
POSTLARVAL STAGES AND GROWTH PATTERNS OF THE SPIDER CRAB PYROMAIA TUBERCULATA (BRACHYURA, MAJIDAE)...
14
ing the effect of temperature on the develop- ment of the shore crab Carcinus maenas (L., 1758) (see Mohamedeen and Hartnoll, 1989), and had provided evidence of mechanisms controlling size variation in the caridean shrimp Palaemon elegans Rathke, 1837 (see Hartnoll and Dalley, 1981). Provided the stage sequence is covered during the rearing protocol, regressions of log intermolt period and log percentage molt increment against size can be readily obtained and used to ex- plain growth patterns. Because straight lines are regarded to fit those relationships, they should be preferred to the so-called Hiatt growth diagram, in which a hyperbola should be used to fit premolt to postmolt size re- gressions (Mauchline, 1976). Only in some exceptional cases can post- larval molt stages be identified from field samples, either by means of inspecting diag- nostic characters allowing their separation or by attributing different stages to discrete nor- mal components in size-frequency distribu- tions (Childress and Price, 1978; Rice, and Hartnoll, 1983; Hartnoll and Rice, 1986). Hence, raising and examining laboratory-held The determination of growth and age in crustaceans is particularly difficult because molting imposes restrictions to tagging pro- cedures and results in loss of any calcified structures that could be potentially used for the estimation of age (Hartnoll, 1982). Indi- rect methods, namely the treatment of size- frequency distributions for modal progression analyses, are particularly common. However, they only provide good results if recruitment is restricted to a short season and population changes due to migration are minimal (Siegel and Wenner, 1985). Results from laboratory rearing would not probably replicate the growth patterns of shapes found in nature, but they provide in- formation on the sequence of stages, varia- tion of size within each stage, molt increment and intermolt period. Useful growth compar- isons can be derived if captivity conditions are assumed to affect growth independently of sex and developmental phase. Due to prac- tical problems of laboratory rearing, however, long stage sequences are seldom obtained and reported. For decapods, long rearing experi- ments had led to interesting insight regard- JOURNAL OF CRUSTACEAN BIOLOGY , 22(2): 314–327, 2002 POSTLARVAL STAGES AND GROWTH PATTERNS OF THE SPIDER CRAB PYROMAIA TUBERCULATA (BRACHYURA, MAJIDAE) FROM LABORATORY-REARED MATERIAL Augusto A. V. Flores, Fernando P. L. Marques, and Maria Lucia Negreiros-Fransozo (AAVF, corresponding author) Instituto do Mar, Laboratório Marítimo da Guia, Faculdade de Ciências da Universidade de Lisboa, Estrada do Guincho, 2750-450, Cascais, Portugal (e-mail: afl[email protected]); (FPLM, MLNF) NEBECC (Group of Studies on Crustacean Biology, Ecology and Culture), Departamento de Zoologia, Instituto de Biociências and Centro de Aquicultura, Universidade Estadual Paulista, UNESP, 18618-000 Botucatu (São Paulo), Brazil ABSTRACT The spider crab Pyromaia tuberculata was introduced into southeastern Brazil; ovigerous mate- rial was collected and reared in the laboratory. Morphologic changes and growth patterns of post- larval development are reported. Results show that within-stage size variation is lowest in mature stages, especially in the case of females in which there is an apparent size threshold for the last ju- venile stages to undergo the puberty molt. A prepuberty molt taking place at the fourth crab stage is indicated by analyzing the allometric growth of the abdomen in females. In contrast, the same procedure using the allometric growth of chelae failed in detecting both the prepuberty and pu- berty molts in males. Conversely to females, which develop a complex brood chamber at the pu- berty molt, the enlargement of chelae was not consistent in all postpuberty males. The short instar sequence of this species, in no case exceeding nine stages, is marked by conspicuous morphologic alterations achieved at each molt. Almost all stages can be identified by examining diagnostic fea- tures of rostrum, abdomen, sternum, and pleopods. 314
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of POSTLARVAL STAGES AND GROWTH PATTERNS OF THE SPIDER CRAB PYROMAIA TUBERCULATA (BRACHYURA, MAJIDAE)...

ing the effect of temperature on the develop-ment of the shore crab Carcinus maenas (L.,1758) (see Mohamedeen and Hartnoll, 1989),and had provided evidence of mechanismscontrolling size variation in the carideanshrimp Palaemon elegans Rathke, 1837 (seeHartnoll and Dalley, 1981). Provided thestage sequence is covered during the rearingprotocol, regressions of log intermolt periodand log percentage molt increment againstsize can be readily obtained and used to ex-plain growth patterns. Because straight linesare regarded to fit those relationships, theyshould be preferred to the so-called Hiattgrowth diagram, in which a hyperbola shouldbe used to fit premolt to postmolt size re-gressions (Mauchline, 1976).
Only in some exceptional cases can post-larval molt stages be identified from fieldsamples, either by means of inspecting diag-nostic characters allowing their separation orby attributing different stages to discrete nor-mal components in size-frequency distribu-tions (Childress and Price, 1978; Rice, andHartnoll, 1983; Hartnoll and Rice, 1986).Hence, raising and examining laboratory-held
The determination of growth and age incrustaceans is particularly difficult becausemolting imposes restrictions to tagging pro-cedures and results in loss of any calcifiedstructures that could be potentially used forthe estimation of age (Hartnoll, 1982). Indi-rect methods, namely the treatment of size-frequency distributions for modal progressionanalyses, are particularly common. However,they only provide good results if recruitmentis restricted to a short season and populationchanges due to migration are minimal (Siegeland Wenner, 1985).
Results from laboratory rearing would notprobably replicate the growth patterns ofshapes found in nature, but they provide in-formation on the sequence of stages, varia-tion of size within each stage, molt incrementand intermolt period. Useful growth compar-isons can be derived if captivity conditionsare assumed to affect growth independentlyof sex and developmental phase. Due to prac-tical problems of laboratory rearing, however,long stage sequences are seldom obtained andreported. For decapods, long rearing experi-ments had led to interesting insight regard-
JOURNAL OF CRUSTACEAN BIOLOGY, 22(2): 314–327, 2002
POSTLARVAL STAGES AND GROWTH PATTERNS OF THE SPIDER CRAB PYROMAIA TUBERCULATA (BRACHYURA, MAJIDAE)
FROM LABORATORY-REARED MATERIAL
Augusto A. V. Flores, Fernando P. L. Marques, and Maria Lucia Negreiros-Fransozo
(AAVF, corresponding author) Instituto do Mar, Laboratório Marítimo da Guia, Faculdade de Ciências daUniversidade de Lisboa, Estrada do Guincho, 2750-450, Cascais, Portugal (e-mail: [email protected]);
(FPLM, MLNF) NEBECC (Group of Studies on Crustacean Biology, Ecology and Culture), Departamento de Zoologia, Instituto de Biociências and Centro de Aquicultura,
Universidade Estadual Paulista, UNESP, 18618-000 Botucatu (São Paulo), Brazil
A B S T R A C T
The spider crab Pyromaia tuberculata was introduced into southeastern Brazil; ovigerous mate-rial was collected and reared in the laboratory. Morphologic changes and growth patterns of post-larval development are reported. Results show that within-stage size variation is lowest in maturestages, especially in the case of females in which there is an apparent size threshold for the last ju-venile stages to undergo the puberty molt. A prepuberty molt taking place at the fourth crab stageis indicated by analyzing the allometric growth of the abdomen in females. In contrast, the sameprocedure using the allometric growth of chelae failed in detecting both the prepuberty and pu-berty molts in males. Conversely to females, which develop a complex brood chamber at the pu-berty molt, the enlargement of chelae was not consistent in all postpuberty males. The short instarsequence of this species, in no case exceeding nine stages, is marked by conspicuous morphologicalterations achieved at each molt. Almost all stages can be identified by examining diagnostic fea-tures of rostrum, abdomen, sternum, and pleopods.
314

individuals is the most feasible method toassess stage sequence and to identify stage-specific morphology. This technique has beencarried out in several studies on brachyurancrabs, but in almost all cases descriptive ac-counts are limited to juvenile stages (e.g.,Shen, 1935; Ingle, 1977; Fransozo, 1986/87;Fransozo and Negreiros-Fransozo, 1987;Negreiros-Fransozo and Fransozo, 1991;Rieger and Nakagawa, 1995; Flores et al.,1998). Apart from identifying the differenti-ation of pleopods in early juvenile stages, de-termining the age and at which stage othercritical postlarval events take place may helpto resolve questions related to the life his-tory of these crustaceans.
In spider crabs, patterns of growth and re-production do not vary beyond certain limitsbecause these crabs are known to enter ter-minal anecdysis after the puberty molt. Whilethere is still some uncertainty if males mayreproduce during preceding molt stages, it isclear that females are only capable of breed-ing after undergoing the puberty molt (Hart-noll, 1965). Life history and reproductiveecology of majids may, however, depend onrelevant well-documented variation of thebreeding period (Hartnoll, 1963), reproduc-tive effort (Bryant and Hartnoll, 1995), tim-ing of gonad maturation, and mating system(Jones and Hartnoll, 1997).
The natural geographic range of Pyromaiatuberculata (Lockington, 1877) extends in theeastern Pacific from California, U.S.A., toPanama, but the species has been introducedin Japan (Sakai, 1976), the Indo-West Pacific(Grifffin and Tranter, 1986), Western Aus-tralia (Morgan, 1990), and southeastern Brazil(Melo et al., 1989). Furota (1996a, b) had al-ready investigated the life cycle of P. tuber-culata in the Bay of Tokyo, including theoverall growth pattern of crabs maintained incaptivity under different temperature condi-tions. Yet, morphologic accounts of reared in-dividuals and detailed analysis of molt incre-ment and intermolt period were not conducted.
This present study provides a morpholog-ical account of P. tuberculata postlarvalstages by laboratory rearing of the offspringobtained from ovigerous females collected insoutheastern Brazil. The growth components,molt increment, and intermolt period are ex-amined in detail according to size, and the re-sults discussed taking into consideration criti-cal molts delimiting different growth phases.
MATERIALS AND METHODS
Rearing.—Two ovigerous females of Pyromaia tubercu-lata were collected by trawling at a depth ranging from35 to 40 m in the Bay of Ubatuba, São Paulo, Brazil(23°25′00″S, 45°00′30″W). These specimens were heldin aquaria until hatching, which took place on 20 and 21May, 2000. Larvae were transferred to 700-ml acrylic jarscontaining 500 ml of filtered sea water. Megalopae wereseparated from cultures of zoeae and, as soon as noticed,specimens in first crab stages were removed and held sep-arately in 100-ml acrylic jars containing 50 ml of filteredsea water.
Both larvae and specimens in early juvenile stageswere fed ad libitum with Artemia nauplii. Individuals inlater stages were fed with Artemia nauplii supplementedwith small pieces of shrimp and/or mussels. Sea waterwas changed, and specimens were checked for moltingand fed daily. All acrylic ware was washed in fresh wa-ter and air dried before being used again the followingday. Rearing was conducted in a temperature-controlledroom at 24° ± 1°C. Average salinity was 32‰ and a 12L: 12D photoperiod was maintained. Specimens were heldin captivity until no further molting was observed for aperiod ranging from 28 to 53 d in crabs that had delayedterminal anecdysis to the ninth stage. Considering the in-termolt period (IP) of crabs that had already molted to thefinal stage, such a time interval is certainly long enoughto ensure that remaining crabs would not molt again. Allspecimens were labeled and fixed in 70% ethanol. Lar-vae, crab stages, and a spent female were deposited in theDecapod Larval Collection at the NEBECC (Núcleo deEstudos em Biologia, Ecologia e Cultivo de Crustáceos),Department of Zoology—IB, Universidade EstadualPaulista, Botucatu, São Paulo, Brazil.
Morphometry.—Exuviae were preferred for morphologicexamination, measurements, and illustrations. Measure-ments included chela height (ChH) for males (measuredclose to the insertion of dactylus), maximum abdomenwidth for females (AW), maximum carapace width (CW),and carapace length (CL, including rostrum) for all stages.For this morphometric analysis, 21 to 28 specimens wereexamined for each of the first eight stages. Only six in-dividuals belonging to the ninth crab stage were avail-able. Measurements were recorded with a precision of0.01 mm using a Zeiss stereomicroscope provided with anAxiovision image analysis system. Percentage molt incre-ment (%MI) was calculated as
%MI = [(CW2 – CW1) × 100]/CW1
where CW1 and CW2 are premolt and postmolt sizes, re-spectively.
Morphology.—The morphological changes of the cara-pace, rostrum, sternum, abdomen, pleopods, and malechelae were assessed. For this purpose, a variable num-ber of specimens, at least five, were examined for eachstage and sex. In cases where enhanced contrast and de-tail were needed, namely while observing small devel-oping pleopods and first appearance of gonopores, dis-sected appendages were stained with chlorazol black. De-pending on the size of structures, illustrations wereprepared using either an Olympus BH-2 microscope sup-plied with phase contrast or a Wild stereomicroscope,both provided with a camera lucida.
FLORES ET AL.: POSTLARVAL DEVELOPMENT OF PYROMAIA TUBERCULATA 315

Statistical Procedures.—In order to test whether there isa recognizable prepuberty molt during juvenile develop-ment, the Somerton’s Mature 2 computer program(Somerton, 1983) was used to locate eventual inflectionpoints in ChH vs. CW (for males) and AW vs. CW (forfemales) loge-linearized allometric relationships. Finalstages were not included in this procedure. The analysisof covariance (ANCOVA, Snedecor and Cochran, 1980)was used to test whether linear regression parameters inlog10 IP vs. CW and log10 %MI vs. CW relationships dif-fer between sexes. Student’s t-tests, corrected in the caseof significantly contrasting variances (Sokal and Rohlf,1995), were run for two-sample comparisons.
RESULTS
Morphologic Changes During Postlarval Ontogeny
Carapace.—The ratio CL/CW did not presentsignificant differences in any of the stages;neither were between-sex differences detectedat any given stage (P > 0.05 in all cases). Themost conspicuous change regards rostralshape, which developed from the widely bifidform characteristic of crab stage 1 to the acuterostrum found in crab stage 5 (Fig. 1).
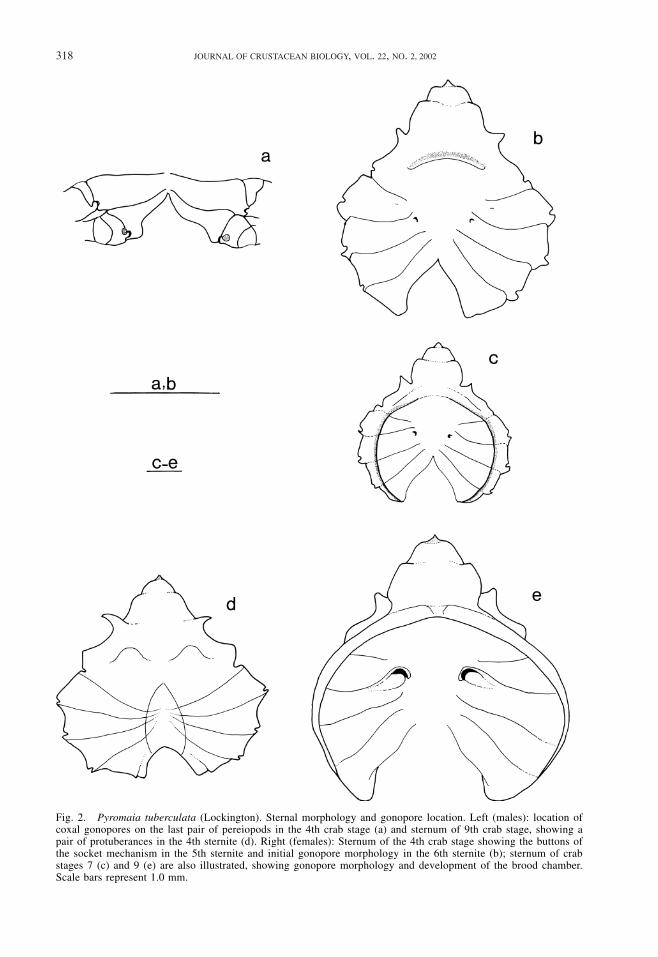
Sternum.—Gonopores were first visible incrab stage 4 for both sexes. In juvenile males,gonopores closely resembled those of theadult, both in shape and location (Fig. 2a), butin females they looked like small scars with-out any apparent opening (Fig. 2b). The but-tons of the abdominal locking mechanism(sensu Hartnoll, 1963) were found in the firstthree crab stages and occasionally in stage 4.Apart from the median depression that fits theabdomen and encloses the gonopods in laterstages, the most noticeable change in the ster-nal morphology of males was the develop-ment of a pair of protuberances on the fourthsternite. They were vestigial in crab stage 4,pronounced in stage 5, and increasingly de-veloped in further stages to the form shownin Fig. 2d. In females, the sternum underwentprofound change, forming together with theabdomen a specialized brood chamber. Dif-ferentiation started in crab stage 4, in whicha curved thickened fringe could be easily dis-tinguished on sternite 4 (Fig. 2b). Alterationsprogressed gradually until stage 7, when thechamber border was now obvious in all in-volved sternites (Fig. 2c). At the pubertymolt, the gonopore complex was formed andthe edge of the brood chamber ventrallyfolded, tightly adjusted to the mature ab-dominal shape (Fig. 2e).
Abdomen.—Main changes of abdominal mor-phology are shown in Fig. 3. In crab stage 1,the abdomen was composed of six somitesand telson. The margins of somites 4 and 5were relatively rounder compared to laterstages. In crab stage 2, somite 6 and telsonwere fused. The sockets of the locking mech-anism could be noticed laterally at approxi-mately the distal margin of former somite 6.From stage 3, the abdominal morphologyclearly differed between sexes. In males, thesecond somite was distinctly narrower in thatstage, and distal somites developed as form-ing a triangular shape from stage 4. Socketsof locking mechanism were no longer visiblein stage 5, and the relative length of somite6 + telson increased steadily through the laterstage sequence. In females, the abdomen wasalready distally wider in stage 3. Somite 5was fused to somite 6 + telson in stage 4,sockets at former somite 6 disappeared instage 5, and fused somites developed gradu-ally to the round shape found in stage 7. Af-ter the puberty molt, fused somites were sig-nificantly wider than long.
Pleopods.—No appendages were found incrab stage 1. In stage 2, appendage buds werepresent in both sexes (Figs. 4, 5). In males(Fig. 4), pleopod differentiation was fast. Instage 3, gonopods were much longer than instage 2 and possessed 4 to 5 setae distributedon their proximal third and distal tip. A lon-gitudinal groove was already present in stage4, and overall morphology of gonopods de-veloped gradually to the adult form, whichwas already found in some specimens dur-ing stage 6. As illustrated in Fig. 4, overallgonopod morphologies of late immature in-dividuals and mature last stage were identi-cal. In females (Fig. 5), small buds in crabstage 2 developed into biramous, longer ap-pendages in stage 3, after which pleopod de-velopment was relatively restrained. Exopodswere always longer than endopods. Fromstage 4 to 7, endopods became 2-segmented,and small setae appeared scattered on theproximal portion of exopods and arranged inrows along the endopods. Abrupt changestook place during the puberty molt, whenpleopods acquired the typical mature pattern,with many long setae arranged on the en-dopods and shorter ones densely aggregatedalong the exopods.
See Tables 1 and 2 for a summary of mor-
316 JOURNAL OF CRUSTACEAN BIOLOGY, VOL. 22, NO. 2, 2002

FLORES ET AL.: POSTLARVAL DEVELOPMENT OF PYROMAIA TUBERCULATA 317
Fig. 1. Pyromaia tuberculata (Lockington). Carapace outlines of 1st to 5th crab stages. Scale bar represents 1.0 mm.
318 JOURNAL OF CRUSTACEAN BIOLOGY, VOL. 22, NO. 2, 2002
Fig. 2. Pyromaia tuberculata (Lockington). Sternal morphology and gonopore location. Left (males): location ofcoxal gonopores on the last pair of pereiopods in the 4th crab stage (a) and sternum of 9th crab stage, showing apair of protuberances in the 4th sternite (d). Right (females): Sternum of the 4th crab stage showing the buttons ofthe socket mechanism in the 5th sternite and initial gonopore morphology in the 6th sternite (b); sternum of crabstages 7 (c) and 9 (e) are also illustrated, showing gonopore morphology and development of the brood chamber.Scale bars represent 1.0 mm.

phological characteristics of male and femalepostlarval stages, respectively.
Growth Patterns
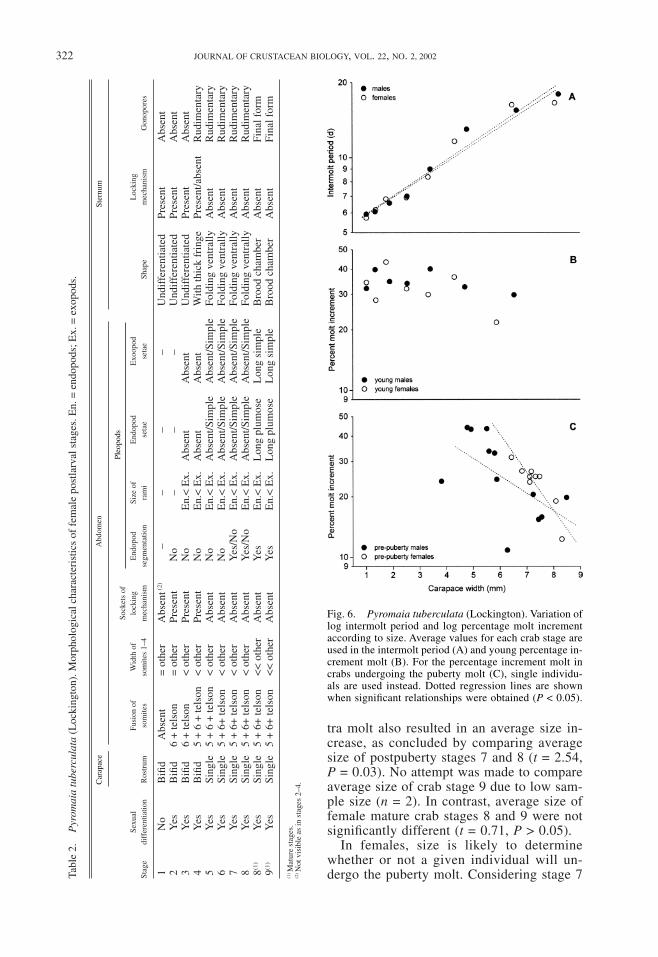
Intermolt Period, Molt Increment and Over-all Growth.—Log IP increased according tosize for both sexes. Linear relationships weresignificant in males (log IP = 0.693 +0.073CW, r = 0.98, P = 0.000) and females(log IP = 0.696 + 0.072CW, r = 0.98, P =0.000). Slopes of regression lines did not dif-fer significantly between sexes (Fig. 6, AN-COVA, F(1,2) = 0.03, P > 0.05). Log %MIslightly decreased according to size in youngcrabs, but linear relationships were not sig-nificant for either of the sexes (P > 0.05). Forcrabs undergoing the puberty molt, significantnegative relationships were obtained formales (log %MI = 1.936 – 0.088CW, r =–0.60, P = 0.04) and females (log %MI =2.695 – 0.182CW, r = –0.90, P = 0.001). Asteeper decrease of %MI could be verified infemales (Fig. 6), but slopes did not differ sta-
tistically (ANCOVA, F(1,17) = 1.18, P > 0.05).Overall growth curves are shown in Fig. 7.Exponential growth was obvious up to stage6, after which growth rates decreased rapidlyto the asymptotic stage 9 average size. Fittinggrowth curves were not attempted due to lackof data independence.
No significant differences were found inaverage size of males and females at anygiven crab stage (P > 0.05 in all compar-isons). Coefficients of variation increasedsteadily up to stage 7 (Fig. 8) but decreasedin stage 8, which included most mature crabs.Size variation for individuals delaying matu-rity to the ninth stage was still lower for fe-males but higher for males.
Allometry.—Bi-log scatterplots for AW vs.CW and ChH vs. CW were plotted for fe-males and males, respectively, and shown inFig. 9. As often found in majids, the abruptchange of the abdominal morphology duringthe puberty molt was reflected in the AW vs.
FLORES ET AL.: POSTLARVAL DEVELOPMENT OF PYROMAIA TUBERCULATA 319
Fig. 3. Pyromaia tuberculata (Lockington). Abdominal morphology. First and second crab stage illustrations areshown on the left of the figure. Sexually differentiated 3rd, 4th, 7th, and 9th crab stages progress towards the right,with male abdomens represented above the females’. Up to the 4th crab stage, scale bars represent 0.5 mm. For 7thand 9th crab stages, scale bars represent 2.0 mm.

CW plot as a discrete scatter enclosing onlyadult specimens. In contrast, no obvious seg-regation was found in the ChH vs. AW scat-terplot for males. In fact, clear differences ofchelar morphology between juveniles andadults were only found in a few examples.
In most cases, distinguishing pre- and post-puberty males was difficult.
The allometric growth of the abdomen infemales showed two different phases withinthe juvenile boundaries (Fig. 9). This inflec-tion point was located at 3.2-mm CW (Table
320 JOURNAL OF CRUSTACEAN BIOLOGY, VOL. 22, NO. 2, 2002
Fig. 4. Pyromaia tuberculata (Lockington). Morphology of the male 1st pleopod. Scale bars represent 0.1 mm forthe 2nd, 3rd, and 4th crab stages. For the 7th and 9th crab stages, scale bars represent 2.0 mm.
Table 1. Pyromaia tuberculata (Lockington). Morphological characteristics of male postlarval stages.
Abdomen
Sockets ofSexual Fusion of Width of locking Setal Locking
Stage differentiation Rostrum somites 3rd somite mechanism Sulcus distribution mechanism Gonopores
1 No Bifid Absent <4th Absent(2) – – Present Absent2 Yes Bifid 6 + telson >4th Present Absent Absent Present Absent3 Yes Bifid 6 + telson >4th Present Absent Sparse Present Absent4 Yes Bifid 6 + telson >>4th Present Present Sparse Present/absent Present5 Yes Single 6 + telson >>4th Absent Present Sparse Absent Present6 Yes Single 6 + telson >>4th Absent Final form Adult-like Absent Present7 Yes Single 6 + telson >>4th Absent Final form Adult-like Absent Present7(1) Yes Single 6 + telson >>4th Absent Final form Adult-like Absent Present8 Yes Single 6 + telson >>4th Absent Final form Adult-like Absent Present8(1) Yes Single 6 + telson >>4th Absent Final form Adult-like Absent Present9(1) Yes Single 6 + telson >>4th Absent Final form Adult-like Absent Present
(1) Mature stages.(2) Not visible as in stages 2–4.
Gonopods
SternumCarapace

3) and should indicate the occurrence of theprepuberty molt, which takes place in crabstage 4. No parallel event can be drawn fromthe allometric pattern of chelae in males.
Adult Stages and Size at Maturity.—De-scriptive statistics of mature crabs are shownin Table 4. Males entered terminal anecdysis
from crab stage 7 to 9, with most individu-als ceasing molting in the eighth stage. Mostfemales reached maturity at stage 8, but someunderwent an extra molt. Size range of laststages was much greater in males than in fe-males, with average size of females beinglarger than males (t = 3.18, P < 0.01). Formales, delaying terminal anecdysis to an ex-
FLORES ET AL.: POSTLARVAL DEVELOPMENT OF PYROMAIA TUBERCULATA 321
Fig. 5. Pyromaia tuberculata (Lockington). Morphology of female pleopods. In all stages, 2nd to 5th pleopods arerepresented from left to right. Scale bars represent 0.1 mm for the 2nd and 3rd crab stages, 0.3 mm for the 4th and7th crab stages, and 2.0 mm for the 9th crab stage.
tra molt also resulted in an average size in-crease, as concluded by comparing averagesize of postpuberty stages 7 and 8 (t = 2.54,P = 0.03). No attempt was made to compareaverage size of crab stage 9 due to low sam-ple size (n = 2). In contrast, average size offemale mature crab stages 8 and 9 were notsignificantly different (t = 0.71, P > 0.05).
In females, size is likely to determinewhether or not a given individual will un-dergo the puberty molt. Considering stage 7
322 JOURNAL OF CRUSTACEAN BIOLOGY, VOL. 22, NO. 2, 2002
Tabl
e 2.
Pyr
omai
a tu
berc
ulat
a(L
ocki
ngto
n). M
orph
olog
ical
cha
ract
eris
tics
of f
emal
e po
stla
rval
sta
ges.
En.
= e
ndop
ods;
Ex.
= e
xopo
ds.
Abd
omen
Sock
ets
ofSe
xual
Fusi
on o
fW
idth
of
lock
ing
End
opod
Size
of
End
opod
Exo
opod
Loc
king
Stag
edi
ffer
entia
tion
Ros
trum
som
ites
som
ites
1–4
mec
hani
smse
gmen
tatio
nra
mi
seta
ese
tae
Shap
em
echa
nism
Gon
opor
es
1N
oB
ifid
Abs
ent
= o
ther
Abs
ent(2
)–
––
–U
ndif
fere
ntia
ted
Pres
ent
Abs
ent
2Y
esB
ifid
6 +
tels
on=
oth
erPr
esen
tN
o–
––
Und
iffe
rent
iate
dPr
esen
tA
bsen
t3
Yes
Bifi
d6
+ te
lson
< o
ther
Pres
ent
No
En.
< E
x.A
bsen
tA
bsen
tU
ndif
fere
ntia
ted
Pres
ent
Abs
ent
4Y
esB
ifid
5 +
6 +
tels
on<
oth
erPr
esen
tN
oE
n.<
Ex.
Abs
ent
Abs
ent
With
thic
k fr
inge
Pres
ent/a
bsen
tR
udim
enta
ry5
Yes
Sing
le5
+ 6
+ te
lson
< o
ther
Abs
ent
No
En.
< E
x.A
bsen
t/Sim
ple
Abs
ent/S
impl
eFo
ldin
g ve
ntra
llyA
bsen
tR
udim
enta
ry6
Yes
Sing
le5
+ 6
+ te
lson
< o
ther
Abs
ent
No
En.
< E
x.A
bsen
t/Sim
ple
Abs
ent/S
impl
eFo
ldin
g ve
ntra
llyA
bsen
tR
udim
enta
ry7
Yes
Sing
le5
+ 6
+ te
lson
< o
ther
Abs
ent
Yes
/No
En.
< E
x.A
bsen
t/Sim
ple
Abs
ent/S
impl
eFo
ldin
g ve
ntra
llyA
bsen
tR
udim
enta
ry8
Yes
Sing
le5
+ 6
+ te
lson
< o
ther
Abs
ent
Yes
/No
En.
< E
x.A
bsen
t/Sim
ple
Abs
ent/S
impl
eFo
ldin
g ve
ntra
llyA
bsen
tR
udim
enta
ry8(1
)Y
esSi
ngle
5 +
6+
tels
on<
< o
ther
Abs
ent
Yes
En.
< E
x.L
ong
plum
ose
Lon
g si
mpl
eB
rood
cha
mbe
rA
bsen
tFi
nal f
orm
9(1)
Yes
Sing
le5
+ 6
+ te
lson
<<
oth
erA
bsen
tY
esE
n.<
Ex.
Lon
g pl
umos
eL
ong
sim
ple
Bro
od c
ham
ber
Abs
ent
Fina
l for
m
(1) M
atur
e st
ages
.(2
) N
ot v
isib
le a
s in
sta
ges
2–4.
Fig. 6. Pyromaia tuberculata (Lockington). Variation oflog intermolt period and log percentage molt incrementaccording to size. Average values for each crab stage areused in the intermolt period (A) and young percentage in-crement molt (B). For the percentage increment molt incrabs undergoing the puberty molt (C), single individu-als are used instead. Dotted regression lines are shownwhen significant relationships were obtained (P < 0.05).
Car
apac
e
Pleo
pods
Ster
num
short compared to other brachyuran crabs.This pattern relies on specific growth featuresand profound morphologic changes at specificmolts of this restricted stage sequence.
Increasing intermolt periods and decreas-ing percentage molt increments are expectedto take place from early juveniles to largestadults (Mauchline, 1977). Yet, there are someexceptions to this trend, more common in thecase of molt increment variation. Hartnoll(1982) pointed out that percentage molt in-crement might not decrease with increasingsize in crustaceans entering terminal anec-dysis. In fact, it increases in the blue crab Cal-linectes sapidus Rathbun, 1896 (see Tagatz,1968) and juveniles of the spider crab Hyascoarctatus Leach, 1815 (see Anger, 1984),while it is apparently constant for the majidPugettia producta (Randall, 1839), as calcu-lated from figures published by O’Brien(1984). Based on growth and postmolt vs.premolt plots, Furota (1996b) stated that per-centage molt increment appears to be constantthrough the crab stages of P. tuberculata. In
FLORES ET AL.: POSTLARVAL DEVELOPMENT OF PYROMAIA TUBERCULATA 323
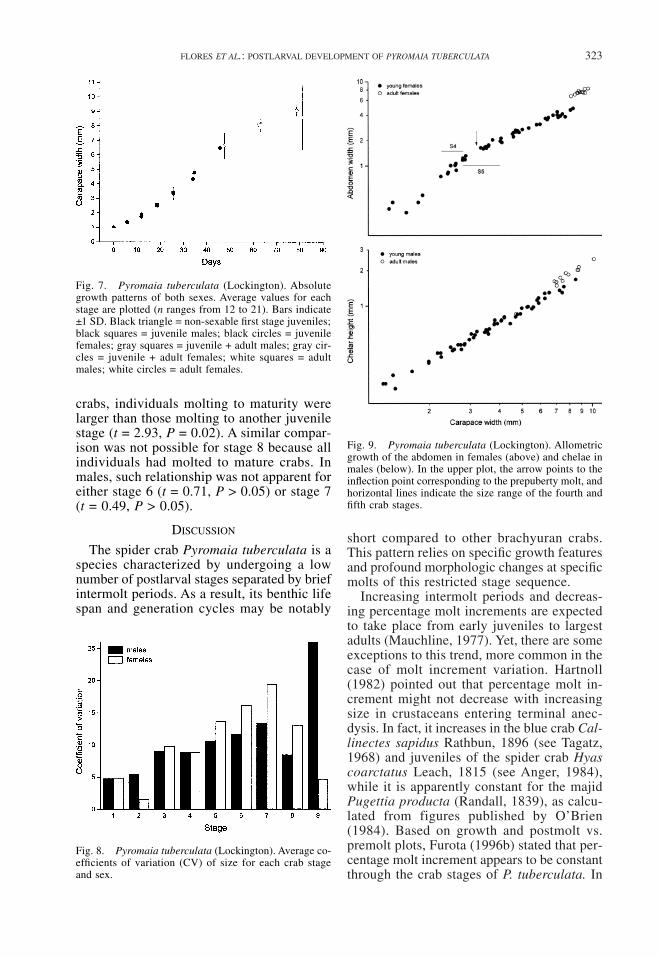
Fig. 7. Pyromaia tuberculata (Lockington). Absolutegrowth patterns of both sexes. Average values for eachstage are plotted (n ranges from 12 to 21). Bars indicate±1 SD. Black triangle = non-sexable first stage juveniles;black squares = juvenile males; black circles = juvenilefemales; gray squares = juvenile + adult males; gray cir-cles = juvenile + adult females; white squares = adultmales; white circles = adult females.
Fig. 9. Pyromaia tuberculata (Lockington). Allometricgrowth of the abdomen in females (above) and chelae inmales (below). In the upper plot, the arrow points to theinflection point corresponding to the prepuberty molt, andhorizontal lines indicate the size range of the fourth andfifth crab stages.
Fig. 8. Pyromaia tuberculata (Lockington). Average co-efficients of variation (CV) of size for each crab stageand sex.
crabs, individuals molting to maturity werelarger than those molting to another juvenilestage (t = 2.93, P = 0.02). A similar compar-ison was not possible for stage 8 because allindividuals had molted to mature crabs. Inmales, such relationship was not apparent foreither stage 6 (t = 0.71, P > 0.05) or stage 7(t = 0.49, P > 0.05).
DISCUSSION
The spider crab Pyromaia tuberculata is aspecies characterized by undergoing a lownumber of postlarval stages separated by briefintermolt periods. As a result, its benthic lifespan and generation cycles may be notably

this study, however, tests of linear regressionsignificance on log% MI vs. CW relationshipsshowed that although not significant for earlystages, percentage molt increment does de-crease with size for individuals undergoingthe puberty molt. Similar to what was foundby Cleaver (1949) for the cancrid Cancermagister Dana, 1852, there is a decrease ofpercentage molt increment associated to sex-ual maturity in P. tuberculata. Cleaver foundthat this trend is more pronounced in females,which, in spite of lacking statistical support,was also observed in the present study.
At bottom-water temperatures close to av-erage annual values in the study region (seeNegreiros-Fransozo et al., 2000), intermoltperiods are relatively short, and completepostlarval development takes place in ap-proximately 80 days. Intermolt duration ac-cording to size is nearly identical for bothsexes. During the juvenile phase, an expo-nential growth was obtained, similar to thatfound by Anger (1984), only decreasing atlate crab stages including mature specimens.Therefore, growth functions would be essen-tially exponential for plots including only in-dividuals achieving terminal anecdysis at thesame stage. Overall growth features andwithin-stage size are similar for both sexes,for what main differences between their post-larval cycles are related to the timing of ter-minal anecdysis. Males may start reproduc-ing at the age of 47–81 days, considering thatadulthood is only attained after the pubertymolt, and females at 62–79 days.
Increasing within-stage size discrepancy, asassessed by comparing coefficients of varia-tion, is not verified through all the stage se-quence as expected if there were no control-ling mechanisms preventing the predictableaccumulation of size variability. Coefficientsof variation decrease considerably at stage 8,when most crabs attain maturity, and at stage9 in the case of later mature females. Con-trol of size variation providing narrow size
ranges at particular ontogenetic phases hadbeen hypothesized to provide adaptive ad-vantages for crustaceans (Hartnoll and Dal-ley, 1981). In P. tuberculata females, sizecontrol at crab stage 8 is likely to be an out-come of a size threshold for molting to ma-turity in prepuberty individuals, similar to theconclusions of O’Brien (1984) for Pugettiaproducta. Under the captivity conditions pro-vided, small stage 7 crabs, roughly smallerthan 7.0-mm CW, underwent an ordinary moltwhile larger specimens molted to maturity.Because percentage molt increments arehigher in the former, size variation of stage8 will necessarily be lower. In males, no suchsize threshold was verified. Small size vari-ation at male stage 8, and also at female stage9, may be solely explained by the smaller per-centage molt increments experienced by crabsduring the puberty molt. Highest variationrecorded for the male stage 9 should be re-garded with caution because the value pre-sented corresponds to the average of only twospecimens. Yet, it should be noted that al-though significant, there is a considerableamount of scatter around the fitted regressionline for the percentage molt increment plotobtained for males undergoing the pubertymolt. This fact, together with early maturityat stage 7 in a significant proportion of thereared population, explains why the sizerange of adult crabs is wider in males.
Results of the present study agree to someextent with previous published work on thisspecies. Using size-frequency distributionsand laboratory-reared specimens, Furota(1996b) found seven postlarval stages infemales and six to eight in males. Thedetermination of stage number matched be-tween field and laboratory evidence, butFurota found contrasting larger crabs in thenatural population. Therefore, it can be con-cluded that extrinsic (environmental) condi-tions are playing a major role in the controlof final crab size by affecting molt incre-
324 JOURNAL OF CRUSTACEAN BIOLOGY, VOL. 22, NO. 2, 2002
Table 3. Results of Somerton’s Mature 2 computer technique to locate the prepuberty molt in juvenile stages ofPyromaia tuberculata (Lockington). AW = abdomen width; ChH = chelar height.
Somerton’s F-test
Sex Dimension Value Cut-point (mm) Growth phase Intercept (log a) Slope (b)
Female AW 42.8* 3.2 Early juvenile –1.95 2.03Late juvenile – 1.22
Male ChH 0.1ns – Juvenile –1.82 1.08
*P = 0.000; ns = not significant.

ments. The number of molts seems to be moreconservative, probably under genetic control,as suggested by Hartnoll et al. (1993). In thepresent study, it was also found that numberof stages to reach maturity is more variablein males than females. Captive stages areabout the same size in both accounts, but thisstudy reveals that individuals may passthrough one extra stage on average. Such adifference may well be a result of genetic di-vergence between these populations. Identi-fying from where this species originatedwhen introduced in southeastern Brazil mayhelp to solve this issue.
Reduced postlarval series involves deepmorphologic alterations at each stage, mak-ing possible the identification of most stagesby means of inspecting certain morphologi-cal characters. Such changes are particularlymarked along the female stage sequence, dur-ing which the abdomen, sternum, andpleopods develop jointly to form a conspic-uous brood chamber in mature specimens,similar to that reported by Hartnoll (1965) inthe closely related inachine Stenorhynchusseticornis (Herbst, 1788). In both sexes, ab-dominal somites 6 and telson are fused in thesecond stage. In the case of males, fusedsomites are relatively longer in later stagesand accommodate developing gonopods. Infemales, somites 5, 6, and telson are all fusedin crab stage 4, which will afterwards developtowards a concave broad structure only ob-served in fully developed crabs.
The prepuberty molt, thus, divides the de-velopment of young into an early juvenilephase until crab stage 4, during which mostremarkable changes take place, and a late ju-venile phase, from stage 5 to later prepubertycrabs, when distal abdominal somites developto a rounder shape. For females that had al-ready undergone the prepuberty molt, thenumber of further juvenile stages seems to de-
pend on the increment required to reach thematurity size threshold. However, more spe-cific studies are needed to verify if such acontention applies. Other studies on the rela-tive growth of spider crabs have shown theoccurrence of the prepuberty molt in malesand females using the relative growth ofchelae (Teissier, 1935; Vernet-Cornubert,1958; Hartnoll, 1963). In P. tuberculatamales, allometric relationships are apparentlyof no use to locate either the prepuberty orthe puberty molt. The relative growth ofchelae, during the juvenile growth phase, isbetter explained by a single regression line,and mature specimens do not appear as a dis-crete group in bi-log plots. Only in some in-dividuals is there a marked development ofchelipeds during the puberty molt, as ob-served in other spider crabs (Hartnoll, 1963,1965). It is likely that both morphotypes co-exist in natural populations. Whether thoseadult crabs differ in their mating success isan interesting question for future research.The morphology of gonopods is also quitesimilar between late juvenile and postpubertystages. In other majids, large prepuberty crabsmay already hold developed testes (Hartnoll,1963) and engage in copula (Wirtz andDiesel, 1983). Hence, there are no groundsto discard successful mating by those indi-viduals.
Breeding in P. tuberculata extends year-round, thus providing continuous oppor-tunistic recruitment of megalopae (Furota,1996a). Larval development takes from 14 to20 days at temperatures from 20° to 25°C(Furota, 1996a; Fransozo and Negreiros-Fransozo, 1997), for what the whole cyclemay take less than four months. Developmentof gonads may start before the puberty moltbecause soft-shell mating shortly followed byegg laying was observed. Less fecund primip-arous females may counterbalance reproduc-tive output by extruding a larger number ofeggs in following ovipositions (Furota,1996b), a reproductive feature also verified inother majids (Somerton and Meyers, 1983;Diesel, 1988; Bryant and Hartnoll, 1995).
Assuming that maximum survival of off-spring is the main variable determining life-history strategies, fast development to matu-rity shortly followed by breeding is predictedto be selected under high mortality conditions(Hartnoll and Gould, 1988). Furota (1996b)highlighted the overall patterns of growth and
FLORES ET AL.: POSTLARVAL DEVELOPMENT OF PYROMAIA TUBERCULATA 325
Table 4. Descriptive statistics of size in adult crabs ofPyromaia tuberculata (Lockington). n = number ofobservations; CW = carapace width.
CW (mm) Overall size
Sex Stage n (%) Mean SD Mean Range
Male 7 5 6.8 1.2 7.8 4.7–10.28 8 8.2 0.79 2 8.6 2.2
Female 8 7 8.9 0.5 8.9 8.1–9.69 4 9.1 0.4

reproduction in P. tuberculata, and showed thatthree generation cycles may take place eachyear. The species is thereby capable of ex-ploiting unstable environments such as the up-per parts of the Bay of Tokyo where hypoxiacauses mass mortality during the summer.Settling breeding populations within a rela-tively short time interval is probably the mostimportant condition determining the successof invading populations. From the first recordof P. tuberculata in southeastern Brazil in 1989(Melo, 1989), the quantity of crabs in benthictrawlings has become increasingly frequent.Monitoring its population dynamics in ade-quate detail is most desirable to aid model-ing the temporal abundance trends of marineexotic invertebrates and assess the impact ofthis particular species on the benthic com-munity. The descriptions presented herein onthe postlarval stages of P. tuberculata can beused to interpret size-frequency distributionsby associating histogram modes to their cor-responding crab stages. This seems particu-larly promising because large molt incrementsmay allow unambiguous separation of normalcomponents in polymodal distributions.
ACKNOWLEDGEMENTS
We are thankful to the crew members of the Oceano-graphic Vessel Progresso for assistance during the col-lecting trips to Ubatuba. We thank Dr. A. Fransozo andthe students of NEBECC for their help during the rear-ing procedure. We thank Dr. J. Paula, Universidade deLisboa, for providing logistic support during a stay atthe Laboratório Maritimo da Guia, Cascais, Portugal.FPLM is indebted to Carla Flores for her kind hospital-ity in Portugal. This work was funded by FAPESP (Fun-dação de Amparo à Pesquisa do Estado de São Paulo,Brazil) as JP-3 postdoctoral grants # 99/082556-5 and #99/10407-1 to FPLM and logistic support (#98/03134-6and #98/09426-9).
LITERATURE CITED
Anger, K. 1984. Development and growth in larval andjuvenile Hyas coarctatus (Decapoda, Majidae) rearedin the laboratory.—Marine Ecology Progress Series 19:115–123.
Bryant, A. D., and R. G. Hartnoll. 1995. Reproductiveinvestment in two spider crabs with different breedingstrategies.—Journal of Experimental Marine Biologyand Ecology 188: 261–275.
Childress, J. J., and M. H. Price. 1978. Growth rate ofthe bathypelagic crustacean Gnathophausia ingens(Mysidacea: Lophogastridae). 1. Dimensional growthand population structure.—Marine Biology 50: 47–62.
Cleaver, F. C. 1949. Preliminary results of the coastalcrab (Cancer magister) investigation.—WashingtonState Department of Fisheries, Biological report 49A:47–82.
Diesel, R. 1988. Male-female association in the spidercrab Inachus phalangium: the influence of female re-
productive stage and size.—Journal of Crustacean Bi-ology 8: 63–69.
Flores, A. A. V., M. L. Negreiros-Fransozo, and A. Fran-sozo. 1998. The megalopa and juvenile developmentof Pachygrapsus transversus (Gibbes, 1850) (De-capoda, Brachyura) compared with other grapsidcrabs.—Crustaceana 71: 197–222.
Fransozo, A. 1986/87. Desenvolvimento dos estágios ju-venis de Sesarma (Holometopus) rectum Randall, 1840(Decapoda, Grapsidae) obtidos em laboratório.—Nat-uralia 11/12: 77–87.
———, and M. L. Negreiros-Fransozo. 1987. Morfolo-gia dos primeiros estágios juvenis de Eriphia gonagra(Fabricius, 1781) e Eurypanopeus abbreviatus (Stimp-son, 1860) (Crustacea, Decapoda, Xanthidae), obtidosem laboratório.—Papéis Avulsos de Zoologia 36:257–277.
———, and ———. 1997. Larval stages of Pyromaia tu-berculata (Lockington, 1877) (Decapoda, Majidae, In-achinae) reared in the laboratory.—Crustaceana 70:304–323.
Furota, T. 1996a. Life cycle studies on the introducedspider crab Pyromaia tuberculata (Lockington)(Brachyura: Majidae). I: Egg and larval stages.—Jour-nal of Crustacean Biology 16: 71–76.
———. 1996b. Life cycle studies on the introducedspider crab Pyromaia tuberculata (Lockington)(Brachyura: Majidae). II: Crab stage and reproduc-tion.—Journal of Crustacean Biology 16: 77–91.
Griffin, D. J. G., and H. A. Tranter. 1986. The DecapodaBrachyura of the Siboga expedition. Part VIII. Maji-dae.—Siboga-expeditie monographie 39: 1–335. E. J.Brill, Leiden.
Hartnoll, R. G. 1963. The biology of Manx spidercrabs.—Proceedings of the Zoological Society of Lon-don 141: 423–496.
———. 1965. The biology of spider crabs: a compari-son of British and Jamaican species.—Crustaceana 9:1–16.
———. 1982. Growth. Pp. 111–196 in D. E. Bliss, ed.-in-chief. The Biology of Crustacea. Vol. 2. L. G. Abele,ed. Embryology, Morphology and Genetics. AcademicPress, New York.
———, A. D. Bryant, and P. Gould. 1993. Size distribu-tion in spider crab populations—spatial and temporalvariation.—Journal of Crustacean Biology 13:647–655.
———, and R. Dalley. 1981. The control of size varia-tion within instars of a crustacean.—Journal of Exper-imental Marine Biology and Ecology 53: 235–239.
———, and P. Gould. 1988. Brachyuran life historystrategies and the optimization of egg production. Pp.1–9 in A. A. Fincham and P. S. Rainbow, eds. Aspectsof Decapod Crustacean Biology, Symposia of the Zo-ological Society of London. Vol. 59. Clarendon Press,Oxford.
———, and A. L. Rice. 1986. Further studies on the bi-ology of the deep-sea spider crab Dorhynchus thom-soni: instar sequence and the annual cycle. Pp. 231–241in P. E. Gibbs, ed. Nineteenth European Marine Biol-ogy Symposium. Cambridge University Press, Cam-bridge.
Ingle, R. W. 1977. The larval and post-larval develop-ment of the scorpion spider crab, Inachus dorsettensis(Pennant) (Family: Majidae), reared in the labora-tory.—Bulletin of the British Museum of Natural His-tory 30: 331–342.
326 JOURNAL OF CRUSTACEAN BIOLOGY, VOL. 22, NO. 2, 2002

Jones, D. R., and R. G. Hartnoll. 1997. Mate selectionand mating behaviour in spider crabs.—Estuarine,Coastal and Shelf Science 44: 185–193.
Mauchline, J. 1976. The Hiatt growth diagram for Crus-tacea.—Marine Biology 35: 79–84.
———. 1977. Growth of shrimps, crabs and lobsters—an assessment.—Journal du Conseil Internationale dela Exploration de la Mer 37: 162–169.
Melo, G. A. S., V. G. Veloso, and M. C. Oliveira. 1989.A fauna de Brachyura (Crustacea, Decapoda) do litoraldo sudeste do Brasil. Lista preliminar.—Nerítica 4:1–31.
Mohamedeen, H., and R. G. Hartnoll. 1989. Larval andpostlarval growth of individually reared specimens ofthe common shore crab Carcinus maenas (L.).—Jour-nal of Experimental Marine Biology and Ecology 134:1–24.
Morgan, G. J. 1990. An introduced Eastern Pacific ma-jid crab from Cockburn Sound, Southwestern Aus-tralia.—Crustaceana 58: 316, 317.
Negreiros-Fransozo, M. L., and A. Fransozo. 1991.Growth and age determination of three juvenile crabspecies (Crustacea, Decapoda, Brachyura).—PapéisAvulsos de Zoologia, São Paulo 37: 277–283.
———, J. M. Nakagaki, and A. L. D. Reigada. 2000. Sea-sonal occurrence of decapods in shallow waters of asubtropical area. Pp. 351–361 in J. C. von Vaupel Kleinand F. R. Schram, eds. The Biodiversity Crisis andCrustacea. A. A. Balkema, The Netherlands.
O’Brien, J. 1984. Precocious maturity of the majid crab,Pugettia producta, parasitized by the Rhizocephalanbarnacle, Heterosaccus califomicus.—Biological Bul-letin 166: 384–395.
Rice, A. L., and R. G. Hartnoll. 1983. Aspects of thebiology of the deep-sea spider crab, Dorhynchus thom-soni (Crustacea: Brachyura).—Journal of Zoology,London 201: 417–431.
Rieger, P. J., and C. Nakagawa. 1995. Desenvolvimentojuvenil de Chasmagnathus granulata Dana, 1851(Crustacea, Decapoda, Grapsidae), em laboratório.—Nauplius 3: 59–74.
Sakai, T. 1976. Crabs of Japan and the Adjacent Seas.Kodansha, Tokyo. 773 pp.
Shen, C. J. 1935. An investigation of the post-larval de-velopment of the shore crab Carcinus maenas, withspecial reference to the external secondary sexual char-acters.—Proceedings of the Zoological Society of Lon-don 1: 1–33.
Siegel, P. R., and A. W. Wenner. 1985. Field and labo-ratory studies of sand crab growth. Pp. 165–170 in A.M. Wenner, ed. Factors in Adult Growth. A. A.Balkema, Rotterdam. 362 pp.
Snedecor, G. W., and W. G. Cochran. 1980. StatisticalMethods. The Iowa State University Press, Ames, Iowa.507 pp.
Sokal, R., and F. J. Rohlf. 1995. Biometry: the Principlesand Practice of Statistics in Biological Research, Thirdedition. 887 pp.
Somerton, D. A. 1983. The size at sexual maturity ofblue king crab Paralithodes platypus, in Alaska.—Fish-ery Bulletin (US) 18: 621–628.
———, and W. S. Meyer. 1983. Fecundity differences be-tween primiparous and multiparous female Alaskan tan-ner crab (Chionoecetes bairdi).—Journal of CrustaceanBiology 3: 183–186.
Tagatz, M. E. 1968. Growth of juvenile blue crabs, Cal-linectes sapidus Rathbun, in the St. Johns River,Florida.—Fishery Bulletin (US) 67: 281–288.
Teissier, G. 1935. Croissance des variants sexuels chezMaïa squinado L.—Travaux de la Station Biologiquede Roscoff 13: 93–130.
Vernet-Cornubert, G. 1958. Biologie générale de Pisatetraodon (Pennant).—Bulletin de l’institut Océan-ographique, Monaco 113: 1–52.
Wirtz, P., and R. Diesel. 1983. The social structure ofInachus phalangium, a spider crab associated with thesea anemone Anemonia sulcata.—Zeitschrift fürTierpsychologie 62: 209–234.
RECEIVED: 26 February 2001.ACCEPTED: 4 September 2001.
FLORES ET AL.: POSTLARVAL DEVELOPMENT OF PYROMAIA TUBERCULATA 327




















