
Comparative cytogenetics of eight species of Cycloramphus (Anura, Cycloramphidae
Upload
independentCategory
view
1download
0
BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofitpublishers, academic institutions, research libraries, and research funders in the common goal of maximizing access tocritical research.
A NEW SPECIES OF BUFO (ANURA: BUFONIDAE) FROMANDEAN PERUAuthor(s): Edgar Lehr, Jennifer B. Pramuk, Mikael LundbergSource: Herpetologica, 61(3):308-318. 2005.Published By: The Herpetologists' LeagueDOI: http://dx.doi.org/10.1655/04-90.1URL: http://www.bioone.org/doi/full/10.1655/04-90.1
BioOne (www.bioone.org) is a nonprofit, online aggregation of core research in thebiological, ecological, and environmental sciences. BioOne provides a sustainable onlineplatform for over 170 journals and books published by nonprofit societies, associations,museums, institutions, and presses.
Your use of this PDF, the BioOne Web site, and all posted and associated contentindicates your acceptance of BioOne’s Terms of Use, available at www.bioone.org/page/terms_of_use.
Usage of BioOne content is strictly limited to personal, educational, and non-commercialuse. Commercial inquiries or rights and permissions requests should be directed to theindividual publisher as copyright holder.

Herpetologica, 61(3), 2005, 308–318� 2005 by The Herpetologists’ League, Inc.
A NEW SPECIES OF BUFO (ANURA: BUFONIDAE)FROM ANDEAN PERU
EDGAR LEHR1,2,3, JENNIFER B. PRAMUK
4, AND MIKAEL LUNDBERG1
1Staatliche Naturhistorische Sammlungen Dresden, Museum fur Tierkunde,Konigsbrucker Landstrasse 159, D-01109 Dresden, Germany
4Department of Integrative Biology, Brigham Young University,401 WIDB Provo, UT 84602, USA.
ABSTRACT: A new species of Bufo tentatively assigned to the Bufo veraguensis group is described fromforests near Paucartambo de Pasco, Peru, at elevations of 2600–3000 m in the Cordillera Oriental of centralPeru. The new species differs from all known bufonids by having numerous large glands with many keratinoustips on the dorsum, large glands on the tibia and forearm, and two large dorsal glands lying between theparotoids. The new species is most similar to Bufo chavin. Both species are compared to each other andB. veraguensis with respect to external characters and skull osteology. The tadpole and call of the new speciesremain unknown.
Key words: Andes; Anura; Bufo chavin; Bufonidae; Bufo multiverrucosus; Bufo veraguensis; New Species;Osteology; Peru
TOADS of the Bufo veraguensis group asdefined by Duellman and Schulte (1992) occuron forested slopes of the Andes in Peru,Bolivia, and northwestern Argentina from1280–3070 m in elevation (Duellman andSchulte, 1992; Lehr et al., 2001). Seven speciesdescribed within the last 15 years have beenassigned to the B. veraguensis group, and cur-rently it is composed of the following 11 taxa:Bufo amboroensis Harvey and Smith, B. arbor-escandens Duellman and Schulte, B. chavinLehr et al., B. fissipes Boulenger, B. incaStejneger (Hoogmoed, 1990: considered a syn-onym of B. veraguensis, but resurrected byDuellman and Schulte, 1992); B. justinianoiHarvey and Smith, B. nesiotes Duellman andToft, B. quechua Gallardo, B. rumbolli Car-rizo, B. veraguensis Schmidt, and B. sp. nov.Lehr, Pramuk and Lundberg, this paper.
Intensive fieldwork during the last fouryears along the Paucartambo de Pasco Valleyin eastern central Peru has revealed severalnew eleutherodactyline anurans (Lehr et al.,2004; Lehr, 2005; Lehr et al., in press) andseveral other species of anurans awaiting de-scription. Among the anurans were specimensof Bufo with distinctive warts on the dorsum
and extremities. Herein, we describe thisnew species.
MATERIALS AND METHODS
The format of this description follows thatof Duellman and Schulte (1992). Field noteswere recorded by M. Lundberg. Specimenscollected in 2000 were preserved in 4% for-malin, those collected after 2000 were pre-served in 96% ethanol, and all are stored in70% ethanol. Specimens were dissected todetermine sex by observation of presence orabsence of ova or testes. Measurements ofadults were made with dial calipers androunded to the nearest 0.1 mm and are asfollow: SVL (snout–vent length); TL (tibialength); FL (foot length, distance from proxi-mal margin of inner metatarsal tubercle to tipof fourth toe); HL (head length, from angle ofjaw to tip of snout); HW (head width, greatestwidth, measured between centers of tympana);ED (eye diameter); TY (tympanum diameter,horizontal); IOD (interorbital distance); EW(eyelid width); IND (internarial distance); E–N (eye–nostril distance, straight line distancebetween anterior corner of orbit and posteriormargin of external naris); PL (parotoid length,horizontal); and PW (parotoid width, greatestwidth). Herein, we use a dash for ranges, anda backslash to indicate separate counts ofopposing sides (left/right) of the same speci-men. Number for glands within dorsolateral
2 PRESENT ADDRESS: Natural History Museum andBiodiversity Research Center, Division of Herpetology,The University of Kansas, Jayhawk Boulevard 1345,Lawrence, KS 66045-7561, USA.
3 CORRESPONDENCE: e-mail, [email protected]
308

row was combined for both sides and for allspecimens and presented as a range. An adultfemale (MTD 44751) was skeletonized follow-ing Simmons (1986). C & S indicates cleared-and-stained. Our terminology for cranial crestsand for webbing formula follows Cei (1980)and Savage and Heyer (1997), respectively.Drawings were made using a stereomicroscopewith drawing tube attachment. Acronyms formuseum collections are those of Leviton et al.(1985) and Frost (2004), with the addition ofMTD (Museum fur Tierkunde Dresden, Ger-many). Comparative data for Bufo chavin weretaken from Lehr et al. (2001), Lehr (2002), andthe specimens examined (Appendix I).
Bufo multiverrucosus sp. nov.
Holotype.—MHNSM 17820 (Fig. 1), anadult female obtained northwest of Auqui-marca (108 449 06.00 S, 758 429 42.60 W) at 2900m on 21 March 2000, Distrito de Paucartambo,Provincia de Pasco, Departamento de Pasco,Peru, by a local collector for M. Lundberg.
Paratypes.—All from Provincia de Pasco,Departamento de Pasco, Peru: nine femaletopotypes (MHNSM 17836 collected on 27April 2000, MHNSM 20653–56, MTD 44747–49, 44751 [skeleton] collected on 11 March2002), one male topotype (MHNSM 20656collected on 11 March 2002); two females(MHNSM 19035–36), one juvenile (MHNSM19037) collected on 7 September 2002 atCillapata (Distrito de Paucartambo, approxi-mately 1.5 km NNE Auquimarca, 108 439 52.00S, 758 429 47.80 W, 2900 m), two juveniles (MTD44752, 45076) collected (respectively) on 1 April2002 and 2 June 2002 at Cillapata; one female(KU 291686) collected on 4 April 2002 atChinche at 3000 m; two juveniles (MHNSM18612–13) collected on 26 June 2002 at km 79along the Ninacaca–Oxapampa road, near theUchun Bridge (Distrito de Huachon, 108 39915.10 S, 758 449 41.20 W, 2600 m); one female(MTD 46378) collected on 18 March 2004 at LaFlorida, approximately 6.6 km east of Quipar-acra, at 2600 m. All specimens were collected bylocal people working with M. Lundberg.
FIG. 1.—Left column: Bufo multiverrucosus (MHNSM 17820, holotype, SVL 60.5 mm) in dorsal and ventral view; rightcolumn: Bufo chavin (MTD 44753, SVL 58.8 mm) in dorsal and ventral view, scale in mm. Photos by F. Hohler.
September 2005] HERPETOLOGICA 309

Diagnosis.—A medium-sized species of Bufoattaining a maximum SVL of 68.9 mm in adultfemales (MTD 44749, Fig. 2). The new speciesis distinguished from all congeners by thefollowing combination of characters: (1) can-thus rostralis distinct, orbitotympanic and post-orbital crests weak; (2) tympanum distinct; (3)parotoid glands moderately large, subtriangu-lar, contacting eye; two large glands on dorsumof head between parotoids; (4) numerous large,keratin-tipped glands on dorsal surfaces ofbody; one dorsolateral row on each side of bodyconsisting of large, nearly round elevated tu-bercles, nearly forming a fold; (5) forearm andtibia usually with a large, elevated, elongategland; outer dorsal margin of foot with a row oflarge, elevated, elongate glands; outer dorsalmargin of hand with one small, elevated, tri-angular gland; (6) ventrolateral granules en-larged and elevated; (7) tarsal fold absent; (8)webbing on toes fleshy; (9) first finger shorterthan second; (10) males without vocal sacs orvocal slits; Fingers I and II in breeding malesbearing small keratinous spines.
The presence of large glands on the dorsumof the head between the parotoids, largeglands on tibia and forearm and numerouskeratin-tipped glands on dorsal surfaces of thebody immediately distinguishes Bufo multi-verrucosus from all the members of the genus.Bufo multiverrucosus differs from B. vera-guensis by having a dorsolateral row of en-larged tubercles, by having visible tympana,and by having Finger I shorter than Finger II.Bufo multiverrucosus is most similar to thecentral Peruvian species B. chavin (Fig. 1).Bufo multiverrucosus differs from B. chavin asfollows (characteristics of B. chavin in paren-theses): dorsum with many, large, elevated,conical glands, all with keratinous tips, skinspiny (few large, slightly elevated, ovoidglands, glands with keratinous tips restrictedto head and neck, skin smooth); two largeglands situated dorsally on head, contactingeach other and the parotoids, nearly forminga row (no large glands between parotoids);glands on tibia and forearm with keratinoustips, difficult to distinguish from surroundingglands (glands on tibia and forearm entire andclearly discernable from surrounding glands);arms with many glands situated laterally andventrolaterally, distinct row of glands notdiscernible (outer surface of arm with a ven-
trolateral row of glands sometimes forming anentire gland, clearly distinguishable); uppereyelid with large tubercles (upper eyelidusually without tubercles); dorsolateral rowof elevated glands round to ovoid in dorsalview, subovoid in profile, 5–11 on each side(�x 5 8 6 1, n 5 14), bearing keratinous tips,narrowly separated from parotoids or contact-ing parotoids, narrowly separated from eachother, nearly forming a dorsolateral fold,extending to inguinal region (elongated, ele-vated glands, 3–8 on each side [�x 5 5 6 1, n 57] without keratinous tips, beginning aboveinsertion of forearm and extending to inguinalregion); flanks with many small tubercles (withfew scattered tubercles); parotoid contactingeye (narrowly separated from eye); verticaldistance between lower margin of eye and up-per lip as large as vertical diameter of tympa-num (less than vertical diameter of tympanum);anterior end of parotoid broad, contactingupper eyelid (narrow, narrowly separated fromupper eyelid); parotoid glands subtriangular indorsal and dorsolateral views (ovoid in dorsalview, subtriangular in dorsolateral view); skinon throat and chest strongly granular (weaklygranular); venter strongly granular (weakly
FIG. 2.—Dorsolateral and ventral view of female Bufomultiverrucosus (MTD 44749, paratype, SVL 68.9 mm) inlife. Photos by M. Lundberg.
310 HERPETOLOGICA [Vol. 61, No. 3

areolate); fingers and toes with broad lateralfringes (narrow lateral fringes); webbing be-tween fingers clearly discernable (fleshy, vesti-gial); narrow middorsal stripe absent (palecream to yellow middorsal stripe usuallypresent); venter predominately pale brownwith dark brown blotches forming longitudinalstripes (venter predominantly dark brown withpale brown blotches).
Description of the holotype.—Body robust;head broader than long; snout truncate indorsal profile, rounded in lateral profile; can-thus rostralis distinct, covered with smallkeratin-tipped glands, conical in profile;orbitotympanic and postorbital crests weak;dorsum of head flat, skin co-ossified withunderlying cranial bones; interorbital distancegreater than eyelid width; internarial areaconcave; nostrils slightly protuberant, directedlaterally; loreal region concave; lips rounded;small V-shaped notch at symphysis of upperjaw; oral ricti positioned below posteriormargin of tympanum; tympanum diametersmaller than eye diameter; tympanic annulusindistinct, dorsal and posteroventral marginspartly covered by parotoids, anterolateralmargin covered by postorbital crest; skin ondorsum with numerous large, round, elevatedglands; dorsal glands ovoid in profile, each
bearing many keratinous tips; dorsal skinbetween large glands covered with smallglands, conical in profile, each bearing a singlekeratinous tip; parotoid glands ovoid in dorsalview, subtriangular in dorsolateral view, nearlyas long as wide, widest posteriorly, descendingonto side of head with anterior edge contactingeye; numerous large, elevated, elongate glands(greatest length 6.1 mm) irregularly distribut-ed on dorsum, each bearing several keratinoustips; glands larger and more densely arrangedmiddorsally between neck and cloacal open-ing, decreasing in size and density laterally;two large, elongate glands (greatest length7.4 mm/6.8 mm) dorsally on head betweenparotoids, contacting each other and theparotoids; upper eyelid with conical keratin-tipped tubercles; dorsolateral row of nearlyround, elevated glands, subovoid in profile (11/8, greatest length 3.9 mm), narrowly separatedfrom each other, nearly forming dorsolateralfold, contacting tympanum anteriorly, extend-ing to inguinal region and increasing in sizeposteriorly; ventrolateral row of five small ele-vated, round glands present dextrally (sinistralrow not discernible from surrounding tuber-cles) beginning posterior to insertion of fore-limb, extending to inguinal region; numeroussmall tubercles on flanks; skin of throat, chest,and venter granular; granules on venter largerthan those on chest and throat; arms relativelyshort and robust; hands broad with relativelyshort fingers; relative length of adpressedfingers 1 , 2 , 4 , 3; webbing fleshy; web-bing formula of fingers I2þ–2II2–3�III3–3IV;lateral fringes broad; tips of digits terminatingin indistinct discs; subarticular tubercles roundin ventral aspect and in profile; supernumer-ary tubercles 60–70% as large as subarticulartubercles; palmar tubercles large and round,flat in profile, about three times size of thenartubercle; thenar tubercle ovoid, round inprofile (Fig. 3); limbs moderately short androbust; tibia length shorter than foot length;tarsal fold absent; inner metatarsal tubercleovoid, conical in profile, two times the size ofouter metatarsal tubercle; outer metatarsaltubercle ovoid, flat in profile; toes moderatelyshort; relative length of adpressed toes 1 , 2 ,3 , 5 , 4; fleshy webbing; webbing formula oftoes I1þ–2�II1þ–3�III1þ–3�IV3þ–2þV; broadlateral fringes present; subarticular tuber-cles small and round, conical in profile;
FIG. 3.—Ventral view of right foot and hand of holotypeof Bufo multiverrucosus (MHNSM 20028). Drawings byJ. Pramuk.
September 2005] HERPETOLOGICA 311

supernumerary tubercles 80–90% as large assubarticular tubercles (Fig. 3); dorsal surfaceof each forearm with large, elevated, elongategland; gland on left forearm (length 10.6 mm)nearly entire, gland on right forearm dividedinto row of glands (length 10.4 mm); outerlateral surfaces of forearms with many round-to-ovoid tubercles, conical in profile; outerdorsal margin of hand with one small, elevatedgland; dorsal surface of tibia with one large,elevated, elongate gland (length 13.5 mm/12.7mm); ventral surface and ventrolateral outersurface of foot with many round to ovoidglands, conical in profile; outer dorsal marginof foot with large, elevated, elongate glandsforming row and one distinct, elevated, tri-angular gland on base of fifth toe (length 6.1mm/5.0 mm); cloacal opening protuberant,directed ventrally near upper level of thighs;choanae small, ovoid; maxillary, premaxillary,mandibular, and vomerine teeth absent;tongue elongate and ovoid, nearly twice aslong as wide, about equal in width throughoutits length, free posteriorly for about one thirdits length.
Measurements (in mm) of holotype: SVL:60.5; TL: 18.9; FL: 20.8; HL: 19.0; HW: 21.5;ED: 5.1; TY: 3.1; IOD: 6.4; EW: 4.7; IND: 4.4;E–N: 4.3; PL: 9.0; PW: 9.4.
Comparative cranial osteology.—The skullof Bufo multiverrucosus is broadly roundedanteriorly and is wider than long with thegreatest width being at the level of thequadratojugals (Fig. 4A–C). Of 30 species ofBufo examined (Appendix 1), the skull ofthe new species is most similar in structureto the crania of B. chavin (Fig. 5A–C) andB. veraguensis (Fig. 5D–F). Relative to otherBufo, species of the B. veraguensis group havelightly (B. chavin) to moderately ossified(B. veraguensis, and B. multiverrucosus) skulls;the latter two species have slight dermal orna-mentation on the dorsal surface of the nasalsand frontoparietals, with the ornamentation ofB. multiverrucosus being the most developed.All three species also have the dorsal surface ofthe sphenethmoid exposed, a consequence oftheir relatively light cranial ossification com-pared to other Bufo.
In dorsal view, in Bufo multiverrucosus andB. chavin, the premaxillae and anterodorsalmargins of the maxillae are visible throughouttheir entire length (only the lateral-most edges
of the premaxillae are visible in B. veraguensis;Fig. 5D). In addition, the posterior margins ofthe nasals of B. multiverrucosus and B. chavinextend posteriorly to a slightly greater extentthan do those of B. veraguensis and, as such,compose slightly greater proportions of themargins of the orbits. In dorsal view, the orbitsof B. multiverrucosus and B. chavin areobliquely ovoid; whereas, in B. veraguensis,they are relatively more round.
In ventral view, the premaxillae and maxillaeof Bufo chavin and B. multiverrucosusare deeper and more robust than those ofB. veraguensis. Moreover, the maxillae ofB. chavin and B. multiverrucosus are angledposterolaterally to a greater degree from theirpoint of contact with the lateral margins of thepremaxillae than they are in B. veraguensis,yielding a relatively more smoothly archedappearance to the maxillary arcade of the lat-ter species. In ventral view, B. chavin andB. multiverrucosus have a ridge present on thedorsal surface of the parasphenoid that extendsfrom the point of contact of the medial ramus ofthe pterygoid and parasphenoid to the medialpoint of the parasphenoid. A ridge on theparasphenoid also is present in B. veraguensis;however, it is only present medially and doesnot extend to the point where the pterygoidcontacts the parasphenoid. In ventral view, theanterior edge of the cultriform process of theparasphenoid is more acuminate in B. vera-guensis than it is in B. multiverrucosus and B.chavin. In addition, the vomers are more robustin B. veraguensis and have distinctive post-choanal and prechoanal processes; whereas, inB. multiverrucosus and B. chavin, the vomersare more slender and lack well-developedpostchoanal processes.
In lateral view, the quadratojugals of Bufochavin, B. multiverrucosus and B. veraguensisare moderately well developed and overlapslightly less than the posterior 1/4 of themaxillae. Bufo multiverrucosus and B. chavinboth share the presence a dorsal expansion ofthe pars facialis of the maxilla that contacts theventrolateral surface of the nasal, whereas theprocess is absent in B. veraguensis. The overallshape of the skull of Bufo multiverrucosus ismost similar to that of B. chavin; both specieshave similarly shaped and flattened frontopa-rietals that broadly contact the medial surfacesof the otic rami of the squamosals. In contrast,
312 HERPETOLOGICA [Vol. 61, No. 3
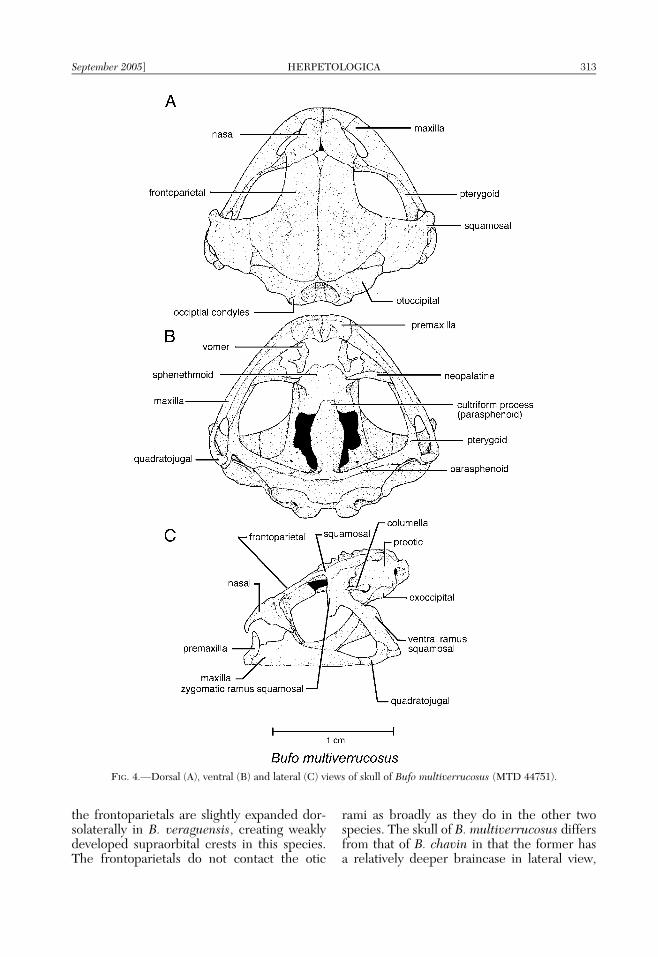
the frontoparietals are slightly expanded dor-solaterally in B. veraguensis, creating weaklydeveloped supraorbital crests in this species.The frontoparietals do not contact the otic
rami as broadly as they do in the other twospecies. The skull of B. multiverrucosus differsfrom that of B. chavin in that the former hasa relatively deeper braincase in lateral view,
FIG. 4.—Dorsal (A), ventral (B) and lateral (C) views of skull of Bufo multiverrucosus (MTD 44751).
September 2005] HERPETOLOGICA 313

greater exostosing of the dorsal surface of theskull, and a less acuminate anterior edge of thecultriform process of the parasphenoid.
Coloration of holotype in preservative.—Dorsum brown with irregular dark brownblotches and stripes; two pale brown, ovoidblotches at middorsum; pale brownish orangestripe extending from dorsal surface of parotoidacross dorsolateral row of tubercles to inguinal
region on both sides; flanks dorsolaterally andventrolaterally with dark brown, irregularstripes beginning behind tympanum and ex-tending with interruptions to inguinal region;remaining areas of flanks dark brown with smallbrownish orange blotches; inguinal region andregion posterior to axilla cream; tympanumdark brown, narrow dark brown stripe extend-ing from posterior corner of eye to junction of
FIG. 5.—Left column: dorsal (A), ventral (B) and lateral (C) view of skull of B. chavin (MTD 43786), right column:dorsal (D), ventral (E), and lateral (F) views of skull of B. veraguensis (MTD 44751).
314 HERPETOLOGICA [Vol. 61, No. 3
jaw on both sides of head; upper lip below eyecream with dark brown bars; throat, chest andventer cream with dark brown blotches; darkbrown blotches on venter forming two longitu-dinal, slightly curved stripes extending fromchest across venter to thighs on both sides; darkbrown blotch on middle of thighs below cloaca;ventral surfaces of hands and feet gray.
Coloration of holotype in life.—As describedabove except that the orange and cream col-oration are more intense. Iris greenish brownwith fine, black reticulations, pupil borderedby olive-yellow ringlet.
Variation.—The type series is colored andpattered as described above. Dorsal groundcoloration in life varies from nearly entirelydark brown (MHNSM 17836) to orange brown(MTD 44747). Ventral ground coloration var-ies from cream, pale brown to brownish orange(MTD 44747) with a variable ventral patternconsisting of small, dark brown blotches(MHNSM 20655, MTD 44747), dark brownblotches forming prominent stripes (MHNSM20653, MTD 44750) or venter and throatnearly entirely dark brown (MHNDM 20654,20654). Variation in the type series is mostconspicuous in number, size and distributionof glands and tubercles on the dorsum.Specimen MHNSM 20653 has the fewestglands (11); MHNSM 17820 has the greatestnumber of glands (19). MHNSM 17836 hasthe most prominent postorbital and preorbitalcrests. There is no obvious sexual dimorphismin skin texture. The single male (MHNSM20656, SVL 47.7 mm) lacks a vocal sac andvocal slits and has Finger I and II slightly
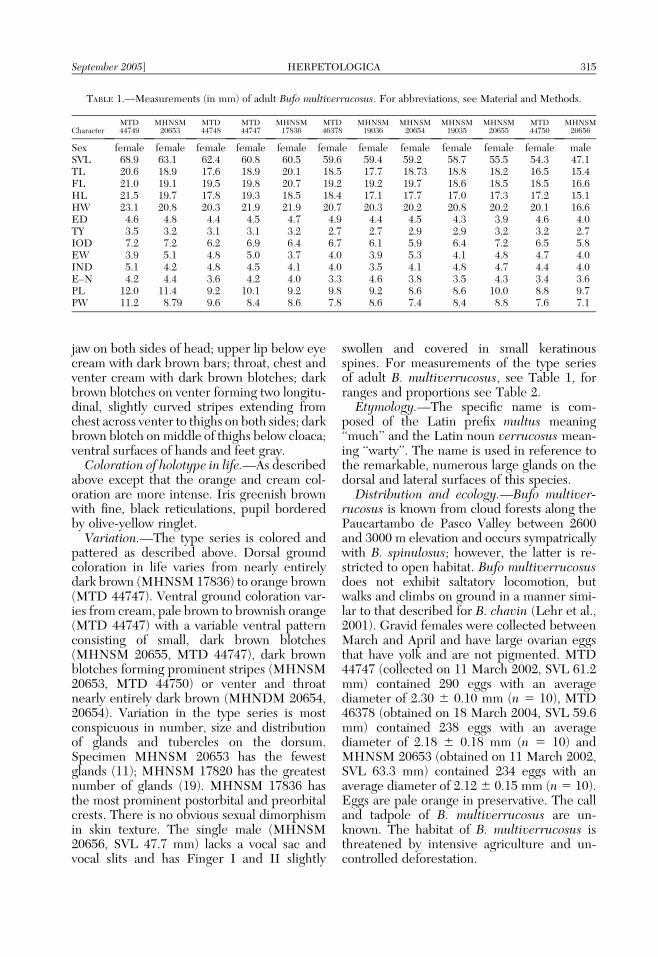
swollen and covered in small keratinousspines. For measurements of the type seriesof adult B. multiverrucosus, see Table 1, forranges and proportions see Table 2.
Etymology.—The specific name is com-posed of the Latin prefix multus meaning‘‘much’’ and the Latin noun verrucosus mean-ing ‘‘warty’’. The name is used in reference tothe remarkable, numerous large glands on thedorsal and lateral surfaces of this species.
Distribution and ecology.—Bufo multiver-rucosus is known from cloud forests along thePaucartambo de Pasco Valley between 2600and 3000 m elevation and occurs sympatricallywith B. spinulosus; however, the latter is re-stricted to open habitat. Bufo multiverrucosusdoes not exhibit saltatory locomotion, butwalks and climbs on ground in a manner simi-lar to that described for B. chavin (Lehr et al.,2001). Gravid females were collected betweenMarch and April and have large ovarian eggsthat have yolk and are not pigmented. MTD44747 (collected on 11 March 2002, SVL 61.2mm) contained 290 eggs with an averagediameter of 2.30 6 0.10 mm (n 5 10), MTD46378 (obtained on 18 March 2004, SVL 59.6mm) contained 238 eggs with an averagediameter of 2.18 6 0.18 mm (n 5 10) andMHNSM 20653 (obtained on 11 March 2002,SVL 63.3 mm) contained 234 eggs with anaverage diameter of 2.12 6 0.15 mm (n 5 10).Eggs are pale orange in preservative. The calland tadpole of B. multiverrucosus are un-known. The habitat of B. multiverrucosus isthreatened by intensive agriculture and un-controlled deforestation.
TABLE 1.—Measurements (in mm) of adult Bufo multiverrucosus. For abbreviations, see Material and Methods.
CharacterMTD44749
MHNSM20653
MTD44748
MTD44747
MHNSM17836
MTD46378
MHNSM19036
MHNSM20654
MHNSM19035
MHNSM20655
MTD44750
MHNSM20656
Sex female female female female female female female female female female female maleSVL 68.9 63.1 62.4 60.8 60.5 59.6 59.4 59.2 58.7 55.5 54.3 47.1TL 20.6 18.9 17.6 18.9 20.1 18.5 17.7 18.73 18.8 18.2 16.5 15.4FL 21.0 19.1 19.5 19.8 20.7 19.2 19.2 19.7 18.6 18.5 18.5 16.6HL 21.5 19.7 17.8 19.3 18.5 18.4 17.1 17.7 17.0 17.3 17.2 15.1HW 23.1 20.8 20.3 21.9 21.9 20.7 20.3 20.2 20.8 20.2 20.1 16.6ED 4.6 4.8 4.4 4.5 4.7 4.9 4.4 4.5 4.3 3.9 4.6 4.0TY 3.5 3.2 3.1 3.1 3.2 2.7 2.7 2.9 2.9 3.2 3.2 2.7IOD 7.2 7.2 6.2 6.9 6.4 6.7 6.1 5.9 6.4 7.2 6.5 5.8EW 3.9 5.1 4.8 5.0 3.7 4.0 3.9 5.3 4.1 4.8 4.7 4.0IND 5.1 4.2 4.8 4.5 4.1 4.0 3.5 4.1 4.8 4.7 4.4 4.0E–N 4.2 4.4 3.6 4.2 4.0 3.3 4.6 3.8 3.5 4.3 3.4 3.6PL 12.0 11.4 9.2 10.1 9.2 9.8 9.2 8.6 8.6 10.0 8.8 9.7PW 11.2 8.79 9.6 8.4 8.6 7.8 8.6 7.4 8.4 8.8 7.6 7.1
September 2005] HERPETOLOGICA 315

REMARKS
Bufo multiverrucosus and B. chavin havelarge, elevated, elongate glands on each fore-arm and tibia. Glands on the extremities arefound in several, unrelated groups of Bufo.Within South America, species with glandsappear to be restricted to high elevations andare only known for the Bufo veraguensis groupand the Bufo spinulosus group (e.g.,B. cophotis and a new Bufo sp. from Bolivia[I. De la Riva, personal communications]),while glands on extremities also are present inthe Bufo alvarius group (e.g., Bufo alvariusGirard) endemic to lower elevations of south-western United States and Mexico and in theBufo bufo group (e.g., Bufo torrenticolaMatsui) from Japan. These glands may secretebufotoxin and likely are used as defenseagainst predation. Bufo chavin and B. multi-verrucosus share similar ecologies: gravidfemales of B. chavin and B. multiverrucosuscontained as many as 286 eggs (Lehr et al.,2001) and 290 eggs, respectively. Males ofboth species lack vocal slits.
Phylogenetic analysis of morphological dataincluding 38 species of Bufo, indicate thatB. chavin and B. multiverrucosus are closelyrelated to B. veraguensis (Pramuk and Lehr, inpress). Another example for two very similar
species from the localities mentioned above isthat of Phrynopus juninensis (Shreve) (Auqui-marca) and P. kauneorum Lehr, Aguilar, andKohler (Palma Pampa). Recent molecular andmorphological comparisons including 15 spe-cies of Phrynopus demonstrate that both aresister species. Both species pairs of Bufo andPhrynopus indicate that both faunal commu-nities may have derived from ancestral faunassharing similar biogeographic histories. Thedistance (airline) between the type localities ofBufo multiverrucosus (Auquimarca) and B.chavin (Palma Pampa) is 96 km (Fig. 6).
Forests of the eastern Andes contain a richdiversity of species. Within the past eightyears, intensive fieldwork in eastern centralPeru, has yielded 22 new species (19 anurans, 3reptiles) that have been described by Lehr andcolleagues—there yet remain many scientifi-cally unexplored regions in the eastern Andes.
RESUMEN
Se describe una nueva especie de Bufotentativamente asignada al grupo Bufoveraguensis procedente de bosques cerca aPaucartambo, Pasco, Peru a elevaciones de2600–3000 m en la Cordillera Oriental de Perucentral. La nueva especie difiere de todas lasespecies descritas por tener glandulas grandesy numerosas con muchas puntas queratiniza-das en el dorso, grandes glandulas en la tibia yantebrazo, y dos grandes glandulas dorsalesque estan entre las parotoides. La nueva
TABLE 2.—Ranges (in mm) and proportions of adultfemales of Bufo multiverrucosus. For abbreviations, see
Material and Methods.
Characters (n 5 12)
SVL 54.0–68.9TL 16.5–20.6FL 18.5–21.0HL 17.7–21.5HW 20.1–23.1ED 3.4–5.1TY 2.7–3.5IOD 6.0–7.2EW 3.7–5.1IND 3.5–5.1E–N 3.4–4.6PL 8.6–11.4PW 6.8–11.2TL/SVL 0.28–0.33FL/SVL 0.30–0.34HL/SVL 0.29–0.32HW/SVL 0.33–0.37HW/HL 1.06–1.22E–N/ED 0.67–1.10EW/IOD 0.54–0.90TY/ED 0.55–0.82
FIG. 6.—Map showing type localities of Bufo chavin(Palma Pampa, indicated with a square) and B. multi-verrucosus (Auquimarca, indicated with a star).
316 HERPETOLOGICA [Vol. 61, No. 3

especie es muy similar a Bufo chavin. Secomparan ambas especies junto a B. veraguen-sis con respecto a los caracteres externos yosteologıa del craneo. Se desconoce el rena-cuajo y canto de la nueva especie.
Acknowledgments.—Loan of material was kindly pro-vided by G. Kohler, J. H. Cordova, and L. Trueb. Portionsof the research were supported by a postdoctoral researchgrant given to E. Lehr by the Alexander von Humboldt-Foundation. C. Aguilar translated the abstract intoSpanish. We thank R. Acero for collecting and exportpermits issued by the Ministerio de Agricultura (INRE-NA), Lima, Peru.
LITERATURE CITED
CEI, J. M. 1980. Amphibians of Argentina. MonitoreZoologico Italiano (N. S.) Monographs 2:1–609.
DUELLMAN, W. E., AND R. SCHULTE. 1992. Descriptionof a new species of Bufo from northern Peru withcomments on phenetic groups of South American toads(Anura: Bufonidae). Copeia 1992:162–172.
FROST, D. R. 2004. Amphibian Species of the World: anOnline Reference. Version 3.0 (22 August, 2004). Elec-tronic Database accessible at http://research.amnh.org/herpetology/amphibia/index.html. American Muse-um of Natural History, New York, New York, U.S.A.
HOOGMOED, M. S. 1990. Biosystematics of South AmericanBufonidae, with special reference to the Bufo ‘‘typho-nius’’ group. Pp. 113–123. In G. Peters and R. Hutterer(Eds.), Vertebrates in the Tropics. Museum AlexanderKonig, Bonn, Germany.
LEHR, E. 2002. Amphibien und Reptilien in Peru: DieHerpetofauna entlang des 10. Breitengrades von Peru:Arterfassung, Taxonomie, okologische Bemerkungenund biogeographische Beziehungen. Natur und TierVerlag (NTV Wissenschaft), Munster, Germany.
LEHR, E. 2005. A new species of the Eleutherodactylusnigrovittatus group (Anura: Leptodactylidae) fromAndean Peru. Herpetologica. 61:199–208.
LEHR, E., G. KOHLER, C. AGUILAR, AND E. PONCE. 2001.New species of Bufo (Anura: Bufonidae) from CentralPeru. Copeia 2001:216–223.
LEHR, E., M. LUNDBERG AND C. AGUILAR, 2004. A newspecies of Phyllonastes from Peru (Anura: Leptodacty-lidae). Journal of Herpetology 38:214–218.
LEHR, E., M. LUNDBERG AND C. AGUILAR, In press. Threenew species of Phrynopus from central Peru (Amphibia,Anura, Leptodactylidae). Copeia.
LEVITON, A. E., R. H. GIBBS, JR., E. HEAL, AND C. E.DAWSON. 1985. Standards in herpetology and ichthyol-ogy: part I. Standard symbolic codes for institutionalresource collections in herpetology and ichthyology.Copeia 1985:802–832.
PRAMUK, J. B., AND E. LEHR. In press. Taxonomic status ofAtelophryniscus chrysophorus (Anura: Bufonidae) in-ferred from phylogeny. Journal of Herpetology.
SAVAGE, J. M., AND W. R. HEYER. 1997. Digital webbingformulae for anurans: a refinement. HerpetologicalReview 28:131.
SIMMONS, J. E. 1986. A method for prepartation of anuranosteological material. Pp. 37–39. In J. Waddingtonand D. M. Rudkin (Eds.), Proceedings of the 1985
Workshop on Care and Maintenance of Natural HistoryCollections. Life Sciences Miscellaneous Publications,Royal Ontario Museum, Ontario, Canada.
Accepted:27 March 2005Associate Editor: Joseph Mendelson III
APPENDIX I
Comparative Specimens Examined
Bufo chavin: MHNSM 20028 (holotype), MTD 42758–61, 44318, 44753, 43788–801, SMF 80001–008 (para-types): Peru: Departamento Huanuco: Provincia Pachitea:Distrito Chaglla: Palma Pampa (098 539 140 S, 758 539 210W, 3010 m), located approximately 20 km southeast ofChaglla, along the road from Chaglla to Tomayrica; SMF80002 (paratype): Peru: Departamento Huanuco: Provin-cia Pachitea: Distrito Chaglla: Chaglla (098 499 480 S, 758539 320 W, 3070 m).
Bufo veraguensis: KU 164084: Peru: Cuzco (skeleton);KU 139115–31: Peru: Cuzco; KU 163092: Peru: Ayacucho.
Osteological preparations.—Leptocactylidae: Leptodac-tylus ocellatus: KU 289186: Paraguay: Parque NacionalSan Rafael. Bufonidae: Atelophryniscus chrysophorus: KU206733: Honduras: Atlantıda: S. Slope Cerro Bufalo,Quebrada de Oro, 790 m [C&S]; Crepidophryne epiotica:KU 107394: Panama: Bocas del Toro [C&S]; Melano-phryniscus stelzneri: KU 93180: Argentina: San Luis[C&S]; Osornophryne bufoniformis: KU 144116 [C&S],KU 170103: Colombia: Cauca: Paramo Purace; QCAZ7684 [C&S]; Schismaderma carens: KU 195751: SouthAfrica: Transvaal [C&S]; KU 195749: South Africa: Natal;Truebella tothastes: KU 196605: Peru: Ayacucho: Carapa[C&S]. Bufonidae: Bufo: NORTH AMERICA: B. ameri-canus: KU 16444, 16457, 18194, 21146: USA: KansasDouglas Co. Lawrence, Haskell Bottoms; B. alvarius: KU14085, 25204: USA: Arizona; B. boreas: MVZ 142849:Canada: Vancouver Island; B. cognatus: KU 21149: USA:Kansas; B. debilis: KU 14099: USA: Texas; KU 73388:USA: New Mexico; B. exsul: MVZ 137717: USA:California; B. punctatus: KU 73390: USA: New Mexico;KU 9107, 15314, 20984–85, 73391: USA: New Mexico; B.quercicus: KU 19474–77: USA: Mississippi; B. woodhousii:KU 7170 18185, 18209, 18249: USA: Kansas; B. lemur: KU288710 [C&S], USNM 226397: Puerto Rico: Santa Isabel.CENTRAL AMERICA: B. coccifer: KU 68147–48: CostaRica: Puntarenas; KU 68151: Costa Rica: Cartago; B.holdridgei: KU 103462: Costa Rica: Heredia; B. valliceps:KU 59873: Guatemala: El Penten 10 km NNW Chinaga;KU 59874, KU 68155–56: Mexico: Chiapas; KU 52164Nicaragua: Manaqua. EURASIA: B. bufo: CAS 98078:Spain: Madrid; KU 144227: Portugal: Baixo Alentejo; B.macrotis: CAS 153004: Thailand: Nakhon Nayok Province;B. melanostictus: KU 129017: Singapore; KU 153941–42:Java: Central Java; B. stomaticus: KU 200370: India: Orissa,Sambalpur, Barpali. AFRICA: B. regularis: CAS 122180,131374: Kenya; B. xeros: CAS 122241, 122245: Kenya:Eastern Province Marsabit District. SOUTH AMERICA:Bufo crucifer group: B. crucifer: KU 93112: Brazil: EspiritoSanto; Bufo granulosus group: B. granulosus: KU 110431[C&S], 169346 [C&S], 10461, 110460: Colombia: Meta;KU 110462; KU 170090–92: Colombia: Magdalena;
September 2005] HERPETOLOGICA 317

B. mirandaribeiroi: USNM 28934: Brazil: Para to Manaos;Bufo guttatus group: B. blombergi: KU 69843: Colombia:Santander; KU 59763: Ecuador, Collari; B. guttatus: KU167631: Venezuela, Bolıvar; Bufo margaritifer group: B.typhonius: KU 93138: Brazil: Amapa; KU 104756 Ecua-dor: Napo; KU 127511: Brazil: Para; KU 205269: Peru:Madre de Dıos; Bufo marinus group: B. marinus: KU42566: Nicaragua: Managua; KU 69846: Mexico: AltaVerapaz; KU 84935–37: Nicaragua: Rivas; KU 152914:Ecuador: Napo; B. schneider: KU 160307: Argentina:Santiago del Estero; B. poeppigii: KU 183234–35: Bolivia:La Paz; Bufo spinulosus group: B. amabilis: KU 120365,120371, 124587: Ecuador: Loja; B. arequipensis: KU
214793: Peru: Arequipa; B. atacamensis: KU 217351:Chile: Coquimbo; B. chilensis: KU 217363: Chile:Santiago; B. cophotis: KU 218517–18: Peru: Cajamarca[C&S]; 218525–26: Peru: Cajamarca; B. limensis: KU209226: Peru: Lima; B. amabilis: KU 120365, 120365,124587: Ecuador: Loja; B. spinulosus: KU 160270–72:Bolivia: Potosı; KU 163032; Peru: Puno; KU 163036:Peru: Puno; KU 163066, 163074: Peru: Ayacucho;B. vellardi: KU 136053: Peru: Cajamarca; Bufo veraguensisgroup: B. chavin: MTD 43786–87: Peru: Huanuco, PalmaPampa; B. veraguensis: KU 164084: Peru: Cuzco;B. multiverrucosus: MTD 44751: Peru: Pasco, Paucar-tambo de Pasco Valley.
Herpetologica, 61(3), 2005, 318–325� 2005 by The Herpetologists’ League, Inc.
A NEW SPECIES OF THE ELEUTHERODACTYLUS DISCOIDALISGROUP (ANURA: LEPTODACTYLIDAE) FROM ANDEAN
HUMID MONTANE FORESTS OF BOLIVIA
JOSE M. PADIAL1, LUCINDO GONZALES
2, AND IGNACIO DE LA RIVA1,3
1Museo Nacional de Ciencias Naturales, CSIC. C/Jose Gutierrez Abascal 2, 28006 Madrid, Spain2Museo de Historia Natural Noel Kempff Mercado, P.O. Box 2489, Santa Cruz de la Sierra, Bolivia
A new species of the Eleutherodactylus discoidalis group from the Andean humid montane forests ofDepartamento La Paz, northern Bolivia, is described. The new species differs from other species of the groupby having regularly warty dorsal skin, a small tympanum, slightly enlarged, rounded digit discs, and uniformlydark brown coloration. Data on the advertisement call of the new species and an operational key for the speciesof the E. discoidalis group are provided.
Key words: Anura; Bolivia; Eleutherodactylus; Leptodactylidae; New species
THE Eleutherodactylus discoidalis Groupwas defined by Lynch (1976) on the basis ofseveral external characters: first finger longerthan second, head width ,45% of snout–ventlength, no cranial crests, ungual flaps of digitsnot indented, digits not expanded, and toeslacking webbing. He recognized five species:E. cruralis (Boulenger, 1902), E. discoidalis(Peracca, 1895), E. elassodiscus Lynch, 1973,E. granulosus (Boulenger, 1903) and E.nigrovittatus (Andersson, 1945). Subsequent-ly, Lynch, (1989) considered E. granulosusa junior synonym of E. cruralis, and split theformer E. discoidalis Group, rendering it onlywith the two southern species: E. discoidalis(Argentina and Bolivia) and E. cruralis (south-ern Peru and Bolivia) (De la Riva, 1993).
There were two synapomorphies proposed forthis group: 1) conical subarticular tubercles,and 2) supernumerary plantar tubercles. Re-cent studies in poorly surveyed areas of Boliviarevealed two new species of Eleutherodactylusbelonging to this group: E. zongoensis Reichleand Kohler, 1997 and E. ibischi Reichle,Lotters and De la Riva, 2001. Members ofthe E. discoidalis Group are terrestrial andinhabit different ecoregions: the Tucumanianand Yungas Forest (E. discoidalis), the AndeanMontane Rainforest (E. cruralis, E. zongoen-sis), the Amazonian Lowland Rainforest (E.cruralis), and the Inner Andean Dry Forest (E.ibischi and E. cruralis) (De la Riva et al., 2000;Kohler, 2000). Recent fieldwork in the humidmontane forests of Departamento La Paz,Bolivia, yielded new material of this group. Wemade morphological comparisons of some3 CORRESPONDENCE: e-mail, [email protected]
318 HERPETOLOGICA [Vol. 61, No. 3

specimens collected at two localities in MadidiNational Park and Pilon-Lajas Biosphere Re-serve, with specimens of all the species of theE. discoidalis group. We concluded that thesespecimens represent an unknown species,which is described herein.
MATERIALS AND METHODS
Specimens were fixed in 10% formalin andpreserved in 70% ethanol. For morphologicaland color characteristics used in the diagnosisand description, we followed Lynch (1989) andLynch and Duellman (1997). Specimens ex-amined are listed in the Appendix. Recordingequipment included a Sony WM D6C taperecorder and a Sennheiser Me 80 directionalmicrophone. Recordings were obtained on 18December 2003 at the type locality, during theevening and at sunset; air temperature was 21C. Eigth calls were analyzed. The sounds weredigitized at a sampling frequency of 44.1 KHzand 16 bit resolution with a Delta 66 digitizingboard and Peak 3.2 (OSX) software, and editedwith Audacity 1.2.2 (OSX). Praat 4.2.22 (OSX)software was used to obtain numerical in-formation and to generate audiospectrogramsand oscillograms. Frequency information wasobtained through fast Fourier transform (FFT)(width, 1024 points). Digitized calls weredeposited in the Fonoteca Zoologica, MuseoNacional de Ciencias Naturales, Madrid, TrackNumber 263. Measurements were taken witha digital caliper to the nearest 0.01 mm, butfollowing Hayek et al. (2001), for avoidingpseudo precision, we rounded all measure-ments to only one decimal. Abbreviations areas follows: snout–vent length, SVL; head width
(measured at level of rictus), HW; head length(from rictus to tip of snout), HL; interorbitaldistance, IOD; eye diameter (measured hori-zontally), ED; upper eyelid width, EW;tympanic membrane length, TYL; tympanicmembrane height, TYH; eye to nostril dis-tance, EN; tibia length, TL; foot length (fromproximal border of inner metatarsal tubercle totip of fourth toe), FL. Color characteristicswere noted in life. Museum abbreviations referto: Museo de Historia Natural Noel KempffMercado, Santa Cruz de la Sierra, Bolivia(MNK-A [Amphibian Collection]); ColeccionBoliviana de Fauna, La Paz, Bolivia (CBF);Museo de Historia Natural de la UniversidadMayor de San Marcos, Lima, Peru (MHNSM);United States National Museum of NaturalHistory, Smithsonian Institution, Washington,USA (USNM); Centro de Biodiversidad yGenetica, Universidad Mayor de San Simon,Cochabamba, Bolivia (CBG); ZoologischesForchungsinstitut und Museum AlexanderKoenig, Bonn, Germany (ZFMK); and MuseoNacional de Ciencias Naturales, Madrid, Spain(MNCN).
SYSTEMATICS
Eleutherodactylus madidi sp. nov.(Figs. 1, 2)
Holotype.—MNK-A 7856, an adult malefrom Arroyo Huacataya, Serranıa Eslabon,Area Natural de Manejo Integrado Madidi,Provincia Franz Tamayo, Departamento LaPaz, Bolivia (148 209 120 S, 688 059 570 W), ca.
FIG. 1.—Male of Eleutherodactylus madidi sp. nov.,holotype (SVL 28.87 mm). FIG. 2.—Male of Eleutherodactylus madidi sp. nov.,
MNCN 42015 (SVL 28.30 mm).
September 2005] HERPETOLOGICA 319

1500 m, obtained on 17 December 2003 byJ. M. Padial.
Paratypes.—MNK-A 7197 and MNCN42014–15, (adult males) paratopotypes, samedata as the holotype; MNK-A 4137 (adultmale) and MNK-A 4138 (adult female), fromLa Cascada, Biosphere Reserve Pilon Lajas,Provincia Sud Yungas, Departamento La Paz,Bolivia (158 249 000 S, 678 089 590 W), 900 m,collected on 10 July 1999 by L. Gonzales.
Diagnosis.—A member of the E. discoidalisGroup, as defined by Lynch (1989), character-ized by: (1) skin on dorsum heavily warty, wartsregular in size, low, small, rounded, some ofthem enlarged on the posterior part of the headand scapular region; warts on flanks larger andmore elongate than those of dorsum; ventersmooth; posterior surfaces of limbs uniformly
warty; discoidal fold present, barely visible; nodorsolateral folds; postrictal glands well de-veloped; (2) tympanic membrane and annulusdistinct, its length about 2/5 to 1/2 of eyelength; supratympanic fold short; (3) headlarge, longer than wide; snout round tosubacuminate in dorsal and lateral views;canthus rostralis rounded; (4) cranial crestsabsent; upper eyelid tubercles absent; (5)vomerine odontophores situated posteriorme-dial to choanae; (6) males with vocal slits anda single faint nuptial pad on thumb; (7) firstfinger longer than second; subarticulartubercles enlarged and subconical to conicalin profile; supernumerary tubercles small;terminal discs of fingers rounded, not enlarged(Fig. 3); (8) lateral fringes and keels on fingersabsent; (9) ulnar tubercles absent; (10) notubercles on heel and tarsus; (11) inner meta-tarsal tubercle ovoid, outer more rounded,subequal; (12) toes lacking lateral fringes orkeels; webbing absent; fifth toe shorter thanthird; discs of toes moderately enlarged,rounded (Fig. 3); (13) dorsal coloration darkbrown to black, with diffuse dark marks; inpreservative, dorsal regions brownish gray orpurplish gray; snout dark gray with darkermarks; throat cream with dense and finebrownish gray mottling; venter cream withscarce to dense brownish gray fine mottling.
Eleutherodactylus madidi can be distin-guished from E. cruralis by having: dorsal skinuniformly warty (smooth or shagreen with fewto many small, scattered warts in E. cruralis),rounded finger discs (truncate on Fingers IIIand IV), dorsum in life uniformly dark brown(brown color with many well defined dark andlight marks and flecks), and larger size of males(SVL of adult males of E. madidi 27.9–29.4 mm[n 5 5]; SVL of adult males of E. cruralis 18.3–25.0 mm [n 5 35]). Eleutherodactylus madidican be distinguished from E. discoidalis asfollows: vomerine odontophores posterior tochoanae (between choanae in E. discoidalis),finger and toe discs rounded (truncate), dorsalskin uniformly warty (shagreen), finger and toefringes absent (present), and dorsal colorationdark brown with scarce pattern (variable, withintense pattern). Eleutherodactylus madididiffers from E. ibischi by having tympanicmembrane equal or smaller than 1/2 of eyediameter (larger), first finger longer thansecond (equal), rounded finger discs (discs on
FIG. 3.—Hand and foot of the holotype of Eleuther-odactylus madidi sp. nov.
320 HERPETOLOGICA [Vol. 61, No. 3

Fingers III and IV two times wider than thedigits proximal to the disc), and dark browndorsal coloration (light brown to beige).Eleutherodactylus madidi can be distinguishedfrom E. zongoensis by its warty skin on dorsum(dorsum and extremities tuberculate in E.zongoensis), head longer than wide (widerthan long), smaller tympanic membrane (TYL/ED 5 0.41–0.56 in E. madidi [n 5 6], TYL/ED 5 0.60 in E. zongoensis [holotype]), toediscs round and moderately enlarged (trun-cate, small), labial bars present (absent), andventral region cream with fine mottling (uni-formly dark pinkish brown).
The species of the genus Ischnocnema sharemany morphological characters with the spe-cies of the Eleutherodactylus discoidalis Group(e.g., warty skin and lack of T-shaped digitaltips). In Bolivia three species have been re-ported: I. quixensis (Jimenez de la Espada,1872) in the lowland Amazonian rainforests ofDepartamento Pando, I. sanctaecrucis Harvey& Keck, 1995 in Andean cloud forests ofDepartamento Cochabamba and DepartamentoSanta Cruz, and I. sanderi Padial, Reichle andDe la Riva, 2005 in cloud forests of Departa-mento La Paz (De la Riva et al., 2000; Padial etal., 2005). Eleutherodactylus madidi is easy todistinguish from Ischnocnema quixensis and I.saxatilis Duellman, 1990 (from the Andes ofnorthern Peru); these species of Ischnocnemahave irregularly warty and tuberculate dorsalskin, a different coloration, and are much morerobust and larger. Eleutherodactylus madidi issmaller than I. sanctaecrucis (SVL of adultmales of E. madidi 27.9–29.4 mm [n 5 5]; SVLof adult males of I. sanctaecrucis 31.9–35.1 mm[n 5 3]), has slender body and extremities,possess warty skin (tuberculate in I. sanctae-crucis), lacks scarlet marks on dorsum (pres-ent), and lacks toe and finger fringes (present).Eleutherodactylus madidi differs from I. sand-eri, by lacking a W-shaped dorsal scapular fold(present in I. sanderi), lacking finger fringes(present), and having dorsum dark brown withdarker diffuse marks (light brown with welldefined dark marks).
Description of the holotype.—Head slightlynarrower than body, longer than wide; snoutrounded in dorsal view and lateral profile;nostrils slightly protuberant, oriented laterally;canthus rostralis indistinct, rounded in profile;loreal region slightly concave, sloping gradu-
ally to the lips; upper eyelid without tubercles;no cranial crests. Supratympanic fold distinct;tympanic membrane and tympanic annulusdistinct; tympanic membrane nearly round, itslength about 1/2 of eye length; tympanic mem-brane separated from eye by a distance ofalmost half the eye length; three postrictalglands. Choanae rounded, small, anterolateral,not concealed by palatal shelf of the maxillaryarch when roof of mouth is viewed from below;vomerine odontophores triangular, postero-medial to choanae, well developed, their ante-rior margin at the level of the posterior marginof the choanae, their width about 1.3 times thediameter of the choanae; vocal slits present,posterolateral to tongue. Skin of dorsal sur-faces and posterior parts of hind limbs finelywarty, most warts of the same size, with largerones on posterior surfaces of dorsum and onthe flanks; ventral surfaces almost smooth,with a finely granular texture on the throat;a thin middorsal fold from the anterior marginof the snout to the sacral region; two faint,sinuous, parallel occipital folds; no dorsolateralfolds; discoidal fold distinct.
No ulnar tubercles (only white spots);palmar tubercle moderately large, rounded,larger than elongate thenar tubercle; supernu-merary tubercles small, subconical, smallerthan subarticular tubercles; subarticulartubercles large, subconical to round; discs offingers rounded, slightly wider than thecorresponding digits; fingers lacking lateralfringes and keels; nuptial pads absent; relativelength of fingers 2 , 4 , 1 , 3 (Fig. 3).
Heel and tarsus lacking tubercles or folds;inner metatarsal tubercle ovoid, longer thanwide, almost the same size as the conical outer;supernumerary tubercles small, rounded; sub-articular tubercles large, conical, directed to-ward tip of digit; toes lacking lateral fringes andkeels; discs rounded, slightly enlarged, about1.3 times the width of the corresponding digit;relative length of toes 1 , 2 , 5 , 3 , 4 (Fig. 3).
Measurements (in mm).—SVL 28.9; headwidth 9.4; head length 10.5; interorbitaldistance 2.8; eye diameter 3.8; upper eyelidwidth 2.3; tympanic membrane length 2.1;tympanic membrane height 2.2; eye to nostrildistance 3.5; tibia length 15.2; foot length 14.5.For proportions, see Table 1.
Coloration.—In life, dorsum mostly darkbrown, with some dark, diffuse marks; dorsal
September 2005] HERPETOLOGICA 321

surfaces of extremities dark brown with finebrown mottling and diffuse bars; loreal regionand lips lighter, lips with diffuse dark brownbars (Fig. 1); supratympanic fold dark brownto black; ventral regions cream with finebrown mottling, more intense on extremities;inner surfaces of hind limbs brown; a whitishcream line departs from the proximal part ofthe posterior surface of each thigh and goesabove the level of the vent, forming a paren-theses-shaped pattern around it; plantar sur-faces brown, with gray and white tubercles.The coloration in preservative is similar.
Variation.—There is little variation amongspecimens from Eslabon, but some differencesdo exist between them and the specimens from LaCascada. In thespecimens fromEslabon,nonehasa nuptial pad, while the male from La Cascada hasa faint nuptial pad on each thumb; one male fromEslabon (MNCN 42015), has a fine, bright yellowmiddorsal line from the tip of the snout to thecoccyx (Fig. 2). The specimens from La Cascadahad orange tones in the ventral regions, postrictalglands, axillary regions, andgroins. The single malefrom La Cascada has larger thenar and palmartubercles and more developed supernumerarytubercles; the tympanic membrane is less evidentin the two specimens from this locality. The singlefemale (MNK-A 4138) is larger (SVL 33.0 mm)than the males (SVL 27.9–29.4 mm; n 5 5), haslighter dorsal coloration, the head is longerthan wide, and the vomerine odontophores arerounded to slightly oval, low and almost in contact.
Advertisement call.—The call consists ofa single pulsed note of variable duration (meanduration, 988.7 ms; range, 926–1075; n 5 8)and regular pulse repetition, with 29.6 pulsesper call (28–32) (Fig. 4); the pulse rate is 29.9pulses/s (29.3–30.3) and the dominant fre-quency is at 2436.4 Hz (2411.7–2584); thereare no harmonics. The call repetition rate is 4.3calls/minute; often, two calls are emitted to-gether in a sequence and, in this case, the firstcall is shorter than the second. The call of E.madidi is longer and has a much highernumber of pulses than the call of E. cruralis(228–413 ms, 12–17 pulses) and E. ibischi(117–193 ms, 6–8 pulses); the pulse rate ishigher in both E. cruralis (41–53 pulses/s) andE. ibischi (36–43 pulses/s), and the dominantfrequency is higher in E. cruralis (3300–3420Hz) but similar in E. ibischi (2000–2847 Hz)(see Marquez et al., 1995; Reichle et al., 2001).These characteristics allow a clear distinctionof the three species by their call.
Distribution and ecology.—Eleutherodacty-lus madidi inhabits the humid montane forestsof the Andean slopes of northern Bolivia atelevations of 900–1500 m (Fig. 5). The speciesis only known from La Cascada and SerranıaEslabon, two localities of Departmento La Pazseparated by an airline distance of 150 km. It isa terrestrial species that calls mainly at dusk,from natural cavities in the ground, and it is dif-ficult to find. The single female (MNK-A 4138),collected on 10 July (dry season) is gravid.
TABLE 1.—Measurements and proportions of the typespecimens of Eleutherodactylus madidi sp. nov. (M,male; F, female; for other abbreviations, see Material
and Methods).
MNK-A7856 (M)
MNCN42014 (M)
MNCN42015 (M)
MNK-A7197 (M)
MNK-A4137 (M)
MNK-A4138 (F)
SVL 28.9 27.9 28.3 28.5 29.4 33.0HW 9.4 9.9 9.7 9.9 10.8 10.8HL 10.5 12.1 10.9 10.1 10.7 11.3IOD 2.8 2.4 2.5 2.1 2.6 3.0ED 3.8 3.6 4.3 3.7 3.7 4.0EW 2.3 2.6 2.8 2.2 2.8 2.4TYL 2.1 2.0 1.8 1.5 1.6 1.7TYH 2.2 2.3 2.5 2.2 1.6 1.7EN 3.5 3.2 3.3 3.8 3.6 3.6TL 15.2 15.6 14.5 15.2 16.3 16.9FL 14.5 13.7 13.9 14.4 15.0 15.8HL/HW 1.12 1.21 1.13 1.02 0.99 1.03TYL/ED 0.55 0.57 0.43 0.41 0.43 0.42EN/ED 0.93 0.90 0.78 1.01 0.98 0.90IOD/ED 0.74 0.68 0.59 0.56 0.72 0.75TL/SVL 0.53 0.56 0.51 0.55 0.55 0.58
FIG. 4.—Oscillogram and spectrogram of a 1 s section ofthe call of Eleutherodactylus madidi sp. nov.; airtemperature, 21 C.
322 HERPETOLOGICA [Vol. 61, No. 3

Etymology.—The specific name refers tothe Madidi National Park, where the speciesoccurs. Madidi is one of the most importantprotected areas in the world in terms ofbiodiversity, and it holds a highly diverseamphibian fauna, which is still poorly in-ventoried. By naming a frog species after thepark, we aim to call attention on Madidi’sbiological richness and the need of safeguard-ing this natural area for future generations.
DISCUSSION
We conclude that the new species belongs tothe Eleutherodactylus discoidalis Group pro-posed by Lynch (1989); this conclusion is wellsupported by different morphological and callcharacteristics. Although some differences werefound among the specimens from the twolocalities studied, we do not consider that theyrepresent different species; we conclude that thisrepresents simple interpopulational variations.
With E. madidi, the E. discoidalis Group iscomposed of five species sharing the putativesynapomorphies proposed by Lynch (1989).
The five species occur in Bolivia, one inArgentina (E. discoidalis) and one in Peru (E.cruralis). We provide an operational key forthe species of the E. discoidalis Group, but itmust be taken into account that the phyloge-netic relationships and putative monophyly ofthis group are still obscure. Especially, therelationships between the E. discoidalis Groupand other Eleutherodactylus and how thegenus Ischnocnema relates to them, deservefuture attention.
OPERATIONAL KEY TO THE SPECIES OF THE
ELEUTHERODACTYLUS DISCOIDALIS GROUP
1. Vomerine odontophores posterior to choanae - - - - - - 2Vomerine odontophores between choanae - - - - - - - - - - - - - - - -
- - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - E. discoidalis2. Dorsal skin uniformly warty or tuberculate - - - - - - - - - 3
Dorsal skin smooth or shagreen with scatteredirregular tubercles or warts - - - - - - - - - - - - - - - - - - - - - - - - - 4
3. Terminal disc of fingers and toes rounded;tympanic membrane 2/5 to 1/2 of the eyediameter - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - E. madidi
Terminal discs of Fingers III and IV truncate;tympanic membrane evident, large, longerthan 1/2 of the eye diameter - - - - - - - - - E. zongoensis
FIG. 5.—Distribution of Eleutherodactylus madidi sp. nov. in the Amazonian slopes of Departamento La Paz, Bolivia.The square represents the type locality.
September 2005] HERPETOLOGICA 323

4. Tympanic membrane evident, longer than 1/2of eye diameter; discs of Fingers III and IVtwo times wider than the correspondingdigits; dorsum beige - - - - - - - - - - - - - - - - - - - - - - - - E. ibischi
Tympanic membrane evident, shorter than 1/2 ofeye diameter; discs of Fingers III and IV atmost one and a half times wider than thecorresponding digits - - - - - - - - - - - - - - - - - - - - - - - E. cruralis
RESUMEN
Se describe una nueva especie del grupo deEleutherodactylus discoidalis de los bosqueshumedos andinos del Departmento de La Paz,en el norte de Bolivia. La nueva especie se difer-encia de otras especies del grupo por tener pieldorsal uniformemente rugosa, tımpano peque-no, discos digitales redondeados y ligeramenteagrandados y color marron oscuro bastanteuniforme. Se proporciona una clave artificialpara las especies del grupo de E. discoidalis.
Acknowledgments.—For the help provided during manyyears, we are grateful to the personnel at the Museo deHistoria Natural Noel Kempff Mercado, especially to M.Suarez, R. Vespa, A. Justiniano, and R. Montano. R.Aguayo (Centro de Biodiversidad y Genetica), C. Aguilarand J. Cordova (Museo de Historia Natural de laUniversidad Nacional Mayor de San Marcos), J. Aparicio(Coleccion Boliviana de Fauna), W. Bohme and J. Kohler(Zoologisches Forchungsinstitut und Museum AlexanderKoenig), and W. R. Heyer (U. S. National Museum ofNatural History, Smithsonian Institution), provided work-ing facilities and specimens for study. R. Marquez and G.Solıs (Fonoteca Zoologica, Museo Nacional de CienciasNaturales) provided assistance with sound analysis. Wethank R. McDiarmid (U. S. National Museum of NaturalHistory, Smithsonian Institution) and S. Reichle (TheNature Conservancy) for providing funds for parts of thefieldwork, and P. Comas, D. Embert, and P. Macuapa fortheir company during it. The Director of the MadidiNational Park, O. Loayza, kindly accepted our researchproject. We are extremely grateful to the people of SanJose de Uchupiamonas and to all the personnel of theChalalan Ecolodge, for their support, help, and friendship,and we are especially indebted to its manager, G. Mamani,for allowing us to use the outstanding Chalalan facilities asour base camp. Padial’s research was financed by a grant ofthe Mutis programme of the MAE-AECI (Spain), and histrip to visit the museums in USA was funded by an ErnstMayr Travel Grant in Animal Systematics (Museum ofComparative Zoology, Harvard University). This work waspartially funded by the project REN/GLO 2001-1046 ofthe Spanish Ministry of Science and Technology, and bya project of the AECI (Spanish Agency of InternationalCooperation) (I. De la Riva, Principal Investigator) toinventory Madidi’s herpetofauna.
LITERATURE CITED
DE LA RIVA, I. 1993. Sinopsis del genero Eleutherodactylus(Amphibia, Anura, Leptodactylidae) en Bolivia y adicionde tres nuevas especies para el paıs. Revista Espanola deHerpetologıa 7:97–105.
DE LA RIVA, I., J. KOHLER, S. LOTTERS, AND S. REICHLE.2000. Ten years of research on Bolivian amphibians:updated checklist, distribution, taxonomic problems,literature and iconography. Revista Espanola de Her-petologıa 14:19–164.
HAYEK, L. A., W. R. HEYER, AND C. GASCON. 2001. Frogmorphometrics: a cautionary tale. Alytes 18:153–177.
KOHLER, J. 2000. Amphibian diversity in Bolivia: a studywith special reference to montane forest regions.Bonner zoologische Monographien 48:1–243.
LYNCH, J. D. 1976. The species groups of the SouthAmerican frogs of the genus Eleutherodactylus (Lep-todactylidae). Occasional Papers of the Museum ofNatural History, The University of Kansas 61:1–24.
———. 1989. Intrageneric relationships of mainlandEleutherodactylus (Leptodactylidae). I. A review of thefrogs assigned to the Eleutherodactylus discoidalisspecies group. Milwaukee Public Museum, Contribu-tions in Biology and Geology 79:1–25.
LYNCH, J. D., AND W. E. DUELLMAN. 1997. Frogs of thegenus Eleutherodactylus (Leptodactylidae) in westernEcuador: systematics, ecology, and biogeography. TheUniversity of Kansas Natural History Museum SpecialPublications 23:1–236.
MARQUEZ, R., I. DE LA RIVA, AND J. BOSCH. 1995.Advertisement calls of Bolivian Leptodactylidae (Am-phibia, Anura). Journal of Zoology, London 237:213–136.
PADIAL, J. M., S. REICHLE, AND I. DE LA RIVA. 2005. Newspecies of Ischnocnema (Anura, Leptodactylidae) fromthe Andes of Bolivia. Journal of Herpetology 39:186–191.
REICHLE, S., AND J. KOHLER. 1997. A new species ofEleutherodactylus (Anura: Leptodactylidae) from theAndean slopes of Bolivia. Amphibia-Reptilia 18:333–337.
REICHLE, S., S. LOTTERS, AND I. DE LA RIVA. 2001. A newspecies of the discoidalis group of Eleutherodactylus(Anura, Leptodactylidae) from Inner-Andean DryValleys of Bolivia. Journal of Herpetology 35:21–26.
Accepted:28 March 2005Associate Editor: Joseph Mendelson III
APPENDIX
Material Examined
Eleutherodactylus cruralisBOLIVIA: Departamento La Paz: Boqueron, Pilon
Lajas, CBG 792–93; Camino maderero El Chaval, ArroyoMikai, Reserva Pilon Lajas, MNK-A 3759–61; PuertoLinares, 360 m, USNM 281100–30; 5 km W of SanBuenaventura, USNM 280617; Departamento Cocha-bamba: Valle de Sajta, MNK-A 3633; Villa Tunari, MNK-A 1492; Los Guacharos MNK-A 6617–19; Entre Paractito yEl Palmar, ZFMK 72541; Departamento Pando: Florida,Reserva Manuripi, MNK-A 5086; Lago Bay, ReservaManuripi, MNK-A 6120–21; Departamento Beni: Asun-cion, Pilon Lajas, MNK-A 4074; Laguna Azul, ReservaPilon Lajas, MNK-A 3975–76, 3979, 3985–87, 4003; SanLuis Chico, MNK-A 4027; Serranıa del Pilon, MNK-A4182–83, 4209–13. PERU: Departamento Madre de Dios:Puerto Maldonado, 30 km SSW of Tambopata, USNM284267, 343240, 342989–92; Colpa de Guacamayos, ZonaReservada Tambopata-Candamo, USNM 332436–37.
324 HERPETOLOGICA [Vol. 61, No. 3

Eleutherodactylus discoidalis
BOLIVIA: Departamento Tarija: 12.3 km NW of EntreRıos, on the road to Tarija, MNK-A 3877–97.
Eleutherodactylus ibischi
BOLIVIA: Departamento Santa Cruz: Km 68.5 on SantaCruz de la Sierra-Samaipata road, 750 m elevation, CBF3341 (holotype); Km 60 on Santa Cruz de la Sierra-Samaipata road, MNK-A 6612.
Eleutherodactylus zongoensisBOLIVIA: Departamento La Paz: Valle de Zongo, 1250
m, CBF 2503 (holotype).
Ischnocnema quixensis
BOLIVIA: Departamento Pando, CBF 2528–29; RıoNegro, MNK-A 6525–27.
Ischnocnema sanctaecrucis
BOLIVIA: Departamento Santa Cruz: El Chape, 2060m elevation, MNK-A 1198 (holotype); MNCN 42010–13.
Ischnocnema saxatilis
PERU: Departamento San Martın: Ponga de Shilcayo,about 4 km NNW of Tarapoto, 470 m (068 319 S, 768 539),MHNSM 8431 (paratype).
Herpetologica, 61(3), 2005, 325–336� 2005 by The Herpetologists’ League, Inc.
PHYLOGENETIC RELATIONSHIPS OF THE GENUSPROCTOPORUS SENSU STRICTO (SQUAMATA:
GYMNOPHTHALMIDAE), WITH A NEW SPECIESFROM PUNO, SOUTHEASTERN PERU
TIFFANY M. DOAN1,4,5, TODD A. CASTOE
2, AND WILFREDO ARIZABAL ARRIAGA3
1Biology Department, Vassar College, Box 555, 124 Raymond Avenue, Poughkeepsie, NY 12604-0555, USA2Department of Biology, University of Central Florida, 4000 Central Florida Blvd,
Orlando, FL 32816-2368, USA3Museo de Historia National Cusco, Universidad Nacional de San Antonio Abad de Cusco,
Urb. Los Angeles E-1, Cusco, Peru
The genus Proctoporus sensu stricto is a poorly known gymnophthalmid lizard clade distributed across theAndes of southern Peru and Bolivia. Recent collecting efforts in central and southern Peru recoveredspecimens and tissues of all known members of the genus, enabling the first complete phylogeny of the genusto be constructed. In addition, a new species was found in Puno, Peru and is described herein. We analyzedDNA sequences of three mitochondrial genes using maximum parsimony and Bayesian MCMC methods toreconstruct a phylogeny of the group. The phylogeny suggests an ancient split between a newly discoveredlineage from Puno and the remaining species that coincides geographically with its isolated range. Proctoporuspachyurus and P. sucullucu form sister species; P. bolivianus forms a clade with P. unsaacaeþ P. guentheri.The elevationally restricted ranges of all known Proctoporus species likely have contributed to the high speciesdiversity found in southern Peru. Both allopatric and parapatric modes of speciation are proposed to explainthe diversification of Proctoporus species.
Key words: Andes; Gymnophthalmidae; New species; Peru; Phylogeny; Proctoporus; Puno; SouthAmerica; Squamata; Taxonomy
OF THE 31 described species originallyincluded in the genus Proctoporus, five were
recognized to be in the P. pachyurus speciesgroup by Doan and Castoe (2003) and Uzzell(1970). Through a phylogenetic study includ-ing a broad array of gymnophthalmid lizardtaxa, Castoe et al. (2004) found that Proctopo-rus species did not form a monophyletic group,and that members of the P. pachyurus species
4 PRESENT ADDRESS: Department of Biology, CentralConnecticut State University, 1615 Stanley Street, NewBritain, CT 06050, USA.
5 CORRESPONDENCE: e-mail, [email protected]
September 2005] HERPETOLOGICA 325

group did form a monophyletic group distantlyrelated to a majority of other Proctoporus.Based on these data, Doan and Castoe (2005)separated the P. pachyurus group from all otherProctoporus species by removing all other spe-cies from the genus Proctoporus sensu strictoand placing them in separate genera. Allmolecular phylogenetic analyses to date, how-ever, have not sampled the species P. pachyu-rus nor have they included all members of thespecies traditionally allied with P. pachyurus(i.e., all species of Proctoporus sensu stricto).
Proctoporus sensu stricto is distributedthroughout the Andes of central and southernPeru and Bolivia (Doan and Castoe, 2003).Within Proctoporus, four species are restrictedto the extreme high elevations of the centralAndes, occurring 2600–4080 m above sea level(Doan, 2003; T. Doan, personal observation).The fifth species, P. guentheri, occurs at lowerelevations (1000–3200 m; Doan, 2003). Col-
lectively, these montane species inhabit anarray of Andean habitats including cloud forestand puna.
Although specimens of all currently recog-nized species are represented in various mu-seums, tissues for molecular analysis onlyrecently have become available for certainmembers of the genus (Castoe et al., 2004;Doan and Castoe, 2003). Recent field collec-tions in the central and southern PeruvianDepartments of Apurimac, Cusco, Junın, andPuno allowed the completion of a tissue col-lection of the entire genus (see map, Fig. 1),and the first complete molecular phylogeneticreconstruction of the genus. Here we combineour molecular data for Proctoporus with thelarge gymnophthalmid dataset of Castoe et al.(2004) and re-evaluate the monophyly of thenewly redesignated genus. In addition toreconstructing the phylogeny of all knownProctoporus species, our field investigations
FIG. 1.—Map of southern Peru and neighboring countries depicting the sampling localities for Proctoporus species inthis study. Country frontiers are indicated by thicker black lines. Departmental boundaries of Peru are indicated by thingrey lines; departmental names are in italics. The following symbols are used for each species: five-sided star 5 P.pachyurus; four-sided star 5 P. guentheri; pentagon 5 P. sucullucu; X 5 P. unsaacae; circle 5 P. bolivianus; triangle 5P. new species; square 5 P. sp. 3.
326 HERPETOLOGICA [Vol. 61, No. 3

and subsequent analyses revealed a new line-age of Proctoporus. These specimens wereoriginally identified as P. guentheri based ontheir morphology and the fact that they werefirst encountered at a relatively low elevation(2100 m). Subsequent morphological and mo-lecular analyses, however, determined that thislineage is quite distinct from its congenersand that it actually consists of two separatesister species.
MATERIALS AND METHODS
Specimens of Proctoporus were obtainedthroughout the Peruvian range of the genus(Fig. 1; see Doan and Castoe, 2003, for Cuscolocalities). Specimens were collected by hand,euthanized, fixed in 10% formalin, and latertransferred to 70% ethanol for long-term mu-seum storage. The specimens were depositedat the University of Texas at Arlington Col-lection of Vertebrates (UTA) and the Museode Historia Natural, Universidad Nacional deSan Antonio Abad de Cusco (MHNC; formerlyabbreviated UNSAAC) in Peru. Liver tissuewas taken from all individuals and stored intissue lysis buffer (0.5M Tris, 0.25% EDTA,2.5% SDS).
In addition to the specimens collected in thefield, supplemental museum specimens wereexamined from KU, MHNC, USNM, UTA,and the Gabinete de Zoologıa, UniversidadNacional de San Antonio Abad de Cusco (GZ).Museum abbreviations follow Leviton et al.(1985) except for MHNC and GZ. All speci-mens examined are listed in Appendix I. Mea-surements were made with a digital caliper tothe nearest 0.1 mm. All anatomical terms andmethods of taking meristic counts followKizirian (1996) except as modified by Doan(2003) and Doan and Schargel (2003).
In addition to morphological analysis ofspecimens, we reconstructed a three-gene mo-lecular phylogeny of Proctoporus from south-ern Peru and included a recently collectedspecimen of P. bolivianus from Bolivia. Weadded our sequences to the expanded gym-nophthalmid dataset of Castoe et al. (2004;which included data from Doan and Castoe,2003; and Pellegrino et al., 2001). Our mo-lecular sampling of Proctoporus includedmultiple individuals of each described speciesin the genus and three individuals of the newlydiscovered lineage.
Whole cellular DNA was extracted fromliver tissue using the DNeasy DNA extractionkit (Qiagen). A fragment of the mitochondrialNADH dehydrogenase subunit 4 gene andadjacent tRNAs (hereafter referred to collec-tively as ND4) was PCR amplified using theprimer pair ND4 and Leu as in Arevalo et al.(1994) for all specimens that were collected.Additionally, mitochondrial small and largeribosomal subunit genes (12S and 16S) wereamplified for selected specimens as describedin Parkinson (1999) and Parkinson et al.(1997). Positive PCR products were excisedout of agarose electrophoretic gels and puri-fied using the GeneClean III kit (Bio101).Purified PCR products were quantified anddirectly sequenced using the CEQ D Dye Ter-minator Cycle Sequencing (DTCS) QuickStart Kit (Beckman Coulter), run on a Beck-man CEQ2000 automated sequencer.
Raw sequence chromatographs were editedusing Sequencher 4.2 (2004 Gene CodesCorp.). Sequences were added to the existingalignment of gymnophthalmid lizards fromCastoe et al. (2004) and aligned to this datasetby eye. These sequences were later recheckedfor positive alignment based on inferred aminoacid sequence (protein-coding region) inGenedoc (Nicholas and Nicholas, 1997). Gapsin alignment were treated as ambiguities forphylogenetic analyses. The final alignment con-sisted of a combined total of 93 OTU’s and1640 aligned positions: 860 from ND4 (in-cluding tRNAs), 331 from 12S, and 449 from16S. All new sequences were deposited inGenBank under the accession numbers listedin Appendix II.
We reconstructed phylogenies based on themaximum parsimony (MP) criterion in PAUP*v4.0b10 (Swofford, 2002) and Bayesian (Mar-kov Chain Monte Carlo, MCMC) phylogeneticanalysis in MrBayes v3.0b4 (Ronquist andHuelsenbeck, 2003). Phylogenetic inferencewas conducted on the combined concatenateddataset including all three genes. For MPanalyses we conducted equally-weighted par-simony searches using the heuristic strategywith 100 random taxon addition sequencereplicates. Settings for MP analyses were treebisection-reconnection branch swapping,steepest descent off, and MULTREES optionon (Swofford, 2002). We assessed support forclades in MP analyses using 100 nonparamet-
September 2005] HERPETOLOGICA 327

ric bootstrap (Felsenstein, 1985) pseudorepli-cates with 10 random taxon addition sequencereplicates implemented in PAUP*.
ModelTest version 3.0 (Posada and Cran-dall, 1998) was used to infer the best-fit modelof evolution for the combined dataset based onboth AIC and hLTR model selection criteriaavailable in the program (see also Huelsenbeckand Crandall, 1997; Posada and Crandall,2001). Each MCMC run employed the modelselected by ModelTest (see results).
All MCMC phylogenetic reconstructionswere conducted in MrBayes (Ronquist andHuelsenbeck, 2003) with vague priors (as perthe program’s defaults) and model parametersestimated as part of the analyses. Three heatedchains and a single cold chain were used in allMCMC analyses and runs were initiated withrandom trees. Trees were sampled every 100generations and majority-rule consensus phy-lograms and posterior probabilities for nodeswere assembled from all post burn-in sampledtrees. Four independent MCMC runs wereconducted to confirm stationarity was reachedand that no single run was trapped on local(rather than global) optima and that indepen-dent runs converged on similar stationary pa-rameter estimates. Each independent MCMCrun employed a total of 2.5 million gener-ations, 500,000 of which were discarded asburn-in, yielding 2 million post burn-in gen-erations per run.
RESULTS
The combined dataset consisted of 831 con-stant characters and 686 parsimony informativecharacters. Mean uncorrected genetic dis-tances between species ranged between9.3% and 13.8% (Table 1). The parsimonyanalysis resulted in 41,310 equally-parsimoni-ous trees of 6796 steps (CI 5 0.206, RI 5
0.557). The strict consensus of these trees ispresented in Fig. 2A with nodes receiving,50% bootstrap support shown as collapsed.Overall, the results of MP failed to resolvebasal relationships among Proctoporus speciesand also failed to resolve the monophyly ofthe genus.
ModelTest selected the General Time Re-versible (GTR; Tavare, 1986) model withgamma distributed among-site rate variation(G; Yang, 1993) and an estimated proportionof invariant sites (I) as the best-fit model ofevolution (GTRþGþI model) based on bothAIC and hLTR model selection criteria. Thismodel was used for all Bayesian MCMC runs.Independent Bayesian MCMC analyses re-sulted in extremely similar parameter esti-mates (including likelihoods and posteriorprobabilities) and each rapidly ascended toa stationary likelihood plateau. Thus, all post-burn-in generations from the four indepen-dent runs were combined to estimate param-eters and posterior probabilities for clades.The marginal likelihood across all post-burn-inruns (total of 8 million generations, sampledevery 100) had an arithmetic mean of lnL 5�29878.92 and a harmonic mean of lnL 5�29906.74. The mean and 95% credibilityinterval of parameters of the GTRþGþI model,based on results from the combined fourMCMC runs, are as follows: rG-T 5 1.00(1.00–1.00), rC-T 5 3.96 (3.23–4.71), rC-G 50.18 (0.11–0.26), rA-T 5 0.56 (0.44–0.69), rA-G 5 3.99 (3.38–4.69), rA-C 5 0.509 (0.41–0.63), pi(A) 5 0.39 (0.38–0.41), pi(C) 5 0.28(0.26–0.29), pi(G) 5 0.08 (0.08–0.09), pi(T) 50.24 (0.23–0.26), gamma 5 0.57 (0.53–0.62),pInvar. 5 0.43 (0.40–0.46).
The Bayesian MCMC reconstruction isshown in Fig. 2B with clades receiving ,50%posterior probability collapsed. Four distinct
TABLE 1.—Mean genetic distance between species of Proctoporus. Uncorrected sequence divergence is given below thediagonal (bottom-left) and Kimura-2-parameter genetic distance is given above the diagonal (top-right).
P. bolivianus P. guentheri P. pachyurus P. subsolanus P. sp. 3 Laracani P. sucullucu P. unsaacae
P. bolivianus 0.136 0.147 0.157 0.154 0.153 0.145P. guentheri 0.122 0.133 0.136 0.142 0.140 0.100P. pachyurus 0.130 0.119 0.104 0.102 0.106 0.147P. subsolanus 0.138 0.122 0.096 0.101 0.138 0.157P. sp. 3 Laracani 0.137 0.127 0.094 0.094 0.129 0.146P. sucullucu 0.137 0.126 0.097 0.124 0.117 0.154P. unsaacae 0.130 0.093 0.131 0.139 0.131 0.137
328 HERPETOLOGICA [Vol. 61, No. 3

clades are evident in the Bayesian reconstruc-tion. One clade contains an undescribed line-age of the specimens collected from Sandiaand Laracani, both in the Department ofPuno, Peru. A second clade contains P.pachyurus and P. sucullucu. The third cladeis made up of P. bolivianus; the final cladecontains P. guentheri and P. unsaacae, re-covered as the sister clade to P. bolivianus.Monophyly of Proctoporus sensu stricto issupported by 94% posterior probability. Mono-phyly of each individual species is supportedwith 100% posterior probability support ex-cept for P. bolivianus. One individual ofP. bolivianus from Santa Cruz, Bolivia appearsto be distantly related to other members ofits species.
The maximum parsimony and Bayesian re-constructions do not have any nodes in con-flict, but the parsimony tree suffers from poorresolution. Similarly, both of the reconstruc-tions presented here are congruent with thephylogeny of Castoe et al. (2004) at all nodes.
The specimens from Puno clearly representan undescribed clade of Proctoporus. Moreover,the specimens from Sandia and Laracani appeardistantly related to each other with 9.4%uncorrected sequence divergence (Table 1).Additionally, the morphology of the specimensfrom Sandia and Laracani is quite distinct, withdifferences in numbers of supratympanic tem-porals, coloration, and the presence of a lorealscale. These two populations are separated fromeach other by 1000 m in elevation.
FIG. 2.—Results of phylogenetic analyses including all species of Proctoporus based on 1640 bases of mitochondrialDNA sequence per individual. Individuals of a species are indicated by letters in parentheses and coincide with AppendixII and Castoe et al. (2004). (A) Strict consensus of 41,310 equally parsimonious trees from maximum parsimonyphylogenetic analysis with nodes collapsed if bootstrap support ,50%. Bootstrap values for nodes are shown adjacent tonode if �50%. Nodes with 100% bootstrap values are indicated with a gray square. Branch lengths are not informative.(B) Bayesian MCMC phylogenetic reconstruction phylogram based on 8 million post-burn-in generations with nodescollapsed if posterior probability support ,50%. Posterior probability values for nodes are shown adjacent to nodeif �50%. Nodes with 100% posterior probability are indicated with a grey circle.
September 2005] HERPETOLOGICA 329

Herein, we describe the sixth species ofProctoporus sensu stricto from Sandia,Peru. We believe that the specimen fromLaracani represents an additional distinctspecies, but with only one juvenile speci-men collected we cannot construct a di-agnosis adequate to describe that species.Therefore, we leave the Laracani specimenas an undescribed species until furthercollection produces an adequate type seriesfor that lineage (Proctoporus sp. 3).
SYSTEMATIC ACCOUNT
Proctoporus subsolanus sp. nov.
Holotype.—UTA R-52944 (Fig. 3), a gravidadult female, from the town of Sandia(14.342758 S, 69.462748 W), Province ofSandia, Department of Puno, Peru; 2100 m;collected on 19 June 2003 by Tiffany M. Doan.
Paratypes.—UTA R-52946, an adult male;52947, a subadult male; 52948, and MHNC
FIG. 3.—Head of the holotype of Proctoporus subsolanus (UTA R-52944). Top left–dorsal view; Top right–ventral view;Bottom–lateral view. Scale bar 5 5 mm.
330 HERPETOLOGICA [Vol. 61, No. 3

TMD1269, adult females; all from Sandia(14.355098 S, 69.468878 W); 2221 m; collectedon 20 June 2003 by Todd A. Castoe andWilfredo Arizabal Arriaga.
Referred specimens.—MHNC TMD1266,MHNC TMD1268, MHNC TMD1270, samedata as paratypes.
Diagnosis.—(1) frontonasal longer thanfrontal; (2) supraoculars two; (3) superciliaries3–4, first expanded onto dorsal surface of head;(4) palpebral eye-disc made up of a single,undivided scale; (5) supralabials 5–6; (6)infralabials five; (7) dorsal body scalesquadrangular, with high rounded keel; (8)transverse rows of dorsals 33–37; (9) transverseventral rows 22–24; (10) a continuous series ofsmall lateral scales separating dorsals fromventrals; (11) posterior cloacal plate made upof six scales in both sexes; (12) femoral poresper hind limb in males 5–8, in females 0–3;(13) preanal pores absent; (14) subdigitallamellae on Toe IV 15–17; (15) limbs not over-lapping when adpressed against body in adults;(16) lateral body surfaces without ocelli, orwith very few in both sexes.
Specimens of Proctoporus subsolanus showthe presence of an undivided palpebral eye-disc, which identify them as members of thegenus Proctoporus, as opposed to Riama orPetracola (Doan and Castoe, 2005; Uzzell,1970). Proctoporus subsolanus can be distin-guished from P. guentheri and P. pachyurusby having two supraoculars (P. guentheri 3,P. pachyurus 4). It can be distinguished fromP. unsaacae by lacking a continuous series oflateral ocelli. It can be distinguished fromP. sucullucu by limbs not overlapping whenadpressed. It can be distinguished fromP. bolivianus by having the frontonasal muchlonger than frontal (P. bolivianus has frontalequal length to frontonasal). It can be distin-guished from P. sp. 3 by having fewersubdigital lamellae (21 on Toe IV in theundescribed species).
Description of holotype.—Adult female,gravid, snout–vent length (SVL) 43.6 mm,original complete tail 70.2 mm; head scalessmooth, glossy, without striations or rugosities;rostral scale wider than tall, meeting supra-labials on either side at above the height ofsupralabials, and becoming higher medially,in contact with frontonasal, nasals, and firstsupralabials; frontonasal longer than wide, six-
sided, anterior sutures rectangular, posteriorsutures forming a 1108 angle, in contact withnasals, anteromost superciliary, and frontal,longer than frontal; prefrontals absent; frontallonger than wide, pentagonal, in contact withanteromost superciliary on left side only, incontact with first supraocular on left side, withfirst and second supraoculars on right side, incontact with frontoparietals; frontoparietalshexagonal, in contact with first and secondsupraoculars on left and second and thirdsupraoculars on right, in contact with parietalsand interparietal; supraoculars two on left sideand three on right, first supraocular on rightside in contact with first superciliary only, firston left side and second on right side in contactwith first three superciliaries, second on leftside and third on right side in contact withfourth superciliary, parietal, and postocular;interparietal longer than wide, heptagonal, incontact with parietals and occipitals; parietalspolygonal, with posterior sutures in contactwith occipital, lateral sutures diagonally incontact with subequally large temporal, bor-dering postocular anteriorly, second supra-ocular, and frontoparietal; occipitals three,smaller than parietals, median smallest, ex-tending further posteriorly than two lateraloccipitals. Nasal entire with no separate lorealscale on left side, on right side a partial sutureextending from ventral portion of scale intonostril, longer than high, nostril situated inanterior third of scale, in contact with first andsecond supralabials, first superciliary, andfrenocular; four superciliaries, first expandedonto dorsal surface of head; two preoculars,first in contact with first superciliary andfrenocular, second in contact with frenocularand first subocular; frenocular roughly tri-angular, dorsalmost corner in contact with firstsuperciliary, in contact with second and thirdsupralabials, first and second preoculars, andfirst subocular; palpebral eye-disc made up ofa single transparent scale; suboculars three;postoculars two; temporals smooth, glossy,polygonal; supratympanic temporals two;supralabials five, first four supralabials to angleof jaw; infralabials five. Mental wider thanlong, in contact with first infralabial andpostmental posteriorly; postmental single,pentagonal, posterior suture angular withpoint directed posteriorly, in contact with firstand second infralabials and first pair of genials;
September 2005] HERPETOLOGICA 331

three pairs of genials, anterior pair in contactwith second and third infralabials; secondgenials in contact with third and fourthinfralabials; third pair in contact with fourthand fifth infralabials; one pair of chin shields,separated by irregular pregulars; gular scalerows seven; collar fold slightly distinct, con-cealing two rows of small scales; lateral neckscales round, smooth.
Dorsals rectangular, longer than wide, withanterior and posterior margins slightly con-vexly curved, juxtaposed, with single highrounded keel, in 37 transverse rows; someparavertebral scales irregularly arranged; lon-gitudinal dorsal scale rows 20 at fifth trans-verse ventral scale row, 23 at tenth transverseventral scale row, 24 at fifteenth transverseventral scale row; continuous lateral scaleseries, two to three scales wide, smaller thandorsals, partially hidden in lateral fold, re-duced scales at limb insertion regions present;transverse ventral scale rows 22; longitudinalventral scale rows at midbody 12; anteriorpreanal plate scales paired; posterior preanalplate scales six, lateralmost scales small; scaleson tail rectangular, juxtaposed; dorsal anddorsolateral caudal scales with keel, ventraland ventrolateral caudal scales smooth; mid-ventral subcaudal scales wider than adjacentscales, almost square, anteromost midventralsubcaudal scales subimbricate.
Limbs pentadactyl; digits clawed; dorsalbrachial scales polygonal, subequal in size,subimbricate, smooth; ventral brachial scalesroundish, subimbricate, smooth; antebrachialscales polygonal, subequal in size, smooth,ventral antebrachial scales smallest; scales ondorsal surface of manus polygonal, smooth,subimbricate; scales on palmar surface of ma-nus small, rounded, subimbricate, domelike;thenar scales two, smooth laterally, raised intodomes medially; finger length formula IV . III. II . V . I; scales on dorsal surfaces offingers smooth, quadrangular, covering dorsalhalf of digit, overhanging supradigital lamellae,3 on I, 5 on II, 6 on III, 5/6 on IV, 4 on V;subdigital lamellae 5 on I, 8 on II, 9 on III, 12on IV, 6 on V; scales at base of Finger V thickerthan adjacent scales. Scales on anterodorsalsurface of thigh large, polygonal, smooth,subimbricate; scales on posterior surface ofthigh small, rounded, juxtaposed; scales onventral surface of thigh large, rounded, flat,
smooth; femoral pores 2/1; preanal poresabsent; scales on anterior surface of cruspolygonal, smooth, juxtaposed, decreasing insize distally; scales on anterodorsal surface ofcrus rounded, juxtaposed; scales on ventralsurface of crus large, smooth, flat, subimbri-cate; toe length formula IV . III . V . II . I;scales on dorsal surface of digits single,quadrangular, smooth, of varying sizes, over-hanging supradigital lamellae, 3 on I, 5 on II, 7on III, 11 on IV, 7 on V; subdigital lamellaesingle distally, double proximally, 5 on I, 8 onII, 13 on III, 16 on IV, 11 on V; limbsnot overlapping when adpressed against thebody, separated by eight to nine dorsal scalelengths.
Coloration in preservative.—Dorsal surfaceof head dark brown with lighter brown faintmottling; lateral surface of head like dorsalsurface, but with more light mottling, lip ir-regularly barred with cream coloring; ventralsurface of head yellowish cream with clumpsof black stippling on each scale; pregularregion like head but with fainter stippling,medial scales lack stippling; gular region likehead but with denser stippling per scale,forming longitudinal clumps. Dorsal surfaceof body nearly same color as head, but slightlymore grey and with less mottling, two faintlight brown dorsolateral longitudinal stripesoriginate near occiput and extend to forelimbinsertions; lateral surface of body same color-ation as dorsum, fading to more mottling andlighter brown near venter; ventral surface ofbody with yellowish cream ground color,lateralmost scales with black stippling, medial-most scales lack stippling. Limbs similar tobody, dorsal surface of arms darkly mottledwith dark coloration decreasing towards ven-tral surface, ventral surface of arms yellowishcream without black stippling, dorsal surfaceof legs with more mottling than body, ventralsurface of legs with cream ground color anddispersed grey stippling. Dorsal tail colorationlike that of body, ventral surface of tail lightbrown with dark brown mottling anteriorly,ground color of tail becoming orange withbrown mottling posteriorly and distal portionof tail dark brown with lighter mottling (samecoloration as dorsal surface).
Coloration in life.—Dorsal, lateral, andventral surfaces are similar to coloration inpreservative. Dorsolateral longitudinal stripes
332 HERPETOLOGICA [Vol. 61, No. 3

can be seen to extend from occiput onto baseof tail. Ventral surface of head whitish cream;ventral surface of body yellow.
Variation.—Adult females SVL 40.7–42.6mm, adult male SVL 47.3 mm; complete orig-inal tail of adult female 63.4, no adult maleswith complete original tails known. The para-types and referred specimens are very similarto the holotype with the following minorexceptions. Suparoculars on the right side ofthe holotype’s head appear to be anomalous.No other specimens have three supraoculars.UTA R-52946 has only two occipitals that donot meet medially because the interparietalextends posteriorly and fills the space betweenthe occipitals. UTA R-52946, 52947, and52948 have a partial nasal suture extendinginto the nostril from below, but also havea second partial diagonal suture that extendsfrom the junction with the frenocular, espe-cially on the left side. MHNC TMD1266has three superciliaries. In UTA R-52946,52947, and 52948 the frenocular does notmake contact with the first superciliary andthree postoculars are present. In UTAR-52947 there are three preoculars. MHNCTMD1266 and TMD1268 have six supra-labials. In UTA R-52947 and 52948 there aretwo genial pairs and two chin shield pairs; thefirst pair of chin shields is separated by twopregular scales.
Coloration is quite similar among the speci-mens with some having a lighter dorsum andmore distinct longitudinal stripes. Stippling onthe venter is more distinct or greyer in somespecimens and nearly absent in others. In UTAR-52946 lateral surfaces of the body are orangeand the tail and hind limbs are infused withorange. The ventral surface of the body is orangelaterally and the tail is pink on the originalportion and dark grey on the regeneratedportion. UTA R-52946 has one lateral ocellusof black surrounding a white dot anterior to theforelimb insertions. UTA R-52947 has threeindistinct lateral white spots that do not formocelli that begin anterior to the forelimb in-sertion and continue to approximately midbody.
Meristic variation includes: transverse dor-sal rows 33–37; longitudinal dorsal rows atmidbody 21–23; longitudinal ventral rows 10–12; transverse ventral rows 22–24; subdigitallamellae on Finger IV 10–14; subdigitallamellae on Toe IV 15–21.
Sexual dimorphism is slight in this specieswith males having wider heads (relative toSVL) and two males (UTA R-52946 and52947) are more colorful than the females.Femoral pore number is also sexually di-morphic, with males possessing 6–8 femoralpores per leg and females having 0–3.
Distribution.—Proctoporus subsolanus isknown only from the type locality in northernPuno, in the Sandia Province. This area lieswithin the Cordillera de Apolobamba, on theeasternmost flank of the central Andes ofsouthern Peru. Specimens of this species werefound between 2100 and 2221 m. This slopeabruptly drops into the lowland rainforest ofthe Tambopata National Reserve.
Habitat and ecology.—Specimens of thisspecies were found within and around thetown of Sandia often on agricultural terraceswhere maize and other crops were being cul-tivated. These areas were probably once cov-ered in cloud forest before human occupationof the area by the Incan civilization. All spec-imens were found beneath stones on theground. Stomach contents were analyzed infour specimens and consisted of coleopterans,hymenopterans (adult and larval ants), isopods,and some unknown arthropod legs. Two gravidfemales were found to contain two eggs each.
Etymology.—The specific epithet is a Latinadjective meaning eastern. This name refers tothe species occurring on the easternmost flankof the Andes Mountains in southern Peru. TheCordillera de Apolobamba is the final moun-tain range before the mountains descend intothe Amazon Basin.
DISCUSSION
The known range of the genus Proctoporussensu stricto is restricted to the departments ofJunın, Ayacucho, Apurımac, Cusco, and Punoof central and southern Peru and the depart-ments of La Paz, Santa Cruz, and Cochabambaof western Bolivia. Species of Proctoporus areknown to have limited altitudinal ranges, withfour members of the genus inhabiting veryhigh elevations (.4000 m; Doan, 2003; Doanand Castoe, 2003). Proctoporus guentheri andthe new species P. subsolanus inhabit lowerelevations (as low as 1000 m for P. guentheri;Doan, 2003) and forested habitats that differgreatly from those of the high Andean species.
September 2005] HERPETOLOGICA 333

Despite our thorough sampling in CuscoDepartment (see Doan and Castoe, 2003), andless extensive but widespread sampling inApurımac, Junın, and Puno, only one localityof sympatry was found among the six species ofProctoporus. At the village of Ollantaytambo,Cusco, P. sucullucu and P. unsaacae werefound to occur syntopically. At this locality, alldiagnostic characters appear to remain dis-crete between the species and no externalmorphological evidence of introgression isapparent (this is currently being investigatedusing molecular markers by the authors).These preliminary observations suggest thatProctoporus species retain their specific iden-tity even in syntopic situations, implying theexistence of reproductive isolating mecha-nisms other than geographic isolation suffi-cient to prevent observable interspecificgeneflow.
The phylogeny of the genus suggests anancient split between the lineage from north-ern Puno and the remaining species. Procto-porus subsolanus occurs on the cis-Andeanslope where the mountains begin their plungeinto the Amazon Basin. Such a distributionsubstantially isolates them from other mem-bers of the genus with high mountains to thewest and the barren altiplano to the south.
We find no obvious biogeographic explana-tion to account for the patterns of diversityacross the other Proctoporus lineages. Theonly species from central Peru, P. pachyurusis most closely related to P. sucullucu fromApurımac and Cusco. Proctoporus bolivianusforms a sister relationship with P. unsaacae þP. guentheri (Fig. 2). Proctoporus unsaacae isthe most common species in Cusco, occurringat most of the sampled localities in greatabundance. Proctoporus guentheri occurs inthe same general regions as P. bolivianus but atlower elevations (Doan and Castoe, 2003). Theancestor of P. unsaacae and P. guentheri mayhave speciated parapatrically, with populationsat high elevations giving rise to P. unsaacae andthose at lower elevations becoming P. guen-theri. A similar scenario may explain the diver-sification of the newly discovered Puno lineagewhereby sister species (P. subsolanus and P.sp. 3) appear to have partitioned their alti-tudinal distribution. Speciation of other Proc-toporus clades may have been allopatric, buttheir current distributions do not appear
bounded by any obvious extant barriers togene flow. Overall, the level of geographic pop-ulation sampling in the current study providesonly cursory evidence for historical bioge-ographic hypotheses. Currently, we are con-ducting an expanded population genetic studyacross all available populations in the hopes ofelucidating more detailed estimates of histor-ical genetic and demographic patterns.
RESUMEN
El genero Proctoporus sensu stricto es unclado gymnophthalmido poco conocido queesta distribuido en los Andes del Peru y Bolivia.Colecciones recientes en el sur y centro delPeru registraron especımenes y tejidos detodos los miembros del genero y permiten laprimera filogenia completa del genero. Ade-mas, encontramos una nueva especie en Puno,Peru y la describimos aquı. Analizamos lassecuencias de ADN de tres genes mitocon-driales con analisis de parsimonia maxima yBayesian MCMC para construir una filogeniadel grupo. La filogenia indica que una divisionantigua occurio entre el linaje de Puno y elresto de las especies. Esta division coincidegeograficamente con su ambito aislado. Proc-toporus pachyurus y P. sucullucu son especieshermanas; P. bolivianus forma un clado con P.unsaacae y P. guentheri. Los rangos altitudi-nales restringidos de todas las especies con-ocidas de Proctoporus probablamente hancontribuido a la alta diversidad de las especiesde Proctoporus del sur del Peru. Ambos modosde especiacion alopatrico y parapatrico sonpropuestos para explicar la diversificacion delgenero Proctoporus.
Acknowledgments.—This study was partially funded bygrants from Women’s International Science Collaboration(American Association for the Advancement of Science)to TMD and WAA and the Vassar College Collins Fa-culty Research Fund to TMD. We thank J. Degner, R.Ruggiero, and J. Stainkamp for their suggestions on themanuscript. We thank C. Parkinson for providing labora-tory space and resources for molecular aspects of thisproject. We thank J. A. Campbell and C. J. Franklin forcataloging our specimens. We also thank M. RomeroPastor, R. Acero Villanes, and K. Ramirez of the InstitutoNacional de Recursos Naturales of Peru for facilitatingcollecting and export permits and S. J. Orenstein forlaboratory assistance. In addition, we thank the personnelat the following museums for loans of material under theircare: AMNH, GZ, KU, MHNC, USNM, and UTA.
334 HERPETOLOGICA [Vol. 61, No. 3

LITERATURE CITED
AREVALO, E. S., S. K. DAVIS, AND J. W. SITES, JR. 1994.Mitochondrial DNA sequence divergence and phyloge-netic relationships of the Sceloporus grammicus com-plex (Phrynosomatidae) in central Mexico. SystematicBiology 43:387–418.
CASTOE, T. A., T. M. DOAN, AND C. L. PARKINSON. 2004.Data partitions and complex models in Bayesiananalysis: the phylogeny of gymnophthalmid lizards.Systematic Biology 53:448–469.
DOAN, T. M. 2003. A south-to-north biogeographichypothesis for Andean speciation: evidence from thelizard genus Proctoporus (Reptilia, Gymnophthalmi-dae). Journal of Biogeography 30:361–374.
DOAN, T. M., AND T. A. CASTOE. 2003. Using morphologicaland molecular evidence to infer species boundarieswithin Proctoporus bolivianus Werner (Squamata:Gymnophthalmidae). Herpetologica 59:433–450.
DOAN, T. M., AND T. A. CASTOE. 2005. Phylogenetictaxonomy of the Cercosaurini (Squamata: Gymno-phthalmidae), with new genera for species of Neusticu-rus and Proctoporus. Zoological Journal of the LinneanSociety 145:403–416.
DOAN, T. M., AND W. E. SCHARGEL. 2003. Bridging the gapin Proctoporus distribution: a new species (Squamata:Gymnophthalmidae) from the Andes of Venezuela.Herpetologica 59:68–75.
FELSENSTEIN, J. 1985. Confidence limits on phylogenies:an approach using the bootstrap. Evolution 39:783–791.
HUELSENBECK, J. P., AND K. A. CRANDALL. 1997. Phylogenyestimation and hypothesis testing using maximumlikelihood. Annual Review of Ecology and Systematics28:437–466.
KIZIRIAN, D. A. 1996. A review of Ecuadorian Proctoporus(Squamata: Gymnophthalmidae) with descriptions ofnine new species. Herpetological Monographs 10:85–155.
LEVITON, A. E., R. H. GIBBS, JR., E. HEAL, AND C. E.DAWSON. 1985. Standards in herpetology and ichthyol-ogy: part I. Standard symbolic codes for institutionalresource collections in herpetology and ichthyology.Copeia 1985:802–832.
NICHOLAS, K. B., AND H. B. NICHOLAS, JR. 1997. GeneDoc:a tool for editing and annotating multiple sequencealignments. Distributed by the author at: www.cris.com/;Ketchup/genedoc.shtml.
PARKINSON, C. L. 1999. Molecular systematics and bio-geographical history of pitvipers as determined bymitochondrial ribosomal DNA sequences. Copeia 1999:576–586.
PARKINSON, C. L., S. M. MOODY, AND J. E. ALQUIST. 1997.Phylogenetic relationships of the ‘Agkistrodon Complex’based on mitochondrial DNA sequence data. Pp. 63–78.In R. S. Thorpe, W. Wuster, and A. Malhotra (Eds.),Venomous Snakes: Ecology, Evolution, and SnakebiteSymposia of the Zoological Society of London. Clar-endon Press, Oxford, U.K.
PELLEGRINO, K. C. M., M. T. RODRIGUES, Y. YONENAGA-YASSUDA, AND J. W. SITES. 2001. A molecular perspectiveon the evolution of microteiid lizards (Squamata,Gymnophthalmidae), and a new classification for thefamily. Biological Journal of the Linnean Society74:315–338.
POSADA, D., AND K. A. CRANDALL. 1998. Model-Test:testing the model of DNA substitution. Bioinformatics14:817–818.
POSADA, D., AND K. A. CRANDALL. 2001. Selecting the best-fit model of nucleotide substitution. Systematic Biology50:580–601.
RONQUIST, F., AND J. P. HUELSENBECK. 2003. MrBayes 3:Bayesian phylogenetic inference under mixed models.Bioinformatics 19:1572–1574.
SWOFFORD, D. L. 2002. PAUP*. Phylogenetic AnalysisUsing Parsimony (*and Other Methods), Version4.0b10. Sinauer Associates, Sunderland, Massachusetts.
TAVARE, S. 1986. Some probabilistic and statistical prob-lems on the analysis of DNA sequences. Pp. 57–86. In R.M. Miura (Ed.), Some Mathematical Questions inBiology—DNA Sequence Analysis. American MathSociety, Providence, Rhode Island, U.S.A.
UZZELL, T. M. 1970. Teiid lizards of the genus Proctoporusfrom Bolivia and Peru. Postilla 142:1–39.
YANG, Z. 1993. Maximum likelihood estimation of phylog-eny from DNA sequences when substitution rates differover sites: approximate methods. Molecular Biology andEvolution 10:1396–1401.
Accepted: 19 April 2005Associate Editor: Joseph Mendelson III
APPENDIX I
Specimens Used in Morphological Analysis
Specimen localities are given according to museumcatalog information. Cataloged localities and spellingswere used as is without correction of errors.
Proctoporus bolivianus: BOLIVIA: La Paz: Murillo,Valle de Zongo, Estacion Hidroelectrica Cuti, Khuchu(UTA R-39113); Santa Cruz: Caballero, Canton San Juan,Amboro National Park (AMNH R-150695); PERU: Cusco:500 m up road Carizales (UTA R-51485); 800 m up roadCarizales (UTA R-51486–51487); Canchayoc (UTA R-51483–51484); Cerro de Puquin (GZ 0027); Cerro MachuPicchu (UTA R-51509–51511); Cochayoc (MHNCWAA5024, UTA R-51481–51482); Cosnipata (GZ 0043);Nusta Hispana (USNM 60699–60700, 60748 [holotype ofP. obesus]); Ollantaytambo (USNM 49549 [paratype of P.lacertus], 60719); 25 km NNE Paucartambo, AbraAcanacu (KU 163801, 163804, 163810–163811, 163814,163820, 163827, 163830–163831, 163834, 163836,163839–164740, 163842, 163846); 29 km NNE Paucar-tambo, Abra Acanacu (KU 13965); 31 km NNE Paucar-tambo, Abra Acanacu (KU 13958, 13963); Piscacucho(MHNC AC136–AC141, UTA R-51501–51508); Tinco-chaca (USNM 49551 [holotype of P. lacertus]); Torontoy(USNM 60726–60727).
Proctoporus guentheri: BOLIVIA: La Paz: Nor Yungas,Serrania de Bella Vista, 17 km from Carrasco towardsSapecho (UTA R-39114); PERU: Cusco: Chachabamba(UTA R-51518); Chocalloc (UTA R-51512–51515); MachuPicchu (KU 135157–135160, UTA R-51516–51517); 5 kmWSW Santa Isabel (KU 139307, 139309); 6 km NE SantaIsabel (KU 163939); Winaywayna (MHNC WAA5056).
September 2005] HERPETOLOGICA 335

Proctoporus pachyurus: PERU: Junın: Palca (KU135095, KU 181919–181922, 181924, 181927); NW Palca(KU 181917–181918, 181923, 181926); 1 km NW Palca(KU 181925, 181929–181931); 2 km E Palca (KU 181938–181939); 4 km W Palca (KU135096–135097); 5 km W Palca(KU135099).
Proctoporus subsolanus: PERU: Puno: Sandia, town ofSandia (UTA R-52944 [holotype], UTA R-52946, 52947,52948 [paratypes], MHNC TMD1266, TMD1268,TMD1269 [paratype], TMD1270).
Proctoporus sucullucu: PERU: Cusco: Kusilluchayoc(MHNC WAA5006, UTA R-51478); Piscacucho (UTA R-51496 [holotype], 51497–51500 [paratypes]).
Proctoporus unsaacae: PERU: Cusco: Pisac, Viacha(UTA R-51475–51476, 51479); QuelloUno (MHNCAC132, UTA R-51488 [holotype], 51489–51495 [para-types]); Saqsayhuaman (GZ Valdinos3, UTA R-51477);Urcos (UTA R-51480).
Proctoporus sp. 3: PERU: Puno: Sandia, village ofLaracani (UTA R-52945).
APPENDIX II Proctoporus Specimens Sampled for Molecular Analysis with Museum and GenBank Accession Numbers forEach Gene. Letters in parentheses refer to individuals of a species and coincide with letters from Castoe et al. (2004).
Species Locality Museum ND4 12S 16S
Proctoporus bolivianus (h) Bolivia: Santa Cruz: Amboro AMNH R-150695 AY968812 AY968821 AY968828Proctoporus bolivianus (d) Peru: Cusco: Canchayoc UTA R-51483 AY225176Proctoporus bolivianus (f) Peru: Cusco: Canchayoc UTA R-51484 AY225182 AY968820 AY968827Proctoporus bolivianus (c) Peru: Cusco: Carizales UTA R-51486 AY225179Proctoporus bolivianus (b) Peru: Cusco: Carizales UTA R-51487 AY225180 AY507850 AY507868Proctoporus bolivianus (g) Peru: Cusco: Cochayoc UTA R-51481 AY225181Proctoporus bolivianus (e) Peru: Cusco: Cochayoc UTA R-51482 AY225183Proctoporus bolivianus (a) Peru: Cusco: Piscacucho, 3600 m UTA R-51506 AY225175 AY507851 AY507869Proctoporus guentheri (c) Peru: Cusco: Chocalloc UTA R-51512 AY225184Proctoporus guentheri (a) Peru: Cusco: Chocalloc UTA R-51515 AY225185 AY507849 AY507872Proctoporus guentheri (d) Peru: Cusco: Machu Picchu UTA R-51516 AY225168Proctoporus guentheri (b) Peru: Cusco: Machu Picchu UTA R-51517 AY225169 AY507854 AY507873Proctoporus pachyurus (a) Peru: Junın: Muruhuay UTA R-52949 AY968816 AY968824 AY968834Proctoporus pachyurus (b) Peru: Junın: Palca MHNC TMD1203 AY968815 AY968823 AY968829Proctoporus subsolanus (a) Peru: Puno: Sandia UTA R-52944 AY968814 AY968826 AY968833Proctoporus subsolanus (b) Peru: Puno: Sandia UTA R-52946 AY968811 AY968822 AY968831Proctoporus sucullucu (e) Peru: Apurımac: Abancay UTA R-52950 AY968817 AY968830Proctoporus sucullucu (a) Peru: Cusco: Kusilluchayoc UTA R-51478 AY225171 AY507857 AY507878Proctoporus sucullucu (d) Peru: Cusco: Piscacucho, 3048 m UTA R-51500 AY225188Proctoporus sucullucu (b) Peru: Cusco: Piscacucho, 3191 m UTA R-51496 AY225177 AY507858 AY507879Proctoporus sucullucu (c) Peru: Cusco: Piscacucho, 3300 m UTA R-51498 AY225187Proctoporus unsaacae (d) Peru: Cusco: Pisac UTA R-51475 AY225174 AY968819Proctoporus unsaacae (e) Peru: Cusco: Pisac UTA R-51479 AY225172 AY968818Proctoporus unsaacae (b) Peru: Cusco: Quello Uno UTA R-51488 AY225186 AY507859 AY507882Proctoporus unsaacae (c) Peru: Cusco: Quello Uno UTA R-51493 AY225178Proctoporus unsaacae (a) Peru: Cusco: Saqsayhuaman UTA R-51477 AY225170 AY507860 AY507881Proctoporus unsaacae (f) Peru: Cusco: Urcos UTA R-51480 AY225173Proctoporus sp. 3 Peru: Puno: Laracani UTA R-52945 AY968813 AY968825 AY968832
336 HERPETOLOGICA [Vol. 61, No. 3
Copyright © 2022 FDOKUMEN




















