
Ethnologia Europaea Revisited: Launching Future Ethnologies. An Introduction.

PLEASE SCROLL DOWN FOR ARTICLE
This article was downloaded by: [Università della Calabria]On: 22 January 2010Access details: Access Details: [subscription number 909050702]Publisher Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
Italian Journal of ZoologyPublication details, including instructions for authors and subscription information:http://www.informaworld.com/smpp/title~content=t741771159
Hunting Ants in Mediterranean clay soils: Life history of Siagona europaea(Coleoptera, Carabidae)Thomas Bauer a; Federica Talarico b; Antonio Mazzei b; Anita Giglio b; Tullia Zetto-Brandmayr b; PietroBrandmayr b; Oliver Betz c
a Zoological Institute, Christian Albrechts University, Kiel, Germany b Dipartimento di Ecologia,Università di Calabria, Arcavacata di Rende, Italy c Zoological Institute, Eberhard Karls University,Tübingen, Germany
To cite this Article Bauer, Thomas, Talarico, Federica, Mazzei, Antonio, Giglio, Anita, Zetto-Brandmayr, Tullia,Brandmayr, Pietro and Betz, Oliver(2005) 'Hunting Ants in Mediterranean clay soils: Life history of Siagona europaea(Coleoptera, Carabidae)', Italian Journal of Zoology, 72: 1, 33 — 42To link to this Article: DOI: 10.1080/11250000509356650URL: http://dx.doi.org/10.1080/11250000509356650
Full terms and conditions of use: http://www.informaworld.com/terms-and-conditions-of-access.pdf
This article may be used for research, teaching and private study purposes. Any substantial orsystematic reproduction, re-distribution, re-selling, loan or sub-licensing, systematic supply ordistribution in any form to anyone is expressly forbidden.
The publisher does not give any warranty express or implied or make any representation that the contentswill be complete or accurate or up to date. The accuracy of any instructions, formulae and drug dosesshould be independently verified with primary sources. The publisher shall not be liable for any loss,actions, claims, proceedings, demand or costs or damages whatsoever or howsoever caused arising directlyor indirectly in connection with or arising out of the use of this material.

Ital. J. Zool., 72: 33-42 (2005)
Hunting Ants in Mediterranean claysoils: life history of Siagona europaea(Coleoptera, Carabidae)
THOMAS BAUERZoological Institute, Christian Albrechts University,24098 Kiel (Germany)
FEDERICA TALARICOANTONIO MAZZEIANITA GIGLIOTULLIA ZETTO-BRANDMAYRPIETRO BRANDMAYRDipartimento di Ecologia, Università di Calabria,I-87030 Arcavacata di Rende (Italy)
OLIVER BETZZoological Institute, Eberhard Karls University,72076 Tübingen (Germany)
ABSTRACT
In southern Italy, Siagona europaea is common on clay soilsand occurs up to the 38th parallel. Activity recordings have re-vealed that the beetles are strictly nocturnal. They are resistant todesiccation and prefer temperatures above 30 °C. Their flat bodyindicates adaptation to life in clay soils, which, in southern Italy,are deeply fissured from April to September. The behaviour of thebeetles was observed in a transparent system of cavities consist-ing of plastic boxes connected by tubes. The beetles feed exclu-sively on ants that are caught by surprise attack outside theirnests. Ants perceive the beetles as being hostile via chemicalcues. Single ants then normally retreat, whereas groups of ants at-tack immediately. The beetles behave unaggressively againstcospecifics and rest in groups of various numbers in flat soilcrevices. Copulation takes place from the end of April onwards,whereas egg laying occurs from the end of May onwards. The fe-males produce only a few eggs, which are unusually large, andshift them singly in narrow soil cracks. The larvae develop duringthe summer deep in the soil; they are blind and can be character-ized as cave-life forms by their extremely long cerci and antennaethat have a whip-like unsclerotized sticky distal part.
KEY WORDS: Carabidae - Siagona - Ants.
ACKNOWLEDGMENTS
Prof. Dr. Thorsten Assmann, University of Lüneburg, Germany,provided us with living specimens of S. jenissoni and S. dejeanifrom Morocco and Southern Spain. Dr. Donato A. Grasso, Univer-sity of Parma, determined some ant species for us. ChristianaAnagnostou made the drawings; Dr. Heike Betz helped us withthe diagrams; Dr. Teresa Jones improved the English of this pa-per. The visit of Thomas Bauer to the University of Calabria waspartly financed by the Socrates teacher exchange program. Twoanonymous referees helped improve this paper.
(Received 25 Agust 2004 - Accepted 20 October 2004)
INTRODUCTION
Siagona europaea Dejean 1826 is one of ca. 50 de-scribed species within the genus Siagona, belonging tothe Siagonini, a tribe of the Scaritinae. Siagonini seemto be an old distinct group within the Carabidae (Erwin,1985; Galian et al, 1992); however, to date, almostnothing is known about the ecological demands andlife style of the species of this genus, which has a widegeographic distribution including India (Andrewes,1929), Arabia, Africa and the Mediterranean region.Three species are present in southern Europe, S. jenis-soni Dejean (15-17.5 mm), S. dejeani Rambur (20-25mm) and S. europaea Dejean (12-15 mm). Like all Sia-gonini, S. europaea has a strikingly flat body with astalk-like constriction between the pro- and mesothoraxand strong mandibles with large retinacula (Fig.l). Theflatness of the body and thoracic constriction are obvi-ous adaptations to life in narrow soil crevices. The shortbut strong mandibles are well-suited for grasping andchewing arthropod prey with strong cuticles.
In southern Italy, S. europaea occurs in open land onclay soils (Brandmayr & Pizzolotto, 1990) up to a heightof about 230 m a.s.l. and in Calabria approx. to the 38th
parallel. In early spring, when soil moisture is high, thebeetles are found under stones. From mid-April on-wards, when the soil dries out and becomes deeply fis-sured, they retreat into deeper crevices, especially dur-ing the hot dry hours of the day. Pitfall catches have re-vealed that the beetles are active on the soil surfacefrom the end of February to the beginning of Novemberand dissections have shown that gonad maturation isreached in early summer and lasts until July (Zetto--Brandmayr & Pizzolotto, 1994). Thus, larval develop-ment should take place during the summer. The larvae,however, have never been found. The intestines of bee-tles caught in pitfalls exclusively contain the remains ofants, indicating that they are myrmecophagous. This hasbeen confirmed by feeding experiments: S. europaeafeed exclusively on ants, if ants are available, but do notclearly differentiate between different ant species (Zetto--Brandmayr et al, 1998). Nevertheless, the closeness ofthe relationship between these beetles and ants remainsunknown. Do they invade ant nests? Does the larval de-velopment of these beetles take place in ant nests?
Because of its hidden life style under stones and in thesoil, it is impossible to observe S. europaea (and otherSiagona spp.) in the field and, thus, most details of itslife history are unknown. We have therefore tried to rearS. europaea in the laboratory in order to observe its be-haviour under semi-natural conditions and to investigatethe habitat demands of the species experimentally.
MATERIALS AND METHODS
Approximately 300 living specimens of S. europaea werecaught from March to June 2003 in Calabria, southern Italy, andkept in laboratory. From February to April, they were collected
Downloaded By: [Università della Calabria] At: 09:11 22 January 2010

34 T. BAUER, F. TAIARICO, A. MAZZEI, A. GIGLIO, T. ZETTO-BRANDMAYR, P. BRANDMAYR, O. BETZ
D
Fig. 1 - Siagona europaea: A, dorsal view; B, ventral view; C, lat-eral view; D, rear view of a male; E, rear view of a female (ven-tral view).
by hand from under stones in fields near Cerenzia (50 km east ofCosenza). From mid-April, the beetles moved too quickly to becollected by hand because of the rising temperature and the in-creasing occurrence of soil fissures. Thus, from May onwards,they were caught in pitfall traps, placed near ant colonies, in ameadow near Squillace (15 km southwest of Catanzaro, 5 kmfrom the Ionian Sea). During collection excursions, also the tem-perature and relative humidity (rH) were measured 1 m above thesoil, on the soil surface, under stones and, from April, at variousdepths in the soil fissures in the habitat of S. europaea. In May2003, we received an additional five males and two females of 5.jenissoni Rambur and two females of S. dejeani Dejean fromsouthern Spain, all of which were included in the observations.
The beetles were kept partly singly and partly in groups of upto 30 specimens in the laboratory on moist plaster of Paris inplastic boxes of various sizes (see below). Because of its adapta-tion to life in crevices, Siagona spp. most of the time tries to re-main in contact with soil structures with its back. If this is impos-sible, the beetles are restless and obviously stressed and do notbehave "normally". We solved this problem by offering, on theplaster soil, pieces of glass that were supported by small plastictubes on one side. Under the obliquely angled glass panes, thebeetles could choose spaces of the preferred width but remainedvisible. For observations of egg-laying behaviour, flat Petri dishesfilled with moist clay we placed in the boxes. The clay surfacewas also partly covered with pieces of glass, so that the beetlesfound narrow crevices for egg laying under the glass.
Observation system
In order to observe beetles and ants under almost natural con-ditions with respect to climate and space, we constructed a vari-able artificial "soil gap" system, consisting of plastic boxes thatwere connected by flexible transparent plastic tubes of variouslengths. The size of the boxes was 20 x 20 x 8 cm or 20 x 10 x 8cm. The bottom of each box was covered with moist plaster andequipped with laterally supported, angled glass panes (seeabove). The walls of the boxes were drilled close to the plasterbottom so that the tubes could be fitted and readily accessed bythe animals. The tubes had an inner diameter of ca. 8 mm, so thatbeetles and ants could pass between the boxes. By compressingthe tubes, the connections could also easily be shut or opened.This highly flexible system allowed the observation and manipu-lation of a wide range of confrontation between the animals.
Ant colonies of the following species were excavated in earlySpring in the habitat of S. europaea, and established in the boxsystem in the laboratory: Camponotus sp. (large species, onecolony), Messor sp. (large species, one colony), M. capitatus (twocolonies), Aphaenogaster ionia (three colonies), Tetramoriumsemilaeve (one colony). The Messor colonies were fed with seeds,
the two other species with meat and honey. Each colony hadthree or more chambers of different sizes that were connected bytubes. In one of them, food and water were offered; the otherswere used by the ants for brood and litter. Each chamber couldalso be connected by tubes with the boxes of single or groups ofbeetles. The rH within the system approached 100%; the tempera-ture corresponded to that in the laboratory and was between 18°C (mean) (beginning of March) and 26 °C (end of June) with aminimum in the night and a maximum during the daytime. Thewindows of the laboratory were covered with black paper, whichwas not however completely tight, so that the animals were ableto recognize dawn and dusk.
Observations
For observations and video recordings, the vessels were illumi-nated with a microscope lamp in a way that the animals could seethe light source but not the observer above it. The light did notseem to affect their behaviour in the closed system. On the con-trary, radiation increased their readiness to act, especially on cold-er days. Ants and beetles were, however, sensitive to vibrations,which caused them to remain motionless. We thus had to avoidtouching the table on which the box systems had been placedand had to remain motionless ourselves during the observations.For recording of behaviour, a video camera (Sony DCR-TRV345EDigital Handycam Hi 8) was used. Observations were initiallymade throughout the day. After some time, it became obvious thatthe best hours for observation were those after dusk, because ofthe natural onset of activity of these nocturnal beetles after sunset(see "Daily activity pattern"). A total of about 200 h from March toJune 2003 were spent in observing and filming inter- and intraspe-cific behaviour. For the observation and filming of the interactionsbetween beetles and ants, the plastic tubes between their boxeswere carefully opened to avoid vibrations. A total of about 100 in-teractions were filmed and many more were observed.
Daily activity pattern
The daily activity pattern was recorded in a climatic chamber ata constant temperature of 25 °C and a light regime of L/D = 18/6.The brightness was set at 10,000 Ix, with an artificial dawn anddusk, i.e. an increase or decrease of brightness in steps of 2,000lx within 1 h by automatically and successively switching on andoff single neon tubes of the illumination system. The beetles wereplaced in vessels of 20 x 20 x 8 cm with a moist plaster of Parisbottom. The middle of the bottom was crossed by a red laserbeam close to the plaster surface. On both sides of the beam,pieces of bark were placed as shelters for the beetles. In phasesof activity, the beetles left the space under the bark and ranaround. Each time a beetle crossed through the beam, a signalwas produced; these signals were counted automatically, summa-rized every 10 minutes and sampled on a PC. According to ourobservations and the signal recordings, the behaviour of the bee-tles was not influenced by the weak red light of the laser: theyneither stopped in the beam nor avoided crossing it when theywere active. The activity of 10 males and 10 females was record-ed synchronously but in two different vessels.
Humidity demands
The preferred humidity was investigated in closed circular hu-midity gradients (diameter: 30 cm; height: 11.5 cm): They consist-ed of 10 symmetrical sectors on the bottom and correspondinglyin the lid (Fig. 2). Two of the bottom sectors on opposite sideswere filled with cotton wool containing distilled water and silicagel. The beetles could move between the sectors on a gauze areaabove the bottom. A piece of supported glass (see above) ineach sector provided tactile shelters for the beetles. Humidity wasmeasured through holes in the lid using testo hygro sticks 605--Hl. Once the apparatus was closed, a humidity gradient between25 and > 81% rH was established within 15 min. Experimentswere carried out in a dark room at 28 °C. The beetles were intro-duced into the closed apparatus and, after 20 minutes, their posi-
Downloaded By: [Università della Calabria] At: 09:11 22 January 2010

LIFE HISTORY OF SIAGONA EUROPAEA 35
Fig. 2 - Humidity gradient.
tion in the sectors was recorded every 5 min by using a red lightlamp; 10 males and 10 females were used for this experiment inseparated gradients. Each experiment lasted ca. 2 h and was re-peated on two following days. The position within the gradientwas recorded for a total of 84 times for every beetle.
Temperature demands
The preferred temperature was investigated in circular tempera-ture gradients (Fig. 3) (Circular climate gradients have the advan-tage that the animals that are tested do not get stuck if they passthe cold or hot maximum of the gradient). They consisted of aring of aluminium, 2 cm thick, 6.5 cm broad and with an outer di-ameter of 60 cm. The ring was bordered by aluminium sheets upto a height of 5 cm. The upper surface of the ring had a layer ofmoist plaster of Paris. The lower surface and the outer surfaces ofthe side sheets were insulated with a neoprene layer. The ringcould be heated with an electric iron on one side and cooledwith a laboratory cooler on the opposite side. The plaster surfaceon which the animals were allowed to move was divided intoeight sectors of equal ground space on each side of the ring. Inevery sector, a glass pane provided tactile shelter for the beetles.The temperature was measured with thermometers positioned onthe floor exactly at the borders between the sectors. The experi-ments were carried out in a dark room with gradients of betweenca. 7 and 44 °C. Twelve males and 12 females were used in sepa-rate gradients; 30 min after placing them in the apparatus, theirposition was recorded every 5 min. One experiment lasted 1-2 hand was repeated on the two following days. The position ofevery beetle was recorded 51 times.
cooling unit
area covered withplaster of Paris
Fig. 3 - Temperature gradient.
Statistics
The distribution in the temperature and in the moisture gradi-ents was tested with the Kolmogorov-Smirnov test for Goodnessof Fit, and the humidity preference of the beetles during the 1st
and 2nd hour of the experiment was tested with a 3 x 3 contin-gency table. A chi-square test was used to test for independenceof humidity preference (i.e. first versus second hour of experi-ment). Differences between observed and expected values wereconsidered to be significant when their standardized residuals hada value of > 2 or < -2 (Sokal & Rohlf, 1995).
The data were evaluated with the Statistics package SPSS 6.1for Windows (Chicago).
RESULTS
Predatory behaviour and interactions with ants
The behaviour of the beetles depended on their nutri-tional state. Satisfied beetles were inactive and stayed intheir shelters (under the glass panes). When they werehungry, they moved around, palpating with their anten-nae. They apparently detected the presence of ants bychemical cues. The waste of ant nests was always care-fully investigated by the beetles and hungry beetles alsofed on dead ants from the refuse (11 observations).When a hungry beetle touched an ant, it immediatelyattacked the ant with a short but fast lunge and tried tograsp it. When the first attack failed, it continued withrapid lunges until it attained a safe grip. Large ants(more than half the size of the beetles-, e.g. workers ofMessor sp. or Camponotus sp.) fought back vigorouslyand often caused a retreat of the beetle. Smaller antswere always drawn into a shelter and immediately eat-en. Eating an ant took several minutes because all bodyparts were "carefully" chewed and ingested. Parts thatfell to the ground during the crushing of an ant (e.g.legs, antennae) were taken up again and also chewedand eaten. The faeces of S. europaea had a cylindricalshape, measured about 2 x 0.5 mm and consisted main-ly of cuticle pieces of ants.
The behaviour of all ant species confronted with S.europaea demonstrated clearly in every case that theyrecognized the beetles as hostile and dangerous bychemical cues. Single ant scouts that entered the box ofa beetle first moved "carefully" around, often hesitatingand palpating with the antennae. They detected a mo-tionless beetle in its shelter always without touching itand at a distance of 1.5-2.5 cm, and then immediatelystopped. Depending on its motivation, the ant thenquickly retreated or attacked the beetle by repeatedlypushing it with its mandibles. The latter behaviour alsoseemed to be an alarm signal (probably supported byalarm pheromones) and attracted and caused attacks ofother ants if they were close by (cf. Hölldobler & Wil-son, 1990). Beetles that were attacked by single ants re-taliated, or, if hungry, tried to catch the ant. When abeetle was attacked by a number of ants, it often re-treated under a glass pane, taking advantage of its flatbody. Here, it usually remained motionless and only re-taliated when ants tried to draw it out by grasping its
Downloaded By: [Università della Calabria] At: 09:11 22 January 2010

36 T. BAUER, F. TALARICO, A. MAZZEI, A. GIGUO, T. ZETTO-BRANDMAYR, P. BRANDMAYR, O. BETZ
legs or antennae. When a beetle was continuously mo-lested, for example, in a shelter close to a nest en-trance, it fled struggling and running rapidly until itfound another narrow space.
When a number of ants of a large colony entered abox of beetles, they immediately attacked them and al-ways tried to bite and to kill them. However, they neversucceeded when the beetles found shelter in flatcrevices. Moreover, large ants (Camponotus, Messor,Aphaenogaster) rarely succeeded in holding a beetle,because of its smooth surface and its strong rapid move-ments. Small ants (Tetramorium sp.) were also able toattack a Siagona in its shelter. They seemed, however, tobe unable to hurt it severely, but caused the beetle toflee by biting and adhering to its tarsi and antennae.
When we opened a tube between an ant colony anda beetle box, the ants immediately occupied the tubeand, after contact with the beetles, they guarded andnarrowed the entrance by filling it with particles. Thebeetles, on the other hand, usually avoided confronta-tions with more than one ant and immediately ran awaywhen they were attacked at colony entrances. In thesomewhat artificial situation of our box and tube sys-tems, we observed in seven cases that beetles, irritatedby vibrations caused by the observer, tried to flee and,in panic, entered ant nests. In every case, they were im-mediately severely attacked. They then struggled andran around at high speed. When they found a narrowspace within the nest, they rested for a while. Other-wise, they continued running and struggling until theyfound a way out.
Dying or dead beetles were treated as prey by theants (two observations with Aphaenogaster ionid). Theants drew the dying beetle into the nest, first licked itssurface for a while and then placed larvae on its sur-face. These observations were made with S. europaeaand partly also with 5. jenissoni. They are likely alsotrue for S. dejeani, although this species is much largerand is thus able to overpower larger ants such as Cam-ponotus sp. and Messor sp. S. dejeani is not as fast as S.europaea but its behaviour seems similar.
Intraspecific behaviour
When the beetles moved around, they often touchedeach other with their antennae and sometimes pushedeach other out of the way but very rarely attacked.They apparently recognized each other as cospecificsby means of chemical cues. This was confirmed by theobservation that dead beetles were immediately eatenby their cospecifics. They probably then loose theirspecies-specific chemical surface bouquet and are treat-ed as prey.
Resting in aggregations within flat crevices in varyingnumbers seems to be part of the natural behaviour ofSiagona beetles. During our collections in early spring,S. europaea was usually found in aggregations of 2-7specimens in the same cavity under stones and this ag-
gregation behaviour was maintained during the breed-ing season in the laboratory: the beetles preferred torest in groups close to each other under the glasspanes. Freshly caught beetles, when released into theboxes, ran around in panic until they met an aggrega-tion of cospecifics under a glass pane, at which pointthey immediately became calm. When we fed such anaggregation with ant brood, two to three beetles oftenfed on the same prey item if they were not especiallyhungry. When we put specimens of the three investigat-ed Siagona species together, they treated each otherpeacefully in aggregations under glass panes. However,in large boxes with more than one shelter, single bee-tles often left such groups, so that the numbers of bee-tles under the glass panes changed regularly.
Aggressive behaviour was only observed in S. eu-ropaea and occurred rarely (five observations). The bee-tles attacked cospecifics by intensively pushing and bit-ing them with the mandibles, so that the attacked speci-men finally retreated. If, however, the combatant wasseized by its legs or antennae, it turned back and thenboth stood in opposition like rams, beating each otherstrongly with their laterally expanded antennae. This last-ed for a few seconds and then both turned away (Fig. 4).
The first copulations were observed in S. jenissoni on28th April and in S. europaea on 9th May, and from thenon continuously. Males and females copulated repeat-edly during the season. Females of S. jenissoni, whenbrought into a group of males, copulated subsequentlywith up to five males. Males of S. jenissoni only occa-sionally competed with each other and tried to pulleach other off the female, but always unagressively.Copulations took between 1 and 2.5 min in bothspecies. Several times, females eliminated a white mass(spermatophore?) from their ovipositor after copulation,turned around and ate it.
Fig. 4 - Ritualized fighting: two beetles that feed on an ant pupabeat each other with the antennae (5. europaea) (drawn after afilm recording).
Downloaded By: [Università della Calabria] At: 09:11 22 January 2010
LIFE HISTORY OF SIAGONA EUROPAEA 37
In both species, the males sometimes showed a strik-ing behaviour. Close to females, they repeatedly turnedtheir body to the sides and to the back rubbing it withslow movements against the soil (Fig. 5). This behaviourwas also observed when beetles under their glass panewere heated by the light of a microscope lamp. We ini-tially interpreted this as a kind of courtship behaviour;however, copulation took place without this behaviourand the behaviour was also performed without copula-tion. Thus, it may be involved with chemically markingthe aggregation site or themselves (cf. Discussion).
Fig. 5 - Rubbing behaviour: the beetle on the left is turning itsback to the ground (S.jenissonî) (drawn after a film recording).
Eggs and larvae
In S. europaea, the first egg was found on 28th May.The eggs measured 3-15-3.4 x 0.85-0.9 mm and wereunusually large for a flat beetle with a body length ofabout 12 mm. Only a few eggs were laid per female.For egg laying, the females extended the tube-like ab-dominal end to a length of ca. 2.5 mm and inserted itinto the narrow space between the clay soil and a glasspane (cf. Materials and Methods). They have movablestyli but never used them for digging as do other Cara-bidae (Bauer, 1974; Adis et al, 1997)
We isolated the eggs, but they did not develop, prob-ably because the temperatures in the laboratory weretoo high and unstable in June (cf. "Discussion").
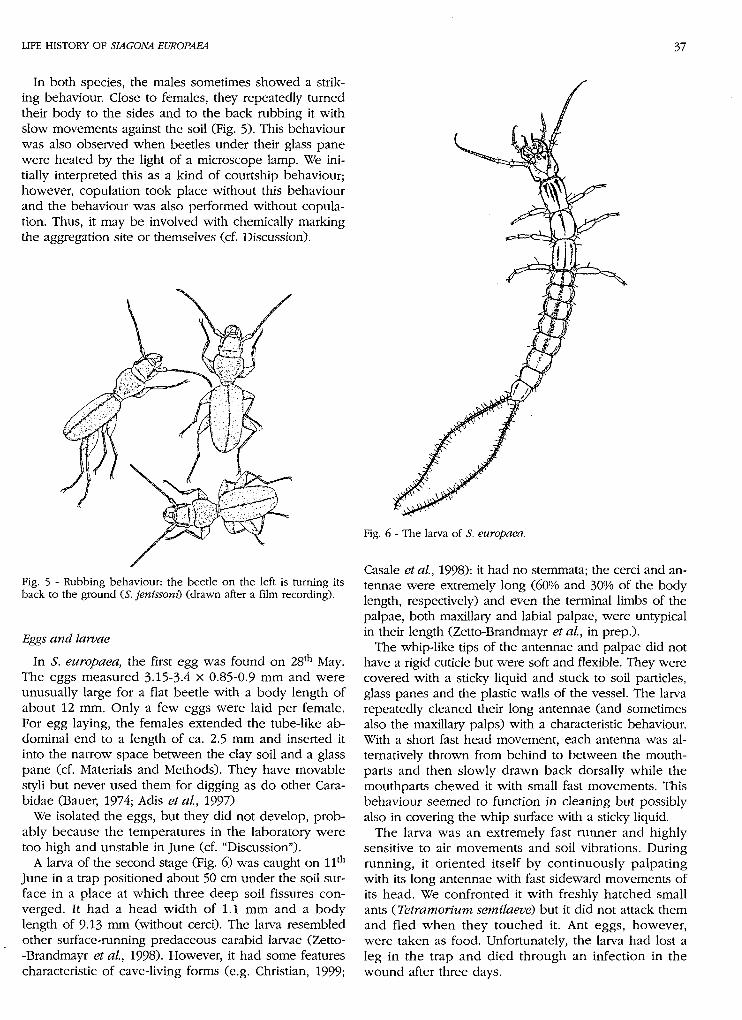
A larva of the second stage (Fig. 6) was caught on 11th
June in a trap positioned about 50 cm under the soil sur-face in a place at which three deep soil fissures con-verged. It had a head width of 1.1 mm and a bodylength of 9-13 mm (without cerci). The larva resembledother surface-running predaceous carabid larvae (Zetto--Brandmayr et al, 1998). However, it had some featurescharacteristic of cave-living forms (e.g. Christian, 1999;
Fig. 6 - The larva of S. europaea.
Cásale et al, 1998): it had no stemmata; the cerci and an-tennae were extremely long (60% and 30% of the bodylength, respectively) and even the terminal limbs of thepalpae, both maxillary and labial palpae, were untypicalin their length (Zetto-Brandmayr et al, in prep.).
The whip-like tips of the antennae and palpae did nothave a rigid cuticle but were soft and flexible. They werecovered with a sticky liquid and stuck to soil particles,glass panes and the plastic walls of the vessel. The larvarepeatedly cleaned their long antennae (and sometimesalso the maxillary palps) with a characteristic behaviour.With a short fast head movement, each antenna was al-ternatively thrown from behind to between the mouth-parts and then slowly drawn back dorsally while themouthparts chewed it with small fast movements. Thisbehaviour seemed to function in cleaning but possiblyalso in covering the whip surface with a sticky liquid.
The larva was an extremely fast runner and highlysensitive to air movements and soil vibrations. Duringrunning, it oriented itself by continuously palpatingwith its long antennae with fast sideward movements ofits head. We confronted it with freshly hatched smallants (Tetramorium semilaevè) but it did not attack themand fled when they touched it. Ant eggs, however,were taken as food. Unfortunately, the larva had lost aleg in the trap and died through an infection in thewound after three days.
Downloaded By: [Università della Calabria] At: 09:11 22 January 2010

38 T. BAUER, F. TALARICO, A. MAZZEI, A. GIGLIO, T. ZETTO-BRANDMAYR, P. BRANDMAYR, O. BETZ
Daily activity pattern and climatic demands
Males and females of S. europaea were observed to bestrictly nocturnal (Fig. 7), indicating that, in their naturalsurroundings, they leave the soil only during the night.The few peaks of activity during the daytime may havebeen attributable to the experimental arrangement: theshelter under the bark pieces did not offer total darkness.
The preferred temperature was between 30 °C and 35°C in both sexes (Fig. 8), a range that is extremely highwhen compared with nocturnal species from central Eu-rope (cf. Discussion).
The distribution of the beetles in the humidity gradientdid not show a clear peak but revealed that they werehighly resistant to desiccation (Fig. 9)- They were activeduring this experiment and often changed their position.
The Kolmogorov-Smirnov test for goodness of fit re-vealed a significant deviation of the sample distributionfrom a uniform distribution with respect to temperaturein = 1340, P < 0.001) and humidity (n = 1448, P < 0.001).
We previously found that species that are sensitive todesiccation tend to avoid the dry sectors after being inthe experimental chamber for some time because they
T 12000
-- 10000
Time [h]
B1200 T
1000-
O)
12
T 12000
-• 10000
- 8000
- • 6000 c
- 4 0 0 0 -j?
- 2000
Fig. 7 - Siagona europaea: activity pattern of males (A) and females (B) over three days
Downloaded By: [Università della Calabria] At: 09:11 22 January 2010

LIFE HISTORY OF SIAGONA EUROPAEA 39
O
n
tri
(A
ñ
100 -80-60-40 -20-
0 -43.5 36.7 34.5 29.5 24.9 20.5 16.5 12.7 7.3
Temperature [°C]
Fig. 8 - Siagona europaea: temperature preference.
soo
100-
80 -
60 -
40 -
«S 2 0 -
Q o1 2 3 4
SectorFig. 9 - Siagona europaea: humidity preference.
dry out during the experiment and are then forced toretreat into the moistest part of the gradient (cf. Bauer,1982). Thus, the comparison of the distribution of thefirst and the second hours in an experiment to deter-mine humidity preferences provides a clear indicationof the species' sensitivity to desiccation. A shift in thedistribution to the moist part was also observed in S.europaea (Table I, Fig. 10); this was statistically signifi-cant (cf. Table I) but weak when compared with otherground beetles. Even during the second hour of the ex-periment, many beetles were still found in the driestsector. The experiments provided clear evidence that S.europaea, although strictly nocturnal, is adapted to ahot, dry environment.
DISCUSSION
Adaptations to habitat
Nearly all phases of the life history of S. europaeatake place in narrow spaces and in darkness. This isprobably also true of the other Siagona species (An-drewes, 1929; Antoine, 1955; Moore, 1972; A. Serrano,personal communication) and may be one reason forthere being almost no information concerning the lifestyle of this genus. Observations of behaviour are onlypossible by keeping the beetles in an artificial "soil sys-tem" as described above, viz. a completely transparentsystem containing flat spaces into which the beetles can
TABLE I - Humidity preference ofS. europaea during the first andsecond hour of the experiments; 5x2 contingency table to test forindependence between levels of humidity and time of exposure.Numbers from top to bottom in each category: observed value, ex-pected value, standardized residual.
Level of humidity 1st hour 2nd hour
204260.5-3.58797.5-1.1
4128.02.5
6159.50.2
331278.5
3.1
317260.5
3.510897.5
1.1
1528.0-2.5
5859.5-0.2
226278.5-3.1
'100-, D First hour [%]• Second hour [%]
2 3 4 5
SectorFig. 10 - Siagona europaea: humidity preference in the first andin the second hour of the experiments.
retreat and rest. Most of the time, the beetles in our ex-periments inserted their flat bodies under the providedglass panes until they made contact with the glass viatheir backs. When they were without this shelter fromabove, they were often in panic and ran around. Flatcrevices were used when S. europaea lay in wait forants and provided a safe shelter when the beetles wereattacked by groups of ants.
Recordings of activity have shown clearly that S. eu-ropaea beetles are nocturnal and avoid light. This isconfirmed by the structure of their compound eyes: S.europaea has small eyes with only about 510 ommatidiain one eye, i.e. 43 ommatidia/mm body length, a valuecharacterizing a nocturnal species (cf. Bauer & Kredler,1993). The pattern of activity explains why we havenever found the beetles on the soil surface in their nat-
Downloaded By: [Università della Calabria] At: 09:11 22 January 2010

40 T. BAUER, F. TALARICO, A. MAZZEI, A. GIGLIO, T. ZETTO-BRANDMAYR, P. BRANDMAYR, O. BETZ
ural environment and could only catch them in traps or,in the spring, under large stones. On the other hand,the presence of narrow spaces seems to be more crucialfor them to behave naturally than does that of totaldarkness. In nature, both components are always com-bined. In our transparent "soil system", the beetles stopmoving when the light is switched on, i.e. brightness in-fluences their behaviour, but only for a short time, asafter a while, they start to move once again.
We had not expected that the beetles would prefertemperatures of more than 30 °C because this seems tobe high for a nocturnal beetle (cf. Thiele, 1977). How-ever, typical habitats of S. europaea on Mediterraneanclay soils provide a wide variety of temperatures at thesame time of day. At the end of May, for example, wemeasured an ambient temperature of 28 °C at 12.30p.m. At the same time, the soil surface temperature was44 °C in the sun, 32 °C in the shade of a stone and 24°C in a soil fissure 20 cm below the surface. This meansthat the beetles can choose a temperature according totheir demands; this may also be possible in the night,when the soil temperatures are still high because of thestrong radiation present in Mediterranean habitats. Tem-perature preferences of more than 30 °C in central Eu-ropean species are only known in diurnal Cicindelidaeand a few Carabidae that use radiation to keep theirbody temperature above the ambient temperature (for asurvey see Thiele, 1977) in order to be ready to fly.
Moreover, resistance to desiccation can easily be in-terpreted as an adaptation to habitat conditions. Duringhot summers, the lack of liquid water may last for daysor longer depending on rainfall. From mid-April on,however, a clay soil is furrowed by deep cracks. At anambient temperature of 28 °C, we measured an rH inthe air of about 48% at a height of 1 m above ground.Only 20 cm deep in a soil fissure, rH was 86% and at 30cm it was >90%, which means that, in dry weather, theatmosphere stays saturated with water vapour deep inthe soil cracks and the beetles can always retreat intoconditions where dangerous water loss can be avoided.
In our experiments, the beetles often changed theirposition between the moistest and the driest sectors (cf.the maxima in Figure 9), probably indicating that Siago-na is also able to absorb water vapour to compensatefor water loss. Thus, this species might not depend onliquid water. However, we have not as yet investigatedthis possibility in greater detail.
Siagona and ants
We knew from former investigations (Zetto-Brand-mayr et al, 1998) that S. europaea is specialized on antsas prey. However, we were unaware of the closeness ofits relationships with this kind of prey. Numerousmyrmecophilous insect species use ants and their nestsas a food source and have developed different inge-nious ways to avoid being attacked by them (for a re-view and examples, see Hölldobler & Wilson, 1990).Siagona europaea does not belong to these species. In
all confrontations observed in our experiments, the antsdiscerned these beetles as an enemy and immediatelyeither retreated or attacked them, even when the bee-tles stayed motionless. The beetles never invaded antnests voluntarily. When they had accidentally enteredthe entrance of an ant nest, they were immediatelyheavily attacked, running around and struggling untilthey had found the way out. Hence, S. europaea isclearly not a myrmecophilous species. It is a myrme-cophagous ant predator that lives at the periphery ofant nests and/or near ant roads, lying in wait for singleants and catching them by surprise.
Intraspecific behaviour
Aggregations in Carabidae have been reported for sev-eral species. Their occurrence seems to be mainly re-stricted to the occupation of winter quarters (for a re-view see Thiele, 1977). Some species, however, aggre-gate all the year round. Greenslade (1963) observed aclumping effect in Nebria brevicollis, Luff (1984, Abstractin 14th Int. Congr. Entomol. Hamburg: 36l) found thatseveral species were significantly aggregated in pitfalltraps and suggested that the beetles defensive secretionsmay act as attractant pheromones at low concentrations.Wautier (1971) found an annual and a diurnal rhythm ofaggregation in two Brachinus species. Aggregation hada peak during the phase of sexual inactivity in winter.The diurnal maximum was in the morning, when thebeetles aggregated under stones, and the clusters dis-persed in the evening, when the beetles individuallysearched for prey. The two Brachinus species seemedto produce a persistent pheromone that maintained theattractivity of their aggregation sites over several years.The biological significance of aggregations can only bespeculated. One advantage might be that the sexes stayclose together, while another might be that a combinedreaction to hostile predators provides a better chance ofsurvival. The latter has been assumed for Brachinusspp., which possess effective defence glands.
The peaceful behaviour in aggregations of Siagonaspecies was unexpected because, outside aggregations,these beetles are extremely fast effective predators whenconfronted with ants. Their strong mandibles are suitedto crushing all the body parts of ants and thus the bee-tles could also inflict substantial damage on each other.However, we never saw this to happen; even the fewaggressive interactions that were observed ended in in-tensive beating with the antennae, resembling a ritual-ized fight. We also regularly observed that the beetlesshared their prey. However, this might be partly a resultof the adequate food supply in the laboratory. Hungrybeetles retreated, turned away or behaved aggressivelyagainst cospecifics while chewing an ant. Nevertheless,these beetles treated each other peacefully, and seemedto recognize each other through chemical cues. Thesecues are not strictly species specific, because specimensof S. jenissoni are also tolerated in aggregations of S. eu-ropaea. In Southern Spain, hundreds of individuals of S.
Downloaded By: [Università della Calabria] At: 09:11 22 January 2010

LIFE HISTORY OF SIAGONA EUROPAEA 41
jenissoni and S. dejeani were observed in aggregationsunder large stones (A. Cásale, personal communication).
Dying beetles are treated as prey and are eaten bytheir cospecifics. This has also been reported for someant species. Ants identify dying and dead cospecifics bya number of chemical breakdown products and mostspecies remove them from their nest (for a review seeHölldobler & Wilson, 1990).
Within the aggregations and especially before copula-tion, the beetles sometimes show the striking turningand friction behaviour described above. They appear tobe attempting to cover their body surface and especiallytheir backs with substances from the floor of theircrevice. The floors of the aggregation sites are alwayscovered with faeces. In Siagona beetles, these faeceshave a typical shape and consist of chitin particles ofants. They probably also contain pheromones. The exis-tence of a fixed action pattern for rubbing the body sur-face on the floor of the crevice (and on other beetles)indicates that covering the body with certain substancesis advantageous. We consider that this advantage resem-bles that in Brachinus species and has to do with de-fence. Ants are able to defend themselves and are thusa "difficult" prey. Moreover, they hunt insects them-selves and even Siagona beetles might serve as preywhen sufficient numbers of ants meet and attack a bee-tle outside flat crevices. On the other hand, we haveobserved that ants always recognize Siagona beetlesfrom some distance as hostile by olfactorial cues. Chem-ical analyses have revealed that some surface sub-stances of the beetles correspond to those of ants (Zet-to-Brandmayr et al., in prep.). This is not unexpectedbecause, as ant-hunters, the beetles come into closecontact with the body of their prey, both during the at-tack and afterwards when they chew part after part ofthe ant body. Hence, their body should be covered witha mixture of different substances, among them a cock-tail of defence substances of different ant species. Anaggregation of Siagona beetles should therefore repre-sent, for any ant (and probably also for other hostilepredators) an intensively and dangerously smelling area,but for the beetles, a safe shelter where they can rest. Ithas still to be tested experimentally whether there arepheromones of the beetles or substances released bythe ants that cause the aggregations in Siagona.
The mating behaviour of Siagona resembles that ofother Carabidae: the male mounts and, if the female isin agreement, copulation takes place. The mating be-haviour in S. jenissoni, however, is unusual for Cara-bidae as the females subsequently copulate with severalmales and both sexes behave peacefully. This seemsnot to be the case in S. europaea. Here, we observedseveral times that females aggressively refused to copu-late and attacked the male; this ended in the ritualizedantennal fighting described above.
Eggs and larvae
The egg of S. europaea is unusually large. Its lengthrepresents more than 26% of the adult body length, a
value much greater than that in most other ground bee-tles. In Elaphrus ssp. for example, egg length is ca. 16%of adult body length (cf. Bauer, 1974). In Broscuscephalotes, however, which produces only a few eggsand is famous for its subterranean brood-care, thelength is ca. 23% (Kempf, 1955).
Although the beetles had been well fed, we foundonly a few eggs and unfortunately did not succeed inkeeping them alive. We suggest that a female producesonly a few eggs and deposits them singly in soil fissuresand probably deeper in the soil. Their large size indi-cates that these eggs must be well hidden and that therisk of being predated is therefore low.
A larva of Siagona sp. (probably S. brunnipes) wasfirst described by Moore (1972). Grebennikov (1999)described the larvae of another unknown species. Theywere collected in Africa and caught in traps on moistclay soil. Moore (1972) proposed that Siagones inhabitdeep cracks in drying mudflats. The described larvaehad one and two stemmata, respectively, on their headsurfaces. The larva of 5. europaea looks similar but hasno stemma. Its most striking feature is the antennaewith their unsclerotized sticky apical part. The cleaningbehaviour of the antennae resembles the cleaning be-haviour of the larva of the carabid Loricera pilicornis.This larva has specialized sticky galeae that are used tocatch springtails and that are also drawn through themouthparts in order to clean and to cover them with aliquid (Bauer & Kredler, 1988).
As opposed to the imagines, the larva of S. europaeais a soft vulnerable cave-dwelling animal but an ex-tremely fast, rapidly reacting runner. It lives deep in soilcracks and does not feed on ants. It probably catchesother soft-bodied soil animals, such as small springtails,that might adhere to the sticky antennae and palpae (cf.Betz & Kölsch, 2004 for such a prey-capture behaviourin the staphylinid larvae of Stenus spp.). Unfortunately,we could observe only the 2nd larval stage. Other stagesof Siagona might have a closer association with antcolonies (cf. Zetto-Brandmayr et al, 1994).
REFERENCES
Adis J., Amorim M. A., Erwin T. E., Bauer Th., 1997 - On ecology,life history and survival strategies of a wing dimorphic groundbeetle (Col.: Carabidae: Odacanthini: Colliuris) inhabiting Cen-tral Amazonian inundation forests. Stud. Neotrop. Fauna & Env-iron, 32: 174-192.
Andrewes H. E., 1929 - Coleoptera, Carabidae. 1. Carabinae. In: J.Stephenson (ed.) The fauna of British India, including Burmaand Ceylon. Taylor and Francis, London.
Antoine M, 1955 - Coleopteres carabiques du Maroc (primier par-tie). La Rose, Paris.
Bauer Th., 1974 - Ethologische, autökologische und ökophysiolo-gische Untersuchungen an Elaphrus cupreus Dft. und Elaphrusriparius L. (Coleoptera, Carabidae). Oecologia, 14: 139-196.
Bauer Th., 1982 - Prey capture in a ground beetle larva. Anim.Behav., 30: 203-208.
Bauer Th, Kredler M, 1988 - Adhesive mouthparts in a groundbeetle larva (Coleoptera, Carabidae, Loricera pilicornis F.) andtheir function during predation. Zool. Anz, 221: 145-156.
Downloaded By: [Università della Calabria] At: 09:11 22 January 2010

42 T. BAUER, F. TALARICO, A. MAZZEI, A. GIGLIO, T. ZETTO-BRANDMAYR, P. BRANDMAYR, O. BETZ
Bauer Th., Kredler M., 1993 - Morphology of the compound eyesas an indicator of life-style in carabid beetles. Can. J. Zool., 71:799-810.
Betz O., Kölsch G., 2004 - The role of adhesion in prey captureand predator defence in arthropods. Arthropod Struct. Dev., 33:3-30.
Brandmayr P., Pizzolotto R., 1990 - Ground beetles coenoses inthe landscape of the Nebrodi Mountains, Sicily (Coleoptera,Carabidae). Il Naturalista Siciliano, 14: 51-64.
Casale A., Vigna Taglianti A., Juberthie C., 1998 - Coleoptera Cara-bidae. In: Juberthie C., Decu V. (eds), Encyclopaedia biospeo-logica, Tome II. Moulis, Bucarest, pp. 836-1373.
Christian E., 1999 - From sand pores to cave systems - subter-ranean biology in Austria. Decheniana (Bonn), 37: 121-126.
Erwin T. L., 1985 - The taxon pulse: a general pattern of lineageradiation and extinction among carabid beetles. In: G. E. Ball(ed.), Taxonomy, phylogeny and zoogeography of beetles andants. Junk, Dordrecht, pp. 435-472.
Galian J., Ortiz A. S., Serrano J., 1992 - A chromosome study oftwelve species of old and middle-aged lineages of Carabidae(Coleoptera). Elytron, 6: 143-150.
Greenlade P. J. M., 1963 - Further notes on aggregation in Cara-bidae (Coleoptera) with especial reference to Nebria brevicollis(F.) Entomol. Mon. Mag., 99: 109-114.
Grebennikov V. V., 1999 - Larvae of the supertribe Siagonitae:genera Siagona LATREILLE and Enceladas BONELLI(Coleoptera: Carabidae). Koleopt. Rdsch., 69: 1-10.
Hölldobler B., Wilson E. O., 1990 - The ants. Harvard UniversityPress, Cambridge (Mass.).
Kempf W., 1955 - Zur Biologie von Broscus cephalotes L. (Cara-bidae). Zool. Anz., 155: 30-33.
Moore B. P., 1972 - Description of the larva of Siagona(Coleoptera, Carabidae). J. Entomol., 41: 155-157.
Sokal R. R., Rohlf F. J., 1995 - Biometry. Freeman, New York.Thiele H. U., 1977 - Carabid beetles in their enviroments. In: D. S.
Farner (ed.), Zoophysiology and ecology, Vol 10. Springer,Berlin, Heidelberg, New York, pp. 1-169.
Wautier V., 1971 - Un phenomene social chez les Coleopteres: legregarisme de Brachinus (Caraboidea; Brachinidae). InsectesSoc, 18: 1-84.
Zetto-Brandmayr T., Giglio A., De Rose E., 1998 - Feeding behav-iour and food preference of Siagona europaea Dejean, amyrmecophagous carabid beetle (Coleoptera Carabidae). InsectSoc. Life, 2: 203-207.
Zetto-Brandmayr T., Marano I., Paarmann W., 1994 - Hypothesesconcerning the preying behaviour of the larvae of Graphipterusserrator Forskal, a myrmecophilous carabid beetle. Ethol. Ecol.Evol., 3: 119-124.
Zetto-Brandmayr T., Pizzolotto R., 1994 - Siagona europaea De-jean: first results from field collecting, life cycle, and the evi-dence of a possible myrmecophagous diet (Coleoptera, Cara-bidae, Siagonini). Entomologist, 113: 120-125.
Downloaded By: [Università della Calabria] At: 09:11 22 January 2010
Copyright © 2022 FDOKUMEN




















