts in [PSI�] propagation by breakingSup35p aggregates. Furthermore, large visible aggregates ofGFP fusion of both Sup35-NM and Ure2p (the determinant ofthe yeast [URE3] prion) have been suggested to be dead-endproducts of prion propagation that lack or have limited seedingcapacity (3, 38, 48).
The appearance of fluorescent NGMC foci in cells of agingcultures shows that a functional Sup35-GFP fusion protein alsocan aggregate to the degree that it forms visible fluorescentfoci, but our data also show that [GPSI�] propagation does notrequire aggregation of NGMC to such an extent. Therefore,fluorescent foci do not necessarily represent [GPSI�] seeds. Infact, wild-type and SSA1-21 cells had similar numbers of focibut different numbers of seeds. Moreover, the large number offoci in stationary-phase cells does not correlate with the smallnumber of seeds estimated to be present in these cells. Thus,most if not all of the NGMC aggregates represented by focilack seeding capacity.
Our data showing that there are many fewer [GPSI�] seedsthan foci are consistent with the conclusions drawn by Kryn-dushkin et al., who proposed that foci seen in cells expressingSup35-GFP fusions represent the largest aggregates and thatthe majority of seeds should be below detection by GFP fluo-rescence (27). Moreover, since detection of aggregates by flu-orescence recovery after photobleaching in the diffuse fluores-cence of log-phase cells implies that most of the fluorescentmaterial is in an aggregated form, the number of aggregatesin these cells must be much higher than the number of fociseen in stationary-phase cells. Thus, most Sup35p aggregates in[PSI�] cells do not function as prion seeds. Alternatively, cur-rent methods for determining [PSI�] seed numbers per cellsignificantly underestimate such numbers.
REFERENCES
1. Allen, K. D., R. D. Wegrzyn, T. A. Chernova, S. Muller, G. P. Newnam, P. A.Winslett, K. B. Wittich, K. D. Wilkinson, and Y. O. Chernoff.. Hsp70 chap-erones as modulators of prion life cycle: novel effects of Ssa and Ssb on theSaccharomyces cerevisiae prion [PSI�]. Genetics, in press.
296 SONG ET AL. EUKARYOT. CELL
on August 5, 2014 by guest
http://ec.asm.org/
Dow
nloaded from

2. Baudin, A., O. Ozier-Kalogeropoulos, A. Denouel, F. Lacroute, and C. Cul-lin. 1993. A simple and efficient method for direct gene deletion in Saccha-romyces cerevisiae. Nucleic Acids Res. 21:3329–3330.
3. Borchsenius, A. S., R. D. Wegrzyn, G. P. Newnam, S. G. Inge-Vechtomov,and Y. O. Chernoff. 2001. Yeast prion protein derivative defective in aggre-gate shearing and production of new ‘seeds’. EMBO J. 20:6683–6691.
4. Bukau, B., and A. L. Horwich. 1998. The Hsp70 and Hsp60 chaperonemachines. Cell 92:351–366.
5. Chernoff, Y. O., S. L. Lindquist, B. Ono, S. G. Inge-Vechtomov, and S. W.Liebman. 1995. Role of the chaperone protein Hsp104 in propagation of theyeast prion-like factor [psi�]. Science 268:880–884.
6. Collins, S. R., A. Douglass, R. D. Vale, and J. S. Weissman. 2004. Mechanismof prion propagation: amyloid growth occurs by monomer addition. PLoSBiol. 2:e321.
7. Cox, B., F. Ness, and M. Tuite. 2003. Analysis of the generation and segre-gation of propagons: entities that propagate the [PSI�] prion in yeast.Genetics 165:23–33.
8. Cox, B. S. 1965. “�” a cytoplasmic suppressor of super-suppressor in yeast.Heredity 20:505–521.
9. Crist, C. G., T. Nakayashiki, H. Kurahashi, and Y. Nakamura. 2003. [PHI�],a novel Sup35-prion variant propagated with non-Gln/Asn oligopeptide re-peats in the absence of the chaperone protein Hsp104. Genes Cells 8:603–618.
10. DePace, A. H., A. Santoso, P. Hillner, and J. S. Weissman. 1998. A criticalrole for amino-terminal glutamine/asparagine repeats in the formation andpropagation of a yeast prion. Cell 93:1241–1252.
11. Derkatch, I. L., M. E. Bradley, P. Zhou, Y. O. Chernoff, and S. W. Liebman.1997. Genetic and environmental factors affecting the de novo appearance ofthe [PSI�] prion in Saccharomyces cerevisiae. Genetics 147:507–519.
12. Edskes, H. K., and R. B. Wickner. 2000. A protein required for priongeneration: [URE3] induction requires the Ras-regulated Mks1 protein.Proc. Natl. Acad. Sci. USA 97:6625–6629.
13. Escher, A., D. J. O’Kane, J. Lee, and A. A. Szalay. 1989. Bacterial luciferasealpha beta fusion protein is fully active as a monomer and highly sensitive invivo to elevated temperature. Proc. Natl. Acad. Sci. USA 86:6528–6532.
14. Fernandez-Bellot, E., E. Guillemet, F. Ness, A. Baudin-Baillieu, L. Ripaud,M. Tuite, and C. Cullin. 2002. The [URE3] phenotype: evidence for asoluble prion in yeast. EMBO Rep. 3:76–81.
15. Ferreira, P. C., F. Ness, S. R. Edwards, B. S. Cox, and M. F. Tuite. 2001. Theelimination of the yeast [PSI�] prion by guanidine hydrochloride is the resultof Hsp104 inactivation. Mol. Microbiol. 40:1357–1369.
16. Gao, B. C., J. Biosca, E. A. Craig, L. E. Greene, and E. Eisenberg. 1991.Uncoating of coated vesicles by yeast hsp70 proteins. J. Biol. Chem. 266:19565–19571.
17. Glover, J. R., A. S. Kowal, E. C. Schirmer, M. M. Patino, J. J. Liu, and S.Lindquist. 1997. Self-seeded fibers formed by Sup35, the protein determi-nant of [PSI�], a heritable prion-like factor of S. cerevisiae. Cell 89:811–819.
18. Glover, J. R., and S. Lindquist. 1998. Hsp104, Hsp70, and Hsp40: a novelchaperone system that rescues previously aggregated proteins. Cell 94:73–82.
19. Hara, H., T. Nakayashiki, C. G. Crist, and Y. Nakamura. 2003. Prion domaininteraction responsible for species discrimination in yeast [PSI�] transmis-sion. Genes Cells 8:925–939.
20. Jones, G. W., and D. C. Masison. 2003. Saccharomyces cerevisiae Hsp70mutations affect [PSI�] prion propagation and cell growth differently andimplicate Hsp40 and tetratricopeptide repeat cochaperones in impairment of[PSI�]. Genetics 163:495–506.
21. Jones, G. W., Y. Song, S. Chung, and D. C. Masison. 2004. Propagation ofyeast [PSI�] prion is impaired by factors that regulate Hsp70 substratebinding. Mol. Cell. Biol. 24:3928–3937.
22. Jung, G., G. Jones, and D. C. Masison. 2002. Amino acid residue 184 of yeastHsp104 chaperone is critical for prion-curing by guanidine, prion propaga-tion, and thermotolerance. Proc. Natl. Acad. Sci. USA 99:9936–9941.
23. Jung, G., G. Jones, R. D. Wegrzyn, and D. C. Masison. 2000. A role forcytosolic hsp70 in yeast [PSI�] prion propagation and [PSI�] as a cellularstress. Genetics 156:559–570.
24. Jung, G., and D. C. Masison. 2001. Guanidine hydrochloride inhibits Hsp104activity in vivo: a possible explanation for its effect in curing yeast prions.Curr. Microbiol. 43:7–10.
25. King, C. Y., and R. Diaz-Avalos. 2004. Protein-only transmission of threeyeast prion strains. Nature 428:319–323.
26. King, C. Y., P. Tittmann, H. Gross, R. Gebert, M. Aebi, and K. Wuthrich.1997. Prion-inducing domain 2–114 of yeast Sup35 protein transforms in vitrointo amyloid-like filaments. Proc. Natl. Acad. Sci. USA 94:6618–6622.
27. Kryndushkin, D. S., I. M. Alexandrov, M. D. Ter-Avanesyan, and V. V.Kushnirov. 2003. Yeast [PSI�] prion aggregates are formed by small Sup35polymers fragmented by Hsp104. J. Biol. Chem. 278:49636–49643.
28. Kushnirov, V. V., and M. D. Ter-Avanesyan. 1998. Structure and replicationof yeast prions. Cell 94:13–16.
29. Liu, J. J., and S. Lindquist. 1999. Oligopeptide-repeat expansions modulate‘protein-only’ inheritance in yeast. Nature 400:573–576.
30. Longtine, M. S., A. McKenzie, 3rd, D. J. Demarini, N. G. Shah, A. Wach, A.Brachat, P. Philippsen, and J. R. Pringle. 1998. Additional modules forversatile and economical PCR-based gene deletion and modification in Sac-charomyces cerevisiae. Yeast 14:953–961.
31. Moriyama, H., H. K. Edskes, and R. B. Wickner. 2000. [URE3] prion prop-agation in Saccharomyces cerevisiae: requirement for chaperone Hsp104 andcuring by overexpressed chaperone Ydj1p. Mol. Cell. Biol. 20:8916–8922.
32. Nelson, R. J., T. Ziegelhoffer, C. Nicolet, M. Werner-Washburne, and E. A.Craig. 1992. The translation machinery and 70 kd heat shock protein coop-erate in protein synthesis. Cell 71:97–105.
33. Ness, F., P. Ferreira, B. S. Cox, and M. F. Tuite. 2002. Guanidine hydro-chloride inhibits the generation of prion “seeds” but not prion proteinaggregation in yeast. Mol. Cell. Biol. 22:5593–5605.
34. Newnam, G. P., R. D. Wegrzyn, S. L. Lindquist, and Y. O. Chernoff. 1999.Antagonistic interactions between yeast chaperones Hsp104 and Hsp70 inprion curing. Mol. Cell. Biol. 19:1325–1333.
35. Parsell, D. A., A. S. Kowal, M. A. Singer, and S. Lindquist. 1994. Proteindisaggregation mediated by heat-shock protein Hsp104. Nature 372:475–478.
36. Patino, M. M., J. J. Liu, J. R. Glover, and S. Lindquist. 1996. Support for theprion hypothesis for inheritance of a phenotypic trait in yeast. Science 273:622–626.
37. Paushkin, S. V., V. V. Kushnirov, V. N. Smirnov, and M. D. Ter-Avanesyan.1996. Propagation of the yeast prion-like [psi�] determinant is mediated byoligomerization of the SUP35-encoded polypeptide chain release factor.EMBO J. 15:3127–3134.
38. Ripaud, L., L. Maillet, and C. Cullin. 2003. The mechanisms of [URE3]prion elimination demonstrate that large aggregates of Ure2p are dead-endproducts. EMBO J. 22:5251–5259.
39. Shorter, J., and S. Lindquist. 2004. Hsp104 catalyzes formation and elimi-nation of self-replicating Sup35 Prion conformers. Science 304:1793–1797.
40. Sikorski, R. S., and P. Hieter. 1989. A system of shuttle vectors and yeasthost strains designed for efficient manipulation of DNA in Saccharomycescerevisiae. Genetics 122:19–27.
41. Speransky, V. V., K. L. Taylor, H. K. Edskes, R. B. Wickner, and A. C.Steven. 2001. Prion filament networks in [URE3] cells of Saccharomycescerevisiae. J. Cell Biol. 153:1327–1336.
42. Stansfield, I., K. M. Jones, V. V. Kushnirov, A. R. Dagkesamanskaya, A. I.Poznyakovski, S. V. Paushkin, C. R. Nierras, B. S. Cox, M. D. Ter-Avane-syan, and M. F. Tuite. 1995. The products of the SUP45 (eRF1) and SUP35genes interact to mediate translation termination in Saccharomyces cerevi-siae. EMBO J. 14:4365–4373.
43. Tanaka, M., P. Chien, N. Naber, R. Cooke, and J. S. Weissman. 2004.Conformational variations in an infectious protein determine prion straindifferences. Nature 428:323–328.
44. Taylor, K. L., N. Cheng, R. W. Williams, A. C. Steven, and R. B. Wickner.1999. Prion domain initiation of amyloid formation in vitro from nativeUre2p. Science 283:1339–1343.
45. Ter-Avanesyan, M. D., V. V. Kushnirov, A. R. Dagkesamanskaya, S. A.Didichenko, Y. O. Chernoff, S. G. Inge-Vechtomov, and V. N. Smirnov. 1993.Deletion analysis of the SUP35 gene of the yeast Saccharomyces cerevisiaereveals two non-overlapping functional regions in the encoded protein. Mol.Microbiol. 7:683–692.
46. Tuite, M. F., C. R. Mundy, and B. S. Cox. 1981. Agents that cause a highfrequency of genetic change from [psi�] to [psi-] in Saccharomyces cerevisiae.Genetics 98:691–711.
47. Wach, A., A. Brachat, R. Pohlmann, and P. Philippsen. 1994. New heterol-ogous modules for classical or PCR-based gene disruptions in Saccharomycescerevisiae. Yeast 10:1793–1808.
48. Wegrzyn, R. D., K. Bapat, G. P. Newnam, A. D. Zink, and Y. O. Chernoff.2001. Mechanism of prion loss after Hsp104 inactivation in yeast. Mol. Cell.Biol. 21:4656–4669.
49. Werner-Washburne, M., D. E. Stone, and E. A. Craig. 1987. Complex inter-actions among members of an essential subfamily of hsp70 genes in Saccha-romyces cerevisiae. Mol. Cell. Biol. 7:2568–2577.
50. Wickner, R. B. 1994. Evidence for a prion analog in S. cerevisiae: the [URE3]non-Mendelian genetic element as an altered URE2 protein. Science 264:566–569.
51. Wickner, R. B., K. L. Taylor, H. K. Edskes, M. L. Maddelein, H. Moriyama,and B. T. Roberts. 2000. Prions of yeast as heritable amyloidoses. J. Struct.Biol. 130:310–322.
52. Wilson, P. G., and M. R. Culbertson. 1988. SUF12 suppressor protein ofyeast. A fusion protein related to the EF-1 family of elongation factors. J.Mol. Biol. 199:559–573.
53. Zhou, P., I. L. Derkatch, and S. W. Liebman. 2001. The relationship betweenvisible intracellular aggregates that appear after overexpression of Sup35 andthe yeast prion-like elements [PSI�] and [PIN�]. Mol. Microbiol. 39:37–46.
54. Zhouravleva, G., L. Frolova, X. Le Goff, R. Le Guellec, S. Inge-Vechtomov,L. Kisselev, and M. Philippe. 1995. Termination of translation in eukaryotesis governed by two interacting polypeptide chain release factors, eRF1 andeRF3. EMBO J. 14:4065–4072.
VOL. 4, 2005 Hsp70 ROLE IN YEAST PRION SEED REPLICATION 297
on August 5, 2014 by guest
http://ec.asm.org/
Dow
nloaded from
Copyright © 2022 FDOKUMEN




















