
Correlated patterns of tracheal compression and convective gas exchange in a carabid beetle

ORIGINAL PAPER
In situ extinction of carabid beetles and community changesin a protected suburban forest during the past century:the ‘‘Bosco Farneto’’ near Trieste (Italy)
Pietro Brandmayr Æ Roberto Pizzolotto ÆGiorgio Colombetta Æ Tullia Zetto
Received: 13 June 2007 / Accepted: 18 March 2008 / Published online: 8 April 2008
� Springer Science+Business Media B.V. 2008
Abstract The carabid beetle species assemblages (14
sample sites) of a 238 ha urban oak forest in Trieste, Italy,
studied in 1983–84 with pitfall traps, were compared with
an historic list of 57 species hand collected by entomolo-
gists in the same forest before it was logged in 1944.Trap
data have been improved by hand collections to get a
species list as complete as the historic one. Multivariate
analysis was used to group the sites (14 plus the historic
list) into three assemblages of brooks, clearings and forests.
After the Second World War logging, the secondary eco-
logical succession resulted in a lower species number, with
a trend to a new equilibrium. Some important forest spe-
cialists, such as Laemostenus venustus, have been lost.
Human pressure and recolonisation by carabids are still
occurring. The in situ extinction of ground beetles near the
end of the last century reached values between 57 and 64%
of the species historically recorded. Consequently, urgent
restoration measures are required for waterside habitats,
forest and land management, to maintain small open areas
or clearings. The study of carabid species assemblages and
habitat affinities seems a useful tool for adaptive manage-
ment of forests affected by human activities, because
changes in carabid species number and type can be easily
related to human disturbance.
Keywords Biodiversity loss � Carabidae �Human disturbance � Land use change � Local extinction
Introduction
Carabid and tiger beetles are increasingly used as valid
biological indicators in an attempt to develop models of
biodiversity variation related to environmental degradation
(Thiele 1977; Stork 1990; Pearson and Cassola 1992; Des-
ender et al. 1994b; Niemela 1996; Brandmayr et al. 1996;
Luff 1996). The coleopteran family Carabidae is rich in
species and well-known, at least in some fundamental eco-
logical and behavioural traits (Lovei and Sunderland 1996).
Carabids and their species assemblages are frequently
used as environmental indicators or ecological indicators
(in the sense of McGeoch 1998). Several studies deal with
their indicator value in climate change (Ashworth 1996;
Gobbi et al. 2006), habitat fragmentation (De Vries 1994,
Spence et al. 1996; Davies and Margules 1998; Magura
et al. 2001; Niemela 2001; Lovei et al. 2006), or evaluate
the influence of stressing factors of management practices
in grasslands (Blake et al. 1996; Rushton et al. 1989), in
croplands (Pfiffner and Niggli 1996; Kromp 1999) and in
logging forests in temperate (Szyszko 1983) or boreal
zones (Niemela et al. 1993). Carabids can, together with
other taxa, be used as biodiversity indicators (Rainio and
Niemela 2003).
Nearly all the studies quoted here are contemporary or
short-time comparisons of sampling plots or sites; they are
snapshots of differences, during short periods of one or two
to three years. For carabids very few exceptions are known;
the best are perhaps the studies of Desender and Turin
(1989) and Desender et al. (1994a) of the decline of open
land species in the fauna of four western European coun-
tries. Nevertheless, exact numbers of species losses related
to specific habitats are very difficult to obtain, probably
because studies of such areas are rarely focused over an
entire century or many decades. Brandmayr and Algieri
P. Brandmayr � R. Pizzolotto (&) � T. Zetto
Dipartimento di Ecologia, Universita della Calabria,
87036 Rende, CS, Italy
e-mail: [email protected]
G. Colombetta
Via Elia 2, Trieste, Italy
123
J Insect Conserv (2009) 13:231–243
DOI 10.1007/s10841-008-9161-6

(2001) found that about 86% of the Italian populations of
the chlaeniine wetland ground beetle Epomis circum-
scriptus were extinct at the end of the twentieth century.
In the city of Trieste (Italy, Friuli-Venezia Giulia
Region), an old Mediterranean suburban forest, long-called
Farneto (or Boschetto) by Triestine citizens, was protected
for centuries because of its importance as a wood reserve
for the garrisons and for supplying natural resources (fungi,
bark, medicinal plants) to inhabitants, its vegetation and
biomass structure was left unchanged until 1943–44, when
the necessities of the Second World War forced the
inhabitants to exploit its extensive wood reserves. After
clear-cutting during the War, part of the hill slope was
artificially reforested, while some other parts recovered
mostly by natural regeneration.
The municipality of Trieste recently guaranteed the
protection of the forest against urbanisation. In 1996 the
Nature-2000 team of the Department of Biology of Trieste
University proposed the creation of a SCI (Site of Com-
munity Interest), and now the Farneto Forest is scheduled
as ‘‘Site of National Interest’’ in the frame of the parallel
protection Programme ‘‘Bioitaly’’, with the SIN-number:
IT3342014.
The Farneto wood area represents a type of synthesis in
space and time of these anthropization processes. In past
times the area was on the outskirts of Trieste, until it was
surrounded progressively by the city but without losing the
ecological function of a semi-natural area connecting the
more natural karstic plateau.
The present research aims to appraise in a historical
perspective the effect of anthropogenic activities and
urbanization on the degradation of the carabid species
assemblages and their ability to recover. The Farneto rep-
resents a unique possibility for this purpose because the site
has been visited by entomologists since the year 1818,
when the naturalists Hoppe and Hornschuch collected a
new lebiine subspecies, Lebia fulvicollis thoracica. Later,
between 1890 and 1930, the forest was the preferred col-
lecting target of a very active group of Triestine
entomologists and the results for carabid beetles were
organized by Giuseppe Muller (1926) in a detailed topo-
graphic catalogue.
The following questions were focused on in detail. (1)
Are carabid species assemblages a valuable record of the
changes observed in a suburban forest during the past
century? (2) Are the species assemblages censused after the
secondary succession (1945–19804) similar to the ancient
ones, or has a new ‘‘equilibrium’’ been reached? (3) How
many carabid species have been lost (become locally
extinct) during the past century? (4) Does human distur-
bance still exist and what should be the policies against it?
(5) Is the study of the ‘‘in situ extinction’’ an useful
approach?
Materials and methods
Study area
The climate of the study area in the city of Trieste belongs
to the submediterranean type. The mean annual tempera-
ture of the Farneto is 1.5�C below the mean of the city
centre (14�C), and is estimated at around 12.5�C at a
medium altitude of 150 m. The precipitation is about
1,000 mm/year.
The Farneto (Figs. 1, 2) covers an area of 238 ha on a
sandstone hill slope (Eocene ‘‘Flysch’’), located at the
eastern periphery of the city. The area extends like a finger
from about the centre of the city (altitude 60 m, and no
more than 1.5 km from the Adriatic Sea) to the border of
the Karst plateau behind Trieste (240 m asl). Its length is
about three kilometres and its maximum width is 700 m.
The slope exposure is mainly N or NNE. The southern
border is marked by the edges of the Chiadino and Cac-
ciatore hills. Two small, more or less permanent streams
run from the hilltops towards the Torrente Grande, which
borders the northern edge of the forest.
The original vegetation was a mixed forest dominated
by Quercus petraea, with a dense Sesleria autumnalis herb
layer (Associazione Sportiva 1994; Poldini 1989). Before
clear-cut logging in 1944, all the layers of the forest
structure were entirely native, with a mean diameter of
major trees (mostly oaks) around 1–1.5 m. Other tree
species were Q. cerris, Castanea sativa, Acer campestre,
Carpinus betulus, Ostrya carpinifolia and, along the
streams, Salix alba and Populus alba. After 1945, in the
post-war period, the area was partly reforested with aus-
trian black pine (Pinus nigra) and with cypress (Cupressus
sempervirens and C. arizonica), but a large part of the
forest was invaded by Robinia pseudoacacia. In 1984 parts
of the forest area were still occupied by remnants of the
open land created after the Second World War, but about
70% of the surface had been reconquered by native trees,
especially oaks, Fraxinus ornus and Ostrya carpinifolia.
This recently cleared open land was frequently crossed by
burnt areas that had retarded the ecological succession.
Some very small patches of humid vegetation with
Phragmites australis were also present.
After 1975, probably due to climate change and conse-
quently milder winters, the understorey shows an
increasing expansion of evergreen bushes, mainly Laurus
nobilis; other native Mediterranean elements such as Rus-
cus aculeatus were always found in the forest.
As a result, about 95% of the original Farneto is now
covered by trees 25–50 years old. At the time of our
1983–84 investigation, artificial pine plantations or natu-
rally-regenerated coppice forest accounted for about 85%
of the entire surface of the Farneto, and trees of the
232 J Insect Conserv (2009) 13:231–243
123

allochtonous species Robinia pseudacacia comprised
more than the 20% of the forest canopy.In 1984, the area
was threatened by pollution of brooks and the main
stream, the ‘‘Torrente Grande’’, and by illegal land use,
e.g. abandoning waste and rubbish.
At the eastern boundary of the Farneto, the open land is
represented by a true old pasture. Open ground was man-
aged through sheep and cattle grazing until the 1950s, when
this type of land use began to decline strongly in the whole
Trieste Province. This old open land still exists despite
Fig. 1 The study area of the Farneto wood hill (13� 210 3000 Est, 45� 380 5000 North)
Fig. 2 The Farneto forest seen
from the edge of the Karst
Plateau near Trieste
J Insect Conserv (2009) 13:231–243 233
123
heavy urbanisation of the surroundings in the last 20 years.
This site was also used for the historical comparison.
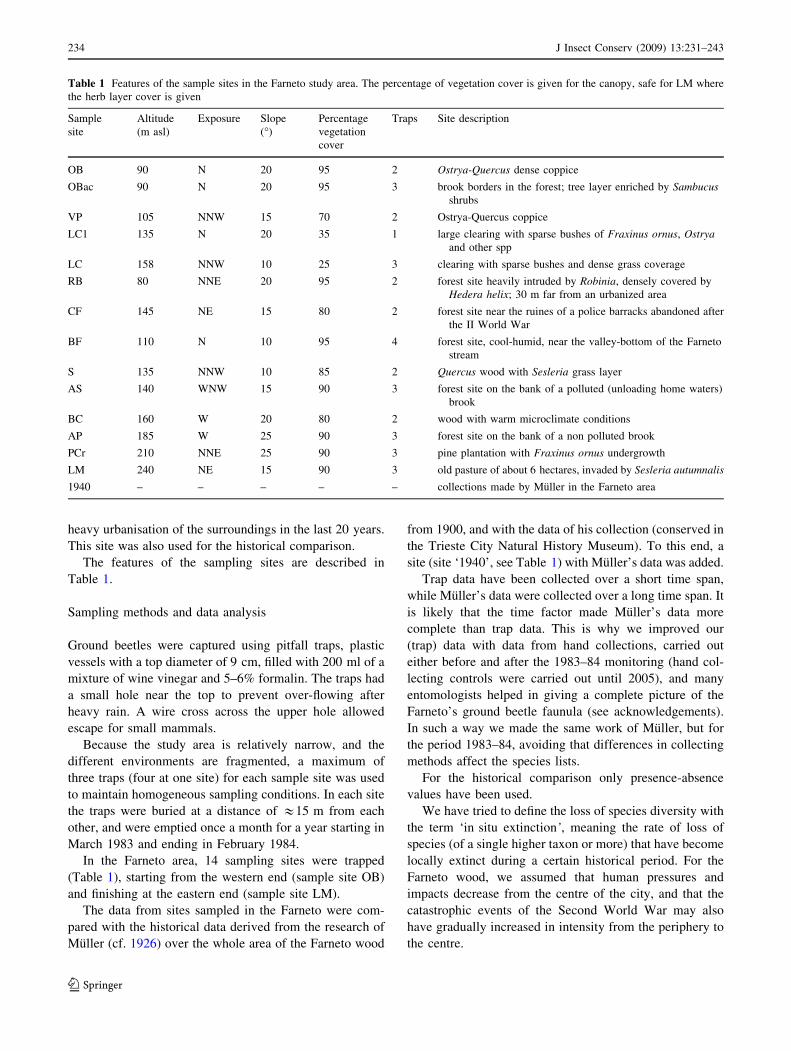
The features of the sampling sites are described in
Table 1.
Sampling methods and data analysis
Ground beetles were captured using pitfall traps, plastic
vessels with a top diameter of 9 cm, filled with 200 ml of a
mixture of wine vinegar and 5–6% formalin. The traps had
a small hole near the top to prevent over-flowing after
heavy rain. A wire cross across the upper hole allowed
escape for small mammals.
Because the study area is relatively narrow, and the
different environments are fragmented, a maximum of
three traps (four at one site) for each sample site was used
to maintain homogeneous sampling conditions. In each site
the traps were buried at a distance of &15 m from each
other, and were emptied once a month for a year starting in
March 1983 and ending in February 1984.
In the Farneto area, 14 sampling sites were trapped
(Table 1), starting from the western end (sample site OB)
and finishing at the eastern end (sample site LM).
The data from sites sampled in the Farneto were com-
pared with the historical data derived from the research of
Muller (cf. 1926) over the whole area of the Farneto wood
from 1900, and with the data of his collection (conserved in
the Trieste City Natural History Museum). To this end, a
site (site ‘1940’, see Table 1) with Muller’s data was added.
Trap data have been collected over a short time span,
while Muller’s data were collected over a long time span. It
is likely that the time factor made Muller’s data more
complete than trap data. This is why we improved our
(trap) data with data from hand collections, carried out
either before and after the 1983–84 monitoring (hand col-
lecting controls were carried out until 2005), and many
entomologists helped in giving a complete picture of the
Farneto’s ground beetle faunula (see acknowledgements).
In such a way we made the same work of Muller, but for
the period 1983–84, avoiding that differences in collecting
methods affect the species lists.
For the historical comparison only presence-absence
values have been used.
We have tried to define the loss of species diversity with
the term ‘in situ extinction’, meaning the rate of loss of
species (of a single higher taxon or more) that have become
locally extinct during a certain historical period. For the
Farneto wood, we assumed that human pressures and
impacts decrease from the centre of the city, and that the
catastrophic events of the Second World War may also
have gradually increased in intensity from the periphery to
the centre.
Table 1 Features of the sample sites in the Farneto study area. The percentage of vegetation cover is given for the canopy, safe for LM where
the herb layer cover is given
Sample
site
Altitude
(m asl)
Exposure Slope
(�)
Percentage
vegetation
cover
Traps Site description
OB 90 N 20 95 2 Ostrya-Quercus dense coppice
OBac 90 N 20 95 3 brook borders in the forest; tree layer enriched by Sambucusshrubs
VP 105 NNW 15 70 2 Ostrya-Quercus coppice
LC1 135 N 20 35 1 large clearing with sparse bushes of Fraxinus ornus, Ostryaand other spp
LC 158 NNW 10 25 3 clearing with sparse bushes and dense grass coverage
RB 80 NNE 20 95 2 forest site heavily intruded by Robinia, densely covered by
Hedera helix; 30 m far from an urbanized area
CF 145 NE 15 80 2 forest site near the ruines of a police barracks abandoned after
the II World War
BF 110 N 10 95 4 forest site, cool-humid, near the valley-bottom of the Farneto
stream
S 135 NNW 10 85 2 Quercus wood with Sesleria grass layer
AS 140 WNW 15 90 3 forest site on the bank of a polluted (unloading home waters)
brook
BC 160 W 20 80 2 wood with warm microclimate conditions
AP 185 W 25 90 3 forest site on the bank of a non polluted brook
PCr 210 NNE 25 90 3 pine plantation with Fraxinus ornus undergrowth
LM 240 NE 15 90 3 old pasture of about 6 hectares, invaded by Sesleria autumnalis
1940 – – – – – collections made by Muller in the Farneto area
234 J Insect Conserv (2009) 13:231–243
123

A species accumulation curve and richness estimation
after 50 randomizations were computed on trap data to give
an estimate of catching efficiency. The estimators were:
Shannon diversity index, Incidence-based Coverage Esti-
mator (ICE) and Coleman estimator (Colwell 2005). ‘ICE
is the sum of the probabilities of encounter for the species
observed, taking into account species present but not
observed’ (Colwell 2005). To compute ICE, a hypothetical
maximum number of species is estimated on the basis of a
threshold (defined by the researcher, i.e. in our case the
species captured only three times) corresponding to the
least frequent species. ICE gives the theoretical species
richness including species not discovered in any sample.
The Coleman estimator estimates species richness on the
basis of all species actually discovered (Colwell 2005).
Data were analysed by cluster analysis (correlation
coefficient and minimum variance, also known as Ward’s
distance), and principal components analysis (StatSoft Inc.
2001).
To evaluate the nature of the changes that have had the
most influence in modifying the carabid assemblages of the
Farneto, the habitat preference of all the 66 taxa found in
the Farneto was determined from the literature or from
interpretation of available datasets. Each of the 66 species
in Table 2 was assigned to one of the following main
ecological classes: forest, pastures (and meadows), river-
side (or wetlands), using the criterion of Thiele (1977), see
also Maelfait et al. (1994); Telfer and Eversham(1996),
and Cuesta et al. (2006), which takes into account the peak
of abundance. Data were ordered as follows: species
recorded before 1940, species collected during the study in
1983–84, new findings in pitfall traps in 1983–84 (probably
not found by other entomologists who were pure hand
collectors) and species found before but not recorded in
1983–84 (defined as: lost).
Results
Catching efficiency
A species accumulation curve and richness estimation after
50 randomizations of pitfall traps data are outlined in
Fig. 3. The observed species curve (S obs.) does not flatten
towards the value of 27 (i.e. the number of species captured
by pitfal traps), but it does not seem to be very steep there.
Perhaps, by visual estimation of the shape of the curve, it
could be hypothesized that the asymptote lies between 30
and 32. Figures predicted by the Coleman curve are very
similar to the observed ones. ICE predicts a higher species
number, but this estimator is also based on hypothetical
species, so it is not only bound to actual ecosystem con-
ditions. More interestingly is the fact that ICE flattens the
more sites are added, and so does the Shannon index,
suggesting that the number of observed species is near the
actual maximum.
The species assemblages 1983–84
In the 14 sites of the Farneto forest, a complex of 27
ground beetle taxa was collected in 35 pit-fall traps
(Table 2). Data collected by hand were also included,
reaching a total of 70 species (i.e. four species have been
added: Paranchus albipes, a riverside species; Trechus
subnotatus, a rare riverside dweller; Clivina collaris, riv-
erside; Laemostenus algerinus, forest dweller).
Among the species found 19 are forest dwellers, 14 are
forest specialists, which are more represented in the eastern
(and upper) side of the forest. The most common forest
specialist is Carabus catenulatus, a typical earthworm
predator, but snail predators are also widespread (Carabus
caelatus, coriaceus, especially in the larval stages). Five
forest species are endemics of south-eastern Europe, and
the rare hygrophilous Trechus subnotatus seems to have
survived the clear-cutting shock of 1944 in at least one
stand. The canopy predators Calosoma sycophanta and
C. inquisitor are both active in many places.
Stands VP, S, CF and RB were at the time degraded or still
partly open land, and this suggests that at least some forest
species recolonized the young succession stages from the
eastern, Karst-facing, side of the area. Figure 4 gives the
activity density of the three most important forest specialists
in order of distance from the city centre. At least two of them
(Abax ater and Carabus caelatus) show a possible coloni-
sation direction from east to west; a small population of the
first could have survived near the Botanical Garden, where
some older trees were left standing.
Data classification and ordination
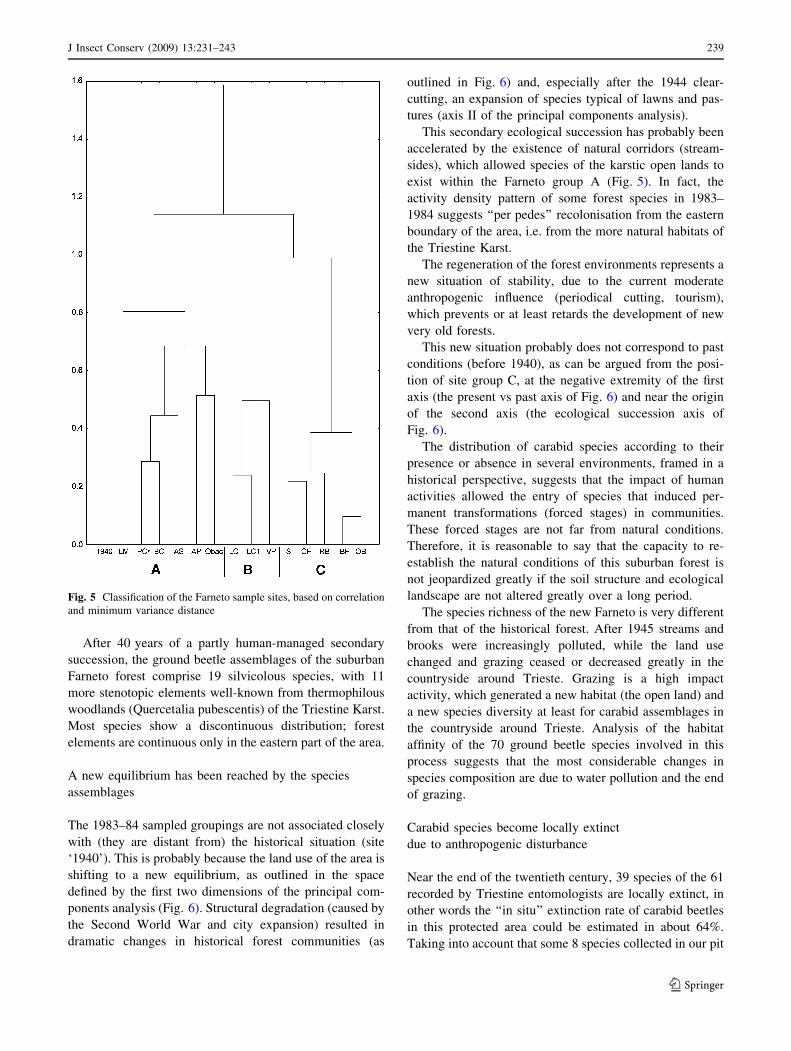
The analysis of the urbanisation phenomenon (the sites of
the Farneto wood area) (Table 2), shows a pattern (Fig. 5)
where site ‘1940’ (based on Muller’s collections) is sepa-
rate from the other groups, probably because it has 39
species not found in the present investigation.
The species distribution in the Farneto wood allows
identification of three groups of sites: group A composed
mainly of sample sites along small brooks, including the
only true pasture of the Farneto wood area; group B
composed of samples from clearings, and the related group
C of the forest samples.
The historical-dynamic relationships of the carabid
beetles species groupings can be deduced partly from
ordination through principal components analysis of the
sites (Fig. 6), where the first two dimensions explain 62%
of the variation.
J Insect Conserv (2009) 13:231–243 235
123

Table 2 Carabid beetle presence in the Farneto study area. Species grouped according to habitat preference (F = forest, forest specialists in
bold, P = pasture, R = riverside). Nomenclature follows Vigna (1993). Sites ordered as in Fig. 5
1940 LM PCr BC AS AP Obac LC LC1 VP S CF RB BF OB
F Carabus coriaceus L. 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
F Carabus catenulatus Scopoli 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
F Harpalus atratus Latreille 1 1 1 1 1 1
F Carabus caelatus Fabr. 1 1 1 1 1 1 1
F Abax ater (Villers) 1 1 1 1 1 1 1 1 1 1
F Myas chalybaeus (Palliardi) 1 1 1 1
F Laemostenus algerinus (Gory) 1
F Calosoma sycophanta (L.) 1 1 1 1
F Steropus melas (Creutzer) 1 1 1 1 1 1 1 1
F Notiophilus rufipes Curtis 1 1 1 1
F Synuchus vivalis (Illiger) 1 1
F Leistus rufomarginatus (Duft.) 1 1 1 1 1
F Calosoma inquisitor (L.) 1 1
F Trechus quadristristus (Schrank) 1
F Molops ovipennis Chaudoir 1
F Laemostenus elongatus (Dej.) 1
F Laemostenus venustus (Dej.) 1
F Philorhizus quadrisignatus (Dej.) 1
F Anillus florentinus Dieck 1
P Calathus glabricollis Dej. 1 1
P Harpalus dimidiatus (Rossi) 1 1 1 1
P Harpalus rubripes (Duft.) 1 1 1
P Anchomenus dorsalis (Pontopp.) 1 1
P Brachinus crepitans (L.) 1 1
P Calathus cinctus Motsch. 1
P Harpalus sulphuripes Germar 1 1
P Harpalus distinguendus (Duft.) 1
P Amara aequestris (Duft.) 1
P Ophonus puncticeps Stephens 1 1
P Scybalicus oblongiusculus (Dej.) 1
P Poecilus koyi (Germar) 1 1
P Lebia cruxminor (L.) 1
P Poecilus cupreus (L.) 1
P Ophonus brevicollis (Serville) 1
P Lamprias fulvicollis (Fabr.) 1
P Amara anthobia Villa 1
P Callistus lunatus (Fabr.) 1
P Amara eurynota (Panzer) 1
P Olisthopus glabricollis (Germar) 1
P Microlestes fulvibasis (Reitt.) 1
P Bradycellus verbasci (Duft.) 1
P Ocydromus tetracolus (Say) 1
P Diachromus germanus (L.) 1
P Harpalus flavicornis Dej. 1
P Amara familiaris Duft. 1
P Harpalus anxius (Duft.) 1
P Lamprias cyanocephala (L.) 1
P Microlestes fissuralis (Reitt.) 1
236 J Insect Conserv (2009) 13:231–243
123
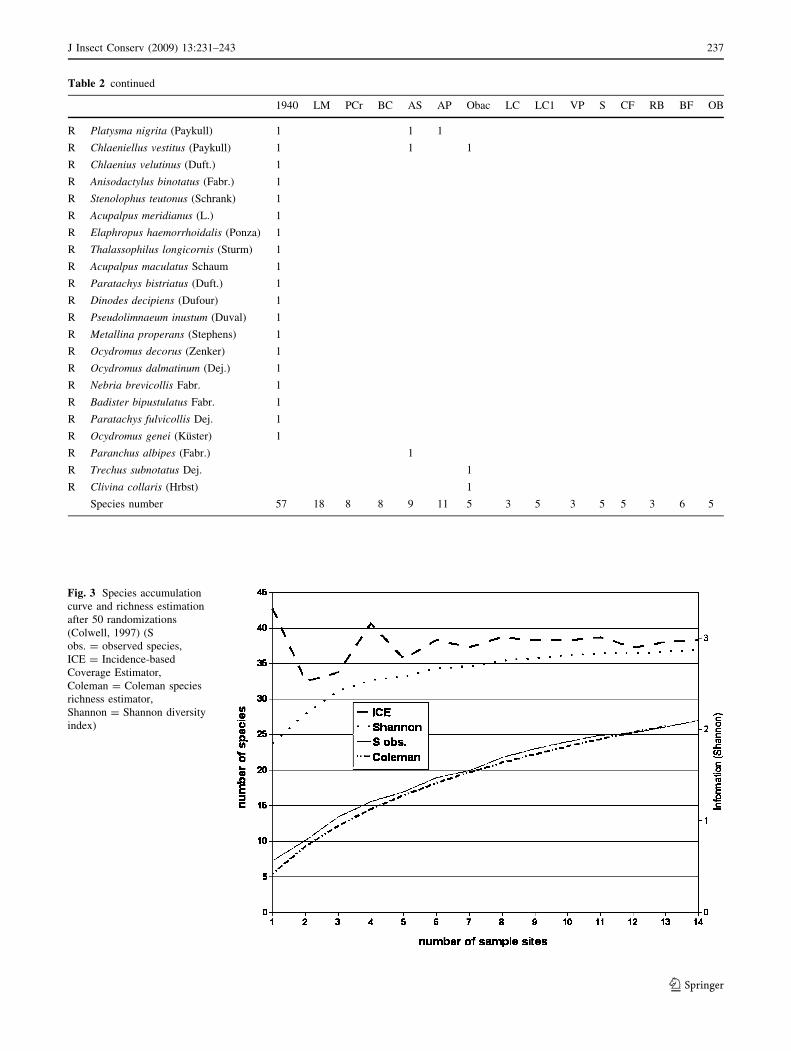
Table 2 continued
1940 LM PCr BC AS AP Obac LC LC1 VP S CF RB BF OB
R Platysma nigrita (Paykull) 1 1 1
R Chlaeniellus vestitus (Paykull) 1 1 1
R Chlaenius velutinus (Duft.) 1
R Anisodactylus binotatus (Fabr.) 1
R Stenolophus teutonus (Schrank) 1
R Acupalpus meridianus (L.) 1
R Elaphropus haemorrhoidalis (Ponza) 1
R Thalassophilus longicornis (Sturm) 1
R Acupalpus maculatus Schaum 1
R Paratachys bistriatus (Duft.) 1
R Dinodes decipiens (Dufour) 1
R Pseudolimnaeum inustum (Duval) 1
R Metallina properans (Stephens) 1
R Ocydromus decorus (Zenker) 1
R Ocydromus dalmatinum (Dej.) 1
R Nebria brevicollis Fabr. 1
R Badister bipustulatus Fabr. 1
R Paratachys fulvicollis Dej. 1
R Ocydromus genei (Kuster) 1
R Paranchus albipes (Fabr.) 1
R Trechus subnotatus Dej. 1
R Clivina collaris (Hrbst) 1
Species number 57 18 8 8 9 11 5 3 5 3 5 5 3 6 5
Fig. 3 Species accumulation
curve and richness estimation
after 50 randomizations
(Colwell, 1997) (S
obs. = observed species,
ICE = Incidence-based
Coverage Estimator,
Coleman = Coleman species
richness estimator,
Shannon = Shannon diversity
index)
J Insect Conserv (2009) 13:231–243 237
123

Along the first axis (Fig. 6), site ‘1940’ is the only site
with positive correlation. Site LM has the second highest
score, and the forest samples lie at the negative extremity. On
the basis of the position of sites LM and ‘1940’ vs. the other
sites, it is possible to interpret the first axis as the axis of the
variation between the historical and present situations.
Along the second axis, sites ‘1940’ and LM are at the
two extremities, while the forest sites of group C (Fig. 6)
are concentrated at the axis origin. In this case, the second
axis represents the variation associated with ecological
succession induced by deforestation.
According to the ecological meaning of the axes, as
explained before, the space defined by the principal com-
ponent analysis is therefore comprised of four quadrants
(Fig. 6): quadrant I, the sub-space of the historical natural
conditions, quadrant II, probably associated with historical
degradation of the environment (a reasonable hypothesis,
because no data are available), quadrant III, associated with
conditions of recovery from degradation, and quadrant IV,
associated with present conditions.
Bearing in mind that the meaning of quadrant II was
inferred by its relative position, an ideal historical track was
drawn on the quadrants of Fig. 6, to outline the relationships
between heavy anthropization and secondary ecological
succession.
Species survival and habitat affinity
Figure 7 shows the change of species on the basis of habitat
affinity. For each group, the habitat affinity is reported as
species number. The highest proportion of species surviving
in 1983 was forest species, whereas both pasture and
waterside components exhibited outstanding losses. Water
pollution is probably the cause of the disappearance of the
most hygrophilic or streamside carabids, which were
reduced from 22 to five species (23%). The pasture and
meadow components were also affected greatly by reduc-
tion: from 24 to 12 (50%). Forest dwellers survived the best:
14 of 15 (93%). It is likely that the new species recorded in
1983 were living in the old Farneto, so the correct reduction
is probably from 19 to 14 (74%). Thus, the lost species (39)
pertain mostly to the waterside (44%) and pasture dwellers
(44%), and minimally to the forest elements (12%). Fur-
thermore, no new waterside species were collected.
Discussion
The answers posed in the introduction follow as subchapters.
Carabid species assemblages are a valuable record
of the changes observed in a suburban forest during
the past century
In the ecological landscape of the Trieste city surroundings
where farming and forestry have modified the natural
conditions, characteristic groupings of carabid species have
developed, which can be distinguished from those of
environments less affected by human activity (Fig. 5).
Fig. 4 Annual Activity Density
(individuals/trap in the standard
period of 10 days. Ordinate). of
three abundant forest species in
the Farneto wood, ordered by
increasing distance from the
centre of Trieste (abscissa,
where the distance is between
brackets). The forest links to the
Karst plateau at its eastern
border through a pasture of
*6–7 hectares (Site LM, right
in the graph). For graphical
reasons the values for
C. caelatus have been weighted
by 10
238 J Insect Conserv (2009) 13:231–243
123
After 40 years of a partly human-managed secondary
succession, the ground beetle assemblages of the suburban
Farneto forest comprise 19 silvicolous species, with 11
more stenotopic elements well-known from thermophilous
woodlands (Quercetalia pubescentis) of the Triestine Karst.
Most species show a discontinuous distribution; forest
elements are continuous only in the eastern part of the area.
A new equilibrium has been reached by the species
assemblages
The 1983–84 sampled groupings are not associated closely
with (they are distant from) the historical situation (site
‘1940’). This is probably because the land use of the area is
shifting to a new equilibrium, as outlined in the space
defined by the first two dimensions of the principal com-
ponents analysis (Fig. 6). Structural degradation (caused by
the Second World War and city expansion) resulted in
dramatic changes in historical forest communities (as
outlined in Fig. 6) and, especially after the 1944 clear-
cutting, an expansion of species typical of lawns and pas-
tures (axis II of the principal components analysis).
This secondary ecological succession has probably been
accelerated by the existence of natural corridors (stream-
sides), which allowed species of the karstic open lands to
exist within the Farneto group A (Fig. 5). In fact, the
activity density pattern of some forest species in 1983–
1984 suggests ‘‘per pedes’’ recolonisation from the eastern
boundary of the area, i.e. from the more natural habitats of
the Triestine Karst.
The regeneration of the forest environments represents a
new situation of stability, due to the current moderate
anthropogenic influence (periodical cutting, tourism),
which prevents or at least retards the development of new
very old forests.
This new situation probably does not correspond to past
conditions (before 1940), as can be argued from the posi-
tion of site group C, at the negative extremity of the first
axis (the present vs past axis of Fig. 6) and near the origin
of the second axis (the ecological succession axis of
Fig. 6).
The distribution of carabid species according to their
presence or absence in several environments, framed in a
historical perspective, suggests that the impact of human
activities allowed the entry of species that induced per-
manent transformations (forced stages) in communities.
These forced stages are not far from natural conditions.
Therefore, it is reasonable to say that the capacity to re-
establish the natural conditions of this suburban forest is
not jeopardized greatly if the soil structure and ecological
landscape are not altered greatly over a long period.
The species richness of the new Farneto is very different
from that of the historical forest. After 1945 streams and
brooks were increasingly polluted, while the land use
changed and grazing ceased or decreased greatly in the
countryside around Trieste. Grazing is a high impact
activity, which generated a new habitat (the open land) and
a new species diversity at least for carabid assemblages in
the countryside around Trieste. Analysis of the habitat
affinity of the 70 ground beetle species involved in this
process suggests that the most considerable changes in
species composition are due to water pollution and the end
of grazing.
Carabid species become locally extinct
due to anthropogenic disturbance
Near the end of the twentieth century, 39 species of the 61
recorded by Triestine entomologists are locally extinct, in
other words the ‘‘in situ’’ extinction rate of carabid beetles
in this protected area could be estimated in about 64%.
Taking into account that some 8 species collected in our pit
Fig. 5 Classification of the Farneto sample sites, based on correlation
and minimum variance distance
J Insect Conserv (2009) 13:231–243 239
123
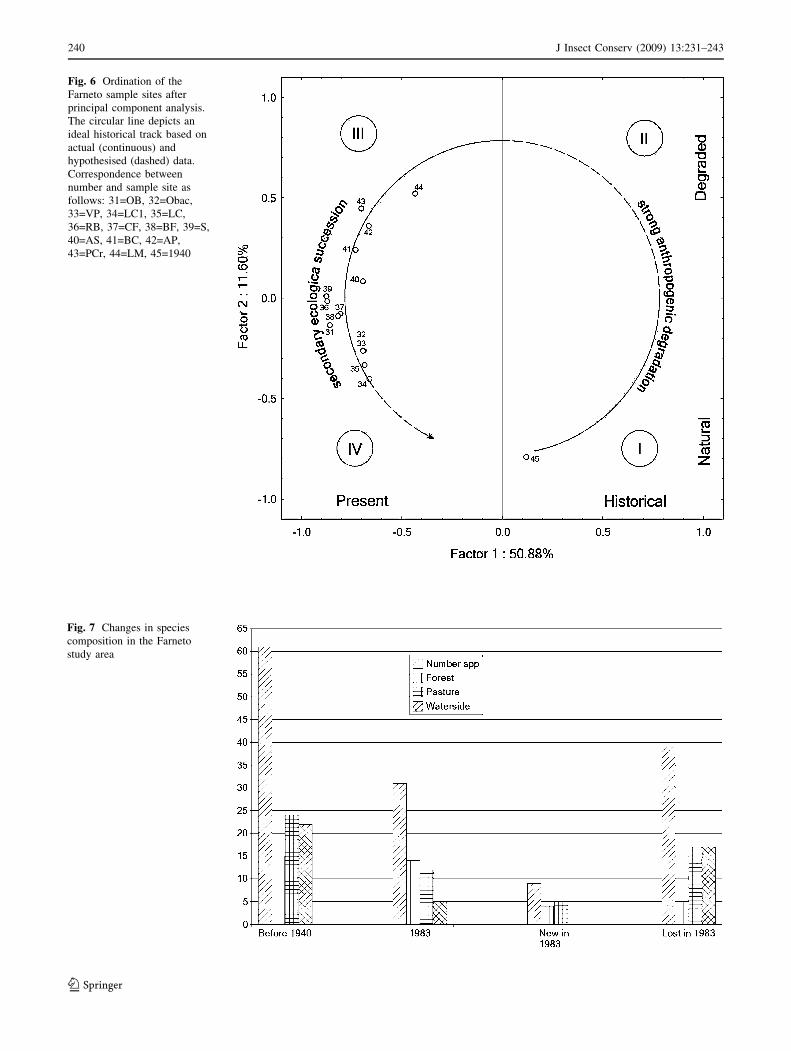
Fig. 6 Ordination of the
Farneto sample sites after
principal component analysis.
The circular line depicts an
ideal historical track based on
actual (continuous) and
hypothesised (dashed) data.
Correspondence between
number and sample site as
follows: 31=OB, 32=Obac,
33=VP, 34=LC1, 35=LC,
36=RB, 37=CF, 38=BF, 39=S,
40=AS, 41=BC, 42=AP,
43=PCr, 44=LM, 45=1940
Fig. 7 Changes in species
composition in the Farneto
study area
240 J Insect Conserv (2009) 13:231–243
123

fall traps may have escaped the hand collecting of previous
entomologists (especially Molops ovipennis, Myas cha-
lybaeus and Carabus caelatus, which are difficult to catch
without traps), a theoretical minimum of extinction may be
39/(61 + 8), approximately 57%. In both cases, more than
half of the carabid species of the Farneto forest disappeared
in the second part of the past century, and this alarming fact
makes theories about species diversity in disturbance gra-
dients oversimplified. It is well-known that the ‘‘stepping
stones of global extinctions are local extinctions’’ (Lovei
2001), and at least four or five of the vanished species (e.g.
Chlaenius velutinus, Pseudolimnaeum inustum) have today
(2006) only one live population in the whole Trieste
Province (personal communications, see acknowledge-
ments). The ultimate cause of extinction of most taxa
(90%) in the United States is habitat destruction or deg-
radation (Wilcove et al. 1998).
Human disturbance is still affecting the study area
Human disturbance in the forest continues and both forest
and open land species seem to avoid the urbanized borders
of the Farneto (Fig. 4). Consequently, in our opinion, the
following measures could stop local species extinction and
favour the original species richness: (1) restoration of
surface water quality and of waterside habitats for inver-
tebrates and vertebrates (the only amphibian surviving in
the forest is the fire salamander Salamandra salamandra);
(2) high-trunk (old-growth) oriented management, at least
in the core area of the forest, and soil restoration especially
through log decay; (3) maintenance of some clearings and
open tracks by horse grazing and/or by mowing.
Some of the forest specialists of the area need conser-
vation measures. Despite appearing less affected by
changes and having the ability to recolonize the original
habitat, some more sensitive species seem locally extinct,
e.g. Procerus gigas, the largest carabid of Europe (which
had a small live population until 1975 in the Botanical
Garden), Anillus florentinus (a deep soil dwelling bem-
bidiine species, highly affected by soil dryness),
Laemostenus venustus (a dendrophilic species connected to
very old chestnut and oak trees, see Pizzolotto et al. 1991),
and others of less importance. The sphodrine carabid L.
venustus is surely extinct in the whole Province of Trieste
and further investigations could clarify if L. venustus may
be considered a dendrophilic indicator of Mediterranean
forests, today well preserved in southern Italy.
Because secondary ecological succession is still going
on, it seems merely theoretical to predict whether the
maximum species number will be equal to, less than or
above the recorded 70. It is reasonable, however, to predict
that after several patches of the original landscape (oak
woods) have re-established, Farneto wood will reach
maximum species diversity, if human disturbance is kept at
a moderate level (tourism, recreation, small clearings).
‘In situ extinction’ as a tool for monitoring the natural
species turnover
The study of the in situ extinction, accompained by the
natural species turnover, is a useful approach for checking
the efficiency of conservation policies in protected areas, or
for evaluating new areas for the status of protection sites.
Species listings remain the most important attribute for site
evaluation (Usher 1986); moreover methods should be
quantitative, including relative abundances (Samways
1995). Many well-known National Natural Reserves, such
as the Monks Wood Reserve in Cambridgeshire, provide
excellent examples of species losses due to the cessation of
traditional management (e.g. butterfly species extinctions
in Peterken 1991). Monitoring carabid species disappear-
ance and/or recolonisation as in the Farneto could be an
optimal way of monitoring to check recovery after clear-
cutting.
The in situ extinction approach is not dependent on the
boundary span of the study area, when a historical census
of reliable biological indicators, such as carabids, has been
carried out, over at least a twenty-year time period. In this
sense, its application ranges from the rehabilitation moni-
toring of contaminated sites to the effects of climate
change on alpine environments, for example.
Conclusion
Further research is needed to elucidate the biological fea-
tures of carabid species (see Pizzolotto 1994) and
parameters of carabid species assemblages (see Penev
1996), to allow comparisons within different areas at large
scales (Dennis and Ruggiero 1996) and develop sound
management practices for suburban forests (the adaptive
management of Hillborn et al. 1995).
As a general remark, it seems more useful to recognize
and reconstruct the causes of disturbance, and relate them
to species assemblages and habitat preference, rather than
solely to the number of species. Carabid beetle studies are a
valid help toward this aim, because changes of carabid
species number and type can be related to human distur-
bance (Niemela et al. 2002; Venn et al. 2003; Ishitani et al.
2003). Furthermore we found that such disturbance can
lead to local species extinction, which could be a serious
biodiversity impoverishment when involving endemic
species.
Acknowledgements The authors are very indebted to several Tri-
estine entomologists, who generously helped with advice and opened
J Insect Conserv (2009) 13:231–243 241
123

their own collections. The late Mr. Carlo Posarini donated the only
specimen of Laemostenus algerinus captured in Trieste after World
War II, and helped in monitoring Procerus gigas for two decades
(1965–1985). Detailed information about the insect diversity of the
Farneto forest was obtained by Dr. Giuseppe Muller’s latest pupils,
Dr. Bruno Millo and Dr. Tiziano De Monte, and by our unforgettable
friend Dr. Guido Calligaris. The Museum of Natural History of the
city of Trieste allowed careful examinations of the two main collec-
tions and of ‘‘inserenda’’.
References
Ashworth AC (1996) The response of arctic carabids (Coleoptera) to
climate change based on the fossil record of the Quaternary
Period. Ann Zool Fenn 33:125–131
Associazione Sportiva del Corpo Forestale (1994) Il bosco Farneto -
Storia, natura e sentieri del ‘‘Boschetto’’ di Trieste. Spring,
Trieste
Blake S, Foster GN, Fischer GEJ, Ligertwood GL (1996) Effects of
management practices on the ground beetle faunas of newly
established wildflower meadows in southern Scotland. Ann Zool
Fenn 33:139–147
Brandmayr P, Algieri MC (2001) Habitat affinities of chlaeniine
species (Coleoptera, Carabidae) in Calabria and the status of
Epomis circumscriptus, evaluated by the ‘‘Cronogeonemie’’
software. In: Brandmayr P, Loevei G, Zetto T, Casale A, Vigna
A (eds) Natural history and applied ecology of Carabid Beetles.
Pensoft, Sofia
Brandmayr P, Cagnin M, Mingozzi A, Pizzolotto R (1996) Map of
zoocoenoses and evaluation for the Menta river dam in
Aspromonte (Calabria, Italy). Z Okol Natur 5:15–28
Colwell RK (2005) EstimateS: Statistical estimation of species
richness and shared species from samples. Version 7.5. User’s
Guide and application published at: http://purl.oclc.org/estimates
Cuesta D, Taboada A, Calvo L, Salgado JM (2006) A preliminary
investigation of ground beetle (Coleoptera: Carabidae) assem-
blages and vegetation community structure in Calluna vulgarisheatlands in NW Spain. Entomol Fenn 11:241–252
Davies KF, Margules CR (1998) Effects of habitat fragmentation on
carabid beetles: experimental evidence. J Animal Ecol 67:460–
471
De Vries HH (1994) Size of habitat and presence of ground beetle
species. In: Desender K, Dufrene M, Loreau M, Luff ML,
Maelfait J-P (eds) Carabid Beetles, ecology and evolution.
Kluwer Academic Publishers, Dordrecht
Dennis JG, Ruggiero MA (1996) Biodiversity inventory: building an
inventory at scales from local to global. In: Szaro RC, Johnston
DW (eds) Biodiversity in managed landscapes: theory and
practice. Oxford University Press, New York
Desender K, Turin H (1989) Loss of habitats and changes in the
composition of the ground and tiger beetle fauna in four west
European countries since 1950 (Coleoptera: Carabidae, Cicin-
delidae). Biol Conserv 48:277–294
Desender K, Dufrene M, Maelfait JP (1994a) Long-term dynamics of
carabid beetles in Belgium: a preliminary analysis on the
influence of changing climate and land use by means of a
database covering more than a century. In: Desender K, Dufrene
M, Loreau M, Luff ML, Maelfait J-P (eds) Carabid Beetles,
ecology and evolution. Kluwer Academic Publishers, Dordrecht
Desender K, Dufrene M, Loreau M, Luff ML, Maelfait J-P (eds)
(1994b) Carabid Beetles, ecology and evolution. Kluwer Aca-
demic Publishers, Dordrecht
Gobbi M, De Bernardi F, Pelfini M, Rossaro B, Brandmayr P (2006)
Epigean Arthropod Succession along a 154-year Glacier
Foreland Chronosequence in the Forni Valley (Central Italian
Alps). Arct Antarct Alp Res 38:357–362
Hillborn R, Walters CJ, Ludwig D (1995) Sustainable exploitation of
renewable resources. Annu Rev Ecol Syst 26:45–67
Ishitani M, Kotze DJ, Niemela J (2003) Changes in carabid beetle
assemblages across an urban-rural gradient in Japan. Ecography
26:481–489
Kromp B (1999) Carabid beetles in sustainable agriculture: a review
on pest control efficacy, cultivation impacts and enhancement.
Agric Ecosyst Environ 74:187–228
Lovei G (2001) Extinctions, modern examples of. In: Encyclopedia of
Biodiversity, 2. Academic Press
Lovei G, Sunderland KD (1996) Ecology and behavior of ground
beetles (Coleoptera: Carabidae). Annu Rev Entomol 41:231–256
Lovei G, Magura T, Tothmeresz B, Kodobocz V (2006) The influence
of matrix and edges on species richness patterns of ground
beetles (Coleoptera: Carabidae) in habitat islands. Glob Ecol
Biogeogr 15:283–289
Luff ML (1996) Use of carabids as environmental indicators in
grasslands and cereals. Ann Zool Fenn 35:185–195
Maelfait J-P, Desender K, Dufrene M (1994) Carabid beetles and
nature conservation research in Belgium: a review. In: Desender
k, Dufrene M, Loreau M, Luff ML,Maelfait J-P (eds) Carabid
Beetles, ecology and evolution. Kluwer Academic Publisher,
Dordrecht, The Netherlands
Magura T, Kodobocz V, Tothmeresz B (2001) Effects of habitat
fragmentation on carabids in forest patches. J Biogeogr 28:129–
138
McGeoch MA (1998) The selection, testing, and application of
terrestrial insects as bioindicators. Biol Rev 73:181–201
Muller G (1926) I Coleotteri della Venezia Giulia. Catalogo ragionato
- Parte I: Adephaga. Mosettigs, Trieste
Niemela J (1996) From systematics to conservation - carabidologists
do it all. Ann Zool Fenn 33:1–4
Niemela J (2001) Carabid beetles (Coleoptera: Carabidae) and habitat
fragmentation: a review. Eur J Entomol 98:127–132
Niemela J, Langor D, Spence JR (1993) Effects of Clear-Cut
Harvesting on Boreal Ground-Beetle Assemblages (Coleoptera:
Carabidae) in Western Canada. Conserv Biol 7:551–561
Niemela J. Kotze D J, Venn S, Penev L, Stoyanov I, Spence J, Hartley
D, de Oca EM (2002) Carabid beetle assemblages (Coleoptera,
Carabidae) across urban-rural gradients: an international com-
parison. Landsc Ecol 17:387–401
Pearson DL, Cassola F (1992) World-wide species richness patterns
of tiger beetles (Coleoptera: Cicindelidae): indicator taxon for
biodiversity and conservation studies. Conserv Biol 6:376–391
Penev L (1996) Large scale variation in carabid assemblages, with
special reference to local fauna concept. Ann Zool Fenn 33:49–64
Peterken GF (1991) Ecological issues in the management of
woodland nature reserves. In: Spellerberg BS, Goldsmith FB,
Morris MG (eds) The scientific management of temperate
communities for conservation. Blackwell Scientific Puclications,
Oxford
Pfiffner L, Niggli U (1996) Effects of bio-dynamic, organic and
conventional farming on ground beetles (Col. Carabidae) and
other epigaeic arthropods in winter wheat. Biol Agric Hortic
12:353–364
Pizzolotto R (1994) Soil arthropods for faunal indices in assessing
changes in natural value resulting from human disturbance. In:
Boyle T, Boyle CEB (eds) Biodiversity, temperate ecosystems
and global change. Sprtinger Verlag, Berlin, Heidelberg, New
York
Pizzolotto R, Mingozzi A, Cagnin M, Tripepi S, Aloise G, Barbieri A,
Scalzo A, Brandmayr P (1991) Effetti della ceduazione periodica
del castagneto sulle comunita di Coleotteri Carabidi, rettili,
uccelli e micromammiferi terricoli. S.IT.E. Atti 12:449–453
242 J Insect Conserv (2009) 13:231–243
123

Poldini L (1989) La vegetazione del Carso isontino e triestino. Lint
Publisher, Trieste
Rainio J, Niemela J (2003) Ground beetles (Coleoptera: Carabidae) as
bioindicators. Biodivers Conserv 12:487–506
Rushton SP, Luff ML, Eyre MD (1989) Effect of pasture improve-
ment and management on the ground beetle and spider
communitites of upland grasslands. J Appl Ecol 26:489–503
Samways MJ (1995) Insect conservation biology. conservation
biology series. Chapman & Hall, London
Spence JR, Langor DW, Niemela J, Carcamo HA, Currie CC (1996)
Northern forestry and ground beetles: the case for concern about
old-growth species. Ann Zool Fenn 33:173–184
StatSoft (2001) STATISTICA (data analysis software system),
version 6. www.statsoft.com
Stork NE (ed) (1990) The role of Ground Beetles in Ecological and
Environmental Studies. Intercept, Andover-Hampshire
Szyszko J (1983) State of Carabidae (Col.) fauna in fresh pine forest
and tentative valorisation of this environment. SGGW-AG
Monographs No. 28. Warsaw Agricultural University Press,
Warsaw, Poland
Telfer MG, Eversham BC (1996) Ecology and conservation of
heatland Carabidae in eastern England. Ann Zool Fenn 33:133–
138
Thiele HU (1977) Carabid Beetles in their environments. Springer,
Berlin, Heidelberg, New York
Usher MB (1986) Wildlife conservation evaluation: attributes, criteria
and values. In: Usher MB (ed) Wildlife conservation evaluation.
Chapman & Hall, London
Venn SJ, Kotze DJ, Niemela J (2003) Urbanization effects on carabid
diversity in boreal forests. Eur J Entomol 100:73–80
Vigna A (1993) Coleoptera Archostemata, Adephaga 1 (Carabidae).
Checklist delle specie della fauna italiana. Calderini, Bologna
Wilcove DS, Rothstein D, Dubow J, Phillips A, Losos E (1998)
Quantifying threats to imperiled species in the United States.
Bioscience 48:607–615
J Insect Conserv (2009) 13:231–243 243
123
Copyright © 2022 FDOKUMEN




















