
Factors affecting the vulnerability of cane toads (Bufo marinus) to predation by ants: PREDATION ON...
14
Factors affecting the vulnerability of cane toads (Bufo marinus) to predation by ants GEORGIA WARD-FEAR, GREGORY P. BROWN and RICHARD SHINE* School of Biological Sciences A08, University of Sydney, NSW 2006, Australia Received 9 September 2009; accepted for publication 28 October 2009Experimental evidence on the determinants of prey vulnerability is scarce, especially for vertebrates in the field. Invasive species offer robust opportunities to explore prey vulnerability, because the intensity of predation on or by such animals has not been eroded by coevolution. Around waterbodies in tropical Australia, native meat ants (Iridomyrmex reburrus) consume many metamorph cane toads (Bufo marinus, an invasive anuran). We document the determinants of toad vulnerability, especially the roles of toad body size and ant density. Larger metamorphs were attacked sooner (because they attracted more ants), but escaped more often. Overall, smaller toads were more likely to be killed. Ant densities influenced toad responses, as well as attack rate and success. Data on the immediate outcomes of attacks underestimate mortality: more than 73% of apparent ‘escapees’ died within 24 h. Because mortality during this period was independent of toad size, predation was less size selective than suggested by immediate outcomes. © 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 99, 738–751. ADDITIONAL KEYWORDS: body size – natural selection – predator–prey – Rhinella marina – size- dependent mortality. INTRODUCTION Few biologists doubt that predation constitutes an important cause of mortality for many species in the wild, that predation is nonrandom with respect to phenotypic traits of the prey as well as environmental conditions, and that such biases impose selection on prey attributes (e.g. phenotypic characteristics, Fox, 1975; movement and habitat use, Sih, 1988; diet and feeding habits, Godin, 1990). Nonetheless, there have been surprisingly few field studies to quantify the determinants of prey vulnerability, especially in ver- tebrates (Shine et al., 2001). Although such studies have documented the effects of attributes, such as size and sex, on prey vulnerability (e.g. Trexler, Tempe & Travis, 1994; Acharya, 1995), the causal mechanisms generating such patterns generally remain specula- tive (Endler, 1986). For most systems involving ver- tebrates as prey, there are logistical and/or ethical obstacles to conducting manipulative studies on the ways in which specific traits affect the outcome of an interaction with a predator (Shine et al., 2001). Another logistical problem is that, if predation selects strongly on specific traits, we expect rapid evolution- ary change to reduce phenotypic variance in those traits, or to change other traits in ways that reduce the fitness penalty (predation vulnerability) of some invariant trait (such as sex; Sih, 1994). Typically, variance in the trait under selection will be decreased rapidly by sustained selection. Lastly, wild verte- brates are difficult to observe over a representative range of times and places of activity, and are likely to be particularly vigilant and easily disturbed under situations of high predation risk (Holomuzuki & Short, 1990). Thus, despite the putative central importance of predation in the ecology of many vertebrate popula- tions, detailed quantitative evidence on the predator– prey interaction is scarce. That scarcity is especially frustrating for traits such as body size, which we might expect to influence predation vulnerability in many systems. Plausibly, a prey animal’s body *Corresponding author. E-mail: [email protected] Biological Journal of the Linnean Society, 2010, 99, 738–751. With 8 figures © 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 99, 738–751 738
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Factors affecting the vulnerability of cane toads (Bufo marinus) to predation by ants: PREDATION ON...

Factors affecting the vulnerability of cane toads(Bufo marinus) to predation by ants
GEORGIA WARD-FEAR, GREGORY P. BROWN and RICHARD SHINE*
School of Biological Sciences A08, University of Sydney, NSW 2006, Australia
Received 9 September 2009; accepted for publication 28 October 2009bij_1395 738..751
Experimental evidence on the determinants of prey vulnerability is scarce, especially for vertebrates in the field.Invasive species offer robust opportunities to explore prey vulnerability, because the intensity of predation on orby such animals has not been eroded by coevolution. Around waterbodies in tropical Australia, native meat ants(Iridomyrmex reburrus) consume many metamorph cane toads (Bufo marinus, an invasive anuran). We documentthe determinants of toad vulnerability, especially the roles of toad body size and ant density. Larger metamorphswere attacked sooner (because they attracted more ants), but escaped more often. Overall, smaller toads were morelikely to be killed. Ant densities influenced toad responses, as well as attack rate and success. Data on theimmediate outcomes of attacks underestimate mortality: more than 73% of apparent ‘escapees’ died within 24 h.Because mortality during this period was independent of toad size, predation was less size selective than suggestedby immediate outcomes. © 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010,99, 738–751.
ADDITIONAL KEYWORDS: body size – natural selection – predator–prey – Rhinella marina – size-dependent mortality.
INTRODUCTION
Few biologists doubt that predation constitutes animportant cause of mortality for many species in thewild, that predation is nonrandom with respect tophenotypic traits of the prey as well as environmentalconditions, and that such biases impose selection onprey attributes (e.g. phenotypic characteristics, Fox,1975; movement and habitat use, Sih, 1988; diet andfeeding habits, Godin, 1990). Nonetheless, there havebeen surprisingly few field studies to quantify thedeterminants of prey vulnerability, especially in ver-tebrates (Shine et al., 2001). Although such studieshave documented the effects of attributes, such as sizeand sex, on prey vulnerability (e.g. Trexler, Tempe &Travis, 1994; Acharya, 1995), the causal mechanismsgenerating such patterns generally remain specula-tive (Endler, 1986). For most systems involving ver-tebrates as prey, there are logistical and/or ethicalobstacles to conducting manipulative studies on the
ways in which specific traits affect the outcome of aninteraction with a predator (Shine et al., 2001).Another logistical problem is that, if predation selectsstrongly on specific traits, we expect rapid evolution-ary change to reduce phenotypic variance in thosetraits, or to change other traits in ways that reducethe fitness penalty (predation vulnerability) of someinvariant trait (such as sex; Sih, 1994). Typically,variance in the trait under selection will be decreasedrapidly by sustained selection. Lastly, wild verte-brates are difficult to observe over a representativerange of times and places of activity, and are likely tobe particularly vigilant and easily disturbed undersituations of high predation risk (Holomuzuki &Short, 1990).
Thus, despite the putative central importance ofpredation in the ecology of many vertebrate popula-tions, detailed quantitative evidence on the predator–prey interaction is scarce. That scarcity is especiallyfrustrating for traits such as body size, which wemight expect to influence predation vulnerabilityin many systems. Plausibly, a prey animal’s body*Corresponding author. E-mail: [email protected]
Biological Journal of the Linnean Society, 2010, 99, 738–751. With 8 figures
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 99, 738–751738

size influences its nutritional value to a predator(thus potentially increasing its likelihood of beingattacked), whilst, at the same time, increasing theprey’s ability to flee and/or to defend itself (Trexleret al., 1994). The most extensive datasets come fromcases in which prey occur in unusual abundance,allowing comparisons of trait distributions before vs.after the predation event (e.g. Quinn & Kinnison,1999 for bears eating salmon; Shine et al. 2001 forcrows eating gartersnakes).
One way to circumvent these problems is to studyan invasive species. First, they often occur at highdensities (Mack et al., 2000). Second, there are noethical issues associated with the death of largenumbers of individuals of such species, as long astreatment is humane. Third, the lack of contactbetween predator and prey over evolutionary timeeliminates the potential for long-term coevolution,and thus the interaction may be more intense thanwe would see in a stable coevolved system where mosttraits have reached evolutionary equilibrium (Endler,1986). We exploited these opportunities to examinethe interaction between an invasive anuran and itsinvertebrate predators in tropical Australia.
MATERIAL AND METHODSTHE STUDY SYSTEM
Native to South and Central America, cane toads(Bufo marinus, Linnaeus 1758) were introduced tonorth-eastern Queensland in 1935 and have sincespread through much of tropical and subtropical Aus-tralia (Lever, 2001; Urban et al., 2007). They reachedthe area of Darwin, in the wet–dry tropics of theNorthern Territory, in 2005 (Phillips et al., 2007). Westudied toads beside isolated waterbodies on the Ade-laide River floodplain, 60 km east of Darwin, in 2008.In this area, toads breed primarily in the late wetseason and early dry season (March through May),with most metamorphs emerging from April throughJune (M. Crossland, pers. comm.). Reflecting fallingwater levels at this time of year, combined with the
cane toad’s preference for spawning in waterbodieswith flat, bare, sun-exposed edges (Hagman & Shine,2006; Semeniuk, Lemckert & Shine, 2007), the youngtoads emerge onto expanses of drying mud at thepond margins (Cohen & Alford, 1993; Pizzatto &Shine, 2008). Desiccation risk precludes dispersal intothe surrounding landscape, such that young toadsaggregate near the water’s edge until wet-seasonrains facilitate dispersal (Child et al., 2008a; Child,Phillips & Shine, 2008b). The bare margins of theseponds are also foraging sites for bearded meat ants(Iridomyrmex reburrus; Shattuck, 1999), a medium-sized (to 10 mm body length) ant that consumes bothdead and live prey (Carrol & Janzen, 1973; Mobbset al., 1978). Unlike many vertebrates (e.g. Phillips &Shine, 2004; Letnic, Webb & Shine, 2008), these antscan tolerate toad toxins (Clerke & Williamson, 1992).
OBSERVATIONAL STUDY OF METAMORPH MORTALITY
Initially, we conducted field surveys to observe inter-actions between cane toads and meat ants in anunmanipulated environment. We surveyed each of sixwaterbodies (see Table 1 for detailed geographicallocations) at 08.00 h, 12.00 h and 16.00 h once permonth from March to June, as well as opportunisti-cally at other times. During these surveys, werecorded the number of ants and metamorphs within2 m of the water’s edge, as well as any ant attacksresulting in mortality of cane toad metamorphs. Todetermine whether mortality was size selective, wemeasured the snout–urostyle length (= SUL) of alldead metamorphs. On days that we saw more thanone metamorph being killed, we also collected arandom sample of live metamorphs to compare theirSULs with those of the victims (using a one-wayANOVA).
MANIPULATIVE TRIALS OF METAMORPH
VULNERABILITY: THE ROLES OF TOAD MORPHOLOGY
AND LOCOMOTOR PERFORMANCE, AND ANT DENSITY
Nonrandom mortality of metamorphs with respect tobody size (see above) might arise via several mecha-
Table 1. Geographical locations (depicted by latitude and longitude references) of waterbodies surveyed in experimentson the Adelaide River floodplain in tropical Australia. ‘Initial size’ shows each waterbody’s area at the beginning of thestudy
Waterbody Latitude Longitude Initial size (m2)
Farm Dam 12°38′24.80″S 131°18′57″E 900Lily 12°34′39″S 131°19′5″E 1700Ropehead 12°36′15.16″S 131°18′77″E 200One Tree 12°36′83″S 131°18′7.16″E 350Trev’s 12°36′38.32″S 131°17′51.56″E 500Slim’s Wallow 12°37′19.87″S 131°18′19.72″E 100
PREDATION ON ANURANS BY ANTS 739
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 99, 738–751

nisms: for example, smaller metamorphs might befound in habitats with higher ant densities, be recog-nized more readily as potential prey by ants, be lesswell defended chemically or be less able to escape ifseized. To identify causation, we need to conductexperiments. We tested the influence of metamorphlocomotor ability and body size, and ant densities, ontoad vulnerability to ant predation as follows. Wecollected 261 toad metamorphs from floodplain water-bodies, such that their body sizes spanned an SULrange of 9–22 mm (i.e. encompassing the range ofbody sizes seen in metamorph populations in thefield). We measured the mass, SUL and right tibialength of each of these animals, and then measuredtheir sprint speeds in the laboratory at 28 °C and 45%humidity.
At the beginning of the sprint speed trial, themetamorph was placed at the start of a plasticrunway (0.2 ¥ 0.2 ¥ 1 m) and encouraged to run bylightly tapping it on the urostyle. Each time themetamorph stopped hopping, it was tapped lightly.We recorded the time taken to reach the 0.33 m,0.66 m and 1.0 m marks on the runway; the fastesttime over 0.33 m was used to calculate the sprintspeed (cm s-1).
We then took these metamorphs into the field andplaced them individually in the centre of a circular0.5 m2 arena at the edge of a waterbody. We selectedsites to span a range of ant densities (simulating therange of natural levels observed) of 5–30 ants per0.5 m2, in increments of five (counted at the beginningof each trial). The arena was surrounded by a wire-mesh fence that was impenetrable to toads, but(because of its mesh size) did not impede ants. At thebeginning of each trial, we measured the temperatureand humidity; then, for the next 2 min, we scored: (1)the number of hops by the metamorph; (2) thenumber of ant encounters (defined as an ant � 5 mmfrom a toad, such that at least its antennae were incontact with the toad); (3) the number of attacks (i.e.cases in which the metamorph was bitten); (4) themetamorph’s response to attack; (5) whether themetamorph was captured or escaped; and (6) obser-vations of ant activity, such as swarming (large-scalerecruitment events) or prey-dropping behaviour.
On conclusion of the trial, we removed the meta-morph from the arena and returned it to the labora-tory to assess its injuries. If, during the trial, themetamorph was deemed unable to escape, we imme-diately terminated the trials and euthanased the toadfor ethical reasons. To assess the extent and locationof the injuries, we examined all toads under a dis-secting microscope and scored the position andnumber of lacerations attributable to ant attack. Wealso collected a sample of metamorphs that we wit-nessed escaping an attack, and compared their sub-
sequent mortality rates with those of a control sample(metamorphs not seen to have been attacked). Overthis 24 h period, the toads were kept under moist,warm conditions in the laboratory.
We used linear regression analyses to explore therelationship between continuous variables (e.g. bodysize vs. hops per minute), and logistic regression toanalyse categorical variables (e.g. body size vs.‘subdued or escaped’).
TESTING HYPOTHESES ABOUT CAUSAL MECHANISMS
IN ANT–TOAD INTERACTIONS
Size-dependent vulnerability to predation is consis-tent with multiple causal mechanisms; for example,body size might influence a metamorph’s microhabi-tat (and thus the probability of encountering an ant),its conspicuousness (and thus the probability ofattracting an ant’s attention) and/or its toxicity orlocomotor ability (and thus its ability to escape ifseized by an ant). To distinguish between thesehypotheses, we designed experiments to test theeffects of body size, toxicity and activity levels on atoad’s vulnerability to ant attack. Prior to runningthese experiments, we compared the rate of antattack directed to live vs. dead metamorphs to testwhether we could use dead metamorphs rather thanlive ones. In these preliminary tests, ants were givena choice between a live and a dead metamorph (of thesame size) in the arena described above. We observedthem for 2 min and counted the attacks on each.
To determine whether a metamorph’s body sizeinfluenced its probability of being attacked, we placeddead metamorphs of three sizes (SUL: ‘small’, 10 mm;‘medium’, 15 mm; ‘large’, 20 mm) at equal distancesfrom ant’s nests and counted the number of ants in a5 cm2 quadrat around each metamorph at intervals of1, 5, 10, 15, 20 and 30 min. We used a similar methodto investigate the potential role of toxicity, usingbeef-mince balls (0.15 g, 15 ¥ 10 ¥ 10 mm) with andwithout a coating of toad toxin as the stimulus. Thetoxin was derived by massaging the parotoid glandsof five juvenile toads, and the resulting material wassmeared onto the beef mince. To evaluate whether atoad’s rate of movement affected its probability ofbeing attacked, we attached a beef-mince meatball(0.15 g, 15 ¥ 10 ¥ 10 mm) to a 30 cm length of cotton,and used the other end of the cotton to ‘bounce’ themeatball around an enclosure (40 ¥ 30 ¥ 20 cm) con-taining 20 naïve meat ants. For each trial, wemanipulated movement rates, with each ‘hop’ movingthe ‘prey item’ 2 cm; the item was moved around a‘figure of eight’ circuit at either low (one bounce every3 s) or high (one bounce every 1 s) activity levels, inrandom order. During these trials we scored: (1) thenumber of ant encounters with the meatball; (2) the
740 G. WARD-FEAR ET AL.
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 99, 738–751

number of ant attacks; and (3) the number of antsthat pursued the prey item.
To analyse the effects of prey toxicity and size onant responses, we ran repeated-measure ANOVAsusing ‘prey’ type (e.g. toxic or not) as the factor, timeas the repeated measure and the numbers of ants asthe dependent variable. We analysed the activity ratedata using one-way ANOVAs on the rates of encoun-ter, attack and pursuit.
RESULTSOBSERVATIONAL STUDY OF METAMORPH MORTALITY
We compared the body sizes of 23 metamorphs killedby ants in the field with those of 23 live metamorphs.The analysis of these data revealed that smallermetamorphs were more likely to be killed(F1,44 = 44.387, P < 0.0001; Fig. 1). Hence, we focusedon prey body size and predator density for our experi-mental manipulations. The temperature and humid-ity during our field trials were not correlatedsignificantly with any of the variables concerningtoad–ant interactions, and so (for simplicity) the
results of the analyses below do not incorporate theweather variables.
MANIPULATIVE TRIALS OF METAMORPH
VULNERABILITY: THE ROLES OF TOAD MORPHOLOGY
AND LOCOMOTOR PERFORMANCE, AND ANT DENSITY
The speed of a metamorph toad was enhancedsignificantly by both body size [SUL: F2,258 = 218.492,P < 0.0001; Fig. 2: speed (cm s-1) = -16.53045 +2.2741575 (SUL), r2 = 0.65] and relative tibialength [SUL: F2,258 = 4.4, P = 0.037; speed (cm s-1) =0.8646574 - 0.2812185 (tibia residual), r2 = 0.02].However, neither locomotor speed nor relative tibialength were correlated significantly with any othervariable measured (all P > 0.05, after body size effectswere included), and so are not considered furtherbelow.
Larger metamorphs exhibited faster movement(Table 2; Fig. 3A), but a toad’s rate of movementwas not influenced significantly by ant density(F1,157 = 3.34, P = 0.07), or by the interaction betweenbody size and ant density (F1,157 = 2.25, P = 0.013).Whether or not a metamorph was attacked wasdependent on its rate of movement (likelihood ratiotest from logistic regression on hops per minute:c2 = 11.74, d.f. = 1, P = 0.0006; Fig. 3B) and antdensity (Table 2): attacks were most likely athigher ant densities, and directed to faster movingmetamorphs.
The probability that an encounter between an antand a toad would result in an attack was negativelycorrelated with toad body size and positively corre-
Figure 1. Body size distributions of metamorph canetoads randomly sampled around three tropical waterbod-ies (A), compared with a concurrently collected sample ofmetamorphs killed by predatory ants (B).
Figure 2. The relationship between the body size of ametamorph cane toad and its maximum speed (as indi-cated by the fastest of three times recorded as the animalran down a 33 cm section of raceway) in the laboratory.
PREDATION ON ANURANS BY ANTS 741
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 99, 738–751

lated with ant density (Table 2; Fig. 4A, B, respec-tively). That is, an ant that encountered a toad wasmore likely to attack if the toad was small and thedensity of foraging ants was high. If a toad wasattacked, its probability of escaping was increased bya larger body size and decreased by a higher antdensity (Table 2; Fig. 4C, D, respectively.)
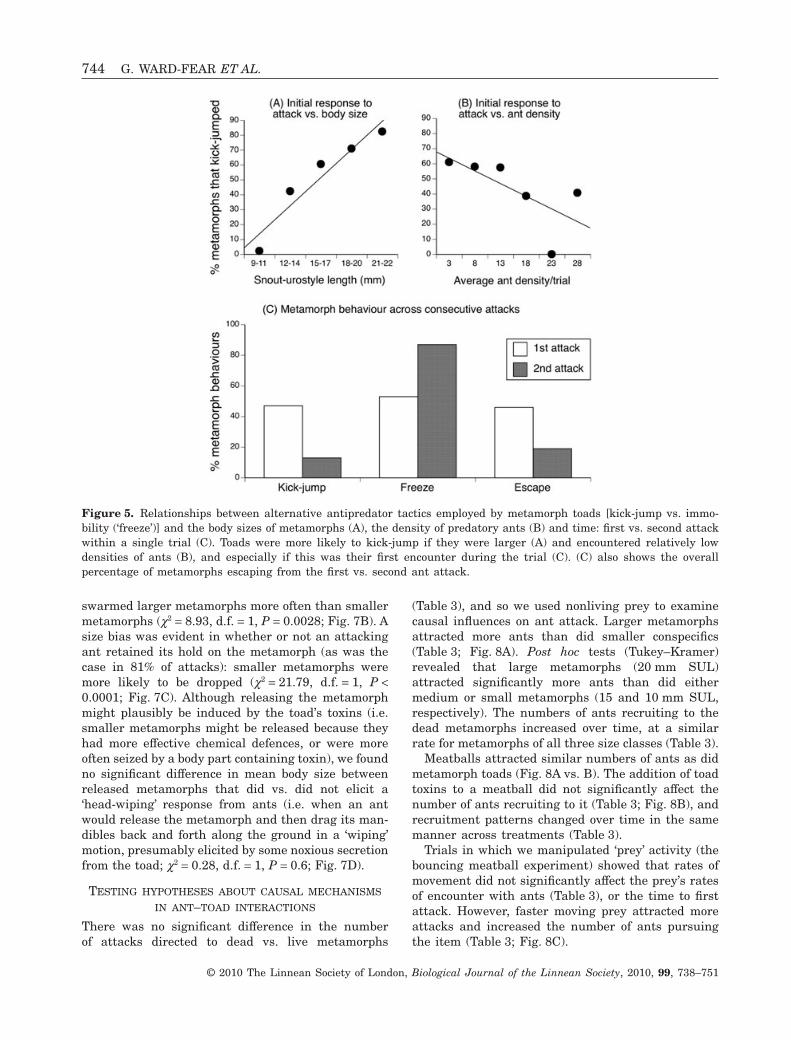
The initial response of a metamorph toad to attackwas dependent on its body size and the predatordensity. Larger metamorphs were more likely to ‘kick-jump’ (i.e. kick and jump at the same time) whenattacked (Table 2; Fig. 5A) and were less likely toexhibit the alternative response of immobility (‘freez-ing’). Regardless of body size, all metamorphs wereless likely to kick-jump and more likely to ‘freeze’when attacked in situations with high ant densities(Table 2; Fig. 5B). A toad’s propensity to kick-jumpalso decreased between the first and second attackswithin a trial (c2 = 14.47, d.f. = 1, P = 0.0001; Fig. 5C).Toads that ‘froze’ rather than kick-jumped were lesslikely to escape (c2 = 4.85, d.f. = 1, P = 0.027).
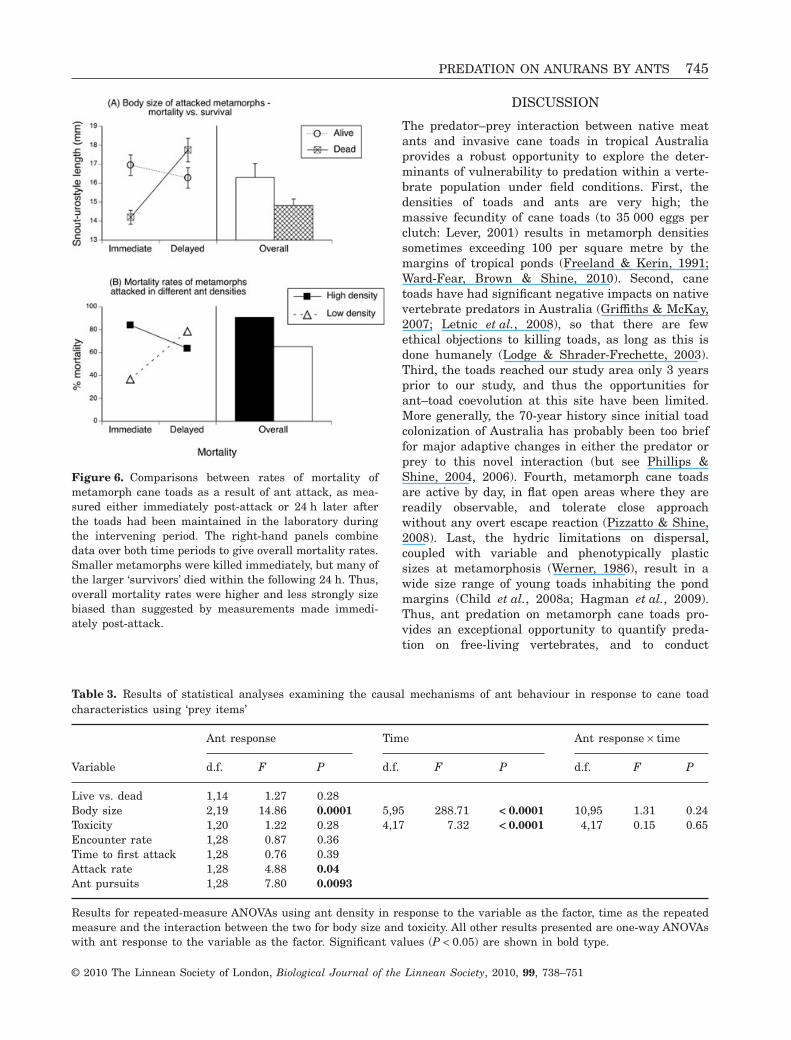
Immediate mortality in the arena trials was sizebiased, with higher mortality rates in smaller meta-morphs (F1,161 = 17.93, P < 0.0001; Fig. 6A). However,this size bias disappeared when analyses were basedon survival rates in the subsequent 24 h (i.e. toadsthat had been attacked but had apparently escaped).The ‘escapee’ metamorphs that died were larger, onaverage, than those that survived through the sameperiod – albeit, not significantly so (F1,151 = 3.15,P = 0.08). Combining our data on immediate mortal-ity with those on subsequent mortality, we are leftwith a marginally nonsignificant trend for smallermetamorphs to be more likely to die as a result of antattack (F1,161 = 3.28, P = 0.07).
Predator density also affected mortality rates: 84%of encounters between toads and ants at high antdensities (> 13 ants per 0.5 m2) led to attack, vs. 51%of encounters at low predator densities (< 8 antsper 0.5 m2; comparing these two rates of attack:c2 = 43.76, d.f. = 1, P < 0.0001). Attack by an antresulted in ‘immediate’ mortality for 84% of meta-morphs at high predator densities, and 37% of meta-morphs at low ant densities (c2 = 37.65, d.f. = 1,P < 0.0001; Fig. 6B). Survival rates of ‘escapee’ meta-morphs 24 h post-attack were not affected by antdensity (64% at high density vs. 79% at low density:c2 = 0.92, d.f. = 1, P = 0.34). Combining the data forthese two periods, the mortality rate of metamorphsattacked at high predator densities was 90.5%, com-pared with 65% for metamorphs at low predatordensities. The overall rate of mortality was higherfor metamorphs attacked at higher ant densities(c2 = 4.18, d.f. = 1, P = 0.04), although the mortalityrates at low ant densities nearly doubled once delayedmortality had been accounted for. Hence, the rate ofT
able
2.R
esu
lts
ofst
atis
tica
lan
alys
esex
amin
ing
the
outc
omes
ofen
cou
nte
rsbe
twee
npr
edat
ory
ants
and
met
amor
phca
ne
toad
s
Var
iabl
e
Bod
ysi
ze(S
UL
)A
nt
den
sity
Reg
ress
ion
equ
atio
nr2
d.f.
For
c2P
d.f.
For
c2P
Rat
eof
mov
emen
t(h
ops/
min
)1,
157
20.0
5<
0.00
011,
157
3.34
0.07
hop
s/m
in=
2.21
2142
5+
0.68
1828
8(S
UL
)0.
17A
ttac
ks/e
nco
un
ters
1,24
18.
160.
0047
1,24
19.
860.
0007
atta
cks/
enco
un
ters
=0.
626
-0.
17(S
UL
)0.
03at
tack
s/en
cou
nte
rs=
0.23
8+
0.01
(an
tde
nsi
ty)
0.05
Was
met
amor
phat
tack
ed?
10.
030.
851
10.4
40.
0012
Esc
aped
atta
ck?
111
.22
0.00
081
12.8
80.
0003
Init
ial
reac
tion
toat
tack
(kic
k-ju
mp
vs.
free
ze)
130
.96
<0.
0001
14.
630.
032
An
alys
esu
sin
gli
nea
rre
gres
sion
betw
een
body
size
,an
tde
nsi
tyan
dco
nti
nu
ous
vari
able
s(t
optw
oro
ws)
.L
inea
req
uat
ion
sal
sopr
esen
ted.
Res
ult
sof
logi
stic
regr
essi
onw
ith
body
size
and
ant
den
sity
agai
nst
cate
gori
cal
vari
able
sgi
vin
gch
i-sq
uar
edva
lues
(bot
tom
thre
ero
ws)
.S
ign
ifica
nt
valu
es(P
<0.
05)
are
show
nin
bold
type
.S
UL
,sn
out–
uro
styl
ele
ngt
h.
742 G. WARD-FEAR ET AL.
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 99, 738–751

toad mortality as a result of ant attack is greatlyunderestimated if conclusions are based on immedi-ate outcomes, and this bias is greatest for observa-tions made at low ant densities.
Our data on ant behaviour clarify the mechanismslinking toad mortality to ant densities. At higherdensities, more ants swarmed onto a recently seizedprey item (c2 = 11.23, d.f. = 1, P = 0.0008; Fig. 7A) and
Figure 3. Rates of movement (hops completed per minute) by metamorph cane toads in the field, prior to experiencingan attack by predatory ants. Larger toads moved more quickly (A), and faster movement was correlated with a higherrate of ant attack (B). Both (A) and (B) show mean values and associated standard errors. In (A), the snout to urostylelength (SUL) has been categorized into increments of 3 mm for clarity in the graphical representation; however, statisticalanalyses (see text) used continuous data for both variables (hops per minute and SUL).
Figure 4. The effects of cane toad body size and ant density on rates at which toads were attacked by meat ants, andon the proportion of toads that escaped these attacks. (A) and (B) show that the percentage of ant encounters that ledto attacks decreased with toad body size and increased with ant density. Once attacked, the percentage of metamorphsthat escaped from the ants increased with body size (C) and decreased with ant density (D). The snout to urostyle lengthhas been categorized into increments of 3 mm for clarity in the graphical representation, but statistical analyses werebased on continuous data. Ant density was scored (in increments of five) at the start of each 2 min trial.
PREDATION ON ANURANS BY ANTS 743
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 99, 738–751
swarmed larger metamorphs more often than smallermetamorphs (c2 = 8.93, d.f. = 1, P = 0.0028; Fig. 7B). Asize bias was evident in whether or not an attackingant retained its hold on the metamorph (as was thecase in 81% of attacks): smaller metamorphs weremore likely to be dropped (c2 = 21.79, d.f. = 1, P <0.0001; Fig. 7C). Although releasing the metamorphmight plausibly be induced by the toad’s toxins (i.e.smaller metamorphs might be released because theyhad more effective chemical defences, or were moreoften seized by a body part containing toxin), we foundno significant difference in mean body size betweenreleased metamorphs that did vs. did not elicit a‘head-wiping’ response from ants (i.e. when an antwould release the metamorph and then drag its man-dibles back and forth along the ground in a ‘wiping’motion, presumably elicited by some noxious secretionfrom the toad; c2 = 0.28, d.f. = 1, P = 0.6; Fig. 7D).
TESTING HYPOTHESES ABOUT CAUSAL MECHANISMS
IN ANT–TOAD INTERACTIONS
There was no significant difference in the numberof attacks directed to dead vs. live metamorphs
(Table 3), and so we used nonliving prey to examinecausal influences on ant attack. Larger metamorphsattracted more ants than did smaller conspecifics(Table 3; Fig. 8A). Post hoc tests (Tukey–Kramer)revealed that large metamorphs (20 mm SUL)attracted significantly more ants than did eithermedium or small metamorphs (15 and 10 mm SUL,respectively). The numbers of ants recruiting to thedead metamorphs increased over time, at a similarrate for metamorphs of all three size classes (Table 3).
Meatballs attracted similar numbers of ants as didmetamorph toads (Fig. 8A vs. B). The addition of toadtoxins to a meatball did not significantly affect thenumber of ants recruiting to it (Table 3; Fig. 8B), andrecruitment patterns changed over time in the samemanner across treatments (Table 3).
Trials in which we manipulated ‘prey’ activity (thebouncing meatball experiment) showed that rates ofmovement did not significantly affect the prey’s ratesof encounter with ants (Table 3), or the time to firstattack. However, faster moving prey attracted moreattacks and increased the number of ants pursuingthe item (Table 3; Fig. 8C).
Figure 5. Relationships between alternative antipredator tactics employed by metamorph toads [kick-jump vs. immo-bility (‘freeze’)] and the body sizes of metamorphs (A), the density of predatory ants (B) and time: first vs. second attackwithin a single trial (C). Toads were more likely to kick-jump if they were larger (A) and encountered relatively lowdensities of ants (B), and especially if this was their first encounter during the trial (C). (C) also shows the overallpercentage of metamorphs escaping from the first vs. second ant attack.
744 G. WARD-FEAR ET AL.
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 99, 738–751
DISCUSSION
The predator–prey interaction between native meatants and invasive cane toads in tropical Australiaprovides a robust opportunity to explore the deter-minants of vulnerability to predation within a verte-brate population under field conditions. First, thedensities of toads and ants are very high; themassive fecundity of cane toads (to 35 000 eggs perclutch: Lever, 2001) results in metamorph densitiessometimes exceeding 100 per square metre by themargins of tropical ponds (Freeland & Kerin, 1991;Ward-Fear, Brown & Shine, 2010). Second, canetoads have had significant negative impacts on nativevertebrate predators in Australia (Griffiths & McKay,2007; Letnic et al., 2008), so that there are fewethical objections to killing toads, as long as this isdone humanely (Lodge & Shrader-Frechette, 2003).Third, the toads reached our study area only 3 yearsprior to our study, and thus the opportunities forant–toad coevolution at this site have been limited.More generally, the 70-year history since initial toadcolonization of Australia has probably been too brieffor major adaptive changes in either the predator orprey to this novel interaction (but see Phillips &Shine, 2004, 2006). Fourth, metamorph cane toadsare active by day, in flat open areas where they arereadily observable, and tolerate close approachwithout any overt escape reaction (Pizzatto & Shine,2008). Last, the hydric limitations on dispersal,coupled with variable and phenotypically plasticsizes at metamorphosis (Werner, 1986), result in awide size range of young toads inhabiting the pondmargins (Child et al., 2008a; Hagman et al., 2009).Thus, ant predation on metamorph cane toads pro-vides an exceptional opportunity to quantify preda-tion on free-living vertebrates, and to conduct
Figure 6. Comparisons between rates of mortality ofmetamorph cane toads as a result of ant attack, as mea-sured either immediately post-attack or 24 h later afterthe toads had been maintained in the laboratory duringthe intervening period. The right-hand panels combinedata over both time periods to give overall mortality rates.Smaller metamorphs were killed immediately, but many ofthe larger ‘survivors’ died within the following 24 h. Thus,overall mortality rates were higher and less strongly sizebiased than suggested by measurements made immedi-ately post-attack.
Table 3. Results of statistical analyses examining the causal mechanisms of ant behaviour in response to cane toadcharacteristics using ‘prey items’
Variable
Ant response Time Ant response ¥ time
d.f. F P d.f. F P d.f. F P
Live vs. dead 1,14 1.27 0.28Body size 2,19 14.86 0.0001 5,95 288.71 < 0.0001 10,95 1.31 0.24Toxicity 1,20 1.22 0.28 4,17 7.32 < 0.0001 4,17 0.15 0.65Encounter rate 1,28 0.87 0.36Time to first attack 1,28 0.76 0.39Attack rate 1,28 4.88 0.04Ant pursuits 1,28 7.80 0.0093
Results for repeated-measure ANOVAs using ant density in response to the variable as the factor, time as the repeatedmeasure and the interaction between the two for body size and toxicity. All other results presented are one-way ANOVAswith ant response to the variable as the factor. Significant values (P < 0.05) are shown in bold type.
PREDATION ON ANURANS BY ANTS 745
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 99, 738–751

experimental tests of hypotheses about the determi-nants of prey vulnerability.
Importantly, the system facilitates detailed mea-surement and experimental manipulation of relevantvariables, with large sample sizes, in ways that wouldbe impossible to achieve with predator–prey interac-tions in most wild vertebrates. Our data confirm thata cane toad’s vulnerability to predation is increased athigher densities of predators, is not affected by thetoad’s locomotor ability (at least over the range exhib-ited by metamorph toads within this population), butis influenced by body size in complex ways. Forexample, a toad’s size influences the number of antsthat it attracts, both directly and as a consequence offaster rates of movements by larger metamorphs.Overall, body size affects toad vulnerability less whenpredator densities are higher, and also affects mortal-ity rates less than would be suggested by observationsmade at the time of encounter. In total, our observa-tions and experiments identify a complex series oflinks that affect prey vulnerability in this system.
One surprising result was the magnitude of delayedmortality: we recorded 69% mortality at the time ofthe attack, increasing to 83% after 24 h. The size biasin toad vulnerability also weakened over this period.
Similar delayed effects of stressful events (includingpredator attacks) may be common; for example, theclassic study of winterkill in New York sparrows byHerman Bumpus (1899) reported that much of themortality occurred after the extreme weather event.In the case of predator attacks, apparently minorinjuries sustained during the encounter may laterprove lethal. For example, deer that sustain lacera-tions during attacks by Komodo dragons often die ofsepticaemia a few days later (Auffenberg, 1981), andpasserine birds struck but not held by pit-vipers oftendie of envenomation within a few minutes (Shineet al., 2002). In addition, some salmon species expe-rience high rates of delayed mortality attributable toinjuries sustained in unsuccessful attacks by harbourseals (Chapman et al., 1991). As meat ants lack asting (Shattuck, 1999), metamorph toads are mostlikely to die from physical injuries, from toxic secre-tions delivered by the ant’s mandibles or the associ-ated shock. The most common site for ants to seizetoads was around the hind limbs, followed by thehead. Serious injuries could be sustained in less than5 s, and we found no mortality bias with injury loca-tion. Thus, the assumption that a toad is unscathed ifit escapes from a brief ant attack is often wrong.
Figure 7. Behavioural response of ants to metamorph cane toads. Ants were more likely to swarm over a recently seizedtoad if ant densities were high (A) and if the prey item was large (B). Smaller toads were more likely to be released afterthe initial seizure than were larger toads (C). Whether or not the ant wiped its head after releasing a toad was unaffectedby toad size (D).
746 G. WARD-FEAR ET AL.
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 99, 738–751

Studies of prey vulnerability in the field shouldmonitor longer term mortality rates following appar-ently unsuccessful predation events.
A metamorph toad’s body size influenced its vulner-ability in complex ways, and interacted with antdensity and the timeframe over which mortality was
evaluated. Our initial field surveys suggested thatsmaller metamorphs were more vulnerable to antattack. Experimental trials in which we standardizedencounter conditions (including ant densities) revealedthe same pattern of size-dependent mortality. Thus,these data falsify one possible explanation for size-dependent mortality – that the bias is driven byontogenetic or size-related shifts in metamorph toadsthat modify the times and/or places they encounterants, and/or the numbers of ants encountered in thesesites. Ontogenetic shifts in habitat use are a commonantipredator strategy that trades off physiological con-straints with predation risk. For example, many fishspecies exhibit ontogenetic shifts from poor (e.g. densealgal clumps) to higher quality foraging habitat. Thisshift correlates with their ontogenetic increase inbody size, and thus lower vulnerability to predation(Dahlgren & Eggleston, 2000). Similar ontogeneticshifts occur in cane toads (Freeland & Kerin, 1991;Cohen & Alford, 1993), but cannot explain the higherpredation rates on smaller metamorph toads. None-theless, such ecological shifts may exacerbate orreduce the magnitude of the ant predation size bias.
Because we measured each toad’s locomotor perfor-mance prior to ant encounter trials, we can alsofalsify another plausible hypothesis: that larger meta-morphs escape predation because of enhanced loco-motor abilities. An extensive literature assumes thatlocomotor performance is under strong selectionbecause of its role in escaping predators (e.g. Hale,1999; Domenici, 2001). Although some field studieshave documented higher survival rates in faster indi-viduals within a cohort, the causal basis of this biasremains obscure (Endler, 1986). Undoubtedly, sometypes of predator–prey interaction place a selectivepremium on prey mobility (e.g. gazelles fleeing chee-tahs), and the same may even be true for cane toadsconfronted with types of predator other than meatants, or encountering predators at a different stage inthe life history. However, some predator–prey rela-tionships confer no adaptive advantage for faster loco-motor performance, because the predator is fasterthan even the fastest prey item (Schwarzkopf &Shine, 1992). Thus, although intuition suggests thatlocomotor speed will affect vulnerability in predator–prey encounters, our data show no such effect. Thisnull result partially reflects the limited range of loco-motor abilities within the cohort of metamorph toadstested. Comparisons with Australian native frogssuggest that the much greater athletic abilities ofsome of those taxa do indeed reduce their vulnerabil-ity to attack by meat ants (Ward-Fear, 2008; Ward-Fear et al., 2009).
In contrast with these null results, metamorphbody size significantly affected other aspects of theant vs. toad encounter. In our trials with live meta-
Figure 8. Effects of prey body size and activity level onthe predatory behaviour of meat ants. When offered achoice of three sizes of dead metamorph toad, the antsrecruited more rapidly to larger toads than to smallerconspecifics (A). Whether or not a meatball was coatedwith toad toxin did not affect the number of ants that wereattracted to it (B). A meatball that was moved rapidly (one‘bounce’ per second) attracted more attacks and pursuitsthan did a meatball that was moved more slowly (one‘bounce’ per 3 s, C).
PREDATION ON ANURANS BY ANTS 747
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 99, 738–751

morphs, smaller toads were more likely to be attacked(Fig. 4A). However, trials with dead metamorphs pro-vided the opposite result: ants recruited faster anddisproportionately more towards larger metamorphs,presumably because a larger object provides morevisual and/or chemical cues to its presence. So whydid smaller metamorphs attract higher rates of antattack? Plausibly, a metamorph’s size may influence(or correlate with) its behaviour or physiology (type oreffectiveness of toxic defence: Hayes et al., 2009) inways that affect ant responses. A smaller body sizeincreased the probability that an ant would release itafter attack, but (based on similar rates of ant ‘head-wiping’ following such release), size-specific toxicitydid not seem to be the major determinant of thisbehaviour. In addition, movement patterns differedbetween smaller and larger metamorphs, but in theopposite direction to that predicted: larger metamor-phs moved more rapidly, thus attracting more ants. Inthe absence of all other stimuli, our ‘bouncing meat-ball’ experiments confirmed this pattern – higheractivity levels (faster movement) attracted more pur-suits and attacks than the lower activity levels(slower movement) analogous to that of smallermetamorphs.
Why, then, did smaller metamorphs sustain moreattacks and hence constitute a disproportionatenumber of ant victims (at least if mortality wasscored at the time of the encounter)? This paradoxcan be clarified by observations of the antipredatorresponses of the metamorphs. A metamorph’s sizeinfluenced its vulnerability not just because of theprobability that it would be attacked, but alsobecause it affected the toad’s probability of escape.Larger metamorphs encountered more ants but weremore likely to respond in a way that prevented theinteraction from becoming an attack. This responsealso allowed them to escape from an attack, thusincreasing their chance of survival, at least in theshort term (Fig. 4C). The ability of larger metamor-phs to escape appears to reflect their physicalstrength (ability to kick-jump away from the ant) aswell as their reliance on this effective tactic. Smallertoads were more likely to remain immobile (‘freeze’)if seized, a tactic that was commonly fatal (81%mortality of immobile metamorphs). Thus, althoughlarger toads showed higher encounter rates withants because of their faster movement, they alsomoved through high-risk areas more rapidly, andwere more likely to escape if seized.
Ontogenetic shifts in antipredator tactics are wide-spread, and may reflect correlated ontogenetic shiftsin the effectiveness of alternative responses (Lima &Dill, 1990). For example, juvenile snakes of somespecies rely on hissing and striking more so than doconspecific adults (Herzog, Bowers & Burghardt,
1992), and the duration of death feigning in somebeetles is determined by body size (Hozumi &Miyatake, 2005). Similarly, ground squirrels showontogenetic shifts in their behavioural response towarning calls made by conspecifics. Naïve juvenileswill ‘freeze’ in response to every adult call, but thisresponse increases in selectivity as soon as the juve-nile individuals are old enough to disperse, regardlessof predator experience (Mateo & Holmes, 1999).
Given the common futility of immobility as adefence against ant attack (only 19% of toads thatadopted this tactic were released by the ants), whydid the small metamorph toads use this tactic sofrequently? The answer may lie in the effectivenessagainst other predators; for example, many verte-brates cannot tolerate toad toxins and thus may ceaseattacking as soon as they detect the toad’s toxins(Crossland, 2001). Refraining from struggling mightreduce the danger inflicted by the predator. In addi-tion, small metamorphs show such limited strengthand speed that attempts at escape via rapid locomo-tion are unlikely to be successful. Finally, immobilityand death feigning are widespread antipredatorresponses in a diverse array of animal species (e.g.birds, Rovee et al., 1976; amphibians, Williams et al.,2000; reptiles, Gehlbach, 1970), and may facilitatesurvival by eliminating the stimuli that encouragefurther attack (Gallup et al., 1971). In keeping withan adaptive interpretation of this behaviour, studiesin insects have reported significant heritability (andthus an opportunity for adaptive shifts in response toselection) in the duration of death feigning behaviour(Prohammer & Wade, 1981; Honma, Oku & Nishida,2006). Immobility in response to attack occurs inseveral amphibian species with toxic skin secretions(including many small bufonids), and may enhancethe survival value of their noxious skin secretions(Brodie, Formanowicz & Brodie, 1978). Such a systemis less likely to function effectively for an invasivespecies encountering a novel predator that may not besusceptible to (or not react to) its toxins, as is appar-ently the case for Bufo marinus encountering Austra-lian meat ants.
Interestingly, metamorph toads changed their anti-predator responses between the first and secondattacks they encountered during a trial (Fig. 5C). Theability to detect, and flexibly respond to, variation inthe type or magnitude of threat posed by encounter-ing a predator often may enhance individual fitness(Lima & Bednekoff, 1999). Threat levels can changeon a seasonal or diel basis (Werner, 1986), and evenminute to minute during an encounter (Dill & Gillett,1991). Metamorphs were more likely to freeze(regardless of body size) during a second attack, pos-sibly reflecting a tactic that enhances fitness (at leastin the toads’ ancestral range) under situations in
748 G. WARD-FEAR ET AL.
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 99, 738–751

which an initial attempt at escape has been unsuc-cessful. Only about one-half of the ants that releasedmetamorphs exhibited head-wiping behaviour afterdoing so, suggesting that toad toxins may not havebeen the only factor in initiating release (or, indeed,may not have been a factor at all). However, althoughthe immobility response was associated with a lowerprobability of toad survival than the kick-jumpresponse, causation is difficult to infer (i.e. perhapsthe toads that kick-jumped were also more likely tosurvive for other reasons).
The density of predatory ants encountered by ametamorph toad also influenced its probability ofattack and escape. These effects are not simple func-tions of cumulative risk, but reflect more complexchanges in the behaviour of both prey and predator.Larger numbers of ants induced metamorph toads(even larger individuals) to ‘freeze’ rather than kick-jump, and induced ants to recruit in large numbers toany prey item that was seized by another ant. Thetoads’ ineffective behavioural response may reflect thelack of coevolution between cane toads and Australianpredatory ants. The end result was a high risk ofmortality to toads and, importantly, the magnitude ofthis risk was underestimated by the field data. Whenwe maintained ‘escapee’ toads in the laboratory, mostdied over the next 24 h. This delayed mortality washigher in toads from the low ant density treatments,perhaps because of the initially higher mortality rates(and thus lower proportion of survivors) from the highant density encounters.
In summary, invasive cane toads in tropical Aus-tralia experience high rates of mortality as a resultof predation by native meat ants. Several aspects ofthe study system facilitated our attempts to clarifythe proximate determinants of toad vulnerability.Although several of these patterns are straightfor-ward and intuitive, our work also revealed unex-pected complexities (e.g. effects of predator density onbehaviour of both predator and prey; changes throughtime post-attack in apparent mortality rates and inthe size selectivity of mortality). Many of theseresults may be unique to the specific system studied,but others (such as links between prey body size,antipredator responses and vulnerability) may bewidespread. For example, antipredator responses ofmany aquatic invertebrates depend on body size. Inlobster populations, the smallest individuals arehighly cryptic in the presence of predators, whereaslarge lobsters are aggressive (Wahle, 1992). Thus,smaller individuals trade off their foraging abilitieswith predation risk, whereas larger individualsutilize better quality resources but are involved inmore aggressive interactions with conspecifics andpredators. Conversely, herons and other wading birdsfeed preferentially on larger individuals of sailfin
mollies (Trexler et al., 1994). As size at maturation isgenetically determined to some extent, herons effec-tively exert selection for a smaller body size on thepopulation (Snelson, 1984).
Other cases of size-biased predation have clear con-sequences at a population level. Some insectivorousbats exert sex-biased predation on moths, as a resultof taking particular size classes (Acharya, 1995).Even closely related predators may differ in the lifehistory stages they target – for example, man-eatinglions often take adult humans, whereas man-eatingleopards concentrate on children (presumably reflect-ing the smaller size of the predator: Peterhans &Gnoske, 2001). In conclusion, interactions betweenpredators and their prey influence animal behaviourand population dynamics in a complex manner.Manipulative studies utilizing ‘model systems’ thatenable the collection of large datasets can help toclarify the underlying determinants of prey vulner-ability and predator behaviour in the wild.
ACKNOWLEDGEMENTS
We thank B. Phillips for advice on experimentaldesign and analysis, and M. Bruny for support.M. Crossland and L. Pizzatto helped with the hus-bandry of metamorphs, and M. Elphick with manu-script preparation. The study was funded by theAustralian Research Council and all procedureswere approved by the University of Sydney AnimalEthics Committee.
REFERENCES
Acharya L. 1995. Sex-biased predation on moths by insec-tivorous bats. Animal Behaviour 49: 1461–1468.
Auffenberg W. 1981. The behavioral ecology of the Komodomonitor. Gainesville, FL: University Press of Florida.
Brodie ED Jr, Formanowicz DR Jr, Brodie ED III. 1978.The development of noxiousness of Bufo americanus tad-poles to aquatic predators. Herpetologica 34: 302–307.
Bumpus H. 1899. The elimination of the unfit as illustratedby the introduced sparrow Passer domesticus. Marine Bio-logical Laboratory, Woods Hole, Massachusetts. Biology Lec-tures 1898: 209–228.
Carrol CR, Janzen DH. 1973. Ecology of foraging ants.Annual Review of Ecology and Systematics 13: 231–256.
Chapman DA, Giorgi M, Hill A, Maule S, McCutcheon D,Park W, Platts K, Pratt J, Scab L, Seeb A, Utter F.1991. Status of Snake River Chinook salmon. Portland, OR:Pacific Northwest Utilities Conference Committee.
Child T, Phillips BL, Brown GP, Shine R. 2008a. Thespatial ecology of cane toads (Bufo marinus) in tropicalAustralia: why do metamorphs stay near the water? AustralEcology 33: 630–640.
PREDATION ON ANURANS BY ANTS 749
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 99, 738–751

Child T, Phillips BL, Shine R. 2008b. Abiotic and bioticinfluences on the dispersal behaviour of metamorph canetoads (Bufo marinus) in tropical Australia. Journal ofExperimental Zoology 390A: 215–224.
Clerke RB, Williamson I. 1992. A note on the predation ofBufo marinus juveniles by the ant Iridomyrmex purpureus.Australian Zoologist 28: 64–67.
Cohen MP, Alford RA. 1993. Growth, survival, and activitypatterns of recently metamorphosed Bufo marinus. WildlifeResearch 20: 1–13.
Crossland MR. 2001. Ability of predatory native Australianfishes to learn to avoid toxic larvae of the introduced toad(Bufo marinus). Journal of Fish Biology 59: 319–329.
Dahlgren C, Eggleston D. 2000. Ecological processes under-lying ontogenetic habitat shifts in a coral reef fish. Ecology81: 2227–2240.
Dill LM, Gillett JF. 1991. The economic logic of barnacleBalanus glandula (Darwin) hiding behavior. Journal ofExperimental Marine Biology and Ecology 153: 115–127.
Domenici P. 2001. The scaling of locomotor performance inpredator–prey encounters: from fish to killer whales. Com-parative Biochemistry and Physiology 131A: 169–182.
Endler JA. 1986. Natural selection in the wild. Princeton,NJ: Princeton University Press.
Fox S. 1975. Natural selection on morphological phenotypesof the lizard Uta stansburiana. Evolution 29: 95–107.
Freeland WJ, Kerin SH. 1991. Ontogenetic alteration ofactivity and habitat selection in Bufo marinus. WildlifeResearch 18: 431–443.
Gallup GC, Nash RF, Donegan NH, McClure MK. 1971.The immobility response: a predator-induced reaction inchickens. Psychological Record 21: 513–519.
Gehlbach FR. 1970. Death-feigning and erratic behaviour inleptotyphlopid, colubrid and elapid snakes. Herpetologica26: 24–34.
Godin J-G. 1990. Diet selection under the risk of predation.In: Hughes RN, ed. Behavioural mechanisms of food selec-tion. New York: Springer, 739–769.
Griffiths AD, McKay JL. 2007. Cane toads reduce the abun-dance and site occupancy of Merten’s water monitor(Varanus mertensi). Wildlife Research 34: 609–615.
Hagman M, Hayes RA, Capon RJ, Shine R. 2009. Alarmcues experienced by cane toad tadpoles affect post-metamorphic morphology and chemical defences. Func-tional Ecology 23: 126–132.
Hagman M, Shine R. 2006. Spawning site selection by feralcane toads (Bufo marinus) at an invasion front in tropicalAustralia. Austral Ecology 31: 551–558.
Hale ME. 1999. Locomotor mechanics during early lifehistory: effects of size and ontogeny on fast-start perfor-mance of salmonid fishes. Journal of Experimental Biology202: 1465–1479.
Hayes RA, Crossland MR, Hagman M, Capon RJ, ShineR. 2009. Ontogenetic variation in the chemical defenses ofcane toads (Bufo marinus): toxin profiles and effects onpredators. Journal of Chemical Ecology 35: 391–399.
Herzog HA, Bowers BA, Burghardt GM. 1992.Development of antipredator responses in snakes: V. Species
differences in ontogenetic trajectories. Developmental Psy-chobiology 25: 199–211.
Holomuzuki JR, Short TM. 1990. Ontogenetic shifts inhabitat use and activity in a stream dwelling isopod. Holo-arctic Ecology 13: 300–307.
Honma A, Oku S, Nishida T. 2006. Adaptive significance ofdeath feigning posture as a specialized inducible defenceagainst gape-limited predators. Proceedings of the RoyalSociety Series B 273: 1631–1636.
Hozumi N, Miyatake T. 2005. Body-size dependent differ-ence in death-feigning behaviour of adult Callosobruchuschinensis. Journal of Insect Behaviour 18: 557–566.
Letnic M, Webb JK, Shine R. 2008. Invasive cane toads(Bufo marinus) cause mass mortality of freshwater croco-diles (Crocodylus johnstoni) in tropical Australia. BiologicalConservation 141: 1773–1782.
Lever C. 2001. The cane toad. The history and ecology of asuccessful colonist. Otley, West Yorkshire: Westbury Aca-demic and Scientific Publishing.
Lima SL, Bednekoff PA. 1999. Temporal variation in dangerdrives antipredator behavior: the predation risk allocationhypothesis. American Naturalist 153: 649–659.
Lima SL, Dill LM. 1990. Behavioral decisions made underthe risk of predation: a review and prospectus. CanadianJournal of Zoology 68: 619–640.
Lodge DM, Shrader-Frechette K. 2003. Nonindigenousspecies: ecological explanation, environmental ethics andpublic policy. Conservation Biology 17: 31–37.
Mack RN, Simberloff D, Lonsdale WM, Evans H, CloutM, Bazzaz F. 2000. Biotic invasions: causes, epidemiology,global consequences and control. Ecological Applications 10:689–710.
Mateo JM, Holmes WG. 1999. Plasticity of alarm-callresponse development in Belding’s ground squirrels (Sper-mophilus beldingi, Sciuridae). Ethology 105: 193–206.
Mobbs CJ, Tedder G, Wade AM, Williams S. 1978. A noteon food and foraging in relation to temperature in the meatant Iridomyrmex purpureus form viridiaeneus. Journal ofthe Australian Entomological Society 17: 193–197.
Peterhans JC, Gnoske TP. 2001. The science of ‘man-eating’ among lions Panthera leo with a reconstruction ofthe natural history of the ‘man-eaters of Tsavo’. Journal ofEast African Natural History 90: 1–40.
Phillips BL, Brown GP, Greenlees MJ, Webb JK, ShineR. 2007. Rapid expansion of the cane toad (Bufo marinus)invasion front in tropical Australia. Austral Ecology 32:169–176.
Phillips BL, Shine R. 2004. Adapting to an invasive species:toxic cane toads induce morphological change in Australiansnakes. Proceedings of the National Academy of Sciences ofthe United States of America 101: 17150–17155.
Phillips BL, Shine R. 2006. An invasive species inducesrapid adaptive change in a native predator: cane toads andblack snakes in Australia. Proceedings of the Royal SocietySeries B 273: 1545–1550.
Pizzatto L, Shine R. 2008. The behavioral ecology of canni-balism in cane toads (Bufo marinus). Behavioral Ecologyand Sociobiology 63: 123–133.
750 G. WARD-FEAR ET AL.
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 99, 738–751

Prohammer LA, Wade MJ. 1981. Geographic and geneticvariation in death-feigning behaviour in the flour beetle,Tribolium castaneum. Behavior Genetics 11: 395–401.
Quinn TP, Kinnison MT. 1999. Size-selective and sex-selective predation by brown bears on sockeye salmon. Oeco-logia 121: 273–282.
Rovee CK, Kaufman LW, Collier GH, Kent GC Jr. 1976.Periodicity of death feigning by domestic fowl in responseto simulated predation. Physiology and Behavior 17: 891–895.
Schwarzkopf L, Shine R. 1992. Costs of reproduction inlizards: escape tactics and susceptibility to predation.Behavioral Ecology and Sociobiology 31: 17–25.
Semeniuk M, Lemckert F, Shine R. 2007. Breeding-siteselection by cane toads (Bufo marinus) and native frogs innorthern NSW, Australia. Wildlife Research 34: 59–66.
Shattuck SO. 1999. Australian ants: their biology and iden-tification. Collingwood, Vic.: CSIRO Publishing.
Shine R, LeMaster MP, Moore IT, Olsson MM, MasonRT. 2001. Bumpus in the snake den: effects of sex, size, andbody condition on mortality of red-sided garter snakes.Evolution 55: 598–604.
Shine R, Sun L, Fitzgerald M, Kearney M. 2002. Acciden-tal altruism in insular pit-vipers (Gloydius shedaoensis,Viperidae). Evolutionary Ecology 16: 541–548.
Sih A. 1988. The effects of predators on habitat use, activityand mating behaviour of a semi-aquatic bug. Animal Behav-iour 36: 1846–1848.
Sih A. 1994. Predation risk and the evolutionary ecology ofreproductive behaviour. Journal of Fish Biology 45A: 111–130.
Snelson FF Jr. 1984. Seasonal maturation and growth of
males in a natural population of Poecilia latipinna. Copeia1984: 296–304.
Trexler JC, Tempe RC, Travis J. 1994. Size-selective pre-dation of sailfin mollies by two species of heron. Oikos 69:250–258.
Urban MC, Phillips BL, Skelly DK, Shine R. 2007. Thecane toad’s (Chaunus [Bufo] marinus) increasing ability toinvade Australia is revealed by a dynamically updatedrange model. Proceedings of the Royal Society Series B 274:1413–1419.
Wahle RA. 1992. Body-size dependent antipredator mecha-nisms of the American lobster. Oikos 65: 52–60.
Ward-Fear G. 2008. Ecological interactions between meta-morph cane toads (Bufo marinus) and meat ants (Iri-domyrmex reburrus) in tropical Australia. BSc (Honours)thesis, School of Biological Sciences, University of Sydney,Australia.
Ward-Fear G, Brown GP, Greenlees M, Shine R. 2009.Maladaptive traits in invasive species: in Australia, canetoads are more vulnerable to predatory ants than are nativefrogs. Functional Ecology 23: 559–568.
Ward-Fear G, Brown GP, Shine R. 2010. Using a nativepredator (the meat ant, Iridomyrmex reburrus) to reduce theabundance of an invasive species (the cane toad, Bufomarinus) in tropical Australia. Journal of Applied Ecologydoi: 10.1111/j.1365-2664.2010.01773.x.
Werner EE. 1986. Amphibian metamorphosis: growth rate,predation risk and the optimal size at transformation.American Naturalist 128: 319–341.
Williams CR, Brodie ED Jr, Tyler MJ, Walker SJ. 2000.Antipredator mechanisms of Australian frogs. Journal ofHerpetology 34: 431–443.
PREDATION ON ANURANS BY ANTS 751
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 99, 738–751




















