
Nest predation on woodland songbirds: when is nest predation density dependent
15
OIKOS 87: 65-74. Copenhagen 1999 Nest predation on woodland songbirds: when is nest predation density dependent? Kenneth A. Schmidt and Christopher J. Whelan Schmidt, K. A. and Whelan, C. J. 1999. Nest predation on woodland songbirds: when is nest predation density dependent? - Oikos 87: 65-74. Nest predation, if density dependent, may structure avian communities by selecting for nest-site characteristics that dilute the predators' perception of nest density (e.g., by increasing the number and types of potential nest sites) thereby reducing losses to predators. We tested for density dependent nest predation at a site dominated by predation from raccoons, Procyon lotor, and blue jays, Cyanocitta cristata. We used cohorts of 20 artificial ground nests placed on experimental grids at four densities (1.7, 7, 65, and 167 nests/ha). Predation was density dependent for one of two temporal replicates. In a second experiment, we tested for density dependent preda- tion on both ground and shrub nests using more realistic nest densities (3.5 and 8.9 nestslha) and excluding avian and small mammal predators by placing nylon screening over the nests. Predation was density dependent on artificial ground nests and density independent on shrub nests. Finally, we analyzed predation on natural shrub nests separating out avian (i.e., jays) and large mammalian predation based on disturbance to the depredated nest. In each of three years there was no evidence for density dependent predation. An outbreak of periodic cicadas in the first experiment and the maturation of a large mulberry crop during the second experiment demon- strated the importance of the distribution and abundance of alternative foods on the response of predators toward heterogeneity in nest abundance. K. A. Schmidt, Dept of Biological Sciences, Unic. of Illinois, 845 W. Taylor, Chicago, IL 60607, USA @resent address: Inst. of Ecosystem Studies, Box AB (Route &A), Millbrook, N Y 12545 [[email protected]]). - C. J. Whelan, Illinois Natural History Survey, Midewin National Tallgrass Prairie, 30071 S. Rt. 53, Wilmlngton, IL 60481, USA. Predation shapes community organization in a variety Under apparent competition and apostatic selec- of organisms (e.g., Kotler 1984, Sih et al. 1985, Holt tion, nest predation, if density dependent, could select and Lawton 1994, Kotler et al. 1994, Navarrete 1996, for nest-site characteristics that would effectively di- Wilson et al. 1996). Within avian communities, Martin lute the predator's perception of nest density by in- (1988a, b, c, 1993, 1996) and others (Ricklefs 1989, creasing the number and types of potential nest sites, Reitsma 1990) have suggested nest predation may be and thereby reducing losses to predators. There is evi- largely responsible for the diversity and composition of dence for this mechanism in the avian community breeding bird species. It has long been known that nest inhabiting mixed conifer drainages at a high elevation predation profoundly affects population productivity site in central Arizona (Martin 1993, 1996), but its (Ricklefs 1969, 1984, Martin 1992), but it was not until generality awaits further testing. Moreover, the neces- the conception of apostatic selection (Clarke 1969, sary assumption that nest predation is density de- Rand 1967, Ricklefs and O'Rourke 1975), and apparent pendent is not well established despite several stu- competition (Holt 1977, 1987, Jefferies and Lawton dies documenting it (e.g., Fretwell 1972, Page et al. 1984, Holt and Kotler 1987) that ecologists postulated 1983, Sugden and Beyersbergen 1986, Martin 1988b, a mechanism linking predation to niche diversification. 1996, Hoi and Winkler 1994). It is the assump- Accepted 14 December 1998 Copyright 0 OIKOS 1999 ISSN 0030-1299 Printed in Ireland - all rights reserved OIKOS 87:l (1999)
Transcript of Nest predation on woodland songbirds: when is nest predation density dependent

OIKOS 87: 65-74. Copenhagen 1999
Nest predation on woodland songbirds: when is nest predation density dependent?
Kenneth A. Schmidt and Christopher J. Whelan
Schmidt, K. A. and Whelan, C. J. 1999. Nest predation on woodland songbirds: when is nest predation density dependent? - Oikos 87: 65-74.
Nest predation, if density dependent, may structure avian communities by selecting for nest-site characteristics that dilute the predators' perception of nest density (e.g., by increasing the number and types of potential nest sites) thereby reducing losses to predators. We tested for density dependent nest predation at a site dominated by predation from raccoons, Procyon lotor, and blue jays, Cyanocitta cristata. We used cohorts of 20 artificial ground nests placed on experimental grids at four densities (1.7, 7, 65, and 167 nests/ha). Predation was density dependent for one of two temporal replicates. In a second experiment, we tested for density dependent preda- tion on both ground and shrub nests using more realistic nest densities (3.5 and 8.9 nestslha) and excluding avian and small mammal predators by placing nylon screening over the nests. Predation was density dependent on artificial ground nests and density independent on shrub nests. Finally, we analyzed predation on natural shrub nests separating out avian (i.e., jays) and large mammalian predation based on disturbance to the depredated nest. In each of three years there was no evidence for density dependent predation. An outbreak of periodic cicadas in the first experiment and the maturation of a large mulberry crop during the second experiment demon- strated the importance of the distribution and abundance of alternative foods on the response of predators toward heterogeneity in nest abundance.
K. A. Schmidt, Dept of Biological Sciences, Unic. of Illinois, 845 W. Taylor, Chicago, IL 60607, USA @resent address: Inst. of Ecosystem Studies, Box AB (Route &A), Millbrook, N Y 12545 [[email protected]]). - C. J. Whelan, Illinois Natural History Survey, Midewin National Tallgrass Prairie, 30071 S. Rt. 53, Wilmlngton, IL 60481, USA.
Predation shapes community organization in a variety Under apparent competition and apostatic selec-of organisms (e.g., Kotler 1984, Sih et al. 1985, Holt tion, nest predation, if density dependent, could select and Lawton 1994, Kotler et al. 1994, Navarrete 1996, for nest-site characteristics that would effectively di- Wilson et al. 1996). Within avian communities, Martin lute the predator's perception of nest density by in- (1988a, b, c, 1993, 1996) and others (Ricklefs 1989, creasing the number and types of potential nest sites, Reitsma 1990) have suggested nest predation may be and thereby reducing losses to predators. There is evi- largely responsible for the diversity and composition of dence for this mechanism in the avian community breeding bird species. It has long been known that nest inhabiting mixed conifer drainages at a high elevation predation profoundly affects population productivity site in central Arizona (Martin 1993, 1996), but its (Ricklefs 1969, 1984, Martin 1992), but it was not until generality awaits further testing. Moreover, the neces- the conception of apostatic selection (Clarke 1969, sary assumption that nest predation is density de-Rand 1967, Ricklefs and O'Rourke 1975), and apparent pendent is not well established despite several stu-competition (Holt 1977, 1987, Jefferies and Lawton dies documenting it (e.g., Fretwell 1972, Page et al. 1984, Holt and Kotler 1987) that ecologists postulated 1983, Sugden and Beyersbergen 1986, Martin 1988b, a mechanism linking predation to niche diversification. 1996, Hoi and Winkler 1994). It is the assump-
Accepted 14 December 1998
Copyright 0OIKOS 1999 ISSN 0030-1299 Printed in Ireland - all rights reserved
OIKOS 87:l (1999)

- --
- - -
- - - -
- - -
tion of density dependent nest predation that is the focus of the present paper.
Density dependent predation requires that predators assess and respond to spatial or temporal heterogeneity in resource (e.g., nest) abundance (Schmidt and Brown 1996). If this premise is correct, we expect to find density dependent (DD) nest predation only when predators can both recognize and respond to higher nest densities. This response may be functional (e.g., increased foraging time for nests; Holt and Kotler 1987), numerical (e.g., aggregation toward patches of higher nest density: Holt 1977), or psychological (e.g., search image formation, Tinbergen 1960, Morgan and Brown 1996). However, whether predators respond to heterogeneity in nest abundance or not is also poten- tially influenced by additional ecological variables, such as the abundance and distribution of alternative re-sources within the environment.
Medium-sized generalist predators (e.g., raccoons, opossums, corvids, gulls) are most likely to exhibit DD nest predation because only they have large enough home ranges and daily movements to be able to detect and respond to differences in nest density. Moreover, they are ecological "cream skimmers", i.e., they can quickly assess and shift their diet toward resources that are locally abundant. Here we describe observational studies and experimental tests for density dependent nest predation within a community of woodland song- birds in northeastern Illinois dominated by predation from raccoons, Procyon lotor, and blue jays, Cyanocitta cristata. In addition, outbreaks of cicadas and mulberry fruit during experimental studies in 1990 and 1995, respectively, enabled us to examine the influence of abundant, alternative food on predation rates and the strength of density dependence.
Methods Our studies were conducted in the deciduous wood- lands of The Morton Arboretum located about 45 km west of Lake Michigan in the western suburbs of Chicago, IL, USA, and at Hidden Lake Forest Pre- serve, an approximately 150-ha woodland abutting the Arboretum. The Arboretum's woodlands have a canopy dominated largely by Acer saccharum, Quercus alba, Q. rubra, Fraxinus americana, and Juglans nigra, and an understory consisting of Acer saplings, small trees (e.g., Ostrya viginiana), and native (e.g., Viburnum dentatum) and exotic (e.g., Loniccra maackii and Rham- nus cathartica) shrubs.
Between 1991 and 1996, we used remote cameras (see Whelan et al. 1994) attached to artificial nests baited with quail (Coturnix japonica) eggs to photograph 137 nest predation events (Table 1). Raccoon and opossum (Didelphis cirginianis) comprised 70% and 11% (all
prior to 1995), respectively, of photos of ground nest predators. (Table 1). Blue jay comprised 9% of photos and sciurids (Sciurus carolinensis, S. nigra, Tamias striatus) only 7%. While Roper (1992) and Haskell (1995) argued that quail eggs underestimate predation by smaller predators (e.g., chipmunks), we believe this is unlikely since quail eggs offered to chipmunks were readily consumed (unpubl. data). In addition, raccoons and jays comprised 44% and 56%, respectively, of predators photographed at shrub nests (n =45; Table 1). While no other species have been photographed depredating shrub nests, we have observed crows and grackles depredating thrush nests. Feral cats, Felis do- mesticus, and snakes (e.g., Thamnophis sirtalis) are rare at the site.
Experiment 1
On 18 June 1990, we established (following the method- ology of Reitsma 1990) plots of 20 artificial nests baited with quail eggs (Coturnix japonica) at two sites: four plots in the East Woods of The Morton Arboretum and three plots at Hidden Lake ( > 1 km apart). In addition to the spatial replicate, we repeated the experiment beginning 20 July, 1990. We concealed nests on the ground under vegetation and monitored them every five days for 15 days. Nests were evenly dispersed within plots and their density altered by varying the internest distance. Densities ranged from 166.7 nestslha (10-m spacing between nests; only one replicate in the East Woods), 65 nestslha (16-m spacing), 6.7 nestslha (50-m spacing) and 1.67 nestslha (100-m spacing). Nests were considered depredated if one or both eggs were missing or damaged. Depredated nests were not replaced. Dur- ing the June replicate there was an emergence of 17- year periodic cicadas (Magicicada sp.). Cicadas were rare or absent during the July replicate.
Experiment 2
In 1995 we revised our methodology to include shrub nests and eliminate avian and small mammal predation
Table 1. Photographs of predators on artificial nests baited with quail eggs.
Ground nests Shrub nests
Predator 1992 1994 1995 1995 1996
Raccoon 36 7 21 15 1 Opossum 7 4 Squirrel 2 Chipmunk 1 - 4 - -
Blue Jay 3 4 2 5 23 Unknown 1 1 Total 48 17 27 20 25

and instead focus on predation by large mammals by screening nests to exclude the smaller predators. More- over, we eliminated the practice of decreasing nest density by increasing the internest distance. This design suffers several methodological problems including: 1) uniform nest distribution (Picman 1988); 2) lack of dimensionality, i.e., linear transects may lead to traplin- ing by predators (see Whelan et al, 1994); 3) increasing the internest distance from 10 to 100 m results in large differences in plot size and thus possibly in the number of predators, habitat structure, etc.
On 5 June 1995 we established four 2.25-ha plots (150 x 150 rn) in the East Woods separated by at least 0.5 km. Each plot was composed of thirty-six 0.0625-ha subplots (25 x 25 m). We baited each plot with artificial wicker nests containing one quail egg. Two plots con- tained ground nests lined with leaves and inconspicu- ously placed in a shallow depression on the ground under a shrub or sapling. We placed leaves over the nest to partially conceal the egg and minimize preda- tion by visually-searching predators (e.g., blue jays). The two remaining plots contained nests placed 0.5-2 m above ground fastened with a wire tie to a shrub or sapling so as to mimic a natural nest. We fitted nylon screening over the top of the nest to prevent access to avian and small mammalian predators. Small mammals are probably capable of chewing through the nest or screening, but they are rare predators (see above), and the condition of depredated nests or nest substrate suggested larger predators (e.g.. claw marks, fur, knocked over shrubs). We did not place screens on ground nests because nests were not fastened down, and might have been difficult to relocate had predators displaced the nests during their efforts to remove the screening. Photographs (see above) indicated that blue jays and rodents were only occasional ground nest predators. Depredated nests were often chewed, and dislodged as far as several meters indicating larger predators. The few depredated ground nests without these indicators were attributed to jays or rodents and not included in the analyses.
We used two densities: 8.9 nestsiha (20 nests/plot) or 3.5 nestslha (8 nests/plot). Equal numbers of nests were placed in each quarter plot (2 and 5 for low and high density treatments, respectively; Fig. I ) , but to mini- mize uniform nest dispersion, we randomly assigned subplots within a quarter plot to receive a nest. No subplot contained more than one nest. Moreover, placement within a subplot was random (Fig. 1). We checked nests each morning and replaced them if depre- dated to hold density constant. The replacement nest was placed in a randomly chosen subplot as just de- scribed. We maintained treatments at a plot for five days, after which we collected all nests. We replicated the experiment three additional times with two days between replicates in which no nests were present, and rotated treatments among the plots following a Latin
OIKOS 87: l (1999)
whole plot: 150 x 150 m
quarter plot 75x75 m
3.5 nestslha 8.8 nestslha
Fig. 1. Diagrammatic representation of experimental setup for experiment 2 (see text). Nests are represented by circles.
Square design. In this way, each treatment was present at one plot per each replicate, and over the course of the experiment, each plot received each treatment once.
The artificial wicker nests do not mimic the appear- ance of real bird nests, but the predator of interest (i.e., raccoon) is nocturnal and searches olfactorily (Whelan et al. 1994), and thus nest appearance should not affect the results. Moreover, all nests regardless of density had the same appearance. We did not attempt to eliminate human scent associated with placing out artificial nests. Although human scent may affect predation rates, these effects are inconsistent (Whelan et al. 1994). Moreover, scent trails were not reliably associated with a reward. From May through July, field personnel walked through all experimental plots in search of nesting songbirds, thus human scent was already associated with the plots.
Large scale patterns: nest predation on a guild of shrub-nesting species
We examined nest predation on the community of nesting woodland songbirds within the Arboretum's East Woods from 1994-1996. Nesting species included wood thrush (see Appendix for scientific names), Amer- ican robin, blue jay, grey catbird, northern cardinal, and rose-breasted grosbeak. The first three species regu- larly nest within the woods, and make up > 90% of discovered nests. The remaining species often nest at woodland edges or in the Arboretum's collections, but also may be found nesting in canopy gaps within the woods. All the species have large, roughly similarly sized and shaped open cup nests placed generally within the shrub layer (1-3 m). Some robin and jay nests were higher, but those above s 7.5 m were not included because their fate could not always be unambiguously determined. Nests of other species were not included because they nest on woodland edges, within different vegetative strata (e.g., on the ground or in the sub- canopy), or their nests have a distinctly different ap- pearance (e.g., red-eyed vireo). Nests were located by systematically searching the woods. The majority were
67

found during or before egg laying and monitored, in general, every 2-5 days. Based on the size and conspic- uousness of the nests, the distribution of singing males (wood thrush), and the small number of nests found after the breeding season, we estimate we found > 90% of the nests for these species within our study area.
To arrive at nest density estimates during the season, we used detailed nest records to determine the date of fledging the final young or the date of depredation. Nest predation was determined by the stage of the nesting cycle at which eggs or young disappeared, the condition of an empty nest, and the presence or behav- ior of nearby adults/fledglings. We assumed predation occurred at the midpoint between nest checks when exact dates were unknown. Fledging dates were esti-mated in the same way, or when records were detailed enough, we determined the probable date of fledging based on the length of the various stages of the nest cycle (from Ehrlich et al. 1988). A small number of nests (spaced throughout the season) were not included because the time we observed them was too brief to determine their stage, or their fate was ambiguous.
To test for density dependent nest predation within a breeding season we divided the season into a series of three-day cohorts. Nest densities were calculated as the mean number of nests initially entering the cohort (active on day 1) and the number active on day 3 (given they were present on day 1). For cohorts with a mini- mum density of 10 nests, we calculated the cohort predation rates as number of nests depredatedlmean density.
Data analysis
We used Mantel-Haenszel tests to compare nest sur-vival distributions and contingency table analyses to compare overall levels of survivorship at the end of experiment 1. The former tests for differences in the rate of nest predation between treatments, while the latter tests for differences at the end of the experimental trial. We used Fisher's exact tests in pair-wise compari- sons of the number of surviving versus depredated nests to locate differences among density treatments at each site. For experiment 2, we used x2 analyses to test for differences in the frequency of nest predation between treatments and between halves of the experiment, i.e., early (pre-mulberry ripening) and late (post-mulberry ripening); see below.
To test for density dependent predation on real bird nests, we used Spearman's rank correlation to test if the proportion of nests depredated was correlated with active nest density. To test for differences in predation rates on real nests between the pre- and post-mulberry ripening, we used Mann-Whitney U-tests.
Results
Experiment I
Nest survivorship was density dependent at both sites for the June replicate (East Woods: Mantel-Haenszel x: = 13.03, P = 0.005; Hidden Lake: Mantel-Haenszel xi = 14.6, P = 0.001), but not the July replicate (all P >0.1). In June, the rate of predation increased from the least dense to the most dense plot at both sites (Fig. 2) with one exception at the East Woods where the 65 nestlha plot exceeded that of the 166.7 nests,lha plot between the census at day 10 and day 15. In July, all nests were depredated in 5 of 7 plots by day 10 of the experiment, and 6 of 7 plots by day 15 (Fig. 2). Similarly, the overall amount of predation differed among the density plots at both sites for the June replicate (Fig. 2; East Woods: x: = 19.9, P <0.001; Hidden Lake: x: = 21.51, P <0.0001), but not the July replicate (where most plots had no nests surviving). For the June replicate, the following pattern of overall nest predation among treatments emerged at the East Woods: 1.67 > 6.7 > 166.7 (P's <0.003) and 65 = 166.7 (P = 0.08); Hidden Lake: 1.67 > 6.7 ( P <0.01) and 6.7 = 5 (P > 0.30).
Experiment 2
Halfway through experiment 2, the Arboretum's Moraceae (mulberry, Morus rubra, and M. alba) collec- tion produced an exceptionally abundant fruit crop that provided alternative food for raccoons. Spotlighting observations found raccoon density was high in the mulberry collection but low in the woods, and raccoon scat collected at this time (Berris 1997) documented their diet consisted predominantly of fruit, particularly Morus spp. The percent of scats containing bird parts (e.g., feathers, egg shell fragments) dropped from 10- 21% in 1994 to 0% in 1995 during this time period (Berris 1997).
More total nests were depredated before maturation of mulberry fruit (222 versus 143). Moreover, a signifi- cantly greater proportion of ground nests were depre- dated in high density plots (Fig. 3) during the pre-maturation period (x: = 8.45, P <0.005), but not during the post-maturation period (x:=0.87, P >0.50). Predation on shrub nests was density independent (Fig. 3) both during pre-maturation (x: = 2.43, P > 0.10) and post-maturation of mulberry fruit (x: = 1.11, P >0.25). We believe that as a consequence of the availability of alternative food, predation rates on both real (see be- low) and artificial nests declined concomitantly and the density dependence nature of predation on ground nests was altered.
To determine if nests were equally depredated over the five days of exposure during a replicate, we tabu-
OIKOS 87:1 (1999) 68

lated the number of depredated nests by treatment per day of exposure (1 -5), pooling all replicates. Predation was spread evenly throughout the exposure period for all treatments (P>0.25) except the high shrub density treatment (xi =24.33, P <0.005). The high shrub den- sity treatment had more nests depredated (n =42) dur- ing the first two days of exposure than the remaining three days of exposure (n = IS), although the effect was seen solely during the second replicate. The lack of significant effects indicates raccoons did not learn to cue in on nest locations or develop search images over the period of a replicate (the significant effect for the high shrub density treatment is in the opposite direc- tion). In addition, raccoons' responses when treatments were switched on a plot were rapid (i.e., the number of nests depredated on day one was similar to all other days), indicating that previous treatments did not influ- ence predation rates.
Predation on a guild of shrub-nesting species
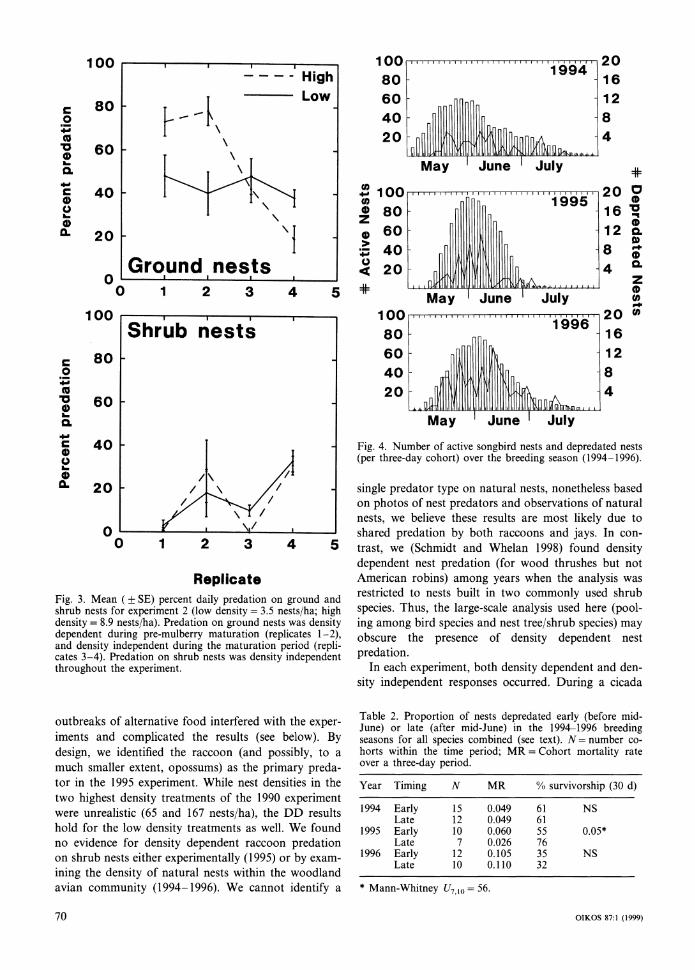
Nest densities and the number of nests depredated (by cohort) throughout the breeding season for the three years of this study are shown in Fig. 4. In no year was
Fig. 2. Percent of nests (n = 20) surviving over a 15-day simulated incubation period for the different nest densities at East Woods and 20 - I
IHidden Lake sites. The 166.7 I
nestslha was used only during I I
the June replicate at the East 0
the proportion of nests depredated significantly corre- lated with nest density (1994: n =27, r, =0.321; 1995: n = 17,rs=0.134; 1996:n=22, rs=0.254,al1 P>0.10).
To examine the impact of early mulberry maturation on nesting songbirds, we compared nest predation rates per cohort starting before and after mid-June (period of mulberry fruit maturation in 1995). For 1994 and 1996, predation rates were nearly identical between the two halves of the breeding season (Table 2), while in 1995 predation rates dropped by 21% during the maturation period (difference in cohort survival rate extrapolated out to a 30-day nesting cycle; Table 2). Thus, there was a concomitant drop in predation rates on artificial and real nests that corresponds to the period of high fruit abundance in 1995. That a similar drop was not seen in other years strongly suggests the exceptionally large mulberry crop and early maturation was the cause of reduced nest predation, rather than this being a sea-sonal phenomenon.
Discussion
We documented density dependent predation on artifi- cial ground nests both in 1990 and 1995, although
HL: June
i \ I I \ I i -'\ \
*-.. \ . -.+-----*
I I
0 5 10 I 5 20
I
EW: July HL: July
- - - A -Woods. Density dependence 0 5 I 0 15 20 0 5 10 15 was statistically significant only in the June replicate. Day Day
20
I I I I - - - - High Low
- l--k\
May I June I July =k- w -\ \
\ I Ground nests ,
I I I I
Shrub nests
-
Replicate Fig. 3. Mean (iSE) percent daily predation on ground and shrub nests for experiment 2 (low density = 3.5 nests/ha; high density = 8.9 nestsiha). Predation on ground nests was density dependent during pre-mulberry maturation (replicates 1-2), and density independent during the maturation period (repli- cates 3-4). Predation on shrub nests was density independent throughout the experiment.
outbreaks of alternative food interfered with the exper- iments and complicated the results (see below). By design, we identified the raccoon (and possibly, to a much smaller extent, opossums) as the primary preda- tor in the 1995 experiment. While nest densities in the two highest density treatments of the 1990 experiment were unrealistic (65 and 167 nestsiha), the DD results hold for the low density treatments as well. We found no evidence for density dependent raccoon predation on shrub nests either experimentally (1995) or by exam- ining the density of natural nests within the woodland avian community (1994-1996). We cannot identify a
May ' June July
Fig. 4. Number of active songbird nests and depredated nests (per three-day cohort) over the breeding season (1994-1996).
single predator type on natural nests, nonetheless based on photos of nest predators and observations of natural nests, we believe these results are most likely due to shared predation by both raccoons and jays. In con- trast, we (Schmidt and Whelan 1998) found density dependent nest predation (for wood thrushes but not American robins) among years when the analysis was restricted to nests built in two commonly used shrub species. Thus, the large-scale analysis used here (pool- ing among bird species and nest treelshrub species) may obscure the presence of density dependent nest predation.
In each experiment, both density dependent and den- sity independent responses occurred. During a cicada
Table 2. Proportion of nests depredated early (before mid- June) or late (after mid-June) in the 19941996 breeding seasons for all species combined (see text). N = number co-horts within the time period; MR = Cohort mortality rate over a three-day period.
Year Timing N MR % survivorship (30 d)
1994 Early 15 0.049 61 N S Late 12 0.049 61
1995 Early 10 0.060 55 0.05* Late 7 0.026 76
1996 Early 12 0.105 35 NS Late 10 0.110 32
* Mann-Whitney I/,,,, = 56

outbreak in 1990, nest predation was DD, but DI afterwards. In contrast, in 1995, nest predation was DD before mulberry maturation and DI during fruit matu- ration. We believe the cicada outbreak in June 1990 did not negate density dependence, but rather unmasked its presence. Predation rates were lower across plots during the cicada outbreak than afterwards (compare June vs July; Fig. 2). This enabled us to resolve differences between density treatments during the first temporal replicate in 1990, but not the second replicate when high predation prohibited any conclusions (e.g., at Hid- den Lake, only one nest survived to the first nest check; Fig. 2). In 1995, the experimental methodology was better suited to resolve possible differences between treatments. In this case, the presence of mulberries appears to have negated density dependent predation (Fig. 3).
Why should nest predation be DD in the presence of cicadas, but not in the presence of mulberries? We believe the answer lies in the distribution of the alterna- tive food with respect to nests. The Moraceae collection lies outside the woods where we monitored nesting densities and performed the experiments. Thus, rac-coons were selecting an alternative habitat to forage in, and did not encounter nests while foraging for mulber- ries. Raccoons foraging in the collections could become satiated to the point that searching for nests was un- profitable, and nests were consumed only if encoun-tered incidentally (Vickery et al. 1992). In contrast, cicadas and nests co-occurred within the woods, and raccoons may have responded to their cumulative for- aging success on both nests and cicadas. Thus, during the cicada outbreak, nest predation remained DD. While our explanation for the apparent shift in density dependence seems reasonable, it requires rigorous ex-perimental testing to eliminate alternative hypotheses.
Other studies have documented changes in density dependence in relation to alternative foods. Dunn (1977) showed predation on tits (Pauus spp.) by weasels significantly increased with tit density, but using multi- ple regression, the effect of tit density was nonsignifi- cant if rodent density (alternative prey for weasels) was included in the model. Similarly, Hogstad (1995) found that nest predation in fieldfares by weasels was density dependent during years with low rodent density, but density independent during years with high rodent density.
One drawback of our experimental design involves the scale of replication; raccoons may be capable of travelling the 500 m between plots during replicates. However, if raccoons were discovering and aggregating towards the high nest density plots we would expect to see more nests depredated at those plots over the five days of a replicate. That we did not indicates that, at least within the time period of a replicate, plots were independent.
Table 3. Open-nest woodland breeding species historically occurring in the Chicago region and either presently occurring or presently absent from The Morton Arboretum. Species occurring historically were inferred based on species' accounts given in Cory (1909) and Ford et al. (1934). Species listed as rare by these authors (e.g., cerulean warbler), not expected based on lack of suitable habitat (e.g., prothonotary warbler), or edge specialists (e.g., indigo bunting, yellow-breasted chat) are not included in the counts. Scientific names are given in the Appendix. Veery. hooded warbler, and black-billed cuckoo are listed as absent but may nest on rare occasion in some years (pers. obs.). Absent species are disproportionately distributed among the groundllow shrub nesters.
Historically: Present Present Today: Present Absent
Ground,llow shrub 1 6* P =0.004 Fisher's
Shrubjsubcanopy 11 2T
* Veery, ovenbird, Kentucky warbler, hooded warbler, whip- poor-will, ruffed grouse. + black-billed cuckoo, least flycatcher.
Community implications
Martin (1988a) predicted that if DD predation is act- ing, fewer species should nest in vegetation layers with the greatest nest predation. Communities of birds in Iowa (Best and Stauffer 1980, Stauffer and Best 1986) and Arizona (Martin 1988b) fit the pattern. In accord with Martin's prediction, at The Morton Arboretum, six of seven ground or low shrub ( < 1 m) nesting species are presently absent in all or most years despite being common historically (Table 3). In contrast, only two of 13 shrub or subcanopy nesters recorded histori- cally are presently absent. Moreover, these changes are not limited to The Morton Arboretum, but have oc- curred in other Chicago area forest preserves (e.g., Wright Woods, MacArthur Woods; De Vore 1996, S. Hickman pers. cornrn.).
These changes to the avian community at The Mor- ton Arboretum (and other woodlands) may be related to factors other than nest predation; loss of suitable breeding habitat (e.g., successional change [Holmes et al. 19861, forest fragmentation [Robbins et al. 1989]), or regional avian population declines (James et al. 1996) are complementary or alternative explanations. How- ever, meso-predator release (Soule et al. 1988), urban- ization (Hoffman and Gottschang 1977, Adams 1994, Engels and Sexton 1994), and fragmentation (Bider 1968, Andren 1992) create environments conducive to important and widespread nest predator species, such as raccoons and jays (Wilcove 1985, Miller and Knight 1993, Picman and Schriml 1994, Whelan et al. 1994, this study). Raccoon sightings during annual spring spotlighting surveys conducted in over 40 counties in Illinois have tripled since 1981 (Bluett 1996), and paral- lels the decline in the estimated annual raccoon harvest in Illinois from > 380000 animals in 1979 to a low in 1990 at about 71 000 animals. Current raccoon densities
OIKOS 87.1 (1999)

in forest preserves around the Chicago region may be as high as one raccoon per 1-2 ha (S. Gehrt pers. comm.), with forest preserves surrounded by urban development, as is The Morton Arboretum, having the highest raccoon densities (S. Gehrt pers. comm.).
When is nest predation density dependent?
We argue that assessment and response by predators to heterogeneity in nest abundance is necessary and suffi- cient to generate DD nest predation. These factors will be influenced by nest density, predators' home range size, and the abundance and distribution of alternative foods. Higher nest density allows foragers to better distinguish between patches of differing quality (Valone and Brown 1989, Schmidt and Brown 1996), and aids the formation of search images which otherwise may be unlikely when densities are low (Tinbergen 1960, Mor- gan and Brown 1996). Moreover, resource density de- termines the benefit of responding and if abundant alternative food is available, a forager may forego its response to the nests in favor of alternative food (Dunn 1977, Hogstad 1995, this study).
Predators with small home ranges (e.g., 1-2 ha or smaller; Tamias, Tamiasciurus, Spermophilus) will be unable to detect larger scale heterogeneity in nest abun- dance because individuals cannot sample the full range of heterogeneity. As a population, these predators may be important, but an individual predator simply en- counters too few nests to likely detect or respond to higher nest densities, unless densities are artificially high (e.g., Reitsma 1990, 1992). As expected, these predators have shown DI responses to nest density (Boag et al. 1984, 07Reilly and Hannon 1989, Reitsma 1990, 1992). Martin's (1988b, 1996) studies with red squirrels pro- vides the exception, but see Reitsma (1990, 1992). How- ever, given that nests were spaced (on linear transects) at least 40 m apart, it is unlikely that an individual squirrel had access to more than a handful of nests, questioning whether red squirrels were solely responsi- ble for the increased predation rates seen at higher nest densities.
Predators with large home ranges (e.g., corvids, gulls, canids, raccoon) can exploit heterogeneity in nest abun- dance by virtue of their access to a large numbers of nests, and their ability to detect resource heterogeneity at large spatial scales. These large predators, in fact, are responsible for nearly all the observed cases of DD nest predation (e.g., Krebs 1971, Fretwell 1972, Dunn 1977, Potts 1980, Page et al. 1983, Sugden and Beyersbergen 1986, Hoi and Winkler 1994, this study). Density inde- pendent responses have occurred as well, but are lim- ited to studies with very low nest densities (e.g., < 1 nest/ha, e'g'9 O'Reil l~and Harmon 1989,AndrCn 1991, Niemuth and B o ~ c e 1995) when predators are likely to have difficulty assessing nest density and/or the cost of
missed opportunity (i.e., not foraging for alternative food) may outweigh the benefit of responding to low nest densities (Dunn 1977, this study). Breeding birds may exploit predator responses by nesting at low densi- ties to avoid DD predation (Fretwell 1972, Knapton 1979). Thus, the over-dispersion of bird nests can be seen as a phenomenon of the "Ghost of Predation Past", a proposition that, unfortunately, may be very difficult to test (see Connell 1980).
Finally, snakes are important nest predators that may have territories large enough (e.g., > 10 ha; Durner and Gates 1989, Plummer and Congdon 1994) to potentially encounter a significant number of nests. However, their low metabolic demand and tendency to remain relative inactive or restricted to a small portion of their home range for up to two weeks (Durner and Gates 1989) limits their capability to quickly respond to higher nest densities. Thus, it is not surprising that studies where snakes have been the dominant nest predator have yielded DI results (Best 1978, Gottfried 1978, Zimmerman 1984).
In conclusion, we have demonstrated strong and density dependent nest predation by large mammalian predators on artificial ground nests. We argue that this may explain the low abundance and the skewed distri- bution of local extinction of ground nesting species observed in and around the study area. The applicabil- ity of these results to similar woodlands elsewhere awaits further testing. Nonetheless, the ability of gener- alist predators to both structure their prey community and respond to anthropogenic effects of habitat alter- ation suggest that such impacts may be a general feature of predator-prey ecology within human altered landscapes.
Acknowledgements - Special thanks go to Steve Dilger, Mike Dilger, Lynnette Koutnik, and Dave Robson for enduring the rigors of field work to collect the nest predation data. We thank Joel Brown, Burt Kotler, Tom Martin, Scott Robinson, and Tom Poulson for stimulating conversation and sugges- tions with the manuscript. We also thank Bob Bluett and Stan Gehrt for sharing information and data on raccoon abundance throughout Illinois, and Scott Hickman for information on breeding bird densities for several Chicago area forest pre- serves. Quail eggs were supplied by Henry Marks and Vivian Patten of the Southeast Poultry Research Laboratory of the Agricultural Research Service, South Atlantic Area, U.S. Dept of Agriculture. This research was supported by grants from Sigma Xi (Grant-in-aid-of-Research), NSF DIG (both to K.A.S.), and the Emily Rodgers Davis Chair Endowment to The Morton Arboretum. This work is in partial fulfillment of the requirements for a doctorate (to K.A.S.) in ecology and evolution in the Dept of Biological Sciences at the Univ. of Illinois at Chicago.
References Adams, L, W, 1994, Urban habitats: a landscape
perspective. - Univ. of Minnesota Press, Minneapolis, MN.

Andren, H. 1991. Predation: an overrated factor for over-dis- persion of birds' nests? - Anim. Behav. 41: 1063-1069.
Andren, H. 1992. Corvid density and nest predation in relation to forest fragmentation: a landscape perspective. - Ecology 73: 794-804.
Berris, L. S. 1997. Mechanisms affecting foraging decisions of the raccoon (Procyon lotor) in northeastern Illinois. - MS dissertation, Univ. of Illinois at Chicago, Chicago.
Best, L. B. 1978. Field sparrow reproductive success and nesting ecology. - Auk 95: 9-22.
Best, L. B. and Stauffer, D. F. 1980. Factors affecting nesting success in riparian bird communities. - Condor 82: 149- 158.
Bider, J. R. 1968. Anllnal activity in uncontrolled terrestrial communities as determined by a sand transect technique. -Ecol. Monogr. 38: 269-308.
Bluett, B. 1996. 1996 Spotlight survey: furbearer program management note 96-3. - Illinois Dept of Natural Re-sources, Div. of Wildlife Services.
Boag, D. A., Reebs, S. G. and Schroeder, M. A. 1984. Egg loss among spruce grouse inhabiting lodgepole pine forests. - Can. J. Zool. 62: 1034-1037.
Clarke, B. 1969. The evidence for apostatic selection. - Hered-ity 24: 347-352.
Connell, J. H. 1980. Diversity and the coevolution of competi- tors, or the ghost of competition past. - Oikos 35: 131- 138.
Cory, k.B. 1909. The birds of Illinois and Wisconsin. - Field Museum of Natural History, Publ. 131, Chicago.
De Vore, S. 1996. Breeding birds of the Ryerson Conservation Area, Lake County. - Meadowlark 5: 53-56.
Dunn. E. 1977. Predation by weasels (Mustela nicalis) on breeding tits (Parus spp.) in relation to the density of tits and rodents. - J. Anim. Ecol. 46: 633-652.
Durner, G. M. and Gates, J. E. 1989. Spatial ecology of black rat snakes of Remington Farms, Maryland. - J. Wildl. Manage. 57: 812-826.
Ehrlich, P. R., Dobson, D. S. and Wheye, D. 1988. The birder's handbook: a field guide to the natural history of North American birds. - Simon and Schuster. New York.
Engels, 7 . M. and Sexton, C. W. 1994. Negative correlation of blue jays and golden-cheeked warblers near an urbanizing area. - Conserv. Biol. 8: 286-290.
Ford, E. R., Sanborn, C. C. and Coursen, C. B. 1934. Birds of the Chicago region. - Chicago Academy of Sciences, Chicago.
Fretwell, S. D. 1972. Populations in a seasonal environment. -Princeton Univ. Press, Princeton, NJ.
Gottfried, B. M. 1978. Anti-predator aggression in birds nest- ing in old field habitats: an experimental analysis. - Con-dor 81: 251-257.
Haskell, D. G. 1995. Forest fragmentation and nest predation: are experiments with Japanese quail eggs misleading? -Auk 112: 767-770.
Hoffman, C. 0, and Gottschang, J. L. 1977. Numbers, distri- bution, and movements of a raccoon population in a surburban residential community. - J. Mammal. 58: 623- 636.
Hogstad, 0 . 1995. Do avian and mammalian predators select for different nest dispersion patterns of Fieldfares Turdus pilaris? A 15-year study. - Ibis 137: 484-489.
Hoi, H and Winkler, H. 1994. Predation on nests: a case of apparent competition. - Oecologia 98: 436-440.
Holmes, R. T., Sherry, T. W. and Sturges, F. W. 1986. Bird community dynamics in a temperate deciduous forest: long-termtreids at Hubbard ~ r ~ o k . Ecol. Monogr. 56: -
201 -220. Holt, R. D. 1977. Predation, apparent competition, and the
structure of prey communities. - Theor. Popul. Biol. 12: 197-229.
Holt. R. D. 1987. Prey communities in patchy environments. - Oikos 50: 276-290.
Holt. R. D. and Kotler, B. P. 1987. Short-term apparent competition. - Am. Nat. 130: 412-430.
Holt, R. D. and Lawton, J. H. 1994. The ecological conse- quences of shared natural enemies. - Annu. Rev. Ecol. Syst. 25: 495-520.
James, F. C., McCulloch, C. E. and Wiedenfeld, D. A. 1996. New approaches to the analyses of population trends in land birds. - Ecology 77: 13-27.
Jefferies, M. J. and Lawton, J. H. 1984. Enemy-free space and the structure of ecological communities. - Biol. J. Linn. SOC. 23: 269-286.
Knapton, R. W. 1979. Optimal size of territory in the clay-col- ored sparrow. Spiza pallidn. - Can. J. Zool. 57: 1358- .---13 10.
Kotler, B. P. 1984. Risk of predation and the structure of desert rodent communities. - Ecology 65: 689-701.
Kotler, B. P., Brown, J. S. and Mitchell, W. A. 1994. The role of predation in shaping the behavior, morphology, and community organisation of desert rodents. - Aust. J. Zool. 42: 449-466.
Krebs, J. R. 1971. Territory and breeding density in the great tit Parus major. - Ecology 52: 2-22.
Martin, T. E. 1988a. Processes organizing open-nesting bird assemblages: competition or nest predation? - Evol. Ecol. 2: 37-50.
Martin, T. E. 1988b. On the advantage of being different: nest predation and the coexistence of bird species. - Proc. Natl. Acad. Sci. USA 85: 2196-2199.
Martin, T. E. 1988~. Habitat and area effects on forest bird assemblages: is nest predation an influence? - Ecology 69: 74-84.
Martin, T. E. 1992. Breeding productivity considerations: what are the appropriate habitat features for management? - In: Hagan, J. M. and Johnson, D. W. (eds), Ecology and conservation of neotropical migrant landbirds. Smithso- nian Inst. Press. Washington, pp. 455-473.
Martin, T. E. 1993. Nest predation and nest sites: new per- spectives on old patterns. - BioScience 43: 523-532.
Martin. T. E. 1996. Fitness costs of resource overlap among coexisting bird species. - Nature 380: 338-340.
Miller, C. K. and Knight, R. 1993. Does nest predator assem- blage affect reproductive success in songbirds? - Condor 95: 712-715.
Morgan, R. A. and Brown, J. S. 1996. Using giving-up densi- ties to detect search images. - Am. Nat. 148: 1059-1074.
Navarrete, S. A. 1996. Variable predation: effects of whelks on a mid-intertidal successional community. - Ecol. Monogr. 66: 301-322.
Niemuth, N. D. and Boyce, M. S. 1995. Spatial and temporal patterns of predation of simulated sage grouse nests at high and low nest densities: an exoerimental studv. - Can. J. Zool. 73: 819-825.
O'Reillv. P and Hannon. S. J. 1989. Predation of simulated wil~bw ptarmigan nests: the influence of density and cover on suatial and temooral uatterns of uredation. - Can. J. z00i.67: 1263-1287. A
Page. G. W.. Stenzel. L. E.. Winkler. D. W. and Swarth. C. -W. 1983. Spacing out at Mono Lake: breeding success, nest density, and predation in the snowy plover. - Auk 100. * m A .
15-L4.
Picman, J. 1988. Experimental study of predation on eggs of ground-nesting birds: effects of habitat and nest distribu- tion. - Condor 90: 124-131.
Picman, J. and Schriml, L. M. 1994. A camera study of temporal patterns of nest predation in different habitats. -Wilson Bull. 106: 456-465.
Plummer, M. V. and Congdon, J. D. 1994. Radiotelemetric study of activity and movement of racers (Coluber consrric- tor) associated with a Carolina bay in South Carolina. -Copeia (1 994):20-26.
Potts, G. R. 1980. The effects of modern agriculture, nest predation, and game management on the population ecol- ogy of partridges, Perdix perdix, and Alectoris rz&. - Adv. Ecol. Res. 11: 1-82.

Rand, A. S. 1967. Predator-prey interactions and the evolution of aspect diversity. - Atas do Simposio sobre a Biotia Amazonica 5: 73-83.
Reitsma, L. R. 1990. Importance of nest predation to avian population regulation and community structure in a north- ern hardwoods forest. - Ph.D. Dissertation, Dartmouth College, Hanover, NH.
Reitsma, L. R. 1992. Is nest predation density dependent? A test using artificial nests. - Can. J. Zool. 70: 2498-2500.
Ricklefs, R. E. 1969. An analysis of nesting mortality in birds. - Smithsonian Contrib. Zool. 9: 1-48.
Ricklefs, R. E. 1984. The optimization of growth rates in altricial birds. - Ecology 65: 1602- 1616.
Ricklefs, R. E. 1989. Nest predation and the species diversity of birds. - Trends Ecol. Evol. 2: 183-186.
Ricklefs. R. E. and O'Rourke, R. 1975. Aspect diversity in moths: a temperate-tropical con~parison. - Evolution 29: 313-324,
Robbins, C. S., Dawson, D. K. and Dowell, B. A. 1989. Habitat area requirements of breeding forest birds of the Middle Atlantic states. - Wildl. Monogr. 103: 1-34.
Roper, J. J. 1992. Nest predation experiments with quail eggs: too much to swallow? - Oikos 65: 528-530.
Schmidt, K. A. and Brown, J. S. 1996. Patch assessment in fox squirrels: the role of resource density. patch size. and boundaries. - Am. Nat. 147: 360-380.
Schmidt, K. A. and Whelan, C. J. 1998. Predator-mediated interactions between guilds of nesting songbirds: experi- mental and observational evidence. - Am. Nat. 152: 393- 402.
Sih, A,, Crowley, P.. McPeek. M., Petranka, J. and Strohmeier, K. 1985. Predation, competition, and prey communities: a review of field ex~eriments. - Annu. Rev. Ecol. Syst. 16: 269-31 1.
Soule, M. E., Bolger, D. T., Alberts, A. C., Sauvajot, R., Wright, J., Sorice, M. and Hill. S. 1988. Recon- structed dynamics of rapid extinctions of chaparral-requir- ing birds in urban habitat islands. - Conserv. Biol. 2: 75-92.
Stauffer. D. F. and Best, L. B. 1986. Nest-site characteristics of open-nesting birds in riparian habitats in Iowa. -
Wilson Bull. 98: 231-242. Sugden, L. G. and Beyersbergen, G. W. 1986. Effect of density
and concealment on American crow predation of simulated duck nests. - J. Wildl. Manage. 50: 9-14.
Tinbergen, L. 1960. The natural control of insects in pine woods. I. Factors influencing the intensity of predation by songbirds. - Arch. Neerl. Zool. 13: 265-343.
Valone, T. J. and Brown, J. S. 1989. Measuring patch assess- ment abilities of desert granivores. - Ecology 70: 1800- 1810.
Vickery, P. D., Hunter, M. L. and Wells, J. F. 1992. Evidence of incidental nest predation and its effects on nests of threatened grassland birds. - Oikos 63: 281-288.
Whelan, C. J.. Dilger, M. L., Robson, D. R., Hallyn. N. H. and Dilger, S. 1994. Effects of olfactory cues on artificial nest experiments. Auk 11 1: 945-952. -
Wilcove, D. S. 1985. Nest predation in forest tracts and the decline of migratory songbirds. - Ecology 66: 121 1 -1264.
Wilson, H. B., Hassell, M. P. and Godfray. H. C. J. 1996. Host-parasitoid food webs: dynamics. persistence, and in- vasion. - Am. Nat. 148: 787-806.
Zimmerman, J. L. 1984. Nest predation and its relationship to habitat and nest density in dickcissels. - Condor 86: 68-72.
Appendix A. Common and scientific names for avian species used in the text.
Ruffed grouse Bonasa urnhellus Woodcock Philohelu minor Whip-poor-will Caprimulgus oocijerus Black-billed cuckoo Coccyzus erythropthalmus Common crow Corcus anlericana Blue jay Cyanocitta crisrata Least flycatcher Empidonax tninimus Wood thrush Hylorichla rnustelina Veery Cathartis fusescerzs Northern cardinal Cardittalis ccrrdinalis Rose-breasted grosbeak Pheucticus ludocicianus Hooded warbler U'ilsoniu citrinn Kentucky warbler Oporornis ,forrnosus Ovenbird Seiurus aurocapillus Yellow-breasted chat Icteria cirens Red-eyed vireo Vireo olicaceus Common grackle Quiscalus quiscula Gray catbird Dumetellu carolinensis
OIKOS 87 1 0999)

You have printed the following article:
Nest Predation on Woodland Songbirds: When Is Nest Predation Density Dependent?Kenneth A. Schmidt; Christopher J. WhelanOikos, Vol. 87, No. 1. (Oct., 1999), pp. 65-74.Stable URL:
http://links.jstor.org/sici?sici=0030-1299%28199910%2987%3A1%3C65%3ANPOWSW%3E2.0.CO%3B2-8
This article references the following linked citations. If you are trying to access articles from anoff-campus location, you may be required to first logon via your library web site to access JSTOR. Pleasevisit your library's website or contact a librarian to learn about options for remote access to JSTOR.
References
Corvid Density and Nest Predation in Relation to Forest Fragmentation: A LandscapePerspectiveHenrik AndrénEcology, Vol. 73, No. 3. (Jun., 1992), pp. 794-804.Stable URL:
http://links.jstor.org/sici?sici=0012-9658%28199206%2973%3A3%3C794%3ACDANPI%3E2.0.CO%3B2-1
Factors Affecting Nesting Success in Riparian Bird CommunitiesLouis B. Best; F. StaufferThe Condor, Vol. 82, No. 2. (May, 1980), pp. 149-158.Stable URL:
http://links.jstor.org/sici?sici=0010-5422%28198005%2982%3A2%3C149%3AFANSIR%3E2.0.CO%3B2-J
Predation by Weasels (Mustela nivalis) on Breeding Tits (Parus Spp.) in Relation to theDensity of Tits and RodentsEuan DunnThe Journal of Animal Ecology, Vol. 46, No. 2. (Jun., 1977), pp. 633-652.Stable URL:
http://links.jstor.org/sici?sici=0021-8790%28197706%2946%3A2%3C633%3APBW%28NO%3E2.0.CO%3B2-5
http://www.jstor.org
LINKED CITATIONS- Page 1 of 5 -

Negative Correlation of Blue Jays and Golden-cheeked Warblers Near an Urbanizing AreaThomas M. Engels; Charles W. SextonConservation Biology, Vol. 8, No. 1. (Mar., 1994), pp. 286-290.Stable URL:
http://links.jstor.org/sici?sici=0888-8892%28199403%298%3A1%3C286%3ANCOBJA%3E2.0.CO%3B2-P
Anti-Predator Aggression in Birds Nesting in Old Field Habitats: An Experimental AnalysisBradley M. GottfriedThe Condor, Vol. 81, No. 3. (Aug., 1979), pp. 251-257.Stable URL:
http://links.jstor.org/sici?sici=0010-5422%28197908%2981%3A3%3C251%3AAAIBNI%3E2.0.CO%3B2-Y
Short-Term Apparent CompetitionRobert D. Holt; Burt P. KotlerThe American Naturalist, Vol. 130, No. 3. (Sep., 1987), pp. 412-430.Stable URL:
http://links.jstor.org/sici?sici=0003-0147%28198709%29130%3A3%3C412%3ASAC%3E2.0.CO%3B2-5
The Ecological Consequences of Shared Natural EnemiesR. D. Holt; J. H. LawtonAnnual Review of Ecology and Systematics, Vol. 25. (1994), pp. 495-520.Stable URL:
http://links.jstor.org/sici?sici=0066-4162%281994%2925%3C495%3ATECOSN%3E2.0.CO%3B2-S
New Approaches to the Analysis of Population Trends in Land BirdsFrances C. James; Charles E. McCulloch; David A. WiedenfeldEcology, Vol. 77, No. 1. (Jan., 1996), pp. 13-27.Stable URL:
http://links.jstor.org/sici?sici=0012-9658%28199601%2977%3A1%3C13%3ANATTAO%3E2.0.CO%3B2-2
Risk of Predation and the Structure of Desert Rodent CommunitiesBurt P. KotlerEcology, Vol. 65, No. 3. (Jun., 1984), pp. 689-701.Stable URL:
http://links.jstor.org/sici?sici=0012-9658%28198406%2965%3A3%3C689%3AROPATS%3E2.0.CO%3B2-C
http://www.jstor.org
LINKED CITATIONS- Page 2 of 5 -

On the Advantage of Being Different: Nest Predation and the Coexistence of Bird SpeciesThomas E. MartinProceedings of the National Academy of Sciences of the United States of America, Vol. 85, No. 7.(Apr. 1, 1988), pp. 2196-2199.Stable URL:
http://links.jstor.org/sici?sici=0027-8424%2819880401%2985%3A7%3C2196%3AOTAOBD%3E2.0.CO%3B2-B
Habitat and Area Effects on Forest Bird Assemblages: Is Nest Predation an Influence?Thomas E. MartinEcology, Vol. 69, No. 1. (Feb., 1988), pp. 74-84.Stable URL:
http://links.jstor.org/sici?sici=0012-9658%28198802%2969%3A1%3C74%3AHAAEOF%3E2.0.CO%3B2-O
Nest Predation and Nest SitesThomas E. MartinBioScience, Vol. 43, No. 8. (Sep., 1993), pp. 523-532.Stable URL:
http://links.jstor.org/sici?sici=0006-3568%28199309%2943%3A8%3C523%3ANPANS%3E2.0.CO%3B2-X
Does Predator Assemblage Affect Reproductive Success in Songbirds?Clinton K. Miller; Richard L. KnightThe Condor, Vol. 95, No. 3. (Aug., 1993), pp. 712-715.Stable URL:
http://links.jstor.org/sici?sici=0010-5422%28199308%2995%3A3%3C712%3ADPAARS%3E2.0.CO%3B2-F
Using Giving-Up Densities to Detect Search ImagesRobert A. Morgan; Joel S. BrownThe American Naturalist, Vol. 148, No. 6. (Dec., 1996), pp. 1059-1074.Stable URL:
http://links.jstor.org/sici?sici=0003-0147%28199612%29148%3A6%3C1059%3AUGDTDS%3E2.0.CO%3B2-R
Experimental Study of Predation on Eggs of Ground-Nesting Birds: Effects of Habitat andNest DistributionJaroslav PicmanThe Condor, Vol. 90, No. 1. (Feb., 1988), pp. 124-131.Stable URL:
http://links.jstor.org/sici?sici=0010-5422%28198802%2990%3A1%3C124%3AESOPOE%3E2.0.CO%3B2-3
http://www.jstor.org
LINKED CITATIONS- Page 3 of 5 -

Radiotelemetric Study of Activity and Movements of Racers (Coluber constrictor) Associatedwith a Carolina Bay in South CarolinaMichael V. Plummer; Justin D. CongdonCopeia, Vol. 1994, No. 1. (Feb. 1, 1994), pp. 20-26.Stable URL:
http://links.jstor.org/sici?sici=0045-8511%2819940201%293%3A1994%3A1%3C20%3ARSOAAM%3E2.0.CO%3B2-8
The Optimization of Growth Rate in Altricial BirdsRobert E. RicklefsEcology, Vol. 65, No. 5. (Oct., 1984), pp. 1602-1616.Stable URL:
http://links.jstor.org/sici?sici=0012-9658%28198410%2965%3A5%3C1602%3ATOOGRI%3E2.0.CO%3B2-T
Aspect Diversity in Moths: A Temperate-Tropical ComparisonRobert E. Ricklefs; Kevin O'RourkeEvolution, Vol. 29, No. 2. (Jun., 1975), pp. 313-324.Stable URL:
http://links.jstor.org/sici?sici=0014-3820%28197506%2929%3A2%3C313%3AADIMAT%3E2.0.CO%3B2-2
Patch Assessment in Fox Squirrels: The Role of Resource Density, Patch Size, and PatchBoundariesKenneth A. Schmidt; Joel S. BrownThe American Naturalist, Vol. 147, No. 3. (Mar., 1996), pp. 360-380.Stable URL:
http://links.jstor.org/sici?sici=0003-0147%28199603%29147%3A3%3C360%3APAIFST%3E2.0.CO%3B2-8
Predator-Mediated Interactions Between and Within Guilds of Nesting Songbirds:Experimental and Observational EvidenceKenneth A. Schmidt; Christopher J. WhelanThe American Naturalist, Vol. 152, No. 3. (Sep., 1998), pp. 393-402.Stable URL:
http://links.jstor.org/sici?sici=0003-0147%28199809%29152%3A3%3C393%3APIBAWG%3E2.0.CO%3B2-N
Predation, Competition, and Prey Communities: A Review of Field ExperimentsAndrew Sih; Philip Crowley; Mark McPeek; James Petranka; Kevin StrohmeierAnnual Review of Ecology and Systematics, Vol. 16. (1985), pp. 269-311.Stable URL:
http://links.jstor.org/sici?sici=0066-4162%281985%2916%3C269%3APCAPCA%3E2.0.CO%3B2-P
http://www.jstor.org
LINKED CITATIONS- Page 4 of 5 -

Measuring Patch Assessment Abilities of Desert GranivoresThomas J. Valone; Joel S. BrownEcology, Vol. 70, No. 6. (Dec., 1989), pp. 1800-1810.Stable URL:
http://links.jstor.org/sici?sici=0012-9658%28198912%2970%3A6%3C1800%3AMPAAOD%3E2.0.CO%3B2-D
Nest Predation in Forest Tracts and the Decline of Migratory SongbirdsDavid S. WilcoveEcology, Vol. 66, No. 4. (Aug., 1985), pp. 1211-1214.Stable URL:
http://links.jstor.org/sici?sici=0012-9658%28198508%2966%3A4%3C1211%3ANPIFTA%3E2.0.CO%3B2-M
Host-Parasitoid Food Webs: Dynamics, Persistence, and InvasionH. B. Wilson; M. P. Hassell; H. C. J. GodfrayThe American Naturalist, Vol. 148, No. 5. (Nov., 1996), pp. 787-806.Stable URL:
http://links.jstor.org/sici?sici=0003-0147%28199611%29148%3A5%3C787%3AHFWDPA%3E2.0.CO%3B2-W
Nest Predation and Its Relationship to Habitat and Nest Density in DickcisselsJohn L. ZimmermanThe Condor, Vol. 86, No. 1. (Feb., 1984), pp. 68-72.Stable URL:
http://links.jstor.org/sici?sici=0010-5422%28198402%2986%3A1%3C68%3ANPAIRT%3E2.0.CO%3B2-X
http://www.jstor.org
LINKED CITATIONS- Page 5 of 5 -




















