
Diet and Foraging Mode of Bufo Marinus and Leptodactylus ocellatus
Upload
fieldmuseumCategory
view
2download
0
Biochemical Genetics, Vol. 12, No. 2, 1974
Genetic Variation and Population Ecology of Some Southeast Asian Frogs of the Genera Bufo and Rana
Robert F. Inger, 1 Harold K. Voris, ~ and Helen H. Voris ~
Received 14 Feb. 1974IFinal 16 Apr. 1974
The relationship between population structure and amount of genetic variation within populations has been investigated in seven species of anurans from South- east Asia. Two populations each of Bufo asper, B. parvus, B. melanostictus, Rana blythi, and R. erythraea and one population each of R. hosei and R. laticeps were studied," all except B. melanostictus and R. erythraea are forest species. Information on distributional characteristics such as general habitat, form of the adult range, the relationships of nonbreeding range to breeding site, the tendency to aggregate at the breeding site, and the size of the breeding aggregation was used to outline major aspect~ of the breeding structure of each species and predict relative amounts of genic variation. Genetic variation was estimated from measures of protein polymorphism as detected through gel electrophoresis. The assays used were liver EST, FUM, GDH, LAP, LDH, MDH, and total protein; muscle EST, GOT, MDH, and total protein; plasma EST and total protein. The pattern of variation of the populations of Bufo species agreed with predictions based on population structure, with B. asper the least polymorphic, B. melanostictus the most polymorphic, and B. parvus intermediate. Interspec(fic differenees were not as great in Rana as in Bufo. Although R. erythraea was more variable than R. blythi in terms of mean number of morphs per zone (as predicted from distributional characteristics), R. blythi possessed polymorphism at more zones. R. hosei was the most polymorphic Rana studied, but distributional data are insuJficient to explain why this was the case.
K E Y W O R D S : e l e c t r o p h o r e s i s ; e c o l o g y ; A s i a n f r o g s ; g e n e t i c v a r i a t i o n .
T h i s r e s e a r c h w a s s u p p o r t e d b y a g r a n t f r o m the N a t i o n a l Sc i ence F o u n d a t i o n ( G B 7845X) . J F i e l d M u s e u m o f N a t u r a l H i s t o r y , C h i c a g o , I l l ino is .
121
© 1974 Plenum Publishing Corporation, 227 West 17th Street, New York, N.Y. 10011. No part of this publica- tion may be reproduced, stored in a retrieval system, or transmitted, in any form or by any means, electronic, mechanical, photocopying, microfilming, recording, or otherwise, without written permission of the publisher.

122 Inger, Voris, and Voris
INTRODUCTION
An intensive quest for explanations of the varying amounts of protein polymorphism observed in natural populations of both plants and animals is currently under way. Investigations have been undertaken on factors such as the relation to morphological (Lakovaara et al., 1972) and cytological (Nair et al., 1971) variation, postulated phylogenetic relations (Hubby and Throckmorton, 1965), geographic patterns in protein variation (Dessauer and Nevo, 1969; Johnson and Selander, 1971; Prakash et al., 1969), and population ecology within species (Petras, 1967). Multispecies studies have, in the main, been directed toward phylogenetic or taxonomic implications of patterns of protein polymorphism and hence of genetic variation.
Attempts to relate interspecific differences in patterns of protein variation to ecological differences have been relatively few. It is this aspect of the general problem that is the focus of our work. The work by Johnson and Selander (1971) on kangaroo rats (Dipodomys), though concerned primarily with other relations, exposed a rough correlation between coarse habitat distribution and genetic variation. But the quality of their ecological information pre- vented more than general comments. The merit of our study, if any, is that our ecological data are stronger and therefore allow deeper investigation of the relationships between ecological and protein (and genetic) variation within groups of related species.
The work on aspects of population structure in relation to genetic variation has its base in Sewall Wright's models. Theoretically, closely related species that are similar in most aspects of their ways of life but differ in terms of the dimensionality (linear vs. two-dimensional) of their adult ranges, effective neighborhood size, effective population size, or other critical factors that would affect gene flow directly should show different patterns of genetic variation. It has been the purpose of this study to relate predictions based on Wright's models and our knowledge of the structure and ecology of certain frog populations to protein variation.
Gel electrophoresis is the most realistic yet practical means of detecting genetic variation currently available (Hubby and Lewontin, 1966), and it has been used to provide our estimates of variation. Dessauer and Nevo (1969) used the same technique to analyze genetic variation in two closely related species of frogs (genus Aeris), but their emphases were macrogeographic patterns and interspecific differences in allelic frequencies.
MATERIALS AND METHODS
Animals Studied
Three species of Bufo and four of Rana, collected at six localities within 22
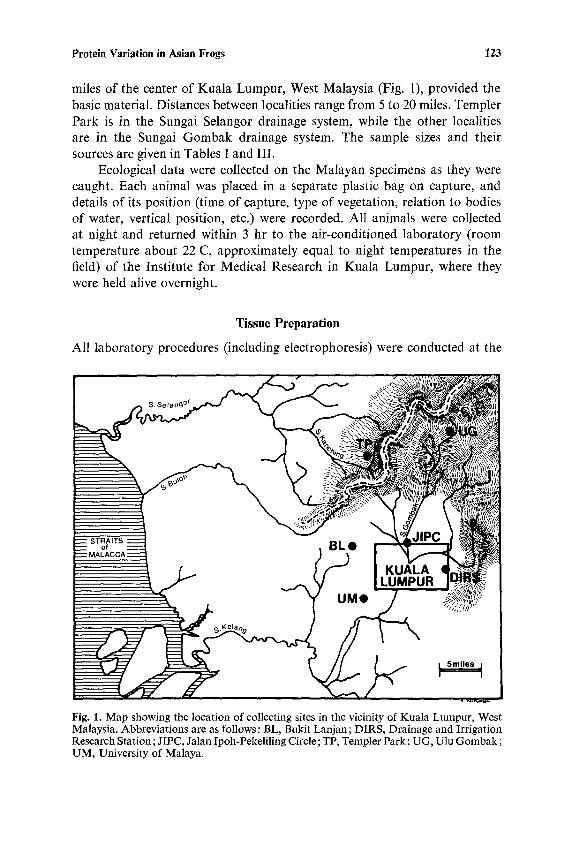
Protein Variation in Asian Frogs 123
miles of the center of Kuala Lumpur, West Malaysia (Fig. 1), provided the basic material. Distances between localities range from 5 to 20 miles. Templer Park is in the Sungai Selangor drainage system, while the other localities are in the Sungai Gombak drainage system. The sample sizes and their sources are given in Tables I and III.
Ecological data were collected on the Malayan specimens as they were caught. Each animal was placed in a separate plastic bag on capture, and details of its position (time of capture, type of vegetation, relation to bodies of water, vertical position, etc.) were recorded. All animals were collected at night and returned within 3 hr to the air-conditioned laboratory (room temperature about 22 C, approximately equal to night temperatures in the field) of the Institute for Medical Research in Kuala Lumpur, where they were held alive overnight.
Tissue Preparation
All laboratory procedures (including electrophoresis) were conducted at the
Fig. 1. Map showing the location of collecting sites in the vicinity of Kuala Lumpur, West Malaysia. Abbreviations are as follows: BL, Bukit Lanjan; DIRS, Drainage and Irrigation Research Station; JIPC, Jalan Ipoh-Pekeliling Circle; TP, Templet Park; UG, Ulu Gombak; UM, University of Malaya.

124 Inger, Voris, and Voris
Institute for Medical Research. Plasma, liver, and muscle samples were pre- pared from each animal within 72 hr of capture. Each specimen was double- pithed, the body cavity was opened, and a blood sample was taken from the heart using a 1-ml syringe and 25-gauge needle rinsed in sodium heparin. 'l-he blood sample was centrifuged, and the plasma was drawn off and frozen in 2-ml polystyrene conical sample containers at approximately -20 C for 1-4 weeks. In the case of very small plasma samples (< 0.05 ml), a 40% sucrose solution containing a small amount of bromophenol blue was added in the ratio 6 parts plasma to 1 part sucrose solution before freezing. This sucrose solution was also added to the other plasma samples after thawing, just prior to electrophoresis. Liver and muscle samples were removed, rinsed in cold 0.7% saline, blotted on filter paper, quick-frozen on dry ice, weighed, and stored frozen for 1-2 days. Then they were placed on cold glass plates, minced with a razor blade, and mixed with either cold 0.075 M tris(hydroxymethyl)aminomethane glycine (pH 8.9) (tris glycine buffer), 4% sucrose, 1% MgC1 z or 0.09 M tris(hydroxymethyl)aminomethane disodium(ethylenedinitrilo)tetraacetate borate (pH 8.4) (trisNa-EDTA borate buffer), 4 ~ sucrose, I% MgC12 in the ratio 1 g tissue to 2 ml buffer. Muscle samples were further macerated by homogenizing with a motorized unit for approximately 30-45 sec using a ground-glass 2-ml tube and pestle suspended in an ice bath. Both liver and muscle were then immediately transferred to 0.2- to 1-ml centrifuge tubes, quick-frozen in -60 C acetone, stored frozen on dry ice for 1-2 days, and thawed, after which a layer of toluene was added and the sample was shaken and centrifuged 3 min at 21,500 g. The supernatant was drawn off and recentrifuged for 5 rain at 21,500 g, and the final supernatant was frozen and stored in the polystyrene sample containers at -20 C for 1-4 weeks prior to electrophoresis.
Electrophoresis Procedures
Samples were thawed a few minutes before application to polyacrylamide gels held in E.C. survey model vertical cells. Five and one-half percent gels were prepared using cyanogum-41 gelling agent (electrophoresis grade of 95% acrylamide and 5% bisaerylamide) and either 0.15 M tris glycine (pH 8.9) buffer or 0.18 M tris Na-EDTA borate (pH 8.4) buffer, diluted 1:1 with distilled water (total volume 280 ml) with 1.1 ml 28% ammonium persulfate and 0.31 ml N,N,N'-tetremethylethylenediamine (TEMED) added to achieve polymerization. Buffer corresponding to that used in the gel was placed full strength in the gel cell buffer chambers. Electrophoresis was carried out at 400 v for 141--3 hr, at - 2 to - 4 C. Running time and milliampere differences (20-140 ma) varied depending on which buffer was used, which in turn was determined by the tissue and the enzyme being assayed. After electrophoresis

Protein Variation in Asian Frogs 125
each gel was cut into two slabs, each containing ten separations. Next the gels were removed to a total protein stain or an enzyme assay incubation mixture.
Enzyme Assay Procedures
The choice of assays was based on the availability of histochemical staining methods and the clarity of the results with amphibian materials. The assay recipes used are modifications of those found in the literature: Dessauer (1966), Shaw and Koen (1968), and Shaw and Prasad (1970).
Esterases (EST) were studied in plasma (P), liver (L), and muscle (M). ]-he esterase gels were run approximately 2 hr using the tris borate buffer system. Lactic dehydrogenase (LDH) was studied in liver (L), and these gels were run approximately 2½ hr using the tris borate buffer system. Glutamate dehydrogenase (GDH) was studied in liver (L), and gels were run approxi- mately 21 hr using the tris glycine buffer system. Malic dehydrogenase (MDH) was studied in liver (L) and muscle (M). The MDH gels were run approximately 2¼ hr using the tris borate buffer system. Fumarase (FUM) was studied in the liver (L) of Bufo species only. These gels were run using tris glycine buffer for 2¼ hr. Glutamic-oxaloacetate transaminase (GOT) was studied in muscle (M), and these gels were run using the tris borate buffer system for 2¼ hr. In GOT gels, bands rapidly diffused a short while after appearing. In these cases, drawings of the band positions were made while the gel aplzeared maximally developed, and later analysis was largely on the basis of the drawings. Leucine aminopeptidase (LAP) was studied in liver (L) in Bufo and in muscle (M) in Rana. Gels were run approximately 2 hr using the tris borate buffer system. Total protein (TP) was studied in plasma (P), muscle (M), and liver (12) in Rana, but only in plasma and muscle in Bufo. These gels were run approximately 2¼ hr using the tris glycine buffer system and then placed for 1 hr in 0.2% amido black stain at 50 C. They were then destained in a mixture of 5 parts methanol, 5 parts water, 1 part glacial acetic acid for 24 hr.
On completion of incubation, all gels were transferred to a preservative (described as destain solution above) for 24 hr and then sealed in plastic bags and photographed. Gels were shipped back to the United States and were scored there 9-16 months after running.
Analysis of Electrophoretic Data
Gels were scanned on a light table or white enamel pan; the former allowed closely spaced high-activity bands to be discriminated, and the latter em- phasized the lighter bands.
Some of the assays (e.g., GDH, MDH, FUM) showed only a single area

126 Inger, Voris, and Voris
of activity consisting of one or more bands. Others (e.g., TP) showed several areas of activity, that is, groups of one or more bands which migrated different distances through the gel, leaving clear spaces between the groups. These broad divisions of activity separated by areas lacking in bands were designated as zones 2 and numbered in ascending order from the origin. Each pattern of bands within a zone was termed a morph. Some tissue-assay combinations showed only a single morph throughout a population, others showed nearly as many morphs as individuals studied. A zone was considered to show variation if more than one morph was present even if a variant was exhibited only by a single individual.
With limited laboratory facilities, frogs were collected, tissues sampled, and gels run in succession. Repeat runs or runs including new combinations of samples were not possible. Identification of genetic homozygotes and heterozygotes was thus impossible in most cases. Because of this limitation and the fact that morphs often could not be determined to be identical between populations, the designation of discrete loci and alleles is inappropriate. In some cases (e.g., MDH and LDH), our zones are almost certainly equivalent to single loci and the morphs observed equivalent to allozymes. However, in others (e.g., esterases), a zone may be comprised of several loci. Thus estimates of variation for such a zone may be inflated disproportionately when com- pared to estimates of alleles per locus found in various literature sources. In this study, zones are definitely the same between populations of the same species, and an inspection of Tables I and III reveals that the number of zones present for interspecific comparisons within each genus does not differ greatly. In Bufo 16 of 24 zones are identifiable in all populations, and in Rana 16 of 19 zones are likewise identifiable. Genetic comparability of zones between species is not implied here, although it is likely the case in many instances.
RESULTS
Summary of Frog Ecology
The information presented here was drawn from observations made in Malaya during 1970 and 1971, and in Borneo during 1962 and 1963. Much of the information has been previously published (Inger, 1966, 1969).
Since the behavior of the Malayan individuals of these species appears generally identical to that of their Bornean counterparts, we rely heavily on the Bornean data in the description of ecology and population structure because they are most extensive.
Bufo asper is a large toad (adult snout-vent length 70-120 +ram) that
2 A similar approach and vocabulary have been used by Ruddle et al. (1966).

Protein Variation in Asian Frogs 127
occurs in primary and secondary rainforests from peninsular Thailand and Burma to Java and Borneo. It is a strictly riparian species living on banks of small to medium-sized streams (width 3-30 m) and large rivers having slow, moderate, or swift currents. The majority of individuals of this species are found within 1.3 m of stream beds, and of the 920 collected in Borneo only four were collected more than 10 m from a stream bed. In Malaya, B. asper was caught an average of 1.97 m from stream beds (n = 59, s = 4.51). Bufo asper does not form aggregations, and individuals are usually scattered along stream banks. For three surveyed forest streams in Borneo, the 727 observa- tions of this species were almost evenly divided among 90-m intervals of the 650-m study area on each stream. Median home ranges varied from 43 to 75 m (n = 106) on these Bornean streams. Though its breeding habits are not well known, B. asper apparently lays its eggs in quiet portions of streams, as transforming young were found at the edges of side pools of Bornean creeks.
Bufo parvus is a small toad (adults 30-45 ram) occurring from peninsular Thailand and Burma through the Malaya peninsula. This form ranges broadly in forests (including some rubber plantations) but breeds only in pools alongside small streams (width <10 m). Many adults (262) were caught dispersed on the forest floor more than 30 m from streams at Bukit Lanjan. At Templer Park, 23 of 25 individuals were obtained from breeding aggre- gations (calling, pairs in amplexus, and eggs observed) in rocky pools beside a small creek. Two lone individuals on the forest floor more than 10 m from the nearest stream completed the sample. Bufo parvus is very similar to Bornean populations of B. biporcatus in morphology and behavior, and our opinion now is that they are probably conspecific. Observations on B. biporcatus carried out over a year in Borneo supplement our knowledge. Of the 191 B. biporcatus collected in Borneo, 47 individuals (25~) were caught more than 10 m from a stream, while all the others were collected along streams in breeding aggregations. On one Bornean stream, Sungai Ensurai, 41 B. biporcatus were seen on 15 occasions beside or in small pools similar to those mentioned above in connection with B. parvus. These pools were concentrated in a strip 15 m long. Males were calling, and pairs in amplexus and strands of eggs were seen. Marked individuals were recaptured at this site at intervals of from 1 to 4 months. No B. biporcatus was seen at any other point along the 650-m study area of this stream on any of the 36 nights of observation spread over a full year.
Bufo melanostictus is a moderate-sized toad; adults of both sexes may exceed 80 mm. Its geographic range extends from India and southern China to Bali and Borneo. It occurs in large numbers in open, disturbed, and culti- vated areas in cities and towns; it is not found in forests. We have no home range data for B. melanostictus, but in damp weather they were conspicuous in Kuala Lumpur and could be seen moving considerable distances (5-10 m

128 Inger, Voris, and Voris
at a time) along and between drainage ditches. Bufo melanostictus breeds the year round in ditches and other standing water in various parts of Southeast Asia and the East Indies (Church, 1960; Smith, 1917). Though high densities form at breeding sites (Smith, 1917), this form is so ubiquitous within any given locale in Southeast Asia that one cannot speak of breeding aggregations in the sense of semi-isolated, widely scattered clumps of individuals.
Rana bIythi is a large frog (adults 85-170 + mm) that occurs in the forests of the Malaya peninsula as well as Sumatra and Borneo. It is a strictly riparian species living on the banks of medium-sized streams and very large rivers with sluggish to swift currents. Half-grown frogs are sometimes found in smaller tributaries (about 1 m wide) a short distance from the mouth. This species rarely moves far from a stream bed, and of 1330 collected in Borneo, only eight individuals were found more than 10 m from a stream bed. In Malaya, R. blythi specimens were caught an average of 2.97 m (n = 93, s = 3.107) from stream beds, and only two individuals were captured more than 10 m from a stream bed. Individuals of R. blythi, though not randomly distributed, do not form clumps or aggregations along stream banks, and the 953 observations made on three Bornean streams were scattered along the 650-m study area of each stream. Median home ranges for adults on these streams varied from 16 to 19 m (n = 175). Eggs are laid in quiet reaches of streams, and tadpoles usually live in pools cut off from the main current.
Rana hosei is a moderate-sized frog (males 50-60 ram, females 85-100 mm) distributed in the Malaya peninsula, Sumatra, Borneo, and Java. Like R. blythi, it is almost exclusively riparian and confined to forests. Rana hosei is found on the banks and low vegetation along medium-sized to large streams (width 10-30+ m). Unlike R. btythi, R. hosei occurs only on moderately to very swift streams. Individuals are usually found within a few meters of stream beds, and less than 2% of the 640 R. hosei specimens collected in Borneo were more than 8 m from a stream. Along the Sungai Gombak in Malaya, none of the individuals for which we have data was caught more than 3 m from a stream bed (n --- 15, J( = 1.17 m, s = 0.8997). This species does not aggregate on small streams (width <20 m), but does form calling aggregations of males on banks of large, swift rivers (Inger, 1969, and un- published observations). Home range size of R. hosei is unknown,
Rana erythraea is a medium-sized frog (adult males 30-45 ram; females 50-75 ram) occurring from eastern India and Burma to Java, Borneo, Sumatra, and the Philippine Islands. It is similar to B. melanostictus in that it is usually found near human habitation in cultivated or disturbed areas and develops large populations even in towns (Alcala, 1955; Inger and Greenberg, 1963). The borders of lakes and ponds, open swamps, paddy fields, weed- choked ditches, and sluggish creeks are typical habitats. Rana erythraea never moves far from standing or flowing water (Alcala, 1955; Boulenger,

Protein Variation in Asian Frogs 129
1912), and all individuals collected in Malaya were caught within 15 m of a pond or stream. Despite its riparian habits, the association of R. erythraea with ponds, rice fields, and ditches means that its range is not linear, as is the case for the other riparian species studied. Rana erythraea breeds the year round in Southeast Asia, Borneo, and the Philippine Islands. Though breed- ing populations are large, R. erythraea does not form specialized breeding aggregations in the sense of semi-isolated clusters of adults that have moved from normal feeding areas to breeding sites.
Rana laticeps is a small rainforest frog (adults 32-40 mm) ranging from Assam to Borneo. At Bukit Lanjan, Malaya, 33 R. laticeps were captured along a small sandy intermittent stream (width < 1 m). Most individuals were found in the stream, and no individual was captured more than 1 m from the stream (J~ distance = 0.07 m, s = 0.192). Of the 39 individuals of this species collected in Borneo, all but five were caught in or beside small (about 1 m) forest streams. The breeding habits of R. latieeps are unknown, but in view of its apparently restricted horizontal distribution, it almost certainly breeds in pools of very small streams.
Electrophoresis Results
For the Bufo populations, the frequency distribution of morphs for each zone is given in Table I. The assays have been clustered into the two enzymatic groups originally designated by Gillespie and Kojima (1968) and the non- enzymatic proteins (Selander and Yang, 1969, p. 665). These groups are represented by five, ten, and eight zones, respectively. As with most other organisms examined previously (e.g., the house mouse; see Hunter and Strachan, 1961), the esterase zones exhibit extensive variation, which in our assays may be in part a function of the presence of the products of several loci within a single zone.
These data have been summarized (Table II) in terms of the number of morphs per zone and the effective number of morphs per zone, rn~. The latter is the reciprocal of the probability of two frogs taken from the same population having the same morph at a particular zone. It is conceptually similar to the effective number of alleles (ne) of Kimura and Crow (1964). Because the effective number of morphs takes into account both the number of morphs and their relative frequencies, it is helpful in comparing popula- tions represented by varying sample sizes.
Within each species, populations are roughly similar in number of morphs and effective number of morphs at zones. Summary data such as the mean number of morphs per zone and the proportion of zones poly- morphic (Table II) further document the similarity of populations within species. The application of the Wilcoxon matched pairs signed-ranks test
C

130 Ingex, Voris, and Voris
C
0
<
t"~ ¢'-,I w-~ . -~ ¢ .3 ¢ ~ t'e3 t ~
¢ '4
~=~

Protein Variation in Asian Frogs 131
0 o
O
~<
O
Q
~D
O
e~
. , ~
~.~ N N
• ~ ~ ~

132 Inger, Voris, and Voris
0
g N
<
m N
u e~
.¢0
' , . 0
0 N
OQ ,--~

Protein Variation in Asian Frogs ]33
. . o ~ ~
~ g g g . o o o
0
~O( 'q
g § g $ ~ o
,.0 ,.0 ,.0
0 0 0 0 0 0 ~
~0 ( '~
0
0
0 0
0
0 0
0
0
0
. : = ~ = >
0
~ 0
N O 0
z ~ , ~
, d
2
0
,.Q
o
i
o ~
. ~ =

134 Inger, Voris, and Voris
%
¢J
0
cD
~3 o ~
~ v
<
c~ ~ ~ ~ , - ~

Protein Variation in Asian Frogs 135
cq
~--~ ~--~ I ~ . ~ c r~ c a ce~ ¢ ' ~ t"-I O 0 ~ ' ~
• =~ ;=
g .
o ~
~3
r ~
O

136 Inger, Voris, and Voris
C~ 0
0
<
Z
0~
. o
<

Protein Variation in Asian Frogs 137
©
°

138 lnger, Voris, and Voris
(Siegel, 1956, p. 75) to the effective number of morphs at zones of each of these population pairs confirms the hypothesis of no difference between populations within species. The same test applied to the number of morphs at zones gives similar results (no difference between populations within species), with the exception of the B. rnelanostictus comparison. Here the disparate sample sizes (30 vs. 10) clearly are responsible for the rejection of the null hypotheses at the 5% level of significance. In particular, the excep- tionally small sample of ten B. melanostictus from JIPC has been struck from further comparisons involving either number of morphs or effective number of morphs due to the low "ceiling" this small sample imposes on these measures.
A comparison of populations between species yields different results. In terms of mean number of morphs per zone and proportion of zones, polymorphic B. asper populations are the least variable, B. melanostictus the most variable, and B. parvus intermediate. It is also evident that B. parvus is more similar to B. asper than it is to B. melanostictus in these measures of variation.
All possible interspecific pairwise comparisons of populations (excluding comparisons involving the exceptionally small sample of B. melanostictus from JIPC) have been made using the Wilcoxon test on the number of morphs and the effective number of morphs (me) at zones. In terms of me, the B. rnelanostictus population at DIRS is significantly more variable at the 5~ level than both the B. asper and the B. parvus populations. Differences between B. asper and B. parvus in me are not significant (P > 0.05). The appli- cation of the Wilcoxon test to the number of morphs at zones gave slightly different results. Bufo melanostictus at DIRS is different from both B. asper populations at the 5% level but different from only one of the B. parvus populations, the one from Bukit Lanjan.
The morph frequency data from the Rana populations are given in Table III. The number of morphs per zone and effective number of morphs for each zone are summarized in Table IV. Neither R. blythi nor R. erythraea shows intraspecific, interpopulation differences in amounts of variation measured by m e and the number of morphs. The results of the Wilcoxon test using both number of morphs and me values at zones indicate no signifi- cant difference between populations within species at the 5~ level.
Rana species are not as different from one another as are those of the Bufo studied. In terms of mean number of morphs per zone, the populations fall into the following order (highest to lowest): R. hosei, R. erythraea (Bukit Lanjan and University of Malaya), R. blythi (Ulu Gombak and Templer Park), and R. laticeps (Table IV). Unfortunately, the values for R. laticeps are not reliable because of lack of data for several critical zones. None of the populations is significantly different from the others at the 5% level, using

Protein Variation in Asian Frogs 139
the Wilcoxon test on the number of morphs or effective number of morphs at zones.
The summary data for Bufo all pointed to the same ordering of the populations in terms of variability. This is not true for the data from species of Rana. Rana erythraea is more variable than R. blythi in terms of the average number of morphs per zone, but the reverse is true with respect to the pro- portion of zones polymorphic per population. This result stems from the fact that R. erythraea has very high variation in the esterases but modest variation among the critical enzymes and nonenzymatic proteins, while R. blythi, although not as variable in the esterases, does show some variation at almost all zones.
DISCUSSION
Wright (see his summary paper: Wright, 1951), in analyzing the relationship between population characteristics and genetic structure, emphasized the significance of population size and shape of range in affecting local differen- tiation and variation. Although information on the populations of amphibians we studied is far from complete, known differences in the distributional patterns of these species can be related to Wright's hypothesis and form the basis of predictions testable by electrophoretic data.
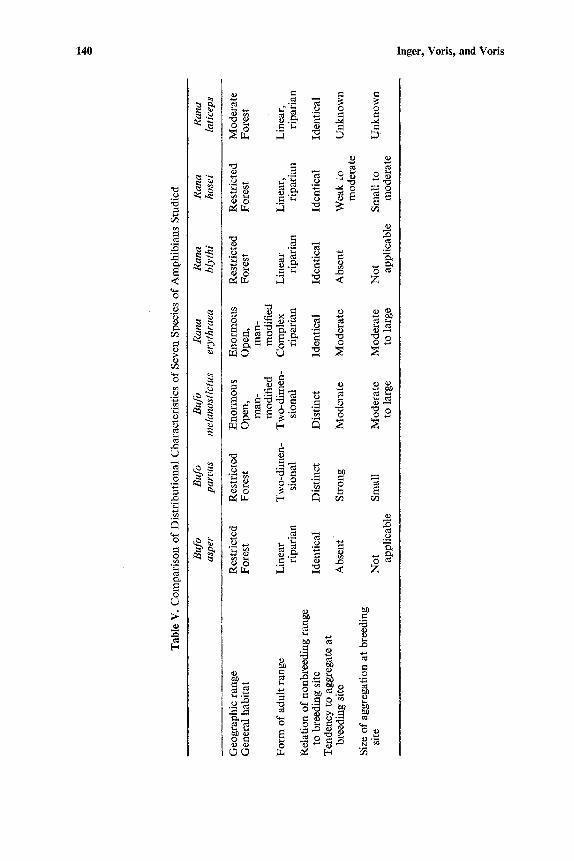
The distributional features of the seven species we studied are presented in the Results section and summarized in Table V. The characteristics of these populations that are most likely to affect the rate and amount of local differen- tiation and the extent of local heterogeneity are the form of the postmetamor- phic range, the relationship of the nonbreeding range to the breeding site, and the tendency to form breeding aggregations.
Linear, riparian ranges increase the likelihood of matings by near neighbors and thus increase the amount of inbreeding. Two-dimensional ranges, on the other hand, at least offer the possibility ofmatings byindividuals from relatively distant parts of the local range. Effective population sizes for two species differing in this respect are related as X~/2:X, according to Wright (1951). Though some fidelity to particular breeding sites shown by frogs and toads having two-dimensional ranges may reduce the difference in effective population sizes, species with linear ranges should exhibit more inbreeding and therefore less heterogeneity within and more differentiation between local populations. If breeding sites are essentially identical to feeding and nonbreeding areas, it appears likely that members of a breeding pair could have been near neighbors, which would contribute to inbreeding and reduction of gene flow. If breeding and nonbreeding areas are not identical, the chances for significant movement of a juvenile frog in the period between its metamorphosis and reproductive activity are enhanced, increasing effective
140 Inger, Voris, and Voris
"t:J
<
r ~
r-,
{3
, . 0
e.,
0
~.~
g
&
M
o . . o
o . o
~ o
&
0
-~o ~E
e~
Z
~D
Z
._=
0
c~

Protein Variation in Asian Frogs 141
population size, reducing the rate of local differentiation, and increasing the amount of heterogeneity within local populations.
Formation of breeding aggregations, by bringing a number of individuals together from a relatively wide area, tends to increase effective population size, reduce the rate of inbreeding and local differentiation, and increase the amount of heterogeneity locally. The degree to which these effects are felt is probably directly related to the size of the aggregation.
The general character of the habitat has an effect on gene flow among populations. Species living in open, man-modified environments have irregu- larly shaped but more or less continuous areas available in the surroundings of Kuala Lumpur and over large sections of Malaya. Forest environments, on the other hand, are becoming increasingly constricted into islands, thus isolating populations of forest species. All other things being equal, gene flow among populations should be greater and local differentiation less for those species living in open, man-modified habitats than for species confined to forests.
The extent of the geographic range of a species, the remaining distri- butional feature of Table V, is likely to influence the amount of regional, as distinguished from local, differentiation. It is also likely to be related to the degree of ecological or physiological adaptability, which may result from various patterns of genetic structure.
Prior to discussing these predictions in terms of the specific populations we studied, it is appropriate to review certain limitations of the data that will necessarily restrict the scope of the discussion from this point forward. For example, the fact that all samples were taken from one geographic area precludes any discussion of the role regional differences may play in the genetics of our frog populations. Second, the quality of the electrophoretic data did not allow identification of morphs between populations, as is neces- sary to obtain estimates of local differentiation. Thus we can compare populations within and between species in terms of relative amounts of genetic variation, but not in terms of the extent of local differentiation.
If the seven species studied are considered in the light of the last five distributional characteristics (Table V) and their potential effects on genetic variation, we would expect B. asper to show more homogeneity within local populations than B. parvus or B. melanostietus. The restriction of B. asper to forests, which now exist only in isolated patches around Kuala Lumpur, certainly limits gene flow among populations. Its linear range, coincidence of breeding and feeding areas, and lack of breeding aggregations should result in high rates of inbreeding and local homogeneity. It differs from B. melano- stictus in all of these distributional features and from B. parvus in all but the restriction to forest. The electrophoretic data substantiate this prediction. Both total and mean number of morphs per zone and in terms of proportion

142 Inger, Voris, and Voris
of polymorphic zones, B. asper shows the least amount of variation. In terms of effective number of morphs (me), B. asper populations are significantly less variable than the B. melanostictus population from DIRS.
We would not expect B. parvus and B. melanostictus to differ greatly in the amount of genetic variation since they share some important charac- teristics: both have a two-dimensional adult range, and both have breeding sites distinct from their nonbreeding ranges (Table V). However, B. parvus has a stronger tendency to form semi-isolated breeding aggregations, which theoretically should increase the effective population size and decrease the inbreeding effect. This trend would be offset by the smaller numbers of B. parvus at breeding sites. Also, the difference between B. parvus and B. melanostictus in the general character of the habitat should result in more homogeneity of local populations in B. parvus. Here the electrophoretic results do not fit as closely to what we predicted. Although the direction of difference is as anticipated, B. parvus with less variation, the magnitude of difference is greater than predicted. In fact, B. parvus populations are closer to B. asper populations in amount of variation (e.g., as measured by mean number of morphs per zone) than to the DIRS B. melanostictus population. This is demonstrated by the fact that both populations of B. parvus were found to be statistically less variable than the DIRS B. melanostictus in terms of effective number of morphs, while neither B. parvus population was found to be significantly different from either of the B. asper populations. Here the lack of more detailed measures of the population structure of B. parvus is likely the reason for the "discrepancies" between prediction and findings.
The species of Rana studied are uniform in the coincidence of breeding and feeding sites. Though all are riparian, the complex range of R. erythraea, its large numbers, and its less restricted habitat should result in its showing more genetic variation within local populations than R. blythi and probably the others, although less is known of their ecology. Rana erythraea shows a moderate tendency to form breeding aggregations, which would further increase genetic variability; R. hosei would be expected to be less variable on this basis since it has a weak tendency to aggregate and forms smaller aggregations; and R. blythi, which does not form breeding aggregations at all, would be least variable. It is not known how this factor affects R. laticeps. In general, more information is needed on the ecology of both R. hosei and R. laticeps in order to evaluate the effect of ecological factors on their genetic variation.
Although the electrophoretic results show that both R. erythraea populations are more variable than either of the R. blythi populations in mean number of morphs per zone, the differences are not significant (P> 0.05). In terms of another measure, proportion of zones polymorphic, R. blythi is more variable than R. erythraea. Surprisingly, the R. hosei population is

Protein Variation in Asian Frogs 143
the most variable Rana population in mean number of morphs. The high variation in the R. hosei population is not expected, and the reason for it is not known.
These data point up two important general conclusions. First, it now seems evident that where our knowledge of the ecology and population structure of the frog species is greatest (e.g., B. asper, B. melanostictus, R.
blythi, and R. erythraea), we may have enough information to predict a fairly realistic outline of the pattern of genetic variation. In species about which we know less, our predictions become less useful, partly because some factors remain unknown in a qualitative sense and partly because our meas- ures of the relative intensity of various factors may be too crude to allow an estimate of net effect. For example, the high variation in the R. hosei popu- lation was not anticipated, nor can it be explained in the context of our current understanding of the ecology and population structure of this species.
Second, the "prediction" approach we have taken to explore the re- lationship between genetic variation and the ecology of species has an import- ant inherent difficulty. We constantly run the risk of making the correct prediction for the wrong reason(s). Of course, the fewer concrete data we have on the critical factors such as neighborhood size and rates of gene flow, the greater the risk. It is evident that the time is rapidly approaching when a major effort is going to have to be made toward obtaining complete and reliable estimates of these critical factors for some populations to provide a base on which to build studies of this type.
ACKNOWLEDGMENTS
Laboratory space for this study was generously supplied by the Medical Ecology Division of the Institute for Medical Research in Kuala Lumpur. Special personal thanks are owed to Dr. Illar Muul of the U.S. Army Medical Research team stationed at the Institute and to Lira Boo Liat, officer-in- charge of the Division, and their respective staffs for help in solving the logisti- cal problems that inevitably come up when doing research in a foreign country. We also thank Diane Goodhart and Thangavelu S/O Subramaniam for assistance in the laboratory. We are grateful to R. C. Lewontin for valuable comments on an early manuscript.
R E F E R E N C E S
Alcala, A. C. (1955). Observations on the life history and ecology of Rana erythraea Schlegel, on Negros Island, Philippines. Silliman J. 2:175.
Boulenger, G. A. (1912). Reptilia and Batraehia. . . in a Vertebrate Fauna of the Malaya Peninsula, Taylor & Francis, London.

144 Inger, Voris, and Voris
Church, G. (1960). Annual and lunar periodicity in the sexual cycle of the Javanese toad, Bufo melanostictus Schneider. Zoologica (Sci. Cont. N. Y. Zool. Soc. Pt. 4) 44:181.
Dessauer, H. C. (1966). Multiple localizations of specific proteins on starch gel electrophero- grams. Rutgers Univ. Serol. Mus. Bull. 36:1.
Dessauer, H. C., and Nevo E. (1969). Geographic variation of blood and liver proteins in cricket frogs. Biochem. Genet. 3:171.
Gillespie, J. H., and Kojima, K. (1968). The degree of polymorphisms in enzymes involved in energy production compared to that in nonspecific enzymes in two Drosophila ananassae populations. Proc. Natl. Aead. Sci. 61:582.
Hubby, J. L., and Lewontin, R. C. (1966). A molecular approach to the study of genic heterozygosity in natural populations. L The number of alleles at different loci in Drosophila pseudoobscura. Genetics 54:577.
Hubby, J. L., and Throckmorton, L. H. (1965). Protein differences in Drosophila. II. Comparative species genetics and evolutionary problems. Genetics 52:203.
Hunter, R. L., and Strachan, D. S. (1961). Esterases of mouse blood. Ann. N. Y. Aead. Sei. 94(1243) :861.
Inger, R. F. (1966). The systematics and zoogeography of the Amphibia of Borneo. Fieldiana Zool. 52:1.
Inger, R. F. (1969). Organization of communities of frogs along small rain forest streams in Sarawak. J. Anita. Ecol. 38:123.
Inger, R. F., and Greenberg, B. (1963). The annual reproductive pattern of the frog Rana erythraea in Sarawak. Physiol. Zool. 36(1):21.
Johnson, W. E., and Selanders, R. K. (1971). Protein variation and systematies in kangaroo rats (genus Dipodomys). Syst. Zool. 20:377.
Kimura, M., and Crow, J. F. (1964). The number of alleles that can be maintained in a finite population. Genetics 49:725.
Koehn, R., and Mitton, J. (1972). Population genetics of marine pelecypods. I. Ecological heterogeneity and evolutionary strategy at an enzyme locus. Am. Naturalist 106(947): 47.
Lakovaara, S., Saura, A., and Falk, C. T. (1972). Genetic distance and evolutionary relationships in the Drosophila obscura group. Evolution 26(2):177.
Lewontin, R. C., and Hubby, J. L. (1966) A molecular approach to the study of genie heterozygosity in natural populations. II. Amount of variation and degree of heterozy- gosity in natural populations of Drosophila pseudoobscura. Genetics 54:595.
Menhinick, E. F. (1964). A comparison of some species diversity indices applied to samples of field insects. Ecology 45(4) :859.
Nair, P. S., Brncic, D., and Kojima, K. (1971). II. Isozyme variations and evolutionary relationships in the mesophragmatica species group of Drosophila. Stud. Genet. (Univ. Texas Pabl.) 6(7103):17.
Petras, M. L. (1963). Genetic control of a serum esterase component in Mus musculus. Proc. Natl. Acad. Sci. 50:112.
Petras, M. L. (1967). Studies of natural populations of Mus. I. Biochemical polymorphisms and their bearing on breeding structure. Evolution 21:259.
Prakash, S., Lewontin, R. C., and Hubby, J, L. (1969). A molecular approach to the study of genie heterozygosity in natural populations. IV. Patterns of genie variation in central, marginal and isolated populations of Drosophila pseudoobscura. Genetics 61(4): 841.
Ruddle, F. H. (1966). Kidney esterases of the mouse (Mus muscalus): Electrophoretic analysis of inbred lines C57B/6J, RF/J and SJL/J. J. Histochem. Cytochem. 14(1):25.
Selander, R. K., and Yang, S. Y. (1969). Protein polymorphism and genie heterozygosity in a wild population of the house mouse (Mus musculus). Genetics 63:653.
Selander, R. K., Yang, S. Y., Lewontin, R. C., and Johnson, W. E. (1970). Genetic variation in the horseshoe crab (Limuluspolyphemus), a phylogenetic "relic." Evolution 24(2) :402.
Shaw, C. R., and Koen, A. L. (1968). Starch gel zone electrophoresis of enzymes. In Smith, I. (ed.), Chromatographic and Electrophoretic Techniques, Vol. II, Interscience, New York.

Protein Variation in Asian Frogs 145
Shaw, C. R., and Prasad, R. (1970). Starch gel electrophoresis of enzymes--A compilation of recipes. Biochem. Genet. 4:297.
Siegel, S. (1956). Nonparametric Statistics for the Behavioral Sciences, McGraw-Hill, New York.
Smith, M. A. (1917). A list of the batrachians at present known to inhabitat Siam. J. Nat. Hist. Soc. Siam 2:226.
Wright, S. (1951). The genetical structure of populations. Ann. Eugen. 15:323.
D
Copyright © 2022 FDOKUMEN




















