
Weber and noise adaptation in the retina of the toad Bufo marinus

A
fttaIuogias©
K
1
td1e2apssviat
0d
Aquatic Toxicology 86 (2008) 447–456
Endosulfan acute toxicity in Bufo bufo gills: Ultrastructuralchanges and nitric oxide synthase localization
Ilaria Bernabo a, Elvira Brunelli a,∗, Cecilia Berg b, Antonella Bonacci a, Sandro Tripepi a
a Department of Ecology, University of Calabria, Via P. Bucci, 87036 Rende (Cosenza), Italyb Department of Environmental Toxicology, Uppsala University, Norbyvagen 18A, 752 36 Uppsala, Sweden
Received 13 September 2007; received in revised form 13 December 2007; accepted 14 December 2007
bstract
Endosulfan is an organochlorine pesticide used in agriculture for a wide range of crops. Endosulfan concentrations of up to 0.7 mg/L can beound in ponds and streams near sprayed agricultural fields. We investigated the short-term toxicity of endosulfan in common toad (Bufo bufo)adpoles after 24, 48, and 96 h of exposure. Acute toxicity was evaluated at nominal concentrations ranging from 0.01 to 0.6 mg/L: concentrationshat could be found after the application of pesticide. Our results show that 0.43 mg/L of endosulfan caused 50% mortality (LC50). The effects ofsublethal endosulfan concentration (0.2 mg/L) on gill apparatus morphology were evaluated by scanning and transmission electron microscopy.
mmunohistochemical methods were also applied to detect the expression pattern of the inducible isoform of nitric oxide synthase (iNOS) in the gillssing the confocal laser scanner microscope. Exposure to 0.2 mg/L of endosulfan caused an apparent increase in mucus production, the occurrencef secretory vesicles and lamellar bodies, a widening of intercellular spaces and additionally there was evidence of an inflammatory response in the
ill apparatus. The morphological alterations occurred after 24 h and were more pronounced after 48 and 96 h of exposure. Altered morphology andncreased mucus secretion indicate impaired gas exchange and osmoregulation in the gills. In addition, there was an increase of iNOS expressionfter 24 and 48 h which may reflect hypoxia and inflammation in the gill epithelium. Our results clearly indicate that short-term exposure to aublethal concentration of endosulfan, near the high end of the environmental range, disrupts gill morphology and function in B. bufo tadpoles.2007 Elsevier B.V. All rights reserved.
fan
cMaaac1
hsabp
eywords: Amphibians; Gills; Ultrastructure; iNOS; Water pollution; Endosul
. Introduction
Amphibians have been increasingly drawing the atten-ion of conservation biologists due to reports of a globalecline in their populations (Wake, 1991; Alford and Richards,999; Houlahan et al., 2000; Kiesecker et al., 2001; Sparlingt al., 2001; Blaustein and Kiesecker, 2002; Stuart et al.,004). Several hypotheses have been proposed to explain themphibian decline: habitat degradation, emerging diseases (i.e.arasites/infections), predation and competition by introducedpecies, increased ultraviolet radiation resulting from atmo-pheric ozone depletion (Beebee and Griffiths, 2005), and aulnerability to waterborne and airborne environmental contam-
nants (Wake, 1991; Davidson et al., 2001, 2002). Amphibiansre particularly exposed to environmental pollutants becausehey have permeable skin, gills and eggs that can absorb chemi-∗ Corresponding author. Tel.: +39 0984492975; fax: +39 0984492986.E-mail address: [email protected] (E. Brunelli).
ota(cFA
166-445X/$ – see front matter © 2007 Elsevier B.V. All rights reserved.oi:10.1016/j.aquatox.2007.12.006
als from the aquatic environment (Duellman and Trueb, 1986;aterna et al., 1995; Venturino et al., 2003). Furthermore, many
mphibians spend the greater part of their larval development inquatic habitats adjacent to agricultural fields where pesticidesre applied; hence, they can be directly exposed to waterbornehemicals during this critical growth period (Carey and Bryant,995; Clark et al., 1998).
Endosulfan (6,7,8,9,10,10-hexachloro-1,5,5a,6,9,9a-hexa-ydro-6,9-ethano-2,4,3-benzodioxathiepin-3-oxide) is a non-ystemic organochlorine insecticide, which was developednd introduced in the mid 1950s (EFSA, 2005) and haseen considered a safer alternative to other organochlorineesticides, such as DDT and Dieldrin, in many countries allver the world since the 1970s. In 2001 endosulfan was amonghe 10 most frequently found pesticide residues on fruitsnd vegetables from member states of the European Union
European Commission, 2004). Spain and Italy are the majoronsumers of endosulfan within the European Union (Germanederal Environment Agency, 2004). In the United States ofmerica approximately 1.4 million pounds of endosulfan active
4 Toxic
itaw1phiivsb(t1daa2btyo
aeivceobdatmlH2etbt
(nor(ienqpita
ttofPrmB
aecca
2
2
liDplaG
2
pi4ld
2
pcTwsdnn1
sgl
48 I. Bernabo et al. / Aquatic
ngredient are used annually (EPA, 2002). Endosulfan concen-rations of 0.7 mg/L have been found near agricultural fieldsnd levels exceeding 0.5 mg/L were discovered in a tidal creekater column following agricultural runoff events (Ernst et al.,991; EPA, 2002). High concentrations of endosulfan were alsoresent in adult and larval Rana lessonae and the hemiclonalybrid R. esculenta specimens collected at an agricultural arean central Italy (Fagotti et al., 2005). This molecule is persistentn the environment and has a tendency to bioaccumulate inarious aquatic organisms (Naqvi and Vaishnavi, 1993). Ithould, therefore, along with its metabolite endosulfan sulphatee included in the list of persistent organic pollutants (POPs)EFSA, 2005). Endosulfan is characterized by strong acuteoxicity to fish and aquatic invertebrates (Johnson and Finley,980; Gopal et al., 1981, 1985; Coats, 1990), and has wellocumented neurotoxic effects in both humans and laboratorynimals (Anand et al., 1986; Singh et al., 1992; Lakshmanand Raju, 1994; Paul and Balasubramaniam, 1997; Giusi et al.,005). Risk assessments for aquatic species were conductedy the US Environmental Protection Agency. They estimatedhat the use of endosulfan, at typical rates in a given number ofears, on tomatoes would result in a 90% probability that 60%f aquatic species would suffer mortality (EPA, 2002).
A concept of ecotoxicology is that deleterious health effectsre preceded by sublethal changes at molecular and cellular lev-ls. Detection of such markers can therefore serve as an earlyndication of contaminant bioavailability or impact, thus pro-iding valuable information regarding the extent, and possibleonsequences of prolonged or continued exposure (Langstont al., 2002). Amphibian gills are, along with the skin, onef the first sites of contact with the environmental pollutantecause of the continuous processing of water. There are fewata regarding the effects of endosulfan on amphibians, andvailable reports have focused mainly on acute toxicity inhe aquatic stage of amphibians, survival time, larval malfor-
ation, and behavioural pattern, both in field studies and inaboratory experiments (Gopal et al., 1981; Berrill et al., 1998;arris et al., 1998, 2000; Lajmanovich et al., 2005; Broomhall,002, 2004). No previous study has examined the effects ofndosulfan on amphibian gill morphology. We therefore inves-igated the effects of endosulfan on the ultrastructure of Bufoufo gills with the aim to develop a sensitive biomarker ofoxicity.
Moreover, we also studied the involvement of nitric oxideNO) through expression in the gills of the inducible isoform ofitric oxide synthase (iNOS) by histochemical analysis. Nitricxide (NO) is an intercellular messenger molecule, which plays aole in many physiological and pathophysiological mechanismsMoncada et al., 1992). The inducible isoform iNOS is expressedn different types of cells, including macrophages, vascularndothelial cells, vascular smooth muscle cells, epithelial cells,eutrophils, and fibroblasts. It catalyzes the formation of largeuantities of NO with cytotoxic/cytostatic effects in response to
athological stimuli. iNOS is known as the isoform involved inmmunological or inflammatory processes. The expression pat-ern of iNOS may reflect the degree of inflammation in a tissuend provide a measure for assessing the inflammatory responset3ii
ology 86 (2008) 447–456
o various chemicals (Kim et al., 2004, 2005). Therefore, induc-ion of iNOS immunoreactivity in the gills serves as an indicatorf toxic and oxidative stress in amphibians and can provide use-ul information on the ability of gills to respond to this stress.revious studies on the role and activity of NO in amphibians inesponse to pesticide exposure have focused only on the epider-is and on the liver (Johnson et al., 2000; Fenoglio et al., 2006;arni et al., 2007).
In this study the effects of endosulfan on B. bufo gills weressessed by analyzing the gill epithelium ultrastructure and byxamining the immunohistochemical expression of iNOS usingonfocal microscopy. In order to identify sublethal exposureoncentrations of endosulfan for B. bufo tadpoles, we conductedrange finding study.
. Materials and methods
.1. Study organisms
The common toad (B. bufo) is an opportunistic species thatives in a wide variety of habitats including agricultural sites. Its classified in the Least concern (LC) IUCN category (2007).uring recent decades, however, a marked decrease in B. bufoopulations has been observed locally in Europe, which mostikely to be due to the loss and alteration of its breeding habitats a result of intensive agricultural practices (Pavignano andiacoma, 1990; Carrier and Beebee, 2003).
.2. Tadpole maintenance and holding conditions
B. bufo tadpoles were hatched from eggs collected from aermanent pond located near Cosenza (Calabria, Southern Italy),n a regional natural reserve. The tadpoles were acclimatized fordays in tap water at a temperature of 22 ± 1 ◦C, and were fed
ettuce ad libitum. The developmental stage of the tadpoles wasetermined according to Gosner (1960).
.3. Exposure to endosulfan
Endosulfan (Chem Service Inc., West Chester, PA, USA,urity 99%) was dissolved in tap water to obtain the nominaloncentrations: 0.01, 0.05, 0.1, 0.2, 0.3, 0.35, 0.4, 0.5, 0.6 mg/L.adpoles of comparable body dimensions at Gosner stage 25ere assigned to the various exposure tanks. This stage corre-
ponds to that used for previous studies on endosulfan toxicity;espite the suggestions of some authors (Berrill et al., 1994)o evidence was available for differences in sensitivity betweenewly hatched and further developed tadpoles (Berrill et al.,998).
The control group was kept in tap water. A static expo-ure system was used in accordance with standard procedureuidelines (ASTM, 1997). After 96 h endosulfan biodegradationevels were observed to be less than 15% of the initial concentra-
ion as shown in other similar studies (Broomhall, 2004). Each0 L glass tank contained 30 tadpoles. For each concentration,ncluding the control, three replicate tanks were used. The exper-ment was conducted in 12:12 h light–dark cycles, median pH
Toxic
7t
lsaf
2
fUupiptCstALct
ao
2
swaaw
bHawsoCflma(ncw
2
d
mcS
3
3
pggdtFme
3a
3The gill apparatus of B. bufo, as that of other anuran species,
is supported by four pairs of gill arches. Each branchial archgives insertion ventrally to the gill tufts and dorsally to the gillfilters (Fig. 1A). Filters are constituted by a main flattened axis to
Table 1The relation between the several concentrations of endosulfan and the mortalityrate of Bufo bufo
Concentration (mg/L) Number of dead tadpoles
0.000 00.010 00.050 00.100 00.200 90.300 180.350 240.400 330.500 540.600 78
Sample size = 90.
Table 2Estimated LC values and confidence limits for Bufo bufo exposed to endosulfan
Point Concentration (mg/L) 95% confidence limits
Lower Upper
LC 1.00 0.149 0.096 0.190LC 5.00 0.203 0.149 0.242LC 10.00 0.240 0.188 0.277LC 15.00 0.268 0.219 0.303LC 50.00 0.430 0.392 0.480
I. Bernabo et al. / Aquatic
.3, and water temperature 22 ± 1 ◦C. During the 96 h exposurehe animals were not fed.
After identifying the lethal concentration, we exposed B. bufoarvae for 24, 48, and 96 h of a sublethal concentration of endo-ulfan (0.2 mg/L) under the experimental conditions describedbove. Two replicates, each containing 35 tadpoles were usedor the control and the exposure group.
.4. Morphological analysis
The animals were anesthetized with tricaine methanesul-onate MS-222 (Sigma–Aldrich Chemical Co., St. Louis, MO,SA) after 24, 48, and 96 h exposure and the gills were removedsing a dissection microscope. Three randomly selected tadpoleser group and time point were used for the SEM and TEM stud-es, respectively. Samples were fixed in 3% glutaraldehyde inhosphate buffer (0.1 M, pH 7.2) for 2 h and then in 1% osmiumetroxide in the same buffer for 2 h at 4 ◦C (Sigma–Aldrichhemical Co.). After dehydration, specimens for the TEM
tudy were immersed in uranyl acetate in increasing concen-rations of acetone, and then embedded in Epon-Araldite (Flukag, Buchs, Switzerland). Ultrathin sections were obtained withKB Nova ultramicrotome, stained with uranyl acetate and leaditrate, coated in Edwards EM 400, and then examined and pho-ographed with Zeiss EM 900 transmission electron microscope.
Samples for the SEM study were dehydrated and driedccording to the critical point method, covered with gold, andbserved by Zeiss DSM 940 scanning electron microscope.
.5. Immunohistochemistry
Three exposed and three control tadpoles were randomlyelected and analyzed at each time point. The sample specimensere fixed for 24 h in Bouin’s fixative (freshly made), dehydrated
nd embedded in paraffin wax. Sagittal 7 �m sections were cutnd mounted on slides. The sections were rinsed with distilledater and phosphate buffered saline (PBS).Immunofluorescence staining was performed as described
y Coons et al. (1955). The sections were immersed in 2%2O2 in 0.1 PBS for 20 min to block endogenous peroxidase
ctivity. After being washed three times in PBS, the sectionsere incubated with normal goat serum for 10 min. Unwashed
ections were incubated overnight at 4 ◦C with a mouse mon-clonal antibody to iNOS (1:100, Sigma–Aldrich Chemicalo); after several washes in PBS, they were incubated withuoresceinisothiocyanate-conjugated g-globulins goat anti-ouse (diluted 1:20, Sigma–Aldrich Chemical Co.) for 30 min
t room temperature. In order to examine non-specific bindingnegative control), the primary antiserum was substituted withon-immune goat serum (diluted 1:50, Sigma–Aldrich Chemi-al Co.) in PBS in control sections. The sections were analyzedith Leica TCS SP2 Confocal Laser Scanning Microscope.
.6. Statistical analysis
The lethal concentration for 50% of the animals (LC50) wasetermined by the use of Finney’s Probit Analysis LC50 Deter-
LLLL
ology 86 (2008) 447–456 449
ination Method (Finney, 1971). The computer analysis wasonducted with LC50 1.00 software developed by EPA (LC50oftware Program Version 1.00, 1999).
. Results
.1. Survival
The nominal 96 h LC50 value for endosulfan in B. bufo tad-oles was 0.430 mg/L. No mortality occurred in the controlroup and the 0.01, 0.05, or 0.1 mg/L endosulfan-containingroups. In all groups the mortality rate was lower than 50%uring the first 24 h period. Table 1 shows the relation betweenhe endosulfan concentration and the mortality rate according toinney’s Probit Analysis using EPA Computer Program. Esti-ated LC50 values and confidence limits for 96 h endosulfan
xposure are presented in Table 2.
.2. Morphological and ultrastructural analysis of the gillpparatus
.2.1. Control group
C 85.00 0.689 0.591 0.900C 90.00 0.771 0.646 1.053C 95.00 0.909 0.737 1.331C 99.00 1.240 0.938 2.070
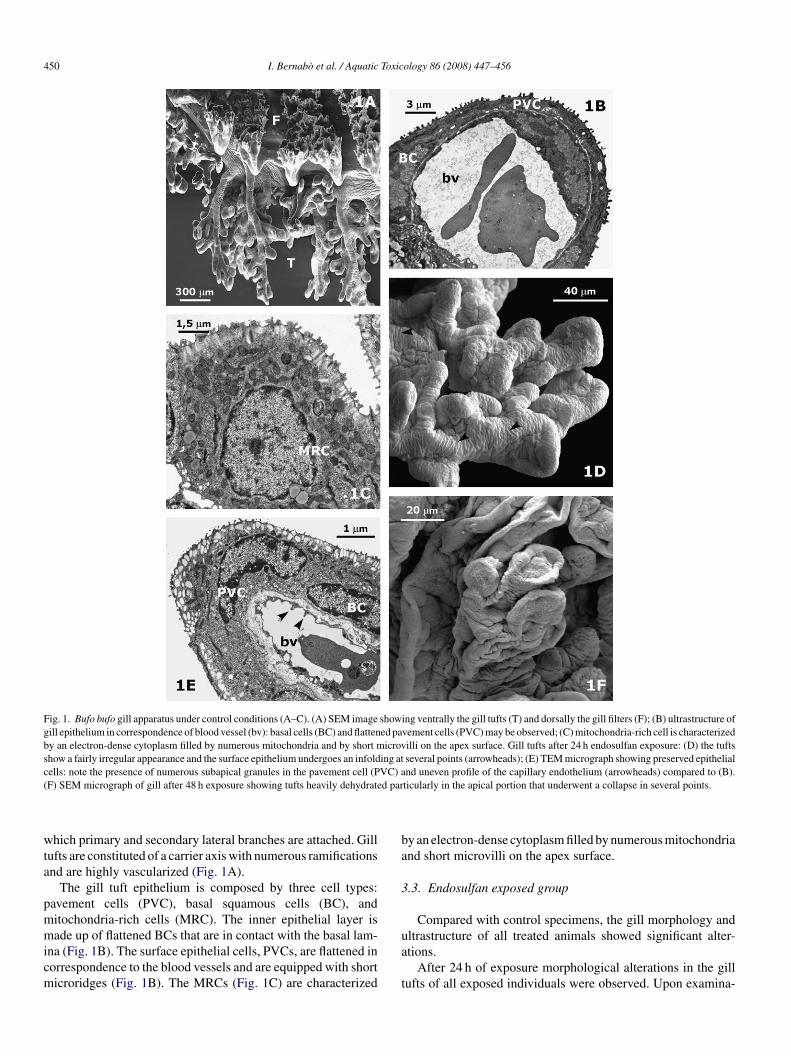
450 I. Bernabo et al. / Aquatic Toxicology 86 (2008) 447–456
Fig. 1. Bufo bufo gill apparatus under control conditions (A–C). (A) SEM image showing ventrally the gill tufts (T) and dorsally the gill filters (F); (B) ultrastructure ofgill epithelium in correspondence of blood vessel (bv): basal cells (BC) and flattened pavement cells (PVC) may be observed; (C) mitochondria-rich cell is characterizedby an electron-dense cytoplasm filled by numerous mitochondria and by short microvilli on the apex surface. Gill tufts after 24 h endosulfan exposure: (D) the tuftsshow a fairly irregular appearance and the surface epithelium undergoes an infolding at several points (arrowheads); (E) TEM micrograph showing preserved epithelialcells: note the presence of numerous subapical granules in the pavement cell (PVC) and uneven profile of the capillary endothelium (arrowheads) compared to (B).( d par
wta
pmmicm
ba
3
F) SEM micrograph of gill after 48 h exposure showing tufts heavily dehydrate
hich primary and secondary lateral branches are attached. Gillufts are constituted of a carrier axis with numerous ramificationsnd are highly vascularized (Fig. 1A).
The gill tuft epithelium is composed by three cell types:avement cells (PVC), basal squamous cells (BC), anditochondria-rich cells (MRC). The inner epithelial layer is
ade up of flattened BCs that are in contact with the basal lam-na (Fig. 1B). The surface epithelial cells, PVCs, are flattened inorrespondence to the blood vessels and are equipped with shorticroridges (Fig. 1B). The MRCs (Fig. 1C) are characterized
ua
t
ticularly in the apical portion that underwent a collapse in several points.
y an electron-dense cytoplasm filled by numerous mitochondriand short microvilli on the apex surface.
.3. Endosulfan exposed group
Compared with control specimens, the gill morphology and
ltrastructure of all treated animals showed significant alter-tions.After 24 h of exposure morphological alterations in the gillufts of all exposed individuals were observed. Upon examina-
Toxic
ttTcc
atslbacog
tcmSpcwt
a
Flwssc
I. Bernabo et al. / Aquatic
ion with the SEM, the tuft showed an irregular appearance andhe surface epithelium was infolded at several points (Fig. 1D).he ultrastructure of the epithelial cells appeared intact: only theapillary endothelium showed an irregular inner surface profileompared with the control (Fig. 1E).
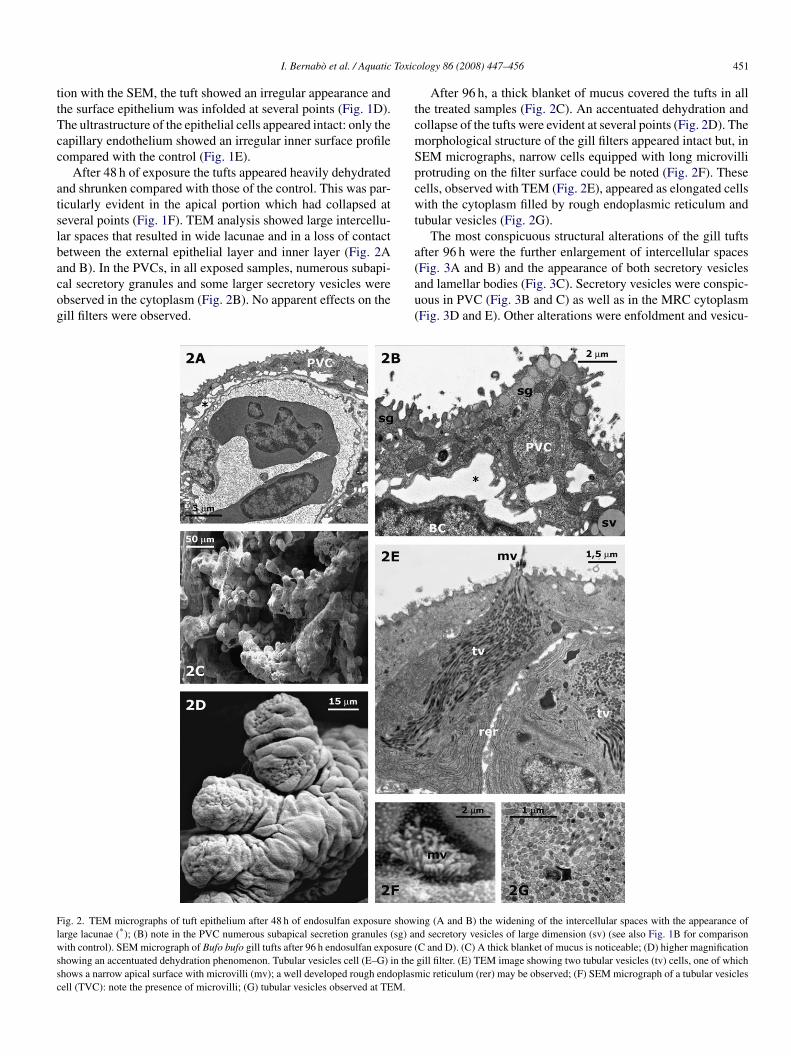
After 48 h of exposure the tufts appeared heavily dehydratednd shrunken compared with those of the control. This was par-icularly evident in the apical portion which had collapsed ateveral points (Fig. 1F). TEM analysis showed large intercellu-ar spaces that resulted in wide lacunae and in a loss of contactetween the external epithelial layer and inner layer (Fig. 2A
nd B). In the PVCs, in all exposed samples, numerous subapi-al secretory granules and some larger secretory vesicles werebserved in the cytoplasm (Fig. 2B). No apparent effects on theill filters were observed.(au(
ig. 2. TEM micrographs of tuft epithelium after 48 h of endosulfan exposure showarge lacunae (*); (B) note in the PVC numerous subapical secretion granules (sg) anith control). SEM micrograph of Bufo bufo gill tufts after 96 h endosulfan exposure (
howing an accentuated dehydration phenomenon. Tubular vesicles cell (E–G) in thehows a narrow apical surface with microvilli (mv); a well developed rough endoplasmell (TVC): note the presence of microvilli; (G) tubular vesicles observed at TEM.
ology 86 (2008) 447–456 451
After 96 h, a thick blanket of mucus covered the tufts in allhe treated samples (Fig. 2C). An accentuated dehydration andollapse of the tufts were evident at several points (Fig. 2D). Theorphological structure of the gill filters appeared intact but, inEM micrographs, narrow cells equipped with long microvillirotruding on the filter surface could be noted (Fig. 2F). Theseells, observed with TEM (Fig. 2E), appeared as elongated cellsith the cytoplasm filled by rough endoplasmic reticulum and
ubular vesicles (Fig. 2G).The most conspicuous structural alterations of the gill tufts
fter 96 h were the further enlargement of intercellular spaces
Fig. 3A and B) and the appearance of both secretory vesiclesnd lamellar bodies (Fig. 3C). Secretory vesicles were conspic-ous in PVC (Fig. 3B and C) as well as in the MRC cytoplasmFig. 3D and E). Other alterations were enfoldment and vesicu-ing (A and B) the widening of the intercellular spaces with the appearance ofd secretory vesicles of large dimension (sv) (see also Fig. 1B for comparisonC and D). (C) A thick blanket of mucus is noticeable; (D) higher magnificationgill filter. (E) TEM image showing two tubular vesicles (tv) cells, one of which
ic reticulum (rer) may be observed; (F) SEM micrograph of a tubular vesicles

452 I. Bernabo et al. / Aquatic Toxicology 86 (2008) 447–456
F othelL ) and
lo
3
l(tittcbmic
le
4
etrcaap
ig. 3. TEM micrographs after 96 h exposure (A–F). Note the alterations of endamellar bodies (lb) and secretory vesicles (sv) are present in pavement cells (C
ation of the endothelium (Fig. 3A and B), as well as appearancef macrophages (Fig. 3F).
.4. iNOS immunolocalization
Immunolocalization of iNOS isoform in the gills revealed noabelling or weak labelling in control samples at all time pointsFig. 4A). In all endosulfan exposed individuals iNOS immunos-aining increased after 24 h compared with control (Fig. 4B). Thentensity of the staining increased at 48 h of exposure at whichime the iNOS reached the maximal expression. This pattern washe same in both tuft and filter epithelium (Fig. 4C). Epithelialells showed a marked gradational reactivity for the enzyme; the
asal cells were heavily stained while, in the surface, they wereoderately or weakly stained (Fig. 4D). Immunopositive cells,ntensely stained, were also recognizable in the lumen of bloodapillaries containing blood cell elements (Fig. 4C and D). The
ifiUA
ium (A: arrowheads) and evident enlargement of the intercellular space (B) (*).in MRC cytoplasm (D and E); macrophage (F).
abelling completely disappeared after 96 h of exposure in allxposed samples (Fig. 4E).
. Discussion
In the present study we investigated for the first time theffects of endosulfan on the gill ultrastructure and on the pat-ern of iNOS expression in the gills of B. bufo tadpoles. Ouresults demonstrate that short-term exposure to a sublethal con-entration of endosulfan (0.2 mg/L) induced iNOS expressionnd seriously affected the structure and ultrastructure of the gillpparatus. The alterations of the gill apparatus became moreronounced with increased exposure time and were most severe
n the tuft which is the main site of respiration, whereas thelters are involved in the feeding mechanism (Hourdry, 1974;chiyama et al., 1990; Brunelli et al., 2004). With regards to thenuran class, the gills apart from having a respiratory function
I. Bernabo et al. / Aquatic Toxicology 86 (2008) 447–456 453
Fig. 4. Confocal micrographs of gill sections labelled with the mouse monoclonal antibody to iNOS (A–E). (A) Control sample reveals no or weak labelling; (B) thelocalization is evident after 24 h in both filter and tuft epithelium. (C and D) The intensity of staining increases at 48 h of exposure when iNOS reaches the maximumo ries (*
a
(titfLiset(spt
dacptW
sfihpma(
alsessiwu
f expression; note immunopositive, intensely stained, cells in the blood capillafter 96 h of exposure.
McIndoe and Smith, 1984), are also involved in osmoregula-ion (Hourdry, 1974; Uchiyama and Yoshizawa, 1992). Takingnto account the important roles of tadpole gills, the alterationshat we observed in B. bufo suggest serious effects of endosul-an in gas exchange and in osmotic and acid–base regulation.imited data are available for comparison with our results to
nvestigate the consequences of pesticides on the respiratoryystem of amphibians. Moreover, there are few studies on theffects of endosulfan in amphibians and additionally about theoxic effects on organs or systems other than the nervous systemEFSA, 2005).) Only one published study looking at Hylidaehowed ultrastructural changes of the gills due to toxicity fromaraquat (Lajmanovich et al., 1998); to our knowledge, this ishe first study of the effects of endosulfan on the amphibian gills.
An apparent increase in mucous secretion was the most evi-ent result of endosulfan application after 24 h and the secretionppeared to increase after 48 and 96 h of exposure. Under normal
onditions, amphibians secrete mucus from the vacuoles of theavement cells; the function of the mucus coating is probablyo facilitate the exchange of gases (Kato and Kurihara, 1988).e suggest that the enhanced mucous secretion could be con-
agWg
) which contain cellular elements (D); (E) the labelling completely disappears
idered as an attempt to counteract uptake of the pollutant. Inshes, increased mucus production in Cirrhinus mrigala gillsas been reported after exposure to malathion, an organophos-horous insecticide (Roy and Munshi, 1991). The excess ofucus, however, impairs respiratory exchange which eventu-
lly can result in hypoxia, as observed in numerous teleosteiFranklin, 1990).
An increase of intercellular spaces, secretory vesicles and theppearance of lamellar bodies were observed in the gill epithe-ium of all treated tadpoles after 48 h. Increased intercellularpaces in the gill epithelium were found in Hylidae tadpoles afterxposure to paraquat (Lajmanovich et al., 1998). These authorsuggested that the increase of intercellular spaces occur by thehift of plasma fluids to intercellular space as a resulting of annflammatory response. The presence of vesicles (e.g. granules)as described in so-called “granule-containing cells” in Trit-rus vittatus gills (Warburg and Rosenberg, 1997). Rosenberg
nd Warburg (1992, 1993) previously described similar round,ranule-containing cells in Pelobates syriacus ventral epidermis.e rarely observed secretory vesicles (i.e. membrane-boundranules according to Warburg and Rosenberg, 1997) in the gill

4 Toxic
envcate
dcpfsaatsmpint
wswawws
idFt
plc(etvtedsaIotaatrwm
b2
dmi2tmat
0it(i4vse
ab
fltaAwe
tnett
A
tf
R
A
A
A
54 I. Bernabo et al. / Aquatic
pithelium of the B. bufo control group. In B. bufo, we couldot describe “granule-containing cells” but we found secretoryesicles (e.g. granules) similar to those found in the “granule-ontaining cells” of urodeles. In B. bufo the secretory vesiclesppear along with lamellar bodies in PVCs and MRCs. In par-icular they appear first in PVC and then in MRC after 96 hxposure.
Lamellar bodies are sub-cellular structures that have beenetected in various cell types under normal and pathologicalonditions. The best known lamellar bodies are produced byneumocyte cells in the lung alveoli to provide the lung sur-actant system (Schmitz. and Muller, 1991). In non-mammals,urfactant appears to act as anti-glue, preventing the adhesion ofdjacent respiratory surfaces that occur when the lungs collapsend cannot be reinflated. Without anti-glue to lower the surfaceension of the fluid intervening between the contacting epithelialurfaces, the process of inspiration after lung collapse would beore difficult (Daniels et al., 1998; Hermida et al., 2002). No
revious study has demonstrated the presence of lamellar bodiesn amphibian gills and the role of these sub-cellular structures isot explainable on the basis of our data. More studies are neededo clarify this topic.
The most remarkable modification noted in the gill filtersas the appearance of tubular vesicle cells after 96 h expo-
ure that were not observed in the control animals. These cellsere observed to have the typical features of secreting cells,
nd have never been described previously. Their ultrastructureas characterized by a narrow opening on the outer surface andell developed RER and Golgi apparatus, features indicating
ecreting activity.We therefore suppose that the development of this cell type
s involved in the increased mucous secretion as a part of aefensive response of the gill epithelium to endosulfan exposure.urther analyses are needed to elucidate the functional role of
his cell.It is well known that iNOS plays an important role in many
athophysiological processes, including various forms of circu-atory shock, the regulation of vascular tone, inflammation, andarcinogenesis (Nathan and Xie, 1994). Fenoglio and colleaguesFenoglio et al., 2006) found an increase in NOS/NADPHxpression in keratinocytes and in MRCs in the epidermis ofhe frog Rana kl. esculenta living in polluted areas. This obser-ation may be indicative of the ability of the frog skin to respondo oxidative stress in polluted areas. In this study a marked differ-nce in the intensity and localization of iNOS is observed underifferent exposure times; in control samples the epithelial cellshow no or an extremely weak labelling. The intensity increasedfter 24 and 48 h when iNOS reached the maximum expression.n our observations, induction of iNOS may reflect the degreef hypoxia and inflammation. Moreover, iNOS is closely relatedo defence mechanisms against infectious organisms (Dezfuli etl., 2003; Kwon et al., 2001). It also regulates the functionalctivity, growth and death of many immune and inflamma-
ory cell types including macrophages (Coleman, 2001). Earliereports have showed that treatment of murine macrophagesith DDT, chlordane, endosulfan and methyl parathion aug-ented the levels of nitrite but this will have to be confirmedA
ology 86 (2008) 447–456
y further investigations (Crittenden et al., 1998; Ayub et al.,003).
Additional effects of endosulfan in the gills included theegeneration of epithelial cells and an increased number ofacrophages. We suggest a short-term effect of the nitric oxide
n regulating the blood flow through the gill (Ebbesson et al.,005) and in contributing to the modulation of an inflamma-ory response. Han et al. (2007) showed that treatment of mouse
acrophages with endosulfan induces the expression of iNOSnd proinflammatory cytokines in mammals, and suggested thathis insecticide has inflammatory potential.
The LC50 value for endosulfan in B. bufo tadpoles was.43 mg/L, which is in the range of previously published stud-es. The endosulfan 96 h LC50 values were 0.0018 mg/L in Ranaigrina (Gopal et al., 1981), 0.123 mg/L in Bufo melanostictusVardia et al., 1984), higher than 2.35 mg/L for metamorphos-ng Rana pipiens tadpoles (Harris et al., 2000), and higher than.7 mg/L for Rana clamitans tadpoles (Harris et al., 1998). Thisariation in LC50 value could result from species differences inensitivity or from differences in testing methodologies (Berrillt al., 1998; Bridges and Semlitsch, 2000).
To conclude, this laboratory study is a relevant preliminarynalysis that has been useful to establish the sensitivity of B.ufo larvae to acute endosulfan exposure.
In our study, mortality and relevant gill alterations resultedrom endosulfan application after exposure to concentrationsower than the concentrations of insecticide found near agricul-ural fields (0.7 mg/L) or in a tidal creek water column followinggricultural runoff (0.5 mg/L) (Ernst et al., 1991; EPA, 2002).s a whole, the study presented here is a preliminary analysis,hich will continue to look at the effects of chronic exposure to
ndosulfan at environmentally relevant concentrations.Furthermore, assessing the risk to amphibians of concentra-
ions potentially encountered in agricultural areas or in theiratural environment and studying sublethal effects of endosulfanxposure is particularly important in light of current conserva-ion efforts to limit the loss of biodiversity and to fully understandhe role of pesticides in the decline of amphibian populations.
cknowledgements
This study was supported in part by a grant provided by “Dot-orato di Ricerca in Biologia Animale”. We thank Enrico Perrottaor technical assistance.
eferences
lford, R.A., Richards, S.J., 1999. Global amphibian decline: a problem inapplied ecology. Annu. Rev. Ecol. Syst. 30, 133–165.
nand, M., Agrawal, A.K., Gopal, K., Sur, R.N., Seth, P.K., 1986. Endosulfanand cholinergic (muscarinic) transmission: effect on electroencephalogramsand [3H] quinuclidinyl benzilate in pigeon brain. Environ. Res. 40, 421–426.
merican Society for Testing Material (ASTM), 1997. Standard practice forconducting acute toxicity tests with fishes, macroinvertebrates, and amphib-
ians. E 729–790. In American Society for Testing and Materials Standards,Philadelphia.yub, S., Verma, J., Das, N., 2003. Effect of endosulfan and malathion on lipidperoxidation, nitrite and TNF-a release by rat peritoneal macrophages. Int.Immunopharmacol. 3, 1819–1828.

Toxic
B
B
B
B
B
B
B
B
B
C
C
C
C
C
C
C
D
D
D
D
D
E
E
E
E
E
E
F
F
F
F
G
G
G
G
G
H
H
H
H
H
H
I
I. Bernabo et al. / Aquatic
arni, S., Boncompagni, E., Grosso, A., Bertone, V., Freitas, I., Fasola, M.,Fenoglio, C., 2007. Evaluation of Rana snk esculenta blood cell responseto chemical stressors in the environment during the larval and adult phases.Aquat. Toxicol. 81, 45–54.
eebee, T.J.C., Griffiths, R.A., 2005. The amphibian decline crisis: a watershedfor conservation biology? Biol. Conserv. 125, 271–285.
errill, M., Bertram, S., McGillivray, L., Kolohon, M., Pauli, B., 1994. Effectsof low concentrations of forest use pesticides on frog embryos and tadpoles.Environ. Toxicol. Chem. 13, 657–664.
errill, M., Coulson, D., McGillivray, L., Pauli, B., 1998. Toxicity of endo-sulfan to aquatic stage of anuran amphibians. Environ. Toxicol. Chem. 9,1738–1744.
laustein, A.R., Kiesecker, J.M., 2002. Complexity in conservation: lessonsfrom the global decline of amphibian populations. Ecol. Lett. 5, 597–608.
ridges, C.M., Semlitsch, R.D., 2000. Variation in pesticide tolerance of tad-poles among and within species of Ranidae and patterns amphibian decline.Conserv. Biol. 14, 490–1499.
roomhall, S., 2002. The effects of endosulfan and variable water temperatureon survivorship and subsequent vulnerability to predation in Litoria citropatadpoles. Aquat. Toxicol. 61, 243–250.
roomhall, S., 2004. Egg temperature modifies predator avoidance and theeffects of the insecticide endosulfan on tadpoles of an Australian frog. J.Appl. Ecol. 41, 105–113.
runelli, E., Perrotta, E., Tripepi, S., 2004. Ultrastructure and development of thegills in Rana dalmatina (Amphibia, Anura). Zoomorphology 123, 203–211.
arey, C., Bryant, C.J., 1995. Possible interrelations among environmentaltoxicants, amphibian development, and decline of amphibian populations.Environ. Health Perspect. 103, 13–17.
arrier, J.A., Beebee, T.J.C., 2003. Recent, substantial, and unexplained declinesof the common toad Bufo bufo in lowland England. Biol. Conserv. 111,395–399.
lark, E.J., Norris, D.O., Jones, R.E., 1998. Interactions of gonadal steroidsand pesticides (DDT, DDE) on gonaduct growth in larval tiger salamanders,Ambystoma tigrinum. Gen. Comp. Endocrinol. 109, 94–105.
oats, J.R., 1990. Mechanisms of toxic action and structure-activity relation-ships for organochlorine and synthetic pyrethroid insecticides. Environ.Health Perspect. 87, 255–262.
oleman, J.W., 2001. Nitric oxide in immunity and inflammation. Int.Immunopharmacol. 1, 1397–1406.
oons, A.H., Leduc, E.H., Connolly, J.M., 1955. Studies on antibody. I. Amethod for the histochemical demonstration of specific antibody and itsapplication to a study of the hyperimmune rabbit. J. Exp. Med. 102, 49–59.
rittenden, P.L., Carr, R., Pruett, S.B., 1998. Immunotoxicological assessmentof methyl parathion in female B6C3F1 mice. J. Toxicol. Environ. Health 54,1–20.
aniels, C.B., Lopatko, O.V., Orgeig, S., 1998. Evolution of surface activityrelated functions of vertebrate pulmonary surfactant. Clin. Exp. Pharmacol.Physiol. 25, 716–721.
avidson, C., Shaffer, H.B., Jennings, M.R., 2001. Declines of the Californiared-legged frog: climate, UVB, habitat and pesticides hypothesis. Ecol. Appl.11, 464–479.
avidson, C., Shaffer, H.B., Jennings, M.R., 2002. Spatial tests of the pesti-cide drift, habitat destruction, UV-B, and climate-change hypotheses forCalifornia amphibian declines. Conserv. Biol. 16, 1588–1601.
ezfuli, B.S., Giari, L., Konecny, R., Jaeger, P., Manera, M., 2003. Immuno-histochemistry, ultrastructure and pathology of gills of Abramis brama fromLake Mondsee, Austria, infected with Ergasilus sieboldi (Copepoda). Dis.Aquat. Org. 53, 257–262.
uellman, W.E., Trueb, L., 1986. Biology of Amphibians. McGraw-Hill, NewYork.
bbesson, L.E., Tipsmark, C.K., Holmqvist, B., Nilsen, T., Andersson, E., Ste-fansson, S.O., Madsen, S.S., 2005. Nitric oxide synthase in the gill of Atlantic
salmon: colocalization with and inhibition of Na+, K+-ATPase. J. Exp. Biol.208, 1011–1017.nvironmental Protection Agency (EPA), 1999. LC50 Software Program, Ver-sion 1. 00. Center for Exposure Assessment Modeling (CEAM) DistributionCenter.
J
ology 86 (2008) 447–456 455
nvironmental Protection Agency (EPA), 2002. Reregistration Eligibility Deci-sion for Endosulfan. July 31, 2002. United States Environmental ProtectionAgency, Office of Prevention, Pesticides, and Toxic Substances.
rnst, W.R., Jonah, P., Doe, K., Julien, G., Hennigar, P., 1991. Toxicity to aquaticorganisms of off-target deposition of endosulfan applied by aircraft. Environ.Toxicol. Chem. 10, 103–114.
uropean Commission, 2004. Monitoring of Pesticide Residues in Products ofPlant Origin in the European Union. Iceland and Lichtenstein, Norway.
uropean Food Safety Authority (EFSA), 2005. Opinion of the scientific panelon contaminants in the food chain on a request from the commission relatedto endosulfan as undesirable substance in animal feed. EFSA J. 234, 1–29.
agotti, A., Morosi, L., Di Rosa, I., Clarioni, R., Simoncelli, F., Pascolini, R., Pel-legrino, R., Guex, G.D., Hotz, H., 2005. Bioaccumulation of organochlorinepesticides in frogs of the Rana esculenta complex in central Italy. AmphibiaReptilia 26, 93–104.
enoglio, C., Grosso, A., Boncompagni, E., Milanesi, G., Gandini, C., Barni, S.,2006. Morphofunctional evidence of changes in principal and mitochondria-rich cells in the epidermis of the frog Rana kl. esculenta living in a pollutedhabitat. Arch. Environ. Contam. Toxicol. 51, 690–702.
inney, D.J., 1971. Probit Analysis, third ed. Cambridge University Press, NewYork, p. 668.
ranklin, G.E., 1990. Surface ultrastructure changes in the gills of sockeyesalmon (Teleostei: Oncorhynchus nerka) during seawater transfer: compar-ison of successful and unsuccessful seawater adaptation. J. Morphol. 206,13–23.
erman Federal Environment Agency, 2004. Draft Dossier Prepared in Supportof a Proposal of Endosulfan to be Considered as a Candidate for Inclu-sion in the UN-ECE LRTAP Protocol on Persistent Organic Pollutants.Umweltbundesamt, Berlin.
iusi, G., Facciolo, R.M., Alo, R., Carelli, A., Madeo, M., Brandmayr, P.,Canonaco, M., 2005. Some environmental contaminants influence motorand feeding behaviors in the Ornate wrasse (Thalassoma pavo) via dis-tinct cerebral histamine receptor subtypes. Environ. Health Perspect. 113,1522–1529.
opal, K., Khanna, R.N., Anand, M., Gupta, G.S.D., 1981. The acute toxicityof endosulfan to freshwater organism. Toxicol. Lett. 7, 453–456.
opal, K., Anand, M., Mehrotra, S., Ray, P.K., 1985. Neurobehavioral changesin fresh water fish Channa punctatus exposed to endosulfan. J. Adv. Zool.5, 74–80.
osner, K.L., 1960. A simplified table for staging anuran embryos and larvaewith notes on identification. Herpetologica 16, 183–190.
an, E.H., Hwang, Y.P., Kim, H.G., Jeong, H.G., 2007. Inflammatory effect ofendosulfan via NF-jB activation in macrophages. Biochem. Biophys. Res.Commun. 355, 860–865.
arris, M.L., Bishop, C.A., Struger, J., Ripley, B., Bogart, J.P., 1998. Thefunctional integrity of northern leopard frog (Rana pipiens) and green frog(Rana clamitans) populations in orchard wetlands. II. Effects of pesticidesand eutrophic conditions on early life stage development. Environ. Toxicol.Chem. 17, 1351–1363.
arris, M.L., Chora, L., Bishop, C.A., Bogart, J.P., 2000. Species- and age-related differences in susceptibility to pesticide exposure for two amphibians,Rana pipiens and Bufo americanus. Bull. Environ. Contam. Toxicol. 64,263–270.
ermida, G.N., Farıas, A., Fiorito, L.E., 2002. Ultrastructural characteristics ofthe lung of Melanophryniscus stelzneri stelzneri (Weyenberg, 1875) (Anura,Bufonidae). Biocell 26, 347–355.
oulahan, J.E., Findlay, C.S., Schmidt, B.R., Meyer, A.H., Kuzmin, S.L., 2000.Quantitative evidence for global amphibian population decline. Nature 404,752–755.
ourdry, J., 1974. Etude des branchies “internes” puis de leur regression aumoment de la metamorphose, chez la larve de Discoglossus pictus (OTTH),Amphibien Anoure. J. Microsc. Paris 20, 165–182.
nternational Union for Conservation of Nature (IUCN)., 2007. The 2007 IUCN
Red List of Threatened Animals. International Union for Conservation ofNature, Gland, Switzerland (available from http://www.redlist.org).ohnson, W.W., Finley, M.T., 1980. Handbook of Acute Toxicity of Chemicalsto Fish and Aquatic Invertebrates. U.S. Department of Interior, Fish andWildlife Service, Washington, DC (Resource Publication 137).

4 Toxic
J
K
K
K
K
K
L
L
L
L
M
M
M
N
N
P
P
R
R
R
S
S
S
S
U
U
V
Venturino, A., Rosenbaum, E., Caballero de Castro, A., 2003. Biomarkers ofeffect in toads and frogs. Biomarkers 8, 167–186.
56 I. Bernabo et al. / Aquatic
ohnson, M.S., Vodela, J.K., Reddy, G., Holladay, S.D., 2000. Fate and thebiochemical effects of 2,4,6-trinitrotoluene exposure to tiger salamanders(Ambystoma tigrinum). Ecotoxicol. Environ. Saf. 46, 186–191.
ato, S., Kurihara, K., 1988. Electron microscopic studies of the external gillepithelium of the salamander, Hynobius dunni Tago, followed throughoutlarval life. Okajimas Folia Anat. Jpn. 65, 177–202.
iesecker, J.M., Blaustein, A.R., Belden, L.K., 2001. Complex causes ofamphibian population declines. Nature 410, 681–684.
im, J.Y., Choi, C.Y., Lee, K.J., Shin, D.W., Jung, K.S., Chung, Y.C., Lee, S.S.,Shin, J.G., Jeong, H.G., 2004. Induction of inducible nitric oxide synthaseand proinflammatory cytokines expression by o,p-DDT in macrophages.Toxicol. Lett. 147, 261–269.
im, J.Y., Oh, K.N., Han, E.H., Kim, D.H., Jeong, T.C., Lee, E.S., Jeong, H.G.,2005. Methoxychlor-induced inducible nitric oxide synthase and proinflam-matory cytokines expression in macrophages via NF-jB, ERK, and p38mitogen-activated protein kinases. Biochem. Biophys. Res. Commun. 333,1234–1240.
won, S., Newcomb, R.L., George, S.C., 2001. Mechanisms of synergisticcytochine-induced nitric oxide production in human alveolar epithelial cells.Nitric Oxide 5, 534–546.
ajmanovich, R.C., Izaguirre, M.F., Casco, V.H., 1998. Paraquat tolerance andalteration of internal gill structure of Scinax nasica tadpoles (Anura: Hyli-dae). Arch. Environ. Contam. Toxicol. 34, 364–369.
ajmanovich, R.C., Cabagna, M., Peltzer, P.M., Stringhini, G.A., Attademo,A.M., 2005. Micronucleus induction in erythrocytes of the Hyla pulchellatadpoles (Amphibia: Hylidae) exposed to insecticide endosulfan. Mutat. Res.587, 67–72.
akshmana, M.K., Raju, T.R., 1994. Endosulfan induces small but significantchanges in the levels of noradrenaline, dopamine and serotonin in the devel-oping rat brain and deficits in the operant learning performance. Toxicology91, 139–150.
angston, W.J., Chesman, B.S., Burt, G.R., Pope, N.D., McEvoy, J., 2002. Met-allothionein in liver of eels Anguilla anguilla from the Thames Estuary: anindicator of environmental quality? Mar. Environ. Res. 53, 263–293.
aterna, E.J., Rabeni, C.F., Lapoint, T.W., 1995. Effects of the syntheticpyrethroid insecticide, esfenvalerate, on larval leopard frogs (Rana spp.).Environ. Toxicol. Chem. 14, 613–622.
cIndoe, R., Smith, D.J., 1984. Functional anatomy of the internal gills of thetadpole of Litoria ewingii (Anura, Hylidae). Zoomorphology 104, 280–291.
oncada, S., Palmer, R.M., Higgs, E.A., 1992. Nitric oxide: physiology, patho-physiology, and pharmacology. Pharm. Rev. 43, 109–142.
aqvi, S.M., Vaishnavi, C., 1993. Bioaccumulation potential and toxicity ofendosulfan insecticide to not-target animals. Comp. Biochem. Physiol. C105, 347–361.
WW
ology 86 (2008) 447–456
athan, C., Xie, Q., 1994. Nitric oxide 3 synthases: roles, tolls, and controls.Cell 78, 915–918.
aul, V., Balasubramaniam, E., 1997. Effects of single and repeated adminis-tration of endosulfan on behaviour and its interaction with centrally actingdrugs in experimental animals: a mini review. Environ. Toxicol. Pharmacol.3, 151–157.
avignano, I., Giacoma, C., 1990. Una valutazione ecologica dello stato di con-servazione delle zone umide minori della pianura Piemontese – Atti VIConv. naz. Ass. “A. Chigi”, Torino (1989). Mus. reg. Sci. Nat. Torino., 91–104.
osenberg, M., Warburg, M.K., 1992. Ultrastructure and histochemistry of ven-tral epidermis in Pelobates syriacus (Anura; Pelobatidae) tadpoles. Biol.Struct. Morphogen. 4, 164–170.
osenberg, M., Warburg, M.K., 1993. The ventral epidermis of Pelobates syri-acus (Anura; Pelobatidae). Isr. J. Zool. 39, 235–243.
oy, P.K., Munshi, J.S.D., 1991. Malathion induced structural and morphometricchanges of gills of a fresh water major carp, Cirrhinus mrigala. J. Environ.Biol. 12, 79–87.
chmitz, G., Muller, G., 1991. Structure and function of lamellar bodies, lipid-protein complexes involved in storage and secretion of cellular lipids. J.Lipid Res. 32, 1539–1570.
ingh, N., Singh, C.P., Kumar, H., Brat, G.K., 1992. Endosulfan poisoning: astudy of 22 cases. J. Assoc. Phys. India 40, 87.
parling, D.W., Fellers, F.M., McDonnell, L., 2001. Pesticides and amphib-ian population declines in California, USA. Environ. Toxicol. Chem. 7,1591–1595.
tuart, S.N., Chanson, J.S., Cox, N.A., Young, B.E., Rodrigues, A.S., Fischman,D.L., Waller, R.W., 2004. Status and trends of amphibian declines andextinctions worldwide. Science 306, 1783–1786.
chiyama, M., Yoshizawa, H., Wakasugi, C., Oguro, C., 1990. Structure ofinternal gills in tadpoles of the crab-eating frog, Rana cancrivora. Zool. Sci.7, 623–630.
chiyama, M., Yoshizawa, H., 1992. Salinity tolerance and structure of externaland internal gills in tadpoles of the crab-eating frog, Rana cancrivora. CellTissue Res. 267, 35–44.
ardia, H.K., Rao, P.S., Durve, V.S., 1984. Sensitivity of toad larvae to 2,4-Dand endosulfan pesticides. Arch. Hydrobiol. 100, 395–400.
ake, D.B., 1991. Declining amphibian populations. Science 253, 60.arburg, M.K., Rosenberg, M., 1997. Structure of gill epithelium in Triturus
vittatus larvae. Ann. Anat. 179, 57–64.
Copyright © 2022 FDOKUMEN




















