
Ultrastructure of the antennal sensorial appendage of larvae of Ophonus ardosiacus (Lutshnik, 1922)...
13
Zoologischer Anzeiger 247 (2008) 209–221 Ultrastructure of the antennal sensorial appendage of larvae of Ophonus ardosiacus (Lutshnik, 1922) (Coleoptera, Carabidae) and possible correlations between size and shape and the larval feeding habits A. Giglio a, , P. Brandmayr a , E.A. Ferrero b , P.G. Giulianini b , E. Perrotta a , F.F. Talarico a , T. Zetto Brandmayr a a Dipartimento di Ecologia, Universita` della Calabria, I- 87036 Arcavacata di Rende (Cs), Italia b Dipartimento di Biologia, Universita` di Trieste, via Giorgieri 7, I-34127 Trieste, Italia Received 26 June 2007; received in revised form 28 November 2007; accepted 3 December 2007 Corresponding Editor: M. Schmitt Abstract The antennal sensory appendage morphology was studied in 19 species of carabid beetle larvae using scanning electron microscopy. Shape and size of the organ has been re-examined in selected taxa and are compared to figure out possible correlations between antennal sensory structures and larval morphoecological types. Morphometric analysis shows no correlation between antennal sensory appendage dimension and the four most common behavioural types. The ultrastructure of this sensory organ is described for larvae of Ophonus ardosiacus and the possible function is discussed. Olfactory function has been indicated as the most probable. The high number of chemosensory neurons of this multiporous organ suggests a possible ability of fine odour discrimination related to prey choice or host location. r 2008 Elsevier GmbH. All rights reserved. Keywords: Larval ground beetles; Olfactory chemoreceptor; Antennal sensory organ; Hyaline vesicle; Larval life styles 1. Introduction Carabid beetles are a family of Coleoptera widely spread in all terrestrial habitat where they are important predators. Previous studies have been suggested that there are correlations between body form and habit (e.g. feeding, locomotory, burrowing, flying) in Carabid beetle adults (Forsythe 1991; Evans 1994). In carabid larvae, it is well known that some larval features (mandible form, general body form, urogomphi mor- phology and feeding behaviour) are correlated with the main selective pressures of the habitat. Zetto Brandmayr et al. (1998) have correlated morphological features, behavioural traits and possible selective pressures by grouping known species into nine main behavioural types (1. soil pore explorers, 2. surface runners, 3. surface walkers, 4. sand diggers, 5. seed feeders, 6. burrow trappers, 7. C-shape Harpalines, 8. parasitoids and predatory symbionts, 9. trichome disc larvae). However, very fragmentary information concerns sen- sory equipment of head appendages of larval stages related to the behaviour, feeding habits and use of space. At present, knowledge about larval sensorial equipment and its possible function regards the rostral processes of Notiophilus biguttatus (Fabricius, 1779) (Altner and Bauer 1982), the adhesive mouthparts of Loricera ARTICLE IN PRESS www.elsevier.de/jcz 0044-5231/$ - see front matter r 2008 Elsevier GmbH. All rights reserved. doi:10.1016/j.jcz.2007.12.001 Corresponding author. Tel.: +390984492982; fax: +390984492981. E-mail address: [email protected] (A. Giglio).
Transcript of Ultrastructure of the antennal sensorial appendage of larvae of Ophonus ardosiacus (Lutshnik, 1922)...

ARTICLE IN PRESS
0044-5231/$ - se
doi:10.1016/j.jc
�Correspondfax: +39098449
E-mail addr
Zoologischer Anzeiger 247 (2008) 209–221
www.elsevier.de/jcz
Ultrastructure of the antennal sensorial appendage of larvae of Ophonusardosiacus (Lutshnik, 1922) (Coleoptera, Carabidae) and possible
correlations between size and shape and the larval feeding habits
A. Giglioa,�, P. Brandmayra, E.A. Ferrerob, P.G. Giulianinib, E. Perrottaa,F.F. Talaricoa, T. Zetto Brandmayra
aDipartimento di Ecologia, Universita della Calabria, I- 87036 Arcavacata di Rende (Cs), ItaliabDipartimento di Biologia, Universita di Trieste, via Giorgieri 7, I-34127 Trieste, Italia
Received 26 June 2007; received in revised form 28 November 2007; accepted 3 December 2007Corresponding Editor: M. Schmitt
Abstract
The antennal sensory appendage morphology was studied in 19 species of carabid beetle larvae using scanningelectron microscopy. Shape and size of the organ has been re-examined in selected taxa and are compared to figure outpossible correlations between antennal sensory structures and larval morphoecological types. Morphometric analysisshows no correlation between antennal sensory appendage dimension and the four most common behavioural types.The ultrastructure of this sensory organ is described for larvae of Ophonus ardosiacus and the possible function isdiscussed. Olfactory function has been indicated as the most probable. The high number of chemosensory neurons ofthis multiporous organ suggests a possible ability of fine odour discrimination related to prey choice or host location.r 2008 Elsevier GmbH. All rights reserved.
Keywords: Larval ground beetles; Olfactory chemoreceptor; Antennal sensory organ; Hyaline vesicle; Larval life styles
1. Introduction
Carabid beetles are a family of Coleoptera widelyspread in all terrestrial habitat where they are importantpredators. Previous studies have been suggested thatthere are correlations between body form and habit (e.g.feeding, locomotory, burrowing, flying) in Carabidbeetle adults (Forsythe 1991; Evans 1994). In carabidlarvae, it is well known that some larval features(mandible form, general body form, urogomphi mor-phology and feeding behaviour) are correlated with the
e front matter r 2008 Elsevier GmbH. All rights reserved.
z.2007.12.001
ing author. Tel.: +390984492982;
2981.
ess: [email protected] (A. Giglio).
main selective pressures of the habitat. Zetto Brandmayret al. (1998) have correlated morphological features,behavioural traits and possible selective pressures bygrouping known species into nine main behaviouraltypes (1. soil pore explorers, 2. surface runners, 3.surface walkers, 4. sand diggers, 5. seed feeders, 6.burrow trappers, 7. C-shape Harpalines, 8. parasitoidsand predatory symbionts, 9. trichome disc larvae).However, very fragmentary information concerns sen-sory equipment of head appendages of larval stagesrelated to the behaviour, feeding habits and use of space.At present, knowledge about larval sensorial equipmentand its possible function regards the rostral processes ofNotiophilus biguttatus (Fabricius, 1779) (Altner andBauer 1982), the adhesive mouthparts of Loricera

ARTICLE IN PRESSA. Giglio et al. / Zoologischer Anzeiger 247 (2008) 209–221210
pilicornis (Fabricius, 1775) (Bauer and Kredler 1988),the highly modified antennae in Siagona europaea
(Dejean, 1826) (Zetto Brandmayr et al. 2007), themaxillary palps in Nebria (Latreille, 1802) (Spenceand Sutcliffe 1982), the general sensory equipment ofthe ant’s brood-eater larva of Graphipterus serrator
(Forsk(al, 1775) (Zetto Brandmayr et al. 1993) andPaussini larvae (Luna de Carvalho 1991; Bousquet 1986;Di Giulio et al. 2003). Recently, labial and maxillarypalps of 22 larval species belonging to six of the nineproposed behavioural types have been described com-paratively (Giglio et al. 2003). Little informations occuron antennal sensory structure. As in Adephagan larvae,carabid beetles antennae are four-segmented with asensory appendage on the third article, whose form andsize show an interesting variation among species and/orhigher taxa.
The present work is a morphometric comparativestudy of this antennal organ in larvae of some European
Table 1. List of carabid species whose larvae were studied
Behavioural
types
Tribe Genus Species
Surface walkers Carabini Carabus lefebvrei (D
Surface runners Nebriini Leistus *fulvibarbis
Nebria kratteri (D
*germari (H
*dahli (Stu
Notiophilini Notiophilus *rufipes (C
Surface walkers Licinini Licinus silphoides (
Soil pore
explorers
Trechini Trechus *quadristria
1781)
Typhlotrechus *bilimeki (S
Pterostichini Laemostenus
(Antisphodrus)
*obtusus (C
Calathus *glabricolli
*fuscipes (G
*mollis (Ma
*erratus (S
Poecilus cupreus (Li
Seed feeders Harpalini Ophonus *ardosiacus
*sabulicola
Harpalus
(Pseudophonus)
rufipes (De
Harpalus
(Harpalus)
*tardus (Pa
An asterisk (*) marks the specimens preserved in our collection.
species of Carabidae in relation to their morphoecologi-cal type. In addition, in order to assess its functional role,ultrastructural analysis of the antennal sensory appen-dage is provided for Ophonus ardosiacus (Lutshnik, 1922)larvae.
2. Materials and methods
We examined 19 species of 13 genera belonging toseven tribes and four behavioural types (Table 1). Theywere chosen to cover a wide range of taxonomic unitsand to include species with different life history andfeeding preferences. Most samples were collected in thefield and preserved in 70% ethanol (Zetto Brandmayrcollection, Department of Ecology, University ofCalabria); only O. ardosiacus and C. lefebvrei larvaewere obtained ‘‘ex ovipositione’’ in laboratory and
Capture site Larval
stages
ejean, 1826) Lab-rearing spring 2006 III
(Dejean, 1826) Trionto (CS) 22.01.95 III
ejean, 1831) Serra San Bruno (VV) 22.12.97 I/III
eer, 1837) Dolomiti 29.06.80 III
rm, 1815) Slovenia, Slavnik Mount
26.11.74
III
urtis, 1829) Trionto (CS) 14.06.94 III
Rossi, 1790) Trionto (CS) 19.05.93 III
tus (Schrank, Friuli, pastures (Magredi)
21.04.79
II
turm, 1847) Grotta delle Torri (TS)
04.07.83
III
haudoir, 1861) Alpi Liguri 17.09.64 I/III
s (Dejean, 1828) Stena Mount, Carso Triestino
15.12.77
II
oeze, 1777) Sicilia, Nebrodi Mounts
25.04.81
I
rsham, 1802) Sicilia, Peloritani Mounts
30.05.82
III
ahlberg, 1827) Friuli, pastures (Magredi)
25.10.78
II/III
nne, 1758) Tarsia (CS), 14.06.00 I
(Lutshnik, 1922) Lab-rearing 76-77 I/III
(Panzer, 1796) Paolo Mounts Faenza,
04.03.77
II/III
geer, 1774) Tarsia (CS) 14.10.99 III
nzer, 1797) Trionto (CS) 28.06.94 I

ARTICLE IN PRESSA. Giglio et al. / Zoologischer Anzeiger 247 (2008) 209–221 211
reared to the third stage under controlled temperatureand photoperiod. According to the method ofZetto Brandmayr (1976, 1978), the larval stages ofO. ardosiacus were reared in laboratory and fed onthe Umbelliferae seeds, mostly Daucus carota. While,C. lefebvrei larval stages were fed on snails (Helix sp.).
For scanning electron microscopy (SEM), antennaewere cut and doubly fixed in 3% glutaraldehyde and 1%osmium tetroxide in 0.1M cacodylate buffer pH 7.2–7.3,dehydrated in a graded ethanol series and critical point-dried. After dehydration, they are mounted on stubs,gold coated and examined in a Cambridge Stereoscan100 scanning electron microscope.
For morphometric measurements, a reduced sample(Table 2) of larvae from 11 species belonging to differentbehavioural types was selected. Photographs were takenwith a stereoscope (Zeiss Stemi SV 11 Apo) andacquired by Matrox PC-VCR software (Windows2000). Measurements were taken using Sigma ScanPro 5 Software (SPSSs Inc.). Differences in morpho-metric measurements were tested using non-parametrictest (U Mann–Whitney test) with Statistica 6.0 software(Statsoft Italia SrL).
Ultrastructural investigations were performed onO. ardosiacus. For light microscopy (LM) observations,the samples were fixed in Bouin and stained bymethylene blue vital (0.5% stock solution diluted 1:100in insect saline before use) to evidence bipolar neurons.The solution was injected by means of a fine tipped glasspipette into the basal antennal joint. Diffusion andsuccessful staining occurred within 1 h. Photographs ofthe preparations were immediately taken, to avoidfading, under a Leitz Dialux phase contrast LM.Permanent mounts were obtained by fixation in 12%W/V ammonium molybdate (Alexandrowicz 1954) and
Table 2. Measurements (mean7standard deviation) of the antenn
width
Behavioural types Species Head wid
Surface walkers Carabus lefebvrei 3.20270.
Surface runners Leistus fulvibarbis 0.99470.
Nebria kratteri 2.20870.
Nebria dahli 1.68270.
Nebria germari 1.24670.
Notiophilus rufipes 0.81670.
Soil pore explorers Laemostenus obtusus 1.16470.
Calathus erratus 1.2470.
Calathus mollis 0.91670.
Seed feeders Ophonus ardosiacus 2.22270.
Ophonus sabulicola 2.14670.
Species are investigated in relation to their life style. Means of 5 specimens
subsequent dehydration in ethanol and clearing inxylene. For transmission electron microscopy (TEM),single antennae were removed from the head andimmediately immersed in 3% glutaraldehyde in 0.1Mcacodylate buffer with 5% sucrose pH 7.2–7.3. Theywere then rinsed in cacodylate buffer, post-fixed in 1%osmium tetroxide in 0.1M cacodylate buffer for 1 h at4 1C and rinsed in the same buffer. Dehydration in agraded ethanol series was followed by embedding inepon-araldite (Electron Microscopy Sciences, FortWashington, PA). Thin sections, cut with an UltratomeIIILKB with glass and diamond knives, were subse-quently stained with uranyl acetate and lead citrate andviewed with a Philips EM208 electron microscope at80 kV.
3. Results
3.1. Morphology and morphometry of antennal
sensory appendage
In carabid beetle larvae the antennae are locatedanterolaterally on the head and articulated in fourantennomeres (Fig. 1A). The third antennomere shows acupuliform sensory appendage (sensorium) varying inshape and size in predator (Figs. 2A–I and 3A–E, G)and seed feeder (Figs. 3F, H, I) larvae.
Table 2 summarizes antennal sensory appendagemeasurements for five individuals (third larval stage)of each species. For comparison, height and width of thesensory appendage are related to head width. Thediagram (Fig. 4) shows that the antennal sensoryappendage of all the species is longer than wide
al sensory appendage base width and height related to head
th (mm) Antennal sensory appendage (mm)
Base w/head w Height/head w
051 0.01770.001 0.02570.001
032 0.02370.004 0.03970.007
138 0.01170.002 0.01470.003
042 0.0170.002 0.01670.001
073 0.01570.002 0.02470.002
069 0.02470.002 0.03470.007
099 0.01170.001 0.01970.002
061 0.01170.001 0.01570.002
135 0.01470.004 0.00470.003
108 0.01570.001 0.01370.001
083 0.01270.002 0.01570.002
(III stage) per species are given.

ARTICLE IN PRESS
an
as
m
hw
nA
B
C
Fig. 1. Ophonus ardosiacus larva: I stage. (A) Head scanning electron micrograph, dorsal view; an: antennae, as: antennal sensory
appendage, hw: head width, m: mandible. (B) Antennal sensory appendage scanning electron micrograph. (C) Antennal sensory
appendage stained by methylene blue vital (light microscopy micrograph); n: neurons. Scale bars: 200mm (A), 20 mm (B, C).
A. Giglio et al. / Zoologischer Anzeiger 247 (2008) 209–221212
except in C. mollis and O. ardosiacus. The sensoryappendage is smaller in C. lefebvrei than in L. fulvibarbis
(Mann–Whitney test: n1 ¼ n2 ¼ 5, Z ¼ 2.61, Po0.01);while in N. rufipes it is significantly wider than in C.
lefebvrei (Mann–Whitney test: n1 ¼ n2 ¼ 5, Z ¼ 2.61,Po0.01) but there is no significant difference about theheight. The width and the length of the sensorium arenot significantly different in Nebria kratteri, Calathus
erratus, Ophonus sabulicola and N. dahli. The sensoryappendage is significantly longer in L. (A.) obtusus thanin C. erratus (Mann–Whitney test: n1 ¼ n2 ¼ 5,Z ¼ �1.98, Po0.01), N. kratteri and O. sabulicola
(Mann–Whitney test: n1 ¼ n2 ¼ 5, Z ¼ �2.19,Po0.01). In two species belonging to the same genus,N. kratteri and N. germari, there are significantdifferences in width (Mann–Whitney test: n1 ¼ n2 ¼ 5,Z ¼ �2.19, Po0.02) and height (Mann–Whitney test:n1 ¼ n2 ¼ 5, Z ¼ �2.61, Po0.01) of the sensoryappendage; while O. sabulicola has a sensory appendagesignificantly wider than O. ardosiacus (Mann–Whitneytest: n1 ¼ n2 ¼ 5, Z ¼ 2.19, Po0.02).
3.2. Ultrastructure of O. ardosiacus sensory
appendage
O. ardosiacus (Fig. 1A) antennal sensory appendage isa dome-shaped organ located in the third antennalsegment, measuring 29.071.2 (N ¼ 5) mm in height and34.471.5 (N ¼ 5) mm in diameter in third larval stage.Observations by SEM (Fig. 1B) and LM (Fig. 1C) revealthat the antennal sensory appendage consists of acupuliform part implanted on the thickened socketalong its basal circumference, so defining the subcupularspace in which clustered cell bodies are located.
Ultrathin sections (Figs. 5 and 6A, B) show a sensoryorgan covered by cuticle, showing a subcuticular spaceand consisting of 20 morphofunctional units. A doubletof sensory cells surrounded by three accessory ensheath-ing cells forms each unit.
The cuticle (Figs. 5 and 6A) of the third antennomereis 800 nm in thickness with: an epicuticular layer, anexocuticle and a multilayered endocuticle. Above thesocket, the cuticle consists only of the epi- and
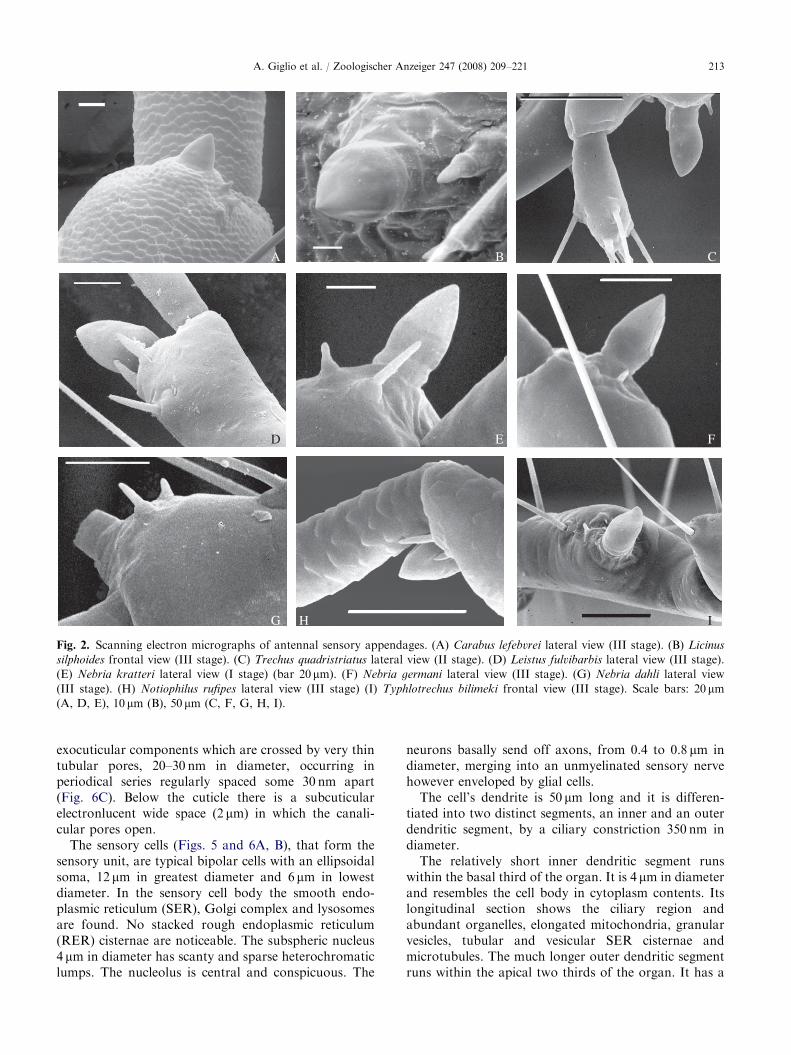
ARTICLE IN PRESS
A B C
D E F
G H I
Fig. 2. Scanning electron micrographs of antennal sensory appendages. (A) Carabus lefebvrei lateral view (III stage). (B) Licinus
silphoides frontal view (III stage). (C) Trechus quadristriatus lateral view (II stage). (D) Leistus fulvibarbis lateral view (III stage).
(E) Nebria kratteri lateral view (I stage) (bar 20mm). (F) Nebria germani lateral view (III stage). (G) Nebria dahli lateral view
(III stage). (H) Notiophilus rufipes lateral view (III stage) (I) Typhlotrechus bilimeki frontal view (III stage). Scale bars: 20mm(A, D, E), 10 mm (B), 50 mm (C, F, G, H, I).
A. Giglio et al. / Zoologischer Anzeiger 247 (2008) 209–221 213
exocuticular components which are crossed by very thintubular pores, 20–30 nm in diameter, occurring inperiodical series regularly spaced some 30 nm apart(Fig. 6C). Below the cuticle there is a subcuticularelectronlucent wide space (2 mm) in which the canali-cular pores open.
The sensory cells (Figs. 5 and 6A, B), that form thesensory unit, are typical bipolar cells with an ellipsoidalsoma, 12 mm in greatest diameter and 6 mm in lowestdiameter. In the sensory cell body the smooth endo-plasmic reticulum (SER), Golgi complex and lysosomesare found. No stacked rough endoplasmic reticulum(RER) cisternae are noticeable. The subspheric nucleus4 mm in diameter has scanty and sparse heterochromaticlumps. The nucleolus is central and conspicuous. The
neurons basally send off axons, from 0.4 to 0.8 mm indiameter, merging into an unmyelinated sensory nervehowever enveloped by glial cells.
The cell’s dendrite is 50 mm long and it is differen-tiated into two distinct segments, an inner and an outerdendritic segment, by a ciliary constriction 350 nm indiameter.
The relatively short inner dendritic segment runswithin the basal third of the organ. It is 4 mm in diameterand resembles the cell body in cytoplasm contents. Itslongitudinal section shows the ciliary region andabundant organelles, elongated mitochondria, granularvesicles, tubular and vesicular SER cisternae andmicrotubules. The much longer outer dendritic segmentruns within the apical two thirds of the organ. It has a
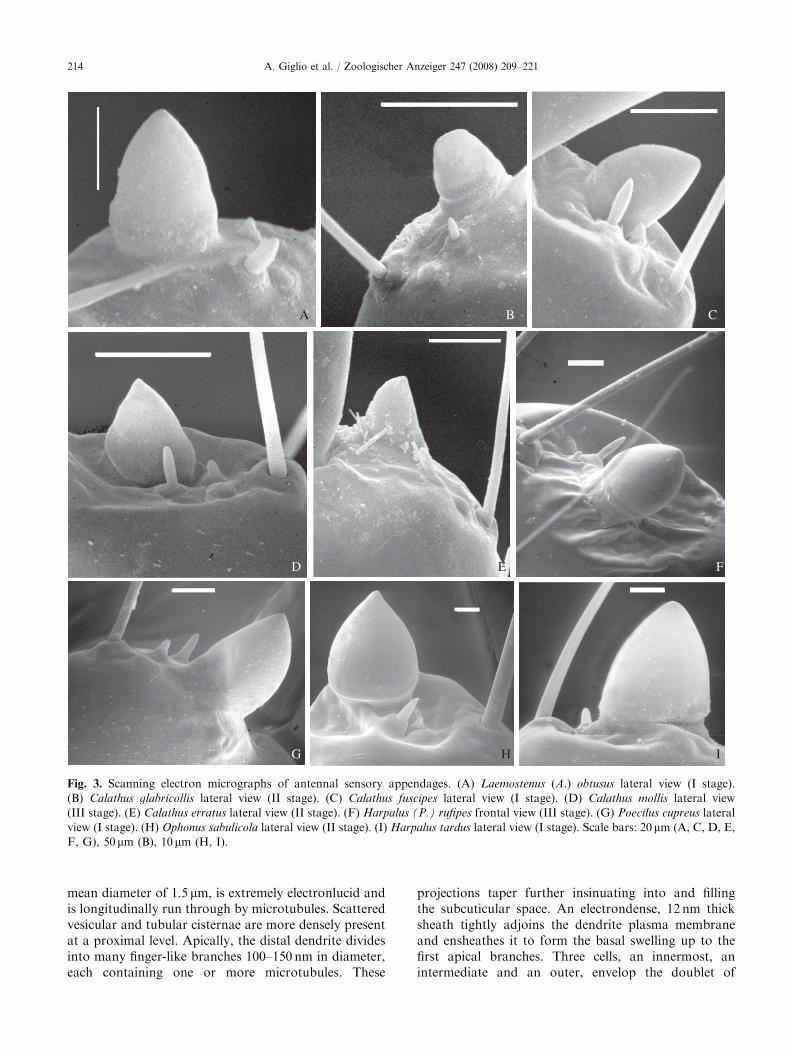
ARTICLE IN PRESS
A B C
D E F
G H I
Fig. 3. Scanning electron micrographs of antennal sensory appendages. (A) Laemostenus (A.) obtusus lateral view (I stage).
(B) Calathus glabricollis lateral view (II stage). (C) Calathus fuscipes lateral view (I stage). (D) Calathus mollis lateral view
(III stage). (E) Calathus erratus lateral view (II stage). (F) Harpalus (P.) rufipes frontal view (III stage). (G) Poecilus cupreus lateral
view (I stage). (H) Ophonus sabulicola lateral view (II stage). (I) Harpalus tardus lateral view (I stage). Scale bars: 20mm (A, C, D, E,
F, G), 50mm (B), 10mm (H, I).
A. Giglio et al. / Zoologischer Anzeiger 247 (2008) 209–221214
mean diameter of 1.5 mm, is extremely electronlucid andis longitudinally run through by microtubules. Scatteredvesicular and tubular cisternae are more densely presentat a proximal level. Apically, the distal dendrite dividesinto many finger-like branches 100–150 nm in diameter,each containing one or more microtubules. These
projections taper further insinuating into and fillingthe subcuticular space. An electrondense, 12 nm thicksheath tightly adjoins the dendrite plasma membraneand ensheathes it to form the basal swelling up to thefirst apical branches. Three cells, an innermost, anintermediate and an outer, envelop the doublet of

ARTICLE IN PRESS
Fig. 4. Measurements of the antennal sensory appendage base width and height related to head width.
A. Giglio et al. / Zoologischer Anzeiger 247 (2008) 209–221 215
sensory neurons (Fig. 5). The innermost or thechogencell has its cell body at the same level of the sensory celldoublets. It is associated with each dendrite doublet andenvelops it up to the proximal end of the outer dendriticsegment by means of a cytoplasmic layer 0.5 mm thick onaverage. This cytoplasmic lamina adheres to the apicalinner dendritic segment by means of septate junctions.Distally, many mitochondria and cytoskeletal fibres11 nm in diameter arranged in longitudinal bundles of300 nm thickness, are present. Between dendrite andaccessory cell, a space or primary receptor lymph cavityoccurs. Proximally, the sinuous finger-like projections ofthe accessory cell envelop the outer dendritic segmentsurface and make its sheath. The cytoplasmic projec-tions of the intermediate and outer enveloping cells,interspersed among the sensory doublets, are typicallyelectrondense and include compact mitochondria, lyso-somes, tubular SER cisternae, microtubules and glyco-gen granules. The intercellular space becomes nearlysealed in places by close apposition of plasma mem-branes which form junctional complexes with gap- andintermediate junctions and septate desmosomes. Distallythere are wide lacunar spaces, so called secondaryreceptor lymph cavities. In these, enveloping celllaminar, 40–60 nm thick, projections parallel andstacked make sinuous and whorled profiles. Moreapically, below the cupola, the projections of theenveloping cells swell into bulbous endings surroundedby many laminar layers. In their cytoplasm manymultilaminar bodies with membranes whorls 2 mm indiameter, become apparent. These myelin-like bodiesmay represent evidence of the high membrane turnoverto be related to the large surface area exposed to thereceptor lymph space.
4. Discussion
The antennal sensory appendage of polyphagancoleopteran larvae was investigated by several authors(Scott and Zacharuk 1971; Roppel et al. 1972; Corbiere-Tichane 1971, 1973; Ryan and Behan 1973; Behan andRyan 1978; Bloom et al. 1981). Some of them alsodescribed the ultrastructure and function of a senseorgan located on the second segment: a modified orcompound sensillum basiconicum in structure andolfactory in function. Corbiere (1969) reported itspresence in Bathysciinae (Coleoptera: Cholevidae), andcalled it ‘‘membranous lobe’’ (lobe membraneux);Zacharuk (1962) in Elateridae named it ‘‘antennalsensory appendage’’; Bloom et al. (1981) and Behanand Ryan (1978) identified it as a ‘‘sensillum placo-deum’’ in Tenebrionidae. In carabid beetles thisstructure, firstly called ‘‘hyaline vesicle’’, has beendescribed by van Emden (1942) and Jeannel (1941,1942). It is a small appendage located in a lateralmembranous area of the third antennal segment. Thedifference in location between Polyphaga and Adephagais related to the different number of antennomeres. Thisvariation was examined more in detail by Beutel et al.(1999), who stated the existence of four antennomeres inneuropterids (e.g. Sialidae), in the basic Archostematacoleopterans, in Adephaga and some scarabeids. Inpolyphagan larvae the majority are three-segmented, butin many families the number is reduced to one or two(Lawrence 1991). In the somewhat aberrant, aquatic andminiaturized Myxophaga two-segmented antennaehave been found. Minelli (2004) re-examines in detailthe number of antennal joints in Coleoptera and gives tothe sensorial appendage a sort of ‘‘key-position’’ in

ARTICLE IN PRESS
ax
in
ic
gc
s1
d
od
e
ep
en
ex
so
ss
lc
lc
tp
cuoc
od
idid
s2
s1
s2
Fig. 5. Schematical drawing summarising the ultrastructural features of Ophonus ardosiacus antennal sensory appendage. Black
rectangle highlights a detail of sensory cell unit. ax: axons cu: cuticle; d: dendrite of sensory cell; e: epidermal cell; en: endocuticle; ep:
epicuticle; ex: exocuticle; gc: glial cell; ic: innermost cell; id: inner dentritic segment; in: intercalary cell; lc: lymph cavity; oc: outer
cell; od: outer dentritic segment; s1, s2: sensory cell; so: socket; ss: subcuticular space; tp: tubular pore.
A. Giglio et al. / Zoologischer Anzeiger 247 (2008) 209–221216
determining their basic number: three, according toLawrence’s idea.
The sensorial appendage varies also in size and lengthfrom a moderately long appendage to a more or lessshort coniform or cupuliform sensorium (Beutel et al.1999), and it is absent in Sialidae, some hydroadephaganfamilies (Gyrinidae, Noteridae) and many polyphagangroups. Also in Carabidae a trend from a four- to athree-jointed antenna is seen comparing ‘‘normal’’ taxawith the autapomorphic status of Helluomorphoides
larvae (antacoria of antennomeres II–III disappears,Bousquet 1987) and with the fully three-jointed antennaof Anthiini derived by fusion (see Table 3).
About ultrastructure, the antennal sensory appendageof O. ardosiacus is a multiporous structure with a clusterof sensory units. The dendrite outer segments are highly
branched and fray off into thin digitiform projectionsthat increase membrane surface receptor sensitivity tohighly diluted odour stimulants. The occurrence of poretubules extending from the pore inner end to contact thedendrite tip membrane is considered evidence of astimulus conducting mechanism to the dendritic mem-brane molecular receptors (Zacharuk 1980; Steinbrecht1987). Multiporous sensilla with high numbers ofchemosensory neurons – the most commonly foundtype on the antennae and proved olfactory in function –have been described (Corbiere-Tichane 1971; Meinecke1975; Zacharuk 1980, 1985; Altner and Prillinger 1980;Chapman 1982). Moreover, the complex multicellularstructure of the organ suggests a possible ability of fineodour discrimination related to prey choice. This featureis convergent to ‘‘lobe membraneux’’ of Speophyes

ARTICLE IN PRESS
od
d
cu
so
id
A
B
C
Fig. 6. Transmission electron micrograph of Ophonus ardosiacus antennal sensory appendage. (A) longitudinal ultrathin section; cu:
cuticle, d: dendrite, so: socket. (B) Longitudinal ultrathin section of inner (id) and outer dendritic segment (od) highly branched. (C).
Longitudinal ultrathin section of cuticular pores (black headarrow). Scale bars: 1.1 mm (A), 1.7 mm (B), 0.09 mm (C).
A. Giglio et al. / Zoologischer Anzeiger 247 (2008) 209–221 217
lucidulus larvae (Coleoptera, Bathysciinae) (Corbiere-Tichane 1973). The pores tubule complex is not reportedby Corbiere-Tichane (1971) and Corbiere (1969) butevidence of pore tubule contacts is technique dependentand may require special fixation (Steinbrecht andMueller 1971).
In carabid larvae, biotic factors (food) are mostimportant in evolution of mandibles form while abioticfactors (soil type and sun radiation) influence generalbody form (Zetto Brandmayr et al. 1998). Moreover,larval species show a difference in number and distribu-tion of the sensorial pattern of maxillary and labialpalps related to feeding habits and use of space (Giglioet al. 2003). More generally, comparing the antennalsensory appendage in the taxa so far described (Table 3and references) it seems that variations in form (conical,reduced or absent) are related to larval feedingbehaviour and mobility. Indeed, a fine olfactive dis-crimination is suggested also by the maximal size of thevesicle found in N. rufipes and L. fulvibarbis (Fig. 4):both are specialized collembolan predators. The sensoryappendage is vestigial or fully reduced in locomotoryless active larvae or stages, e.g. in all the instars ofCarterus calydonius (Brandmayr 1975). This mediterra-nean Harpalinae species shows full-time brood care insubterranean paedotrophic nests, supplied by themother beetle with Daucus seeds. The larvae of thisDitomine remain in the nest, few centimeters away from
the core of the nest, throughout their pre-imaginal life(Brandmayr and Brandmayr Zetto 1974). In parasitoidforms (Brachinus, Pheropsophus, Lebia, Eripus) or antnest predatory inquilines (Graphipterus) the sensoryappendage is wide cupuliform just in the first stage thatis the only highly mobile one in order to reach the host/ant nest, while in the older stages the organ is smaller orquite reduced together with other head appendages(Erwin 1967; Habu and Sadanaga 1965; Liebherr andBall 1990; Zetto Brandmayr et al. 1993; Paarmann1985). In Cicindela larvae, this sense organ is missingand replaced by a sensory area (Breyer 1989). For thisgenus the capability of ambushing the prey by usingburrows as shelters is well known (Schremmer 1979).The sensory appendage is bulbous and as long as theantennomere IV in Paussini myrmecophilus species andconical in Metrius and Ozaenini (Bousquet 1986; DiGiulio et al. 2003).
Morphometric analysis shows no correlation betweenantennal sensory appendage dimension and behaviouraltype. However, there are variations in size of thissensory structure in specialized and unspecializedpredators and active seed feeders that can be related tothe prey-food detection. So, the high number ofchemosensory neurons of this multiporous organsuggests a possible ability of fine odour discrimina-tion related to prey choice or host location. Eventhough, further researches, concerning electrophysiological

ARTICLE IN PRESS
Table 3. Variations in form of the antennal sensory appendage related to the feeding behaviour in carabid’s taxa known and in the
‘‘basic’’ family Trachypachidae
Genus Larval feeding behaviour Antennal sensory appendage References
Cupuliform/conic Reduced/
absent
Modified
Trachypachus
Motschulsky, 1844
Unspecialized predators X Arndt and Beutel 1995
Systolosoma Solier, 1849 Unspecialized Predators X Arndt and Beutel 1995
Omoglymmius
Ganglbauer, 1891
Slime mold feeders?
physogastric
X Grandi 1955; Beutel 1992b
Gehringia Darlington,
1933
Unspecialized predators X Lindroth 1960
Metrius Eschscholtz,
1829
Termites-feeder X Bousquet 1986; Beutel
1992a; Di Giulio et al. 2003
Paussus Linneus, 1775 Myrmecophilous X Di Giulio et al. 2003
Notiophilus� Dumeril,
1806
Collembola-feeders X Altner and Bauer 1982;
Luff 1993
Nebria� Latreille, 1802 Small prey feeders X Luff 1993
Leistus� Froehlich, 1799 Collembola-feeders X Daly and Ryan 1979; Luff
1993
Omophron Latreille,
1802
Unspecialized predators X Beutel 1991
Carabus� Linneus, 1758 Earthworm or snail-feeder X Luff 1993
Opisthius Kirby, 1837 Unspecialized predators ‘‘Inconspicuous’’ Lindroth 1960
Promecognathus
Chaudoir, 1846
Unspecialized predators X Bousquet and Smetana
1986
Cicindela Linneus, 1758 Ambush predators in
burrows
X Luff 1993
Loricera Latreille, 1802 Collembola-feeders X Bauer and Kredler 1988
Elaphrus Fabricius, 1775 Collembola-feeders X Luff 1993
Siagona Latreille, 1804 Ant predator? X Zetto Brandmayr et al.
2007
Scarites Fabricius, 1775 Hard prey feeders, sand
diggers
X Luff 1993
Broscus Panzer, 1813 Unspecialized predators X Luff 1993
Pseudomorpha Kirby,
1825
Ant predator, eruciform X Thompson 1979; Erwin
1981
Sphallomorpha
Westwood, 1841
Ambush ant predator X Moore 1974
Trechus� Clairville, 1806 Unspecialized predators X Luff 1993
Typhlotrechus� Mueller,
1913
Unspecialized predators X Luff 1993
Bembidion Stephens,
1828
Unspecialized predators X Luff 1993
Morion Latreille, 1806 Under-bark predators (?) X Thompson 1979
Poecilus� Bonelli, 1810 Unspecialized predators X Luff 1993
Pterostichus Bonelli,
1810
Unspecialized predators X Luff 1993
Laemostenus� Bonelli,1810
Unspecialized predators X Casale 1988
Calathus� Bonelli, 1810 Unspecialized predators X Luff 1993
Carterus Dejean, 1829 Seed feeder, physogastric X Brandmayr 1975
Ophonus� Stephens,1828
Seed feeders X X Brandmayr et al. 1980; Luff
1993
Harpalus� Latreille,1802 s. l.
Seed feeders or predatory X Brandmayr et al. 1980; Luff
1993
Licinus� Latreille, 1802 Snail-feeders X Luff 1993
A. Giglio et al. / Zoologischer Anzeiger 247 (2008) 209–221218

ARTICLE IN PRESS
Table 3. (continued )
Genus Larval feeding behaviour Antennal sensory appendage References
Cupuliform/conic Reduced/
absent
Modified
Graphipterus Forsk(al,
1775
Ant brood predators I stage II–III stages Zetto Brandmayr et al.
1993
Chlaenius Bonelli, 1810 Unspecialized predators X Habu and Sadanaga 1965;
Bonacci et al. 2005
Helluomorphoides Ball,
1951
Ant praedator? ? Thompson 1979; Bousquet
1987
Thermophilum
Basilewski, 1950
Ants and ant brood X Uscidda 1974
Eripus Dejean, 1829 Ectoparasitoids X Liebherr and Ball 1990
Cymindis Latreille, 1806 Unspecialized predators X Luff 1993
Brachinus Weber, 1801 Ectoparasitoids I stage II–III stages Erwin 1967
Pheropsophus Solier,
1833
Ectoparasitoids X Habu and Sadanaga 1965
Lebia Latreille, 1802 Ectoparasitoids I stage II–III stages Habu and Sadanaga 1965;
Luff 1993
�indicate taxa selected in order to give an idea of variation, not for an exhaustive survey. In the seed-eating Ophonus and related taxa we may find
normally developed sensorial vesicles (Ophonus s. str., Metophonus) but also modified, perhaps less-functional forms (Brandmayr et al. 1980). A
similar, kidney-shaped sensorial appendage has been found in Systolosoma lateritium.
A. Giglio et al. / Zoologischer Anzeiger 247 (2008) 209–221 219
response, are needed to correlate the sensory structure withthe specific odour stimulants.
Acknowledgements
The authors are grateful to Dr. Stefano Sponza forthe helpful discussions in statistical analysis. Thisresearch was funded by a MIUR (ex 60%) grant‘‘Ricerche eco-etologiche e morfofunzionali in Coleot-teri carabidi’’ to T. Zetto.
References
Alexandrowicz, J.S., 1954. Notes on the nervous system in the
Stomatopoda. IV. Muscle receptor organs. Pubbl. Staz.
Zool. Napoli 25, 94–111.
Altner, H., Bauer, T., 1982. Ultrastructure of a specialized,
thrust-sensitive, insect mechanoreceptor: stimulus-transmit-
ting structures and sensory apparatus in the rostral horns of
Notiophilus biguttatus. Cell Tissue Res. 226, 337–354.
Altner, H., Prillinger, L., 1980. Ultrastructure of invertebrate
chemo- thermo, and hygroreceptors and its functional
significance. Int. Rev. Cytol. 67, 69–138.
Arndt, E., Beutel, R.G., 1995. Larval morphology of
Systolosoma Solier and Trachypachus Motschulsky (Co-
leoptera: Trachypachidae) with phylogenetic considera-
tions. Entomol. Scand. 26, 439–446.
Bauer, T., Kredler, M., 1988. Adhesive mouthparts in a
ground beetle larva (Coleoptera, Carabidae, Loricera
pilicornis F.) and their function during predation. Zool.
Anz. 221, 145–156.
Behan, M., Ryan, M.F., 1978. Ultrastructure of antennal
sensory receptors of Tribolium larvae (Coleoptera, Teneb-
rionidae). Int. J. Ins. Morphol. Embryol. 7 (3), 221–236.
Beutel, R.G., 1991. Larval head structures of Omophron and
their implications for the relationships of Omophronini
(Coleoptera: Carabidae). Entomol. Scand. 22, 55–67.
Beutel, R.G., 1992a. Study on the systematic position of
Metriini based on characters of the larval head (Coleoptera:
Carabidae). Syst. Entomol. 17, 207–218.
Beutel, R.G., 1992b. Larval head structures of Omoglymmius
hamatus and their implications for the relationships of
Rhysodidae (Coleoptera: Adephaga). Entomol. Scand. 23,
169–184.
Beutel, R.G., Maddison, D.R., Haas, A., 1999. Phylogenetic
analysis of Myxophaga (Coleoptera) using larval charac-
ters. Syst. Entomol. 24, 171–192.
Bloom, J.W., Zacharuk, R.Y., Holodniuk, A.E., 1981.
Ultrastructure of a terminal chordotonal sensillum in larval
antennae of the yellow mealworm, Tenebrio molitor L. Can.
J. Zool. 59 (3), 515–524.
Bonacci, T., Zetto Brandmayr, T., Brandmayr, P., 2005.
Descrizione della larva di Chlaenius chrysocephalus (Coleoptera
Carabidae). Naturalista Sicil. S. IV, XXIX (3–4), 169–176.
Bousquet, Y., 1986. Description of first-instar larva of Metrius
contractus Eschscholtz (Coleoptera, Carabidae) with re-
marks about phylogenetic relationships and ranking of the
genus Metrius Eschscholtz. Can. Entomol. 118, 373–388.
Bousquet, Y., 1987. Description of the larva of Hellumor-
phoides praestus bicolour Harris with comments on the
relationships of the Helluonini (Coleoptera: Carabidae).
Can. Entomol. 119, 921–930.
Bousquet, Y., Smetana, A., 1986. A description of the first
instar larva of Promecognathus Chaudoir (Coleoptera:
Carabidae). Syst. Entomol. 11, 25–31.

ARTICLE IN PRESSA. Giglio et al. / Zoologischer Anzeiger 247 (2008) 209–221220
Brandmayr, P., 1975. Note morfologiche sugli studi preimma-
ginali di Carterus (Sabienus) calydonius Rossi (Coleoptera,
Carabidae). Boll. Soc Entomol. It. 107, 9–19.
Brandmayr, P., Brandmayr Zetto, T., 1974. Sulle cure
parentali e su altri aspetti della biologia di Carterus
(Sabienus) calydonius Rossi, con alcune considerazioni
sui fenomeni di cura della prole sino ad oggi riscontrati in
Carabidi (Coleoptera, Carabidae). Redia 55, 143–175.
Brandmayr, P., Ferrero, E., Zetto Brandmayr, T., 1980. Larval
versus imaginal taxonomy and the systematic status of the
ground beetle taxa Harpalus and Ophonus (Coleoptera:
Carabidae: Harpalini). Entomol. Gen. 6 (2/4), 335–353.
Breyer, S., 1989. Skelet und Muskulatur des Kopfes der Larve
von Cicindela campestris L. (Coleoptera, Cicindelidae).
Stuttgarter Beitr. Naturk., Ser. A 438 (60), 1–60.
Casale, A., 1988. Revisione degli Sphodrina (Coleoptera,
Carabidae, Sphodrini). Monografie V, Museo regionale di
Scienze Naturali, Torino.
Chapman, R.F., 1982. Chemoreception: the significance of
receptor numbers. Adv. Ins. Physiol. 16, 247–356.
Corbiere, G., 1969. Ultrastructure et electrophysiologie du
lobe membraneux de l’antenne chez la larve du Speophyes
lucidulus (Coleoptere). J. Ins. Physiol. 15, 1759–1765.
Corbiere-Tichane, G., 1971. Structure nerveuse enigmatique dans
l’antenne de la larve du Speophyes lucidulus Delar. (Coleoptere
cavernicole de la sousfamille des Bathysciinae). Etude au
microscope eletronique. J. Microsc. 10 (2), 191–202.
Corbiere-Tichane, G., 1973. Sur les structures sensorielles et
leurs fonctions chez la larve de Speophyes lucidulus. Ann.
Speleol. 28 (2), 247–265.
Daly, P.J., Ryan, M.F., 1979. Ultrastructure of antennal
sensilla of Nebria brevicollis (Fab.) (Coleoptera: Carabi-
dae). Int. J. Ins. Morphol. Embryol. 8, 169–181.
Di Giulio, A., Fattorini, S., Kaupp, A., Taglianti, A.V., Nagel,
P., 2003. Review of competing hypoteses of phylogenetic
relationships of Paussinae (Coleoptera, Carabidae) based
on larval characters. Syst. Entomol. 28 (4), 509–537.
Emden van, F.I., 1942. A key to the genera of larval Carabidae
(Col.). Trans. R. Entomol. Soc. London 92, 1–99.
Erwin, T.L., 1967. Bombardier beetles (Coleoptera, Carabi-
dae) of North America: Part II. Biology and behavior of
Brachinus pallidus Erwin in California. The Coleopterist’s
Bull., 41–55.
Erwin, T.L., 1981. A synopsis of the immature stages of
Pseudomorphini (Coleoptera: Carabidae) with notes on
tribal affinities and behaviour in relation to life with ants.
The Coleopterist’s Bull. 35 (1), 53–68.
Evans, M.E.G., 1994. The carabid body plan: a functional
interpretation. In: Desender, K., et al. (Eds.), Carabid
Beetles: Ecology and Evolution. Kluwer Academic Publish-
ers, Netherlands, pp. 25–31.
Forsythe, T.G., 1991. Feeding and locomotory functions in
relation to body form in five species of ground beetle
(Coleoptera: Carabidae). J. Zool. Soc. London 223, 233–263.
Giglio, A., Ferrero, E.A., Perrotta, E., Tripepi, S., Zetto
Brandmayr, T., 2003. Ultrastructure and comparative
morphology of mouth-part sensilla in ground beetle larvae
(Insecta, Coleoptera, Carabidae). Zool. Anz. 242, 277–292.
Grandi, G., 1955. Rhysodes germari Ganglb. (Coleoptera
Rhysodidae). Documenti morfologici ed eto-ecologici (XXI
contributo alla conoscenza degli insetti a regime specializ-
zato). Boll. dell’Ist. di Entomol. dell’Univ. Bologna 21,
179–196.
Habu, A., Sadanaga, K., 1965. Illustrations for identification
of larvae of the Carabidae found in cultivated fields and
paddy-fields. Bull. Nat. Inst. Agric. Sci. Japan IIIC (19),
81–216.
Jeannel, R., 1941. Faune de France. Coleopteres Carabique 39,
Lechevalier, Paris.
Jeannel, R., 1942. Faune de France. Coleopteres Carabique 40,
Lechevalier, Paris.
Lawrence, J.F., 1991. Order Coleoptera. In: Stehr, F.W. (Ed.),
Immature Insects, vol. 2. Kendall/Hunt, Dubuque, Iowa,
pp. 144–658.
Liebherr, J.K., Ball, G.E., 1990. The first instar larva of Eripus
oaxacanus Straneo & Ball (Coleoptera: Carabidae: Pelecii-
ni): indicator of affinity or convergence? Syst. Entomol. 15,
69–79.
Lindroth, C.H., 1960. The larvae of Trachypachus Mtsch.,
Gehringia Darl., and Opisthius Kby. (Col. Carabidae).
Opusc. Entomol. XXV (1–2), 30–42.
Luff, M.L., 1993. The Carabidae (Coleoptera) larvae of
Fennoscandia and Denmark. Fauna Entomol. Scand. 27,
1–187.
Luna de Carvalho, E., 1991. Revisao do estudo das larvas de
Carabıdeos Paussinae e de subfamılias afins (Coleoptera,
Adephaga). Elytron 5, 285–310.
Meinecke, C.C., 1975. Riechsensillen und Systematik der
Lamellicornia (Insecta, Coleoptera). Zoomorphologie 82,
1–42.
Minelli, A., 2004. A segmental analysis of the beetle antenna.
Studi Trent. Sci. Nat., Acta Biol. 81, 91–101.
Moore, B.P., 1974. The larval habits of two species of
Sphallomorpha westwood (Coleoptera: Carabidae, Pseudo-
morphinae). J. Aust. Entomol. Soc. 13, 179–183.
Paarmann, W., 1985. Larvae preying on ant broods: an
adaptation of the desert carabid beetle Graphipterus
serrator Forsk(al (Col., Carabidae) to arid environments.
J. Arid Environ. 9, 210–214.
Roppel, R.M., Arbogast, R.T., Zeigler, J.A., 1972. Antennal
sensilla of the larval sawtoothed grain beetle, Oryzaephilus
surinamensis (Coleoptera, Cucujidae). Rev. Can. Biol. 31
(1), 9–20.
Ryan, M.F., Behan, M., 1973. The sensory receptors of
Tribolium larvae. Physiol. Zool. 46 (3), 238–244.
Schremmer, F., 1979. Ethokologische Beobachtungen zum
Wohnrohrenbau bei Larven der mitteleuropaischen San-
dlaufkafer-Art Cicindela silvicola (Coleoptera, Cicindeli-
dae). Entomol. Gen. 5 (3), 201–219.
Scott, D.A., Zacharuk, R.Y., 1971. Fine structure of the
dendritic junction body region of the antennal sensory cone
in a larval elaterid (Coleoptera). Can. J. Zool. 49 (6),
817–821.
Spence, J.R., Sutcliffe, J.F., 1982. Structure and function of
feeding in larvae of Nebria (Coleoptera: Carabidae). Can. J.
Zool. 60, 2382–2394.
Steinbrecht, R.A., 1987. Functional morphology of phero-
mone-sensitive sensilla. In: Prestwich, G.D., Blomquist,
G.J. (Eds.), Pheromone Biochemistry. Academic Press,
New York, pp. 353–384.

ARTICLE IN PRESSA. Giglio et al. / Zoologischer Anzeiger 247 (2008) 209–221 221
Steinbrecht, R.A., Mueller, B., 1971. On the stimulus
conducting structures in insect olfactory receptors. Z.
Zellforsch. Mikrosk. Anat. 117, 570–575.
Thompson, R.G., 1979. Larvae of North American Carabidae
with a key to the tribes. In: Erwin, T.L., Ball, G.E.,
Whitehead, D.R. (Eds.), Carabid Beetles: Their Evolution,
Natural History and Classification. Dr W. Junk BV
Publishers, The Hague, pp. 209–291.
Uscidda, C., 1974. La larva di Megacephala euphratica Latr. e
Dej. e Thermophila sexmaculata (Fabr.). ‘‘Studi Sassaresi’’
Sez. III- Ann. della Fac. di Agraria dell’Univ. Sassari, 22.
Zacharuk, R.Y., 1962. Sense organs of the head of larvae of
some Elateridae (Coleoptera): their distribution, structure
and innervation. J. Morphol. 111, 1–33.
Zacharuk, R.Y., 1980. Ultrastructure and function of insect
chemosensilla. Ann. Rev. Entomol. 25, 27–47.
Zacharuk, R.Y., 1985. Antennae and sensilla. In: Kerkut,
G.A., Gilbert, L.S. (Eds.), Comparative Insect Physiology,
Biochemistry and Pharmacology, vol. 6. Pergamon Press,
Oxford, pp. 1–69.
Zetto Brandmayr, T., 1976. Nutrizione e allevamento di
Carabidi esclusivamente fitofagi: spermofagia larvale di
Ophonus ardosiacus Lutsh. Redia 59, 197–206.
Zetto Brandmayr, T., 1978. Studi sulla fitofagia nei Carabidi:
spermofagia larvale di Ophonus ardosiacus Lutsh. (Coleop-
tera, Carabidae). Atti XI Congresso It. Entomol. Portici-
Sorrento, 59–64.
Zetto Brandmayr, T., Brandmayr, P., Marano, I., Paarmann,
W., 1993. The larva of Graphipterus serrator (Forskal 1775)
(Coleoptera Carabidae Graphipterini): description and
functional morphology. Trop. Zool. 6, 299–312.
Zetto Brandmayr, T., Giglio, A., Marano, I., Brandmayr, P.,
1998. Morphofunctional and ecological features in
carabid (Coleoptera) larvae. Mus. Reg. Sci. Nat. Torino,
449–490.
Zetto Brandmayr, T., Mazzei, A., Talarico, F., Giglio, A.,
Bauer, T., Brandmayr, P., 2007. The larva of Siagona
europaea Dejean, 1826: morphology and collecting techni-
que for a subterranean blind ‘‘running ant killer’’ (Coleop-
tera: Carabidae). It. J. Zool. 74 (3), 239–245.




















