
Transsexual limb transplants in fiddler crabs and expression of novel sensory capabilities
10
Transsexual Limb Transplants in Fiddler Crabs and Expression of Novel Sensory Capabilities MARC J. WEISSBURG, 1 * CHARLES D. DERBY, 2 OMAR JOHNSON, 2 BRIAN MCALVIN, 1 AND JAMES M. MOFFETT JR. 2 1 School of Biology, Georgia Institute of Technology, Atlanta, Georgia 30332 2 Department of Biology and Center for Behavioral Neuroscience, Georgia State University, Atlanta, Georgia 30303 ABSTRACT We used transsexual limb transplants in fiddler crabs to examine how peripheral sensory structures interact with the central nervous system (CNS) to produce a sexually dimorphic behavior. Female and male chemosensory feeding claws were transplanted onto male hosts in place of nonfeeding, nonchemosensory claws. Successfully transplanted claws retain donor morphologies and contain chemosensory neurons. Neurons in successfully transplanted fe- male feeding claws express the enhanced sensitivity to chemical cues seen in female, but not male, neurons in claws of normal animals. When chemically stimulated, the transplanted claws evoke feeding behavior not observed in normal males, even though the sensory neurons in the transplanted limb project to the host’s sexually dimorphic neuropil not known to receive chemosensory input. Behavioral sensitivity is directly related to the sensitivity of peripheral neurons in the transplanted feeding claw. Thus, the interactions between periph- eral neurons and their targets may restructure the CNS so that novel sensory capabilities are expressed, and this can produce sexually dimorphic behaviors. J. Comp. Neurol. 440:311–320, 2001. © 2001 Wiley-Liss, Inc. Indexing terms: sexual dimorphism; limb transplants; fiddler crabs; sensory capabilities; neuronal plasticity The properties and function of sensory systems depend on characteristics of both the peripheral sensory elements and the central nervous system (CNS). Sensitivity and dynamic range, among other properties of receptor neu- rons, set limits on responses to physical stimuli such as light, sound, and chemical cues (see, e.g., Atema et al., 1988; Weissburg and Derby, 1995; Lehrer, 1997). Projec- tion patterns of sensory afferents map various stimulus features onto a spatial dimension in many sensory sys- tems (see, e.g., Konishi, 1986; Hayama and Caprio, 1989; Newland et al., 2000). In the CNS, modality-specific orga- nization of the neuropil (see, e.g., Murphey et al., 1989; Killian et al., 1993; Newland et al., 2000) and spatial organization of distinct cell types (Goldsmith, 1991; Mei- sami, 1991) occur that presumably reflect the need for particular patterns of connectivity necessary for proper sensory functioning. Although it is possible to describe such features as a property of either the CNS or the periphery, in many cases it is the interaction between peripheral sensory elements and their targets that affects the structure and operation of the CNS. This observation is supported by a variety of transplantation experiments that induce axons to grow into foreign neuropil, which demonstrate that sensory neurons have the capacity to influence patterns of organization in the CNS (Schneider- man et al., 1986; Sur et al., 1990; Scalia et al., 1995; Ro ¨ssler et al., 1999; Yaka et al., 1999). It is difficult to assess fully the significance of peripheral effects, because the ability to experimentally link sensory elements with given properties to specific CNS elements is rare, particularly in systems in which corresponding be- havioral and physiological studies are possible. Thus, it is fairly difficult to establish the extent to which a peripheral Omar Johnson’s current address is: 5145 Cascade Hills, Atlanta, GA 30331. James M. Moffett Jr.’s current address is: Department of Botany and Microbiology, Auburn University, Auburn, AL 36849. *Correspondence to: Marc J. Weissburg, School of Biology, Georgia In- stitute of Technology, Atlanta, GA 30332. Received 3 May 2001; Revised 10 August 2001; Accepted 27 August 2001 THE JOURNAL OF COMPARATIVE NEUROLOGY 440:311–320 (2001) © 2001 WILEY-LISS, INC.
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Transsexual limb transplants in fiddler crabs and expression of novel sensory capabilities

Transsexual Limb Transplants in FiddlerCrabs and Expression of Novel
Sensory Capabilities
MARC J. WEISSBURG,1* CHARLES D. DERBY,2 OMAR JOHNSON,2
BRIAN MCALVIN,1AND JAMES M. MOFFETT JR.2
1School of Biology, Georgia Institute of Technology, Atlanta, Georgia 303322Department of Biology and Center for Behavioral Neuroscience, Georgia State University,
Atlanta, Georgia 30303
ABSTRACTWe used transsexual limb transplants in fiddler crabs to examine how peripheral sensory
structures interact with the central nervous system (CNS) to produce a sexually dimorphicbehavior. Female and male chemosensory feeding claws were transplanted onto male hosts inplace of nonfeeding, nonchemosensory claws. Successfully transplanted claws retain donormorphologies and contain chemosensory neurons. Neurons in successfully transplanted fe-male feeding claws express the enhanced sensitivity to chemical cues seen in female, but notmale, neurons in claws of normal animals. When chemically stimulated, the transplantedclaws evoke feeding behavior not observed in normal males, even though the sensory neuronsin the transplanted limb project to the host’s sexually dimorphic neuropil not known toreceive chemosensory input. Behavioral sensitivity is directly related to the sensitivity ofperipheral neurons in the transplanted feeding claw. Thus, the interactions between periph-eral neurons and their targets may restructure the CNS so that novel sensory capabilities areexpressed, and this can produce sexually dimorphic behaviors. J. Comp. Neurol. 440:311–320,2001. © 2001 Wiley-Liss, Inc.
Indexing terms: sexual dimorphism; limb transplants; fiddler crabs; sensory capabilities; neuronal
plasticity
The properties and function of sensory systems dependon characteristics of both the peripheral sensory elementsand the central nervous system (CNS). Sensitivity anddynamic range, among other properties of receptor neu-rons, set limits on responses to physical stimuli such aslight, sound, and chemical cues (see, e.g., Atema et al.,1988; Weissburg and Derby, 1995; Lehrer, 1997). Projec-tion patterns of sensory afferents map various stimulusfeatures onto a spatial dimension in many sensory sys-tems (see, e.g., Konishi, 1986; Hayama and Caprio, 1989;Newland et al., 2000). In the CNS, modality-specific orga-nization of the neuropil (see, e.g., Murphey et al., 1989;Killian et al., 1993; Newland et al., 2000) and spatialorganization of distinct cell types (Goldsmith, 1991; Mei-sami, 1991) occur that presumably reflect the need forparticular patterns of connectivity necessary for propersensory functioning. Although it is possible to describesuch features as a property of either the CNS or theperiphery, in many cases it is the interaction betweenperipheral sensory elements and their targets that affectsthe structure and operation of the CNS. This observation
is supported by a variety of transplantation experimentsthat induce axons to grow into foreign neuropil, whichdemonstrate that sensory neurons have the capacity toinfluence patterns of organization in the CNS (Schneider-man et al., 1986; Sur et al., 1990; Scalia et al., 1995;Rossler et al., 1999; Yaka et al., 1999).
It is difficult to assess fully the significance of peripheraleffects, because the ability to experimentally link sensoryelements with given properties to specific CNS elements israre, particularly in systems in which corresponding be-havioral and physiological studies are possible. Thus, it isfairly difficult to establish the extent to which a peripheral
Omar Johnson’s current address is: 5145 Cascade Hills, Atlanta, GA30331.
James M. Moffett Jr.’s current address is: Department of Botany andMicrobiology, Auburn University, Auburn, AL 36849.
*Correspondence to: Marc J. Weissburg, School of Biology, Georgia In-stitute of Technology, Atlanta, GA 30332.
Received 3 May 2001; Revised 10 August 2001; Accepted 27 August 2001
THE JOURNAL OF COMPARATIVE NEUROLOGY 440:311–320 (2001)
© 2001 WILEY-LISS, INC.

structure, either through its specific properties or via itsinfluence on the organization of the CNS, dictates a givenlevel of sensory performance. To examine the extent towhich the periphery can legislate behavior, we asked asimple question: Can the transplantation of sexually di-morphic appendages make connections with novel targetsin the host CNS to produce behavior normally seen only inthe donor organism?
Fiddler crabs (genus Uca) are sexually dimorphic crus-taceans especially suitable for transplantation studies.Males possess a single hypertrophied chela (the majorclaw), which is used for communication and defense, and asingle feeding (or minor) claw. Females have two feedingclaws (Crane, 1975). Chemosensory neurons in femaleclaws express greater sensitivity to chemical stimuli thanthe neurons in male feeding claws (Weissburg and Derby,1995; Weissburg, 1999), and this disparity appears, inpart, to regulate sex-specific behavioral thresholds ob-served during feeding (Weissburg, 1993). Chemosensoryneurons are known to be present in the feeding claws ofmales and females; male major claws have never beenshown to detect chemical cues in either physiological(Weissburg and Derby, 1995) or extensive behavioral (Va-liela et al., 1974; Robertson et al., 1981; Weissburg andZimmer-Faust, 1991; Weissburg, 1993) investigations.
In this study, we transplanted female feeding clawsonto the site of the major claw of host males to determinewhether behavioral sensitivity characteristic of femaleswas contingent on the identity of the transplanted limb orthe host CNS. Thus, we ask questions concerning both thedegree of autonomy expressed by developing limbs and theextent to which neurons in these limbs subsequently in-fluence the ability of the CNS to process novel sensoryinputs. We find that, indeed, sensory neurons from trans-planted feeding claws retain their characteristic sensitiv-ity, successfully invade foreign neuropil to establish func-tional connections, and produce behavior that ispredictable from the properties of the donor limb.
MATERIALS AND METHODS
Transplantation
The sand fiddler crab, Uca pugilator, was used in thesestudies. Crabs were obtained from a commercial supplierand shipped to our laboratory. There they were held in 45liter aquaria containing 25 ppt seawater and natural sandsubstrates. A microprocessor-controlled timer was used todrain and fill the tanks to simulate tidal conditions nor-mally experienced by animals in the field. Donor animalswere induced to autotomize feeding claws by squeezing onthe merus of the appendage with a forceps. Six walkinglegs were removed by the same method in order to hastenmolting (Hopkins et al., 1999). Thereafter, crabs were keptindividually in small bowls with micrometer-filtered, ster-ile artificial seawater (ASW) media in an incubator at26°C and a 12:12 hour L:D cycle.
Regenerating crustacean limbs develop through a seriesof stages into a bud-like structure, the limb bud, duringthe intermolt period following limb loss (Hopkins et al.,1999). The limb bud matures into a fully formed limbwhen the animal molts. Following automization, we mon-itored the development of this bud in the donors, and,when it was approximately 2 mm long, animals weretransferred to ASW supplemented with 30 mg/liter genta-
micin and 4 ml/liter Fungizone for 3 days prior to surgery.At this same time, host males were size matched withdonors and also placed in antibiotic ASW. Immediatelyprior to surgery, donor and hosts were chilled to quies-cence, placed ventral side up, and immobilized with wax.The limb bud of the donor crab was removed at the coxa-basis joint, and the hard cuticle of the ischium wasstripped away. Simultaneously, a second person preparedthe host crab by inducing autotomy of the major claw andsix walking legs, and breaking through the autotomymembrane that seals the wound after limb loss. The budwas carefully transferred to the host and oriented in thesocket, using a small amount of cyanoacrylate adhesive toseal the bud onto the host. The host was returned toantibiotic ASW and held in the incubator for another 1week. Thereafter, the animals were kept in sterile ASWuntil they molted. An appreciable proportion (see Results)of animals showed apparent regression or lack of develop-ment of the transplant, so on the first postsurgical moltonly a small, undifferentiated bud (the blastema) waspresent. These animals were allowed to feed ad libitum for1 week and then subjected to another round of autotomy oftheir walking legs to induce a second postsurgical molt.
Using these techniques, we transplanted 448 feedingclaws from female donors and 235 feeding claws donatedfrom males. These experiments were performed in thesummers of 1998, 1999, and 2000, because animals areavailable only seasonally.
To facilitate morphological and functional characteriza-tion of transplanted feeding claws, we performed a varietyof other manipulations to produce regenerating majorclaws. Transplant controls (sham-operated males) werecreated by autotomizing the major claw followed by re-moving the developing limb bud when it reached approx-imately 2 mm in size. The subsequent molt produced ananimal with a developing limb bud on the manipulatedside, which is the same condition experienced by animalsthat failed to develop a claw from a transplanted bud onthe first postsurgical molt. In addition, we autotomizedmajor claws and allowed them to regenerate naturally.Table 1 lists the various experimental treatments, thedescriptive terms used throughout the text, and a briefsummary of their properties.
Neuronatomical studies
In fiddler crabs, all of the nerves emanating from theclaw on a given body side project to the ipsilateral firstthoracic hemiganglion. The left and right hemiganglia area bilateral pair of structures that are part of the fused-ringthoracic ganglion, which contains local processing regionscorresponding to each of the five limb-bearing thoracicsegments.
We performed two different types of anatomical studieson animals receiving transplants. One group of animalswas prepared for transmission electron microscopy (TEM)using protocols described fully by Weissburg et al. (1996).The CNS was fixed in situ using a standard crustaceanformaldehyde– glutaraldeyde fixative, postfixed in 2%osmium tetroxide, and dehydrated in a graded ethanolseries. The hemiganglion receiving the axons from suc-cessfully transplanted claws and the contralateral hemi-ganglion, which receives axons from the native claw, werecleared in propylene oxide and embedded in Araldite-Eponresin. The entire nerve bundle was thin sectioned (thick-ness 0.075 mm) as it entered the ganglion in order to
312 M.J. WEISSBURG ET AL.

visualize the axons that projected to this neuropil on boththe transplanted and the native claw sides. Single photo-graphs of the entire root were taken at low magnification(3200–300) with a Siemens 102 transmission electronmicroscope.
Dye-tracing studies were performed on three host crabsbearing transplanted feeding claws using the carbocya-nine dye DiI protocol developed for crustaceans (Sande-man and Sandeman, 1994). The entire fused-ring thoracicganglion was removed and fixed using standard protocols(Weissburg et al., 1996). A small crystal of DiI was placedon the cut end of the nerve that contains all the axons fromthe transplanted claw, and this preparation was allowedto remain in the dark for at least 6 months at roomtemperature, at which time visual inspections indicatedthat the DiI had stopped migrating. The specimen wascleared in meglumine iothalamate for easier visualizationunder light microscopy (Zill et al., 1993). The whole mountwas then viewed and photographed on a Zeiss confocalmicroscope using LSM 510 software. A series of stackedimages was taken using a 53 objective, and then eachseries was collapsed to produce a single two-dimensionalimage.
Physiology of chemosensory neurons
Methods were used as described by Weissburg andDerby (1995). Briefly, claws were autotomized, stripped ofall cuticle proximal to the merus to expose the nerve andartery, and placed into a glass and Teflon olfactometer.The claw was perfused by delivering oxygenated artificialsaline into the artery via a glass cannula, and stimuliwere supplied using a computer-controlled solenoid valvethat injected 0.5 ml of stimulus into ASW flowing over theclaw. A fine-tipped, saline-filled microelectrode was at-tached by suction to axons in an en passant configuration.Chemoresponsive neurons were identified using “plantmix” (PM) as a search stimulus. PM is an equimolar so-lution of five compounds (D-galactose, D-glucose,D-maltose, D-sucrose, and L-serine) that occur in the nat-ural food of fiddler crabs (Darley, 1977). Neurons werepresented with PM at concentrations from 10–5 to 10–1 M,in ascending order, with a 1 min interstimulus interval.Signals were recorded and amplified using conventionalAC differential techniques and digitally stored with com-mercial software (DataWave; Longmount Co.). Recordingstypically contained spike waveforms from one to threecells, which were sorted off line into individual units onthe basis of various waveform features (rising and fallingslope, amplitude, width) using the DataWave software
package. The numbers of spikes were quantified over 4.5seconds following stimulus introduction and corrected forresponses to the ASW control. These techniques were thesame as those employed in earlier studies with this ani-mal (Weissburg and Derby, 1995; Weissburg, 1999), whichfacilitated direct comparisons with the physiological prop-erties of normal male and female chemosensory neurons.
Behavior
A syringe equipped with an 18-gauge needle was used todeliver a single drop of chemical stimulus onto the distaltip of the feeding claw in order to elicit feeding. Five toseven drops (approximately 10 ml each) of a given stimuluswere delivered alternately to each claw while the behaviorof animals was videotaped under dim red light. Stimuliwere ASW and PM at doses from 10–3 to 10–1 M, presentedin random order. The interstimulus interval was 1minute, contingent on the requirement that animals ex-hibited no feeding activity for 30 seconds prior to stimu-lation. Sensitivity of each claw was assessed by reviewingthe video records and scoring the frequency with which agiven stimulus elicited feeding, defined as occurring whenthe animal lifted either claw from the sediment surface toits mouth. A given stimulus presentation was excludedfrom analysis if the video records revealed that the stim-ulus did not arrive squarely on the distal tip of the claw, orif feeding occurred more than 10 seconds after stimula-tion. These occurrences were rare, and we selected thefirst five stimulations that met our criteria for statisticalanalysis. An individual who was not aware of the presen-tation order of the stimulus intensities analyzed the videorecords. The response frequency across individuals cannotbe normally distributed because the resulting frequenciesfall into a small number of discrete states (e.g., 0, 0.2, 0.4,etc.). Consequently, these data cannot be analyzed byanalysis of variance. However, we can account for thepairwise nature of the design by using log-likelihood teststo examine the frequency of responses for an animal’snative vs. transplanted side for each experimental group(e.g., animals receiving feeding claws from male or femaledonors).
RESULTS
Morphology of transplanted claws
Developing limb buds transplanted to the site of a malemajor claw generated limbs morphologically indistin-guishable from normal feeding claws on the first or second
TABLE 1. Summary Properties of the Various Types of Transplanted Claws Generated in This Study1
Treatment group N Description Physiological and behavioral properties
Successful transplants 101 Feeding claws from male or female donors that developinto feeding claws on the first or second postsurgicalmolt when transplanted to the host major claw site
Contain chemosensory neurons and elicit feeding;sensitivity controlled by donor sex
Unsuccessful transplants 31 Feeding claws from male or female donors that fail todevelop into a feeding claw when transplanted to thehost major claw site, such that the male regenerates amajor claw
No physiological sensitivity detected; do not elicitfeeding behavior
Transplant controls 26 Autotomized major claws removed again during the limbbud stage and then allowed to regenerate
No physiological sensitivity detected; do not elicitfeeding behavior
Regenerating male claws 16 Autotomized major claws allowed to regenerate naturally No physiological sensitivity detected; do not elicitfeeding behavior
Native claws — The feeding claw (one in males, two in females) normallypresent in adult crabs
Contain chemosensory neurons and elicit feeding;properties sex-specific
1N, number of claws produced. The nomenclature used for treatment group represents the terms used throughout the text to refer to claws of a particular type.
313TRANSSEXUAL LIMB TRANSPLANTS IN UCA
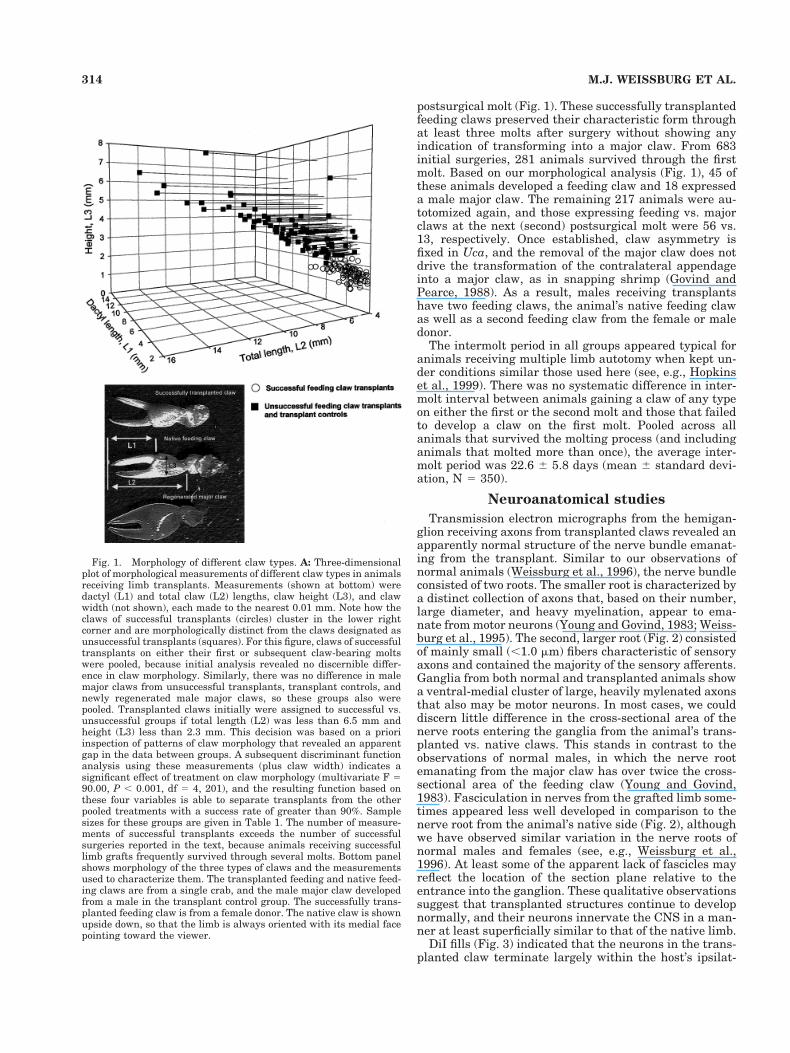
postsurgical molt (Fig. 1). These successfully transplantedfeeding claws preserved their characteristic form throughat least three molts after surgery without showing anyindication of transforming into a major claw. From 683initial surgeries, 281 animals survived through the firstmolt. Based on our morphological analysis (Fig. 1), 45 ofthese animals developed a feeding claw and 18 expresseda male major claw. The remaining 217 animals were au-totomized again, and those expressing feeding vs. majorclaws at the next (second) postsurgical molt were 56 vs.13, respectively. Once established, claw asymmetry isfixed in Uca, and the removal of the major claw does notdrive the transformation of the contralateral appendageinto a major claw, as in snapping shrimp (Govind andPearce, 1988). As a result, males receiving transplantshave two feeding claws, the animal’s native feeding clawas well as a second feeding claw from the female or maledonor.
The intermolt period in all groups appeared typical foranimals receiving multiple limb autotomy when kept un-der conditions similar those used here (see, e.g., Hopkinset al., 1999). There was no systematic difference in inter-molt interval between animals gaining a claw of any typeon either the first or the second molt and those that failedto develop a claw on the first molt. Pooled across allanimals that survived the molting process (and includinganimals that molted more than once), the average inter-molt period was 22.6 6 5.8 days (mean 6 standard devi-ation, N 5 350).
Neuroanatomical studies
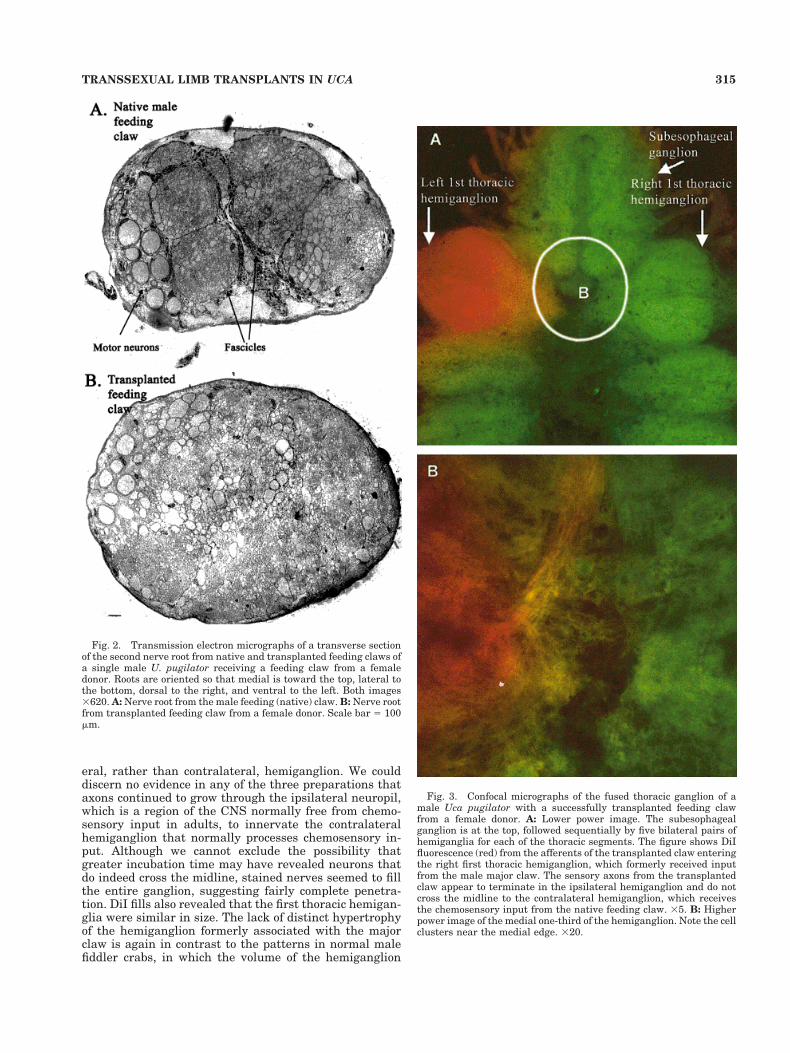
Transmission electron micrographs from the hemigan-glion receiving axons from transplanted claws revealed anapparently normal structure of the nerve bundle emanat-ing from the transplant. Similar to our observations ofnormal animals (Weissburg et al., 1996), the nerve bundleconsisted of two roots. The smaller root is characterized bya distinct collection of axons that, based on their number,large diameter, and heavy myelination, appear to ema-nate from motor neurons (Young and Govind, 1983; Weiss-burg et al., 1995). The second, larger root (Fig. 2) consistedof mainly small (,1.0 mm) fibers characteristic of sensoryaxons and contained the majority of the sensory afferents.Ganglia from both normal and transplanted animals showa ventral-medial cluster of large, heavily mylenated axonsthat also may be motor neurons. In most cases, we coulddiscern little difference in the cross-sectional area of thenerve roots entering the ganglia from the animal’s trans-planted vs. native claws. This stands in contrast to theobservations of normal males, in which the nerve rootemanating from the major claw has over twice the cross-sectional area of the feeding claw (Young and Govind,1983). Fasciculation in nerves from the grafted limb some-times appeared less well developed in comparison to thenerve root from the animal’s native side (Fig. 2), althoughwe have observed similar variation in the nerve roots ofnormal males and females (see, e.g., Weissburg et al.,1996). At least some of the apparent lack of fascicles mayreflect the location of the section plane relative to theentrance into the ganglion. These qualitative observationssuggest that transplanted structures continue to developnormally, and their neurons innervate the CNS in a man-ner at least superficially similar to that of the native limb.
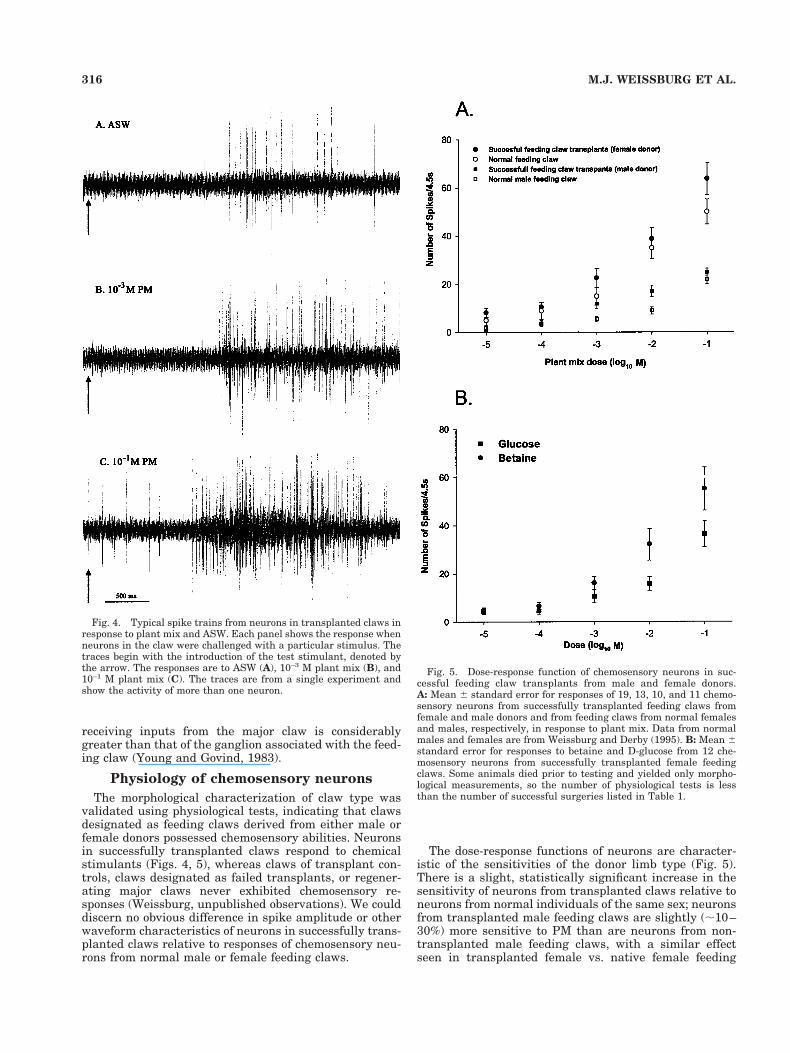
DiI fills (Fig. 3) indicated that the neurons in the trans-planted claw terminate largely within the host’s ipsilat-
Fig. 1. Morphology of different claw types. A: Three-dimensionalplot of morphological measurements of different claw types in animalsreceiving limb transplants. Measurements (shown at bottom) weredactyl (L1) and total claw (L2) lengths, claw height (L3), and clawwidth (not shown), each made to the nearest 0.01 mm. Note how theclaws of successful transplants (circles) cluster in the lower rightcorner and are morphologically distinct from the claws designated asunsuccessful transplants (squares). For this figure, claws of successfultransplants on either their first or subsequent claw-bearing moltswere pooled, because initial analysis revealed no discernible differ-ence in claw morphology. Similarly, there was no difference in malemajor claws from unsuccessful transplants, transplant controls, andnewly regenerated male major claws, so these groups also werepooled. Transplanted claws initially were assigned to successful vs.unsuccessful groups if total length (L2) was less than 6.5 mm andheight (L3) less than 2.3 mm. This decision was based on a prioriinspection of patterns of claw morphology that revealed an apparentgap in the data between groups. A subsequent discriminant functionanalysis using these measurements (plus claw width) indicates asignificant effect of treatment on claw morphology (multivariate F 590.00, P , 0.001, df 5 4, 201), and the resulting function based onthese four variables is able to separate transplants from the otherpooled treatments with a success rate of greater than 90%. Samplesizes for these groups are given in Table 1. The number of measure-ments of successful transplants exceeds the number of successfulsurgeries reported in the text, because animals receiving successfullimb grafts frequently survived through several molts. Bottom panelshows morphology of the three types of claws and the measurementsused to characterize them. The transplanted feeding and native feed-ing claws are from a single crab, and the male major claw developedfrom a male in the transplant control group. The successfully trans-planted feeding claw is from a female donor. The native claw is shownupside down, so that the limb is always oriented with its medial facepointing toward the viewer.
314 M.J. WEISSBURG ET AL.
eral, rather than contralateral, hemiganglion. We coulddiscern no evidence in any of the three preparations thataxons continued to grow through the ipsilateral neuropil,which is a region of the CNS normally free from chemo-sensory input in adults, to innervate the contralateralhemiganglion that normally processes chemosensory in-put. Although we cannot exclude the possibility thatgreater incubation time may have revealed neurons thatdo indeed cross the midline, stained nerves seemed to fillthe entire ganglion, suggesting fairly complete penetra-tion. DiI fills also revealed that the first thoracic hemigan-glia were similar in size. The lack of distinct hypertrophyof the hemiganglion formerly associated with the majorclaw is again in contrast to the patterns in normal malefiddler crabs, in which the volume of the hemiganglion
Fig. 2. Transmission electron micrographs of a transverse sectionof the second nerve root from native and transplanted feeding claws ofa single male U. pugilator receiving a feeding claw from a femaledonor. Roots are oriented so that medial is toward the top, lateral tothe bottom, dorsal to the right, and ventral to the left. Both images3620. A: Nerve root from the male feeding (native) claw. B: Nerve rootfrom transplanted feeding claw from a female donor. Scale bar 5 100mm.
Fig. 3. Confocal micrographs of the fused thoracic ganglion of amale Uca pugilator with a successfully transplanted feeding clawfrom a female donor. A: Lower power image. The subesophagealganglion is at the top, followed sequentially by five bilateral pairs ofhemiganglia for each of the thoracic segments. The figure shows DiIfluorescence (red) from the afferents of the transplanted claw enteringthe right first thoracic hemiganglion, which formerly received inputfrom the male major claw. The sensory axons from the transplantedclaw appear to terminate in the ipsilateral hemiganglion and do notcross the midline to the contralateral hemiganglion, which receivesthe chemosensory input from the native feeding claw. 35. B: Higherpower image of the medial one-third of the hemiganglion. Note the cellclusters near the medial edge. 320.
315TRANSSEXUAL LIMB TRANSPLANTS IN UCA
receiving inputs from the major claw is considerablygreater than that of the ganglion associated with the feed-ing claw (Young and Govind, 1983).
Physiology of chemosensory neurons
The morphological characterization of claw type wasvalidated using physiological tests, indicating that clawsdesignated as feeding claws derived from either male orfemale donors possessed chemosensory abilities. Neuronsin successfully transplanted claws respond to chemicalstimulants (Figs. 4, 5), whereas claws of transplant con-trols, claws designated as failed transplants, or regener-ating major claws never exhibited chemosensory re-sponses (Weissburg, unpublished observations). We coulddiscern no obvious difference in spike amplitude or otherwaveform characteristics of neurons in successfully trans-planted claws relative to responses of chemosensory neu-rons from normal male or female feeding claws.
The dose-response functions of neurons are character-istic of the sensitivities of the donor limb type (Fig. 5).There is a slight, statistically significant increase in thesensitivity of neurons from transplanted claws relative toneurons from normal individuals of the same sex; neuronsfrom transplanted male feeding claws are slightly (;10–30%) more sensitive to PM than are neurons from non-transplanted male feeding claws, with a similar effectseen in transplanted female vs. native female feeding
Fig. 4. Typical spike trains from neurons in transplanted claws inresponse to plant mix and ASW. Each panel shows the response whenneurons in the claw were challenged with a particular stimulus. Thetraces begin with the introduction of the test stimulant, denoted bythe arrow. The responses are to ASW (A), 10–3 M plant mix (B), and10–1 M plant mix (C). The traces are from a single experiment andshow the activity of more than one neuron.
Fig. 5. Dose-response function of chemosensory neurons in suc-cessful feeding claw transplants from male and female donors.A: Mean 6 standard error for responses of 19, 13, 10, and 11 chemo-sensory neurons from successfully transplanted feeding claws fromfemale and male donors and from feeding claws from normal femalesand males, respectively, in response to plant mix. Data from normalmales and females are from Weissburg and Derby (1995). B: Mean 6standard error for responses to betaine and D-glucose from 12 che-mosensory neurons from successfully transplanted female feedingclaws. Some animals died prior to testing and yielded only morpho-logical measurements, so the number of physiological tests is lessthan the number of successful surgeries listed in Table 1.
316 M.J. WEISSBURG ET AL.
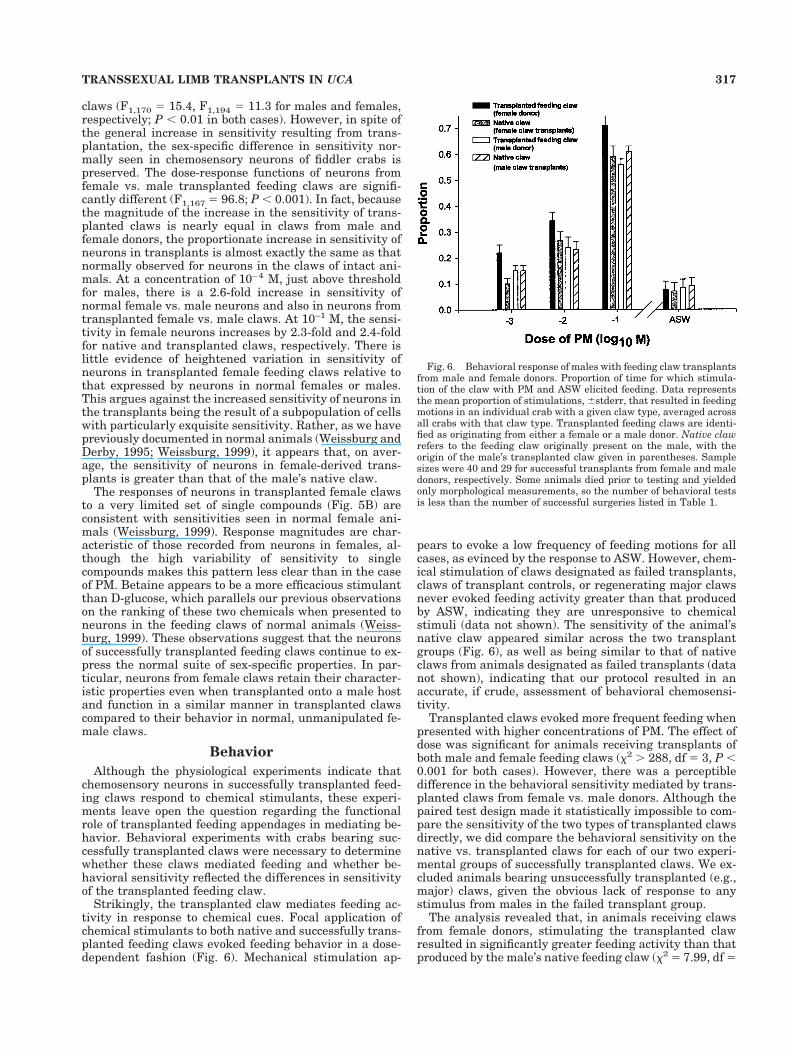
claws (F1,170 5 15.4, F1,194 5 11.3 for males and females,respectively; P , 0.01 in both cases). However, in spite ofthe general increase in sensitivity resulting from trans-plantation, the sex-specific difference in sensitivity nor-mally seen in chemosensory neurons of fiddler crabs ispreserved. The dose-response functions of neurons fromfemale vs. male transplanted feeding claws are signifi-cantly different (F1,167 5 96.8; P , 0.001). In fact, becausethe magnitude of the increase in the sensitivity of trans-planted claws is nearly equal in claws from male andfemale donors, the proportionate increase in sensitivity ofneurons in transplants is almost exactly the same as thatnormally observed for neurons in the claws of intact ani-mals. At a concentration of 10–4 M, just above thresholdfor males, there is a 2.6-fold increase in sensitivity ofnormal female vs. male neurons and also in neurons fromtransplanted female vs. male claws. At 10–1 M, the sensi-tivity in female neurons increases by 2.3-fold and 2.4-foldfor native and transplanted claws, respectively. There islittle evidence of heightened variation in sensitivity ofneurons in transplanted female feeding claws relative tothat expressed by neurons in normal females or males.This argues against the increased sensitivity of neurons inthe transplants being the result of a subpopulation of cellswith particularly exquisite sensitivity. Rather, as we havepreviously documented in normal animals (Weissburg andDerby, 1995; Weissburg, 1999), it appears that, on aver-age, the sensitivity of neurons in female-derived trans-plants is greater than that of the male’s native claw.
The responses of neurons in transplanted female clawsto a very limited set of single compounds (Fig. 5B) areconsistent with sensitivities seen in normal female ani-mals (Weissburg, 1999). Response magnitudes are char-acteristic of those recorded from neurons in females, al-though the high variability of sensitivity to singlecompounds makes this pattern less clear than in the caseof PM. Betaine appears to be a more efficacious stimulantthan D-glucose, which parallels our previous observationson the ranking of these two chemicals when presented toneurons in the feeding claws of normal animals (Weiss-burg, 1999). These observations suggest that the neuronsof successfully transplanted feeding claws continue to ex-press the normal suite of sex-specific properties. In par-ticular, neurons from female claws retain their character-istic properties even when transplanted onto a male hostand function in a similar manner in transplanted clawscompared to their behavior in normal, unmanipulated fe-male claws.
Behavior
Although the physiological experiments indicate thatchemosensory neurons in successfully transplanted feed-ing claws respond to chemical stimulants, these experi-ments leave open the question regarding the functionalrole of transplanted feeding appendages in mediating be-havior. Behavioral experiments with crabs bearing suc-cessfully transplanted claws were necessary to determinewhether these claws mediated feeding and whether be-havioral sensitivity reflected the differences in sensitivityof the transplanted feeding claw.
Strikingly, the transplanted claw mediates feeding ac-tivity in response to chemical cues. Focal application ofchemical stimulants to both native and successfully trans-planted feeding claws evoked feeding behavior in a dose-dependent fashion (Fig. 6). Mechanical stimulation ap-
pears to evoke a low frequency of feeding motions for allcases, as evinced by the response to ASW. However, chem-ical stimulation of claws designated as failed transplants,claws of transplant controls, or regenerating major clawsnever evoked feeding activity greater than that producedby ASW, indicating they are unresponsive to chemicalstimuli (data not shown). The sensitivity of the animal’snative claw appeared similar across the two transplantgroups (Fig. 6), as well as being similar to that of nativeclaws from animals designated as failed transplants (datanot shown), indicating that our protocol resulted in anaccurate, if crude, assessment of behavioral chemosensi-tivity.
Transplanted claws evoked more frequent feeding whenpresented with higher concentrations of PM. The effect ofdose was significant for animals receiving transplants ofboth male and female feeding claws (x2 . 288, df 5 3, P ,0.001 for both cases). However, there was a perceptibledifference in the behavioral sensitivity mediated by trans-planted claws from female vs. male donors. Although thepaired test design made it statistically impossible to com-pare the sensitivity of the two types of transplanted clawsdirectly, we did compare the behavioral sensitivity on thenative vs. transplanted claws for each of our two experi-mental groups of successfully transplanted claws. We ex-cluded animals bearing unsuccessfully transplanted (e.g.,major) claws, given the obvious lack of response to anystimulus from males in the failed transplant group.
The analysis revealed that, in animals receiving clawsfrom female donors, stimulating the transplanted clawresulted in significantly greater feeding activity than thatproduced by the male’s native feeding claw (x2 5 7.99, df 5
Fig. 6. Behavioral response of males with feeding claw transplantsfrom male and female donors. Proportion of time for which stimula-tion of the claw with PM and ASW elicited feeding. Data representsthe mean proportion of stimulations, 6stderr, that resulted in feedingmotions in an individual crab with a given claw type, averaged acrossall crabs with that claw type. Transplanted feeding claws are identi-fied as originating from either a female or a male donor. Native clawrefers to the feeding claw originally present on the male, with theorigin of the male’s transplanted claw given in parentheses. Samplesizes were 40 and 29 for successful transplants from female and maledonors, respectively. Some animals died prior to testing and yieldedonly morphological measurements, so the number of behavioral testsis less than the number of successful surgeries listed in Table 1.
317TRANSSEXUAL LIMB TRANSPLANTS IN UCA

1, P , 0.05). Corrected for response to the ASW controls,the magnitude of this increase ranged from nearly three-fold at 10–3 M to 1.20-fold at 10–1 M. Individual compari-sons indicated that the transsexually grafted feedingclaws evoked significantly greater feeding activity at allconcentrations of PM (maximum likelihood G . 6.1, df 51, P , 0.05, for all comparisons), but not when ASW wasthe stimulus (G 5 4.9, df 5 1, P . 0.05). In contrast, wecould detect no increase in behavioral sensitivity for ani-mals receiving transplants of male feeding claws com-pared to the sensitivity of the male’s native side (x2 50.28, df 5 1, P . 0.05). That is, the transplantation of afemale claw specifically and significantly increased thesensitivity of the behavioral response to chemical stimu-lants relative to the sensitivity evoked by the male’s na-tive feeding claw.
DISCUSSION
Our experiments indicate that the natural regenerativecapacities of crustaceans result in the ability to equip amale crab with additional chemosensory feeding append-ages, rather than their normal complement of one. Devel-oping limb buds from either male or female donors can besuccessfully grafted to males in place of their major claws,to equip males with a second chemosensory feeding claw(Fig. 1). Basic patterns of neuroanatomy, including thelack of bilateral asymmetry in the size of the first thoracicganglion (Figs. 2, 3), also are consistent with the proper-ties of the feeding claw, rather than that of the major clawthat formerly occupied the transplant site. The trans-planted feeding claw appears to maintain its physiologicalcompetence and sex-specific properties (Figs. 4, 5). Thisindicates that transplanted limbs seem to be largely au-tonomous and to legislate their own development even ina foreign developmental context. Ingrowing chemosensoryaxons from transplanted feeding claws make functionalconnections in the CNS normally devoted to processingnonchemosensory input emanating from the male’s majorclaw and, instead, evoke feeding behavior (Fig. 6). Thus,even though the transplanted neurons project to a neuro-pil that does not appear to process chemosensory input,their interactions within this foreign neuropil are suffi-cient to drive the appropriate behavior. In fact, sex-specific sensitivities of neurons in the transplanted feed-ing claw translate into greater behavioral sensitivity ofmales receiving transsexual limb transplants than that ofmales receiving transplants from a male donor (Fig. 6).
Reorganization underlying transplantation
Several transplant studies have suggested that neuronstransplanted to foreign locations successfully navigate tothe CNS, presumably using molecular markers to findtheir way. Proprio- and tactile receptors can be induced toproject to their segmentally homologous neuropil whentransplanted ectopically to a foreign location (Killian etal., 1993). In moths receiving transsexual antennal grafts,sensory axons maintain their sex-specific sensory projec-tions and form sex-specific olfactory glomeruli in the sameposition that they occupy in normal animals (Rossler etal., 1999). These and other experiments in vertebrates andinvertebrates (see, e.g., Sur et al., 1990; Passani et al.,1991) suggest that positional cues are often available,even if not specifically needed, to mediate pathfinding of aparticular group of axons during normal development.
Similarly, ingrowing chemosensory axons from trans-planted claws in fiddler crabs appear to experience appro-priate molecular markers and terminate in the ipsilateralganglion even though this area of neuropil does not nor-mally accommodate this modality.
Once within this ganglion, chemosensory axons estab-lish connections sufficient to drive appropriate chemosen-sory behavior. Ectopically transplanted neurons oftenmaintain largely normal projection patterns in foreignneuropil (Murphey et al., 1983; Edwards, 1988). The fidel-ity with which neurons arborize in particular spatial re-gions of the ganglion allows them to form synapses withneurons with which they normally associate even thoughthese connections are established in an unfamiliar neuralsubstrate (Killian et al., 1993). Occasionally, transplantedneurons form novel connections with different partnersthat have dendrites in the same region where the trans-planted neurons arborize but which are not found in theneuropil normally occupied by the transplants (Murpheyet al., 1983). This suggests that particular patterns ofconnectivity that underlie specific neural functioning mayoccur even when neurons project to a novel region of theCNS, although the behavioral consequences of these con-nections have been largely unexplored. Clearly, trans-planted chemosensory axons in fiddler crabs are able toform functional connections sufficient to produce normalbehavior even when induced to grow into a different hemi-ganglion.
We interpret the behavioral experiments as also sug-gesting that novel sensory capabilities may be conferredon CNS elements when innervated by sensory neurons notnormally associated with a given neuropil. In fiddlercrabs, receptor-rich feeding claws are able to drive feedingbehavior when transplanted to the site of mechanosensorymale major claws. Cross-modal connections have beenproduced in other systems, indicating that sensory neu-rons may utilize novel targets when deprived of theirnormal postsynaptic partners (Sur et al., 1990; Scalia etal., 1995; Yaka et al., 1999). However, to our knowledge,this is the first demonstration that CNS elements withrewired connections can successfully process sensory stim-uli resulting in behavioral discrimination of stimulus in-tensity.
The supposition that our transplant experiments repre-sent cross-modal reorganization is based on strong evi-dence that the hemiganglion receiving transplanted che-mosensory axons does not participate in chemosensoryprocessing. This is certainly the case in males over themajority of their lives. A variety of evidence also suggeststhat, in spite of the fact that the claw dimorphism isinducible, the default condition of this neuropil is to pro-cess input from the nonchemosensory major claw inadults. Prior to the development of the claw asymmetry inmales, which occurs in 1–2 mm animals recently moltedfrom their oceanic larval phase, both claws are major.Damage to one of the claws during this stage sets theasymmetry; the damaged claw develops into a feedingclaw on the subsequent molt (Morgan, 1924; Yamaguchi,1977).
Indeed, we very rarely have seen small juveniles thathave fortuitously escaped damage to either claw and con-sequently sport two major claws rather than the tradi-tional dimorphic arrangement. In such animals, neither oftheir major claws appears to be chemosensory based onphysiological or behavioral assays (Weissburg, unpub-
318 M.J. WEISSBURG ET AL.

lished observations). These males probably use chemore-ceptors on the tips of their walking legs (Weissburg andDerby, 1995) to sense chemical cues. Even if the hemigan-glia innervated by the major claw of adults possess relicsassociated with chemosensory input from prior life stages,reorganization of the thoracic ganglion by ingrowing che-mosensory axons from the transplanted feeding clawwould be necessary.
The reduction in size of hemiganglia associated withtransplanted feeding claws, but that previously receivedinput from major claws, is the most immediate conse-quence of the changes in the neuronal properties resultingfrom limb transplantation. There are other potentialchanges as well, which are associated with a variety ofdifferences in the organization of these two claw types. Forinstance, major and feeding claws differ significantly inthe number of axons and the size-frequency distribution oftheir axonal population impinging on the CNS. Axonswith diameters greater than 1 mm make up approximately69% vs. 85% of the population projecting to the major vs.feeding claw hemiganglia, respectively (Young and Gov-ind, 1983; Weissburg et al., 1996). The diameters of motorneuron axons and somata are 30–60% larger in majorthan in feeding claws (Young and Govind, 1983). Finally,the modality-specific patterns of organization repeatedlyobserved in many other animals argue that ingrowingchemosensory axons would arborize in regions differentfrom those of the mechanosensors that normally carrysensory information into the major claw hemiganglion.
Peripheral vs. central effects in theproduction of sex-specific behavior
Successfully transplanted feeding claws not only aremorphologically indistinguishable from normal feedingclaws but contain sensory neurons that remain faithful totheir donor origin, even when males receive transsexuallimb grafts. In other invertebrates, individual sensoryneurons (Possidente and Murphey, 1989; Passani et al.,1991) or muscle types (Trinkaus-Randall and Mittenthal,1978) that are sexually dimorphic retain their propertiesin transplantation experiments, although experimentalproduction of sexually dimorphic appendages appearsrare. The physiological experiments showing that sensorylimbs maintain the sensitivity characteristic of the donorare consistent with the greater behavioral sensitivity ex-pressed by males receiving feeding claws from female, butnot male, donors. Using a common CNS substrate, themore sensitive chemoreceptors on the transsexuallygrafted feeding claw evoked more frequent feeding behav-ior. This implies that, regardless of any differences in CNSelements between males and females, changes in the prop-erties of the periphery may directly modulate quantitativesex-specific differences in behavioral chemosensitivity.
The ability of chemosensory systems to regain functionfollowing tissue transplantation appears robust, in thatexperiments in both vertebrates (Hendricks et al., 1994;Goheen et al., 1995) and invertebrates (Schneiderman etal., 1986; Murphey et al., 1983; Rossler et al., 1999) showthat chemosensory neurons reestablish connections in ol-factory neuropil following transplantation and mediateolfactory-induced behaviors. The attraction of male mothsto female sex attractants has been successfully recapitu-lated in females by transplantation of male antennal disksinto developing females (Schneiderman et al., 1986; Williset al., 1995). This is similar to our results; both fiddler
crabs and moths show induction of sex specificity viatransplantation of peripheral chemosensory structures.However, unlike the case in fiddler crabs, neurons in bothnative and transplanted antennae of moths project into achemosensory neuropil, whereas neurons in the trans-planted claw of fiddler crabs terminate in a sexually di-morphic region of the CNS that does not receive chemicalinput in normal (adult) crabs prior to the limb graft. Theextent to which this sex specificity relates to the intrinsicproperties of the sensory neurons, rather than potentialsex-specific patterns of organization in the CNS as a resultof interactions with these sensory neurons, remains to beexplored.
Neural plasticity and sensory processing
Neuronal guidance and target selection are presumed toreflect mechanisms necessary to ensure that precise andstereotyped synaptic connections are established, creatingthe neural architecture required for normal operation ofthe CNS. The chemosensory axons in transplanted feedingclaws of fiddler crabs may be able to reorganize localcircuits in the neuropil that normally receive input fromthe major claw, such that the ability to process chemosen-sory signals emerges. Such plasticity has been suggestedfor other systems (Sur et al., 1990; Yaka et al., 1999).However, it has also been argued that processing deficitsarise from patterns of neural architecture that are insuf-ficient for particular tasks but cannot be modified by in-vasion of novel sensory afferents (Scalia et al., 1995).
Alternatively, such a precise pattern of connections maybe unnecessary for the relatively simple process of inten-sity discrimination, and existing circuitry may be suffi-cient. Experiments in moths have suggested that the nor-mal glomerular organization of olfactory neuropil is notnecessary for upwind flight in response to odor plumesand that this motor program can be triggered in moths ofreduced anatomical complexity (Willis et al., 1995). Ratsrecover olfactory ability following transplantation, al-though they are not as proficient at olfactory location ofitems as are normal animals, perhaps because transplan-tation does not result in completely natural patterns ofconnectivity and levels of glomerulization (Goheen et al.,1995; Evers et al., 1996). We are eager to see whether thetypical pattern of neuropil organization is maintained infiddler crabs receiving transplanted appendages andwhether this impacts on more challenging tasks, such asdiscrimination of odor qualities.
ACKNOWLEDGMENTS
We thank H.S. Cate and M. Mulkeen for their technicalassistance at various phases of this study, P.J.H. Harrisonfor confocal microscopy, and C.K Govind for electron mi-croscopy. Comments by P. Steullet, S. Cromarty, and S.Pallas greatly improved this work. We thank J. Thompsonfor his prodigious memory and P.M. Hopkins for advice onthe induction of molting. Two anonymous reviewers pro-vided comments that helped to improve the text. Thesestudies were supported by NIH grants to M.J.W.(DC02731) and C.D.D. (DC00312).
LITERATURE CITED
Atema J, Fay RR, Popper AN, Tavolga WN. 1988. Sensory biology ofaquatic animals. New York: Springer-Verlag.
319TRANSSEXUAL LIMB TRANSPLANTS IN UCA

Crane J. 1975. Fiddler crabs of the world (Ocypodidae: genus Uca). Prince-ton, NJ: Princeton University Press.
Darley WM. 1977. Biochemical composition of diatoms. In: Werner D,editor. Biology of diatoms. Oxford: Blackwell Scientific. p 3–23.
Edwards JS. 1988. Sensory regeneration in arthropods: implications ofhomoeosis and of ectopic sensilla. Am Zool 28:1155–1164.
Evers SM, Hendricks KR, Kott JN, Lee ME, Gregov ME, Westrum LE.1996. Recovery of olfactory function in thirteen-day-old rats after ol-factory bulb transplantation but not after olfactory bulb ablation. BrainRes 724:129–135.
Goheen BL, Kott JN, Anderson NL, Kim A, Westrum LE. 1995. Hostprimary axons make synaptic contacts in transplanted olfactory bulb.J Comp Neurol 352:203–212.
Goldsmith TL. 1991. Photoreception and vision. In: Prosser CL, editor.Neural and integrative animal physiology. New York: Wiley-Liss, Inc.p 171–246.
Govind CK, Pearce J. 1988. Remodeling of nerves during claw reversal inadult snapping shrimp. J Comp Neurol 268:121–130.
Hayama T, Caprio JC. 1989. Lobule structure and somatotopic organiza-tion of the medullary facial lobe in the channel catfish Ictalurus punc-tatus. J Comp Neurol 285:9–17.
Hendricks KR, Kott JN, Gooden MD, Lee ME, Evers SM, Goheen BL,Westrum LE. 1994. Recovery of olfactory behavior. II. Neonatal olfac-tory bulb transplants enhance the rate of behavioral recovery. BrainRes 648:135–147.
Hopkins PM, Chung AC-K, Durica DS. 1999. Limb regeneration in thefiddler crab, Uca pugilator: histological, physiological and molecularconsiderations. Am Zool 39:513–526.
Killian KA, Merritt DJ, Murphey RK. 1993. Transplantation of neuronsreveals processing areas and rules for synaptic connectivity in thecricket nervous system. J Neurobiol 24:1187–1206.
Konishi M. 1986. Centrally synthesized maps of sensory space. TrendsNeurosci 9:163–168.
Lehrer M. 1997. Orientation and communication in Arthropods. Basel:Birkhauser Verlag.
Meisami E. 1991. Chemoreception. In: Prosser CL, editor. Neural andintegrative animal physiology. New York: Wiley-Liss, Inc. p 335–434.
Morgan TH. 1924. The artificial induction of symmetrical claws in malefiddler crabs. Am Naturalist 58:289–295.
Murphey RK, Bacon JP, Sakaguchi DS, Johnson SE. 1983. Transplanta-tion of cricket sensory neurons to ectopic locations: arborizations andsynaptic connections. J Neurosci 3:659–672.
Murphey RK, Possidente D, Pollack G, Merritt DJ. 1989. Modality specificaxonal projections in the flies Phormia and Drosophila. J Comp Neurol290:185–200.
Newland PL, Rogers SM, Gaaboub I, Matheson T. 2000. Parallel somato-topic maps of gustatory and mechanosensory nerons in the centralnervous system of an insect. J Comp Neurol 425:82–96.
Passani MB, Peinado A, Engleman H, Baptista CA, Macagno E. 1991.Normally unused positional cues guide ectopic afferents in the leechCNS. J Neurosci 11:3868–3876.
Possidente DR, Murphey RK. 1989. Genetic control of sexually dimorphic
axon morphology in Drosophila sensory neurons. Dev Biol 132:448–457.
Robertson JR, Fudge JA, Vermeer GK. 1981. Chemical and live feedingstimulants of the sand fiddler crab Uca pugilator (Bosc, 1802). J ExpMar Biol Ecol 53:47–64.
Rossler W, Randolph PW, Tolbert LP, Hildebrand JG. 1999. Axons ofolfactory receptor cells of transexually grafted antennae induce devel-opment of sexually dimorphic glomerui in Manduca sexta. J Neurobiol38:521–541.
Sandeman DC, Sandeman RE. 1994. Electrical responses and synapticconnection of giant serotonin-immunoreactive neurons in crayfish ol-factory and accessory lobes. J Comp Neurol 341:130–144.
Scalia F, Grant AC, Reyes M, Lettvin JY. 1995. Functional properties ofregenerating optic axons terminating in the olfactory cortex. Brain Res685:187–197.
Schneiderman AM, Hildebrand JG, Brennan MM, Tumlinson JH. 1986.Trans-sexually grafted antennae alter pheromone directed behavior ina moth. Nature 323:801–803.
Sur M, Pallas SL, Roe AW. 1990. Cross modal plasticity in cortical devel-opment: differentiation and specification of the sensory neocortex.Trends Neurosci 13:227–233.
Trinkaus-Randall V, Mittenthal JE. 1978. Intra- and interspecific trans-plants of limb buds in Crustacea: a new method for studying centraland peripheral interactions crustacean limbs. J Exp Zool 204:275–281.
Valiela I, Babiec DF, Atherton W, Setziger S, Krebs C. 1974. Some conse-quences of sexual dimorphism: feeding in male and female fiddlercrabs. Biol Bull 147:652–660.
Weissburg MJ. 1993. Sex and the single forager: gender-specific energymaximizing strategies in fiddler crabs. Ecology 74:279–291.
Weissburg MJ. 1999. Tuning breadth and sex-specificity in chemosensoryneurons of Uca pugnax. J Comp Physiol 185:229–238.
Weissburg MJ, Derby CD. 1995. Regulation of sex-specific feeding behaviorin fiddler crabs: physiological properties of chemoreceptor neurons inthe claws and legs of males and females. J Comp Physiol A176:513–526.
Weissburg MJ, Zimmer-Faust RK. 1991. Ontogeny vs. phylogeny in deter-mining patterns of chemoreception: initial studies with fiddler crabs.Biol Bull 181:205–215.
Weissburg MJ, Pearce J, Govind CK, Derby CD. 1996. Sexually dimporphicpatterns of neural organization in the feeding appendages of fiddlercrabs. Cell Tissue Res 286:155–166.
Willis MA, Butler MA, Tolbert LP. 1995. Normal glomerular organizationof the olfactory lobe is not necessary for odor guided locomotion. J CompPhysiol A176:205–216.
Yaka R, Yinon U, Wollberg Z. 1999. Auditory activation of cortical visualareas in cats after early visual deprivation. Eur J Neurosci 11:1301–1312.
Yamaguchi T. 1977. Studies in the handedness of the fiddler crab, Ucalactea. Biol Bull 152:424–436.
Young RE, Govind CK. 1983. Neural asymmetry in male fiddler crabs.Brain Res 280:251–262.
Zill SN, Frazier F, Macfarland DL, Fish SE. 1993. Characterization ofinsect sense organs and optical clearing of whole mount preparationsusing diI in fixed tissue. J Exp Biol 175:299–303.
320 M.J. WEISSBURG ET AL.




















