
Antidiarrhoeal Activity of Musa paradisiaca Sap in Wistar Rats
Upload
independentCategory
view
1download
0
B
R
Sp
SQ1
NS
h
•••••
a
ARRAA
KAAMIIA
1
tiebp
Q2
h0
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
ARTICLE IN PRESSG ModelBR 9132 1–9
Behavioural Brain Research xxx (2014) xxx–xxx
Contents lists available at ScienceDirect
Behavioural Brain Research
jou rn al hom epage: www.elsev ier .com/ locate /bbr
esearch report
ubgroup-dependent effects of voluntary alcohol intake on behavioralrofiles in outbred Wistar rats
hima Momeni ∗, Erika Romaneuropharmacology, Addiction & Behaviour, Department of Pharmaceutical Biosciences, Uppsala University, P.O. Box 591, SE-751 24</postal>, Uppsala,weden
i g h l i g h t s
Large individual differences in alcohol intake were demonstrated.High-drinking rats displayed an alcohol deprivation effect.High-drinking animals showed a lower anxiety-like behavior after alcohol intake.High-drinking animals may consume alcohol for its anxiolytic properties.The present study highlights the importance of studying subgroups.
r t i c l e i n f o
rticle history:eceived 26 May 2014eceived in revised form 26 August 2014ccepted 30 August 2014vailable online xxx
eywords:nxietylcoholultivariate concentric square field (MCSF)
ntermittent accessndividual differencelcohol deprivation effect
a b s t r a c t
Experimental animal models are critical for understanding the genetic, environmental and neurobio-logical underpinnings of alcohol use disorders. Limited studies investigate alcohol-induced effects onbehavior using free-choice paradigms. The aims of the present experiment were to study voluntary alco-hol intake using a modified intermittent access paradigm, investigate the effects of voluntary alcoholintake on behavioral profiles in water- and alcohol-drinking rats, and select extreme low- and high-drinking animals for a more detailed behavioral characterization. Sixty outbred male Wistar rats wererandomized into water and alcohol groups. Behavioral profiles in the multivariate concentric squarefieldTM (MCSF) test were assessed prior to and after voluntary alcohol intake. The animals had intermit-tent access to 20% alcohol and water for three consecutive days per week for seven weeks. The resultsrevealed increased alcohol intake over time. No major alcohol-induced differences on behavior profileswere found when comparing water- and alcohol-drinking animals. The high-drinking animals displayedan alcohol deprivation effect, which was not found in the low-drinking animals. High-drinking rats had
lower risk-taking behavior prior to alcohol access and lower anxiety-like behavior after voluntary alcoholintake compared to low-drinking rats. In conclusion, the modified intermittent access paradigm may beuseful for pharmacological manipulation of alcohol intake. With regard to behavior, the present findingshighlights the importance of studying subgroup-dependent differences and add to the complexity ofindividual differences in behavioral traits of relevance to the vulnerability for excessive alcohol intake.39
40
41
42
43
. Introduction
Alcohol use disorders (AUD) are heterogeneous with regardo etiology, vulnerability for addiction and response to treatmentnvolving complex interactions between multiple genes [1–3] and
Please cite this article in press as: Momeni S, Roman E. Subgroup-depin outbred Wistar rats. Behav Brain Res (2014), http://dx.doi.org/10.10
nvironmental factors [4]. Many different risk factors for AUD haveeen identified including comorbid psychiatric disorders [5] andersonality traits such as impulsivity [6,7]. In addition, clusters of
∗ Corresponding author. Tel.: +46 18 4714620; fax: +46 18 4714253.E-mail address: [email protected] (S. Momeni).
ttp://dx.doi.org/10.1016/j.bbr.2014.08.058166-4328/© 2014 Published by Elsevier B.V.
44
45
46
© 2014 Published by Elsevier B.V.
personality traits brought together into typologies have been linkedto vulnerability for AUD [8]. Due to numerous involving factors, westill do not know why some individuals are more vulnerable forAUD than others and therefore knowledge of individual differencesin vulnerability is needed.
Experimental animal models are critical for understanding thegenetic, environmental and neurobiological underpinnings of AUD.One of the most consistent findings is that animals showing a
endent effects of voluntary alcohol intake on behavioral profiles16/j.bbr.2014.08.058
high anxiety-like behavior also show higher alcohol intake [9–12],but contrasting results have also been reported indicating on thecomplexity [13–15]. In most studies on this topic forced admin-istration paradigms are used, where the animal is administrated
47
48
49
50

ARTICLE IN PRESSG ModelBBR 9132 1–9
2 S. Momeni, E. Roman / Behavioural Brain Research xxx (2014) xxx–xxx
Fig. 1. The experimental outline. The animals were first tested in the MCSF, then randomly assigned into water- and alcohol-drinking groups and introduced to a two-bottlef days pa lcoho( ed ana
atbiohiihoitFspadeosab[
haeTcwtttlwIwap
2
2
Lah(a
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
ree-choice paradigm with intermittent access to 20% alcohol for three consecutive
gain tested in the MCSF, after which the experiment ended. Based on the average aLD), intermediate-drinking (ID) and high-drinking (HD) subgroups for more detail
specific dose of alcohol. The limitation with this procedure ishat individual predisposition to excessive alcohol intake cannote investigated. We have recently shown that risk-taking behav-
or displayed minor associations with voluntary alcohol intake inutbred rats while animals showing a high risk-assessing behaviorad a higher alcohol intake and preference, and increased alcohol
ntake over time compared to low risk-assessing rats. These find-ngs indicate that certain aspects of risk-related behaviors imply onigher vulnerability for alcohol intake [16]. Moreover, the literaturen alcohol-induced effects on behavior after voluntary drinkings limited and contrasting results have been reported likely dueo different intake paradigms, duration of intake and tests used.or instance, rats having access to 6% alcohol solution for 51 con-ecutive days spent more time on the open arms of the elevatedlus maze compared to the water controls [17]. Using a 1 h-limitedccess paradigm with 10% sweetened alcohol for 35 days, alcoholrinking animals had decreased activity on the open arms in thelevated plus maze [18]. Furthermore, when profiling adolescentutbred rats prior to and after a period of alcohol access in order totudy alcohol-induced behavior modulation over time no evidentlcohol-induced effects were found and the predictive value of theehavioral profiling in relation to later alcohol intake was weak19].
The aim of this study was to analyze the effects of voluntary alco-ol intake on behavior profiles in adult male rats. We here introduce
free-choice intermittent access paradigm modified from the mod-ls used by Wise [20] and [21], summarized by Carnicella et al. [22].he animals had free access to 20% alcohol and water during threeonsecutive days followed by a four-day intermission with access toater only. The behavioral profiling of the animals was made using
he multivariate concentric square fieldTM (MCSF) [23,24] wherehe first trial was conducted before the seven-week period of accesso alcohol and the second trial was conducted 3–7 days after theast alcohol session. Alcohol-induced effects on behavioral profiles
ere investigated by comparing water- and alcohol-drinking rats.n addition, subgroups of extreme low- and high-drinking animals
ere selected based on the alcohol intake during the last week ofccess for a more detailed characterization of behavioral profilesrior to and after voluntary alcohol intake.
. Material and methods
.1. Animals and housing
Sixty outbred male Wistar rats (RccHanTM:WI) from Harlanaboratories B.V., Horst, The Netherlands were delivered to the
Please cite this article in press as: Momeni S, Roman E. Subgroup-depin outbred Wistar rats. Behav Brain Res (2014), http://dx.doi.org/10.1
nimal facility at seven weeks of age. The rats were group-oused (three rats per cage) in transparent polysulfone cages59 cm × 38 cm × 20 cm) containing wood-chip bedding materialnd two paper sheets (40 cm × 60 cm; Cellstoff, Papyrus) for
er week for seven weeks. After the period of alcohol access, the animals were oncel intake during the last week of access, the animals were divided into low-drinkinglysis of behavioral profiles in LD and HD rats prior to and after access to alcohol.
enrichment purpose. The cages were placed in temperature-(21 ± 1 ◦C) and humidity-controlled (50 ± 10%) housing cabinetswith a reversed 12 h light/dark cycle (lights off at 07.00 h), and witha masking background noise. The rats were maintained on standardrat chow (R36; Lantmännen, Kimstad, Sweden) and water ad libi-tum. All animal experiments were approved by the Uppsala AnimalEthical Committee and followed the guidelines of the Swedish Leg-islation on Animal Experimentation (Animal Welfare Act SFS1998:56) and the European Communities Council Directive (86/609/EEC).
2.2. Experimental procedures
The outline of the experimental procedure is illustrated in Fig. 1.Upon arrival, the rats were undisturbed for two weeks to allowacclimatization to the animal facility and the reversed light/darkcycle. The week following the acclimatization period the rats werehandled daily using a procedure similar to that of previous exper-iments [24,25], consisting of individual handling, weighing, andadaptation to the transportation bucket that was used to trans-port the animals from the home cage to the testing room. The ratswere then tested in the MCSF test. Subsequently, after the MCSFtest, the animals were given access to alcohol solution and waterin the home cage for seven weeks. Finally, 3–7 days after the lastalcohol session, the animals were once again tested in the MCSFtest with the same conditions as in the first MCSF trial.
2.2.1. The multivariate concentric square fieldTM (MCSF)The MCSF test is an ethologically founded test and unlike many
other tests, it is unprejudiced in the sense that the animal is exposedto stimuli with different quality and can freely choose betweendifferent environments designed to include opportunity for explo-ration risk assessment, risk taking and shelter seeking. In this waya behavioral profile is generated. The details of the MCSF arenahave been described elsewhere [23,24] and in brief in Fig. S1. Thetest was performed in a room separate from the housing room,with the same conditions with regard to temperature, humidityand masking background noise to those in the housing room. Theanimals were tested using a running schedule to avoid disturb-ance in the home cages and to avoid order biases. All observationswere carried out during the dark period of the light/dark cycle. Thearena was divided into ten different zones, which forms the basisof the description and the variables of the animals’ performancein this test. The animals were started in the center zone facingthe wall with no openings, between the center and bridge (Fig.S1). Each animal was tested for 20 min. After each test the floorwas wiped with a cloth containing 10% alcohol solution and was
endent effects of voluntary alcohol intake on behavioral profiles016/j.bbr.2014.08.058
allowed to dry before the next animal was placed in the arena. Theapproximate light conditions (lux) were as following: dark cornerroom < 1; center approximately 20; corridors and hurdle 10–20;slope approximately 30; and bridge: 600–650.
141
142
143
144

ING ModelB
ral Bra
agdTto((rIitgiip
2
a(pufOw2thmbtnWacoc
aeaaaai
2
tattmgotTranmt
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
210
211
212
213
214
215
216
217
218
219
220
221
222
223
224
225
226
227
228
229
230
231
232
233
234
235
236
237
238
239
240
241
242
243
244
245
246
247
248
249
250
251
252
253
254
255
256
257
258
259
260
ARTICLEBR 9132 1–9
S. Momeni, E. Roman / Behaviou
The behavior was recorded by a video camera placed above therena and observed from an adjacent room. The number of rearings,roomings and stretched attend postures (SAPs) from the corri-ors to the center was manually recorded by direct observation.he number of fecal boli, urinations and number of head dips intohe hole board on the hurdle was noted after each trial. A blindedbserver scored the behavior manually using the program Score 3.3Copyright Soldis, Uppsala, Sweden). Latency (LAT, s), frequencyFRQ) and duration (DUR, s) of visits to each different zone wereegistered. Ethovision version 2.3 (Noldus Information Technologync., Wageningen, The Netherlands) was used for automatic track-ng in order to obtain mean velocity (cm/s) and total distance (cm)ravelled. An operational categorization of the various parametersenerated from the MCSF with regard to function (i.e., general activ-ty, exploration, risk assessment, risk taking and shelter seeking)s used in the interpretation of results. In addition a rank-orderrocedure referred to as the trend analysis is used [26].
.2.2. Voluntary alcohol intake and preferenceThe animals were randomly assigned into water (n = 30)
nd alcohol (n = 30) groups and placed individually in cages42 cm × 26 cm × 18 cm) with wooden houses for enrichment pur-ose. The alcohol-drinking animals were introduced to alcoholsing an intermittent two-bottle free-choice paradigm, modifiedrom Wise [20] and [21], and summarized by Carnicella et al. [22].n Tuesdays, Wednesdays, and Thursdays every week, the ratsere given 24 h access to one bottle of water and one bottle of
0% alcohol (v/v). On the other days, the animals had access towo bottles of water. Bottle positions were changed for each alco-ol session to avoid position preference. Alcohol solutions wereade from 96% ethanol (Solveco Etanol A 96%; Solveco AB, Rosers-
erg, Sweden) and tap water. Water and alcohol solution at roomemperature was available in 150 ml plastic bottles with ball-valveipples (Scanbur AB, Sollentuna, Sweden) with minimal spillage.ater and alcohol intake was measured at the end of each 24 h
ccess period by weighing the bottles. Alcohol preference was cal-ulated as alcohol intake (g) divided by the total fluid intake (g). Inrder to minimize disturbance during intake measures, cages werehanged and animals were weighed on Friday afternoons.
Based on the average alcohol intake during the last week ofccess, the alcohol-drinking animals were divided into three differ-nt groups; low-, intermediate- and high-drinking animals (LD, IDnd HD, respectively; n = 10 rats/group). This classification enabled
more detailed description of drinking patterns in the three groupss well as detailed analyses of behavioral profiles prior to and afterccess to alcohol in rats with a high and low voluntary alcoholntake, respectively.
.2.3. Statistical analysesStatistica 10.0 (StatSoft Inc., Tulsa, OK) was used for all statis-
ical analyses. Differences were considered statistically significantt p ≤ 0.05. Most data were not normally distributed according tohe Shapiro–Wilk’s W test. The non-parametric Mann–Whitney U-est was used to compare descriptive MCSF parameters and intake
easures in water- and alcohol-drinking groups, and LD and HDroups. To compare behavior parameters in the first and the sec-nd MCSF trial and to compare fluid intake during week seven tohat during week one, the Wilcoxon matched pair test was used.he Spearman Rank Order Correlation was used for analysis of cor-elations. Further analysis of MCSF data was done by using the trend
Please cite this article in press as: Momeni S, Roman E. Subgroup-depin outbred Wistar rats. Behav Brain Res (2014), http://dx.doi.org/10.10
nalysis [26]. Due to the ranking in the trend analysis, the data isormally distributed and comparisons between the groups wereade using one-way ANOVA and over time comparing the first and
he second MCSF trial using repeated measurements ANOVA.
PRESSin Research xxx (2014) xxx–xxx 3
3. Results
3.1. Alcohol intake and preference using the modifiedintermittent two-bottle free-choice paradigm
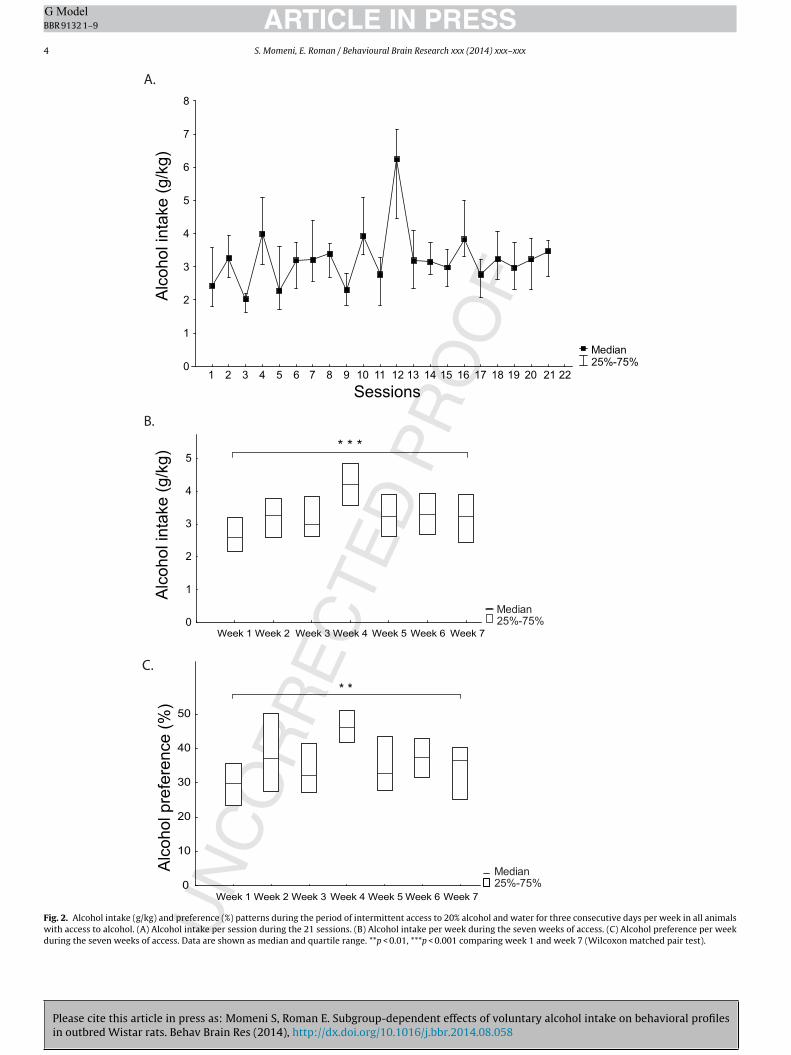
The animals had free-choice intermittent access to 20% alcoholevery Tuesday, Wednesday and Thursday. The pattern of alcoholintake (g/kg) per session and per week is shown in Fig. 2A and B,respectively. The median alcohol intake increased over time witha significant difference [Z = 3.5, p < 0.001] between the first and thelast week of alcohol access (Fig. 2B). Weekly alcohol preference(%) is shown in Fig. 2C. The median alcohol preference reflectedthe alcohol intake and increased over time with a significant differ-ence [Z = 2.6, p < 0.01] between the first and the last week of alcoholaccess (Fig. 2C). The alcohol intake during session 12 displayed amarked difference compared to the other sessions, affecting intakeand preference during week four. During this time the experimentalperson was changed for practical reasons but both experimenterswere female [27].
The median (quartile range, QR) total fluid intake (g/kg) alsoincreased over time with a significant difference [Z = 2.6, p < 0.01]between the first (55.1 (9.6)) and the last (57.7 (15.3)) week of alco-hol access. The total fluid intake did not differ between the alcohol-and water-drinking animals (data not shown). No body weight dif-ference between the alcohol and water group was found (data notshown).
3.2. Behavioral profiles in water- and alcohol-drinking rats
3.2.1. Trial IThe animals were tested in the MCSF and randomly divided
into water- and alcohol-drinking groups prior to alcohol access.The descriptive parameters from the 20-min trial in the MCSF areshown in Table S1. Only minor differences between the groups werenoted. The animals assigned to alcohol access had higher durationper visit in the hurdle and made more nose pokes into the hurdlehole board compared to animals assigned to drink water. No otherdifferences were found. The results from the MCSF trend analysis ofthe functional categories general activity, exploratory activity, riskassessment, risk taking and shelter seeking revealed no significantdifferences between animals assigned to drink water and alcohol(data not shown).
3.2.2. Trial IIThe animals were repeatedly tested after the seven-week period
of alcohol access and the descriptive parameters are shown in TableS1. Besides a higher number of nose pokes into the hurdle holeboard in alcohol-drinking rats compared to water-drinking rats,no differences were found when comparing the water- and thealcohol-drinking animals. The results from the MCSF trend analysisrevealed no significant group differences (data not shown).
3.2.3. Trial I and trial II over timeComparing trial I with trial II over time, differences within the
water- and alcohol-drinking groups were found (Table S1). Differ-ences over time were similar in both groups with the exception ofrisk-taking behavior in the central circle, which was significantlylower in the alcohol-drinking group only.
3.3. Alcohol intake and preference in low-drinking,intermediate-drinking and high-drinking rats
endent effects of voluntary alcohol intake on behavioral profiles16/j.bbr.2014.08.058
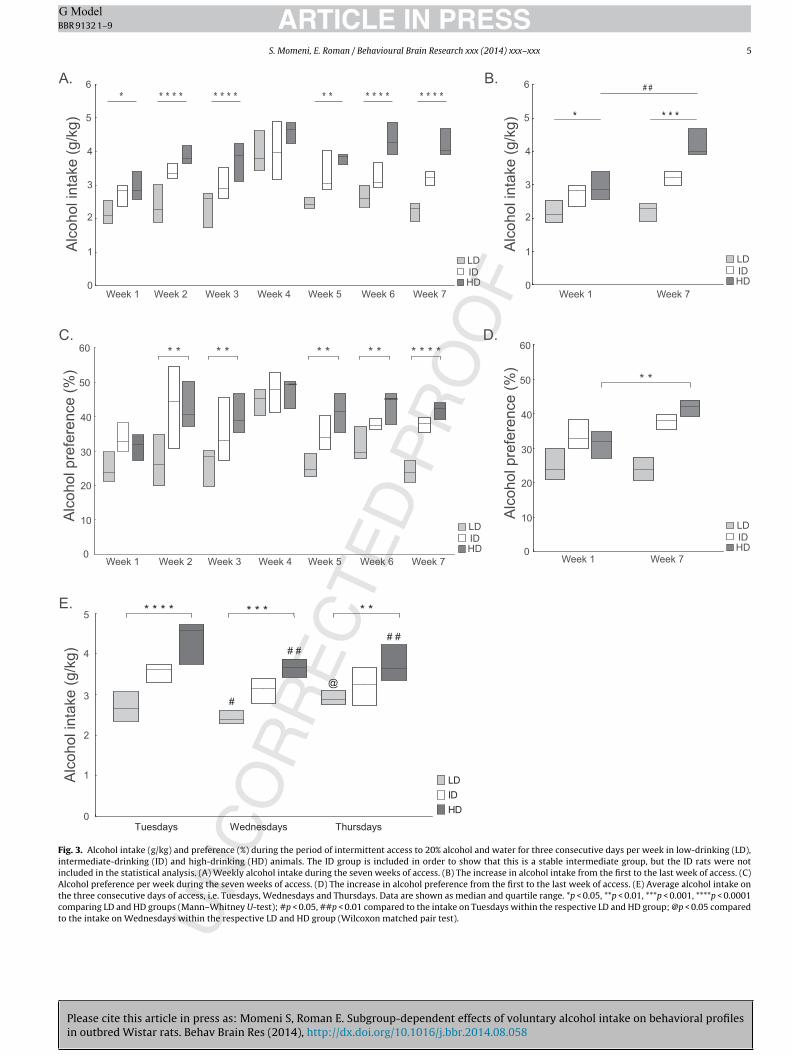
The animals were split by a tertiary split into low-drinking (LD),intermediate-drinking (ID) and high-drinking (HD) groups basedon the average intake during the last week of access to alcohol.The ID animals are shown to illustrate how this group relates to
261
262
263
264
Please cite this article in press as: Momeni S, Roman E. Subgroup-dependent effects of voluntary alcohol intake on behavioral profilesin outbred Wistar rats. Behav Brain Res (2014), http://dx.doi.org/10.1016/j.bbr.2014.08.058
ARTICLE IN PRESSG ModelBBR 9132 1–9
4 S. Momeni, E. Roman / Behavioural Brain Research xxx (2014) xxx–xxx
A.
B.
C.
Fig. 2. Alcohol intake (g/kg) and preference (%) patterns during the period of intermittent access to 20% alcohol and water for three consecutive days per week in all animalswith access to alcohol. (A) Alcohol intake per session during the 21 sessions. (B) Alcohol intake per week during the seven weeks of access. (C) Alcohol preference per weekduring the seven weeks of access. Data are shown as median and quartile range. **p < 0.01, ***p < 0.001 comparing week 1 and week 7 (Wilcoxon matched pair test).
Please cite this article in press as: Momeni S, Roman E. Subgroup-dependent effects of voluntary alcohol intake on behavioral profilesin outbred Wistar rats. Behav Brain Res (2014), http://dx.doi.org/10.1016/j.bbr.2014.08.058
ARTICLE IN PRESSG ModelBBR 9132 1–9
S. Momeni, E. Roman / Behavioural Brain Research xxx (2014) xxx–xxx 5
Fig. 3. Alcohol intake (g/kg) and preference (%) during the period of intermittent access to 20% alcohol and water for three consecutive days per week in low-drinking (LD),intermediate-drinking (ID) and high-drinking (HD) animals. The ID group is included in order to show that this is a stable intermediate group, but the ID rats were notincluded in the statistical analysis. (A) Weekly alcohol intake during the seven weeks of access. (B) The increase in alcohol intake from the first to the last week of access. (C)Alcohol preference per week during the seven weeks of access. (D) The increase in alcohol preference from the first to the last week of access. (E) Average alcohol intake onthe three consecutive days of access, i.e. Tuesdays, Wednesdays and Thursdays. Data are shown as median and quartile range. *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001comparing LD and HD groups (Mann–Whitney U-test); #p < 0.05, ##p < 0.01 compared to the intake on Tuesdays within the respective LD and HD group; @p < 0.05 comparedto the intake on Wednesdays within the respective LD and HD group (Wilcoxon matched pair test).

ING ModelB
6 ral Bra
taAhwatmsmdmaHt
ohwwi
da[coattlr
snNb
3
3
iOtithshNMeiassf
3
sTitp
265
266
267
268
269
270
271
272
273
274
275
276
277
278
279
280
281
282
283
284
285
286
287
288
289
290
291
292
293
294
295
296
297
298
299
300
301
302
303
304
305
306
307
308
309
310
311
312
313
314
315
316
317
318
319
320
321
322
323
324
325
326
327
328
329
330
331
332
333
334
335
336
337
338
339
340
341
342
343
344
345
346
347
348
349
350
351
352
353
354
355
356
357
358
359
360
361
362
363
364
365
366
367
368
369
370
371
372
373
374
375
376
377
378
379
380
381
382
383
384
ARTICLEBR 9132 1–9
S. Momeni, E. Roman / Behaviou
he LD and HD groups, respectively. However, in the statisticalnalysis focus is on the extreme groups, i.e. LD and HD animals.s shown in Fig. 3A, the weekly alcohol intake was significantlyigher in the HD animals compared to the LD animals during alleeks of alcohol access except for week four. When comparing the
lcohol intake week one to week seven, a significant increase overime was seen in the HD group only [Z = 2.7, p < 0.01] (Fig. 3B). The
edian (min-max) weekly cumulative alcohol intake (g/kg) aftereven weeks of alcohol access was 18.6 (16.1–20.4) for the LD ani-als and 28.4 (24.6–33.1) for the HD animals with a significant
ifference between the two groups [U = 00.0, Z = 3.7, p < 0.0001]. Theedian (min–max) alcohol load (g) after seven weeks of alcohol
ccess was 53.8 (41.5–76.1) and 76.9 (63.3–88.7) for the LD andD animals, respectively, with a significant difference between the
wo groups [U = 7.0, Z = 3.2, p < 0.001].The alcohol preference (Fig. 3C) followed a pattern similar to that
f the alcohol intake and the weekly preference was significantlyigher in the HD group during all weeks of alcohol access excepteek one and week four. When comparing the alcohol preferenceeek one to week seven, a significant increase over time was seen
n the HD group only [Z = 2.7, p < 0.01] (Fig. 3D).The average alcohol intake on Tuesdays, Wednesdays and Thurs-
ays is shown in Fig. 3E. The HD animals had a significantly higherlcohol intake on Tuesdays [U = 0.0, Z = 3.7, p < 0.0001], WednesdaysU = 4, Z = 3.4, p = 0.0001], and Thursdays [U = 10.0, Z = 3.0, p = 0.001]ompared to the LD animals. In the HD group, the alcohol intaken Tuesdays was significantly higher than that on Wednesdaysnd Thursdays, respectively; 30% higher on Tuesdays comparedo Wednesdays and 20% higher compared to Thursdays. The pat-ern was somewhat different in the LD group with a significantlyower intake on Wednesdays relative to Tuesdays and Thursdays,espectively.
The weekly water intake (g/kg) and total fluid intake (g/kg) ishown in Table S2. During week five the water intake was sig-ificantly lower in the HD animals compared to the LD animals.o other significant differences were found. No weight differenceetween the LD and the HD rats was found (data not shown).
.4. Behavioral profiles in low-drinking and high-drinking rats
.4.1. Trial IThe descriptive parameters from the 20-min trial in the MCSF
n LD and HD animals prior to alcohol access are shown in Table S3.nly a few differences between the groups were found. Notably,
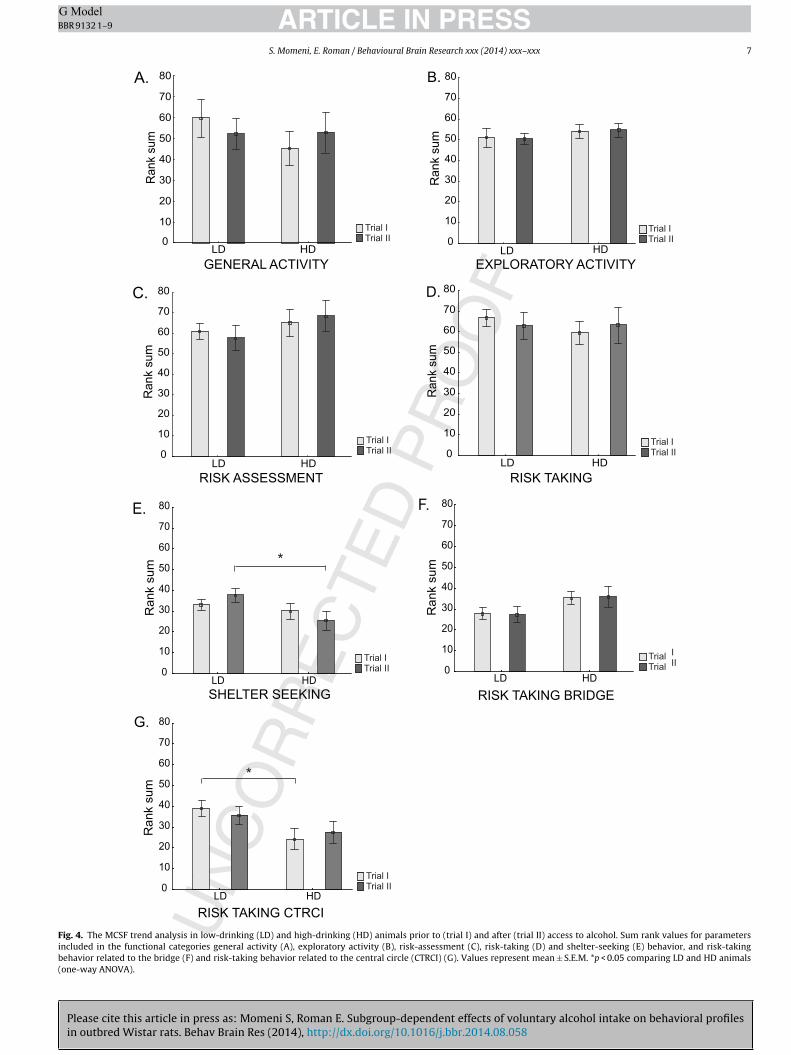
hese differences were related to the functional category risk tak-ng. The HD animals spent significantly longer time per visit inhe center compared to the LD animals. Moreover, the HD ratsad a shorter duration, percentage duration and a trend towards ahorter duration per visit (p = 0.06) in the central circle, indicatingigher thigmotactic behavior in the HD rats relative to the LD rats.o other significant differences were found. The results from theCSF trend analysis of the functional categories general activity,
xploratory activity, risk assessment, risk taking and shelter seek-ng (Fig. 4A–E) revealed no significant differences between the LDnd the HD groups. However, when the risk-taking category wasplit into bridge- and central circle-related parameters (Fig. 4F–G)ignificantly lower risk-taking behavior in the central circle wasound in HD relative to LD rats [F = 5.6, p = 0.03] (Fig. 4G).
.4.2. Trial IIThe descriptive parameters from the second MCSF trial, after the
even-week long period of access to alcohol, are shown in Table S3.
Please cite this article in press as: Momeni S, Roman E. Subgroup-depin outbred Wistar rats. Behav Brain Res (2014), http://dx.doi.org/10.1
he HD animals had a shorter duration and percentage durationn the dark corner room (DCR, i.e. the sheltered area) compared tohe LD animals. No other significant differences were found. Inter-retations about aspects of anxiety-like behavior revealed a lower
PRESSin Research xxx (2014) xxx–xxx
anxiety-like behavior in the HD group compared to the LD group[U = 22.0, Z = 2.07, p = 0.04]. The results from the MCSF trend analy-sis of the functional categories general activity, exploratory activity,risk assessment, risk taking and shelter seeking (Fig. 4) revealedlower shelter-seeking behavior in HD relative to LD rats [F = 4.5,p = 0.05] (Fig. 4E).
3.4.3. Trial I and trial II over timeComparing trial I and trial II over time revealed differences
within the LD and HD group, respectively (Table S3). HD rats spentmore time in the corridors in the second trial and LD animals madefewer visits to and spent less time in the hurdle in the second rela-tive to the first trial. Activity in the central circle was significantlylower in LD animals in trial II than in trial I. With regard to shelterseeking, LD animals spent more time and longer time per visit inthe DCR in the second trial. Anxiety-like behavior increased in theLD animals only in trial II relative to trial I. The results from theMCSF trend analysis of the functional categories general activity,exploratory activity, risk assessment, risk taking and shelter seek-ing (Fig. 4A–G) revealed no significant differences over time withineither the LD or the HD group.
4. Discussion
When subgrouping animals based on the alcohol intake dur-ing the last week of alcohol access using the modified intermittentaccess paradigm, the HD group not only had a higher alcohol intakeand preference compared to the LD group, but also had an increasedalcohol intake over time, which was not seen in the LD group. More-over, the HD group displayed a higher intake on Tuesdays relativeto Wednesdays and Thursdays, indicating on an alcohol depriva-tion effect, which was not found in the LD group. With regard tobehavior, the MCSF is used as a complementary methodologicalpossibility to understand mechanisms underlying a broader behav-ioral repertoire rather than focusing on any particular behavior.Minor overall differences in alcohol-induced effects on behavioralprofiles were found when comparing water- and alcohol-drinkinganimals. However, when focusing on the most extreme subgroupsamong the alcohol-drinking animals, the HD animals were char-acterized by lower risk-taking behavior prior to alcohol access andlower anxiety-like behavior after the period of alcohol consumptionrelative to LD rats.
4.1. Alcohol intake using three consecutive days of intermittentaccess
One aspect of human drinking patterns is high alcohol intake onconsecutive days, especially on weekends [28]. The modified modelused in this study, with alcohol access during three consecutivedays followed by a deprivation period for four days was introducedto mimic human weekend consumption. Using the modified inter-mittent access paradigm with three consecutive days of accessto 20% alcohol for seven weeks, the median alcohol intake dur-ing the last week of access in all alcohol-drinking animals was3.2 g/kg (1.8–5.8 g/kg) and the preference was 36.4% (16.0–46.2%).This is in agreement with previous studies using intermittent accessparadigms in which alcohol consumption up to 6 g/kg has beendescribed [22,29–32] and higher compared to studies in whichWistar rats were given continuous alcohol access [20,31,33]. More-over, compared to our recent study [16] in which the animals hadintermittent access to 20% alcohol on Mondays, Wednesdays andFridays the modified model used herein resulted in a higher alcohol
endent effects of voluntary alcohol intake on behavioral profiles016/j.bbr.2014.08.058
intake and a higher preference as well as an increased intake andpreference over time. One likely explanation for the higher intakeand preference found is the choice of Wistar sub-strain used in thisstudy [16,30].
385
386
387
388
Please cite this article in press as: Momeni S, Roman E. Subgroup-dependent effects of voluntary alcohol intake on behavioral profilesin outbred Wistar rats. Behav Brain Res (2014), http://dx.doi.org/10.1016/j.bbr.2014.08.058
ARTICLE IN PRESSG ModelBBR 9132 1–9
S. Momeni, E. Roman / Behavioural Brain Research xxx (2014) xxx–xxx 7
Fig. 4. The MCSF trend analysis in low-drinking (LD) and high-drinking (HD) animals prior to (trial I) and after (trial II) access to alcohol. Sum rank values for parametersincluded in the functional categories general activity (A), exploratory activity (B), risk-assessment (C), risk-taking (D) and shelter-seeking (E) behavior, and risk-takingbehavior related to the bridge (F) and risk-taking behavior related to the central circle (CTRCI) (G). Values represent mean ± S.E.M. *p < 0.05 comparing LD and HD animals(one-way ANOVA).

ING ModelB
8 ral Bra
amiihgtmsaoeihsgndnHttw[spfapaibild
4
aaiiaihcciu
aiiotavnecde
389
390
391
392
393
394
395
396
397
398
399
400
401
402
403
404
405
406
407
408
409
410
411
412
413
414
415
416
417
418
419
420
421
422
423
424
425
426
427
428
429
430
431
432
433
434
435
436
437
438
439
440
441
442
443
444
445
446
447
448
449
450
451
452
453
454
455
456
457
458
459
460
461
462
463
464
465
466
467
468
469
470
471
472
473
474
475
476
477
478
479
480
481
482
483
484
485
486
487
488
489
490
491
492
493
494
495
496
497
498
499
500
501
502
503
504
505
ARTICLEBR 9132 1–9
S. Momeni, E. Roman / Behaviou
A large individual variation in alcohol intake was found amongll alcohol-drinking animals, which enabled subgrouping of ani-als with high, intermediate and low alcohol intake. This finding
s in line with previous studies [22,32]. Since one part of this studys devoted to the introduction of and the characterization of alco-ol intake using the modified intermittent access paradigm the IDroup is shown in the figures despite being excluded in the sta-istical analysis, since the behavioral analysis was focused on the
ost extreme subgroups. However, this selection shows that theubgroups have a stable alcohol intake throughout the period ofccess to alcohol with preference stabilizing after the forth weekf access. The HD animals had significantly higher intake and pref-rence compared to the LD animals, and also significantly higherntake over time compared to the LD group in which the alco-ol intake was similar throughout the period of alcohol access. Ashown herein, not only the alcohol intake differed between theseroups, but also the drinking pattern. The HD animals had a sig-ificantly higher intake on Tuesdays, after the four day alcoholeprivation, compared to Wednesdays and Thursdays, which wasot seen in the LD group. This enhanced intake indicates that theD animals were more sensitive to the four days of interruption
o alcohol access, i.e. in the HD animals, this modified intermit-ent paradigm gave rise to an alcohol deprivation effect [34,35],hich is typically not seen when using intermittent access models
22] and to our knowledge only has been reported in one previoustudy [36]. The alcohol deprivation effect found herein was not asronounced as that described for some Wistar rats when deprivedrom alcohol after about two months of continuous access under
4-bottle free-choice paradigm [35]. However, this alcohol accessaradigm, with pronounced subgroup-dependent effects, includinglcohol deprivation effects, has clinical validity and may be usefuln future investigations of molecular, neuronal, neurochemical, orehavioral adaptations related to excessive alcohol intake as both
ndividuals that escalate their intake over time (HD animals) andow-drinking individuals (LD animals) without escalation are to beistinguished [22].
.2. Behavior in water- and alcohol-drinking rats
Overall, the predictive value of the behavior prior to alcoholccess on later alcohol intake was weak when comparing water-nd alcohol-drinking animals, which is in line with previous stud-es using the same behavior test to predict voluntary alcohol intaken adolescent animals [19]. We have previously shown that whennimals were split into groups based on risk-assessment behaviorn the MCSF prior to alcohol access, the high risk-assessing ratsad a higher alcohol intake when later given access to alcoholompared to the low-risk-assessing rats [16]. Herein, no asso-iation between risk-assessment behavior and voluntary alcoholntake was found, which may relate to the different study designsed.
The lack of differences in MCSF performance between water-nd alcohol-drinking rats following a period of voluntary alcoholntake is in line with previous studies in which other behav-oral tests were used [37,38] and also following after a periodf abstinence[38]. However, previous studies have revealed con-rasting results with both increased activity [17] and decreasedctivity [18] on the open arms of the elevated plus maze followingoluntary alcohol intake relative to water-drinking controls. Sinceo group-housed water-drinking control group was included the
Please cite this article in press as: Momeni S, Roman E. Subgroup-depin outbred Wistar rats. Behav Brain Res (2014), http://dx.doi.org/10.1
ffects of single housing masking potential alcohol-induced effectsannot be ruled out. It should also be emphasized that this studyesign represents non-dependent rats, which to a large extent mayxplain differences as compared to other findings in Ref. [22].
PRESSin Research xxx (2014) xxx–xxx
4.3. Behavior in low-drinking and high-drinking animals
Despite a lack of differences in MCSF performance betweenwater- and alcohol-drinking rats after the period with access toalcohol, differences within the alcohol-drinking animals emergedwhen studying the subgroups of LD and HD animals in more detail.The HD animals had a shorter duration in the central circle prior toalcohol access compared to the LD animals. As previous results haverevealed differences in risk-taking behavior in the central circleversus on the bridge [26,39], the functional category risk taking wasalso divided into performance on the bridge and in the central circle,respectively. The HD animals showed a lower risk-taking behaviorin central circle-related parameters compared to the LD animals.When repeatedly tested after the period with intermittent accessto alcohol, the HD animals spent shorter time in the DCR, had loweranxiety-like behavior as indicated by the higher risk/shelter indexand lower shelter-seeking behavior in the trend analysis, which alltogether is interpreted as lower anxiety-like behavior compared tothe LD rats. In addition, anxiety-like behavior increased over timein the LD animals only.
Considering the differences between LD and HD rats prior toalcohol access, an evident profile predisposing HD rats cannotbe found in the MCSF trend analysis. However, when taking thefindings together, the fact that the HD rats were characterizedby lower risk-taking behavior prior to alcohol access and loweranxiety-like behavior after alcohol consumption, compared to theLD rats, may indicate that the HD animals consumed alcoholfor its anxiolytic properties likely due to alcohol-induced neu-roadaptive changes. This finding is in line with previous studiesshowing the association of anxiety-like behavior and high alco-hol consumption (e.g. [10,12]) but contrasts our recent findingof risk-assessment behavior being associated with high volun-tary alcohol intake [16] indicating on the complexity in assessingbehavioral predispositions to alcohol intake as well as alcohol-induced effects on behavior. However, these results demonstratethat even if differences cannot be seen between the alcohol- andwater-drinking groups, there are individual differences among thealcohol-drinking animals that become evident when studying sub-groups of animals in more detail.
5. Conclusions
In this study, the effects of voluntary alcohol intake on behav-ior in non-dependent outbred Wistar rats were investigated. Largeindividual differences in alcohol intake were demonstrated whenusing the modified intermittent access paradigm. This enableddetailed analyses of subgroups of low and high alcohol-drinkingrats, respectively. An alcohol deprivation effect was observed inHD animals only, which may be of importance in further phar-macological studies. Moreover, the HD animals were interpretedas having lower anxiety-like behavior after alcohol intake. Thus,this subgroup of animals may consume alcohol for its anxiolyticproperties. Taken together, the present study adds to the complexliterature on interactions between voluntary alcohol intake andbehavioral traits but also highlights the importance of studyingsubgroup-dependent differences in alcohol intake and behav-ior.
Author contributions
endent effects of voluntary alcohol intake on behavioral profiles016/j.bbr.2014.08.058
Conceived and designed the experiment: ER; Performed theexperiment: SM; Analyzed the data: SM; Interpreted the resultsand wrote the manuscript: SM, ER.
506
507
508

ING ModelB
ral Bra
A
amsR&a
A
t
R
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
509
510
511
512
513
514
515
516
517
518
519
520
521
522
523
524
525
526
527
528
529
530
531
532
533
534
535
536
537
538
539
540
541
542
543
544
545
546
547
548
549
550
551
552
553
554
555
556
557
558
559
560
561
562
563
564
565
566
567
568
569
570
571
572
573
574
575
576
577
578
579
580
581
582
583
584
585
586
587
588
589
590
591
592
593
594
595
596
597
598
599
600
601
602
603
604
605
606
607
608
609
610
611
612
613
614
615
616
617
618
619
620
621
622
623
624
625
626
ARTICLEBR 9132 1–9
S. Momeni, E. Roman / Behaviou
cknowledgements
The authors would like to thank Dr. Lena Bergström for valu-ble input on the experimental design and for comments on theanuscript, and Ms. Marita Berg for technical assistance. Financial
upport from the Alcohol Research Council of the Swedish Alcoholetailing Monopoly, and the Magnus Bergvall, Åke Wiberg, Fredrik
Ingrid Thuring (ER) and Facias (ER, SM) Foundations is gratefullycknowledged.
ppendix A. Supplementary data
Supplementary data associated with this article can be found, inhe online version, at http://dx.doi.org/10.1016/j.bbr.2014.08.058.
eferences
[1] Ducci F, Goldman D. The genetic basis of addictive disorders. Psychiatr ClinNorth Am 2012;35:495–519.
[2] Kalsi G, Prescott CA, Kendler KS, Riley BP. Unraveling the molecular mechanismsof alcohol dependence. Trends Genet 2009;25:49–55.
[3] Morozova TV, Goldman D, Mackay TF, Anholt RR. The genetic basis of alco-holism: multiple phenotypes, many genes, complex networks. Genome Biol2012;13:239.
[4] De Bellis MD. Developmental traumatology: a contributory mechanismfor alcohol and substance use disorders. Psychoneuroendocrinology2002;27:155–70.
[5] Kushner MG, Abrams K, Borchardt C. The relationship between anxiety disor-ders and alcohol use disorders: a review of major perspectives and findings.Clin Psychol Rev 2000;20:149–71.
[6] Aragues M, Jurado R, Quinto R, Rubio G. Laboratory paradigms of impulsivityand alcohol dependence: a review. Eur Addict Res 2011;17:64–71.
[7] Lejuez CW, Magidson JF, Mitchell SH, Sinha R, Stevens MC, de Wit H. Behav-ioral and biological indicators of impulsivity in the development of alcohol use,problems, and disorders. Alcohol Clin Exp Res 2010;34:1334–45.
[8] Leggio L, Kenna GA, Fenton M, Bonenfant E, Swift RM. Typologies of alco-hol dependence. From Jellinek to genetics and beyond. Neuropsychol Rev2009;19:115–29.
[9] Bahi A. Individual differences in elevated plus-maze exploration predictedhigher ethanol consumption and preference in outbred mice. PharmacolBiochem Behav 2013;105:83–8.
10] Hayton SJ, Mahoney MK, Olmstead MC. Behavioral traits predicting alcoholdrinking in outbred rats: an investigation of anxiety, novelty seeking, and cog-nitive flexibility. Alcohol Clin Exp Res 2012;36:594–603.
11] McKinzie DL, Sajdyk TJ, McBride WJ, Murphy JM, Lumeng L, Li TK, et al. Acousticstartle and fear-potentiated startle in alcohol-preferring (P) and -nonpreferring(NP) lines of rats. Pharmacol Biochem Behav 2000;65:691–6.
12] Spanagel R, Montkowski A, Allingham K, Stohr T, Shoaib M, Holsboer F,et al. Anxiety: a potential predictor of vulnerability to the initiation ofethanol self-administration in rats. Psychopharmacology (Berl) 1995;122:369–73.
13] Henniger MS, Spanagel R, Wigger A, Landgraf R, Holter SM. Alcohol self-administration in two rat lines selectively bred for extremes in anxiety-relatedbehavior. Neuropsychopharmacology 2002;26:729–36.
14] Koros E, Piasecki J, Kostowski W, Bienkowski P. Saccharin drinking rather thanopen field behaviour predicts initial ethanol acceptance in Wistar rats. AlcoholAlcohol 1998;33:131–40.
Please cite this article in press as: Momeni S, Roman E. Subgroup-depin outbred Wistar rats. Behav Brain Res (2014), http://dx.doi.org/10.10
15] Langen B, Fink H. Anxiety as a predictor of alcohol preference in rats? ProgNeuro-Psychoph 2004;28:961–8.
16] Momeni S, Sharif M, Agren G, Roman E. Individual differences in risk-relatedbehaviors and voluntary alcohol intake in outbred Wistar rats. Behav Pharmacol2014;25:206–15.
[
PRESSin Research xxx (2014) xxx–xxx 9
17] Pohorecky LA. Psychosocial stress and chronic ethanol ingestion in male rats:effects on elevated plus maze behavior and ultrasonic vocalizations. PhysiolBehav 2008;94:432–47.
18] Silvestre JS, Pallares M, Nadal R, Ferre N. Opposite effects of ethanol andketamine in the elevated plus-maze test in Wistar rats undergoing a chronicoral voluntary consumption procedure. J Psychopharmacol 2002;16:305–12.
19] Palm S, Daoura L, Roman E, Nylander I. Effects of rearing conditions onbehaviour and endogenous opioids in rats with alcohol access during adoles-cence. PloS one 2013;8:e76591.
20] Wise RA. Voluntary ethanol intake in rats following exposure to ethanol onvarious schedules. Psychopharmacologia 1973;29:203–10.
21] Wayner MJ, Greenberg I, Tartaglione R, Nolley D, Fraley S, Cott A. A new factoraffecting the consumption of ethyl alcohol and other sapid fluids. Physiol Behav1972;8:345–62.
22] Carnicella S, Ron D, Barak S. Intermittent ethanol access schedule in rats as apreclinical model of alcohol abuse. Alcohol 2014;48:243–52.
23] Meyerson BJ, Augustsson H, Berg M, Roman E. The Concentric Square Field: amultivariate test arena for analysis of explorative strategies. Behav Brain Res2006;168:100–13.
24] Roman E, Colombo G. Lower risk taking and exploratory behavior in alcohol-preferring sP rats than in alcohol non-preferring sNP rats in the multivariateconcentric square field (MCSF) test. Behav Brain Res 2009;205:249–58.
25] Roman E, Stewart RB, Bertholomey ML, Jensen ML, Colombo G, Hyytia P, et al.Behavioral profiling of multiple pairs of rats selectively bred for high and lowalcohol intake using the MCSF test. Addict Biol 2012;17:33–46.
26] Meyerson BJ, Jurek B, Roman E. A rank-order procedure applied to an ethoex-perimental behavior model - the multivariate concentric square field (MCSF)test. JBBS 2013;3:350–61.
27] Sorge RE, Martin LJ, Isbester KA, Sotocinal SG, Rosen S, Tuttle AH, et al. Olfac-tory exposure to males, including men, causes stress and related analgesia inrodents. Nat Methods 2014;11:629–32.
28] Room R, Makela P, Benegal V, Greenfield TK, Hettige S, Tumwesigye NM, et al.Times to drink: cross-cultural variations in drinking in the rhythm of the week.Int J Public Health 2012;57:107–17.
29] Hargreaves GA, Monds L, Gunasekaran N, Dawson B, McGregor IS. Intermittentaccess to beer promotes binge-like drinking in adolescent but not adult Wistarrats. Alcohol 2009;43:305–14.
30] Palm S, Roman E, Nylander I. Differences in voluntary ethanol consumption inWistar rats from five different suppliers. Alcohol 2011;45:607–14.
31] Simms JA, Steensland P, Medina B, Abernathy KE, Chandler LJ, Wise R, et al.Intermittent access to 20% ethanol induces high ethanol consumption in Long-Evans and Wistar rats. Alcohol Clin Exp Res 2008;32:1816–23.
32] Steensland P, Fredriksson I, Holst S, Feltmann K, Franck J, Schilstrom B, et al.The monoamine stabilizer (−)-OSU6162 attenuates voluntary ethanol intakeand ethanol-induced dopamine output in nucleus accumbens. Biol Psychiatry2012;72:823–31.
33] Nylander I, Roman E. Is the rodent maternal separation model a valid andeffective model for studies on the early-life impact on ethanol consumption?Psychopharmacology (Berl) 2013;229:555–69.
34] Sinclair JD, Senter RJ. Development of an alcohol-deprivation effect in rats. Q JStud Alcohol 1968;29:863–7.
35] Vengeliene V, Bilbao A, Spanagel R. The alcohol deprivation effect modelfor studying relapse behavior: a comparison between rats and mice. Alcohol2014;48:313–20.
36] Mill DJ, Bito-Onon JJ, Simms JA, Li R, Bartlett SE. Fischer rats consume 20%ethanol in a long-term intermittent-access two-bottle-choice paradigm. PloSOne 2013;8:e79824.
37] Butler TR, Ariwodola OJ, Weiner JL. The impact of social isolation on HPA axisfunction, anxiety-like behaviors, and ethanol drinking. Front Integr Neurosci2014;7(102).
38] George O, Sanders C, Freiling J, Grigoryan E, Vu S, Allen CD, et al. Recruitmentof medial prefrontal cortex neurons during alcohol withdrawal predicts cog-
endent effects of voluntary alcohol intake on behavioral profiles16/j.bbr.2014.08.058
nitive impairment and excessive alcohol drinking. Proc Natl Acad Sci U S A2012;109:18156–61.
39] Palm S, Momeni S, Lundberg S, Nylander I, Roman E. Risk-assessment andrisk-taking behavior predict potassium- and amphetamine-induced dopamineresponse in the dorsal striatum of rats. Front Behav Neurosci 2014;8:236.
627
628
629
630
631
Copyright © 2022 FDOKUMEN




















