
Elephants, Ants and Superpowers: Nigeria's Relations with ...
Upload
independentCategory
view
1download
0
s
allynightined
ng time Thisdaytime
lesde
) dont laet leacutecheacuteesa journeacutee
C R Biologies 327 (2004) 509ndash517
Ethology Eacutethologie
Sugary food robbing in ants a case of temporal cleptobiosi
Freddie-Jeanne Richardabc Alain Dejeanalowast Jean-Paul Lachaudbd
a Laboratoire drsquoeacutevolution et diversiteacute biologique (CNRSndashUMR 5174) universiteacute Toulouse-3 118 route de Narbonne31062 Toulouse cedex France
b Centre de recherches sur la cognition animale (CNRSndashUMR 5169) universiteacute Toulouse-3 118 route de Narbonne31062 Toulouse cedex France
c Institut de recherches en biologie de lrsquoinsecte (CNRSndashUMR 6035) faculteacute des sciences et techniquesParc de Grandmont 37200 Tours France
d El Colegio de la Frontera Sur (ECOSUR) Apdo Postal 36 30700 Tapachula Chiapas Mexico
Received 8 July 2003 accepted 16 March 2004
Available online 13 May 2004
Presented by Claude Combes
Abstract
This study reports new information on interactions betweenEctatomma tuberculatum (Ponerinae) andCrematogaster limataparabiotica (Myrmicinae) Workers of these sympatric arboreal ant species forage on the same pioneer trees DiurnEctatomma preyed onCrematogaster workers that avoided overt aggression by respecting a lsquosafe distancersquo AtCrematogaster initiated raids within theEctatomma nests that they apparently left with their abdomen empty then remanear the nest entrances where they successfully intercepted 752 of the returningEctatomma foragers (N = 322) Certainintercepted workers rapidly resumed their return trip Others (391) were stopped explored and licked during a loby the Crematogaster Most of them were carrying between their mandibles a droplet of liquid food that was stolenrelationship that appears to be a typical case of interspecific cleptobiosis whose expression varies during thedemonstrates for the first time sugary-food robbing instead of prey robbing in antsTo cite this article F-J Richard etal C R Biologies 327 (2004) 2004 Acadeacutemie des sciences Published by Elsevier SAS All rights reserved
Reacutesumeacute
Vol drsquoaliments sucreacutes chez les fourmis un cas de cleptobiose temporelle Les ouvriegraveres drsquoEctatomma tuberculatum(Ponerinae) etCrematogaster limata parabiotica (Myrmicinae) fourragent sur les mecircmes arbres pionniers De jourCrematogaster qui eacutevitent toute interaction agressive directe de la part desEctatomma en respectant une laquo distanceseacutecuriteacute raquo peuvent leur servir de proie De nuit lesCrematogaster initient des raids agrave lrsquointeacuterieur des nids drsquoEctatomma enressortent avec lrsquoabdomen apparemment vide puis restent pregraves de lrsquoentreacutee du nid ougrave elles interceptent 752 desEctatommaqui rentrent (N = 322) Certaines de ces derniegraveres reprennent rapidement leur trajet de retour Les autres (391 plupart transportaient entre leurs mandibules une gouttelette de liquide sucreacute qui leur fut voleacutee ont eacuteteacute exploreacuteespendant de longues peacuteriodes Cette cleptobiose interspeacutecifique dont lrsquoexpression varie en fonction du moment de l
Corresponding authorE-mail address dejeancictfr (A Dejean)
1631-0691$ ndash see front matter 2004 Acadeacutemie des sciences Published by Elsevier SAS All rights reserveddoi101016jcrvi200403002
510 F-J Richard et al C R Biologies 327 (2004) 509ndash517
permet de deacutecrire pour la premiegravere fois lrsquoexistence de vols drsquoaliments sucreacutes et non de proies chez des fourmisPour citer cetarticle F-J Richard et al C R Biologies 327 (2004) 2004 Acadeacutemie des sciences Published by Elsevier SAS All rights reserved
Keywords ants cleptobiosis interspecificrelationships French GuianaCrematogaster limata parabiotica Ectatomma tuberculatum
Mots-cleacutes fourmis cleptobiose interactions heacuteteacuterospeacutecifiques Guyane Crematogaster limata parabiotica Ectatomma tuberculatum
lep-ali-cesfeacute-deresles
types leom-
di-le
tioneses-
esesciteacutede
aitsess en
ia-a
tre-artbre
us-es dour-
ra-ion
r
sns)nt
flo-des
exis-ian-ten-do-
elaisauxou-des
cestam-
rbresrale-
acelles
et
ex-resp-auxtre
ndant
-
s les
Version franccedilaise abreacutegeacutee
La cleptobiose (eacutegalement connue comme laquo ktoparasitisme raquo ou laquo piraterie raquo) concerne le vol drsquoments entre individus de mecircme espegravece ou drsquoespegravediffeacuterentes Cette forme de compeacutetition par interrence constitue un moyen important par lequelnombreuses espegraveces parviennent agrave obtenir dessources alimentaires limiteacutees tout en reacuteduisantcoucircts lieacutes agrave leur recherche et agrave leur obtention Cede strateacutegie alimentaire largement reacutepandu danmonde animal et abondamment deacutecrit chez de nbreux verteacutebreacutes a eacutegalement eacuteteacute rapporteacute chezvers inverteacutebreacutes et notamment des fourmis ougravecas de la poneacuterine neacuteotropicaleEctatomma ruidumest un des exemples le mieux eacutetudieacute Lrsquoobservafortuite au Panama il y a une quinzaine drsquoanneacutedrsquoune relation de type parasitique entre une autrepegravece drsquoEctatomma (E tuberculatum) et la petite myr-micine Crematogaster limata au cours de laquellles Crematogaster peacuteneacutetraient agrave lrsquointeacuterieur des niddrsquoEctatomma et grimpaient sur le dos des ouvriegraverpour les leacutecher sans subir drsquoagression nous a inagrave eacutetudier de plus pregraves ce genre drsquoassociation afinveacuterifier si une relation de type cleptobiotique nrsquoeacutetpas en jeu et drsquoen deacutetailler les diffeacuterentes phaLes observations reacutealiseacutees en saison des pluieGuyane (Petit Saut) ont concerneacute 36 nids drsquoE tu-berculatum dont au moins 10 preacutesentaient lrsquoassoction avecCrematogaster La majoriteacute des donneacuteeseacuteteacute collecteacutee sur le nid le plus actif Les enregisments ont porteacute sur divers paramegravetres lieacutes drsquoune pagrave lrsquoactiviteacute de fourragement des deux espegraveces (nomde fourrageuses drsquoEctatomma sortant et entrant dnid flux journalier deCrematogaster pregraves des entreacuteedes nids drsquoEctatomma rythme journalier drsquoexploitation des sources sucreacutees produites par des nymphmembracides taux comparatif de preacutesence des fmis au niveau des nectaires extrafloraux drsquoInga thi-baudiana et sur 30 groupes de nymphes de Membcidae) et drsquoautre part agrave leurs interactions (proport
-
e
de fourrageuses drsquoEctatomma arrecircteacutees lors du retouau nid nombre drsquoouvriegraveres deCrematogaster explo-rant les ouvriegraveres drsquoEctatomma et parties du corpconcerneacutees par lrsquoexploration dureacutee des interactioBien que preacutesentant un pic drsquoactiviteacute plus importaen peacuteriode nocturneE tuberculatum est active toutela journeacutee et exploite en continu les nectaires extraraux et les membracides Comme chez la plupartautres poneacuterines les eacutechanges trophallactiques nrsquotent pas chezE tuberculatum et les fourrageuses qureacutecoltent des liquides les transportent entre leurs mdibules (ces liquides adhegraverent gracircce aux forces desion superficielle) avant de les redistribuer par pseutrophallaxie agrave drsquoautres fourrageuses assurant un rdans le transport (299 des cas) ou directementcongeacutenegraveres agrave lrsquointeacuterieur du nid Jusqursquoagrave 618 desvriegraveres rentrant au nid transportent ainsi des liquisucreacutes (N = 377) Sur les sites alimentairesE tuber-culatum rentre en compeacutetition avec drsquoautres espegravede fourmis pour la reacutecolte des sources sucreacutees noment avec lesC limata parabiotica qui exploitent lesmecircmes espegraveces de membracides sur les mecircmes aet durant les mecircmes creacuteneaux horaires mais geacuteneacutement sur des branches diffeacuterentesEctatomma tuber-culatum est connue pour ecirctre une preacutedatrice efficdrsquoun grand nombre drsquoautres fourmis parmi lesquele genreCrematogaster est largement repreacutesenteacute Djour lesC limata parabiotica eacutevitent manifestementout contact direct avec lesEctatomma ndash utilisant lesmecircmes heacutemiptegraveres mais en alternance ou bienploitant les individus les plus eacuteloigneacutes des ouvriegravedrsquoEctatomma et srsquoeacutecartant degraves que lrsquoune drsquoelles se raproche ndash et servent mecircme freacutequemment de proiesEctatomma Les interactions comportementales enles deux espegraveces changent de faccedilon drastique pela nuit lrsquoagressiviteacute normalement preacutesenteacutee par lesEc-tatomma vis-agrave-vis desCrematogaster disparaissant totalement LesCrematogaster nrsquoeacutevitent plus alors lecontact avec lesEctatomma et mecircme lrsquoinitient lrsquoin-teraction se reacutealisant en deux phases distinctes Danun premier temps en deacutebut de peacuteriode nocturne
F-J Richard et al C R Biologies 327 (2004) 509ndash517 511
eacute-plusres-rra-nm-
pregraveseurecircts)
ettequ
s ndashter-er-aire- De
la
srsquoapur-esantdui-ar-
aseagecu-si-
de li
-id-eadntre-andntes
-ates
st
se
nt
ily
ic-
nd
n
and-
y one-
d
nathe
e
-ib-o-
unktun-
instthers
ityour-identctic
s ofsion
ver-
Crematogaster effectuent de veacuteritables raids agrave lrsquointrieur des nids de la poneacuterine Elles en ressortenttard sans avoir apparemment voleacute de nourrituretent pregraves de lrsquoentreacutee et se mettent agrave arrecircter les fougeuses drsquoEctatomma lors de leur retour au nid tout efaisant vibrer leur abdomen recrutant ainsi de nobreuses congeacutenegraveres Sur 322 ouvriegraveres drsquoEctatommarentrant ainsi au nid 752 ont eacuteteacute arrecircteacutees dontde la moitieacute reprend rapidement le chemin vers lnid Dans tous les autres cas (126 sur les 242 arrles petites ouvriegraveres deCrematogaster grimpent sur ledos de la poneacuterine la legravechent et lorsqursquoune gouttelde miellat est preacutesente entre ses mandibules ndash ceest le cas pour 881 des ouvriegraveres ainsi exploreacuteela volent Ces interactions de type laquo cleptobiose inspeacutecifique raquo preacutesentent ainsi lrsquooriginaliteacute de concner des aliments sucreacutes et non des proies contrment aux autres cas rapporteacutes dans la litteacuteratureplus son expression est temporelle car limiteacutee agravepeacuteriode nocturne et facultative lesCrematogaster nedeacutependant pas strictement de cette strateacutegie pourprovisionner Cette temporaliteacute du pheacutenomegravene porait srsquoexpliquer par la forte augmentation drsquoactiviteacute dEctatomma en deacutebut de peacuteriode nocturne provoquune grande confusion agrave lrsquoentreacutee des nids ce qui reacuterait ou mecircme annulerait le filtrage reacutealiseacute par les gdiennes et autoriserait ainsi les raids deCrematogasteravec un minimum de risques drsquoagression Cette phpermettrait agrave ces derniegraveres drsquoacqueacuterir un camouflchimique par absorption passive drsquohydrocarbonesticulaires camouflage qui serait par la suite intenfieacute lors des leacutechages des fourrageuses chargeacuteesquides sucreacutes
1 Introduction
Cleptobiosis kleptoparasitism food robbing thievery and piracy concern the stealing of food by indivuals of the same or different species This widesprform of interference competition is an importameans by which many animals obtain limitedsources while reducing the costs of searching forhandling food[1ndash3] This foraging strategy has beeextensively described for a large range of vertebraincluding fishes[45] birds[267] and mammals[89] Numerous cases of intra- and interspecific cleptobiosis have also been reported in marine invertebr[10] spiders[1112] and insects such as thrips[13]
i
-
-
wasps[14] bees[15] and ants In ants since the firdescriptions by Wroughton in 1892 (in[16]) and thefirst use of the term lsquocleptobiosisrsquo in its current senby Forel[17] numerous cases have been reported[18ndash21] particularly that of the Neotropical ponerine aEctatomma ruidum [22ndash27]
Aiming to further the understanding of the daforaging rhythms ofEctatomma tuberculatum Whe-eler [28] provided a fortuitous account of a parasitlike association between this species andCremato-gaster limata She observed particularly at dawn adusk that hundreds ofC limata file into the nestentrances ofE tuberculatum When encountering aE tuberculatum worker the smallerC limata climbedup one of the larger antrsquos legs and onto its thoraxhead while theE tuberculatum did not react aggressively to the intruders As Wheeler[28] did not noteany aggressiveness we decided to conduct a studthis relationship hypothesizing that the intriguing bhaviour of theCrematogaster workers could be relateto cleptobiosis
2 Materials and methods
Data were collected in Petit Saut French Guiamostly in July 2000 The mean temperature duringdaytime was 285C falling to 228C at night Themean relative humidity was 85ndash90 during daytimall throughout the periods of observation
Ectatomma tuberculatum is a monogynous (or occasionally polygynous) ponerine ant widely distruted in Neotropical regions Colonies have polydmous nests excavated at the base of a tree trNest entrances are characterized by an externalnel (made with rough vegetal fibres) leaning agathe base of the supporting tree The opening oftunnel is generally guarded by specialized workeWorkers concentrate most of their foraging activon the tree situated above their nest plus neighbing trees and shrubs They solitarily gather a wvariety of small invertebrates and exploit differesugary sources As for most Ponerinae trophallaexchanges cannot occur inEctatomma So foragerscarry between their outstretched mandibles dropletliquid substances that adhere thanks to surface tenstrengths[2228ndash33]
We noted the position of 36Ectatomma neststhanks to their characteristic entrance tunnels Ne
512 F-J Richard et al C R Biologies 327 (2004) 509ndash517
ari-er-hathad
a-c--
0-ad weingux
talthe50
ofans
asne ond100
iednd
by
st
ura-
00
nest
thesed
tedrted
t
r ofes
e
ctar
al
-ndst
theless due to problems of accessibility and great vation in the coloniesrsquo levels of activity detailed obsvations were focused on the most active colony thad four nest entrances Ten out of the 36 nestsrelationships withCrematogaster limata parabioticaa myrmicine species very frequent in pioneer formtions with polydomous nests built in hollowed strutures or ant gardens[3435] The difference in size between the species was great (meanEctatomma workerlengthplusmn SE 95 plusmn 08 mmN = 20 Crematogaster35plusmn 006 mmN = 20)
As the Ectatomma circadian rhythm of activityvaries seasonally[31] we conducted a series of 1min scans over an 8-day-long period until we hcovered each hour out of the 24 During each scanchecked the number of workers entering and leavthe nest of one colony We also estimated the flof Crematogaster workers around theEctatomma nestentrances by using a grid ranging from 0 to 4 (0 toabsence of workers 1 one or two workers nearnest entrance 2 up to ten workers 3 10 toworkers 4 more than 50 workers)
We also studied the rhythm of the exploitationsugary sources Firstly we conducted 10-min scper hour concerningEctatomma workers attendingmembracid nymphs (Hemiptera) on a small treewell as Crematogaster workers on several sites ithe area where we also registered the presencEctatomma (six 3-h time periods between 0800 a0200) Secondly we noted the presence of ants onleaves of 20 juvenileInga thibaudiana (Mimosoideae)and 30 groups of membracid nymphs in the studarea during three periods at night in the morning ain the afternoon
We quantified the cleptobiotic events performedCrematogaster workers through the proportions ofEc-tatomma foragers stopped while returning to their nethe number ofCrematogaster individuals exploringan Ectatomma worker the body part concerned (iemouthparts legs abdomen and thorax) and the dtion of the interaction
3 Results
31 Circadian rhythm of activity
Although continuously activeEctatomma workerspresented two peaks of activity at night around 20
f
Fig 1 (A) Circadian rhythm of activity ofEctatomma tuberculatumthrough the mean number of workers entering and exiting theper 1-h time periods (meanplusmn SE) (B) Estimation of the flux ofCre-matogaster limata parabiotica workers at theE tuberculatum nestentrance by using a grid ranging from 0 (absence ofCrematogasterworkers) to 4 (more than 100Crematogaster workers)
and 0300 Nocturnally between 1900 and 0500number of workers returning to their nest surpasthat of workers exiting the nest (Fig 1A) During acontinuous 24-h-long observation period we nothat 618 of the workers entering the nest transposugary liquids between their mandibles (N = 377) Onthe foraging sitesEctatomma competed with other anspecies includingCrematogaster for the exploitationof membracid honeydew and the extrafloral nectaInga In spite of a reduced level of activity in thmorning Ectatomma workers attended membracidall day long (Fig 2A and C) The nocturnal increasin the number of ants exploitingInga extrafloral nectarcorresponds to the increase in the production of neat night[36]
Crematogaster workers also displayed both diurnand nocturnal activity on the foraging sites (Fig 2BndashD) Around theEctatomma nest entrances their activity level was high at night mostly between 2100 a0400 whenEctatomma foragers returned to their ne(Fig 1B)
F-J Richard et al C R Biologies 327 (2004) 509ndash517 513
fntsg
Fig 2 (A) Dynamics of the exploitation of membracid nymphs byEctatomma tuberculatum workers at one site during a 24-h cycle (B) Thesame forCrematogaster limata parabiotica workers recorded at different sites between 0800 and 0200 (C) Percentages of groups omembracid nymphs attended by ants during three periods of the day (other speciesCamponotus spp 100 of the groups attended by aat 1000 and 2300 933 at 1500 hours) (D) Percentages of leaves ofInga thibaudiana (Mimosoideae) noted with ant workers exploitinthe extrafloral nectaries during three periods of the day (other species mostlyCamponotus spp but also someGigantiops destructor andCephalotes sp in the morning total of 39 7 and 72 of the leaves with ants at 1000 1500 and 2300 respectively)
theen-me
t ofm
the
to- Al-tedof
ps
cidve-eytlytothsedof
eenypec-
tledewr fortheion
32 Ants attending membracids
In the studied area bothCrematogaster and Ec-tatomma foragers attended membracid nymphs onsame trees during the same time periods but gerally on different branches They shared the saparts of the branches on only 25 occasions ou194 noted during the study of their circadian rhythof activity In such cases theCrematogaster attendedthe membracid individuals the most distant fromEctatomma foragers and they moved when anEc-tatomma approached The latter frequently triedcapture theCrematogaster workers 15 attempts occurred on the 25 branches shared by both speciesthough most of them successfully escaped we noduring a series of observations that 14 prey out27 retrieved byE tuberculatum foragers were ants10 of themCrematogaster Also four retrieved preywere membracids originating from attended groutwo complete predatory sequences being observed
During trophobiotic relationshipsEctatommaworkers palpated the lateral parts of the membranymphs with their antennae in an up and down moment eliciting the secretion of honeydew As thstood above the nymphs with their mandibles slighopen they recuperated the honeydew that adheredtheir mandibles thanks to surface tension strengWhile repeating this behaviour the workers opentheir mandibles wider and wider as the dropletshoneydew increased in size (Fig 3A) Then in 701of the cases (N = 127) eachEctatomma forager re-trieved the droplet of honeydew accumulated betwits mandibles after having been replaced or not ba nestmate (252 and 449 of the cases restively) In the other situations (299)Ectatommaworkers specialized in honeydew transport shutbetween the individuals that collected the honeydfrom the membracids and the nest (one transporte4ndash7 collectors) The transfer of honeydew fromcollector to the transporter followed an antennat
514 F-J Richard et al C R Biologies 327 (2004) 509ndash517
ed
Fig 3 (A) Ectatomma tuberculatum worker attending membracid nymphs and gathering a honeydew droplet between its outstretchmandibles (B) E tuberculatum worker stopped by twoCrematogaster limata parabiotica workers inspecting its mouthparts (C) Workerof C limata parabiotica licking a part of the honeydew droplet from aE tuberculatum forager
s ofenidsItewen
ab-
n aredthe
ost-p
nest
re-spedrs at-idly
initiated by the transporter that also made a serie3ndash4 back and forth movements with its body Ththe collecting worker again attended the membracwhile the transporter looked for other collectorsreturned to the nest when the droplet of honeydcompletely filled the space separating its wide-opmandibles
33 Relationships between Crematogaster limataparabioticaand Ectatomma tuberculatum
While completely absent in the vicinity of theEc-tatomma nest entrances during the daytime (Fig 1B)at duskCrematogaster workers entered theEctatommanests without any opposition as the guards weresent As already reported[28] the column ofCre-
matogaster passed through the entrance tunnels iraid-like fashion The beginning of such raids occurbetween 1800 and 2245 which corresponds topeak of nocturnal activity ofEctatomma (Fig 1A) Af-ter these raids theCrematogaster workers exited thetunnel and remained around the nest entrance almall night long (Fig 1B) They never had a swollen abdomen nor any evidence of having filled their croThen they stopped returningEctatomma foragers atthe base of the supporting tree trunk around theentrance or on surrounding branchesCrematogasterworkers waved their abdomens at the approach of aturningEctatomma forager but no liquid droplet wanoted at the tip of their abdomens They also taptheir abdomens on the substrate These behavioutracted nestmates that immediately arrived and rap
F-J Richard et al C R Biologies 327 (2004) 509ndash517 515
ed
80
42r-ringhereng
id
-
)
e-sed
ual
eto
dies
to or
hentter
heer-
n
mersquoy
edn-
fniv-hethe
din-
-reted
y-t
histheur
d-
on-il-
gr
tsr anri-
c-
cal-l or
aggregated around theEctatomma CertainCremato-gaster individuals stayed in front of theEctatommathat remained immobile while up to 12 others climbon theEctatomma (32plusmn 24 N = 98)
A study conducted on 322Ectatomma foragers re-turning to their nest permitted us to note that only(248) were not intercepted by theCrematogasterworkers prior to the tunnel entrance Among the 2Ectatomma intercepted 116 (36 of all returning foagers) were stopped and remained immobile dua short time before resuming their return trip to tnest The remaining 126 individuals (391) westopped explored and licked during a relatively lotime (gt1 min) by theCrematogaster workers Thisincluded 111 individuals carrying a droplet of liqufood that was stolen (345)
Of the 316Crematogaster workers thoroughly observed after the interception of anEctatomma for-ager laden with a droplet of liquid food 165 (522focused their exploration on theEctatomma mouth-parts (seeFig 3B and C) Each individual lickedthe honeydew only during a few seconds (66 plusmn54 s N = 70) When the honeydew droplet was dpleted theEctatomma workers remained motionlesmandibles open maxilo-labial mouthparts extendallowing theCrematogaster workers to lick them dur-ing a long time (25plusmn 145 minN = 15)
4 Discussion
We confirm therefore the existence of the unusrelationship betweenE tuberculatum andC limata asreported in central Panama[28] Nevertheless somnew information and significant differences deservebe pointed out
First Ectatomma workers constantly attendegroups of membracid nymphs thanks to two strategWorkers collected honeydew that they transportedthe nest once they were replaced by a nestmatecertain workers collected the honeydew that was ttransported to the nest by other workers The lacooperative behaviour known in the Formicinae[3]presents similarities with the lsquorelay transportrsquo and tlsquoprey chain transfer behaviourrsquo noted for other ponine ants during predation[3738]
Second we noted a dramatic difference inCremato-gaster-Ectatomma interspecific relationships betwee
day and night Although sharing branches with theCrematogaster workers maintained a lsquosafe distancfrom Ectatomma in the foraging areas during the dathe latter being able to prey on them (see also[33])On the contrary at night theCrematogaster work-ers no longer avoided direct contact withE tubercu-latum but initiated the interactions They conducttheir raids on theEctatomma nest entrances then egaged in contact with theEctatomma foragers return-ing to their nest stopping and exploring them
Third Wheeler[28] noted that the behaviour oCrematogaster workers during raids was limited itime and consisted of a kind of lsquoshampooingrsquo dering nutrition from the cuticular secretions andor texcess fluids (nectar and honeydew) trapped onmandibles ofEctatomma workers This author notethat such behaviour first described between anquiline ant and its host[39] could account for a mutualistic association of the lsquocleaner-fishrsquo type wheboth species derive benefits Even if we again nothat Crematogaster workers licked the different bodparts of theEctatomma they mostly stopped and explored theEctatomma retrieving a honeydew drople(881 of the exploredE tuberculatum foragers) andrecruited nestmates in order to rob this droplet Tresulted therefore in a parasitic relationship oflsquocleptobioticrsquo form This nocturnal robbing behavioonly complements the food supply of theCremato-gaster that gather most of their sugary food by attening hemiptera or exploiting extrafloral nectaries
Fourth other cases of cleptobiosis in ants ccerned prey robbing with nevertheless a certain simitude with the present study For exampleMyrmeco-cystus mimicus workers thoroughly inspect returninforagers ofPogonomyrmex spp before robbing theiprey [20] The originality of the cleptobiotic activityof C limata parabiotica consists in exploiting the kindof liquid transport limited to arboreal ponerine anIndeed they carry droplets of liquid between theimandibles which are then imbibed by nestmatesadaptation to the lack of trophallaxis in most Ponenae[4041]
Finally we questioned the origin of the lower noturnal aggressiveness ofEctatomma workers towardsCrematogaster The immobility of the Ectatommaworkers could be triggered by a secreted chemiwhile the Crematogaster wave their abdomens Indeed secretions from the mandibular metapleura
516 F-J Richard et al C R Biologies 327 (2004) 509ndash517
d
id-
plet
he
ur-
ack
at-o-be
p-ac-p-
ire
-oure
nebio-hem
sisceso-e
ACa--the
entGIFs raquo
ds
ty
lm
a-bo5ndash
c02)
man
rrsquos
of-im
ter-star
ction9)
n-
02)
of82
iest
ar-
nd0
Dufour glands act as repellents in severalCremato-gaster species[42ndash44] and such an effect was notein the lestobioticSolenopsis fugax [45] Similarly apacifying action on the host occurs when an indivual of the parasiticFormicoxenus provancheri raisesits abdomen and extrudes the sting producing a droof volatile liquid before engaging in lsquoshampooingrsquo be-haviour withMyrmica incompleta [46]
Nevertheless why does this not work during tdaytime As theCrematogaster lsquoraidsrsquo coincide withthe dusk peak in activity ofEctatomma (1900ndash2100Fig 1A) another hypothesis can be formulated Ding this period the heavy flow ofEctatomma foragersentering and leaving the tunnel drives the guards binto the nest This permits the smallCrematogasterworkers to creep inside the tunnel without beingtacked In this situation they can modify the compsition of their cuticular hydrocarbons enough toaccepted by the nextEctatomma foragers returningto their nest and so to lure them A similar decetive mechanism based on chemical camouflagequisition was proposed to explain intraspecific cletobiosis inE ruidum [27] Later newCrematogasterindividuals can enter the tunnel or they can acqucuticular compounds directly from an interceptedEc-tatomma worker by climbing on it and licking its cuticle this corresponding to the lsquoshampooingrsquo behavidescribed by Wheeler[28] This hypothesis might bsupported by the fact thatC limata parabiotica has theability to share nests and trails with different formiciand ponerine ants in a phenomenon called lsquoparasisrsquo while these ants never share nests between t[343546]
In conclusion we noted a new kind of cleptobioin ants concerning the robbing of sugary substanand not prey robbing as previously known Temprally limited to night time and facultative it implies thpresence ofEctatomma nests onCrematogaster terri-tories
Acknowledgements
We are grateful to Dr Jacques HC Delabie (UPLaboratoacuterio de Mirmecologia Convecircnio UESCEPLAC Itabuna Bahia Brazil) for the identifiction of the ants and to AndreaDejean for proofreading the manuscript This work was supported by
French laquo Ministegravere de lrsquoEacutecologie et du Deacuteveloppemdurable raquo (SOFT program research agreementECOFOR No 98 and the laquo Tropical Ecosystemprogram No 02-E2002)
References
[1] E Curio in The Ethology ofPredation Springer-VerlagBerlin 1976 242 p
[2] HJ Brockmann CJ Barnard Kleptoparasitism in birAnim Behav 27 (1979) 487ndash514
[3] B Houmllldobler EO Wilson in The Ants Harvard UniversiPress Cambridge 1990 732 p
[4] TJ Pitcher Functions of shoaling behaviour in teleosts inJ Pitcher (Ed) The Behaviour of Teleost Fishes Croom HeLondon 1986 pp 294ndash337
[5] WJ Dominey AM Snyder Kleptoparasitism of freshwter crabs by cichlid fishes endemic to Lake Barombi MCameroon West Africa Environ Biol Fishes 22 (1988) 15160
[6] JR Garrido CG Sarasa M Fernandez-Cruz Intraspecifikleptoparasitism in the cattle egret J Field Ornithol 73 (20185ndash190
[7] CE Khatchikian M Favero AI Vassalo Kleptoparasitisby brown-hooded gulls and grey-hooded gulls on Americoystercatchers Waterbirds 25 (2002) 137ndash141
[8] MS Di Bitetti CH Janson Social foraging and the findeshare in capuchin monkeysCebus apella Anim Behav 62(2001) 47ndash56
[9] OP Honer B Wachter ML East H Hofer The responsespotted hyenas to long-term changes in prey populations functional response and interspecific kleptoparasitism J AnEcol 71 (2002) 236ndash246
[10] S Morissette JH Himmelman Subtidal food thieves inactions of four invertabrate kleptoparasites with the seaLeptasterias polaris Anim Behav 60 (2000) 531ndash543
[11] Y Heacutenaut J-P Lachaud Araigneacutees cleptoparasites seacutelede lrsquohocircte par sa toile Ann Soc Entomol Fr (NS) 35 (199148ndash153
[12] M Whitehouse I Agnarsson T Miyashita D Smith K Cagialosi T Masumoto D Li Y HeacutenautArgyrodes phylogenysociality and interspecific interactions J Arachnol 30 (20238ndash245
[13] B Crespi P Abbot The behavioral ecology and evolutionkleptoparasitism in Australian gall thrips Fla Entomol(1999) 147ndash164
[14] FE Kurczewski MG Spofford Alternative nesting strategin Ammophila urnaria (Hymenoptera Sphecidae) J NaHist 32 (1998) 99ndash106
[15] C Michener in The Social Behavior of the Bees A Compative Study The Belknap Pressof Harvard University PressCambridge 1974 404 p
[16] WM Wheeler in Ants Their Structure Development aBehaviour Columbia University Press New York 191663 p
F-J Richard et al C R Biologies 327 (2004) 509ndash517 517
)
in al 20
ing
89ndash
mic
ry
d)
n andash
5ndash
verinendash
hes
n
Ac-
ns
D
-
reyerae)
s
la-theera
c-eaery
n
ri-t 35
on
es-
ra
m
nta-
89)
de-
re
-
es
ific40
[17] A Forel Fourmis termitophages LestobioseAtta tardigradasous-genres drsquoEuponera Ann Soc Entomol Belg 45 (1901389ndash398
[18] T Abe On the food sharing among four species of antssandy grassland I Food and foraging behavior Jap J Eco(1971) 219ndash230
[19] U Maschwitz M MuhlenbergCamponotus rufoglaucus einewegelagernde Ameise Zool Anz 191 (1973) 364ndash368
[20] B Houmllldobler Food robbing inants a form of interferencecompetition Oecologia 69 (1986) 12ndash15
[21] T Yamaguchi Intraspecific competition through food robbin the harvester antMessor aciculatus (Fr Smith) and itsconsequences on colony survival Insectes Soc 42 (1995)101
[22] NA Weber Two common ponerine ants of possible econosignificanceEctatomma tuberculatum (Olivier) andE ruidumRoger Proc Entomol Soc Washington 48 (1946) 1ndash16
[23] MD Breed P Abel TJ Bleuze SE Denton Thievehome ranges and nestmate recognition inEctatomma ruidumOecologia 84 (1990) 117ndash121
[24] MD Breed LE Snyder TL Lynn JA Morhart Acquirechemical camouflage in a tropical ant Anim Behav 44 (1992519ndash523
[25] MD Breed TP McGlynn EM Stocker ANKlein Thiefworkers and variation in nestmate recognition behavior iponerine antEctatomma ruidum Insectes Soc 46 (1999) 327331
[26] I Perfecto JH Vandermeer Cleptobiosis in the antEc-tatomma ruidum in Nicaragua Insectes Soc 40 (1993) 29299
[27] JM Jeral MD Breed BE Hibbard Thief ants hareduced quantities of cuticular compounds in a poneant Ectatomma ruidum Physiol Entomol 22 (1997) 207211
[28] DE WheelerEctatomma tuberculatum foraging biology andassociation withCrematogaster (Hymenoptera Formicidae)Ann Entomol Soc Am 79 (1986) 300ndash303
[29] WL Jr Brown Contributions toward a reclassification of tFormicidae II Tribe Ectatommini (Hymenoptera) Bull MuComp Zool 118 (1958) 173ndash362
[30] A Dejean J-P Lachaud Growth-related changes in predatiobehavior in incipient colonies of the ponerine antEctatommatuberculatum (Olivier) Insectes Soc 39 (1992) 129ndash143
[31] J Valenzuela-Gonzaacutelez A Loacutepez-Meacutendez J-P Lachaudtivity patterns and foraging activity in nests ofEctatommatuberculatum (Hymenoptera Formicidae) in cacao plantatioSouthwest Entomol 20 (1995) 507ndash515
[32] RR da Hora R Feacuteneacuteron J Valenzuela ME FavilaFresneau Queen-size dimorphism in the antEctatomma
tuberculatum (Hymenoptera Formicidae Ponerinae) Sociobiology 38 (2001) 407ndash420
[33] G Ibarra-Nuacutentildeez JA Garciacutea JA Loacutepez J-P Lachaud Panalysis in the diet of some ponerine ants (HymenoptFormicidae) and web-building spiders (Araneae) in coffeplantations in Chiapas Mexico Sociobiology 37 (2001723ndash755
[34] DW Davidson Ecological studies of Neotropical ant gardenEcology 69 (1988) 1138ndash1152
[35] A Dejean B Corbara J Orivel RR Snelling JHC Debie M Belin-Depoux The importance of ant gardens inpioneer vegetal formations of French Guiana (HymenoptFormicidae) Sociobiology 35 (2000) 425ndash439
[36] LM Pascal EF Motte-Florac DB McKey Secretory strutures on the leaf rachis of Caesalpinieae and Mimosoid(Leguminosae) implication for the evolution of nectaglands Am J Bot 87 (2000) 327ndash338
[37] B Schatz J-P Lachaud G Beugnon Polyethism withihunters of the ponerine antEctatomma ruidum InsectesSoc 43 (1996) 111ndash118
[38] WM Wheeler The compound and mixed nests of Amecan ants Part I Observations on a new guest ant Am Na(1901) 431ndash448
[39] B Houmllldobler Liquid food transmission and antennatisignals in ponerine ants Isr J Entomol 19 (1985) 89ndash99
[40] J Liebig J Heinze B Houmllldobler Trophallaxis and aggrsion in the ponerine antPonera coartata implications forthe evolution of liquid food exchange in the HymenopteEthology 103 (1997) 707ndash722
[41] C Bevan A Birch H Caswell Ant insect repellent froblack cocktail ants J Chem Soc 961 (1961) 488
[42] AB Attygalle B Siegel O Vostrowsky HJ BestmanU Maschwitz Chemical composition and function of mepleural gland secretion of the antCrematogaster deformisSmith (Hymenoptera Myrmicinae) J Chem Ecol 15 (19317ndash328
[43] D Daloze M Kaisin C Detrain JM Pasteels Chemicalfense in the three European species ofCrematogaster antsExperientia 47 (1991) 1082ndash1089
[44] B Houmllldobler Chemische Strategie beim Nahrungserwerb deDiebameise (Solenopsis fugax Latr) und der Pharaoameis(Monomorium pharaonis L) Oecologia 11 (1973) 371ndash380
[45] A Lenoir C Errard A Francoeur R Loiselle Relations entre la fourmi parasiteFormicoxenus provancheri et son hocircteMyrmica incompleta Donneacutees biologiques et eacutethologiqu(Hym Formicidae) Insectes Soc 39 (1992) 81ndash97
[46] J Orivel C Errard A Dejean Ant gardens interspecrecognition in parabiotic species Behav Ecol Sociobiol(1997) 87ndash93

510 F-J Richard et al C R Biologies 327 (2004) 509ndash517
permet de deacutecrire pour la premiegravere fois lrsquoexistence de vols drsquoaliments sucreacutes et non de proies chez des fourmisPour citer cetarticle F-J Richard et al C R Biologies 327 (2004) 2004 Acadeacutemie des sciences Published by Elsevier SAS All rights reserved
Keywords ants cleptobiosis interspecificrelationships French GuianaCrematogaster limata parabiotica Ectatomma tuberculatum
Mots-cleacutes fourmis cleptobiose interactions heacuteteacuterospeacutecifiques Guyane Crematogaster limata parabiotica Ectatomma tuberculatum
lep-ali-cesfeacute-deresles
types leom-
di-le
tioneses-
esesciteacutede
aitsess en
ia-a
tre-artbre
us-es dour-
ra-ion
r
sns)nt
flo-des
exis-ian-ten-do-
elaisauxou-des
cestam-
rbresrale-
acelles
et
ex-resp-auxtre
ndant
-
s les
Version franccedilaise abreacutegeacutee
La cleptobiose (eacutegalement connue comme laquo ktoparasitisme raquo ou laquo piraterie raquo) concerne le vol drsquoments entre individus de mecircme espegravece ou drsquoespegravediffeacuterentes Cette forme de compeacutetition par interrence constitue un moyen important par lequelnombreuses espegraveces parviennent agrave obtenir dessources alimentaires limiteacutees tout en reacuteduisantcoucircts lieacutes agrave leur recherche et agrave leur obtention Cede strateacutegie alimentaire largement reacutepandu danmonde animal et abondamment deacutecrit chez de nbreux verteacutebreacutes a eacutegalement eacuteteacute rapporteacute chezvers inverteacutebreacutes et notamment des fourmis ougravecas de la poneacuterine neacuteotropicaleEctatomma ruidumest un des exemples le mieux eacutetudieacute Lrsquoobservafortuite au Panama il y a une quinzaine drsquoanneacutedrsquoune relation de type parasitique entre une autrepegravece drsquoEctatomma (E tuberculatum) et la petite myr-micine Crematogaster limata au cours de laquellles Crematogaster peacuteneacutetraient agrave lrsquointeacuterieur des niddrsquoEctatomma et grimpaient sur le dos des ouvriegraverpour les leacutecher sans subir drsquoagression nous a inagrave eacutetudier de plus pregraves ce genre drsquoassociation afinveacuterifier si une relation de type cleptobiotique nrsquoeacutetpas en jeu et drsquoen deacutetailler les diffeacuterentes phaLes observations reacutealiseacutees en saison des pluieGuyane (Petit Saut) ont concerneacute 36 nids drsquoE tu-berculatum dont au moins 10 preacutesentaient lrsquoassoction avecCrematogaster La majoriteacute des donneacuteeseacuteteacute collecteacutee sur le nid le plus actif Les enregisments ont porteacute sur divers paramegravetres lieacutes drsquoune pagrave lrsquoactiviteacute de fourragement des deux espegraveces (nomde fourrageuses drsquoEctatomma sortant et entrant dnid flux journalier deCrematogaster pregraves des entreacuteedes nids drsquoEctatomma rythme journalier drsquoexploitation des sources sucreacutees produites par des nymphmembracides taux comparatif de preacutesence des fmis au niveau des nectaires extrafloraux drsquoInga thi-baudiana et sur 30 groupes de nymphes de Membcidae) et drsquoautre part agrave leurs interactions (proport
-
e
de fourrageuses drsquoEctatomma arrecircteacutees lors du retouau nid nombre drsquoouvriegraveres deCrematogaster explo-rant les ouvriegraveres drsquoEctatomma et parties du corpconcerneacutees par lrsquoexploration dureacutee des interactioBien que preacutesentant un pic drsquoactiviteacute plus importaen peacuteriode nocturneE tuberculatum est active toutela journeacutee et exploite en continu les nectaires extraraux et les membracides Comme chez la plupartautres poneacuterines les eacutechanges trophallactiques nrsquotent pas chezE tuberculatum et les fourrageuses qureacutecoltent des liquides les transportent entre leurs mdibules (ces liquides adhegraverent gracircce aux forces desion superficielle) avant de les redistribuer par pseutrophallaxie agrave drsquoautres fourrageuses assurant un rdans le transport (299 des cas) ou directementcongeacutenegraveres agrave lrsquointeacuterieur du nid Jusqursquoagrave 618 desvriegraveres rentrant au nid transportent ainsi des liquisucreacutes (N = 377) Sur les sites alimentairesE tuber-culatum rentre en compeacutetition avec drsquoautres espegravede fourmis pour la reacutecolte des sources sucreacutees noment avec lesC limata parabiotica qui exploitent lesmecircmes espegraveces de membracides sur les mecircmes aet durant les mecircmes creacuteneaux horaires mais geacuteneacutement sur des branches diffeacuterentesEctatomma tuber-culatum est connue pour ecirctre une preacutedatrice efficdrsquoun grand nombre drsquoautres fourmis parmi lesquele genreCrematogaster est largement repreacutesenteacute Djour lesC limata parabiotica eacutevitent manifestementout contact direct avec lesEctatomma ndash utilisant lesmecircmes heacutemiptegraveres mais en alternance ou bienploitant les individus les plus eacuteloigneacutes des ouvriegravedrsquoEctatomma et srsquoeacutecartant degraves que lrsquoune drsquoelles se raproche ndash et servent mecircme freacutequemment de proiesEctatomma Les interactions comportementales enles deux espegraveces changent de faccedilon drastique pela nuit lrsquoagressiviteacute normalement preacutesenteacutee par lesEc-tatomma vis-agrave-vis desCrematogaster disparaissant totalement LesCrematogaster nrsquoeacutevitent plus alors lecontact avec lesEctatomma et mecircme lrsquoinitient lrsquoin-teraction se reacutealisant en deux phases distinctes Danun premier temps en deacutebut de peacuteriode nocturne
F-J Richard et al C R Biologies 327 (2004) 509ndash517 511
eacute-plusres-rra-nm-
pregraveseurecircts)
ettequ
s ndashter-er-aire- De
la
srsquoapur-esantdui-ar-
aseagecu-si-
de li
-id-eadntre-andntes
-ates
st
se
nt
ily
ic-
nd
n
and-
y one-
d
nathe
e
-ib-o-
unktun-
instthers
ityour-identctic
s ofsion
ver-
Crematogaster effectuent de veacuteritables raids agrave lrsquointrieur des nids de la poneacuterine Elles en ressortenttard sans avoir apparemment voleacute de nourrituretent pregraves de lrsquoentreacutee et se mettent agrave arrecircter les fougeuses drsquoEctatomma lors de leur retour au nid tout efaisant vibrer leur abdomen recrutant ainsi de nobreuses congeacutenegraveres Sur 322 ouvriegraveres drsquoEctatommarentrant ainsi au nid 752 ont eacuteteacute arrecircteacutees dontde la moitieacute reprend rapidement le chemin vers lnid Dans tous les autres cas (126 sur les 242 arrles petites ouvriegraveres deCrematogaster grimpent sur ledos de la poneacuterine la legravechent et lorsqursquoune gouttelde miellat est preacutesente entre ses mandibules ndash ceest le cas pour 881 des ouvriegraveres ainsi exploreacuteela volent Ces interactions de type laquo cleptobiose inspeacutecifique raquo preacutesentent ainsi lrsquooriginaliteacute de concner des aliments sucreacutes et non des proies contrment aux autres cas rapporteacutes dans la litteacuteratureplus son expression est temporelle car limiteacutee agravepeacuteriode nocturne et facultative lesCrematogaster nedeacutependant pas strictement de cette strateacutegie pourprovisionner Cette temporaliteacute du pheacutenomegravene porait srsquoexpliquer par la forte augmentation drsquoactiviteacute dEctatomma en deacutebut de peacuteriode nocturne provoquune grande confusion agrave lrsquoentreacutee des nids ce qui reacuterait ou mecircme annulerait le filtrage reacutealiseacute par les gdiennes et autoriserait ainsi les raids deCrematogasteravec un minimum de risques drsquoagression Cette phpermettrait agrave ces derniegraveres drsquoacqueacuterir un camouflchimique par absorption passive drsquohydrocarbonesticulaires camouflage qui serait par la suite intenfieacute lors des leacutechages des fourrageuses chargeacuteesquides sucreacutes
1 Introduction
Cleptobiosis kleptoparasitism food robbing thievery and piracy concern the stealing of food by indivuals of the same or different species This widesprform of interference competition is an importameans by which many animals obtain limitedsources while reducing the costs of searching forhandling food[1ndash3] This foraging strategy has beeextensively described for a large range of vertebraincluding fishes[45] birds[267] and mammals[89] Numerous cases of intra- and interspecific cleptobiosis have also been reported in marine invertebr[10] spiders[1112] and insects such as thrips[13]
i
-
-
wasps[14] bees[15] and ants In ants since the firdescriptions by Wroughton in 1892 (in[16]) and thefirst use of the term lsquocleptobiosisrsquo in its current senby Forel[17] numerous cases have been reported[18ndash21] particularly that of the Neotropical ponerine aEctatomma ruidum [22ndash27]
Aiming to further the understanding of the daforaging rhythms ofEctatomma tuberculatum Whe-eler [28] provided a fortuitous account of a parasitlike association between this species andCremato-gaster limata She observed particularly at dawn adusk that hundreds ofC limata file into the nestentrances ofE tuberculatum When encountering aE tuberculatum worker the smallerC limata climbedup one of the larger antrsquos legs and onto its thoraxhead while theE tuberculatum did not react aggressively to the intruders As Wheeler[28] did not noteany aggressiveness we decided to conduct a studthis relationship hypothesizing that the intriguing bhaviour of theCrematogaster workers could be relateto cleptobiosis
2 Materials and methods
Data were collected in Petit Saut French Guiamostly in July 2000 The mean temperature duringdaytime was 285C falling to 228C at night Themean relative humidity was 85ndash90 during daytimall throughout the periods of observation
Ectatomma tuberculatum is a monogynous (or occasionally polygynous) ponerine ant widely distruted in Neotropical regions Colonies have polydmous nests excavated at the base of a tree trNest entrances are characterized by an externalnel (made with rough vegetal fibres) leaning agathe base of the supporting tree The opening oftunnel is generally guarded by specialized workeWorkers concentrate most of their foraging activon the tree situated above their nest plus neighbing trees and shrubs They solitarily gather a wvariety of small invertebrates and exploit differesugary sources As for most Ponerinae trophallaexchanges cannot occur inEctatomma So foragerscarry between their outstretched mandibles dropletliquid substances that adhere thanks to surface tenstrengths[2228ndash33]
We noted the position of 36Ectatomma neststhanks to their characteristic entrance tunnels Ne
512 F-J Richard et al C R Biologies 327 (2004) 509ndash517
ari-er-hathad
a-c--
0-ad weingux
talthe50
ofans
asne ond100
iednd
by
st
ura-
00
nest
thesed
tedrted
t
r ofes
e
ctar
al
-ndst
theless due to problems of accessibility and great vation in the coloniesrsquo levels of activity detailed obsvations were focused on the most active colony thad four nest entrances Ten out of the 36 nestsrelationships withCrematogaster limata parabioticaa myrmicine species very frequent in pioneer formtions with polydomous nests built in hollowed strutures or ant gardens[3435] The difference in size between the species was great (meanEctatomma workerlengthplusmn SE 95 plusmn 08 mmN = 20 Crematogaster35plusmn 006 mmN = 20)
As the Ectatomma circadian rhythm of activityvaries seasonally[31] we conducted a series of 1min scans over an 8-day-long period until we hcovered each hour out of the 24 During each scanchecked the number of workers entering and leavthe nest of one colony We also estimated the flof Crematogaster workers around theEctatomma nestentrances by using a grid ranging from 0 to 4 (0 toabsence of workers 1 one or two workers nearnest entrance 2 up to ten workers 3 10 toworkers 4 more than 50 workers)
We also studied the rhythm of the exploitationsugary sources Firstly we conducted 10-min scper hour concerningEctatomma workers attendingmembracid nymphs (Hemiptera) on a small treewell as Crematogaster workers on several sites ithe area where we also registered the presencEctatomma (six 3-h time periods between 0800 a0200) Secondly we noted the presence of ants onleaves of 20 juvenileInga thibaudiana (Mimosoideae)and 30 groups of membracid nymphs in the studarea during three periods at night in the morning ain the afternoon
We quantified the cleptobiotic events performedCrematogaster workers through the proportions ofEc-tatomma foragers stopped while returning to their nethe number ofCrematogaster individuals exploringan Ectatomma worker the body part concerned (iemouthparts legs abdomen and thorax) and the dtion of the interaction
3 Results
31 Circadian rhythm of activity
Although continuously activeEctatomma workerspresented two peaks of activity at night around 20
f
Fig 1 (A) Circadian rhythm of activity ofEctatomma tuberculatumthrough the mean number of workers entering and exiting theper 1-h time periods (meanplusmn SE) (B) Estimation of the flux ofCre-matogaster limata parabiotica workers at theE tuberculatum nestentrance by using a grid ranging from 0 (absence ofCrematogasterworkers) to 4 (more than 100Crematogaster workers)
and 0300 Nocturnally between 1900 and 0500number of workers returning to their nest surpasthat of workers exiting the nest (Fig 1A) During acontinuous 24-h-long observation period we nothat 618 of the workers entering the nest transposugary liquids between their mandibles (N = 377) Onthe foraging sitesEctatomma competed with other anspecies includingCrematogaster for the exploitationof membracid honeydew and the extrafloral nectaInga In spite of a reduced level of activity in thmorning Ectatomma workers attended membracidall day long (Fig 2A and C) The nocturnal increasin the number of ants exploitingInga extrafloral nectarcorresponds to the increase in the production of neat night[36]
Crematogaster workers also displayed both diurnand nocturnal activity on the foraging sites (Fig 2BndashD) Around theEctatomma nest entrances their activity level was high at night mostly between 2100 a0400 whenEctatomma foragers returned to their ne(Fig 1B)
F-J Richard et al C R Biologies 327 (2004) 509ndash517 513
fntsg
Fig 2 (A) Dynamics of the exploitation of membracid nymphs byEctatomma tuberculatum workers at one site during a 24-h cycle (B) Thesame forCrematogaster limata parabiotica workers recorded at different sites between 0800 and 0200 (C) Percentages of groups omembracid nymphs attended by ants during three periods of the day (other speciesCamponotus spp 100 of the groups attended by aat 1000 and 2300 933 at 1500 hours) (D) Percentages of leaves ofInga thibaudiana (Mimosoideae) noted with ant workers exploitinthe extrafloral nectaries during three periods of the day (other species mostlyCamponotus spp but also someGigantiops destructor andCephalotes sp in the morning total of 39 7 and 72 of the leaves with ants at 1000 1500 and 2300 respectively)
theen-me
t ofm
the
to- Al-tedof
ps
cidve-eytlytothsedof
eenypec-
tledewr fortheion
32 Ants attending membracids
In the studied area bothCrematogaster and Ec-tatomma foragers attended membracid nymphs onsame trees during the same time periods but gerally on different branches They shared the saparts of the branches on only 25 occasions ou194 noted during the study of their circadian rhythof activity In such cases theCrematogaster attendedthe membracid individuals the most distant fromEctatomma foragers and they moved when anEc-tatomma approached The latter frequently triedcapture theCrematogaster workers 15 attempts occurred on the 25 branches shared by both speciesthough most of them successfully escaped we noduring a series of observations that 14 prey out27 retrieved byE tuberculatum foragers were ants10 of themCrematogaster Also four retrieved preywere membracids originating from attended groutwo complete predatory sequences being observed
During trophobiotic relationshipsEctatommaworkers palpated the lateral parts of the membranymphs with their antennae in an up and down moment eliciting the secretion of honeydew As thstood above the nymphs with their mandibles slighopen they recuperated the honeydew that adheredtheir mandibles thanks to surface tension strengWhile repeating this behaviour the workers opentheir mandibles wider and wider as the dropletshoneydew increased in size (Fig 3A) Then in 701of the cases (N = 127) eachEctatomma forager re-trieved the droplet of honeydew accumulated betwits mandibles after having been replaced or not ba nestmate (252 and 449 of the cases restively) In the other situations (299)Ectatommaworkers specialized in honeydew transport shutbetween the individuals that collected the honeydfrom the membracids and the nest (one transporte4ndash7 collectors) The transfer of honeydew fromcollector to the transporter followed an antennat
514 F-J Richard et al C R Biologies 327 (2004) 509ndash517
ed
Fig 3 (A) Ectatomma tuberculatum worker attending membracid nymphs and gathering a honeydew droplet between its outstretchmandibles (B) E tuberculatum worker stopped by twoCrematogaster limata parabiotica workers inspecting its mouthparts (C) Workerof C limata parabiotica licking a part of the honeydew droplet from aE tuberculatum forager
s ofenidsItewen
ab-
n aredthe
ost-p
nest
re-spedrs at-idly
initiated by the transporter that also made a serie3ndash4 back and forth movements with its body Ththe collecting worker again attended the membracwhile the transporter looked for other collectorsreturned to the nest when the droplet of honeydcompletely filled the space separating its wide-opmandibles
33 Relationships between Crematogaster limataparabioticaand Ectatomma tuberculatum
While completely absent in the vicinity of theEc-tatomma nest entrances during the daytime (Fig 1B)at duskCrematogaster workers entered theEctatommanests without any opposition as the guards weresent As already reported[28] the column ofCre-
matogaster passed through the entrance tunnels iraid-like fashion The beginning of such raids occurbetween 1800 and 2245 which corresponds topeak of nocturnal activity ofEctatomma (Fig 1A) Af-ter these raids theCrematogaster workers exited thetunnel and remained around the nest entrance almall night long (Fig 1B) They never had a swollen abdomen nor any evidence of having filled their croThen they stopped returningEctatomma foragers atthe base of the supporting tree trunk around theentrance or on surrounding branchesCrematogasterworkers waved their abdomens at the approach of aturningEctatomma forager but no liquid droplet wanoted at the tip of their abdomens They also taptheir abdomens on the substrate These behavioutracted nestmates that immediately arrived and rap
F-J Richard et al C R Biologies 327 (2004) 509ndash517 515
ed
80
42r-ringhereng
id
-
)
e-sed
ual
eto
dies
to or
hentter
heer-
n
mersquoy
edn-
fniv-hethe
din-
-reted
y-t
histheur
d-
on-il-
gr
tsr anri-
c-
cal-l or
aggregated around theEctatomma CertainCremato-gaster individuals stayed in front of theEctatommathat remained immobile while up to 12 others climbon theEctatomma (32plusmn 24 N = 98)
A study conducted on 322Ectatomma foragers re-turning to their nest permitted us to note that only(248) were not intercepted by theCrematogasterworkers prior to the tunnel entrance Among the 2Ectatomma intercepted 116 (36 of all returning foagers) were stopped and remained immobile dua short time before resuming their return trip to tnest The remaining 126 individuals (391) westopped explored and licked during a relatively lotime (gt1 min) by theCrematogaster workers Thisincluded 111 individuals carrying a droplet of liqufood that was stolen (345)
Of the 316Crematogaster workers thoroughly observed after the interception of anEctatomma for-ager laden with a droplet of liquid food 165 (522focused their exploration on theEctatomma mouth-parts (seeFig 3B and C) Each individual lickedthe honeydew only during a few seconds (66 plusmn54 s N = 70) When the honeydew droplet was dpleted theEctatomma workers remained motionlesmandibles open maxilo-labial mouthparts extendallowing theCrematogaster workers to lick them dur-ing a long time (25plusmn 145 minN = 15)
4 Discussion
We confirm therefore the existence of the unusrelationship betweenE tuberculatum andC limata asreported in central Panama[28] Nevertheless somnew information and significant differences deservebe pointed out
First Ectatomma workers constantly attendegroups of membracid nymphs thanks to two strategWorkers collected honeydew that they transportedthe nest once they were replaced by a nestmatecertain workers collected the honeydew that was ttransported to the nest by other workers The lacooperative behaviour known in the Formicinae[3]presents similarities with the lsquorelay transportrsquo and tlsquoprey chain transfer behaviourrsquo noted for other ponine ants during predation[3738]
Second we noted a dramatic difference inCremato-gaster-Ectatomma interspecific relationships betwee
day and night Although sharing branches with theCrematogaster workers maintained a lsquosafe distancfrom Ectatomma in the foraging areas during the dathe latter being able to prey on them (see also[33])On the contrary at night theCrematogaster work-ers no longer avoided direct contact withE tubercu-latum but initiated the interactions They conducttheir raids on theEctatomma nest entrances then egaged in contact with theEctatomma foragers return-ing to their nest stopping and exploring them
Third Wheeler[28] noted that the behaviour oCrematogaster workers during raids was limited itime and consisted of a kind of lsquoshampooingrsquo dering nutrition from the cuticular secretions andor texcess fluids (nectar and honeydew) trapped onmandibles ofEctatomma workers This author notethat such behaviour first described between anquiline ant and its host[39] could account for a mutualistic association of the lsquocleaner-fishrsquo type wheboth species derive benefits Even if we again nothat Crematogaster workers licked the different bodparts of theEctatomma they mostly stopped and explored theEctatomma retrieving a honeydew drople(881 of the exploredE tuberculatum foragers) andrecruited nestmates in order to rob this droplet Tresulted therefore in a parasitic relationship oflsquocleptobioticrsquo form This nocturnal robbing behavioonly complements the food supply of theCremato-gaster that gather most of their sugary food by attening hemiptera or exploiting extrafloral nectaries
Fourth other cases of cleptobiosis in ants ccerned prey robbing with nevertheless a certain simitude with the present study For exampleMyrmeco-cystus mimicus workers thoroughly inspect returninforagers ofPogonomyrmex spp before robbing theiprey [20] The originality of the cleptobiotic activityof C limata parabiotica consists in exploiting the kindof liquid transport limited to arboreal ponerine anIndeed they carry droplets of liquid between theimandibles which are then imbibed by nestmatesadaptation to the lack of trophallaxis in most Ponenae[4041]
Finally we questioned the origin of the lower noturnal aggressiveness ofEctatomma workers towardsCrematogaster The immobility of the Ectatommaworkers could be triggered by a secreted chemiwhile the Crematogaster wave their abdomens Indeed secretions from the mandibular metapleura
516 F-J Richard et al C R Biologies 327 (2004) 509ndash517
d
id-
plet
he
ur-
ack
at-o-be
p-ac-p-
ire
-oure
nebio-hem
sisceso-e
ACa--the
entGIFs raquo
ds
ty
lm
a-bo5ndash
c02)
man
rrsquos
of-im
ter-star
ction9)
n-
02)
of82
iest
ar-
nd0
Dufour glands act as repellents in severalCremato-gaster species[42ndash44] and such an effect was notein the lestobioticSolenopsis fugax [45] Similarly apacifying action on the host occurs when an indivual of the parasiticFormicoxenus provancheri raisesits abdomen and extrudes the sting producing a droof volatile liquid before engaging in lsquoshampooingrsquo be-haviour withMyrmica incompleta [46]
Nevertheless why does this not work during tdaytime As theCrematogaster lsquoraidsrsquo coincide withthe dusk peak in activity ofEctatomma (1900ndash2100Fig 1A) another hypothesis can be formulated Ding this period the heavy flow ofEctatomma foragersentering and leaving the tunnel drives the guards binto the nest This permits the smallCrematogasterworkers to creep inside the tunnel without beingtacked In this situation they can modify the compsition of their cuticular hydrocarbons enough toaccepted by the nextEctatomma foragers returningto their nest and so to lure them A similar decetive mechanism based on chemical camouflagequisition was proposed to explain intraspecific cletobiosis inE ruidum [27] Later newCrematogasterindividuals can enter the tunnel or they can acqucuticular compounds directly from an interceptedEc-tatomma worker by climbing on it and licking its cuticle this corresponding to the lsquoshampooingrsquo behavidescribed by Wheeler[28] This hypothesis might bsupported by the fact thatC limata parabiotica has theability to share nests and trails with different formiciand ponerine ants in a phenomenon called lsquoparasisrsquo while these ants never share nests between t[343546]
In conclusion we noted a new kind of cleptobioin ants concerning the robbing of sugary substanand not prey robbing as previously known Temprally limited to night time and facultative it implies thpresence ofEctatomma nests onCrematogaster terri-tories
Acknowledgements
We are grateful to Dr Jacques HC Delabie (UPLaboratoacuterio de Mirmecologia Convecircnio UESCEPLAC Itabuna Bahia Brazil) for the identifiction of the ants and to AndreaDejean for proofreading the manuscript This work was supported by
French laquo Ministegravere de lrsquoEacutecologie et du Deacuteveloppemdurable raquo (SOFT program research agreementECOFOR No 98 and the laquo Tropical Ecosystemprogram No 02-E2002)
References
[1] E Curio in The Ethology ofPredation Springer-VerlagBerlin 1976 242 p
[2] HJ Brockmann CJ Barnard Kleptoparasitism in birAnim Behav 27 (1979) 487ndash514
[3] B Houmllldobler EO Wilson in The Ants Harvard UniversiPress Cambridge 1990 732 p
[4] TJ Pitcher Functions of shoaling behaviour in teleosts inJ Pitcher (Ed) The Behaviour of Teleost Fishes Croom HeLondon 1986 pp 294ndash337
[5] WJ Dominey AM Snyder Kleptoparasitism of freshwter crabs by cichlid fishes endemic to Lake Barombi MCameroon West Africa Environ Biol Fishes 22 (1988) 15160
[6] JR Garrido CG Sarasa M Fernandez-Cruz Intraspecifikleptoparasitism in the cattle egret J Field Ornithol 73 (20185ndash190
[7] CE Khatchikian M Favero AI Vassalo Kleptoparasitisby brown-hooded gulls and grey-hooded gulls on Americoystercatchers Waterbirds 25 (2002) 137ndash141
[8] MS Di Bitetti CH Janson Social foraging and the findeshare in capuchin monkeysCebus apella Anim Behav 62(2001) 47ndash56
[9] OP Honer B Wachter ML East H Hofer The responsespotted hyenas to long-term changes in prey populations functional response and interspecific kleptoparasitism J AnEcol 71 (2002) 236ndash246
[10] S Morissette JH Himmelman Subtidal food thieves inactions of four invertabrate kleptoparasites with the seaLeptasterias polaris Anim Behav 60 (2000) 531ndash543
[11] Y Heacutenaut J-P Lachaud Araigneacutees cleptoparasites seacutelede lrsquohocircte par sa toile Ann Soc Entomol Fr (NS) 35 (199148ndash153
[12] M Whitehouse I Agnarsson T Miyashita D Smith K Cagialosi T Masumoto D Li Y HeacutenautArgyrodes phylogenysociality and interspecific interactions J Arachnol 30 (20238ndash245
[13] B Crespi P Abbot The behavioral ecology and evolutionkleptoparasitism in Australian gall thrips Fla Entomol(1999) 147ndash164
[14] FE Kurczewski MG Spofford Alternative nesting strategin Ammophila urnaria (Hymenoptera Sphecidae) J NaHist 32 (1998) 99ndash106
[15] C Michener in The Social Behavior of the Bees A Compative Study The Belknap Pressof Harvard University PressCambridge 1974 404 p
[16] WM Wheeler in Ants Their Structure Development aBehaviour Columbia University Press New York 191663 p
F-J Richard et al C R Biologies 327 (2004) 509ndash517 517
)
in al 20
ing
89ndash
mic
ry
d)
n andash
5ndash
verinendash
hes
n
Ac-
ns
D
-
reyerae)
s
la-theera
c-eaery
n
ri-t 35
on
es-
ra
m
nta-
89)
de-
re
-
es
ific40
[17] A Forel Fourmis termitophages LestobioseAtta tardigradasous-genres drsquoEuponera Ann Soc Entomol Belg 45 (1901389ndash398
[18] T Abe On the food sharing among four species of antssandy grassland I Food and foraging behavior Jap J Eco(1971) 219ndash230
[19] U Maschwitz M MuhlenbergCamponotus rufoglaucus einewegelagernde Ameise Zool Anz 191 (1973) 364ndash368
[20] B Houmllldobler Food robbing inants a form of interferencecompetition Oecologia 69 (1986) 12ndash15
[21] T Yamaguchi Intraspecific competition through food robbin the harvester antMessor aciculatus (Fr Smith) and itsconsequences on colony survival Insectes Soc 42 (1995)101
[22] NA Weber Two common ponerine ants of possible econosignificanceEctatomma tuberculatum (Olivier) andE ruidumRoger Proc Entomol Soc Washington 48 (1946) 1ndash16
[23] MD Breed P Abel TJ Bleuze SE Denton Thievehome ranges and nestmate recognition inEctatomma ruidumOecologia 84 (1990) 117ndash121
[24] MD Breed LE Snyder TL Lynn JA Morhart Acquirechemical camouflage in a tropical ant Anim Behav 44 (1992519ndash523
[25] MD Breed TP McGlynn EM Stocker ANKlein Thiefworkers and variation in nestmate recognition behavior iponerine antEctatomma ruidum Insectes Soc 46 (1999) 327331
[26] I Perfecto JH Vandermeer Cleptobiosis in the antEc-tatomma ruidum in Nicaragua Insectes Soc 40 (1993) 29299
[27] JM Jeral MD Breed BE Hibbard Thief ants hareduced quantities of cuticular compounds in a poneant Ectatomma ruidum Physiol Entomol 22 (1997) 207211
[28] DE WheelerEctatomma tuberculatum foraging biology andassociation withCrematogaster (Hymenoptera Formicidae)Ann Entomol Soc Am 79 (1986) 300ndash303
[29] WL Jr Brown Contributions toward a reclassification of tFormicidae II Tribe Ectatommini (Hymenoptera) Bull MuComp Zool 118 (1958) 173ndash362
[30] A Dejean J-P Lachaud Growth-related changes in predatiobehavior in incipient colonies of the ponerine antEctatommatuberculatum (Olivier) Insectes Soc 39 (1992) 129ndash143
[31] J Valenzuela-Gonzaacutelez A Loacutepez-Meacutendez J-P Lachaudtivity patterns and foraging activity in nests ofEctatommatuberculatum (Hymenoptera Formicidae) in cacao plantatioSouthwest Entomol 20 (1995) 507ndash515
[32] RR da Hora R Feacuteneacuteron J Valenzuela ME FavilaFresneau Queen-size dimorphism in the antEctatomma
tuberculatum (Hymenoptera Formicidae Ponerinae) Sociobiology 38 (2001) 407ndash420
[33] G Ibarra-Nuacutentildeez JA Garciacutea JA Loacutepez J-P Lachaud Panalysis in the diet of some ponerine ants (HymenoptFormicidae) and web-building spiders (Araneae) in coffeplantations in Chiapas Mexico Sociobiology 37 (2001723ndash755
[34] DW Davidson Ecological studies of Neotropical ant gardenEcology 69 (1988) 1138ndash1152
[35] A Dejean B Corbara J Orivel RR Snelling JHC Debie M Belin-Depoux The importance of ant gardens inpioneer vegetal formations of French Guiana (HymenoptFormicidae) Sociobiology 35 (2000) 425ndash439
[36] LM Pascal EF Motte-Florac DB McKey Secretory strutures on the leaf rachis of Caesalpinieae and Mimosoid(Leguminosae) implication for the evolution of nectaglands Am J Bot 87 (2000) 327ndash338
[37] B Schatz J-P Lachaud G Beugnon Polyethism withihunters of the ponerine antEctatomma ruidum InsectesSoc 43 (1996) 111ndash118
[38] WM Wheeler The compound and mixed nests of Amecan ants Part I Observations on a new guest ant Am Na(1901) 431ndash448
[39] B Houmllldobler Liquid food transmission and antennatisignals in ponerine ants Isr J Entomol 19 (1985) 89ndash99
[40] J Liebig J Heinze B Houmllldobler Trophallaxis and aggrsion in the ponerine antPonera coartata implications forthe evolution of liquid food exchange in the HymenopteEthology 103 (1997) 707ndash722
[41] C Bevan A Birch H Caswell Ant insect repellent froblack cocktail ants J Chem Soc 961 (1961) 488
[42] AB Attygalle B Siegel O Vostrowsky HJ BestmanU Maschwitz Chemical composition and function of mepleural gland secretion of the antCrematogaster deformisSmith (Hymenoptera Myrmicinae) J Chem Ecol 15 (19317ndash328
[43] D Daloze M Kaisin C Detrain JM Pasteels Chemicalfense in the three European species ofCrematogaster antsExperientia 47 (1991) 1082ndash1089
[44] B Houmllldobler Chemische Strategie beim Nahrungserwerb deDiebameise (Solenopsis fugax Latr) und der Pharaoameis(Monomorium pharaonis L) Oecologia 11 (1973) 371ndash380
[45] A Lenoir C Errard A Francoeur R Loiselle Relations entre la fourmi parasiteFormicoxenus provancheri et son hocircteMyrmica incompleta Donneacutees biologiques et eacutethologiqu(Hym Formicidae) Insectes Soc 39 (1992) 81ndash97
[46] J Orivel C Errard A Dejean Ant gardens interspecrecognition in parabiotic species Behav Ecol Sociobiol(1997) 87ndash93

F-J Richard et al C R Biologies 327 (2004) 509ndash517 511
eacute-plusres-rra-nm-
pregraveseurecircts)
ettequ
s ndashter-er-aire- De
la
srsquoapur-esantdui-ar-
aseagecu-si-
de li
-id-eadntre-andntes
-ates
st
se
nt
ily
ic-
nd
n
and-
y one-
d
nathe
e
-ib-o-
unktun-
instthers
ityour-identctic
s ofsion
ver-
Crematogaster effectuent de veacuteritables raids agrave lrsquointrieur des nids de la poneacuterine Elles en ressortenttard sans avoir apparemment voleacute de nourrituretent pregraves de lrsquoentreacutee et se mettent agrave arrecircter les fougeuses drsquoEctatomma lors de leur retour au nid tout efaisant vibrer leur abdomen recrutant ainsi de nobreuses congeacutenegraveres Sur 322 ouvriegraveres drsquoEctatommarentrant ainsi au nid 752 ont eacuteteacute arrecircteacutees dontde la moitieacute reprend rapidement le chemin vers lnid Dans tous les autres cas (126 sur les 242 arrles petites ouvriegraveres deCrematogaster grimpent sur ledos de la poneacuterine la legravechent et lorsqursquoune gouttelde miellat est preacutesente entre ses mandibules ndash ceest le cas pour 881 des ouvriegraveres ainsi exploreacuteela volent Ces interactions de type laquo cleptobiose inspeacutecifique raquo preacutesentent ainsi lrsquooriginaliteacute de concner des aliments sucreacutes et non des proies contrment aux autres cas rapporteacutes dans la litteacuteratureplus son expression est temporelle car limiteacutee agravepeacuteriode nocturne et facultative lesCrematogaster nedeacutependant pas strictement de cette strateacutegie pourprovisionner Cette temporaliteacute du pheacutenomegravene porait srsquoexpliquer par la forte augmentation drsquoactiviteacute dEctatomma en deacutebut de peacuteriode nocturne provoquune grande confusion agrave lrsquoentreacutee des nids ce qui reacuterait ou mecircme annulerait le filtrage reacutealiseacute par les gdiennes et autoriserait ainsi les raids deCrematogasteravec un minimum de risques drsquoagression Cette phpermettrait agrave ces derniegraveres drsquoacqueacuterir un camouflchimique par absorption passive drsquohydrocarbonesticulaires camouflage qui serait par la suite intenfieacute lors des leacutechages des fourrageuses chargeacuteesquides sucreacutes
1 Introduction
Cleptobiosis kleptoparasitism food robbing thievery and piracy concern the stealing of food by indivuals of the same or different species This widesprform of interference competition is an importameans by which many animals obtain limitedsources while reducing the costs of searching forhandling food[1ndash3] This foraging strategy has beeextensively described for a large range of vertebraincluding fishes[45] birds[267] and mammals[89] Numerous cases of intra- and interspecific cleptobiosis have also been reported in marine invertebr[10] spiders[1112] and insects such as thrips[13]
i
-
-
wasps[14] bees[15] and ants In ants since the firdescriptions by Wroughton in 1892 (in[16]) and thefirst use of the term lsquocleptobiosisrsquo in its current senby Forel[17] numerous cases have been reported[18ndash21] particularly that of the Neotropical ponerine aEctatomma ruidum [22ndash27]
Aiming to further the understanding of the daforaging rhythms ofEctatomma tuberculatum Whe-eler [28] provided a fortuitous account of a parasitlike association between this species andCremato-gaster limata She observed particularly at dawn adusk that hundreds ofC limata file into the nestentrances ofE tuberculatum When encountering aE tuberculatum worker the smallerC limata climbedup one of the larger antrsquos legs and onto its thoraxhead while theE tuberculatum did not react aggressively to the intruders As Wheeler[28] did not noteany aggressiveness we decided to conduct a studthis relationship hypothesizing that the intriguing bhaviour of theCrematogaster workers could be relateto cleptobiosis
2 Materials and methods
Data were collected in Petit Saut French Guiamostly in July 2000 The mean temperature duringdaytime was 285C falling to 228C at night Themean relative humidity was 85ndash90 during daytimall throughout the periods of observation
Ectatomma tuberculatum is a monogynous (or occasionally polygynous) ponerine ant widely distruted in Neotropical regions Colonies have polydmous nests excavated at the base of a tree trNest entrances are characterized by an externalnel (made with rough vegetal fibres) leaning agathe base of the supporting tree The opening oftunnel is generally guarded by specialized workeWorkers concentrate most of their foraging activon the tree situated above their nest plus neighbing trees and shrubs They solitarily gather a wvariety of small invertebrates and exploit differesugary sources As for most Ponerinae trophallaexchanges cannot occur inEctatomma So foragerscarry between their outstretched mandibles dropletliquid substances that adhere thanks to surface tenstrengths[2228ndash33]
We noted the position of 36Ectatomma neststhanks to their characteristic entrance tunnels Ne
512 F-J Richard et al C R Biologies 327 (2004) 509ndash517
ari-er-hathad
a-c--
0-ad weingux
talthe50
ofans
asne ond100
iednd
by
st
ura-
00
nest
thesed
tedrted
t
r ofes
e
ctar
al
-ndst
theless due to problems of accessibility and great vation in the coloniesrsquo levels of activity detailed obsvations were focused on the most active colony thad four nest entrances Ten out of the 36 nestsrelationships withCrematogaster limata parabioticaa myrmicine species very frequent in pioneer formtions with polydomous nests built in hollowed strutures or ant gardens[3435] The difference in size between the species was great (meanEctatomma workerlengthplusmn SE 95 plusmn 08 mmN = 20 Crematogaster35plusmn 006 mmN = 20)
As the Ectatomma circadian rhythm of activityvaries seasonally[31] we conducted a series of 1min scans over an 8-day-long period until we hcovered each hour out of the 24 During each scanchecked the number of workers entering and leavthe nest of one colony We also estimated the flof Crematogaster workers around theEctatomma nestentrances by using a grid ranging from 0 to 4 (0 toabsence of workers 1 one or two workers nearnest entrance 2 up to ten workers 3 10 toworkers 4 more than 50 workers)
We also studied the rhythm of the exploitationsugary sources Firstly we conducted 10-min scper hour concerningEctatomma workers attendingmembracid nymphs (Hemiptera) on a small treewell as Crematogaster workers on several sites ithe area where we also registered the presencEctatomma (six 3-h time periods between 0800 a0200) Secondly we noted the presence of ants onleaves of 20 juvenileInga thibaudiana (Mimosoideae)and 30 groups of membracid nymphs in the studarea during three periods at night in the morning ain the afternoon
We quantified the cleptobiotic events performedCrematogaster workers through the proportions ofEc-tatomma foragers stopped while returning to their nethe number ofCrematogaster individuals exploringan Ectatomma worker the body part concerned (iemouthparts legs abdomen and thorax) and the dtion of the interaction
3 Results
31 Circadian rhythm of activity
Although continuously activeEctatomma workerspresented two peaks of activity at night around 20
f
Fig 1 (A) Circadian rhythm of activity ofEctatomma tuberculatumthrough the mean number of workers entering and exiting theper 1-h time periods (meanplusmn SE) (B) Estimation of the flux ofCre-matogaster limata parabiotica workers at theE tuberculatum nestentrance by using a grid ranging from 0 (absence ofCrematogasterworkers) to 4 (more than 100Crematogaster workers)
and 0300 Nocturnally between 1900 and 0500number of workers returning to their nest surpasthat of workers exiting the nest (Fig 1A) During acontinuous 24-h-long observation period we nothat 618 of the workers entering the nest transposugary liquids between their mandibles (N = 377) Onthe foraging sitesEctatomma competed with other anspecies includingCrematogaster for the exploitationof membracid honeydew and the extrafloral nectaInga In spite of a reduced level of activity in thmorning Ectatomma workers attended membracidall day long (Fig 2A and C) The nocturnal increasin the number of ants exploitingInga extrafloral nectarcorresponds to the increase in the production of neat night[36]
Crematogaster workers also displayed both diurnand nocturnal activity on the foraging sites (Fig 2BndashD) Around theEctatomma nest entrances their activity level was high at night mostly between 2100 a0400 whenEctatomma foragers returned to their ne(Fig 1B)
F-J Richard et al C R Biologies 327 (2004) 509ndash517 513
fntsg
Fig 2 (A) Dynamics of the exploitation of membracid nymphs byEctatomma tuberculatum workers at one site during a 24-h cycle (B) Thesame forCrematogaster limata parabiotica workers recorded at different sites between 0800 and 0200 (C) Percentages of groups omembracid nymphs attended by ants during three periods of the day (other speciesCamponotus spp 100 of the groups attended by aat 1000 and 2300 933 at 1500 hours) (D) Percentages of leaves ofInga thibaudiana (Mimosoideae) noted with ant workers exploitinthe extrafloral nectaries during three periods of the day (other species mostlyCamponotus spp but also someGigantiops destructor andCephalotes sp in the morning total of 39 7 and 72 of the leaves with ants at 1000 1500 and 2300 respectively)
theen-me
t ofm
the
to- Al-tedof
ps
cidve-eytlytothsedof
eenypec-
tledewr fortheion
32 Ants attending membracids
In the studied area bothCrematogaster and Ec-tatomma foragers attended membracid nymphs onsame trees during the same time periods but gerally on different branches They shared the saparts of the branches on only 25 occasions ou194 noted during the study of their circadian rhythof activity In such cases theCrematogaster attendedthe membracid individuals the most distant fromEctatomma foragers and they moved when anEc-tatomma approached The latter frequently triedcapture theCrematogaster workers 15 attempts occurred on the 25 branches shared by both speciesthough most of them successfully escaped we noduring a series of observations that 14 prey out27 retrieved byE tuberculatum foragers were ants10 of themCrematogaster Also four retrieved preywere membracids originating from attended groutwo complete predatory sequences being observed
During trophobiotic relationshipsEctatommaworkers palpated the lateral parts of the membranymphs with their antennae in an up and down moment eliciting the secretion of honeydew As thstood above the nymphs with their mandibles slighopen they recuperated the honeydew that adheredtheir mandibles thanks to surface tension strengWhile repeating this behaviour the workers opentheir mandibles wider and wider as the dropletshoneydew increased in size (Fig 3A) Then in 701of the cases (N = 127) eachEctatomma forager re-trieved the droplet of honeydew accumulated betwits mandibles after having been replaced or not ba nestmate (252 and 449 of the cases restively) In the other situations (299)Ectatommaworkers specialized in honeydew transport shutbetween the individuals that collected the honeydfrom the membracids and the nest (one transporte4ndash7 collectors) The transfer of honeydew fromcollector to the transporter followed an antennat
514 F-J Richard et al C R Biologies 327 (2004) 509ndash517
ed
Fig 3 (A) Ectatomma tuberculatum worker attending membracid nymphs and gathering a honeydew droplet between its outstretchmandibles (B) E tuberculatum worker stopped by twoCrematogaster limata parabiotica workers inspecting its mouthparts (C) Workerof C limata parabiotica licking a part of the honeydew droplet from aE tuberculatum forager
s ofenidsItewen
ab-
n aredthe
ost-p
nest
re-spedrs at-idly
initiated by the transporter that also made a serie3ndash4 back and forth movements with its body Ththe collecting worker again attended the membracwhile the transporter looked for other collectorsreturned to the nest when the droplet of honeydcompletely filled the space separating its wide-opmandibles
33 Relationships between Crematogaster limataparabioticaand Ectatomma tuberculatum
While completely absent in the vicinity of theEc-tatomma nest entrances during the daytime (Fig 1B)at duskCrematogaster workers entered theEctatommanests without any opposition as the guards weresent As already reported[28] the column ofCre-
matogaster passed through the entrance tunnels iraid-like fashion The beginning of such raids occurbetween 1800 and 2245 which corresponds topeak of nocturnal activity ofEctatomma (Fig 1A) Af-ter these raids theCrematogaster workers exited thetunnel and remained around the nest entrance almall night long (Fig 1B) They never had a swollen abdomen nor any evidence of having filled their croThen they stopped returningEctatomma foragers atthe base of the supporting tree trunk around theentrance or on surrounding branchesCrematogasterworkers waved their abdomens at the approach of aturningEctatomma forager but no liquid droplet wanoted at the tip of their abdomens They also taptheir abdomens on the substrate These behavioutracted nestmates that immediately arrived and rap
F-J Richard et al C R Biologies 327 (2004) 509ndash517 515
ed
80
42r-ringhereng
id
-
)
e-sed
ual
eto
dies
to or
hentter
heer-
n
mersquoy
edn-
fniv-hethe
din-
-reted
y-t
histheur
d-
on-il-
gr
tsr anri-
c-
cal-l or
aggregated around theEctatomma CertainCremato-gaster individuals stayed in front of theEctatommathat remained immobile while up to 12 others climbon theEctatomma (32plusmn 24 N = 98)
A study conducted on 322Ectatomma foragers re-turning to their nest permitted us to note that only(248) were not intercepted by theCrematogasterworkers prior to the tunnel entrance Among the 2Ectatomma intercepted 116 (36 of all returning foagers) were stopped and remained immobile dua short time before resuming their return trip to tnest The remaining 126 individuals (391) westopped explored and licked during a relatively lotime (gt1 min) by theCrematogaster workers Thisincluded 111 individuals carrying a droplet of liqufood that was stolen (345)
Of the 316Crematogaster workers thoroughly observed after the interception of anEctatomma for-ager laden with a droplet of liquid food 165 (522focused their exploration on theEctatomma mouth-parts (seeFig 3B and C) Each individual lickedthe honeydew only during a few seconds (66 plusmn54 s N = 70) When the honeydew droplet was dpleted theEctatomma workers remained motionlesmandibles open maxilo-labial mouthparts extendallowing theCrematogaster workers to lick them dur-ing a long time (25plusmn 145 minN = 15)
4 Discussion
We confirm therefore the existence of the unusrelationship betweenE tuberculatum andC limata asreported in central Panama[28] Nevertheless somnew information and significant differences deservebe pointed out
First Ectatomma workers constantly attendegroups of membracid nymphs thanks to two strategWorkers collected honeydew that they transportedthe nest once they were replaced by a nestmatecertain workers collected the honeydew that was ttransported to the nest by other workers The lacooperative behaviour known in the Formicinae[3]presents similarities with the lsquorelay transportrsquo and tlsquoprey chain transfer behaviourrsquo noted for other ponine ants during predation[3738]
Second we noted a dramatic difference inCremato-gaster-Ectatomma interspecific relationships betwee
day and night Although sharing branches with theCrematogaster workers maintained a lsquosafe distancfrom Ectatomma in the foraging areas during the dathe latter being able to prey on them (see also[33])On the contrary at night theCrematogaster work-ers no longer avoided direct contact withE tubercu-latum but initiated the interactions They conducttheir raids on theEctatomma nest entrances then egaged in contact with theEctatomma foragers return-ing to their nest stopping and exploring them
Third Wheeler[28] noted that the behaviour oCrematogaster workers during raids was limited itime and consisted of a kind of lsquoshampooingrsquo dering nutrition from the cuticular secretions andor texcess fluids (nectar and honeydew) trapped onmandibles ofEctatomma workers This author notethat such behaviour first described between anquiline ant and its host[39] could account for a mutualistic association of the lsquocleaner-fishrsquo type wheboth species derive benefits Even if we again nothat Crematogaster workers licked the different bodparts of theEctatomma they mostly stopped and explored theEctatomma retrieving a honeydew drople(881 of the exploredE tuberculatum foragers) andrecruited nestmates in order to rob this droplet Tresulted therefore in a parasitic relationship oflsquocleptobioticrsquo form This nocturnal robbing behavioonly complements the food supply of theCremato-gaster that gather most of their sugary food by attening hemiptera or exploiting extrafloral nectaries
Fourth other cases of cleptobiosis in ants ccerned prey robbing with nevertheless a certain simitude with the present study For exampleMyrmeco-cystus mimicus workers thoroughly inspect returninforagers ofPogonomyrmex spp before robbing theiprey [20] The originality of the cleptobiotic activityof C limata parabiotica consists in exploiting the kindof liquid transport limited to arboreal ponerine anIndeed they carry droplets of liquid between theimandibles which are then imbibed by nestmatesadaptation to the lack of trophallaxis in most Ponenae[4041]
Finally we questioned the origin of the lower noturnal aggressiveness ofEctatomma workers towardsCrematogaster The immobility of the Ectatommaworkers could be triggered by a secreted chemiwhile the Crematogaster wave their abdomens Indeed secretions from the mandibular metapleura
516 F-J Richard et al C R Biologies 327 (2004) 509ndash517
d
id-
plet
he
ur-
ack
at-o-be
p-ac-p-
ire
-oure
nebio-hem
sisceso-e
ACa--the
entGIFs raquo
ds
ty
lm
a-bo5ndash
c02)
man
rrsquos
of-im
ter-star
ction9)
n-
02)
of82
iest
ar-
nd0
Dufour glands act as repellents in severalCremato-gaster species[42ndash44] and such an effect was notein the lestobioticSolenopsis fugax [45] Similarly apacifying action on the host occurs when an indivual of the parasiticFormicoxenus provancheri raisesits abdomen and extrudes the sting producing a droof volatile liquid before engaging in lsquoshampooingrsquo be-haviour withMyrmica incompleta [46]
Nevertheless why does this not work during tdaytime As theCrematogaster lsquoraidsrsquo coincide withthe dusk peak in activity ofEctatomma (1900ndash2100Fig 1A) another hypothesis can be formulated Ding this period the heavy flow ofEctatomma foragersentering and leaving the tunnel drives the guards binto the nest This permits the smallCrematogasterworkers to creep inside the tunnel without beingtacked In this situation they can modify the compsition of their cuticular hydrocarbons enough toaccepted by the nextEctatomma foragers returningto their nest and so to lure them A similar decetive mechanism based on chemical camouflagequisition was proposed to explain intraspecific cletobiosis inE ruidum [27] Later newCrematogasterindividuals can enter the tunnel or they can acqucuticular compounds directly from an interceptedEc-tatomma worker by climbing on it and licking its cuticle this corresponding to the lsquoshampooingrsquo behavidescribed by Wheeler[28] This hypothesis might bsupported by the fact thatC limata parabiotica has theability to share nests and trails with different formiciand ponerine ants in a phenomenon called lsquoparasisrsquo while these ants never share nests between t[343546]
In conclusion we noted a new kind of cleptobioin ants concerning the robbing of sugary substanand not prey robbing as previously known Temprally limited to night time and facultative it implies thpresence ofEctatomma nests onCrematogaster terri-tories
Acknowledgements
We are grateful to Dr Jacques HC Delabie (UPLaboratoacuterio de Mirmecologia Convecircnio UESCEPLAC Itabuna Bahia Brazil) for the identifiction of the ants and to AndreaDejean for proofreading the manuscript This work was supported by
French laquo Ministegravere de lrsquoEacutecologie et du Deacuteveloppemdurable raquo (SOFT program research agreementECOFOR No 98 and the laquo Tropical Ecosystemprogram No 02-E2002)
References
[1] E Curio in The Ethology ofPredation Springer-VerlagBerlin 1976 242 p
[2] HJ Brockmann CJ Barnard Kleptoparasitism in birAnim Behav 27 (1979) 487ndash514
[3] B Houmllldobler EO Wilson in The Ants Harvard UniversiPress Cambridge 1990 732 p
[4] TJ Pitcher Functions of shoaling behaviour in teleosts inJ Pitcher (Ed) The Behaviour of Teleost Fishes Croom HeLondon 1986 pp 294ndash337
[5] WJ Dominey AM Snyder Kleptoparasitism of freshwter crabs by cichlid fishes endemic to Lake Barombi MCameroon West Africa Environ Biol Fishes 22 (1988) 15160
[6] JR Garrido CG Sarasa M Fernandez-Cruz Intraspecifikleptoparasitism in the cattle egret J Field Ornithol 73 (20185ndash190
[7] CE Khatchikian M Favero AI Vassalo Kleptoparasitisby brown-hooded gulls and grey-hooded gulls on Americoystercatchers Waterbirds 25 (2002) 137ndash141
[8] MS Di Bitetti CH Janson Social foraging and the findeshare in capuchin monkeysCebus apella Anim Behav 62(2001) 47ndash56
[9] OP Honer B Wachter ML East H Hofer The responsespotted hyenas to long-term changes in prey populations functional response and interspecific kleptoparasitism J AnEcol 71 (2002) 236ndash246
[10] S Morissette JH Himmelman Subtidal food thieves inactions of four invertabrate kleptoparasites with the seaLeptasterias polaris Anim Behav 60 (2000) 531ndash543
[11] Y Heacutenaut J-P Lachaud Araigneacutees cleptoparasites seacutelede lrsquohocircte par sa toile Ann Soc Entomol Fr (NS) 35 (199148ndash153
[12] M Whitehouse I Agnarsson T Miyashita D Smith K Cagialosi T Masumoto D Li Y HeacutenautArgyrodes phylogenysociality and interspecific interactions J Arachnol 30 (20238ndash245
[13] B Crespi P Abbot The behavioral ecology and evolutionkleptoparasitism in Australian gall thrips Fla Entomol(1999) 147ndash164
[14] FE Kurczewski MG Spofford Alternative nesting strategin Ammophila urnaria (Hymenoptera Sphecidae) J NaHist 32 (1998) 99ndash106
[15] C Michener in The Social Behavior of the Bees A Compative Study The Belknap Pressof Harvard University PressCambridge 1974 404 p
[16] WM Wheeler in Ants Their Structure Development aBehaviour Columbia University Press New York 191663 p
F-J Richard et al C R Biologies 327 (2004) 509ndash517 517
)
in al 20
ing
89ndash
mic
ry
d)
n andash
5ndash
verinendash
hes
n
Ac-
ns
D
-
reyerae)
s
la-theera
c-eaery
n
ri-t 35
on
es-
ra
m
nta-
89)
de-
re
-
es
ific40
[17] A Forel Fourmis termitophages LestobioseAtta tardigradasous-genres drsquoEuponera Ann Soc Entomol Belg 45 (1901389ndash398
[18] T Abe On the food sharing among four species of antssandy grassland I Food and foraging behavior Jap J Eco(1971) 219ndash230
[19] U Maschwitz M MuhlenbergCamponotus rufoglaucus einewegelagernde Ameise Zool Anz 191 (1973) 364ndash368
[20] B Houmllldobler Food robbing inants a form of interferencecompetition Oecologia 69 (1986) 12ndash15
[21] T Yamaguchi Intraspecific competition through food robbin the harvester antMessor aciculatus (Fr Smith) and itsconsequences on colony survival Insectes Soc 42 (1995)101
[22] NA Weber Two common ponerine ants of possible econosignificanceEctatomma tuberculatum (Olivier) andE ruidumRoger Proc Entomol Soc Washington 48 (1946) 1ndash16
[23] MD Breed P Abel TJ Bleuze SE Denton Thievehome ranges and nestmate recognition inEctatomma ruidumOecologia 84 (1990) 117ndash121
[24] MD Breed LE Snyder TL Lynn JA Morhart Acquirechemical camouflage in a tropical ant Anim Behav 44 (1992519ndash523
[25] MD Breed TP McGlynn EM Stocker ANKlein Thiefworkers and variation in nestmate recognition behavior iponerine antEctatomma ruidum Insectes Soc 46 (1999) 327331
[26] I Perfecto JH Vandermeer Cleptobiosis in the antEc-tatomma ruidum in Nicaragua Insectes Soc 40 (1993) 29299
[27] JM Jeral MD Breed BE Hibbard Thief ants hareduced quantities of cuticular compounds in a poneant Ectatomma ruidum Physiol Entomol 22 (1997) 207211
[28] DE WheelerEctatomma tuberculatum foraging biology andassociation withCrematogaster (Hymenoptera Formicidae)Ann Entomol Soc Am 79 (1986) 300ndash303
[29] WL Jr Brown Contributions toward a reclassification of tFormicidae II Tribe Ectatommini (Hymenoptera) Bull MuComp Zool 118 (1958) 173ndash362
[30] A Dejean J-P Lachaud Growth-related changes in predatiobehavior in incipient colonies of the ponerine antEctatommatuberculatum (Olivier) Insectes Soc 39 (1992) 129ndash143
[31] J Valenzuela-Gonzaacutelez A Loacutepez-Meacutendez J-P Lachaudtivity patterns and foraging activity in nests ofEctatommatuberculatum (Hymenoptera Formicidae) in cacao plantatioSouthwest Entomol 20 (1995) 507ndash515
[32] RR da Hora R Feacuteneacuteron J Valenzuela ME FavilaFresneau Queen-size dimorphism in the antEctatomma
tuberculatum (Hymenoptera Formicidae Ponerinae) Sociobiology 38 (2001) 407ndash420
[33] G Ibarra-Nuacutentildeez JA Garciacutea JA Loacutepez J-P Lachaud Panalysis in the diet of some ponerine ants (HymenoptFormicidae) and web-building spiders (Araneae) in coffeplantations in Chiapas Mexico Sociobiology 37 (2001723ndash755
[34] DW Davidson Ecological studies of Neotropical ant gardenEcology 69 (1988) 1138ndash1152
[35] A Dejean B Corbara J Orivel RR Snelling JHC Debie M Belin-Depoux The importance of ant gardens inpioneer vegetal formations of French Guiana (HymenoptFormicidae) Sociobiology 35 (2000) 425ndash439
[36] LM Pascal EF Motte-Florac DB McKey Secretory strutures on the leaf rachis of Caesalpinieae and Mimosoid(Leguminosae) implication for the evolution of nectaglands Am J Bot 87 (2000) 327ndash338
[37] B Schatz J-P Lachaud G Beugnon Polyethism withihunters of the ponerine antEctatomma ruidum InsectesSoc 43 (1996) 111ndash118
[38] WM Wheeler The compound and mixed nests of Amecan ants Part I Observations on a new guest ant Am Na(1901) 431ndash448
[39] B Houmllldobler Liquid food transmission and antennatisignals in ponerine ants Isr J Entomol 19 (1985) 89ndash99
[40] J Liebig J Heinze B Houmllldobler Trophallaxis and aggrsion in the ponerine antPonera coartata implications forthe evolution of liquid food exchange in the HymenopteEthology 103 (1997) 707ndash722
[41] C Bevan A Birch H Caswell Ant insect repellent froblack cocktail ants J Chem Soc 961 (1961) 488
[42] AB Attygalle B Siegel O Vostrowsky HJ BestmanU Maschwitz Chemical composition and function of mepleural gland secretion of the antCrematogaster deformisSmith (Hymenoptera Myrmicinae) J Chem Ecol 15 (19317ndash328
[43] D Daloze M Kaisin C Detrain JM Pasteels Chemicalfense in the three European species ofCrematogaster antsExperientia 47 (1991) 1082ndash1089
[44] B Houmllldobler Chemische Strategie beim Nahrungserwerb deDiebameise (Solenopsis fugax Latr) und der Pharaoameis(Monomorium pharaonis L) Oecologia 11 (1973) 371ndash380
[45] A Lenoir C Errard A Francoeur R Loiselle Relations entre la fourmi parasiteFormicoxenus provancheri et son hocircteMyrmica incompleta Donneacutees biologiques et eacutethologiqu(Hym Formicidae) Insectes Soc 39 (1992) 81ndash97
[46] J Orivel C Errard A Dejean Ant gardens interspecrecognition in parabiotic species Behav Ecol Sociobiol(1997) 87ndash93
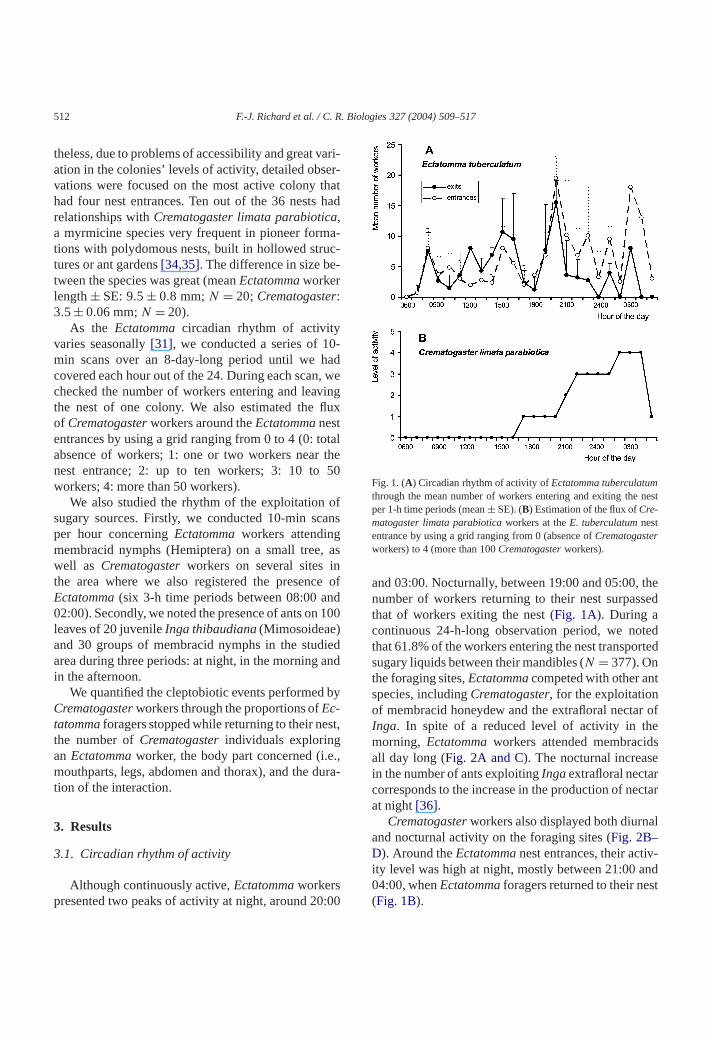
512 F-J Richard et al C R Biologies 327 (2004) 509ndash517
ari-er-hathad
a-c--
0-ad weingux
talthe50
ofans
asne ond100
iednd
by
st
ura-
00
nest
thesed
tedrted
t
r ofes
e
ctar
al
-ndst
theless due to problems of accessibility and great vation in the coloniesrsquo levels of activity detailed obsvations were focused on the most active colony thad four nest entrances Ten out of the 36 nestsrelationships withCrematogaster limata parabioticaa myrmicine species very frequent in pioneer formtions with polydomous nests built in hollowed strutures or ant gardens[3435] The difference in size between the species was great (meanEctatomma workerlengthplusmn SE 95 plusmn 08 mmN = 20 Crematogaster35plusmn 006 mmN = 20)
As the Ectatomma circadian rhythm of activityvaries seasonally[31] we conducted a series of 1min scans over an 8-day-long period until we hcovered each hour out of the 24 During each scanchecked the number of workers entering and leavthe nest of one colony We also estimated the flof Crematogaster workers around theEctatomma nestentrances by using a grid ranging from 0 to 4 (0 toabsence of workers 1 one or two workers nearnest entrance 2 up to ten workers 3 10 toworkers 4 more than 50 workers)
We also studied the rhythm of the exploitationsugary sources Firstly we conducted 10-min scper hour concerningEctatomma workers attendingmembracid nymphs (Hemiptera) on a small treewell as Crematogaster workers on several sites ithe area where we also registered the presencEctatomma (six 3-h time periods between 0800 a0200) Secondly we noted the presence of ants onleaves of 20 juvenileInga thibaudiana (Mimosoideae)and 30 groups of membracid nymphs in the studarea during three periods at night in the morning ain the afternoon
We quantified the cleptobiotic events performedCrematogaster workers through the proportions ofEc-tatomma foragers stopped while returning to their nethe number ofCrematogaster individuals exploringan Ectatomma worker the body part concerned (iemouthparts legs abdomen and thorax) and the dtion of the interaction
3 Results
31 Circadian rhythm of activity
Although continuously activeEctatomma workerspresented two peaks of activity at night around 20
f
Fig 1 (A) Circadian rhythm of activity ofEctatomma tuberculatumthrough the mean number of workers entering and exiting theper 1-h time periods (meanplusmn SE) (B) Estimation of the flux ofCre-matogaster limata parabiotica workers at theE tuberculatum nestentrance by using a grid ranging from 0 (absence ofCrematogasterworkers) to 4 (more than 100Crematogaster workers)
and 0300 Nocturnally between 1900 and 0500number of workers returning to their nest surpasthat of workers exiting the nest (Fig 1A) During acontinuous 24-h-long observation period we nothat 618 of the workers entering the nest transposugary liquids between their mandibles (N = 377) Onthe foraging sitesEctatomma competed with other anspecies includingCrematogaster for the exploitationof membracid honeydew and the extrafloral nectaInga In spite of a reduced level of activity in thmorning Ectatomma workers attended membracidall day long (Fig 2A and C) The nocturnal increasin the number of ants exploitingInga extrafloral nectarcorresponds to the increase in the production of neat night[36]
Crematogaster workers also displayed both diurnand nocturnal activity on the foraging sites (Fig 2BndashD) Around theEctatomma nest entrances their activity level was high at night mostly between 2100 a0400 whenEctatomma foragers returned to their ne(Fig 1B)
F-J Richard et al C R Biologies 327 (2004) 509ndash517 513
fntsg
Fig 2 (A) Dynamics of the exploitation of membracid nymphs byEctatomma tuberculatum workers at one site during a 24-h cycle (B) Thesame forCrematogaster limata parabiotica workers recorded at different sites between 0800 and 0200 (C) Percentages of groups omembracid nymphs attended by ants during three periods of the day (other speciesCamponotus spp 100 of the groups attended by aat 1000 and 2300 933 at 1500 hours) (D) Percentages of leaves ofInga thibaudiana (Mimosoideae) noted with ant workers exploitinthe extrafloral nectaries during three periods of the day (other species mostlyCamponotus spp but also someGigantiops destructor andCephalotes sp in the morning total of 39 7 and 72 of the leaves with ants at 1000 1500 and 2300 respectively)
theen-me
t ofm
the
to- Al-tedof
ps
cidve-eytlytothsedof
eenypec-
tledewr fortheion
32 Ants attending membracids
In the studied area bothCrematogaster and Ec-tatomma foragers attended membracid nymphs onsame trees during the same time periods but gerally on different branches They shared the saparts of the branches on only 25 occasions ou194 noted during the study of their circadian rhythof activity In such cases theCrematogaster attendedthe membracid individuals the most distant fromEctatomma foragers and they moved when anEc-tatomma approached The latter frequently triedcapture theCrematogaster workers 15 attempts occurred on the 25 branches shared by both speciesthough most of them successfully escaped we noduring a series of observations that 14 prey out27 retrieved byE tuberculatum foragers were ants10 of themCrematogaster Also four retrieved preywere membracids originating from attended groutwo complete predatory sequences being observed
During trophobiotic relationshipsEctatommaworkers palpated the lateral parts of the membranymphs with their antennae in an up and down moment eliciting the secretion of honeydew As thstood above the nymphs with their mandibles slighopen they recuperated the honeydew that adheredtheir mandibles thanks to surface tension strengWhile repeating this behaviour the workers opentheir mandibles wider and wider as the dropletshoneydew increased in size (Fig 3A) Then in 701of the cases (N = 127) eachEctatomma forager re-trieved the droplet of honeydew accumulated betwits mandibles after having been replaced or not ba nestmate (252 and 449 of the cases restively) In the other situations (299)Ectatommaworkers specialized in honeydew transport shutbetween the individuals that collected the honeydfrom the membracids and the nest (one transporte4ndash7 collectors) The transfer of honeydew fromcollector to the transporter followed an antennat
514 F-J Richard et al C R Biologies 327 (2004) 509ndash517
ed
Fig 3 (A) Ectatomma tuberculatum worker attending membracid nymphs and gathering a honeydew droplet between its outstretchmandibles (B) E tuberculatum worker stopped by twoCrematogaster limata parabiotica workers inspecting its mouthparts (C) Workerof C limata parabiotica licking a part of the honeydew droplet from aE tuberculatum forager
s ofenidsItewen
ab-
n aredthe
ost-p
nest
re-spedrs at-idly
initiated by the transporter that also made a serie3ndash4 back and forth movements with its body Ththe collecting worker again attended the membracwhile the transporter looked for other collectorsreturned to the nest when the droplet of honeydcompletely filled the space separating its wide-opmandibles
33 Relationships between Crematogaster limataparabioticaand Ectatomma tuberculatum
While completely absent in the vicinity of theEc-tatomma nest entrances during the daytime (Fig 1B)at duskCrematogaster workers entered theEctatommanests without any opposition as the guards weresent As already reported[28] the column ofCre-
matogaster passed through the entrance tunnels iraid-like fashion The beginning of such raids occurbetween 1800 and 2245 which corresponds topeak of nocturnal activity ofEctatomma (Fig 1A) Af-ter these raids theCrematogaster workers exited thetunnel and remained around the nest entrance almall night long (Fig 1B) They never had a swollen abdomen nor any evidence of having filled their croThen they stopped returningEctatomma foragers atthe base of the supporting tree trunk around theentrance or on surrounding branchesCrematogasterworkers waved their abdomens at the approach of aturningEctatomma forager but no liquid droplet wanoted at the tip of their abdomens They also taptheir abdomens on the substrate These behavioutracted nestmates that immediately arrived and rap
F-J Richard et al C R Biologies 327 (2004) 509ndash517 515
ed
80
42r-ringhereng
id
-
)
e-sed
ual
eto
dies
to or
hentter
heer-
n
mersquoy
edn-
fniv-hethe
din-
-reted
y-t
histheur
d-
on-il-
gr
tsr anri-
c-
cal-l or
aggregated around theEctatomma CertainCremato-gaster individuals stayed in front of theEctatommathat remained immobile while up to 12 others climbon theEctatomma (32plusmn 24 N = 98)
A study conducted on 322Ectatomma foragers re-turning to their nest permitted us to note that only(248) were not intercepted by theCrematogasterworkers prior to the tunnel entrance Among the 2Ectatomma intercepted 116 (36 of all returning foagers) were stopped and remained immobile dua short time before resuming their return trip to tnest The remaining 126 individuals (391) westopped explored and licked during a relatively lotime (gt1 min) by theCrematogaster workers Thisincluded 111 individuals carrying a droplet of liqufood that was stolen (345)
Of the 316Crematogaster workers thoroughly observed after the interception of anEctatomma for-ager laden with a droplet of liquid food 165 (522focused their exploration on theEctatomma mouth-parts (seeFig 3B and C) Each individual lickedthe honeydew only during a few seconds (66 plusmn54 s N = 70) When the honeydew droplet was dpleted theEctatomma workers remained motionlesmandibles open maxilo-labial mouthparts extendallowing theCrematogaster workers to lick them dur-ing a long time (25plusmn 145 minN = 15)
4 Discussion
We confirm therefore the existence of the unusrelationship betweenE tuberculatum andC limata asreported in central Panama[28] Nevertheless somnew information and significant differences deservebe pointed out
First Ectatomma workers constantly attendegroups of membracid nymphs thanks to two strategWorkers collected honeydew that they transportedthe nest once they were replaced by a nestmatecertain workers collected the honeydew that was ttransported to the nest by other workers The lacooperative behaviour known in the Formicinae[3]presents similarities with the lsquorelay transportrsquo and tlsquoprey chain transfer behaviourrsquo noted for other ponine ants during predation[3738]
Second we noted a dramatic difference inCremato-gaster-Ectatomma interspecific relationships betwee
day and night Although sharing branches with theCrematogaster workers maintained a lsquosafe distancfrom Ectatomma in the foraging areas during the dathe latter being able to prey on them (see also[33])On the contrary at night theCrematogaster work-ers no longer avoided direct contact withE tubercu-latum but initiated the interactions They conducttheir raids on theEctatomma nest entrances then egaged in contact with theEctatomma foragers return-ing to their nest stopping and exploring them
Third Wheeler[28] noted that the behaviour oCrematogaster workers during raids was limited itime and consisted of a kind of lsquoshampooingrsquo dering nutrition from the cuticular secretions andor texcess fluids (nectar and honeydew) trapped onmandibles ofEctatomma workers This author notethat such behaviour first described between anquiline ant and its host[39] could account for a mutualistic association of the lsquocleaner-fishrsquo type wheboth species derive benefits Even if we again nothat Crematogaster workers licked the different bodparts of theEctatomma they mostly stopped and explored theEctatomma retrieving a honeydew drople(881 of the exploredE tuberculatum foragers) andrecruited nestmates in order to rob this droplet Tresulted therefore in a parasitic relationship oflsquocleptobioticrsquo form This nocturnal robbing behavioonly complements the food supply of theCremato-gaster that gather most of their sugary food by attening hemiptera or exploiting extrafloral nectaries
Fourth other cases of cleptobiosis in ants ccerned prey robbing with nevertheless a certain simitude with the present study For exampleMyrmeco-cystus mimicus workers thoroughly inspect returninforagers ofPogonomyrmex spp before robbing theiprey [20] The originality of the cleptobiotic activityof C limata parabiotica consists in exploiting the kindof liquid transport limited to arboreal ponerine anIndeed they carry droplets of liquid between theimandibles which are then imbibed by nestmatesadaptation to the lack of trophallaxis in most Ponenae[4041]
Finally we questioned the origin of the lower noturnal aggressiveness ofEctatomma workers towardsCrematogaster The immobility of the Ectatommaworkers could be triggered by a secreted chemiwhile the Crematogaster wave their abdomens Indeed secretions from the mandibular metapleura
516 F-J Richard et al C R Biologies 327 (2004) 509ndash517
d
id-
plet
he
ur-
ack
at-o-be
p-ac-p-
ire
-oure
nebio-hem
sisceso-e
ACa--the
entGIFs raquo
ds
ty
lm
a-bo5ndash
c02)
man
rrsquos
of-im
ter-star
ction9)
n-
02)
of82
iest
ar-
nd0
Dufour glands act as repellents in severalCremato-gaster species[42ndash44] and such an effect was notein the lestobioticSolenopsis fugax [45] Similarly apacifying action on the host occurs when an indivual of the parasiticFormicoxenus provancheri raisesits abdomen and extrudes the sting producing a droof volatile liquid before engaging in lsquoshampooingrsquo be-haviour withMyrmica incompleta [46]
Nevertheless why does this not work during tdaytime As theCrematogaster lsquoraidsrsquo coincide withthe dusk peak in activity ofEctatomma (1900ndash2100Fig 1A) another hypothesis can be formulated Ding this period the heavy flow ofEctatomma foragersentering and leaving the tunnel drives the guards binto the nest This permits the smallCrematogasterworkers to creep inside the tunnel without beingtacked In this situation they can modify the compsition of their cuticular hydrocarbons enough toaccepted by the nextEctatomma foragers returningto their nest and so to lure them A similar decetive mechanism based on chemical camouflagequisition was proposed to explain intraspecific cletobiosis inE ruidum [27] Later newCrematogasterindividuals can enter the tunnel or they can acqucuticular compounds directly from an interceptedEc-tatomma worker by climbing on it and licking its cuticle this corresponding to the lsquoshampooingrsquo behavidescribed by Wheeler[28] This hypothesis might bsupported by the fact thatC limata parabiotica has theability to share nests and trails with different formiciand ponerine ants in a phenomenon called lsquoparasisrsquo while these ants never share nests between t[343546]
In conclusion we noted a new kind of cleptobioin ants concerning the robbing of sugary substanand not prey robbing as previously known Temprally limited to night time and facultative it implies thpresence ofEctatomma nests onCrematogaster terri-tories
Acknowledgements
We are grateful to Dr Jacques HC Delabie (UPLaboratoacuterio de Mirmecologia Convecircnio UESCEPLAC Itabuna Bahia Brazil) for the identifiction of the ants and to AndreaDejean for proofreading the manuscript This work was supported by
French laquo Ministegravere de lrsquoEacutecologie et du Deacuteveloppemdurable raquo (SOFT program research agreementECOFOR No 98 and the laquo Tropical Ecosystemprogram No 02-E2002)
References
[1] E Curio in The Ethology ofPredation Springer-VerlagBerlin 1976 242 p
[2] HJ Brockmann CJ Barnard Kleptoparasitism in birAnim Behav 27 (1979) 487ndash514
[3] B Houmllldobler EO Wilson in The Ants Harvard UniversiPress Cambridge 1990 732 p
[4] TJ Pitcher Functions of shoaling behaviour in teleosts inJ Pitcher (Ed) The Behaviour of Teleost Fishes Croom HeLondon 1986 pp 294ndash337
[5] WJ Dominey AM Snyder Kleptoparasitism of freshwter crabs by cichlid fishes endemic to Lake Barombi MCameroon West Africa Environ Biol Fishes 22 (1988) 15160
[6] JR Garrido CG Sarasa M Fernandez-Cruz Intraspecifikleptoparasitism in the cattle egret J Field Ornithol 73 (20185ndash190
[7] CE Khatchikian M Favero AI Vassalo Kleptoparasitisby brown-hooded gulls and grey-hooded gulls on Americoystercatchers Waterbirds 25 (2002) 137ndash141
[8] MS Di Bitetti CH Janson Social foraging and the findeshare in capuchin monkeysCebus apella Anim Behav 62(2001) 47ndash56
[9] OP Honer B Wachter ML East H Hofer The responsespotted hyenas to long-term changes in prey populations functional response and interspecific kleptoparasitism J AnEcol 71 (2002) 236ndash246
[10] S Morissette JH Himmelman Subtidal food thieves inactions of four invertabrate kleptoparasites with the seaLeptasterias polaris Anim Behav 60 (2000) 531ndash543
[11] Y Heacutenaut J-P Lachaud Araigneacutees cleptoparasites seacutelede lrsquohocircte par sa toile Ann Soc Entomol Fr (NS) 35 (199148ndash153
[12] M Whitehouse I Agnarsson T Miyashita D Smith K Cagialosi T Masumoto D Li Y HeacutenautArgyrodes phylogenysociality and interspecific interactions J Arachnol 30 (20238ndash245
[13] B Crespi P Abbot The behavioral ecology and evolutionkleptoparasitism in Australian gall thrips Fla Entomol(1999) 147ndash164
[14] FE Kurczewski MG Spofford Alternative nesting strategin Ammophila urnaria (Hymenoptera Sphecidae) J NaHist 32 (1998) 99ndash106
[15] C Michener in The Social Behavior of the Bees A Compative Study The Belknap Pressof Harvard University PressCambridge 1974 404 p
[16] WM Wheeler in Ants Their Structure Development aBehaviour Columbia University Press New York 191663 p
F-J Richard et al C R Biologies 327 (2004) 509ndash517 517
)
in al 20
ing
89ndash
mic
ry
d)
n andash
5ndash
verinendash
hes
n
Ac-
ns
D
-
reyerae)
s
la-theera
c-eaery
n
ri-t 35
on
es-
ra
m
nta-
89)
de-
re
-
es
ific40
[17] A Forel Fourmis termitophages LestobioseAtta tardigradasous-genres drsquoEuponera Ann Soc Entomol Belg 45 (1901389ndash398
[18] T Abe On the food sharing among four species of antssandy grassland I Food and foraging behavior Jap J Eco(1971) 219ndash230
[19] U Maschwitz M MuhlenbergCamponotus rufoglaucus einewegelagernde Ameise Zool Anz 191 (1973) 364ndash368
[20] B Houmllldobler Food robbing inants a form of interferencecompetition Oecologia 69 (1986) 12ndash15
[21] T Yamaguchi Intraspecific competition through food robbin the harvester antMessor aciculatus (Fr Smith) and itsconsequences on colony survival Insectes Soc 42 (1995)101
[22] NA Weber Two common ponerine ants of possible econosignificanceEctatomma tuberculatum (Olivier) andE ruidumRoger Proc Entomol Soc Washington 48 (1946) 1ndash16
[23] MD Breed P Abel TJ Bleuze SE Denton Thievehome ranges and nestmate recognition inEctatomma ruidumOecologia 84 (1990) 117ndash121
[24] MD Breed LE Snyder TL Lynn JA Morhart Acquirechemical camouflage in a tropical ant Anim Behav 44 (1992519ndash523
[25] MD Breed TP McGlynn EM Stocker ANKlein Thiefworkers and variation in nestmate recognition behavior iponerine antEctatomma ruidum Insectes Soc 46 (1999) 327331
[26] I Perfecto JH Vandermeer Cleptobiosis in the antEc-tatomma ruidum in Nicaragua Insectes Soc 40 (1993) 29299
[27] JM Jeral MD Breed BE Hibbard Thief ants hareduced quantities of cuticular compounds in a poneant Ectatomma ruidum Physiol Entomol 22 (1997) 207211
[28] DE WheelerEctatomma tuberculatum foraging biology andassociation withCrematogaster (Hymenoptera Formicidae)Ann Entomol Soc Am 79 (1986) 300ndash303
[29] WL Jr Brown Contributions toward a reclassification of tFormicidae II Tribe Ectatommini (Hymenoptera) Bull MuComp Zool 118 (1958) 173ndash362
[30] A Dejean J-P Lachaud Growth-related changes in predatiobehavior in incipient colonies of the ponerine antEctatommatuberculatum (Olivier) Insectes Soc 39 (1992) 129ndash143
[31] J Valenzuela-Gonzaacutelez A Loacutepez-Meacutendez J-P Lachaudtivity patterns and foraging activity in nests ofEctatommatuberculatum (Hymenoptera Formicidae) in cacao plantatioSouthwest Entomol 20 (1995) 507ndash515
[32] RR da Hora R Feacuteneacuteron J Valenzuela ME FavilaFresneau Queen-size dimorphism in the antEctatomma
tuberculatum (Hymenoptera Formicidae Ponerinae) Sociobiology 38 (2001) 407ndash420
[33] G Ibarra-Nuacutentildeez JA Garciacutea JA Loacutepez J-P Lachaud Panalysis in the diet of some ponerine ants (HymenoptFormicidae) and web-building spiders (Araneae) in coffeplantations in Chiapas Mexico Sociobiology 37 (2001723ndash755
[34] DW Davidson Ecological studies of Neotropical ant gardenEcology 69 (1988) 1138ndash1152
[35] A Dejean B Corbara J Orivel RR Snelling JHC Debie M Belin-Depoux The importance of ant gardens inpioneer vegetal formations of French Guiana (HymenoptFormicidae) Sociobiology 35 (2000) 425ndash439
[36] LM Pascal EF Motte-Florac DB McKey Secretory strutures on the leaf rachis of Caesalpinieae and Mimosoid(Leguminosae) implication for the evolution of nectaglands Am J Bot 87 (2000) 327ndash338
[37] B Schatz J-P Lachaud G Beugnon Polyethism withihunters of the ponerine antEctatomma ruidum InsectesSoc 43 (1996) 111ndash118
[38] WM Wheeler The compound and mixed nests of Amecan ants Part I Observations on a new guest ant Am Na(1901) 431ndash448
[39] B Houmllldobler Liquid food transmission and antennatisignals in ponerine ants Isr J Entomol 19 (1985) 89ndash99
[40] J Liebig J Heinze B Houmllldobler Trophallaxis and aggrsion in the ponerine antPonera coartata implications forthe evolution of liquid food exchange in the HymenopteEthology 103 (1997) 707ndash722
[41] C Bevan A Birch H Caswell Ant insect repellent froblack cocktail ants J Chem Soc 961 (1961) 488
[42] AB Attygalle B Siegel O Vostrowsky HJ BestmanU Maschwitz Chemical composition and function of mepleural gland secretion of the antCrematogaster deformisSmith (Hymenoptera Myrmicinae) J Chem Ecol 15 (19317ndash328
[43] D Daloze M Kaisin C Detrain JM Pasteels Chemicalfense in the three European species ofCrematogaster antsExperientia 47 (1991) 1082ndash1089
[44] B Houmllldobler Chemische Strategie beim Nahrungserwerb deDiebameise (Solenopsis fugax Latr) und der Pharaoameis(Monomorium pharaonis L) Oecologia 11 (1973) 371ndash380
[45] A Lenoir C Errard A Francoeur R Loiselle Relations entre la fourmi parasiteFormicoxenus provancheri et son hocircteMyrmica incompleta Donneacutees biologiques et eacutethologiqu(Hym Formicidae) Insectes Soc 39 (1992) 81ndash97
[46] J Orivel C Errard A Dejean Ant gardens interspecrecognition in parabiotic species Behav Ecol Sociobiol(1997) 87ndash93
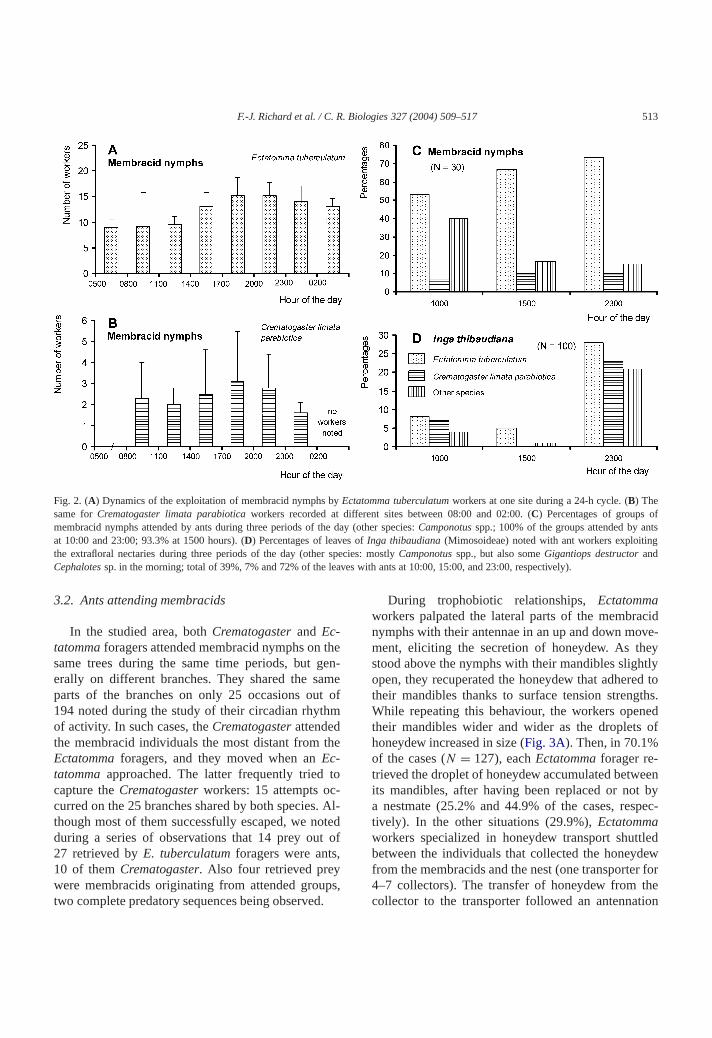
F-J Richard et al C R Biologies 327 (2004) 509ndash517 513
fntsg
Fig 2 (A) Dynamics of the exploitation of membracid nymphs byEctatomma tuberculatum workers at one site during a 24-h cycle (B) Thesame forCrematogaster limata parabiotica workers recorded at different sites between 0800 and 0200 (C) Percentages of groups omembracid nymphs attended by ants during three periods of the day (other speciesCamponotus spp 100 of the groups attended by aat 1000 and 2300 933 at 1500 hours) (D) Percentages of leaves ofInga thibaudiana (Mimosoideae) noted with ant workers exploitinthe extrafloral nectaries during three periods of the day (other species mostlyCamponotus spp but also someGigantiops destructor andCephalotes sp in the morning total of 39 7 and 72 of the leaves with ants at 1000 1500 and 2300 respectively)
theen-me
t ofm
the
to- Al-tedof
ps
cidve-eytlytothsedof
eenypec-
tledewr fortheion
32 Ants attending membracids
In the studied area bothCrematogaster and Ec-tatomma foragers attended membracid nymphs onsame trees during the same time periods but gerally on different branches They shared the saparts of the branches on only 25 occasions ou194 noted during the study of their circadian rhythof activity In such cases theCrematogaster attendedthe membracid individuals the most distant fromEctatomma foragers and they moved when anEc-tatomma approached The latter frequently triedcapture theCrematogaster workers 15 attempts occurred on the 25 branches shared by both speciesthough most of them successfully escaped we noduring a series of observations that 14 prey out27 retrieved byE tuberculatum foragers were ants10 of themCrematogaster Also four retrieved preywere membracids originating from attended groutwo complete predatory sequences being observed
During trophobiotic relationshipsEctatommaworkers palpated the lateral parts of the membranymphs with their antennae in an up and down moment eliciting the secretion of honeydew As thstood above the nymphs with their mandibles slighopen they recuperated the honeydew that adheredtheir mandibles thanks to surface tension strengWhile repeating this behaviour the workers opentheir mandibles wider and wider as the dropletshoneydew increased in size (Fig 3A) Then in 701of the cases (N = 127) eachEctatomma forager re-trieved the droplet of honeydew accumulated betwits mandibles after having been replaced or not ba nestmate (252 and 449 of the cases restively) In the other situations (299)Ectatommaworkers specialized in honeydew transport shutbetween the individuals that collected the honeydfrom the membracids and the nest (one transporte4ndash7 collectors) The transfer of honeydew fromcollector to the transporter followed an antennat
514 F-J Richard et al C R Biologies 327 (2004) 509ndash517
ed
Fig 3 (A) Ectatomma tuberculatum worker attending membracid nymphs and gathering a honeydew droplet between its outstretchmandibles (B) E tuberculatum worker stopped by twoCrematogaster limata parabiotica workers inspecting its mouthparts (C) Workerof C limata parabiotica licking a part of the honeydew droplet from aE tuberculatum forager
s ofenidsItewen
ab-
n aredthe
ost-p
nest
re-spedrs at-idly
initiated by the transporter that also made a serie3ndash4 back and forth movements with its body Ththe collecting worker again attended the membracwhile the transporter looked for other collectorsreturned to the nest when the droplet of honeydcompletely filled the space separating its wide-opmandibles
33 Relationships between Crematogaster limataparabioticaand Ectatomma tuberculatum
While completely absent in the vicinity of theEc-tatomma nest entrances during the daytime (Fig 1B)at duskCrematogaster workers entered theEctatommanests without any opposition as the guards weresent As already reported[28] the column ofCre-
matogaster passed through the entrance tunnels iraid-like fashion The beginning of such raids occurbetween 1800 and 2245 which corresponds topeak of nocturnal activity ofEctatomma (Fig 1A) Af-ter these raids theCrematogaster workers exited thetunnel and remained around the nest entrance almall night long (Fig 1B) They never had a swollen abdomen nor any evidence of having filled their croThen they stopped returningEctatomma foragers atthe base of the supporting tree trunk around theentrance or on surrounding branchesCrematogasterworkers waved their abdomens at the approach of aturningEctatomma forager but no liquid droplet wanoted at the tip of their abdomens They also taptheir abdomens on the substrate These behavioutracted nestmates that immediately arrived and rap
F-J Richard et al C R Biologies 327 (2004) 509ndash517 515
ed
80
42r-ringhereng
id
-
)
e-sed
ual
eto
dies
to or
hentter
heer-
n
mersquoy
edn-
fniv-hethe
din-
-reted
y-t
histheur
d-
on-il-
gr
tsr anri-
c-
cal-l or
aggregated around theEctatomma CertainCremato-gaster individuals stayed in front of theEctatommathat remained immobile while up to 12 others climbon theEctatomma (32plusmn 24 N = 98)
A study conducted on 322Ectatomma foragers re-turning to their nest permitted us to note that only(248) were not intercepted by theCrematogasterworkers prior to the tunnel entrance Among the 2Ectatomma intercepted 116 (36 of all returning foagers) were stopped and remained immobile dua short time before resuming their return trip to tnest The remaining 126 individuals (391) westopped explored and licked during a relatively lotime (gt1 min) by theCrematogaster workers Thisincluded 111 individuals carrying a droplet of liqufood that was stolen (345)
Of the 316Crematogaster workers thoroughly observed after the interception of anEctatomma for-ager laden with a droplet of liquid food 165 (522focused their exploration on theEctatomma mouth-parts (seeFig 3B and C) Each individual lickedthe honeydew only during a few seconds (66 plusmn54 s N = 70) When the honeydew droplet was dpleted theEctatomma workers remained motionlesmandibles open maxilo-labial mouthparts extendallowing theCrematogaster workers to lick them dur-ing a long time (25plusmn 145 minN = 15)
4 Discussion
We confirm therefore the existence of the unusrelationship betweenE tuberculatum andC limata asreported in central Panama[28] Nevertheless somnew information and significant differences deservebe pointed out
First Ectatomma workers constantly attendegroups of membracid nymphs thanks to two strategWorkers collected honeydew that they transportedthe nest once they were replaced by a nestmatecertain workers collected the honeydew that was ttransported to the nest by other workers The lacooperative behaviour known in the Formicinae[3]presents similarities with the lsquorelay transportrsquo and tlsquoprey chain transfer behaviourrsquo noted for other ponine ants during predation[3738]
Second we noted a dramatic difference inCremato-gaster-Ectatomma interspecific relationships betwee
day and night Although sharing branches with theCrematogaster workers maintained a lsquosafe distancfrom Ectatomma in the foraging areas during the dathe latter being able to prey on them (see also[33])On the contrary at night theCrematogaster work-ers no longer avoided direct contact withE tubercu-latum but initiated the interactions They conducttheir raids on theEctatomma nest entrances then egaged in contact with theEctatomma foragers return-ing to their nest stopping and exploring them
Third Wheeler[28] noted that the behaviour oCrematogaster workers during raids was limited itime and consisted of a kind of lsquoshampooingrsquo dering nutrition from the cuticular secretions andor texcess fluids (nectar and honeydew) trapped onmandibles ofEctatomma workers This author notethat such behaviour first described between anquiline ant and its host[39] could account for a mutualistic association of the lsquocleaner-fishrsquo type wheboth species derive benefits Even if we again nothat Crematogaster workers licked the different bodparts of theEctatomma they mostly stopped and explored theEctatomma retrieving a honeydew drople(881 of the exploredE tuberculatum foragers) andrecruited nestmates in order to rob this droplet Tresulted therefore in a parasitic relationship oflsquocleptobioticrsquo form This nocturnal robbing behavioonly complements the food supply of theCremato-gaster that gather most of their sugary food by attening hemiptera or exploiting extrafloral nectaries
Fourth other cases of cleptobiosis in ants ccerned prey robbing with nevertheless a certain simitude with the present study For exampleMyrmeco-cystus mimicus workers thoroughly inspect returninforagers ofPogonomyrmex spp before robbing theiprey [20] The originality of the cleptobiotic activityof C limata parabiotica consists in exploiting the kindof liquid transport limited to arboreal ponerine anIndeed they carry droplets of liquid between theimandibles which are then imbibed by nestmatesadaptation to the lack of trophallaxis in most Ponenae[4041]
Finally we questioned the origin of the lower noturnal aggressiveness ofEctatomma workers towardsCrematogaster The immobility of the Ectatommaworkers could be triggered by a secreted chemiwhile the Crematogaster wave their abdomens Indeed secretions from the mandibular metapleura
516 F-J Richard et al C R Biologies 327 (2004) 509ndash517
d
id-
plet
he
ur-
ack
at-o-be
p-ac-p-
ire
-oure
nebio-hem
sisceso-e
ACa--the
entGIFs raquo
ds
ty
lm
a-bo5ndash
c02)
man
rrsquos
of-im
ter-star
ction9)
n-
02)
of82
iest
ar-
nd0
Dufour glands act as repellents in severalCremato-gaster species[42ndash44] and such an effect was notein the lestobioticSolenopsis fugax [45] Similarly apacifying action on the host occurs when an indivual of the parasiticFormicoxenus provancheri raisesits abdomen and extrudes the sting producing a droof volatile liquid before engaging in lsquoshampooingrsquo be-haviour withMyrmica incompleta [46]
Nevertheless why does this not work during tdaytime As theCrematogaster lsquoraidsrsquo coincide withthe dusk peak in activity ofEctatomma (1900ndash2100Fig 1A) another hypothesis can be formulated Ding this period the heavy flow ofEctatomma foragersentering and leaving the tunnel drives the guards binto the nest This permits the smallCrematogasterworkers to creep inside the tunnel without beingtacked In this situation they can modify the compsition of their cuticular hydrocarbons enough toaccepted by the nextEctatomma foragers returningto their nest and so to lure them A similar decetive mechanism based on chemical camouflagequisition was proposed to explain intraspecific cletobiosis inE ruidum [27] Later newCrematogasterindividuals can enter the tunnel or they can acqucuticular compounds directly from an interceptedEc-tatomma worker by climbing on it and licking its cuticle this corresponding to the lsquoshampooingrsquo behavidescribed by Wheeler[28] This hypothesis might bsupported by the fact thatC limata parabiotica has theability to share nests and trails with different formiciand ponerine ants in a phenomenon called lsquoparasisrsquo while these ants never share nests between t[343546]
In conclusion we noted a new kind of cleptobioin ants concerning the robbing of sugary substanand not prey robbing as previously known Temprally limited to night time and facultative it implies thpresence ofEctatomma nests onCrematogaster terri-tories
Acknowledgements
We are grateful to Dr Jacques HC Delabie (UPLaboratoacuterio de Mirmecologia Convecircnio UESCEPLAC Itabuna Bahia Brazil) for the identifiction of the ants and to AndreaDejean for proofreading the manuscript This work was supported by
French laquo Ministegravere de lrsquoEacutecologie et du Deacuteveloppemdurable raquo (SOFT program research agreementECOFOR No 98 and the laquo Tropical Ecosystemprogram No 02-E2002)
References
[1] E Curio in The Ethology ofPredation Springer-VerlagBerlin 1976 242 p
[2] HJ Brockmann CJ Barnard Kleptoparasitism in birAnim Behav 27 (1979) 487ndash514
[3] B Houmllldobler EO Wilson in The Ants Harvard UniversiPress Cambridge 1990 732 p
[4] TJ Pitcher Functions of shoaling behaviour in teleosts inJ Pitcher (Ed) The Behaviour of Teleost Fishes Croom HeLondon 1986 pp 294ndash337
[5] WJ Dominey AM Snyder Kleptoparasitism of freshwter crabs by cichlid fishes endemic to Lake Barombi MCameroon West Africa Environ Biol Fishes 22 (1988) 15160
[6] JR Garrido CG Sarasa M Fernandez-Cruz Intraspecifikleptoparasitism in the cattle egret J Field Ornithol 73 (20185ndash190
[7] CE Khatchikian M Favero AI Vassalo Kleptoparasitisby brown-hooded gulls and grey-hooded gulls on Americoystercatchers Waterbirds 25 (2002) 137ndash141
[8] MS Di Bitetti CH Janson Social foraging and the findeshare in capuchin monkeysCebus apella Anim Behav 62(2001) 47ndash56
[9] OP Honer B Wachter ML East H Hofer The responsespotted hyenas to long-term changes in prey populations functional response and interspecific kleptoparasitism J AnEcol 71 (2002) 236ndash246
[10] S Morissette JH Himmelman Subtidal food thieves inactions of four invertabrate kleptoparasites with the seaLeptasterias polaris Anim Behav 60 (2000) 531ndash543
[11] Y Heacutenaut J-P Lachaud Araigneacutees cleptoparasites seacutelede lrsquohocircte par sa toile Ann Soc Entomol Fr (NS) 35 (199148ndash153
[12] M Whitehouse I Agnarsson T Miyashita D Smith K Cagialosi T Masumoto D Li Y HeacutenautArgyrodes phylogenysociality and interspecific interactions J Arachnol 30 (20238ndash245
[13] B Crespi P Abbot The behavioral ecology and evolutionkleptoparasitism in Australian gall thrips Fla Entomol(1999) 147ndash164
[14] FE Kurczewski MG Spofford Alternative nesting strategin Ammophila urnaria (Hymenoptera Sphecidae) J NaHist 32 (1998) 99ndash106
[15] C Michener in The Social Behavior of the Bees A Compative Study The Belknap Pressof Harvard University PressCambridge 1974 404 p
[16] WM Wheeler in Ants Their Structure Development aBehaviour Columbia University Press New York 191663 p
F-J Richard et al C R Biologies 327 (2004) 509ndash517 517
)
in al 20
ing
89ndash
mic
ry
d)
n andash
5ndash
verinendash
hes
n
Ac-
ns
D
-
reyerae)
s
la-theera
c-eaery
n
ri-t 35
on
es-
ra
m
nta-
89)
de-
re
-
es
ific40
[17] A Forel Fourmis termitophages LestobioseAtta tardigradasous-genres drsquoEuponera Ann Soc Entomol Belg 45 (1901389ndash398
[18] T Abe On the food sharing among four species of antssandy grassland I Food and foraging behavior Jap J Eco(1971) 219ndash230
[19] U Maschwitz M MuhlenbergCamponotus rufoglaucus einewegelagernde Ameise Zool Anz 191 (1973) 364ndash368
[20] B Houmllldobler Food robbing inants a form of interferencecompetition Oecologia 69 (1986) 12ndash15
[21] T Yamaguchi Intraspecific competition through food robbin the harvester antMessor aciculatus (Fr Smith) and itsconsequences on colony survival Insectes Soc 42 (1995)101
[22] NA Weber Two common ponerine ants of possible econosignificanceEctatomma tuberculatum (Olivier) andE ruidumRoger Proc Entomol Soc Washington 48 (1946) 1ndash16
[23] MD Breed P Abel TJ Bleuze SE Denton Thievehome ranges and nestmate recognition inEctatomma ruidumOecologia 84 (1990) 117ndash121
[24] MD Breed LE Snyder TL Lynn JA Morhart Acquirechemical camouflage in a tropical ant Anim Behav 44 (1992519ndash523
[25] MD Breed TP McGlynn EM Stocker ANKlein Thiefworkers and variation in nestmate recognition behavior iponerine antEctatomma ruidum Insectes Soc 46 (1999) 327331
[26] I Perfecto JH Vandermeer Cleptobiosis in the antEc-tatomma ruidum in Nicaragua Insectes Soc 40 (1993) 29299
[27] JM Jeral MD Breed BE Hibbard Thief ants hareduced quantities of cuticular compounds in a poneant Ectatomma ruidum Physiol Entomol 22 (1997) 207211
[28] DE WheelerEctatomma tuberculatum foraging biology andassociation withCrematogaster (Hymenoptera Formicidae)Ann Entomol Soc Am 79 (1986) 300ndash303
[29] WL Jr Brown Contributions toward a reclassification of tFormicidae II Tribe Ectatommini (Hymenoptera) Bull MuComp Zool 118 (1958) 173ndash362
[30] A Dejean J-P Lachaud Growth-related changes in predatiobehavior in incipient colonies of the ponerine antEctatommatuberculatum (Olivier) Insectes Soc 39 (1992) 129ndash143
[31] J Valenzuela-Gonzaacutelez A Loacutepez-Meacutendez J-P Lachaudtivity patterns and foraging activity in nests ofEctatommatuberculatum (Hymenoptera Formicidae) in cacao plantatioSouthwest Entomol 20 (1995) 507ndash515
[32] RR da Hora R Feacuteneacuteron J Valenzuela ME FavilaFresneau Queen-size dimorphism in the antEctatomma
tuberculatum (Hymenoptera Formicidae Ponerinae) Sociobiology 38 (2001) 407ndash420
[33] G Ibarra-Nuacutentildeez JA Garciacutea JA Loacutepez J-P Lachaud Panalysis in the diet of some ponerine ants (HymenoptFormicidae) and web-building spiders (Araneae) in coffeplantations in Chiapas Mexico Sociobiology 37 (2001723ndash755
[34] DW Davidson Ecological studies of Neotropical ant gardenEcology 69 (1988) 1138ndash1152
[35] A Dejean B Corbara J Orivel RR Snelling JHC Debie M Belin-Depoux The importance of ant gardens inpioneer vegetal formations of French Guiana (HymenoptFormicidae) Sociobiology 35 (2000) 425ndash439
[36] LM Pascal EF Motte-Florac DB McKey Secretory strutures on the leaf rachis of Caesalpinieae and Mimosoid(Leguminosae) implication for the evolution of nectaglands Am J Bot 87 (2000) 327ndash338
[37] B Schatz J-P Lachaud G Beugnon Polyethism withihunters of the ponerine antEctatomma ruidum InsectesSoc 43 (1996) 111ndash118
[38] WM Wheeler The compound and mixed nests of Amecan ants Part I Observations on a new guest ant Am Na(1901) 431ndash448
[39] B Houmllldobler Liquid food transmission and antennatisignals in ponerine ants Isr J Entomol 19 (1985) 89ndash99
[40] J Liebig J Heinze B Houmllldobler Trophallaxis and aggrsion in the ponerine antPonera coartata implications forthe evolution of liquid food exchange in the HymenopteEthology 103 (1997) 707ndash722
[41] C Bevan A Birch H Caswell Ant insect repellent froblack cocktail ants J Chem Soc 961 (1961) 488
[42] AB Attygalle B Siegel O Vostrowsky HJ BestmanU Maschwitz Chemical composition and function of mepleural gland secretion of the antCrematogaster deformisSmith (Hymenoptera Myrmicinae) J Chem Ecol 15 (19317ndash328
[43] D Daloze M Kaisin C Detrain JM Pasteels Chemicalfense in the three European species ofCrematogaster antsExperientia 47 (1991) 1082ndash1089
[44] B Houmllldobler Chemische Strategie beim Nahrungserwerb deDiebameise (Solenopsis fugax Latr) und der Pharaoameis(Monomorium pharaonis L) Oecologia 11 (1973) 371ndash380
[45] A Lenoir C Errard A Francoeur R Loiselle Relations entre la fourmi parasiteFormicoxenus provancheri et son hocircteMyrmica incompleta Donneacutees biologiques et eacutethologiqu(Hym Formicidae) Insectes Soc 39 (1992) 81ndash97
[46] J Orivel C Errard A Dejean Ant gardens interspecrecognition in parabiotic species Behav Ecol Sociobiol(1997) 87ndash93

514 F-J Richard et al C R Biologies 327 (2004) 509ndash517
ed
Fig 3 (A) Ectatomma tuberculatum worker attending membracid nymphs and gathering a honeydew droplet between its outstretchmandibles (B) E tuberculatum worker stopped by twoCrematogaster limata parabiotica workers inspecting its mouthparts (C) Workerof C limata parabiotica licking a part of the honeydew droplet from aE tuberculatum forager
s ofenidsItewen
ab-
n aredthe
ost-p
nest
re-spedrs at-idly
initiated by the transporter that also made a serie3ndash4 back and forth movements with its body Ththe collecting worker again attended the membracwhile the transporter looked for other collectorsreturned to the nest when the droplet of honeydcompletely filled the space separating its wide-opmandibles
33 Relationships between Crematogaster limataparabioticaand Ectatomma tuberculatum
While completely absent in the vicinity of theEc-tatomma nest entrances during the daytime (Fig 1B)at duskCrematogaster workers entered theEctatommanests without any opposition as the guards weresent As already reported[28] the column ofCre-
matogaster passed through the entrance tunnels iraid-like fashion The beginning of such raids occurbetween 1800 and 2245 which corresponds topeak of nocturnal activity ofEctatomma (Fig 1A) Af-ter these raids theCrematogaster workers exited thetunnel and remained around the nest entrance almall night long (Fig 1B) They never had a swollen abdomen nor any evidence of having filled their croThen they stopped returningEctatomma foragers atthe base of the supporting tree trunk around theentrance or on surrounding branchesCrematogasterworkers waved their abdomens at the approach of aturningEctatomma forager but no liquid droplet wanoted at the tip of their abdomens They also taptheir abdomens on the substrate These behavioutracted nestmates that immediately arrived and rap
F-J Richard et al C R Biologies 327 (2004) 509ndash517 515
ed
80
42r-ringhereng
id
-
)
e-sed
ual
eto
dies
to or
hentter
heer-
n
mersquoy
edn-
fniv-hethe
din-
-reted
y-t
histheur
d-
on-il-
gr
tsr anri-
c-
cal-l or
aggregated around theEctatomma CertainCremato-gaster individuals stayed in front of theEctatommathat remained immobile while up to 12 others climbon theEctatomma (32plusmn 24 N = 98)
A study conducted on 322Ectatomma foragers re-turning to their nest permitted us to note that only(248) were not intercepted by theCrematogasterworkers prior to the tunnel entrance Among the 2Ectatomma intercepted 116 (36 of all returning foagers) were stopped and remained immobile dua short time before resuming their return trip to tnest The remaining 126 individuals (391) westopped explored and licked during a relatively lotime (gt1 min) by theCrematogaster workers Thisincluded 111 individuals carrying a droplet of liqufood that was stolen (345)
Of the 316Crematogaster workers thoroughly observed after the interception of anEctatomma for-ager laden with a droplet of liquid food 165 (522focused their exploration on theEctatomma mouth-parts (seeFig 3B and C) Each individual lickedthe honeydew only during a few seconds (66 plusmn54 s N = 70) When the honeydew droplet was dpleted theEctatomma workers remained motionlesmandibles open maxilo-labial mouthparts extendallowing theCrematogaster workers to lick them dur-ing a long time (25plusmn 145 minN = 15)
4 Discussion
We confirm therefore the existence of the unusrelationship betweenE tuberculatum andC limata asreported in central Panama[28] Nevertheless somnew information and significant differences deservebe pointed out
First Ectatomma workers constantly attendegroups of membracid nymphs thanks to two strategWorkers collected honeydew that they transportedthe nest once they were replaced by a nestmatecertain workers collected the honeydew that was ttransported to the nest by other workers The lacooperative behaviour known in the Formicinae[3]presents similarities with the lsquorelay transportrsquo and tlsquoprey chain transfer behaviourrsquo noted for other ponine ants during predation[3738]
Second we noted a dramatic difference inCremato-gaster-Ectatomma interspecific relationships betwee
day and night Although sharing branches with theCrematogaster workers maintained a lsquosafe distancfrom Ectatomma in the foraging areas during the dathe latter being able to prey on them (see also[33])On the contrary at night theCrematogaster work-ers no longer avoided direct contact withE tubercu-latum but initiated the interactions They conducttheir raids on theEctatomma nest entrances then egaged in contact with theEctatomma foragers return-ing to their nest stopping and exploring them
Third Wheeler[28] noted that the behaviour oCrematogaster workers during raids was limited itime and consisted of a kind of lsquoshampooingrsquo dering nutrition from the cuticular secretions andor texcess fluids (nectar and honeydew) trapped onmandibles ofEctatomma workers This author notethat such behaviour first described between anquiline ant and its host[39] could account for a mutualistic association of the lsquocleaner-fishrsquo type wheboth species derive benefits Even if we again nothat Crematogaster workers licked the different bodparts of theEctatomma they mostly stopped and explored theEctatomma retrieving a honeydew drople(881 of the exploredE tuberculatum foragers) andrecruited nestmates in order to rob this droplet Tresulted therefore in a parasitic relationship oflsquocleptobioticrsquo form This nocturnal robbing behavioonly complements the food supply of theCremato-gaster that gather most of their sugary food by attening hemiptera or exploiting extrafloral nectaries
Fourth other cases of cleptobiosis in ants ccerned prey robbing with nevertheless a certain simitude with the present study For exampleMyrmeco-cystus mimicus workers thoroughly inspect returninforagers ofPogonomyrmex spp before robbing theiprey [20] The originality of the cleptobiotic activityof C limata parabiotica consists in exploiting the kindof liquid transport limited to arboreal ponerine anIndeed they carry droplets of liquid between theimandibles which are then imbibed by nestmatesadaptation to the lack of trophallaxis in most Ponenae[4041]
Finally we questioned the origin of the lower noturnal aggressiveness ofEctatomma workers towardsCrematogaster The immobility of the Ectatommaworkers could be triggered by a secreted chemiwhile the Crematogaster wave their abdomens Indeed secretions from the mandibular metapleura
516 F-J Richard et al C R Biologies 327 (2004) 509ndash517
d
id-
plet
he
ur-
ack
at-o-be
p-ac-p-
ire
-oure
nebio-hem
sisceso-e
ACa--the
entGIFs raquo
ds
ty
lm
a-bo5ndash
c02)
man
rrsquos
of-im
ter-star
ction9)
n-
02)
of82
iest
ar-
nd0
Dufour glands act as repellents in severalCremato-gaster species[42ndash44] and such an effect was notein the lestobioticSolenopsis fugax [45] Similarly apacifying action on the host occurs when an indivual of the parasiticFormicoxenus provancheri raisesits abdomen and extrudes the sting producing a droof volatile liquid before engaging in lsquoshampooingrsquo be-haviour withMyrmica incompleta [46]
Nevertheless why does this not work during tdaytime As theCrematogaster lsquoraidsrsquo coincide withthe dusk peak in activity ofEctatomma (1900ndash2100Fig 1A) another hypothesis can be formulated Ding this period the heavy flow ofEctatomma foragersentering and leaving the tunnel drives the guards binto the nest This permits the smallCrematogasterworkers to creep inside the tunnel without beingtacked In this situation they can modify the compsition of their cuticular hydrocarbons enough toaccepted by the nextEctatomma foragers returningto their nest and so to lure them A similar decetive mechanism based on chemical camouflagequisition was proposed to explain intraspecific cletobiosis inE ruidum [27] Later newCrematogasterindividuals can enter the tunnel or they can acqucuticular compounds directly from an interceptedEc-tatomma worker by climbing on it and licking its cuticle this corresponding to the lsquoshampooingrsquo behavidescribed by Wheeler[28] This hypothesis might bsupported by the fact thatC limata parabiotica has theability to share nests and trails with different formiciand ponerine ants in a phenomenon called lsquoparasisrsquo while these ants never share nests between t[343546]
In conclusion we noted a new kind of cleptobioin ants concerning the robbing of sugary substanand not prey robbing as previously known Temprally limited to night time and facultative it implies thpresence ofEctatomma nests onCrematogaster terri-tories
Acknowledgements
We are grateful to Dr Jacques HC Delabie (UPLaboratoacuterio de Mirmecologia Convecircnio UESCEPLAC Itabuna Bahia Brazil) for the identifiction of the ants and to AndreaDejean for proofreading the manuscript This work was supported by
French laquo Ministegravere de lrsquoEacutecologie et du Deacuteveloppemdurable raquo (SOFT program research agreementECOFOR No 98 and the laquo Tropical Ecosystemprogram No 02-E2002)
References
[1] E Curio in The Ethology ofPredation Springer-VerlagBerlin 1976 242 p
[2] HJ Brockmann CJ Barnard Kleptoparasitism in birAnim Behav 27 (1979) 487ndash514
[3] B Houmllldobler EO Wilson in The Ants Harvard UniversiPress Cambridge 1990 732 p
[4] TJ Pitcher Functions of shoaling behaviour in teleosts inJ Pitcher (Ed) The Behaviour of Teleost Fishes Croom HeLondon 1986 pp 294ndash337
[5] WJ Dominey AM Snyder Kleptoparasitism of freshwter crabs by cichlid fishes endemic to Lake Barombi MCameroon West Africa Environ Biol Fishes 22 (1988) 15160
[6] JR Garrido CG Sarasa M Fernandez-Cruz Intraspecifikleptoparasitism in the cattle egret J Field Ornithol 73 (20185ndash190
[7] CE Khatchikian M Favero AI Vassalo Kleptoparasitisby brown-hooded gulls and grey-hooded gulls on Americoystercatchers Waterbirds 25 (2002) 137ndash141
[8] MS Di Bitetti CH Janson Social foraging and the findeshare in capuchin monkeysCebus apella Anim Behav 62(2001) 47ndash56
[9] OP Honer B Wachter ML East H Hofer The responsespotted hyenas to long-term changes in prey populations functional response and interspecific kleptoparasitism J AnEcol 71 (2002) 236ndash246
[10] S Morissette JH Himmelman Subtidal food thieves inactions of four invertabrate kleptoparasites with the seaLeptasterias polaris Anim Behav 60 (2000) 531ndash543
[11] Y Heacutenaut J-P Lachaud Araigneacutees cleptoparasites seacutelede lrsquohocircte par sa toile Ann Soc Entomol Fr (NS) 35 (199148ndash153
[12] M Whitehouse I Agnarsson T Miyashita D Smith K Cagialosi T Masumoto D Li Y HeacutenautArgyrodes phylogenysociality and interspecific interactions J Arachnol 30 (20238ndash245
[13] B Crespi P Abbot The behavioral ecology and evolutionkleptoparasitism in Australian gall thrips Fla Entomol(1999) 147ndash164
[14] FE Kurczewski MG Spofford Alternative nesting strategin Ammophila urnaria (Hymenoptera Sphecidae) J NaHist 32 (1998) 99ndash106
[15] C Michener in The Social Behavior of the Bees A Compative Study The Belknap Pressof Harvard University PressCambridge 1974 404 p
[16] WM Wheeler in Ants Their Structure Development aBehaviour Columbia University Press New York 191663 p
F-J Richard et al C R Biologies 327 (2004) 509ndash517 517
)
in al 20
ing
89ndash
mic
ry
d)
n andash
5ndash
verinendash
hes
n
Ac-
ns
D
-
reyerae)
s
la-theera
c-eaery
n
ri-t 35
on
es-
ra
m
nta-
89)
de-
re
-
es
ific40
[17] A Forel Fourmis termitophages LestobioseAtta tardigradasous-genres drsquoEuponera Ann Soc Entomol Belg 45 (1901389ndash398
[18] T Abe On the food sharing among four species of antssandy grassland I Food and foraging behavior Jap J Eco(1971) 219ndash230
[19] U Maschwitz M MuhlenbergCamponotus rufoglaucus einewegelagernde Ameise Zool Anz 191 (1973) 364ndash368
[20] B Houmllldobler Food robbing inants a form of interferencecompetition Oecologia 69 (1986) 12ndash15
[21] T Yamaguchi Intraspecific competition through food robbin the harvester antMessor aciculatus (Fr Smith) and itsconsequences on colony survival Insectes Soc 42 (1995)101
[22] NA Weber Two common ponerine ants of possible econosignificanceEctatomma tuberculatum (Olivier) andE ruidumRoger Proc Entomol Soc Washington 48 (1946) 1ndash16
[23] MD Breed P Abel TJ Bleuze SE Denton Thievehome ranges and nestmate recognition inEctatomma ruidumOecologia 84 (1990) 117ndash121
[24] MD Breed LE Snyder TL Lynn JA Morhart Acquirechemical camouflage in a tropical ant Anim Behav 44 (1992519ndash523
[25] MD Breed TP McGlynn EM Stocker ANKlein Thiefworkers and variation in nestmate recognition behavior iponerine antEctatomma ruidum Insectes Soc 46 (1999) 327331
[26] I Perfecto JH Vandermeer Cleptobiosis in the antEc-tatomma ruidum in Nicaragua Insectes Soc 40 (1993) 29299
[27] JM Jeral MD Breed BE Hibbard Thief ants hareduced quantities of cuticular compounds in a poneant Ectatomma ruidum Physiol Entomol 22 (1997) 207211
[28] DE WheelerEctatomma tuberculatum foraging biology andassociation withCrematogaster (Hymenoptera Formicidae)Ann Entomol Soc Am 79 (1986) 300ndash303
[29] WL Jr Brown Contributions toward a reclassification of tFormicidae II Tribe Ectatommini (Hymenoptera) Bull MuComp Zool 118 (1958) 173ndash362
[30] A Dejean J-P Lachaud Growth-related changes in predatiobehavior in incipient colonies of the ponerine antEctatommatuberculatum (Olivier) Insectes Soc 39 (1992) 129ndash143
[31] J Valenzuela-Gonzaacutelez A Loacutepez-Meacutendez J-P Lachaudtivity patterns and foraging activity in nests ofEctatommatuberculatum (Hymenoptera Formicidae) in cacao plantatioSouthwest Entomol 20 (1995) 507ndash515
[32] RR da Hora R Feacuteneacuteron J Valenzuela ME FavilaFresneau Queen-size dimorphism in the antEctatomma
tuberculatum (Hymenoptera Formicidae Ponerinae) Sociobiology 38 (2001) 407ndash420
[33] G Ibarra-Nuacutentildeez JA Garciacutea JA Loacutepez J-P Lachaud Panalysis in the diet of some ponerine ants (HymenoptFormicidae) and web-building spiders (Araneae) in coffeplantations in Chiapas Mexico Sociobiology 37 (2001723ndash755
[34] DW Davidson Ecological studies of Neotropical ant gardenEcology 69 (1988) 1138ndash1152
[35] A Dejean B Corbara J Orivel RR Snelling JHC Debie M Belin-Depoux The importance of ant gardens inpioneer vegetal formations of French Guiana (HymenoptFormicidae) Sociobiology 35 (2000) 425ndash439
[36] LM Pascal EF Motte-Florac DB McKey Secretory strutures on the leaf rachis of Caesalpinieae and Mimosoid(Leguminosae) implication for the evolution of nectaglands Am J Bot 87 (2000) 327ndash338
[37] B Schatz J-P Lachaud G Beugnon Polyethism withihunters of the ponerine antEctatomma ruidum InsectesSoc 43 (1996) 111ndash118
[38] WM Wheeler The compound and mixed nests of Amecan ants Part I Observations on a new guest ant Am Na(1901) 431ndash448
[39] B Houmllldobler Liquid food transmission and antennatisignals in ponerine ants Isr J Entomol 19 (1985) 89ndash99
[40] J Liebig J Heinze B Houmllldobler Trophallaxis and aggrsion in the ponerine antPonera coartata implications forthe evolution of liquid food exchange in the HymenopteEthology 103 (1997) 707ndash722
[41] C Bevan A Birch H Caswell Ant insect repellent froblack cocktail ants J Chem Soc 961 (1961) 488
[42] AB Attygalle B Siegel O Vostrowsky HJ BestmanU Maschwitz Chemical composition and function of mepleural gland secretion of the antCrematogaster deformisSmith (Hymenoptera Myrmicinae) J Chem Ecol 15 (19317ndash328
[43] D Daloze M Kaisin C Detrain JM Pasteels Chemicalfense in the three European species ofCrematogaster antsExperientia 47 (1991) 1082ndash1089
[44] B Houmllldobler Chemische Strategie beim Nahrungserwerb deDiebameise (Solenopsis fugax Latr) und der Pharaoameis(Monomorium pharaonis L) Oecologia 11 (1973) 371ndash380
[45] A Lenoir C Errard A Francoeur R Loiselle Relations entre la fourmi parasiteFormicoxenus provancheri et son hocircteMyrmica incompleta Donneacutees biologiques et eacutethologiqu(Hym Formicidae) Insectes Soc 39 (1992) 81ndash97
[46] J Orivel C Errard A Dejean Ant gardens interspecrecognition in parabiotic species Behav Ecol Sociobiol(1997) 87ndash93

F-J Richard et al C R Biologies 327 (2004) 509ndash517 515
ed
80
42r-ringhereng
id
-
)
e-sed
ual
eto
dies
to or
hentter
heer-
n
mersquoy
edn-
fniv-hethe
din-
-reted
y-t
histheur
d-
on-il-
gr
tsr anri-
c-
cal-l or
aggregated around theEctatomma CertainCremato-gaster individuals stayed in front of theEctatommathat remained immobile while up to 12 others climbon theEctatomma (32plusmn 24 N = 98)
A study conducted on 322Ectatomma foragers re-turning to their nest permitted us to note that only(248) were not intercepted by theCrematogasterworkers prior to the tunnel entrance Among the 2Ectatomma intercepted 116 (36 of all returning foagers) were stopped and remained immobile dua short time before resuming their return trip to tnest The remaining 126 individuals (391) westopped explored and licked during a relatively lotime (gt1 min) by theCrematogaster workers Thisincluded 111 individuals carrying a droplet of liqufood that was stolen (345)
Of the 316Crematogaster workers thoroughly observed after the interception of anEctatomma for-ager laden with a droplet of liquid food 165 (522focused their exploration on theEctatomma mouth-parts (seeFig 3B and C) Each individual lickedthe honeydew only during a few seconds (66 plusmn54 s N = 70) When the honeydew droplet was dpleted theEctatomma workers remained motionlesmandibles open maxilo-labial mouthparts extendallowing theCrematogaster workers to lick them dur-ing a long time (25plusmn 145 minN = 15)
4 Discussion
We confirm therefore the existence of the unusrelationship betweenE tuberculatum andC limata asreported in central Panama[28] Nevertheless somnew information and significant differences deservebe pointed out
First Ectatomma workers constantly attendegroups of membracid nymphs thanks to two strategWorkers collected honeydew that they transportedthe nest once they were replaced by a nestmatecertain workers collected the honeydew that was ttransported to the nest by other workers The lacooperative behaviour known in the Formicinae[3]presents similarities with the lsquorelay transportrsquo and tlsquoprey chain transfer behaviourrsquo noted for other ponine ants during predation[3738]
Second we noted a dramatic difference inCremato-gaster-Ectatomma interspecific relationships betwee
day and night Although sharing branches with theCrematogaster workers maintained a lsquosafe distancfrom Ectatomma in the foraging areas during the dathe latter being able to prey on them (see also[33])On the contrary at night theCrematogaster work-ers no longer avoided direct contact withE tubercu-latum but initiated the interactions They conducttheir raids on theEctatomma nest entrances then egaged in contact with theEctatomma foragers return-ing to their nest stopping and exploring them
Third Wheeler[28] noted that the behaviour oCrematogaster workers during raids was limited itime and consisted of a kind of lsquoshampooingrsquo dering nutrition from the cuticular secretions andor texcess fluids (nectar and honeydew) trapped onmandibles ofEctatomma workers This author notethat such behaviour first described between anquiline ant and its host[39] could account for a mutualistic association of the lsquocleaner-fishrsquo type wheboth species derive benefits Even if we again nothat Crematogaster workers licked the different bodparts of theEctatomma they mostly stopped and explored theEctatomma retrieving a honeydew drople(881 of the exploredE tuberculatum foragers) andrecruited nestmates in order to rob this droplet Tresulted therefore in a parasitic relationship oflsquocleptobioticrsquo form This nocturnal robbing behavioonly complements the food supply of theCremato-gaster that gather most of their sugary food by attening hemiptera or exploiting extrafloral nectaries
Fourth other cases of cleptobiosis in ants ccerned prey robbing with nevertheless a certain simitude with the present study For exampleMyrmeco-cystus mimicus workers thoroughly inspect returninforagers ofPogonomyrmex spp before robbing theiprey [20] The originality of the cleptobiotic activityof C limata parabiotica consists in exploiting the kindof liquid transport limited to arboreal ponerine anIndeed they carry droplets of liquid between theimandibles which are then imbibed by nestmatesadaptation to the lack of trophallaxis in most Ponenae[4041]
Finally we questioned the origin of the lower noturnal aggressiveness ofEctatomma workers towardsCrematogaster The immobility of the Ectatommaworkers could be triggered by a secreted chemiwhile the Crematogaster wave their abdomens Indeed secretions from the mandibular metapleura
516 F-J Richard et al C R Biologies 327 (2004) 509ndash517
d
id-
plet
he
ur-
ack
at-o-be
p-ac-p-
ire
-oure
nebio-hem
sisceso-e
ACa--the
entGIFs raquo
ds
ty
lm
a-bo5ndash
c02)
man
rrsquos
of-im
ter-star
ction9)
n-
02)
of82
iest
ar-
nd0
Dufour glands act as repellents in severalCremato-gaster species[42ndash44] and such an effect was notein the lestobioticSolenopsis fugax [45] Similarly apacifying action on the host occurs when an indivual of the parasiticFormicoxenus provancheri raisesits abdomen and extrudes the sting producing a droof volatile liquid before engaging in lsquoshampooingrsquo be-haviour withMyrmica incompleta [46]
Nevertheless why does this not work during tdaytime As theCrematogaster lsquoraidsrsquo coincide withthe dusk peak in activity ofEctatomma (1900ndash2100Fig 1A) another hypothesis can be formulated Ding this period the heavy flow ofEctatomma foragersentering and leaving the tunnel drives the guards binto the nest This permits the smallCrematogasterworkers to creep inside the tunnel without beingtacked In this situation they can modify the compsition of their cuticular hydrocarbons enough toaccepted by the nextEctatomma foragers returningto their nest and so to lure them A similar decetive mechanism based on chemical camouflagequisition was proposed to explain intraspecific cletobiosis inE ruidum [27] Later newCrematogasterindividuals can enter the tunnel or they can acqucuticular compounds directly from an interceptedEc-tatomma worker by climbing on it and licking its cuticle this corresponding to the lsquoshampooingrsquo behavidescribed by Wheeler[28] This hypothesis might bsupported by the fact thatC limata parabiotica has theability to share nests and trails with different formiciand ponerine ants in a phenomenon called lsquoparasisrsquo while these ants never share nests between t[343546]
In conclusion we noted a new kind of cleptobioin ants concerning the robbing of sugary substanand not prey robbing as previously known Temprally limited to night time and facultative it implies thpresence ofEctatomma nests onCrematogaster terri-tories
Acknowledgements
We are grateful to Dr Jacques HC Delabie (UPLaboratoacuterio de Mirmecologia Convecircnio UESCEPLAC Itabuna Bahia Brazil) for the identifiction of the ants and to AndreaDejean for proofreading the manuscript This work was supported by
French laquo Ministegravere de lrsquoEacutecologie et du Deacuteveloppemdurable raquo (SOFT program research agreementECOFOR No 98 and the laquo Tropical Ecosystemprogram No 02-E2002)
References
[1] E Curio in The Ethology ofPredation Springer-VerlagBerlin 1976 242 p
[2] HJ Brockmann CJ Barnard Kleptoparasitism in birAnim Behav 27 (1979) 487ndash514
[3] B Houmllldobler EO Wilson in The Ants Harvard UniversiPress Cambridge 1990 732 p
[4] TJ Pitcher Functions of shoaling behaviour in teleosts inJ Pitcher (Ed) The Behaviour of Teleost Fishes Croom HeLondon 1986 pp 294ndash337
[5] WJ Dominey AM Snyder Kleptoparasitism of freshwter crabs by cichlid fishes endemic to Lake Barombi MCameroon West Africa Environ Biol Fishes 22 (1988) 15160
[6] JR Garrido CG Sarasa M Fernandez-Cruz Intraspecifikleptoparasitism in the cattle egret J Field Ornithol 73 (20185ndash190
[7] CE Khatchikian M Favero AI Vassalo Kleptoparasitisby brown-hooded gulls and grey-hooded gulls on Americoystercatchers Waterbirds 25 (2002) 137ndash141
[8] MS Di Bitetti CH Janson Social foraging and the findeshare in capuchin monkeysCebus apella Anim Behav 62(2001) 47ndash56
[9] OP Honer B Wachter ML East H Hofer The responsespotted hyenas to long-term changes in prey populations functional response and interspecific kleptoparasitism J AnEcol 71 (2002) 236ndash246
[10] S Morissette JH Himmelman Subtidal food thieves inactions of four invertabrate kleptoparasites with the seaLeptasterias polaris Anim Behav 60 (2000) 531ndash543
[11] Y Heacutenaut J-P Lachaud Araigneacutees cleptoparasites seacutelede lrsquohocircte par sa toile Ann Soc Entomol Fr (NS) 35 (199148ndash153
[12] M Whitehouse I Agnarsson T Miyashita D Smith K Cagialosi T Masumoto D Li Y HeacutenautArgyrodes phylogenysociality and interspecific interactions J Arachnol 30 (20238ndash245
[13] B Crespi P Abbot The behavioral ecology and evolutionkleptoparasitism in Australian gall thrips Fla Entomol(1999) 147ndash164
[14] FE Kurczewski MG Spofford Alternative nesting strategin Ammophila urnaria (Hymenoptera Sphecidae) J NaHist 32 (1998) 99ndash106
[15] C Michener in The Social Behavior of the Bees A Compative Study The Belknap Pressof Harvard University PressCambridge 1974 404 p
[16] WM Wheeler in Ants Their Structure Development aBehaviour Columbia University Press New York 191663 p
F-J Richard et al C R Biologies 327 (2004) 509ndash517 517
)
in al 20
ing
89ndash
mic
ry
d)
n andash
5ndash
verinendash
hes
n
Ac-
ns
D
-
reyerae)
s
la-theera
c-eaery
n
ri-t 35
on
es-
ra
m
nta-
89)
de-
re
-
es
ific40
[17] A Forel Fourmis termitophages LestobioseAtta tardigradasous-genres drsquoEuponera Ann Soc Entomol Belg 45 (1901389ndash398
[18] T Abe On the food sharing among four species of antssandy grassland I Food and foraging behavior Jap J Eco(1971) 219ndash230
[19] U Maschwitz M MuhlenbergCamponotus rufoglaucus einewegelagernde Ameise Zool Anz 191 (1973) 364ndash368
[20] B Houmllldobler Food robbing inants a form of interferencecompetition Oecologia 69 (1986) 12ndash15
[21] T Yamaguchi Intraspecific competition through food robbin the harvester antMessor aciculatus (Fr Smith) and itsconsequences on colony survival Insectes Soc 42 (1995)101
[22] NA Weber Two common ponerine ants of possible econosignificanceEctatomma tuberculatum (Olivier) andE ruidumRoger Proc Entomol Soc Washington 48 (1946) 1ndash16
[23] MD Breed P Abel TJ Bleuze SE Denton Thievehome ranges and nestmate recognition inEctatomma ruidumOecologia 84 (1990) 117ndash121
[24] MD Breed LE Snyder TL Lynn JA Morhart Acquirechemical camouflage in a tropical ant Anim Behav 44 (1992519ndash523
[25] MD Breed TP McGlynn EM Stocker ANKlein Thiefworkers and variation in nestmate recognition behavior iponerine antEctatomma ruidum Insectes Soc 46 (1999) 327331
[26] I Perfecto JH Vandermeer Cleptobiosis in the antEc-tatomma ruidum in Nicaragua Insectes Soc 40 (1993) 29299
[27] JM Jeral MD Breed BE Hibbard Thief ants hareduced quantities of cuticular compounds in a poneant Ectatomma ruidum Physiol Entomol 22 (1997) 207211
[28] DE WheelerEctatomma tuberculatum foraging biology andassociation withCrematogaster (Hymenoptera Formicidae)Ann Entomol Soc Am 79 (1986) 300ndash303
[29] WL Jr Brown Contributions toward a reclassification of tFormicidae II Tribe Ectatommini (Hymenoptera) Bull MuComp Zool 118 (1958) 173ndash362
[30] A Dejean J-P Lachaud Growth-related changes in predatiobehavior in incipient colonies of the ponerine antEctatommatuberculatum (Olivier) Insectes Soc 39 (1992) 129ndash143
[31] J Valenzuela-Gonzaacutelez A Loacutepez-Meacutendez J-P Lachaudtivity patterns and foraging activity in nests ofEctatommatuberculatum (Hymenoptera Formicidae) in cacao plantatioSouthwest Entomol 20 (1995) 507ndash515
[32] RR da Hora R Feacuteneacuteron J Valenzuela ME FavilaFresneau Queen-size dimorphism in the antEctatomma
tuberculatum (Hymenoptera Formicidae Ponerinae) Sociobiology 38 (2001) 407ndash420
[33] G Ibarra-Nuacutentildeez JA Garciacutea JA Loacutepez J-P Lachaud Panalysis in the diet of some ponerine ants (HymenoptFormicidae) and web-building spiders (Araneae) in coffeplantations in Chiapas Mexico Sociobiology 37 (2001723ndash755
[34] DW Davidson Ecological studies of Neotropical ant gardenEcology 69 (1988) 1138ndash1152
[35] A Dejean B Corbara J Orivel RR Snelling JHC Debie M Belin-Depoux The importance of ant gardens inpioneer vegetal formations of French Guiana (HymenoptFormicidae) Sociobiology 35 (2000) 425ndash439
[36] LM Pascal EF Motte-Florac DB McKey Secretory strutures on the leaf rachis of Caesalpinieae and Mimosoid(Leguminosae) implication for the evolution of nectaglands Am J Bot 87 (2000) 327ndash338
[37] B Schatz J-P Lachaud G Beugnon Polyethism withihunters of the ponerine antEctatomma ruidum InsectesSoc 43 (1996) 111ndash118
[38] WM Wheeler The compound and mixed nests of Amecan ants Part I Observations on a new guest ant Am Na(1901) 431ndash448
[39] B Houmllldobler Liquid food transmission and antennatisignals in ponerine ants Isr J Entomol 19 (1985) 89ndash99
[40] J Liebig J Heinze B Houmllldobler Trophallaxis and aggrsion in the ponerine antPonera coartata implications forthe evolution of liquid food exchange in the HymenopteEthology 103 (1997) 707ndash722
[41] C Bevan A Birch H Caswell Ant insect repellent froblack cocktail ants J Chem Soc 961 (1961) 488
[42] AB Attygalle B Siegel O Vostrowsky HJ BestmanU Maschwitz Chemical composition and function of mepleural gland secretion of the antCrematogaster deformisSmith (Hymenoptera Myrmicinae) J Chem Ecol 15 (19317ndash328
[43] D Daloze M Kaisin C Detrain JM Pasteels Chemicalfense in the three European species ofCrematogaster antsExperientia 47 (1991) 1082ndash1089
[44] B Houmllldobler Chemische Strategie beim Nahrungserwerb deDiebameise (Solenopsis fugax Latr) und der Pharaoameis(Monomorium pharaonis L) Oecologia 11 (1973) 371ndash380
[45] A Lenoir C Errard A Francoeur R Loiselle Relations entre la fourmi parasiteFormicoxenus provancheri et son hocircteMyrmica incompleta Donneacutees biologiques et eacutethologiqu(Hym Formicidae) Insectes Soc 39 (1992) 81ndash97
[46] J Orivel C Errard A Dejean Ant gardens interspecrecognition in parabiotic species Behav Ecol Sociobiol(1997) 87ndash93

516 F-J Richard et al C R Biologies 327 (2004) 509ndash517
d
id-
plet
he
ur-
ack
at-o-be
p-ac-p-
ire
-oure
nebio-hem
sisceso-e
ACa--the
entGIFs raquo
ds
ty
lm
a-bo5ndash
c02)
man
rrsquos
of-im
ter-star
ction9)
n-
02)
of82
iest
ar-
nd0
Dufour glands act as repellents in severalCremato-gaster species[42ndash44] and such an effect was notein the lestobioticSolenopsis fugax [45] Similarly apacifying action on the host occurs when an indivual of the parasiticFormicoxenus provancheri raisesits abdomen and extrudes the sting producing a droof volatile liquid before engaging in lsquoshampooingrsquo be-haviour withMyrmica incompleta [46]
Nevertheless why does this not work during tdaytime As theCrematogaster lsquoraidsrsquo coincide withthe dusk peak in activity ofEctatomma (1900ndash2100Fig 1A) another hypothesis can be formulated Ding this period the heavy flow ofEctatomma foragersentering and leaving the tunnel drives the guards binto the nest This permits the smallCrematogasterworkers to creep inside the tunnel without beingtacked In this situation they can modify the compsition of their cuticular hydrocarbons enough toaccepted by the nextEctatomma foragers returningto their nest and so to lure them A similar decetive mechanism based on chemical camouflagequisition was proposed to explain intraspecific cletobiosis inE ruidum [27] Later newCrematogasterindividuals can enter the tunnel or they can acqucuticular compounds directly from an interceptedEc-tatomma worker by climbing on it and licking its cuticle this corresponding to the lsquoshampooingrsquo behavidescribed by Wheeler[28] This hypothesis might bsupported by the fact thatC limata parabiotica has theability to share nests and trails with different formiciand ponerine ants in a phenomenon called lsquoparasisrsquo while these ants never share nests between t[343546]
In conclusion we noted a new kind of cleptobioin ants concerning the robbing of sugary substanand not prey robbing as previously known Temprally limited to night time and facultative it implies thpresence ofEctatomma nests onCrematogaster terri-tories
Acknowledgements
We are grateful to Dr Jacques HC Delabie (UPLaboratoacuterio de Mirmecologia Convecircnio UESCEPLAC Itabuna Bahia Brazil) for the identifiction of the ants and to AndreaDejean for proofreading the manuscript This work was supported by
French laquo Ministegravere de lrsquoEacutecologie et du Deacuteveloppemdurable raquo (SOFT program research agreementECOFOR No 98 and the laquo Tropical Ecosystemprogram No 02-E2002)
References
[1] E Curio in The Ethology ofPredation Springer-VerlagBerlin 1976 242 p
[2] HJ Brockmann CJ Barnard Kleptoparasitism in birAnim Behav 27 (1979) 487ndash514
[3] B Houmllldobler EO Wilson in The Ants Harvard UniversiPress Cambridge 1990 732 p
[4] TJ Pitcher Functions of shoaling behaviour in teleosts inJ Pitcher (Ed) The Behaviour of Teleost Fishes Croom HeLondon 1986 pp 294ndash337
[5] WJ Dominey AM Snyder Kleptoparasitism of freshwter crabs by cichlid fishes endemic to Lake Barombi MCameroon West Africa Environ Biol Fishes 22 (1988) 15160
[6] JR Garrido CG Sarasa M Fernandez-Cruz Intraspecifikleptoparasitism in the cattle egret J Field Ornithol 73 (20185ndash190
[7] CE Khatchikian M Favero AI Vassalo Kleptoparasitisby brown-hooded gulls and grey-hooded gulls on Americoystercatchers Waterbirds 25 (2002) 137ndash141
[8] MS Di Bitetti CH Janson Social foraging and the findeshare in capuchin monkeysCebus apella Anim Behav 62(2001) 47ndash56
[9] OP Honer B Wachter ML East H Hofer The responsespotted hyenas to long-term changes in prey populations functional response and interspecific kleptoparasitism J AnEcol 71 (2002) 236ndash246
[10] S Morissette JH Himmelman Subtidal food thieves inactions of four invertabrate kleptoparasites with the seaLeptasterias polaris Anim Behav 60 (2000) 531ndash543
[11] Y Heacutenaut J-P Lachaud Araigneacutees cleptoparasites seacutelede lrsquohocircte par sa toile Ann Soc Entomol Fr (NS) 35 (199148ndash153
[12] M Whitehouse I Agnarsson T Miyashita D Smith K Cagialosi T Masumoto D Li Y HeacutenautArgyrodes phylogenysociality and interspecific interactions J Arachnol 30 (20238ndash245
[13] B Crespi P Abbot The behavioral ecology and evolutionkleptoparasitism in Australian gall thrips Fla Entomol(1999) 147ndash164
[14] FE Kurczewski MG Spofford Alternative nesting strategin Ammophila urnaria (Hymenoptera Sphecidae) J NaHist 32 (1998) 99ndash106
[15] C Michener in The Social Behavior of the Bees A Compative Study The Belknap Pressof Harvard University PressCambridge 1974 404 p
[16] WM Wheeler in Ants Their Structure Development aBehaviour Columbia University Press New York 191663 p
F-J Richard et al C R Biologies 327 (2004) 509ndash517 517
)
in al 20
ing
89ndash
mic
ry
d)
n andash
5ndash
verinendash
hes
n
Ac-
ns
D
-
reyerae)
s
la-theera
c-eaery
n
ri-t 35
on
es-
ra
m
nta-
89)
de-
re
-
es
ific40
[17] A Forel Fourmis termitophages LestobioseAtta tardigradasous-genres drsquoEuponera Ann Soc Entomol Belg 45 (1901389ndash398
[18] T Abe On the food sharing among four species of antssandy grassland I Food and foraging behavior Jap J Eco(1971) 219ndash230
[19] U Maschwitz M MuhlenbergCamponotus rufoglaucus einewegelagernde Ameise Zool Anz 191 (1973) 364ndash368
[20] B Houmllldobler Food robbing inants a form of interferencecompetition Oecologia 69 (1986) 12ndash15
[21] T Yamaguchi Intraspecific competition through food robbin the harvester antMessor aciculatus (Fr Smith) and itsconsequences on colony survival Insectes Soc 42 (1995)101
[22] NA Weber Two common ponerine ants of possible econosignificanceEctatomma tuberculatum (Olivier) andE ruidumRoger Proc Entomol Soc Washington 48 (1946) 1ndash16
[23] MD Breed P Abel TJ Bleuze SE Denton Thievehome ranges and nestmate recognition inEctatomma ruidumOecologia 84 (1990) 117ndash121
[24] MD Breed LE Snyder TL Lynn JA Morhart Acquirechemical camouflage in a tropical ant Anim Behav 44 (1992519ndash523
[25] MD Breed TP McGlynn EM Stocker ANKlein Thiefworkers and variation in nestmate recognition behavior iponerine antEctatomma ruidum Insectes Soc 46 (1999) 327331
[26] I Perfecto JH Vandermeer Cleptobiosis in the antEc-tatomma ruidum in Nicaragua Insectes Soc 40 (1993) 29299
[27] JM Jeral MD Breed BE Hibbard Thief ants hareduced quantities of cuticular compounds in a poneant Ectatomma ruidum Physiol Entomol 22 (1997) 207211
[28] DE WheelerEctatomma tuberculatum foraging biology andassociation withCrematogaster (Hymenoptera Formicidae)Ann Entomol Soc Am 79 (1986) 300ndash303
[29] WL Jr Brown Contributions toward a reclassification of tFormicidae II Tribe Ectatommini (Hymenoptera) Bull MuComp Zool 118 (1958) 173ndash362
[30] A Dejean J-P Lachaud Growth-related changes in predatiobehavior in incipient colonies of the ponerine antEctatommatuberculatum (Olivier) Insectes Soc 39 (1992) 129ndash143
[31] J Valenzuela-Gonzaacutelez A Loacutepez-Meacutendez J-P Lachaudtivity patterns and foraging activity in nests ofEctatommatuberculatum (Hymenoptera Formicidae) in cacao plantatioSouthwest Entomol 20 (1995) 507ndash515
[32] RR da Hora R Feacuteneacuteron J Valenzuela ME FavilaFresneau Queen-size dimorphism in the antEctatomma
tuberculatum (Hymenoptera Formicidae Ponerinae) Sociobiology 38 (2001) 407ndash420
[33] G Ibarra-Nuacutentildeez JA Garciacutea JA Loacutepez J-P Lachaud Panalysis in the diet of some ponerine ants (HymenoptFormicidae) and web-building spiders (Araneae) in coffeplantations in Chiapas Mexico Sociobiology 37 (2001723ndash755
[34] DW Davidson Ecological studies of Neotropical ant gardenEcology 69 (1988) 1138ndash1152
[35] A Dejean B Corbara J Orivel RR Snelling JHC Debie M Belin-Depoux The importance of ant gardens inpioneer vegetal formations of French Guiana (HymenoptFormicidae) Sociobiology 35 (2000) 425ndash439
[36] LM Pascal EF Motte-Florac DB McKey Secretory strutures on the leaf rachis of Caesalpinieae and Mimosoid(Leguminosae) implication for the evolution of nectaglands Am J Bot 87 (2000) 327ndash338
[37] B Schatz J-P Lachaud G Beugnon Polyethism withihunters of the ponerine antEctatomma ruidum InsectesSoc 43 (1996) 111ndash118
[38] WM Wheeler The compound and mixed nests of Amecan ants Part I Observations on a new guest ant Am Na(1901) 431ndash448
[39] B Houmllldobler Liquid food transmission and antennatisignals in ponerine ants Isr J Entomol 19 (1985) 89ndash99
[40] J Liebig J Heinze B Houmllldobler Trophallaxis and aggrsion in the ponerine antPonera coartata implications forthe evolution of liquid food exchange in the HymenopteEthology 103 (1997) 707ndash722
[41] C Bevan A Birch H Caswell Ant insect repellent froblack cocktail ants J Chem Soc 961 (1961) 488
[42] AB Attygalle B Siegel O Vostrowsky HJ BestmanU Maschwitz Chemical composition and function of mepleural gland secretion of the antCrematogaster deformisSmith (Hymenoptera Myrmicinae) J Chem Ecol 15 (19317ndash328
[43] D Daloze M Kaisin C Detrain JM Pasteels Chemicalfense in the three European species ofCrematogaster antsExperientia 47 (1991) 1082ndash1089
[44] B Houmllldobler Chemische Strategie beim Nahrungserwerb deDiebameise (Solenopsis fugax Latr) und der Pharaoameis(Monomorium pharaonis L) Oecologia 11 (1973) 371ndash380
[45] A Lenoir C Errard A Francoeur R Loiselle Relations entre la fourmi parasiteFormicoxenus provancheri et son hocircteMyrmica incompleta Donneacutees biologiques et eacutethologiqu(Hym Formicidae) Insectes Soc 39 (1992) 81ndash97
[46] J Orivel C Errard A Dejean Ant gardens interspecrecognition in parabiotic species Behav Ecol Sociobiol(1997) 87ndash93

F-J Richard et al C R Biologies 327 (2004) 509ndash517 517
)
in al 20
ing
89ndash
mic
ry
d)
n andash
5ndash
verinendash
hes
n
Ac-
ns
D
-
reyerae)
s
la-theera
c-eaery
n
ri-t 35
on
es-
ra
m
nta-
89)
de-
re
-
es
ific40
[17] A Forel Fourmis termitophages LestobioseAtta tardigradasous-genres drsquoEuponera Ann Soc Entomol Belg 45 (1901389ndash398
[18] T Abe On the food sharing among four species of antssandy grassland I Food and foraging behavior Jap J Eco(1971) 219ndash230
[19] U Maschwitz M MuhlenbergCamponotus rufoglaucus einewegelagernde Ameise Zool Anz 191 (1973) 364ndash368
[20] B Houmllldobler Food robbing inants a form of interferencecompetition Oecologia 69 (1986) 12ndash15
[21] T Yamaguchi Intraspecific competition through food robbin the harvester antMessor aciculatus (Fr Smith) and itsconsequences on colony survival Insectes Soc 42 (1995)101
[22] NA Weber Two common ponerine ants of possible econosignificanceEctatomma tuberculatum (Olivier) andE ruidumRoger Proc Entomol Soc Washington 48 (1946) 1ndash16
[23] MD Breed P Abel TJ Bleuze SE Denton Thievehome ranges and nestmate recognition inEctatomma ruidumOecologia 84 (1990) 117ndash121
[24] MD Breed LE Snyder TL Lynn JA Morhart Acquirechemical camouflage in a tropical ant Anim Behav 44 (1992519ndash523
[25] MD Breed TP McGlynn EM Stocker ANKlein Thiefworkers and variation in nestmate recognition behavior iponerine antEctatomma ruidum Insectes Soc 46 (1999) 327331
[26] I Perfecto JH Vandermeer Cleptobiosis in the antEc-tatomma ruidum in Nicaragua Insectes Soc 40 (1993) 29299
[27] JM Jeral MD Breed BE Hibbard Thief ants hareduced quantities of cuticular compounds in a poneant Ectatomma ruidum Physiol Entomol 22 (1997) 207211
[28] DE WheelerEctatomma tuberculatum foraging biology andassociation withCrematogaster (Hymenoptera Formicidae)Ann Entomol Soc Am 79 (1986) 300ndash303
[29] WL Jr Brown Contributions toward a reclassification of tFormicidae II Tribe Ectatommini (Hymenoptera) Bull MuComp Zool 118 (1958) 173ndash362
[30] A Dejean J-P Lachaud Growth-related changes in predatiobehavior in incipient colonies of the ponerine antEctatommatuberculatum (Olivier) Insectes Soc 39 (1992) 129ndash143
[31] J Valenzuela-Gonzaacutelez A Loacutepez-Meacutendez J-P Lachaudtivity patterns and foraging activity in nests ofEctatommatuberculatum (Hymenoptera Formicidae) in cacao plantatioSouthwest Entomol 20 (1995) 507ndash515
[32] RR da Hora R Feacuteneacuteron J Valenzuela ME FavilaFresneau Queen-size dimorphism in the antEctatomma
tuberculatum (Hymenoptera Formicidae Ponerinae) Sociobiology 38 (2001) 407ndash420
[33] G Ibarra-Nuacutentildeez JA Garciacutea JA Loacutepez J-P Lachaud Panalysis in the diet of some ponerine ants (HymenoptFormicidae) and web-building spiders (Araneae) in coffeplantations in Chiapas Mexico Sociobiology 37 (2001723ndash755
[34] DW Davidson Ecological studies of Neotropical ant gardenEcology 69 (1988) 1138ndash1152
[35] A Dejean B Corbara J Orivel RR Snelling JHC Debie M Belin-Depoux The importance of ant gardens inpioneer vegetal formations of French Guiana (HymenoptFormicidae) Sociobiology 35 (2000) 425ndash439
[36] LM Pascal EF Motte-Florac DB McKey Secretory strutures on the leaf rachis of Caesalpinieae and Mimosoid(Leguminosae) implication for the evolution of nectaglands Am J Bot 87 (2000) 327ndash338
[37] B Schatz J-P Lachaud G Beugnon Polyethism withihunters of the ponerine antEctatomma ruidum InsectesSoc 43 (1996) 111ndash118
[38] WM Wheeler The compound and mixed nests of Amecan ants Part I Observations on a new guest ant Am Na(1901) 431ndash448
[39] B Houmllldobler Liquid food transmission and antennatisignals in ponerine ants Isr J Entomol 19 (1985) 89ndash99
[40] J Liebig J Heinze B Houmllldobler Trophallaxis and aggrsion in the ponerine antPonera coartata implications forthe evolution of liquid food exchange in the HymenopteEthology 103 (1997) 707ndash722
[41] C Bevan A Birch H Caswell Ant insect repellent froblack cocktail ants J Chem Soc 961 (1961) 488
[42] AB Attygalle B Siegel O Vostrowsky HJ BestmanU Maschwitz Chemical composition and function of mepleural gland secretion of the antCrematogaster deformisSmith (Hymenoptera Myrmicinae) J Chem Ecol 15 (19317ndash328
[43] D Daloze M Kaisin C Detrain JM Pasteels Chemicalfense in the three European species ofCrematogaster antsExperientia 47 (1991) 1082ndash1089
[44] B Houmllldobler Chemische Strategie beim Nahrungserwerb deDiebameise (Solenopsis fugax Latr) und der Pharaoameis(Monomorium pharaonis L) Oecologia 11 (1973) 371ndash380
[45] A Lenoir C Errard A Francoeur R Loiselle Relations entre la fourmi parasiteFormicoxenus provancheri et son hocircteMyrmica incompleta Donneacutees biologiques et eacutethologiqu(Hym Formicidae) Insectes Soc 39 (1992) 81ndash97
[46] J Orivel C Errard A Dejean Ant gardens interspecrecognition in parabiotic species Behav Ecol Sociobiol(1997) 87ndash93
Copyright © 2022 FDOKUMEN




















