
Carabus (Mesocarabus) dufourii (Autor: J.M. Barea) Los Carábidos (Coleoptera, Adephaga: Carabidae

PLEASE SCROLL DOWN FOR ARTICLE
This article was downloaded by: [Universita Della Calabria]On: 8 November 2010Access details: Access Details: [subscription number 917158269]Publisher Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
Ethology Ecology & EvolutionPublication details, including instructions for authors and subscription information:http://www.informaworld.com/smpp/title~content=t916668712
Electrophysiological and behavioural analyses on prey searching in themyrmecophagous carabid beetle Siagona europaea Dejean 1826(Coleoptera Carabidae)F. Talaricoa; P. G. Giulianinib; P. Brandmayra; A. Giglioa; C. Masalac; G. Sollaic; T. Zetto Brandmayra; P.Solaric
a Dipartimento di Ecologia, Università della Calabria, Via P. Bucci, Arcavacata di Rende, Cosenza, Italyb Dipartimento di Scienze della Vita, Università di Trieste, Via Giorgieri 7, Trieste, Italy c Dipartimentodi Biologia Sperimentale, Università di Cagliari, Cittadella Universitaria di Monserrato, SS 554 km4,500, Monserrato, Cagliari, Italy
Online publication date: 05 November 2010
To cite this Article Talarico, F. , Giulianini, P. G. , Brandmayr, P. , Giglio, A. , Masala, C. , Sollai, G. , Zetto Brandmayr, T.and Solari, P.(2010) 'Electrophysiological and behavioural analyses on prey searching in the myrmecophagous carabidbeetle Siagona europaea Dejean 1826 (Coleoptera Carabidae)', Ethology Ecology & Evolution, 22: 4, 375 — 384To link to this Article: DOI: 10.1080/03949370.2010.510044URL: http://dx.doi.org/10.1080/03949370.2010.510044
Full terms and conditions of use: http://www.informaworld.com/terms-and-conditions-of-access.pdf
This article may be used for research, teaching and private study purposes. Any substantial orsystematic reproduction, re-distribution, re-selling, loan or sub-licensing, systematic supply ordistribution in any form to anyone is expressly forbidden.
The publisher does not give any warranty express or implied or make any representation that the contentswill be complete or accurate or up to date. The accuracy of any instructions, formulae and drug dosesshould be independently verified with primary sources. The publisher shall not be liable for any loss,actions, claims, proceedings, demand or costs or damages whatsoever or howsoever caused arising directlyor indirectly in connection with or arising out of the use of this material.

Ethology Ecology & Evolution 22: 375–384, 2010
ISSN 0394-9370 print/ISSN 1828-7131 online© 2010 Dipartimento di Biologia Evoluzionistica dell’Università, Firenze, ItaliaDOI: 10.1080/03949370.2010.510044http://www.informaworld.com
TEEE0394-93701828-7131Ethology Ecology & Evolution, Vol. 22, No. 4, Jul 2010: pp. 0–0Ethology Ecology & EvolutionElectrophysiological and behavioural analyses on prey searching in the myrmecophagous carabid beetle Siagona europaea Dejean 1826 (Coleoptera Carabidae)
Prey searching in F. Talarico et al.F. TALARICO 1,4, P.G. GIULIANINI 2, P. BRANDMAYR 1, A. GIGLIO 1, C. MASALA 3, G. SOLLAI 3, T. ZETTO BRANDMAYR 1 and P. SOLARI 3
1 Dipartimento di Ecologia, Università della Calabria, Via P. Bucci I-87036 Arcavacata diRende (Cosenza), Italy2 Dipartimento di Scienze della Vita, Università di Trieste, Via Giorgieri 7, I-34127 Trieste,Italy3 Dipartimento di Biologia Sperimentale, Università di Cagliari, Cittadella Universitaria diMonserrato, SS 554 km 4,500, I-09042 Monserrato (Cagliari), Italy
Received 18 December 2009, accepted 13 May 2010
In the present study, electrophysiological and behavioural tests were performedon Siagona europaea Dejean 1826 adults to investigate the importance of chemicalcues in ant location. Olfactory responses of labial palp to terpenoids (citral, limoneneand geraniol), carboxylic acids (acetic and formic acids), aliphatic ketones (heptanone)and aldehydes (hexanal) were tested by means of an electropalpogram (EPG) bio-assay. Our electrophysiological results showed that only acetic and formic acids evokedEPG responses. Y-shape olfactometer bioassays showed that adults were attracted byacetic and formic acids. The role of these chemicals on prey detection is discussed.
KEY WORDS: labial palps, predation, acetic acid, formic acid, ants.
INTRODUCTION
Predators use a variety of stimuli in order to locate prey. It is well known that ininsects for many specialised predators the first step is to discover the prey’s habitat.Since final location of prey is commonly achieved by means of its taste and odour,then in this phase the sensory system plays a crucial role. The adaptive value of senseorgans involved in prey detection was investigated in a variety of insect taxa (ALTNER &PRILLINGER 1980; ZACHARUK 1980, 1985; ALTNER et al. 1983; FORSYTHE 1983; KEIL 1997,1998; BLAND et al. 1998; MITCHELL et al. 1999; SHANBHAG et al. 1999, 2000). Someground beetles find their food by random search: diurnal species use vision (MORWINSKY
& BAUER 1997), while other species use olfactory cues or contact sense organs. It is appar-ent that the distribution pattern of olfactory receptors is related to the specific searching
4 Corresponding author: Federica Talarico, Dipartimento di Ecologia, Università della Calabria,Via P. Bucci cubo 4B, I-87036 Arcavacata di Rende (Cosenza), Italy (E-mail: [email protected]).
Downloaded By: [Universita Della Calabria] At: 15:09 8 November 2010

376 F. Talarico et al.
behaviour of each species. Those using tactile and olfactory mechanisms locate theprey through sensilla on the antennae and the labial and maxillary palps (WHEATER
1989; KIELTY et al. 1996; LÖVEI & SUNDERLAND 1996). In carabid beetles, ultrastruc-tural, electrophysiological and behavioural investigations show that prey recognitionand habitat detection are mostly mediated by olfactory cues, involving mainly anten-nal receptors (DALY & RYAN 1979; EVANS 1983; KIELTY et al. 1996; MERIVEE et al. 2000,2001, 2002, 2004, 2005; MILIUS et al. 2006; MUST et al. 2006a, 2006b; DI GIULIO et al.2009).
Siagona europaea is an exclusive myrmecophage of both adult ant and brood(ZETTO BRANDMAYR & PIZZOLOTTO 1994; ZETTO BRANDMAYR et al. 1998; BAUER et al.2005). It has a wide distribution around the Mediterranean Sea, in Africa and India,preferring in Italy the open lands of the Mediterranean biome (sclerophylls). In South-ern Italy, S. europaea occurs in pastures and abandoned fields only on clay soils up toabout 250 m a.s.l., while in the Calabria Region it reaches approx. 450 m a.s.l. In earlyspring, when soil moisture is high, the beetles are found under stones. From mid-Aprilonwards, when the soil dries out and becomes deeply fissured, they retreat into deepercrevices, especially during the hot dry hours of the day. Their activity is mainly noctur-nal as shown by behavioural recordings and by the structure of their compound eyes,with a value of ommatidia/mm body length typical for a nocturnal species (BAUER &KREDLER 1993; ZETTO BRANDMAYR & PIZZOLOTTO 1994; ZETTO BRANDMAYR et al. 2000a,2000b; BAUER et al. 2005; GIGLIO et al. 2005; TALARICO et al. 2007). The preying behav-iour is marked by a strong involvement of the antennae and labial palps during thesearching phase (ZETTO BRANDMAYR et al. 2000b, 2002, 2007). The carabid beetle scansthe surrounding area and the soil by flicking the antennae and by tapping of the labialpalps on the moist clay surface of the cracks. Therefore, in the absence of visual cues alarge and diverse sensory complement, including chemical and tactile, is instrumentalto predation. The labial palp of S. europaea presents five types of sensilla: three types ofsensilla basiconica on the apical sensory area and two types, one sensillum trichodeumand one coeloconicum, on the external palp surface. On morphological grounds thes. basiconica type 1 were considered as olfactory, the type 2 as gustatory, the type 3and the s. trichodeum as mechanoreceptive, and the s. coeloconicum as a thermo/hygroreceptor (GIGLIO et al. 2010). This sensorial pattern proves that the labial palpplays a key role as sensorial appendage. The aim of the present research is to evaluatethe involvement of the labial palp in prey searching. Therefore, we performed (i) elec-trophysiological recordings to investigate the olfactory responses of the labial palp and(ii) behavioural bioassays to measure the detection capabilities of specimens and theattractive role of tested substances.
MATERIALS AND METHODS
Insects
S. europaea specimens were collected in southern Italy (Calabria) from open fields and pas-tures (250 m a.s.l.) (Squillace, Catanzaro) by bait-traps in three different years: in spring 2003and 2004 for electrophysiological analysis and in spring 2009 for behavioural analysis.
All beetles were maintained in the laboratory in 5-litre plastic boxes with humidified soilunder standardised condition (light regime of L/D = 16/8 hr, 70% RH and at a day/night tempera-ture of 25–20 °C) and were fed with live ants (Tapinoma nigerrimum, Messor sp.). One weekbefore use in behavioural bioassay, insects were removed from plastic boxes, placed separately in200 ml boxes and were not fed. The same insects were not tested on consecutive days.
Downloaded By: [Universita Della Calabria] At: 15:09 8 November 2010

Prey searching in Siagona europaea 377
Electrophysiology analyses
Electropalpogram recording technique. Electropalpogram (EPGs) recordings, which detectthe summed potentials of olfactory receptor neurons in the palp, were performed on single labialpalps, one per beetle, excised and positioned in such a way as to expose the largest olfactory sur-face to the stimulus air flow (RIESGO-ESCOVAR et al. 1995; VETTER et al. 2006).
The recording electrode, a glass micropipette (20 μm in average tip diameter) filled with a50 mM NaCl aqueous solution containing an Ag/AgCl wire, was gently pressed against the palp; asimilar Ag/AgCl wire, inserted into the base of the palpal shaft, acted as the reference electrode.EPGs were recorded with a high input impedance (1015 Ω) electrometer (WPI DUO 773), band-pass filtered (DC to 1 kHz), digitised by means of the Axon Digidata 1200B A/D converter (10,000Hz) and stored on computer for further analyses. Amplitudes and waveforms of EPGs were calcu-lated by means of the Axoscope 8.1 software.
Odour delivery system. An air-stimulus control unit (model CS-55, Syntech, Hilversum, theNetherlands) was used for air and odour delivery with a constant flow rate (1500 ml/min) ofcharcoal-filtered and humidified air continuously blown over the labial palp through the openend of a glass tube (8 mm in diameter, 15 cm long), positioned 15 mm from the palp. Duringodour stimulation, the stimulus-bearing airflow (500 ml/min) was switched for 2 sec to a Pasteurpipette (15 cm long), containing a pleated filter paper strip impregnated with the stimulus,inserted at a right angle into a small port in the wall of the main airstream tube: the stimuluscartridge headspace air was thus added to the main continuous clean air flow. Stimuli were pre-sented in a randomised sequence, separated by interstimulus intervals of at least 2 min. Eachstimulus was tested more than once to verify the reproducibility of the responses. The air con-taining the stimulus was removed from the experimental arena by means of a suction pumpoperating at a flow rate slightly higher than that of stimulation.
The glass pipette, which was kept plugged at both ends when not in use, and kept frozen at− 20 °C between test sessions, contained a renewable strip of filter paper soaked with the odourcompound that allowed for a series of repeated stimulations without any appreciable concentra-tion decay. The filter paper was anyway replaced weekly.
Stimulating compounds. The following compounds were tested as stimuli: (a) citral, limoneneand geraniol (terpenoids), formic acid (carboxylic acid), found in ant secretion (HÖLLDOBLER &WILSON 1990); (b) heptanone (aliphatic ketone) and hexanal (aldehyde), commonly tested in olfactionstudies in insects (THIERY & MARION-POLL 1998; STOPFER et al. 2003); and (c) acetic acid (carboxylicacid), commonly used as a trap bait to catch carabid beetles (THIELE 1977). All chemicals used wereobtained from Sigma Chemicals (purity grade ≥ 98%, except for citral ≥ 95%). Each chemical wasfirst dissolved (100 μg/ml) in dichloromethane (CH2Cl2) and then a 5 μl volume of solution was pipet-ted onto the pleated strip of filter paper (80 × 5 mm), to yield a final dosage of 0.5 μg for each com-pound effectively loaded on filter paper at the highest dosage tested. The CH2Cl2 was evaporatedbefore the experiments started. To obtain the other (lower) concentrations, decadic dilutions from theformer were prepared in CH2Cl2 and 5 μl of each solution was pipetted onto the filter paper strip withthe same procedure, in order to obtain a 0.05 or 0.005 μg load on each filter paper; each compoundwas tested in increasing sequence.
Preliminary experiments indicated that blank filter paper, after CH2Cl2 evaporation, is notstimulating. In each experiment, before each stimulation the response to air as a control formeasuring the presence of a possible mechanoreceptor response component was tested and itsvalue subtracted from the EPG value obtained in response to the test stimulation.
Behavioural bioassays
The experimental device consisted of a Y-shaped olfactometer made of transparent plex-iglas (2.5 cm diameter; length of common tube and two side arms = 12 cm; 8 cm height). At theend of the lateral arms, supports were placed for the substances to be tested. At the beginning
Downloaded By: [Universita Della Calabria] At: 15:09 8 November 2010

378 F. Talarico et al.
of each trial, two electric fans provided a slight and continuous one-way current of air (velocity2 m/sec) moving from the two lateral arms to the central one and then going out through the holein its end wall.
In this experiment, we tested only chemicals (acetic and formic acids) that evoked anelectrophysiological response. To test volatile chemicals, 5 μl of a test mixture or of the puresolvent (dichloromethane) was applied on a piece of filter paper (2 × 0.5 mm) and placed ateach end of the Y tube arms. The concentrations tested were prepared as for the electrophysio-logical tests. Acetic and formic acids were dissolved in dichloromethane in order to obtain 0.5,0.05 or 0.005 μg of chemicals. Each compound was tested in increasing sequences. Beetles wereallowed to choose between: (i) in the first trials, acetic acid dissolved in dichloromethane versusdichloromethane; (ii) in the second trials, formic acid dissolved in dichloromethane versusdichloromethane (control); and (iii) in the third trials, acetic acid versus formic acid, both dis-solved in dichloromethane (control).
Ten replicates (each using a different individual) were used for each experiment and a totalof 90 trials were performed. The animal to be tested was put in the base on the common arm andafter a 2 min settling-down period the activity was recorded for 10 min. A choice was recordedwhen a beetle ran in one of two arms and remained there for at least 1 min. If the beetle stayed inthe common tube or at the junction of the two arms after 10 min, the test was stopped and thebeetle was considered to be a non-responding individual. Before the beginning of each test thefilter paper imbued with treatment solution and control were exposed to the air of a fan for 2 minin order to let the solvent evaporate. Tests were conducted during May 2009 in red light conditionsto avoid influence of phototaxis at 27 ± 2 °C temperature, 60–70% relative humidity. At the end ofeach trial, the tested beetle was removed, the experimental device was cleaned and a new filterpaper was used. Each beetle was tested only once.
Statistical analyses
The paired Student’s t-test (Statistical software package) with a 95% confidence level (P ≤0.05) was used to assess differences in EPG amplitudes, by comparing results obtained for eachcompound with respective air controls.
For behavioural bioassays, the preference for acetic acid and formic acid in each pair-wisecombination was tested using the Sign test.
RESULTS
Electrophysiological experiments
As assessed by EPG responsiveness of the labial palps in S. europaea, among thetested compounds only the two carboxylic acids, acetic and formic, were found to beeffective. In fact, at all the tested doses they evoked negative EPGs (depolarisingdownward deflections) of fairly constant and reproducible time course and character-istic shape in a dose-dependent manner (Fig. 1). In detail, as shown in Fig. 2, aceticacid evoked EPG responses of 7.86 ± 1.49 and 3.06 ± 0.44 mV (n = 9) at the two high-est doses, 0.5 and 0.05 μg, respectively, which were significantly higher with respectto that elicited by the pure air control (− 0.22 ± 0.10 n = 9; P < 0.001). At 0.005 μg,acetic acid was not effective at all. Besides, at the same doses formic acid elicited aresponse from the labial palps of 10.43 ± 1.75, 4.67 ± 0.90 and 0.98 ± 0.17 mV (n = 9),significantly different with respect to the air control (0.20 ± 0.15 mV, n = 9). As for theremaining compounds, they did not elicit any response distinguishable from thecontrol.
Downloaded By: [Universita Della Calabria] At: 15:09 8 November 2010
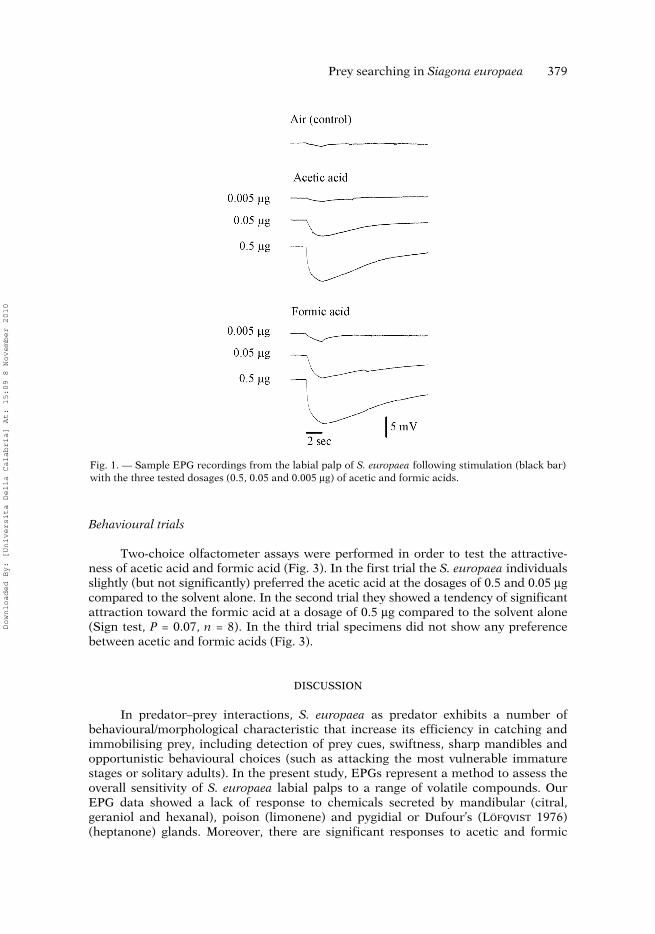
Prey searching in Siagona europaea 379
Behavioural trials
Two-choice olfactometer assays were performed in order to test the attractive-ness of acetic acid and formic acid (Fig. 3). In the first trial the S. europaea individualsslightly (but not significantly) preferred the acetic acid at the dosages of 0.5 and 0.05 μgcompared to the solvent alone. In the second trial they showed a tendency of significantattraction toward the formic acid at a dosage of 0.5 μg compared to the solvent alone(Sign test, P = 0.07, n = 8). In the third trial specimens did not show any preferencebetween acetic and formic acids (Fig. 3).
DISCUSSION
In predator–prey interactions, S. europaea as predator exhibits a number ofbehavioural/morphological characteristic that increase its efficiency in catching andimmobilising prey, including detection of prey cues, swiftness, sharp mandibles andopportunistic behavioural choices (such as attacking the most vulnerable immaturestages or solitary adults). In the present study, EPGs represent a method to assess theoverall sensitivity of S. europaea labial palps to a range of volatile compounds. OurEPG data showed a lack of response to chemicals secreted by mandibular (citral,geraniol and hexanal), poison (limonene) and pygidial or Dufour’s (LÖFQVIST 1976)(heptanone) glands. Moreover, there are significant responses to acetic and formic
Fig. 1. — Sample EPG recordings from the labial palp of S. europaea following stimulation (black bar)with the three tested dosages (0.5, 0.05 and 0.005 μg) of acetic and formic acids.
Downloaded By: [Universita Della Calabria] At: 15:09 8 November 2010
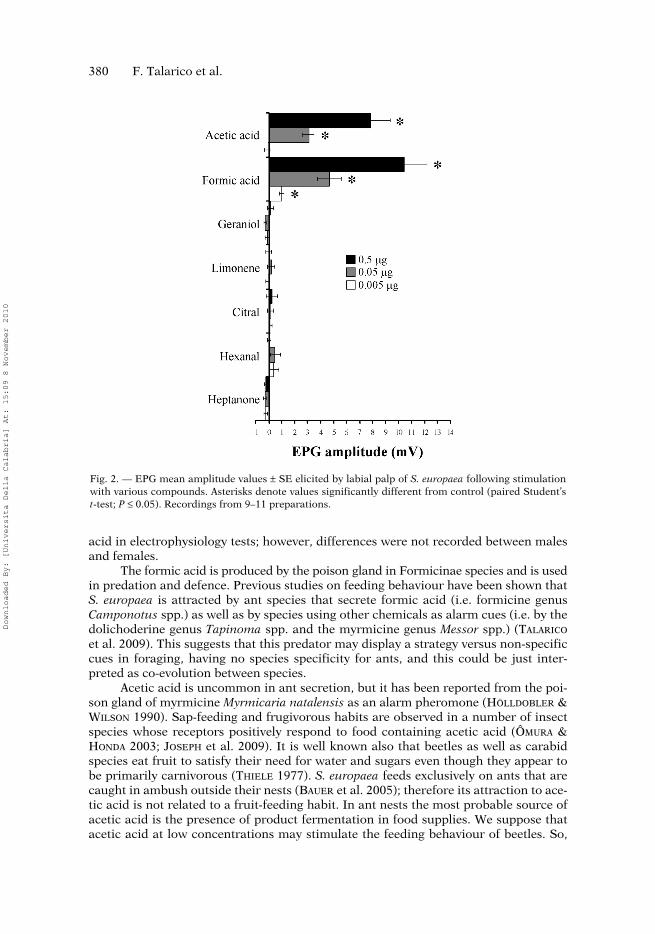
380 F. Talarico et al.
acid in electrophysiology tests; however, differences were not recorded between malesand females.
The formic acid is produced by the poison gland in Formicinae species and is usedin predation and defence. Previous studies on feeding behaviour have been shown thatS. europaea is attracted by ant species that secrete formic acid (i.e. formicine genusCamponotus spp.) as well as by species using other chemicals as alarm cues (i.e. by thedolichoderine genus Tapinoma spp. and the myrmicine genus Messor spp.) (TALARICO
et al. 2009). This suggests that this predator may display a strategy versus non-specificcues in foraging, having no species specificity for ants, and this could be just inter-preted as co-evolution between species.
Acetic acid is uncommon in ant secretion, but it has been reported from the poi-son gland of myrmicine Myrmicaria natalensis as an alarm pheromone (HÖLLDOBLER &WILSON 1990). Sap-feeding and frugivorous habits are observed in a number of insectspecies whose receptors positively respond to food containing acetic acid (ÔMURA &HONDA 2003; JOSEPH et al. 2009). It is well known also that beetles as well as carabidspecies eat fruit to satisfy their need for water and sugars even though they appear tobe primarily carnivorous (THIELE 1977). S. europaea feeds exclusively on ants that arecaught in ambush outside their nests (BAUER et al. 2005); therefore its attraction to ace-tic acid is not related to a fruit-feeding habit. In ant nests the most probable source ofacetic acid is the presence of product fermentation in food supplies. We suppose thatacetic acid at low concentrations may stimulate the feeding behaviour of beetles. So,
Fig. 2. — EPG mean amplitude values ± SE elicited by labial palp of S. europaea following stimulationwith various compounds. Asterisks denote values significantly different from control (paired Student’st-test; P ≤ 0.05). Recordings from 9–11 preparations.
Downloaded By: [Universita Della Calabria] At: 15:09 8 November 2010

Prey searching in Siagona europaea 381
this myrmecophagous species, exploring deep crevices, may smell acetic acid from thefood chamber of ant nests and catch the ants by surprise attacks.
Results from the behavioural studies clearly show that beetles were able to detectat a long range both formic and acetic acid and that these chemicals stimulate foragingbehaviour. The common trend is that in ground beetles the antennae are involved inprey detection. MERIVEE et al. (2007, 2008) have shown that antennal contact chemore-ceptors of some ground beetles, such as Pterostichus oblongopunctatus and P. aethiops,use a sugar-sensitive neuron which is involved in seed predation. However, electro-physiological tests suggest that final location of prey in S. europaea is achieved bymeans of chemical cues detected by the palp olfactory system. Indeed, this species doesnot show the features of a visual hunter, having small eyes typical of nocturnal species(BAUER et al. 2005; TALARICO et al. 2007).
In conclusion, electrophysiological recordings show that the labial palps ofS. europaea are responsive to volatile stimulation with both acetic and formic acid andmay play a critical role in prey search. These results are strengthened by behaviouralexperiments, thus confirming that this Carabid is primarily an olfactory hunter capa-ble of recognising a large spectrum of ant prey. However, further research is needed toinvestigate the role of antenna and maxillary palp on prey searching.
ACKNOWLEDGEMENTS
This work was supported by the Italian Ministero dell’Università e della Ricerca (MiUR).The authors are grateful to Professors E. Ferrero (Department of Life Sciences, University of
Fig. 3. — Attraction of S. europaea to acetic acid and formic acid samples (at dosage from 0.5 to 0.005 μgdissolved in 5 μl dichloromethane) presented in a Y-tube olfactometer. Beetles that made no choicewere excluded from the analysis.
0.005 µg acetic acid 0.005 µg formic acid
0.05 µg formic acid
0.5 µg formic acid
0.005 µg formic acid
0.05 µg formic acid
0.5 µg formic acid
0.005 µg acetic acid
0.05 µg acetic acid
0.5 µg acetic acid
0.05 µg acetic acid
0.5 µg acetic acid
solvent
solvent
solvent
solvent
solvent
solvent
–50 0 50Frequency of decisions (%)
100
Downloaded By: [Universita Della Calabria] At: 15:09 8 November 2010

382 F. Talarico et al.
Trieste), R. Crnjar and A. Liscia (Department of Experimental Biology, University of Cagliari) forhelpful suggestions. Experiments comply with the ‘Principles of animal care’, Publication No.86-23, revised 1985, of the National Institute of Health, and also with the current laws of the EC.
REFERENCES
ALTNER H. & PRILLINGER L. 1980. Ultrastructure of invertebrate chemo- thermo, and hygrorecep-tors and its functional significance. International Review of Cytology: 69–138.
ALTNER H., SCHALLER-SELZER L., STETTER H. & WOHLRAB I. 1983. Poreless sensilla with inflexiblesockets. Cell and Tissue Research 234: 279–307.
BAUER T. & KREDLER M. 1993. Morphology of the compound eyes as an indicator of life-style incarabid beetles. Canadian Journal of Zoology 71: 799–810.
BAUER T., TALARICO F., MAZZEI A., GIGLIO A., ZETTO BRANDMAYR T., BRANDMAYR P. & BETZ O. 2005.Hunting ants in Mediterranean clay soils: life history of Siagona europaea (Coleoptera,Carabidae). Italian Journal of Zoology 72: 33–42.
BLAND R.G., SLANEY D.P. & WEINSTEIN P. 1998. Mouthpart sensilla of cave species of AustralianParatemnopteryx cockroaches (Blatteria: Blattellidae). International Journal of InsectMorphology & Embryology 27: 291–300.
DALY P.J. & RYAN M.F. 1979. Ultrastructure of antennal sensilla of Nebria brevicollis (Fab.)(Coleoptera: Carabidae). International Journal of Insect Morphology and Embryology 8:169–181.
DI GIULIO A., ROSSI STACCONI M.V. & ROMANI R. 2009. Fine structure of the antennal glands of theant nest beetle Paussus favieri (Coleoptera, Carabidae, Paussini). Arthropod Structure &Development 38: 293–302.
EVANS W.G. 1983. Habitat selection in the Carabidae. The Coleopterists Bulletin 37: 164–167.FORSYTHE T.G. 1983. Mouthparts and feeding of certain ground beetles (Coleoptera: Carabidae).
Zoological Journal of the Linnean Society 79: 319–376.GIGLIO A., FERRERO E.A., PERROTTA E., TALARICO F.F. & BRANDMAYR T.Z. 2010. Sensory structures
involved in prey detection on the labial palp of the ant-hunting beetle Siagona europaea Dejean1826 (Coleoptera, Carabidae). Acta Zoologica 91: 328–334.
GIGLIO A., FERRERO E.A. & ZETTO BRANDMAYR T. 2005. Ultrastructural identification of the anten-nal gland complement in Siagona europaea Dejean 1826, a myrmecophagous carabid bee-tle. Acta Zoologica 86: 195–203.
HÖLLDOBLER B. & WILSON E.O. 1990. The ants. Berlin, Heidelberg: Springer-Verlag.JOSEPH R.M., DEVINENI A.V., KING I.F.G. & HEBERLEIN U. 2009. Oviposition preference for and
positional avoidance of acetic acid provide a model for competing behavioral drives inDrosophila. Proceedings of the National Academy of Sciences of the USA 106: 11352–11357.
KEIL T.A. 1997. Functional morphology of insect mechanoreceptors. Microscopy Research andTechnique 39: 506–531.
KEIL T.A. 1998. The structure of integumental mechanoreceptors, pp. 385–404. In: Harrison F.W.,Ed. Microscopic anatomy of invertebrates. New York: Wiley-Liss.
KIELTY J., ALLEN-WILLIAMS L., UNDERWOOD N. & EASTWOOD E. 1996. Behavioral responses of threespecies of ground beetle (Coleoptera: Carabidae) to olfactory cues associated with prey andhabitat. Journal of Insect Behavior 9: 237–250.
LÖFQVIST J. 1976. Formic acid and saturated hydrocarbons as alarm pheromones for the antFormica rufa. Journal of Insect Physiology 22: 1331–1346.
LÖVEI G.L. & SUNDERLAND K.D. 1996. Ecology and behavior of ground beetles (Coleoptera:Carabidae). Annual Review of Entomology 41: 231–256.
MERIVEE E., MÄRTMANN H., MUST A., MILIUS M., WILLIAMS I. & MÄND M. 2008. Electrophysiologicalresponses from neurons of antennal taste sensilla in the polyphagous predatory groundbeetle Pterostichus oblongopunctatus (Fabricius 1787) to plant sugars and amino acids.Journal of Insect Physiology 54: 1213–1219.
Downloaded By: [Universita Della Calabria] At: 15:09 8 November 2010

Prey searching in Siagona europaea 383
MERIVEE E., MUST A., MILIUS M. & LUIK A. 2007. Electrophysiological identification of the sugarcell in antennal taste sensilla of the predatory ground beetle Pterostichus aethiops. Journalof Insect Physiology 53: 377–384.
MERIVEE E., PLOOMI A., LUIK A., RAHI M. & SAMMELSELG V. 2001. Antennal sensilla of the groundbeetle Platynus dorsalis (Pontoppidan, 1763) (Coleoptera, Carabidae). Microscopy Researchand Technique 55: 339–349.
MERIVEE E., PLOOMI A., MILIUS M., LUIK A. & HEIDEMAA M. 2005. Electrophysiological identifica-tion of antennal pH receptors in the ground beetle Pterostichus oblongopunctatus. Physio-logical Entomology 30: 122–133.
MERIVEE E., PLOOMI A., RAHI M., BRESCIANI J., RAVN H. P., LUIK A. & SAMMELSELG V. 2002. Anten-nal sensilla of the ground beetle Bembidion properans Steph. (Coleoptera, Carabidae).Micron 33: 429–440.
MERIVEE E., PLOOMI A., RAHI M., LUIK A. & SAMMELSELG V. 2000. Antennal sensilla of the groundbeetle Bembidion lampros Hbst (Coleoptera, Carabidae). Acta Zoologica 81: 339–350.
MERIVEE E., RENOU M., MAND M., LUIK A., HEIDEMAA M. & PLOOMI A. 2004. Electrophysiologicalresponses to salts from antennal chaetoid taste sensilla of the ground beetle Pterostichusaethiops. Journal of Insect Physiology 50: 1001–1013.
MILIUS M., MERIVEE E., WILLIAMS I., LUIK A., MAND M. & MUST A. 2006. A new method for electro-physiological identification of antennal pH receptor cells in ground beetles: the example ofPterostichus aethiops (Panzer, 1796) (Coleoptera, Carabidae). Journal of Insect Physiology52: 960–967.
MITCHELL B.K., ITAGAKI H. & MARIE-PASCALE R. 1999. Peripheral and central structures involved ininsect gustation. Microscopy Research and Technique 47: 401–415.
MORWINSKY T. & BAUER T. 1997. Prediction of life style by eye morphology in Bembidion species(Coleoptera; Carabidae). Pedobiologia 41: 472–480.
MUST A., MERIVEE E., LUIK A., MAND M. & HEIDEMAA M. 2006a. Responses of antennal campani-form sensilla to rapid temperature changes in ground beetles of the tribe platynini withdifferent habitat preferences and daily activity rhythms. Journal of Insect Physiology 52:506–513.
MUST A., MERIVEE E., MAND M., LUIK A. & HEIDEMAA M. 2006b. Electrophysiological responses ofthe antennal campaniform sensilla to rapid changes of temperature in the ground beetlesPterostichus oblongopunctatus and Poecilus cupreus (Tribe Pterostichini) with differentecological preferences. Physiological Entomology 31: 278–285.
ÔMURA H. & HONDA K. 2003. Feeding responses of adult butterflies, Nymphalis xanthomelas,Kaniska canace and Vanessa indica, to components in tree sap and rotting fruits: synergis-tic effects of ethanol and acetic acid on sugar responsiveness. Journal of Insect Physiology49: 1031–1038.
RIESGO-ESCOVAR J., RAHA D. & CARLSON J.R. 1995. Requirement for a phospholipase C in odorresponse: overlap between olfaction and vision in Drosophila. Neurobiology 92: 2864–2868.
SHANBHAG S.R., MÜLLER B. & STEINBRECHT R.A. 1999. Atlas of olfactory organs of Drosophila mela-nogaster: 1. Types, external organization, innervation and distribution of olfactory sensilla.International Journal of Insect Morphology and Embryology 28: 377–397.
SHANBHAG S.R., MÜLLER B. & STEINBRECHT R.A. 2000. Atlas of olfactory organs of Drosophila mela-nogaster: 2. Internal organization and cellular architecture of olfactory sensilla. ArthropodStructure & Development 29: 211–229.
STOPFER M., JAYARAMAN V. & LAURENT G. 2003. Intensity versus identity coding in an olfactorysystem. Neuron 39: 991–1004.
TALARICO F., BONACCI T., BRANDMAYR P., DALPOZZO R., DE NINO A., GIGLIO A., TAGARELLI A. & ZETTO
BRANDMAYR T. 2009. Avoiding ant detection in Siagona europaea Dejean 1826 (ColeopteraCarabidae): an evolutionary step towards true myrmecophily. Ethology Ecology & Evolution21: 45–61.
TALARICO F., ROMEO M., MASSOLO A., BRANDMAYR P. & ZETTO T. 2007. Morphometry and eyemorphology in three species of Carabus (Coleoptera: Carabidae) in relation to habitatdemands. Journal of Zoological Systematics and Evolutionary Research 45: 33–38.
THIELE H.-U. 1977. Carabid beetles in their environments. Berlin: Springer-Verlag.
Downloaded By: [Universita Della Calabria] At: 15:09 8 November 2010

384 F. Talarico et al.
THIERY D. & MARION-POLL F. 1998. Electroantennogram responses of Douglas-fir seed chalcids toplant volatiles. Journal of Insect Physiology 44: 483–490.
VETTER R.S., SAGE A.E., JUSTUS K.A., CARDÈ R.T. & GALIZIA C.G. 2006. Temporal integrity of an air-nborne odor stimulus is greatly affected by physical aspects of the odor delivery system.Chemical Senses 31: 359–369.
WHEATER C.P. 1989. Prey detection by some predatory Coleoptera (Carabidae and Staphylinidae).Journal Zoological Society of London 218: 171–185.
ZACHARUK R.Y. 1980. Ultrastructure and function of insect chemosensilla. Annual Review ofEntomology 25: 27–47.
ZACHARUK R.Y. 1985. Antennae and sensilla, pp. 1–69. In: Kerkut G.A. & Gilbert L.I., Eds. Com-parative insect physiology, biochemistry and pharmacology. Oxford: Pergamon Press.
ZETTO BRANDMAYR T., BONACCI T., DE ROSE E., GIGLIO A. & TALARICO F. 2002. Interaction betweenants and Siagona europaea Dejean (Coleoptera, Carabidae) inside the nest: tollerance orhostility? Insect Social Life 4: 227–232.
ZETTO BRANDMAYR T., DALPOZZO R., DE NINO A., DE ROSE E., GIGLIO A., PROCOPIO A., SINDONA G. &TALARICO F.F. 2000a. Siagona europaea Dejean (Coleoptera, Carabidae) an ant predatorliving with ants: hypothesis about chemical mimicry. Insect Social Life 3: 201–208.
ZETTO BRANDMAYR T., DE ROSE E., GIGLIO A. & PIZZOLOTTO R. 2000b. Morphological and behaviouralaspects favouring ant predation in Siagona europaea (Coleoptera, Carabidae), pp. 103–109.In: Brandmayr P. et al., Eds. Natural history and applied ecology of carabid beetles. Sofia:Pensoft.
ZETTO BRANDMAYR T., GIGLIO A. & DE ROSE E. 1998. Feeding behaviour and food preference ofSiagona europaea Dejean, a myrmecophagous carabid beetle (Coleoptera, Carabidae).Insect Social Life 2: 203–207.
ZETTO BRANDMAYR T., MAZZEI A., TALARICO F., GIGLIO A., BAUER T. & BRANDMAYR P. 2007. The larvaof Siagona europaea Dejean, 1826: morphology and collecting technique for a subterraneanblind “running ant killer” (Coleoptera, Carabidae). Italian Journal of Zoology 74: 239–245.
ZETTO BRANDMAYR T. & PIZZOLOTTO R. 1994. Siagona europaea Dejean: first result from fieldcollecting, life cycle and the evidence of a possible myrmecophagous diet (Coleoptera,Carabidae, Siagonini). The Entomologist 113: 120–125.
Downloaded By: [Universita Della Calabria] At: 15:09 8 November 2010
Copyright © 2022 FDOKUMEN




















