
Management of Nonsustained Ventricular Tachycardia Guided By Electrophysiological Testing
Upload
independentCategory
view
2download
0
ANTENNAL CONTACT CHEMORECEPTION IN GROUND BEETLES
(COLEOPTERA: CARABIDAE)
JOOKSIKLASTE (COLEOPTERA: CARABIDAE)ANTENNAALNE KONTAKTNE KEMORETSEPTSIOON
MARIT KOMENDANT
A Thesis
submitted for the degree of Doctor of Philosophy in Entomology
Väitekiri
filosoofiadoktori kraadi taotlemiseks entomoloogia erialal
Tartu 2011

EESTI MAAÜLIKOOLESTONIAN UNIVERSITY OF LIFE SCIENCES

ANTENNAL CONTACT CHEMORECEPTION
IN GROUND BEETLES
COLEOPTERA: CARABIDAE
JOOKSIKLASTE (COLEOPTERA: CARABIDAE)ANTENNAALNE KONTAKTNE KEMORETSEPTSIOON
MARIT KOMENDANT
A Th esis
submitted for the degree of Doctor of Philosophy in Entomology
Väitekiri
fi losoofi adoktori kraadi taotlemiseks entomoloogia erialal
Tartu 2011

Institute of Agricultural and Environmental Sciences
Estonian University of Life Sciences
According to the verdict No 80 of March 21, 2011, the Doctoral
Committee of the Agricultural and Natural Sciences of the Estonian
University of Life Sciences has accepted this thesis for the defence of the
degree of Doctor of Philosophy in Entomology.
Opponent: Prof. Habil. Dr. Vincas Būda
Institute of Ecology,
Vilnius University, Lithuania
Supervisors: Dr. Enno Merivee
Estonian University of Life Sciences
Prof. Anne Luik
Estonian University of Life Sciences
Defence of this thesis:
Estonian University of Life Sciences, Karl Ernst von Baer House, Veski
4, Tartu, on May 6, 2011, at 10.00.
Th e English language was edited by Ingrid H. Williams and the Estonian
by Tiina Halling.
Publication of this thesis is granted by the Estonian University of Life
Sciences and by the Doctoral School of Earth Sciences and Ecology
created under the auspices of European Social Fund.
© Marit Komendant 2011
ISBN 978-9949-484-00-3

5
CONTENTS
LIST OF ORIGINAL PUBLICATIONS .......................................... 7
1. INTRODUCTION ....................................................................... 8
2. AIMS OF THE STUDY ............................................................. 12
3. MATERIAL AND METHODS ................................................. 13
3.1. Test beetles ............................................................................. 13
3.2. Electrophysiology ................................................................... 13
3.3. Chemicals and preparation of stimulating solutions ................ 14
3.4. Feeding bioassays (V) ............................................................. 15
3.5. Data analysis ........................................................................... 15
4. RESULTS .................................................................................... 17
4.1. Taste sensilla on the antennal fl agellum of P. oblongopunctatus and
P. aethiops .............................................................................. 17
4.2. Spike shapes and amplitudes evoked by tested chemicals ........ 17
4.3. Eff ect of stimulus pH to the responses of the electrolyte-sensitive
chemoreceptor neurons innervating antennal taste sensilla of P.
aethiops and P. oblongopunctatus ............................................. 18
4.3.1. Response to 100 mmol l -1 acetate buff ers at pH 3–6 ..... 18
4.3.2. Response to 100 mmol l -1 phosphate buff ers at
pH 5.8–8.1 (11.1) ......................................................... 19
4.3.3. Response to 100 mmol l -1 Na+-salts with and without
addition of 10 mmol l -1 NaOH..................................... 19
4.4. Plant sugar sensitivity in ground beetles ................................. 20
4.4.1. Electrophysiological identifi cation of the sugar-sensitive
neuron in antennal taste sensilla of P. aethiops .............. 20
4.4.2. Electrophysiological responses of the chemoreceptor
neurons from antennal taste sensilla of P.
oblongopunctatus to plant sugars and amino acids ......... 21
4.4.2.1. Response of antennal taste neurons to sugars ... 21
4.4.2.2. Response of antennal taste neurons to amino
acids ................................................................ 22
4.5. Response of antennal taste neurons to plant alkaloids and
glucosides ............................................................................... 22
4.5.1. Electrophysiological identifi cation of the alkaloid-sensitive
neuron in antennal taste sensilla of P. oblongopunctatus .. 22

6
4.5.2. Stimulating eff ect of plant alkaloids and glucosides on
the responses of the antennal chemoreceptor neurons ... 23
4.5.3. Peripheral inhibition of the salt- and pH-sensitive
neurons by some plant secondary compounds .............. 23
4.5.4. Eff ect of quinine hydrochloride on the feeding response
of P. oblongopunctatus ................................................... 24
5. DISCUSSION .............................................................................. 25
5.1. pH sensitivity in ground beetles (I, II) .................................... 25
5.1.1. Th e occurrence of two chemoreceptor neurons in insect
contact chemoreceptors responding to salts ................... 25
5.1.2. Th e eff ect of stimulus pH to the salt- and pH-sensitive
neurons in antennal taste sensilla of P. aethiops and P.
oblongopunctatus ........................................................... 26
5.1.3. pH preference in ground beetles .................................... 27
5.2. Sensitivity of ground beetles to plant sugars and amino acids
(III, IV) ................................................................................. 28
5.2.1. Response of the chemoreceptor neurons from antennal
taste sensilla to sugars ................................................... 28
5.2.2. Response of the chemoreceptor neurons from antennal
taste sensilla to amino acids .......................................... 30
5.3. Response of the chemoreceptor neurons in the antennal taste sensilla
of P. oblongopunctatus to plant secondary compounds ............ 31
5.3.1. Stimulating eff ect of plant alkaloids and glucosides on the
responses of the antennal chemoreceptor neurons ......... 31
5.3.2. Peripheral inhibition of the salt- and pH-sensitive neurons
by some plant secondary compounds ............................ 33
CONCLUSIONS ............................................................................. 35
REFERENCES ................................................................................ 37
SUMMARY IN ESTONIAN ......................................................... 46
ACKNOWLEDGEMENTS ............................................................ 51
PUBLICATIONS ............................................................................ 53
CURRICULUM VITAE ............................................................... 129
ELULOOKIRJELDUS .................................................................. 130
LIST OF PUBLICATIONS ........................................................... 131

7
LIST OF ORIGINAL PUBLICATIONS
Th is thesis is a review of the following papers, which are referred to by
Roman numerals in the text. Th e papers are reproduced by kind permission
of the following journals: Physiological Entomology (I) and Journal
of Insect Physiology (II, III, IV).
I Merivee, E., P loomi, A., Milius, M., Luik, A., Heidemaa,
M., 2005. Electrophysiological identifi cation of antennal pH
receptors in the ground beetle Pterostichus oblongopunctatus. Physiological Entomology 30(2), 122–33.
II Milius, M., Merivee, E., Williams, I., Luik, A., Mänd, M., Must,
A., 2006. A new method for electrophysiological identifi cation
of antennal pH receptor cells in ground beetles: the example of
Pterostichus aethiops (Panzer, 1796) (Coleoptera, Carabidae). .
Journal of Insect Physiology 52, 960–967.
III Merivee, E., Must, A., Milius, M., Luik, A., 2007.
Electrophysiological identifi cation of the sugar cell in antennal
taste sensilla of the predatory ground beetle Pterostichus aethiops. Journal of Insect Physiology 53(4), 377–384.
IV Merivee, E., Märtmann, H., Must, A., Milius, M., Williams, I.,
Mänd, M., 2008. Electrophysiological responses from neurons
of antennal taste sensilla in the polyphagous predatory ground
beetle Pterostichus oblongopunctatus (Fabricius 1787) to plant
sugars and amino acids. Journal of Insect Physiology 54, 1213–
1219.
V Komendant, M., Merivee, E., Must, A., Tooming, E., Williams,
I., Luik, A. Electrophysiological response of the chemoreceptor
neurons in the antennal taste sensilla to plant alkaloids and
glucosides in the granivorous ground beetle Pterostichus
oblongopunctatus. (in review)
Table 1. Author’s contribution to each article (%)
I II III IV VIdea and design 10% 80% 25% 0% 80%Data acquisition 50% 100% 50% 40% 100%Data analysis, statistics 50% 80% 50% 40% 80%Writing 10% 80% 15% 0% 90%

8
1. INTRODUCTION
Th e ground beetles (Coleoptera, Carabidae) with 40,000 species belong to
the most numerous families of beetles, their distribution being cosmopoli-
tan. Approximately 275 species have been found in Estonia (Haberman,
1968; Silfverberg, 2004). Binding of ground beetles to certain habitat
types and microhabitats is determined by numerous biotic and abiotic
factors such as food conditions, vegetation type, presence and distribution
of competitors, life history and season, landscape characteristics, agricul-
tural cultivation impacts, temperature and humidity conditions, light
intensity, and others. Favourite wintering sites are well aerated. Habitat
choice in many ground beetles is so specifi c that they are often used to
characterize habitats (Th iele, 1977; Lövei and Sunderland, 1996; Ings
and Hartley, 1999; Cole et al., 2002; Blake et al., 2003; Eyre and Luff ,
2004; Purtauf et al., 2005).
Chemically mediated habitat selection has been shown to occur in several
species of ground beetle by Evans (1982, 1983, 1984, 1988). By behavioural
experiments, he demonstrated that olfactory neurons receptive to methyl
esters of palmitic and oleic acid emitted by the mat-forming blue-green
algae Oscillatoria animalis Agardh and Oscillatoria subbrevis Schmidle are
present in sensillae on the antennae of those Bembidiini associated with
the shores of saline lakes. Salt content and pH of the soil may also have
importance in determining the distribution of ground beetles (Th iele,
1977; Paje and Mossakowski, 1984; Hoback et al., 2000; Irmler, 2001,
2003; Magura, 2002). Behavioural ablation experiments have shown that
presumptive contact chemoreceptive sensilla are located on the ground
beetles’ antennae (Paje and Mossakowski, 1984; Hoback et al., 2000).
Scanning electron microscope studies show that the antennae of ground
beetles are well equipped with various chemoreceptors. In addition, to
numerous small basiconic olfactory sensilla, approximately 56 to 70 larger
bristle-like contact chemosensilla (35–200 μm in length) have been found
on the antennal fl agellum of adults in several genera (Kim and Yamasaki,
1996; Merivee et al., 2000, 2001, 2002). In the ground beetle Nebria
brevicollis these large sensilla are each innervated by one mechanoreceptive
and four chemoreceptive neurons (Daly and Ryan, 1979). A similar set
of neurons is typical for contact chemoreceptors in various insect groups
(Morita and Shiraishi, 1985; Chapman, 1998; Jørgensen et al., 2006).

9
By electrophysiological single sensillum stimulations the occurrence of
two chemoreceptor neurons responding to salts has been demonstrated
in the antennal taste bristles of the ground beetle Pterostichus aethiops. One of them, producing spikes with negative polarity was highly sensitive
to both ionic composition and concentration of the tested nine salt solu-
tions showing a phasic-tonic type of reaction with a pronounced phasic
component. Th e stimulating eff ect was dominated by the cations involved,
and in most cases, monovalent cations were more eff ective stimuli than
divalent cations. Th is neuron was classifi ed as the salt-sensitive neuron.
Th e specifi c function for the second neuron (the “A-cell”) generating two-
tipped spikes with positive polarity still remains to be explained. It was
observed, however, that alkaline salts usually evoked substantially more spikes in the second neuron than acid salts, but the ionic composition of the salts also aff ected the responses of the second neuron (Merivee et al., 2004). Th us, the “A-cell” with positive spikes seems to be the most probable candidate for pH sensitivity in ground beetles, but no electro-physiogical evidence is available. Th e special function of the third and fourth chemoreceptor neurons of the sensillum also needs to be clarifi ed in ground beetles.
Data on food items are available for approximately 1100 species and sub-species of ground beetles of the world. A total of 73% are carnivorous, 19% are omnivorous and 8% are exclusively herbivorous (Th iele, 1977; Larochelle, 1990; Toft and Bilde, 2002). Of these, many species predate on seeds (Honek et al., 2003). Ground beetles use a variety of external stimuli in order to locate food. Behavioural tests and observations have indicated that visual, tactile, olfactory and gustatory cues may be involved (Wheater, 1989; Kielty et al., 1996; Toft and Bilde, 2002). Various sugars and amino acids function as strong phagostimulants in most phytophagous insects equipped with specialized receptor neurons for these chemicals (Chapman, 1998; Hansen-Delkeskamp, 1998; Schoonhoven and van Loon, 2002; Chapman, 2003) suggesting that these chemicals may have a phagostimulatory role in herbivorous and omnivorous ground beetles,
too. Adults of the genus Pterostichus appear to be primarily carnivorous,
feeding on small prey animals, but in many species, plant matter also
forms part of their diet, and, in some the proportion of animal to plant
matter eaten varies with the season (Th iele, 1977; Hengeveld, 1980; Lövei
and Sunderland, 1996). Johnson and Cameron (1969) collected data on
the extent of herbivory in more than 150 species of ground beetle. Some
species in the genera Pterostichus, including Pterostichus oblongopunctatus,

10
Amara, Agonum and Harpalus consume conifer seeds and young seedlings
immediately following germination. In Sweden, P. oblongopunctatus and
Calathus micropterus are the most important predators of Pinus sylvestris seed. In the fi eld, seed predation typically resulted in >20% seed mortal-
ity, reaching even higher levels (up to 60%) on some occasions and most
predation (81% of all damaged seeds) was due to ground beetles (Heikkilä,
1977; Nystrand and Granström, 2000). Th erefore, as in other phytopha-
gous insects, ground beetles feeding exclusively or partly on plant material,
may also have plant sugar- and amino acid-sensitive neurons associated
with their feeding habits although no experimental data is available.
In addition to detecting phagostimulants, most animals, including her-bivorous insects, possess chemoreceptor neurons responding to a diverse range of feeding deterrent compounds. When stimulated, deterrent neurons reduce or fully stop feeding activity (Dethier, 1980; Schoonhoven and van Loon, 2002; Chapman, 2003; Wang et al., 2004). Natural deterrent stimuli recognized by chemoreceptor neurons of herbivorous insects con-stitute the largest and most structurally diverse class of gustatory stimuli. It has been shown that plant glucosides, for example, sinigrin and salicin, and various alkaloids like nicotine (pyrrolidine group), strychnine and quinine (quinoline group), caff eine (purine group) and several others stimulate the deterrent neuron in many herbivorous insects. Th reshold concentrations for compounds that stimulate a deterrent neuron vary from 10-7 to 10-2 mol l-1, but the majority of these are in the range of 10-4 to 10-2 mol l-1. Th erefore, deterrent neurons fulfi l a central role in host-plant recognition and have, since their discovery (Ishikawa 1966), attracted much interest (Glendinning, 2002; Ryan, 2002; Schoonhoven and van Loon, 2002; Chapman, 2003; Jørgensen et al., 2007).
Herbivory, including pre- and post-dispersal plant seed predation, is ex-
tremely common in virtually all ecosystems. Seeds with their very high
nutrient values per unit volume are extremely attractive to granivores as
food (Janzen, 1971). To counterbalance the eff ects of herbivory, plants
have developed a broad spectrum of secondary metabolites involved in
plant defense, which are collectively known as antiherbivory compounds
and can be classifi ed into three main sub-groups: nitrogen compounds
(including alkaloids, cyanogenic glycosides and glucosinolates), terpenoids,
and phenolics. Th ese compounds can act as feeding deterrents or toxins
with various modes of action to herbivores, or reduce plant digestibility
(Janzen, 1971; Jolivet, 1998; Ryan, 2002; Chapman, 2003; Hulme and

11
Benkman, 2004; Davis et al., 2008). Seeds are sources of some of the
most toxic natural products known to humans and the secondary chemi-
cals in seeds present formidable challenges to granivores. However, seed
chemical defenses can only be assessed with reference to specifi c target
organisms since a secondary chemical may not be equally toxic to all
granivores. Some seeds are so toxic that all seed-predators avoid them
(Harborne, 1993; Hulme and Benkman, 2004). Most toxins are usually
in small concentrations (<5%) in the seed and some alkaloids, for exam-
ple, can be lethal at concentrations as low as 0.1% (Hatzold et al., 1983;
Harborne, 1993). Unfortunately, there is no evidence that plant secondary
compounds cause feeding deterrency in ground beetles. However, due to
selective seed predation by ground beetles (Toft and Bilde, 2002; Tooley
and Brust, 2002), the presence of a deterrent neuron in their antennal
contact chemoreceptors should be expected.

12
2. AIMS OF THE STUDY
Th e aims of this study were:
1) by electrophysiological experiments, to test the response of the
chemoreceptor neurons in the antennal taste sensilla of the
ground beetle P. oblongopunctatus to acetate and phosphate
buff ers over a wide range of pH (3–11) in order to clarify the
eff ect of stimulus pH on the neurons, and to clarify the possible
existence of a pH-specifi c neuron in this species (I);
2) using various alkalized and non-alkalized salts as stimuli, to
test the electrophysiological response of the antennal contact
chemoreceptor neurons of the ground beetle P. aethiops to
stimulus pH in order to collect additional data on functioning
of a specifi c pH-sensitive neuron in ground beetles (II);
3) by electrophysiological experiments, to determine whether sugar-
and amino acid-sensitive receptor neurons are present in the
antennal taste sensilla of P. aethiops (III) and P. oblongopunctatus (IV) by testing for stimulatory eff ects of various sugars of live
and decaying plant origin, and amino acids on these sensilla;
4) to test the stimulatory eff ect of some toxic plant secondary
compounds (alkaloids and glucosides) on the chemoreceptor
neurons of the antennal taste sensilla of the granivorous P. oblongopunctatus in order to identify the specifi c feeding deterrent
neuron stimulated by these chemicals (V).

13
3. MATERIAL AND METHODS
3.1. Test beetles
Adult P. oblongopunctatus and P. aethiops were collected from a local
population in southern Estonia. Th ey were kept in 30×20×10 cm plastic
boxes fi lled with moistened moss and pieces of brown-rotted wood in a
refrigerator at 5–6 °C. Th ree to four days prior to the experiments, the bee-
tles were transferred to room temperature (20–23 °C), fed with moistened
cat food (Kitekat, Master Foods, Poland, I, II; Friskies Vitality+, Nestle´
Purina, Hungary, III, IV, V) and given clean water to drink every day.
3.2. Electrophysiology
Each beetle to be tested was restrained securely in a conical tube made
of thin sheet-aluminium of a size that allowed the head and antennae
to protrude from the narrower end. Th e wider rear end of the tube was
blocked with plasticine to prevent the beetle from retreating out of the
tube. Its antennae were fi xed horizontally to the edge of an aluminium
stand with special clamps and beeswax, so that the horizontally located
large contact chemoreceptors (chaetoid taste sensilla) were visible from
above under a light microscope, and easily accessible for micromanipula-
tion from the side. Care was taken not to contaminate the taste sensilla
by contact with instruments or beeswax. Each prepared test beetle was
placed in a Faraday cage (15×10×5 cm) for electrophysiology.
Spikes from taste neurons of the taste sensilla were recorded by the conven-tional single sensillum tip-recording technique (Hodgson et al., 1955). To achieve a good signal-to-noise ratio, the indiff erent tungsten microelectrode was inserted into the base of the fl agellum free of muscular tissue. It was connected to an AC amplifi er via a calibration source, and earthed. Th e recording glass micropipette fi lled with stimulating solution was brought into contact with the tip of the sensillum by means of a micromanipula-tor under visual control through a light microscope at a magnifi cation of 300×. Immediately before each stimulation, 2–3 drops of solution were squeezed out of the micropipette tip by a syringe connected to the rear end of the micropipette by means of a silicone tube, and adsorbed onto a piece of fi lter paper attached to another micromanipulator, in order to

14
avoid concentration changes in the capillary tip due to evaporation. Spikes picked up by the recording electrode were fi ltered with a bandwidth set at 100–2000 Hz, amplifi ed (input impedance 10 GV, amplifi cation factor 2000×), monitored on an oscilloscope screen, and relayed to a computer via an analogue-to-digital input board DAS-1401 (Keithley, Taunton, MA, USA) for data acquisition, storage, and analysis using TestPoint software (Capital Equipment Corp., Billerica, MA, USA) at a sampling rate of 10 kHz. Responses were recorded from neurons of one (II) to fi fteen (I, III, IV, V) randomly selected sensilla on the fl agellomeres 1–9 of each test beetle. Each sensillum was stimulated once only with one solution (IV). In most cases, however, each sensillum was stimulated twice, with control (10 mmol l-1 electrolyte) and test stimulus solution, respectively. In each subsequent test beetle, the order of stimulation was reversed (I, II, III, V). During each 1 s (IV) or 5 s (I, II, III, V) stimulation period, spikes picked up by the recording electrode were counted and analysed. Responses of the sensilla to mechanical stimulation were also tested, especially in those rare cases when it was not clear whether a certain class of recorded spikes originated from the chemosensory or mechanosensory neuron. Th e tip of the sensillum was rapidly moved from its resting position toward the antennal shaft by 15–20°, held in this position for 2–3 s, and then rapidly moved back to its initial position using the recording electrode.
3.3. Chemicals and preparation of stimulating solutions
To explain the eff ect of stimulus pH to the responses of antennal chemo-receptor neurons, 100 mmol l-1 acetate and phosphate buff ers in the pH range 3–8.1 were prepared. For some experiments, the pH of the phosphate buff er was raised to 11.1 with NaOH. To produce stimulating buff ers of approximately the same pH but of diff erent concentrations of ions, the 100 mmol l-1 phosphate buff er (pH 7.6) was diluted in distilled water to 10 mmol l-1 with pH 7.8 (I). To test for the presence of the pH neuron in the antennal taste bristles of the ground beetle P. aethiops more precisely, three solutions of 100 mmol l-1 Na+-salts, Na
2HPO
4, NaCl and NaH
2PO
4 were prepared, acid, neutral
and alkaline, respectively; these served as control stimuli. Test stimuli were prepared by adding small amounts of NaOH to the 100 mmol l-1 salts resulting in three electrolyte mixtures with 1 mmol l-1 NaOH and three mixtures with 10 mmol l-1 NaOH. Addition of NaOH to these salt solu-tions does not change their ionic composition and the Na+ concentration

15
increases only to a small extent (1% and 10%, respectively). At the same time, the pH level of these solutions rises substantially. Th e stimulating eff ect of these sodium salt solutions with and without NaOH was expected to be approximately similar for the salt-sensitive neuron but not for the pH-sensitive neuron if present (Table 1, II). Th e fi nal pH of the stimulat-ing solutions was measured with a pH meter E6115 (Evikon, Estonia).Sugars (III; Table 1, IV), amino acids (Table 1, IV), alkaloids and gluco-sides (Table 1, V) were dissolved in 10 mmol l-1 electrolyte (KCl, NaCl, choline chloride) to give good electrical conductivity. Stimulating chemicals were kept at 5 °C for less than a week. Chemicals were obtained from AppliChem (Germany) and Sigma-Aldrich company.
3.4. Feeding bioassays (V)
Two-choice feeding experiments were carried out in 110-mm diameter
glass Petri dishes. To ensure high air humidity inside the dishes, each
was lined with a moistened Whatman fi lter paper circle (Schleicher and
Schuell, England). Two pine seeds (Pinus sibirica), treated for 20 min with
distilled water (control) and 0.01 to 50 mmol l-1 quinine hydrochloride,
respectively, were placed in the centre of each Petri dish, 50 mm from each
other. Only seeds with seed coats removed were used in the experiments.
Test beetles, one in each Petri dish, were allowed to choose between, and
feed on the seeds for 24 h (20 °C, 14:10 light:dark). After 24 h, the beetles
were removed and the percentage of predation on control seeds and seeds
treated with alkaloid, was visually determined. At each concentration of
quinine hydrochloride, the feeding preference of the beetles was tested
in fi ve replications (10 beetles in each).
3.5. Data analysis
Automated classifi cation and counting of action potentials from taste
sensilla was not an appropriate method for data analysis because two ac-
tion potentials frequently coincided to produce an irregular waveform.
Instead, spikes from several chemosensory neurons innervating anten-
nal taste sensilla tested were visually distinguished by their polarity and
amplitude (Figs. 1, III, IV, V), and counted, using TestPoint software.
Firing rates (spikes/sec) were analysed by counting the number of spikes
within 5 s after stimulus onset. Mean fi ring rates and their standard errors

16
were calculated. t-test for paired samples (I, II, III,), t-test for independ-
ent samples (IV), Wilcoxon test (V) and ANOVA, Tukey test (IV) were
used to determine the signifi cance of diff erences between means. Spike
amplitude analysis was performed in order to explain which specifi c neuron
was stimulated by the chemical classes tested (IV, V).

17
4. RESULTS
4.1. Taste sensilla on the antennal fl agellum of P. oblongopunctatus and P. aethiops
Large blunt-tipped 100–200 μm long taste bristles are located in whorls
in the distal part of fl agellomeres. Th ere are six on the fi rst fl agellomere
and seven on the following fl agellomeres counting from the beetle’s head.
In addition to those, there are six bristles symmetrically at the tip of the
terminal fl agellomere. Hence, the total number of taste bristles on each
antennal fl agellum is 68. All these bristles seemed to respond in a similar
manner to the stimuli tested.
4.2. Spike shapes and amplitudes evoked by tested chemicals
Spike waveform analysis showed that spikes from four diff erent chemore-
ceptor neurons innervating antennal taste sensilla of the ground beetles P.
oblongopunctatus and P. aethiops could be distinguished in the recordings
obtained in response to tested chemicals. Spikes with negative polarity,
1–2.5 mV in amplitude, arose from the salt-sensitive neuron. Th is neuron
generated spikes in a phasic-tonic manner and had an extremely short
response latency lasting less than 10 ms (Fig. 1, III; Fig. 1, V). Two-tipped
spikes with positive polarity, 2–8 mV in amplitude, were generated by
the pH-sensitive neuron (Fig. 1, I, III; Figs. 1 and 2, IV). Compared to
the spikes from the salt-sensitive neuron, these positive spikes appeared
with a much longer delay (Fig. 1, II; Fig. 1A, I, V). Th e shape of these
impulses is complex because they are composed of two separate electrical
events in quick succession and most probably produced by two separate
spike generation sites of the same neuron. Minor diff erences in the time
interval between these two events occuring among antennal taste sensilla
may cause considerable diff erences in the shape of the resulting double-
spike. Usually, the double-spikes of the pH-sensitive neuron described are
two-tipped (Fig. 2A and B, IV). In some sensilla, however, only a small
characteristic notch or discontinuity in the rising phase of these impulses
indicates their complex nature (Fig. 2C and D, IV). Th e sugar-sensitive
neuron also produced spikes with negative polarity, but smaller in ampli-
tude than those of the salt-sensitive neuron (Fig. 1B, III, IV; Fig. 1J, V).
Th e fourth chemoreceptor neuron of the taste sensillum was stimulated
by some alkaloids. Th e negative spikes it produced were variable in size

18
(Fig.1 B–E, V) but diff erent from those originating from the salt- and
sugar-sensitive neurons (Fig. 2, V).
Impulse activity of the mechanosensory neuron belonging to these taste
sensilla and responding to bending of the bristle was recorded from some
intact bristles as well as from bristles with a broken tip. Spikes produced
by the mechanosensory neuron were small but variable in size, and nega-
tive in polarity (Fig. 1C and D, IV).
4.3. Eff ect of stimulus pH to the responses of the electrolyte-sensitive chemoreceptor neurons innervating antennal taste
sensilla of P. aethiops and P. oblongopunctatus
Two chemoreceptor neurons from fl agellar taste bristles of P. aethiops and
P. oblongopunctatus may respond to stimulation with various electrolyte
solutions in a phasic-tonic manner. According to their response charac-
teristics these neurons were classifi ed as salt- (cation-) and pH-sensitive
neuron distinguished by the amplitude, shape and polarity of spikes they
generated (Fig. 1, II). Th e phasic component of the response was more pro-
nounced in the salt-sensitive neuron compared to that of the pH-sensitive
neuron (Figs. 2 and 3, II). Due to large variations in responsiveness of
the pH-sensitive neuron from diff erent taste bristles and diff erent samples
of beetles to the same stimulus (Fig. 4, A-cell, I), stimulating solutions
were tested in pairs to allow a more precise comparison of the responses
to the tested stimuli.
4.3.1. Response to 100 mmol l-1 acetate buff ers at pH 3–6
Th e pH-sensitive neuron did not respond to acetate buff er at pH 3 and
only a very weak response was observed at pH 4; fi rst second response
below 1 spikes/s (Fig. 2A, A-cell, I). Th e number of spikes generated by
the salt-sensitive neuron at pH 3 and 4 was much higher, fi rst second
responses 5.7 and 9.5 spikes/s, respectively (Fig. 2A, B-cell, I). Accord-
ing to how much the pH level of the stimulating buff ers increased fur-
ther, more spikes by the pH- and salt-sensitive neurons were produced.
However, even at pH 6, the mean fi ring rate of the pH-sensitive neuron
remained below 5 spikes/s while that of the salt-sensitive neuron reached
23 spikes/s (Fig. 2B,C, I).

19
4.3.2. Response to 100 mmol l-1 phosphate buff ers at pH 5.8–8.1 (11.1)
In most cases, phosphate buff ers were remarkably more stimulating for
the pH-sensitive neuron compared to acetate buff ers; at pH 5.8, the neu-
ron produced a mean of 8.1–10.5 spikes/s (Fig. 4A,B, A-cell, I). When
buff er pH was increased from 5.8 to 7, the fi rst second fi ring rate of the
pH-sensitive neuron increased by 50%, from 10.5 to 15.8 spikes/s. Even
larger increases in the response, reaching up to 300–800%, were observed
during next four seconds of the response after stimulus onset (Fig. 4A, A-
cell, I). In contrast, the response of the salt-sensitive neuron to these two
phosphate buff ers was approximately equal (Fig. 4A, B-cell, I). However,
the salt-sensitive neuron responded to the larger jumps in stimulus pH.
For example, when buff er pH was increased by three pH units, from 8.1
to 11.1, both of the neurons responded with signifi cant fi ring rate increase
whereby relative change in the response of the pH-sensitive neuron was
considerably larger compared to that of the salt-sensitive neuron, the fi r-
ing rate increased during the fi rst second of the response by 1350% and
25%, respectively (Fig. 4C, A- and B-cell, respectively, I).
4.3.3. Response to 100 mmol l-1 Na+-salts with and without addition of 10 mmol l-1 NaOH
Th e limitation of the buff er series method used for testing the responses
of the chemoreceptor neurons to stimulus pH is that buff ers with diff erent
pH vary largely in both pH and ionic concentrations of the component
salts. Th e method of alkalized salts reported here allows the preparation
of stimulating salt solutions over a wide range of pH whereas changes in
ionic concentrations of the component salts are minimal.
Remarkable diff erences between the pH- and salt-sensitive neurons re-
garding their responsiveness to the pH of the stimulating salt solutions
were demonstrated by electrophysiological experiments with alkalized
and non-alkalized salts. In the pH-sensitive neuron, rates of fi ring evoked
by the mixture of 100 mmol l-1 Na+-salts and 10 mmol l-1 NaOH were
signifi cantly higher compared to those caused by the salts alone (Fig. 3,
II). Th e stimulating eff ect of NaOH to the pH neuron was highest in the
mixture with NaCl (pH 9.6), fi ring rate during the fi rst second of the

20
response was 31 spikes/s and it did not fall below 17 spikes/s also during
the next several seconds recorded. Th e diff erence compared to the response
caused by 100 mmol l-1 NaCl alone (pH 7.6) was 10–22 times depend-
ing on the response time. Th e mixtures of 10 mmol l-1 NaOH with 100
mmol l-1 NaH2PO
4 (pH 5.8) and Na
2HPO
4 (pH 10.6) were 2–7 times
more eff ective stimuli for the pH-sensitive neuron compared to these non-
alkalized salts (pH 4.6 and 8.6, respectively). In contrast, the presence or
absence of 10 mmol l-1 NaOH in the stimulating salt solution had only
little or no eff ect on the spike production of the salt-sensitive neuron.
4.4. Plant sugar sensitivity in ground beetles
4.4.1. Electrophysiological identifi cation of the sugar-sensitive neuron in antennal taste sensilla of P. aethiops
In the ground beetle P. aethiops, by single sensillum tip recording tech-
nique, in addition to the salt- and pH-sensitive neurons, the third chemo-
receptor neuron of the antennal taste sensilla was electrophysiologically
identifi ed as a sugar-sensitive neuron. Th is neuron generated spikes of
considerably smaller amplitude than those of the salt- and pH-sensitive
neurons (Fig. 1B, III), and phasic-tonically responded to sucrose and glu-
cose over the range of 1–1000 mmol l-1 tested. Th e responses of the sugar-
sensitive neuron to stimulating solutions containing glucose or sucrose
were signifi cantly stronger compared to the control solution (p<0.05, t-test
for paired samples; Figs. 2A and 3A, III). Responses were concentration
dependent, with sucrose evoking more spikes than glucose. During the
fi rst second of the response, mean rates of fi ring of the sugar-sensitive
neuron reached up to 19 and 37 spikes/s when stimulated with 1000 mmol
l-1 glucose and sucrose, respectively. Th ereafter, the rates of fi ring quickly
fell below 5 spikes/s (sucrose), or close to zero (glucose).
In contrast, the responses of the salt-sensitive neuron were not signifi cantly
aff ected by the glucose content of the stimulating solutions (p>0.05, t-test
for paired samples; Fig. 2B, III). It was observed, that sucrose had a little
stimulating eff ect to the fi ring rate of the neuron, however (p<0.05, t-test
for paired samples; Fig. 3B, III). Th e responses of the pH-sensitive neuron
were also aff ected to a small degree by the content of sucrose and glucose
in the stimulating solutions, but in a more complicated manner. Firing
rate of the neuron increased to some degree at low concentrations (1 and

21
10 mmol 11) of the sugars (p<0.05, t-test for paired samples). At higher
concentrations (100 and 1000 mmol l-1) of the sugars the opposite was
observed, however: fi ring rates of the pH-sensitive neuron were slightly
suppressed by both glucose and sucrose compared to the responses evoked
by 10 mmol l-1 KCl (control) alone (p<0.05, t-test for paired samples;
Figs. 2 and 3, III).
4.4.2. Electrophysiological responses of the chemoreceptor neurons from antennal taste sensilla of P. oblongopunctatus to
plant sugars and amino acids
Choline chloride at a concentration of 10 mmol l-1 served as the control
stimulus, and as the electrolyte in stimulating mixtures with sugars and
amino acids to ensure a good signal-to-noise ratio. Typically, choline
chloride evoked only the salt-sensitive neuron to discharge with a relatively
low rate of fi ring (approximately 15 spikes/s) during the fi rst second after
stimulus onset (Fig. 3A, IV). Only in rare cases were a few spikes from
other sensory neurons of the sensillum observed (Fig. 1A; Table 1, IV).
Th erefore, the stimulating properties of choline chloride were suitable for
classifying and counting action potentials of the responses evoked by the
mixtures of this electrolyte with sugars and amino acids.
4.4.2.1. Response of antennal taste neurons to sugars
Responses of antennal taste neurons to stimulation by various 100 mmol
l-1 sugars in mixtures with 10 mmol l-1 choline chloride, varied with
the sugar tested. Only three out of twelve sugars, maltose, sucrose and
glucose, evoked strong phasic-tonic responses from the sugar-sensitive
neuron (Fig. 3B–E; Table 1, IV). Th e mean rates of fi ring of the neuron
during the fi rst second after 100 mmol l-1 stimulation reached 49.9, 46.5
and 20.9 spikes/s, respectively. Dose/response curves of these three active
sugars are demonstrated in Fig. 4 (IV). In the range of 1–1000 mmol l-1,
the response to glucose was approximately two times lower than that of
maltose and sucrose (p<0.05, ANOVA, Tukey test). In contrast, no dif-
ferences were found between responses to maltose and sucrose (p>0.05,
ANOVA, Tukey test). Th e remaining nine 100 mmol l-1 sugars had only a
small or no stimulatory eff ect on the neuron. To test for possible seasonal
variation in the functioning of the sugar-sensitive neuron, its responses

22
to 100 mmol l-1 sucrose were compared in active and hibernating beetles
in July (46.5 spikes/s) and February (48.8 spikes/s), respectively but no
diff erences between their responses were observed (t = -0.57; d.f. = 18;
p= 0.57; t-test for independent samples).
4.4.2.2. Response of antennal taste neurons to amino acids
Responses of antennal taste neurons to the seven 10 and 100 mmol l-1
L-amino acids in a mixture with 10 mmol l-1 choline chloride varied with
the type and concentration of amino acid tested. At 10 mmol l-1, none of
the amino acids stimulated the neurons. At 100 mmol l-1, these chemicals
evoked only a few spikes (Table 1, IV) with amplitudes smaller than those
generated by the salt-sensitive neuron (Fig. 3G–I, IV). No initial phasic
component in the sequence of these spikes was observed. Frequently,
they appeared with some delay in the responses, sometimes 5–10 s after
stimulus onset (Fig. 3J, IV). Spike amplitude analysis showed that these
small impulses were generated by the 4th chemosensory neuron rather
than by the sugar-sensitive neuron. Th ough amplitudes of spikes induced
by sugars (sucrose) and amino acids varied and overlapped to a large extent
their histograms had diff erent maxima (Fig. 5, IV), indicating that two
neurons may be involved. Since the mean numbers of these small spikes
during the fi rst second after stimulus onset did not exceed 3 spikes/s, this
neuron has probably no specialized receptor sites for amino acids, at least
for those which were tested.
4.5. Response of antennal taste neurons to plant alkaloids and glucosides
4.5.1. Electrophysiological identifi cation of the alkaloid-sensitive neuron in antennal taste sensilla of P. oblongopunctatus
Some tested alkaloids (quinine, caff eine) and an alkaloid salt (quinine
hydrochloride) induced a neuron of the taste sensilla to produce spikes
with negative polarity, but with a considerably longer period of latency
compared to the spikes of the salt- and sugar-sensitive neurons (Fig. 1B–E,
V). Spike amplitude histograms show two maxima, indicating that spikes
evoked by salt (NaCl), sugar (sucrose) and alkaloid (quinine hydrochloride)
were generated by three diff erent neurons, however, the salt-, sugar- and

23
alkaloid-sensitive neuron, respectively (Fig. 2, V). Spike amplitudes of
the chemoreceptor neurons from diff erent taste sensilla varied to a some
extent, from 0.7 to 2 mV (Fig. 1C,E, V). As a result, in most cases, spike
amplitudes generated by the alkaloid-sensitive neuron were shorter than
those of the salt-sensitive neuron (Fig. 1B,C, V). In some cases, however,
opposite spike amplitude ratios of the neurons were observed (Fig. 1D, V).
4.5.2. Stimulating eff ect of plant alkaloids and glucosides on the responses of the antennal chemoreceptor neurons
Only two of the tested chemicals (Table 1, V), 1 mmol l-1 quinine and 10
mmol l-1quinine hydrochloride, were highly stimulating for the antennal
alkaloid-sensitive neuron, evoking 8.9 and 22.8 spikes/s, respectively. Th e
stimulatory eff ect of 10 mmol l-1 caff eine was very weak, 2.4 spikes/s. In
contrast, a slight inhibiting eff ect of strychnine and sinigrin to the activity
of the neuron was observed. Respective fi ring rates were very low, how-
ever, below 2 spikes/s (Table 1, V). Other plant chemicals tested had little
eff ect on spike production of the alkaloid-sensitive neuron. Phasic-tonic
response of the alkaloid-sensitive neuron to quinine hydrochloride was
concentration dependent over the range of 0.001 to 50 mmol l-1 with the
response threshold at 0.01 mmol l-1, and maximum rate of fi ring equal to
67 spikes/s at 50 mmol l-1 (Figs. 3 and 4, V). However, compared to the
spike trains of the salt-sensitive neuron (Figs. 1 and 5A–F, V), the phasic
component of the response produced by the alkaloid-sensitive neuron was
less pronounced (Fig. 1B–E; Fig. 4; Fig. 5C–F, V).
4.5.3. Peripheral inhibition of the salt- and pH-sensitive neurons by some plant secondary compounds
In addition to stimulation of the alkaloid-sensitive neuron, both quinine
and quinine hydrochloride strongly inhibited spike generation by the salt-
and pH-sensitive neurons when presented in mixtures with 10 mmol l-1
NaCl (Table 1, V). Increasing the quinine hydrochloride concentration
while keeping the concentration of NaCl constant at 10 mmol l-1 caused
a dose-dependent decrease in spike generation by the salt-sensitive neuron
(Figs. 3–5, V). In most tested sensilla, at concentrations above 10 mmol
l-1 quinine hydrochloride, only the alkaloid-sensitive neuron fi red while
the fi ring rate of the salt-sensitive neuron of the taste sensillum decreased

24
close to zero during the 5 s adaptation period (Fig. 4, V). In response to
10 mmol l-1 NaCl alone, the mean number of spikes produced by the pH-
sensitive neuron was relatively low compared to that of the salt-sensitive
neuron, and varied to some extent in diff erent sensilla (Table 1, V). At
quinine hydrochloride concentrations above 0.01 mmol l-1, the mean fi ring
rate of the pH-sensitive neuron dropped to zero (Fig. 3, V). It was also
observed that nicotine and salicin, which did not aff ect the fi ring rate of the
alkaloid-sensitive neuron, greatly reduced the response of the pH-sensitive
neuron (Table 1, V). Sinigrin and caff eine suppressed neural activity of
the salt- and pH-sensitive neuron, respectively. Th is suggested that the
antennal taste sensilla of P. oblongopunctatus may detect plant defensive
compounds not only through the activation of specifi c alkaloid-sensitive
neuron but also through inhibition of the salt- and pH-sensitive neurons.
4.5.4. Eff ect of quinine hydrochloride on the feeding response of P. oblongopunctatus
Feeding choice experiments showed that one of the most stimulating
chemicals for the alkaloid-sensitive neuron, quinine hydrochloride, had a
concentration dependent deterrent eff ect on seed predation in the ground
beetle P. oblongopunctatus, with the threshold concentration at 1 mmol l-1.
Th us, the threshold concentration of this alkaloid to evoke a behavioural
response was a hundred times higher than that for an electrophysiological
response of the alkaloid-sensitive neuron. At higher concentrations, the
deterrent eff ect of quinine hydrochloride was very strong. It was observed
that pine seeds treated with 50 mmol l-1 quinine hydrochloride were never
predated by the beetles (Fig. 6, V).

25
5. DISCUSSION
5.1. pH sensitivity in ground beetles (I, II)
5.1.1. Th e occurrence of two chemoreceptor neurons in insect contact chemoreceptors responding to salts
In a number of insects, including the ground beetles P. oblongopunctatus (I) and P. aethiops (Merivee et al., 2004; II) frequently two neurons of
the taste sensilla respond to salt solutions (Dethier and Hanson, 1968;
Haskell and Schoonhoven, 1969; Den Otter, 1972; Mitchell and Schoon-
hoven, 1973; Mitchell and Seabrook, 1973; Blaney, 1974; Hanson, 1987;
Bernays and Chapman, 2001a; Schoonhoven and van Loon, 2002). It is
suggested that they function as a cation and an anion neuron (Ishikawa,
1963; Elizarov, 1978; Hanson, 1987; Schoonhoven and van Loon, 2002).
Temporal patterns of fi ring and the concentration/response curves are
quite diff erent for the two neurons. In the cation neuron with a phasic-
tonic type of reaction, the magnitude of the response is proportional to
the logarithm of salt concentration over the dynamic range (Evans and
Mellon, 1962; Hanson, 1987). It is believed that the stimulating eff ect
of salts to taste receptors is dominated by the cations involved, and that
monovalent cations are more eff ective stimuli than divalent cations (Evans
and Mellon, 1962; Schoonhoven and van Loon, 2002). Th is neuron type
corresponds to the salt-sensitive neuron of the antennal taste bristles in
Pterostichus (Merivee et al., 2004).
Specifi c stimuli for the second salt neuron are yet to be clarifi ed: it has
been named the ‘anion neuron’ on the basis of a supposed sensitivity to
salt anions or the ‘fi fth neuron’ (Ishikawa, 1963; Dethier and Hanson,
1968; Hanson, 1987; Liscia et al., 1997; Liscia and Solari, 2000). Th is
neuron is characteristically less responsive to diff erences in salt concentra-
tion (Dethier, 1977; Liscia et al., 1997), similar to that observed in the
pH-sensitive neuron of P. oblongopunctatus. In the strict sense of the term,
it is not correct to give the name ‘salt neuron’ to a neuron that does not
respond to changes in salt concentration. On the other hand, a study in
Grammia geneura shows increasing activity in two neurons in response
to potassium and sodium chlorides at concentrations ranging from 10 to
1000 mmol l-1 (Bernays and Chapman, 2001a). Dethier (1977) was the fi rst
to realize that it is not at all certain that the best stimulus for the second
salt neuron is indeed salt. Electrophysiological evidence is provided that

26
the so-called ‘fi fth’ neuron in taste chemosensilla of blowfl ies responds
to deterrent compounds, such as quinine, amiloride, nicotine and caf-
feine (Liscia and Solari, 2000). Very little attention has been paid to this
neuron. Th is is unfortunate because this lack of information handicaps
any attempt to construct an overview of the control of food detection
and habitat selection in insects on the basis of their chemosensory input.
5.1.2. Th e eff ect of stimulus pH to the salt- and pH-sensitive neurons in antennal taste sensilla of P. aethiops and P.
oblongopunctatus
Results of the electrophysiological experiments conducted with acid, neu-
tral and alkaline salts (Merivee et al., 2004), acetate and phosphate buff ers
(I), and alkalized and non-alkalized salts (II) demonstrated that, in ad-
dition to the salt-sensitive neuron, the existence of a pH-sensitive neuron
in antennal chaetoid taste sensilla of the ground beetles P. aethiops and P.
oblongopunctatus is possible. A strong relationship between stimulus pH
and spike production by the antennal pH-sensitive neuron was found:
signifi cantly higher rates of fi ring were recorded at higher pH values com-
pared to those observed at lower pH of the electrolyte solutions tested at
pH ranged from 3 to 11. However, the methods of various salts and buff ers
used to test the eff ect of pH on contact chemoreceptor neurons in ground
beetles have limitations in that these stimuli with diff erent pH vary largely
in both pH and ionic concentration of the component salts leaving the
exact stimulating role of these two factors for the neurons open (Merivee
et al., 2004; I). Th erefore, a method of alkalized (with NaOH) Na+ salts
was developed allowing preparation of stimulating solutions over a wide
range of pH with no diff erences in their ionic composition and minimal
diff erences in their salt concentration. Using this method for stimulation
of antennal contact chemoreceptors in P. aethiops (II), 2.5–10-fold increase
in the fi rst second fi ring rate of the pH-sensitive neuron in response to the
mixture of various 100 mmol l-1 salts and 10 mmol l-1 NaOH (pH was
observed (4–30 spikes/s) compared to that of 100 mmol l-1 NaCl alone
(1–8 spikes/s). Th us, the high stimulating eff ect of alkalized salts to the
pH-sensitive neuron could be attributed to pH alone as it was increased
by 1.3 to 2.0 pH units depending on the salt. In comparison, fi ring rate
of the salt-sensitive neuron changed by only 16–27%.

27
If the response of the pH-sensitive neuron depends on solution pH level only, there should be equivalent responses to buff ers at diff erent con-centrations at the same pH. However, the results do not confi rm this: 10 mmol l-1 phosphate buff er had a greater stimulating eff ect on the pH-sensitive neuron of P. oblongopunctatus than 100 mmol l-1 buff er (I). Th is phenomenon can be explained by interaction of diff erent chemical stimuli to chemosensory neurons. Other chemicals, as well as the pH of the stimulating solution, may infl uence the responses of insect taste receptors to one or another stimulating compound (Hanson, 1987; Liscia et al., 1997; Bernays et al., 1998; Liscia and Solari, 2000; Bernays and Chapman, 2001b; Schoonhoven and van Loon, 2002). Presumably, salts of stimulating buff er solutions at high concentration suppressed the neural activity of the pH-sensitive neuron when stimulated with 100 mmol l-1 phosphate buff er in 10 and 100 mmol l-1 phosphate buff er experiments in P. oblongopunctatus.
5.1.3. pH preference in ground beetles
Preferences for particular soil pH, varying between 3 and 9, in ground
beetles were discovered using laboratory choice experiments some decades
ago (Krogerus, 1960; Paje and Mossakowski, 1984). More recently, in
ecological
fi eld experiments, a correlation between the abundance of some ground
beetles and soil pH has been found (Irmler, 2001, 2003). Our results
suggest that in P. aethiops and P. oblongopunctus in their preferred acid
forest habitats and overwintering sites in brown-rotted wood at pH 3–5,
the antennal pH-sensitive neuron does not discharge or discharges at
very low frequency with the fi rst s fi ring rate close to 1 spike/s or lower.
Areas with a higher pH seem to be unfavourable to this insect and when
contacted the pH-sensitive neuron signals with a stronger response. Th is
may occur, for example, in places with decaying plant material on the soil
surface. Many of the materials composted contain signifi cant amounts
of protein, which is converted to ammonia toxic to many organisms and
serving as the primary substrate in the nitrifi cation processes. Nitrifi cation
processes have been observed at pH levels ranging from 6.6 to 9.7 (Odell
et al., 1996). Obviously, these ground beetles avoid places with a high pH.
Adults of P. aethiops and P. oblongopunctus may spend seven to eight
months, from September to April, in their preferred overwintering sites in

28
brown-rotted wood in Estonia. Fungi often lower the pH of the extracel-
lular medium through secretion of organic acids and thus establish a pH
gradient around the mycelium. Th e organic acid oxalate is produced by
most, if not all, wood-degrading fungi. Usually, a pH of 3–6 is observed
in both white- and brown-rotted wood. Stable moisture content is quaran-
teed as a result of metabolic water production during the wood-degrading
process. For cultures of the white-rot fungus Merulius tremellosus incubated
in air, the optimum water content was 2 g/1 g of wood. Moister conditions
were less favourable to delignifi cation, possibly because the water impeded
aeration (Reid, 1985; Varela and Tien, 2003). Similar prolonged stability
of water content and aeration is probably also quaranteed in brown-rotting
wood which seems to be a favourable microenvironment of ground bee-
tles for their prolonged hibernation. Brown-rotted wood may off er good
protection for hibernating ground beetles against entomopathogenic fungi
and parasites which play an important role in their population dynamic.
Up to 41% parasitism by nematodes and ectoparasitic fungi was found
on 14 species of Bembidion in Norway depending on habitat selection
of the host (Andersen and Skorping, 1991). Eggs of P. oblongopunctatus
suff ered 83% mortality in fresh litter but only 7% in sterilized soil (Hees-
sen, 1981). Although predators, parasites, and pathogens aff ect all ground
beetle developmental stages, quantitative data remain scarce. So, antennal
pH receptors of the ground beetles seem to be a powerful tool in their
selection for favourable acid habitats and microhabitats crucial to survival.
5.2. Sensitivity of ground beetles to plant sugars and amino acids (III, IV)
5.2.1. Response of the chemoreceptor neurons from antennal taste sensilla to sugars
Th ree out of the twelve sugars tested in P. aethiops and P. oblongopunctatus, maltose, sucrose and glucose, evoked strong responses from the sugar-sensitive neurons while the others had little or no eff ect. Most stimulating were the two disaccharides, maltose and sucrose (III, IV), and this is in agreement with results for a number of other insect species (Mitchell and Gregory, 1979; Schoonhoven and van Loon, 2002; Albert, 2003; Sandoval and Albert, 2007). Sucrose is a disaccharide consisting of two monosac-charides, α-glucose and β-fructose, joined by a glycosidic bond between carbon atom 1 of the glucose unit and carbon atom 2 of the fructose unit.

29
Maltose is composed of two molecules of glucose joined together through α-1.4-glycoside linkage. Th us, the presence of a terminal α-glucose unit seems to be a common and important feature of the molecular structure determining the stimulatory properties of these disaccharides in Pteros-tichus. Other monosaccharide units in the molecular structure of sucrose and maltose were of no importance in this respect as the sugar-sensitive neuron responses to these two disaccharides were similar. In contrast, the trisaccharide raffi nose has a non-terminal α-glucose unit and does not evoke a response from the antennal sugar-sensitive neuron in this species (IV). In the red turnip beetle, Entomoscelis americana, maltose (α-glucose-1.4-glucose) was at least an order of magnitude more eff ective in activating the maxillary sugar-sensitive neuron than trehalose (α-glucose-1.1-glucose) illustrating the importance of the 1.4 linkage in these disac-charide molecules (Mitchell and Gregory, 1979). Compared to maltose and sucrose, the third active sugar glucose was approximately two times less eff ective at stimulating the antennal sugar-sensitive neuron of both P. oblongopunctatus and P. aethiops (III, IV). Th is can be partly explained by the fact that glucose exists in at least two anomeric forms, α and β. Th ese two forms interconvert over a timescale of hours in aqueous solu-tion, to a fi nal stable ratio of α:β 36:64, in a process called mutarotation. So the physiologically active α-anomere forms only 36% of the glucose solution which is refl ected in its relatively low dose/response curve. Th e order of stimulatory eff ectiveness of sugars may vary in diff erent insect species or even be diff erent for receptors on various parts of the body (Blaney, 1974; Rüth, 1976; Bernays and Chapman, 2001; Schoonhoven and van Loon, 2002).
No insect so far investigated responds to all sugars, and no single sugar
is a major phagostimulant for all insects. For a given insect species, only
some sugars elicit a response to a greater or lesser extent while other sug-
ars elicit no response. Plant sugars which were found to evoke responses
from the antennal contact chemoreceptors in P. oblongopunctatus, i.e.,
maltose, sucrose and glucose (IV), function as strong phagostimulants
for most phytophagous insects, fl ies, cockroaches and others, equipped
with specialized receptors to detect sugars (Dethier et al., 1956; Blaney,
1974; Rüth, 1976; Amakawa, 2001; Liscia et al., 2002; Schoonhoven and
van Loon, 2002; Chapman, 2003; Sandoval and Albert, 2007). Th is sug-
gests that these sugars may have the same function in P. oblongopunctatus, probably due to its partial herbivory. Although, in contrast to sucrose and
glucose, maltose is uncommon in nature, it can be found in considerable

30
amounts in germinating seeds as a product of the enzymatic degradation
of starch. Obviously, high sensitivity of P. oblongopunctatus to maltose
is related to predation by the species on conifer seeds (Heikkilä, 1977;
Nystrand and Granström, 2000).
Several carbohydrates, in addition to glucose, such as cellobiose, arabinose,
xylose, mannose, rhamnose and galactose are known as components of
cellulose and hemicelluloses. Th ey are released by brown-rot fungi dur-
ing enzymatic wood decay. All water-soluble carbohydrates are ultimately
consumed, so that brown-rotted wood consists almost entirely of modifi ed
lignin. Brown-rot fungi appear to depolymerize wood in the early stages
of decay much more rapidly than the decay products can be metabolized.
Th e occurrence of excess wood decomposition products may help explain
the frequent presence of other wood scavengers in brown-rotted wood
(Eriksson et al., 1990; Zabel and Morrell, 1992; Valášková and Baldrian,
2006). As wood decomposes, it passes through a range of decay stages,
each colonized by a succession of diverse saproxylic insect assemblages
(Grove, 2002). Wood-decay products may serve as chemical cues for these
saproxylic insects and be involved in this colonization process although
no electrophysiological or behavioural evidence is yet available. Some
ground beetles, including P. oblongopunctatus, prefer brown-rotted wood
for their prolonged hibernation (Haberman, 1968; Lindroth, 1986). Our
results showed, however, that P. oblongopunctatus was not able to sense
water-soluble wood sugars released by brown-rot fungi, except for glucose
(IV), indicating that these chemicals are not involved in the search for
hibernation sites by this species, although others may be.
5.2.2. Response of the chemoreceptor neurons from antennal taste sensilla to amino acids
Since amino acids, in addition to sugars, stimulate feeding behaviour in various phytophagous insects these animals have amino acid-specifi c neurons in their taste sensilla. Amino acids and sugars may stimulate the same neuron. Such a versatile receptor neuron is present in the polypha-gous caterpillar of Grammia geneura. Th ese insects have, in addition to an amino acid sensitive neuron in their lateral sensillum styloconicum, a neuron in another sensillum which responds to seven (out of twenty) amino acids. Th is latter neuron can also be stimulated by sucrose, glucose, and trehalose (Schoonhoven and van Loon, 2002; Chapman, 2003). Because

31
of its multiple specifi city this neuron was named a ‘‘phagostimulatory neuron’’, rather than a ‘‘sugar’’ or ‘‘amino acid’’ neuron (Bernays and Chapman, 2001). However, in P. oblongopunctatus, the seven amino acids tested (IV) had only little or no stimulatory eff ect on the neurons of the antennal taste sensilla suggesting that these neurons have no ability to perceive these amino acids. Th e weakly stimulatory eff ect (below 3 spikes/s) of some 100 mmol l-1 amino acids on the 4th chemosensory neuron of these sensilla can be characterized as non-specifi c, or modulating to the responses of non-target chemosensory neurons, the phenomenon well known in insect chemoreceptor functioning (Hanson, 1987; Liscia et al., 1997; Liscia and Solari, 2000; Bernays and Chapman, 2001; Chapman, 2003; Schoonhoven and van Loon, 2002; I; III).
5.3. Response of the chemoreceptor neurons in the antennal taste sensilla of P. oblongopunctatus to plant secondary compounds
5.3.1 Stimulating eff ect of plant alkaloids and glucosides on the responses of the antennal chemoreceptor neurons
Th e experimental data presented show that the “fourth” chemoreceptor
neuron in the antennal taste sensilla of the ground beetle P. oblongopunc-tatus (IV) may respond to some plant alkaloids and alkaloid salts such
as quinine and quinine hydrochloride (V). Th is fi nding seems to be in
agreement with literature data on the feeding habits of this species. P. oblongopunctatus is one of the most common and widespread ground
beetles in the forests of northern Europe (Lindroth, 1985; Niemelä et al.,
1994). In most stands of Pinus sylvestris, seed predation results in 20% seed
mortality, although occasionally it reaches 60%. Post-dispersal pine seed
predation in the Swedish boreal forests studied was due mainly to the two
ground beetle species P. oblongopunctatus and Calathus micropterus. Both
are able to consume conifer seeds. Th e larger species, P. oblongopunctatus, is considered to be the more important pine seed predator of the two
(Heikkilä, 1977; Nystrand and Granström, 2000).
Like many herbivores, numerous insect seed-predators have biochemical
adaptations to deal with toxic plant secondary compounds, including vari-
ous detoxifi cation and sequestration mechanisms (Jolivet, 1998; Hulme
and Benkman, 2004; Schoonhoven et al., 2005; Després et al., 2007).
However, the diffi culties of dealing with more than a few diff erent kinds

32
of defensive compounds has favoured specialization in seed-predators
and helps to explain why a large proportion of them are specialist feed-
ers (Hulme and Benkman, 2004). Th e omnivorous ground beetle P. oblongopunctatus predates on many small animals, most frequently on
Coleoptera, Diptera, Collembola, Acari etc. (Larochelle, 1990), as well
as on conifer seeds (Heikkilä, 1977; Nystrand and Granström, 2000).
Probably, the species feeds on seeds of various plants but respective data
are not available. Selective seed predation has been observed in many other
ground beetles, however (Toft and Bilde, 2002; Tooley and Brust, 2002).
To discriminate between toxic and nontoxic seeds, P. oblongopunctatus
possesses a specifi c feeding deterrent neuron innervating its antennal
chaetoid taste sensilla (V). A strong stimulatory eff ect of quinine (qui-
noline group) and quinine hydrochloride to this neuron was observed.
Th e threshold concentration for the latter at 0.01 mmol l-1 was close to
the upper limit of the threshold concentrations for the plant secondary
compounds that stimulate a deterrent neuron in other herbivorous insects
(Schoonhoven and van Loon, 2002; Chapman, 2003; Schoonhoven et al.,
2005), probably due to the fact that toxin concentration in seeds is usually
much higher than in other parts of the plant (Janzen, 1971; Hulme and
Benkman, 2004; Davis et al., 2008). Th e response of the neuron to qui-
nine hydrochloride was concentration dependent with maximum rates of
fi ring reaching up to 70 spikes/s at 50 mmol l-1. In two-choice behavioural
experiments, quinine hydrochloride had a strong concentration dependent
deterrent eff ect on feeding of P. oblongopunctatus beetles with the response
threshold at 1 mmol l-1. Th us, the threshold concentration of this alkaloid
to evoke a behavioural response was a hundred times higher than that
for an electrophysiological response of the alkaloid-sensitive neuron. In
contrast to quinine hydrochloride, the response of the alkaloid-sensitive
neuron to caff eine (purine group) was very weak, only 2.4 spikes/s at 10
mmol l-1 concentration. Th e eff ect of another quinoline alkaloid, strych-
nine, and sinigrin, to the neuron was rather inhibitory. No response of
the neuron to tested pyrrolidine alkaloid, nicotine, and salicin (glucoside)
was observed suggesting that the antennal alkaloid-sensitive neuron of P.
oblongopunctatus is not equally sensitive to all plant defensive chemicals.
Nevertheless, the list of toxic compounds capable of stimulating the an-
tennal alkaloid-sensitive neuron of P. oblongopunctatus is likely to grow
longer as more plant defence chemicals are tested (V).

33
Quinoline alkaloids are among the most potent insect feeding deter-
rents occurring mainly in Rutaceous and Rubiaceous plants. Th ey can be
present in virtually every part of the plant or just a specifi c part like the
bark, rhizome, leaf or seed (Openshaw, 1967; Michael, 2003; Aniszewski,
2007). Th ese alkaloids represent a large and structurally diverse group
of plant defensive compounds. When contacted, some of them may be
detected by stimulation of a specifi c antennal alkaloid-sensitive neuron of
the ground beetles predating on seeds, similarly to the quinine-sensitivity
of P. oblongopunctatus. Th e ability of granivores to perceive plant defensive
compounds allows them to avoid toxic seeds, and survive. High sensitiv-
ity of P. oblongopunctatus to plant sugars, commonly known as strong
phagostimulants for most phytophagous insects, seems to be also related
to its partially granivorous feeding habits. Th e disaccharide maltose was
one of the most stimulatory sugars for the sugar-sensitive neuron also
innervating antennal taste sensilla of P. oblongopunctatus (IV).
5.3.2. Peripheral inhibition of the salt- and pH-sensitive neurons by some plant secondary compounds
While stimulating the alkaloid-sensitive neuron, quinine hydrochloride strongly reduced spike production by the salt- and pH-sensitive neurons which also innervate these sensilla in Pterostichus spp. (Merivee et al., 2004; I, II). Increasing the quinine hydrochloride concentration, while keeping the concentration of NaCl constant, caused a dose-dependent decrease in spike generation by the salt-sensitive neuron. At 10 mmol l-1 and higher concentrations of quinine hydrochloride, predominant fi ring of the alkaloid-sensitive neuron was observed while the fi ring rate of other neu-rons dropped almost to zero. It was also observed that several tested plant secondary compounds, salicin, sinigrin, caff eine and nicotine, which had only little or no eff ect on the fi ring rate of the alkaloid-sensitive neuron, would greatly reduce the responses of the salt- and pH-sensitive neurons of the antennal taste sensillum of P. oblongopunctatus. Th ese results suggest that the antennal taste sensilla in P. oblongopunctatus may detect toxic and deterrent plant secondary compounds not only through activation of the specifi c alkaloid-sensitive neuron but also through inhibition of taste neurons activated by salts and stimulus pH; this is similar to deterrent inhibition of taste neurons activated by sugars and water in Drosophila (Meunier et al., 2003). Since foodplant acceptability to herbivores depends on the balance between stimulation and deterrency, similar interactions

34
between stimulating chemicals at peripheral taste neurons of herbivorous insects are of widespread occurrence. Plant toxins and deterrents may inhibit sugar neurons (Frazier, 1986; van Loon, 1990; Messchendorp et al., 1996; Bernays and Chapman, 2000; Schoonhoven and van Loon, 2002; Meunier et al., 2003; Schoonhoven et al., 2005), water neurons (Meunier et al., 2003), and sugars and salts may inhibit deterrent neurons (Simmonds and Blaney, 1983; Shields and Mitchell, 1995; Glendinning et al., 2000; Schoonhoven and van Loon, 2002; Schoonhoven et al., 2005). Deterrent compounds that on their own do not stimulate any neuron within a sensillum may also decrease the responsiveness of a neuron re-sponding to a nutrient, as exemplifi ed by sinigrin inhibiting the inositol neuron in Heliothis virescens and H. subfl exa (Bernays and Chapman, 2000; Schoonhoven and van Loon, 2002).
Survival of most terrestrial animals requires ability to detect environmental
NaCl, a nutrient essential for fl uid and electrolyte homeostasis and for a
multitude of other physiological processes (Lindemann, 1996; Contreras
and Lundy, 2000). Th e gustatory system allows insects to detect and ingest
salt, to discriminate between diff erent salts, and to avoid high salt concen-
trations. However, there are only few data that relate input from inorganic
salts to feeding behaviour in phytophagous insects. In some insects, salts
have a deterrent eff ect on feeding (Chapman, 2003; Schoonhoven and
van Loon, 2002). In contrast, larvae of Drosophila show dose-dependent
responses to NaCl: the appetitive responses to low concentration gradu-
ally turn into aversion as concentration is increased up to 200 mmol l-1
and higher (Miyakawa, 1982; Liu et al., 2003; Niewalda et al., 2008).
Some other insects also respond positively to dilute salts. Cockroaches
and housefl ies prefer dilute sodium and potassium chlorides and refuse
distilled water. In the two-choice situation, many animals give a bimodal
response to sodium chloride. Acceptance at low concentrations switches to
rejection at high concentrations (Dethier, 1977). In these cases, inorganic
salt at low concentration may act as a phagostimulant. Th erefore, NaCl
at 10 mmol l-1 may also have a phagostimulatory eff ect in P. oblongopunc-tatus although no behavioural data is available. Th is assumption could
explain the inhibition of spike production of the salt-sensitive neuron
by some toxic compounds in P. oblongopunctatus (V) as being similar to
how plant deterrents inhibit activity of other phagostimulatory neurons
in other phytophagous insects.

35
6. CONCLUSIONS
1. For the fi rst time, a specifi c pH-sensitive chemoreceptor neuron was
identifi ed in insect taste sensilla, most probably related to habitat and
microhabitat selection of the species (P. aethiops and P. oblongopunc-tatus; I, II);
2. For the fi rst time, a sugar-sensitive chemoreceptor neuron responding
to 1–1000 mmol l-1 sucrose and glucose was identifi ed in the taste
sensilla of ground beetles. Th e phasic-tonic response of the neuron
to these sugars was concentration dependent (P. aethiops, III);
3. Th e disacharides with an a-1.4-glycoside linkage, sucrose and maltose,
were the two most stimulatory sugars out of 12 tested for the antennal
sugar-sensitive neuron of P. oblongopunctatus evoking 70 spikes/s at
1000 mmol l-1. Th e stimulatory eff ect of glucose was approximately
two times lower. Due to the partial herbivory of P. oblongopunctatus
these plant sugars are probably involved in its search for food, for
example, for conifer seeds (IV);
4. None of the water-soluble sugars released by brown-rot fungi during
enzymatic wood decay (cellobiose, arabinose, xylose, mannose, rham-
nose and galactose) stimulated the antennal sugar-sensitive neuron in
P. oblongopunctatus (IV);
5. Th e weak stimulating eff ect (below 3 spikes/s) of some 100 mmol
l-1 amino acids (methionine, serine, alanine, glutamine) to the 4th
chemosensory neuron of the antennal taste sensilla of P. oblongopunc-tatus was characterized as non-specifi c, or modulating the responses
of non-target chemosensory neurons (IV);
6. For the fi rst time, the occurrence of an alkaloid-sensitive feeding
deterrent neuron in the antennal taste sensilla of ground beetles (P.
oblongopunctatus) was documented (V);
7. Some tested compounds (quinine and quinine hydrochloride) that
stimulate the alkaloid-sensitive neuron in P. oblongopunctatus may
strongly inhibit activity of other neurons (salt- and pH-sensitive neu-
ron) of the sensillum (V);

36
8. Several tested plant secondary compounds, salicin, sinigrin, caff eine
and nicotine, which had only little or no eff ect on the fi ring rate of
the alkaloid-sensitive neuron, through peripheral inhibition, would
greatly reduce the responses of other chemoreceptor neurons of the
sensillum in P. oblongopunctatus (V).

37
REFERENCES
Albert, P.J., 2003. Electrophysiological responses to sucrose from a gusta-
tory sensillum on the larval maxillary palp of the spruce budworm,
Choristoneura fumiferana (Clem.) (Lepidoptera: Tortricidae). Journal
of Insect Physiology 49, 733–738.
Amakawa, T., 2001. Eff ects of age and blood sugar levels on the proboscis
extension of the blow fl y Phormia regina. Journal of Insect Physiology
47, 195–203.
Andersen, J., Skorping, A., 1991. Parasites of carabid beetles: prevalence
depends on habitat selection of the host. Canadian Journal of Zool-
ogy 69, 1216–1220.
Anizewski, T., 2007. Alkaloids–secrets of life. Alkaloid chemistry, biologi-
cal signifi cance, applications and ecological role. Elsevier, Amsterdam,
Boston, Heidelberg, London, New York, Oxford, Paris, San Diego,
San Francisco, Singapore, Sydney, Tokyo, P 334.
Bernays, E.A., Chapman, R.F., 2000. A neurophysiological study of sen-
sitivity to a feeding deterrent in two sister species of Heliothis with
diff erent diet breadths. Journal of Insect Physiology 46, 905–912.
Bernays, E.A., Chapman, R.F., 2001a. Electrophysiological responses
of taste cells to nutrient mixtures in the polyphagous caterpillar of
Grammia geneura. Journal of Comparative Physiology A 187, 205–213.
Bernays, E.A., Chapman, R.F., 2001b. Taste cell responses in the poly-
phagous arctiid, Grammia geneura: towards a general pattern for cat-
erpillars. Journal of Insect Physiology 47, 1029–1043.
Bernays, E.A., Glendinning, J.I., Chapman, R.F., 1998. Plant acids modu-
late chemosensory responses in Manduca sexta larvae. Physiological
Entomology 23, 193–201
Blake, S., McCracken, D.I., Eyre, M.D., Garside, A., Foster, G.N., 2003.
Th e relationship between the classifi cation of Scottish ground bee-
tle assemblages (Coleoptera, Carabidae) and the national vegetation
classifi cation of British plant communities. Ecography 26, 602–616.
Blaney, W.M., 1974. Electrophysiological responses of the terminal sensilla
on the maxillary palps of Locusta migratoria L. to some electrolytes
and non-electrolytes. Journal of Experimental Biology 60, 275–293.
Chapman, R.F., 1998. Th e Insects. Structure and Function. 4th edn.
Cambridge University Press, Cambridge, P 770.
Chapman, R.F., 2003. Contact chemoreception in feeding by phytopha-
gous insects. Annual Review of Entomology 48, 455–484.

38
Cole, L.J., McCracken, D.I., Dennis, P., Downie, I.S., Griffi n, A.L., Foster,
G.N., Murphy, K.J., Waterhouse, T., 2002. Relationships between
agricultural management and ecological groups of ground beetles
(Coleoptera: Carabidae) on Scotish farmland. Agriculture, Ecosystems
and Environment 93, 323–336.
Contreras, R.J., Lundy, R.F., 2000. Gustatory neuron types in the pe-
riphery: a functional perspective. Physiology and Behavior 69, 41–52.
Daly, P.J., Ryan, M.F., 1979. Ultrastructure of antennal sensilla of Nebria brevicollis (Fab.) (Coleoptera: Carabidae). International Journal of
Insect Morphology and Embryology 8, 169–181.
Davis, A.S., Schutte, B.J., Iannuzzi, J., Renner, K. A., 2008. Chemical and
physical defense of weed seeds in relation to soil seedbank persistence.
Weed Science 56, 676–684.
Den Otter, C.J., 1972. Diff erential sensitivity of insect chemoreceptors
to alkali cations. Journal of Insect Physiology 18, 109–131.
Després, L., David, J.-P., Gallet, C., 2007. Th e evolutionary ecology of
insect resistance to plant chemicals. Trends in Ecology and Evolution
22, 298–307.
Dethier, V.G., 1977. Th e taste of salt. American Science 65, 744–751.
Dethier, V.G., 1980. Evolution of receptor sensitivity to secondary plant
substances with special reference to deterrents. American Naturalist
115, 45–66.
Dethier, V.G., Evans, D.R., Rhoades, M.V., 1956. Some factors control-
ling the ingestion of carbohydrates by the blowfl y. Biological Bulletin
111, 204–222.
Dethier, V.G., Hanson, F.E., 1968. Electrophysiological responses of the
blowfl y to sodium salts of fatty acids. Proceedings of the National
Academy of Sciences (U.S.A.) 60, 1269–1303.
Elizarov, Y.A., 1978. Insect Chemoreception. Moscow University Press,
Russia (in Russian).
Eriksson, K.-E.L., Blanchette, R.A., Ander, P., 1990. Microbial and Enzy-
matic Degradation of Wood Components. Springer-Verlag, New York.
Evans, D.R., Mellon, D., 1962. Stimulation of a primary taste receptor
by salts. Journal of General Physiology 45, 651–661.
Evans, W.G., 1982. Oscillatoria sp. (Cyanophyta) mat metabolites im-
plicated in habitat selection in Bembidion obtusidens (Coleopt.: Car-
abidae). Journal of Chemical Ecology 8, 671–678.
Evans, W.G., 1983. Habitat selection in the Carabidae. Th e Coleopterists’
Bulletin 37, 164–167.

39
Evans, W.G., 1984. Odour-mediated responses of Bembidion obtusidens (Coleoptera: Carabidae) in a wind tunnel. Canadian Journal of En-tomology 116, 1653–1658.
Evans, W.G., 1988. Chemically mediated habitat recognition in shore insects (Coleoptera: Carabidae, Hemiptera: Saldidae). Journal of Chemical Ecology 14, 1441–1454.
Eyre, M.D., Luff , M.L., 2004. Ground beetle species (Coleoptera, Car-abidae) associations with land cover variables in northern England and southern Scotland. Ecography 27, 417–426.
Frazier, J.L., 1986. Th e perception of plant allelochemicals that inhibit feeding. In: Brattsten, L.B., Ahmad, S. (Eds.), Molecular aspects of insect-plant associations. Plenum Press, New York, pp. 1–42
Glendinning, J.I., 2002. How do herbivorous insects cope with noxious secondary plant compounds in their diet? Entomologia Experimentalis et Applicata 104,15–25.
Glendinning, J.I., Nelson, N., Bernays, E.A., 2000. How do inositol and glucose modulate feeding in Manduca sexta caterpillars? Journal of Experimental Biology 203, 1299–1315.
Grove, S.J., 2002. Saproxylic insect ecology and the sustainable man-agement of forests. Th e Annual Review of Ecology, Evolution, and Systematics 33, 1–23.
Haberman, H., 1968. Ground beetles of Estonia. Valgus, Tallinn, P 589 (in Estonian).
Hansen-Delkeskamp, E., 1998. Development of specifi c responses in antennal taste hairs after ecdysis. An electrophysiological investigation of the cockroach, Periplaneta brunnea. Journal of Insect Physiology 44, 659–666.
Hanson, F.E., 1987. Chemoreception in the fl y: the search for the liver-wurst receptor. In: Chapman, R.F., Bernays, E.A., Stoff olano, J.G. (Eds.), Perspectives in Chemoreception and Behaviour. Springer, Germany, pp. 99–122.
Harborne, J.B., 1993. Introduction to Ecological Biochemistry. 4th edn. Academic Press, London, San Diego, P. 318.
Haskell, P.T., Schoonhoven, L.M., 1969. Th e function of certain mouth part receptors in relation to feeding in Schistocerca gregaria and Locusta migratoria migratorioides. Entomologia Experimentalis et Applicata 12, 429–440.
Hatzold, T., Elmadfa, I., Gross, R., Wink, M., Hartmann, T., Witte, L., 1983. Quinolizidine alkaloids in seeds of Lupinus mutabilis. Journal of Agricultural and Food Chemistry 31, 934–930.

40
Heessen, H.J.L., 1981. Egg mortality in P. oblongopunctatus (Coleoptera, Carabidae). Oecologia, 50, 233–235.
Heikkilä, R., 1977. Eläimet kylvetyn männyn ja kuusen siemenen tuhooji-na Pohjois-Suomessa (Summary: Destruction caused by animals to sown pine and spruce seed in northern Finland). Metsäntutkimus-laitoksen Julkaisuja 89, 1–35.
Hengeveld, R., 1980. Polyphagy, oligophagy and food specialization in ground beetles (Coleoptera, Carabidae). Netherlands Journal of Zool-ogy 30, 564–584.
Hoback, W.W., Golick, D.A., Svatos, T.M., Spomer, S.M., Higley, L.G., 2000. Salinity and shade preferences result in ovipositional diff er-ences between sympatric tiger beetle species. Ecological Entomology 25, 180–187.
Hodgson, E.S., Lettvin, J.Y., Roeder, K.D., 1955. Physiology of a primary chemoreceptor unit. Science 122, 417–418.
Honek, A., Martinkova, Z., Jarosik, V., 2003. Ground beetles (Carabidae) as seed predators. European Journal of Entomology 100, 531–544.
Hulme, P.E., Benkman, C.W., 2004. Granivory. In: Herrera, C.M., Pell-myr O. (Eds.), Plant-animal interactions. An evolutionary approach. Blackwell Science, Oxford, pp. 132–154
Ings, T.C., Hartley, S.E., 1999. Th e eff ect of habitat structure on carabid communities during the regeneration of a native Scotish forest. Forest Ecology and Management 119, 123–136.
Irmler, U., 2001. Charakterisierung der Laufkäfergemeinschaften schleswigholsteinischer Wälder und Möglichkeiten ihrer ökologischen Bewertung. Angewandte Carabidologie, Supplement 2. Laufkäfer im Wald, 21–32.
Irmler, U., 2003. Th e spatial and temporal pattern of carabid beetles on arable fi elds in northern Germany (Schleswig–Holstein) and their value as ecological indicators. Agriculture, Ecosystems and Environ-ment 98, 141–151.
Ishikawa, S., 1963. Responses of maxillary chemoreceptors in the larvae of the silkworm, Bombyx mori, to stimulation by carbohydrates. Journal of Cellular and Comparative Physiology 61, 99–107.
Ishikawa, S., 1966. Electrical response and function of bitter substance receptor associated with the maxillary sensilla of the larva of the silk-worm, Bombyx mori L. Journal of Cellular and Comparative Physiol-ogy 67, 1–12.
Janzen, D.H., 1971. Seed predation by animals. Annual Review of Ecol-
ogy, Evolution, and Systematics 2, 465–492.

41
Johnson, U.E., Cameron, R.S., 1969. Phytophagous ground beetles. An-nals of the Entomological Society of America 62, 909–914
Jolivet, P., 1998. Interrelationships between insects and plants. CRC Press, Boca Raton, Florida, P. 309.
Jørgensen, K., Almaas, T.J., Marion-Poll, F., Mustaparta, H., 2007. Elec-trophysiological characterization of responses from gustatory receptor neurons of sensilla chaetica in the moth Heliothis virescens. Chemical Senses 32, 863–879.
Jørgensen, K., Kvello, P., Almaas, T.J., Mustaparta, H., 2006. Two closely located areas in the suboesophageal ganglion and the tritocerebrum receive projections of gustatory receptor neurones located on the an-tennae and the proboscis in the moth Heliothis virescens. Th e Journal of Comparative Neurology 496, 121–134.
Kielty, J.P., Allen-Williams, L.J., Underwood, N., Eastwood, E.A., 1996. Behavioural responses of three species of ground beetle (Coleoptera: Carabidae) to olfactory cues associated with prey and habitat. Journal of Insect Physiology 9, 237–250.
Kim, J.L., Yamasaki, T., 1996. Sensilla of Carabus (Isiocarabus) fi duciarius saishutoicus Csiki (Coleoptera: Carabidae). International Journal of Insect Morphology and Embryology, 25, 153–172.
Krogerus, R., 1960. Ökologische Studien über nordische Moorarthro-poden. Societas Scientiarum. Fennica, Commentationes Biologicae, 21 (3), 1–238.
Larochelle, A., 1990. Th e food of the carabid beetles (Coleoptera: Carabi-dae, including Cicindelinae). Fabreries, Supplément 5, 1–132.
Lindemann, B., 1996. Taste reception. Physiological Reviews 76, 719–766.Lindroth, C.H., 1985. Th e Carabidae (Coleoptera) of Fennoscandia and
Denmark. Fauna Entomologica Scandinavica 15, 1–225.Lindroth, C.H., 1986. Th e Carabidae (Coleoptera) of Fennoscandia and
Denmark. Fauna Entomologica Scandinavica 15, 233–497.Liscia, A., Crnjar, R., Masala, C., Sollai, G., Solari, P., 2002. Sugar re-
ception in the blowfl y: a possible Ca++ involvement. Journal of Insect Physiology 48, 693–699.
Liscia, A., Solari, P., 2000. Bitter taste recognition in the blowfl y: elec-trophysiological and behavioral evidence. Physiology and Behaviour 70, 61–65.
Liscia, A., Solari, P., Majone, R., Tomassini Barbarossa, I., Crnjar, R., 1997. Taste receptor mechanisms in the blowfl y: evidence of amiloride-sensitive and insensitive receptor sites. Physiology and Behaviour 62, 875–879.

42
Liu, L., Leonard, A.S., Motto, D.G., Feller, M.A., Price, M.P., Johnson, W.A., Welsh, M.J. 2003. Contribution of Drosophila DEG/ENaC
Genes to Salt Taste. Neuron 39, 133–146.
Lövei, G.L., Sunderland, K.D., 1996. Ecology and behavior of ground
beetles (Coleoptera: Carabidae). Annual Review of Entomology 41,
231–256.
Magura, T., 2002. Carabids and forest edge: spatial pattern and edge
eff ect. Forest Ecology and Management 157, 23–37.
Merivee, E., Ploomi, A., Luik, A., Rahi, M., Sammelselg, V., 2001. An-
tennal sensilla of the ground beetle Platynus dorsalis (Pontoppidan,
1763) (Coleoptera, Carabidae). Microscopy Research and Technique
55, 339–349.
Merivee, E., Ploomi, A., Rahi, M., Bresciani, J., Ravn, H. P., Luik, A.,
Sammelselg, V., 2002. Antennal sensilla of the ground beetle Bembid-ion properans Steph. (Coleoptera, Carabidae). Micron 33, 429–440.
Merivee, E., Ploomi, A., Rahi, M., Luik, A., Sammelselg, V., 2000. Anten-
nal sensilla of the ground beetle Bembidion lampros Hbst (Coleoptera,
Carabidae). Acta Zoologica (Stockholm) 81, 339–350.
Merivee, E., Renou, M., Mänd, M., Luik, A., Heidemaa, M., Ploomi A.,
2004. Electrophysiological responses to salts from antennal chaetoid
taste sensilla of the ground beetle Pterostichus aethiops. Journal of
Insect Physiology 50, 1001–1013.
Messchendorp, L., van Loon, J.J.A., Gols, G.J.Z., 1996. Behavioural and
sensory responses to drimane antifeedants in Pieris brassicae larvae.
Entomologia Experimentalis et Applicata 79, 195–202.
Meunier, N., Marion-Poll, F., Rospars, J.-P., Tanimura, T., 2003. Periph-
eral coding of bitter taste in Drosophila. Journal of Neurobiology 56,
139–152.
Michael, J.P., 2003. Quinoline, quinazoline and acridone alkaloids. Natu-
ral Product Reports Articles 20, 476–493.
Mitchell, B.K., Gregory, P., 1979. Physiology of the maxillary sugar sensi-
tive cell in the red turnip beetle, Entomoscelis americana. Journal of
Comparative Physiology 132, 167–178.
Mitchell, B.K., Schoonhoven, L.M., 1973. Taste receptors in Colorado
beetle larvae. Journal of Insect Physiology 20, 1787–1793.
Mitchell, B.K., Seabrook, W.D., 1973. Electrophysiological investigations
on tarsal chemoreceptors of the spruce budworm, Choristoneura fumif-erana (Lepidoptera). Journal of Insect Physiology, 20, 1209–1218.

43
Miyakawa, Y., 1982. Behavioural evidence for the existence of sugar, salt
and amino acid taste receptor cells and some of their properties in
Drosophila larvae. Journal of Insect Physiology 28, 405–410.
Morita, H., Shiraishi, A., 1985. Chemoreception physiology. In: Kerkut,
G.A., Gilbert, L.I. (Eds.), Comprehensive Insect Physiology, Bio-
chemistry and Pharmacology. Pergamon Press, Oxford, pp. 133–170.
Niemelä, J., Spence, J. R., Langor, D., Haila, Y., Tukia, H., 1994. Logging
and boreal ground-beetle assemblages on two continents: implica-
tions for conservation. In: Gaston, K., Samways, M., New, T. (Eds.),
Perspectives in insect conservation. Intercept Publications, Andover,
pp. 29–50
Niewalda, T., Singhal, N., Fiala, A., Saumweber, T., Wegener S., Gerber,
B., 2008. Salt Processing in Larval Drosophila: Choice, Feeding,
and Learning Shift from Appetitive to Aversive in a Concentration-
Dependent Way. Chemical Senses 33, 685–692.
Nystrand, O., Granström, A., 2000. Predation on Pinus sylvestris seeds
and juvenile seedlings in Swedish boreal forests in relation to stand
disturbance by logging. Journal of Applied Ecology 37, 449–463.
Odell, L.H., Kirmeyer, G.J., Wilczak, A., Jasangelo, J.G., Marcinko, J.P.,
Wolfe, R.L., 1996. Controlling nitrifi cation in chloraminated systems.
Journal AWWA 88, 86–98.
Openshaw, H.T., 1967. Quinoline alkaloids other than those of Cinchona.
In Manske R.H.F. (Ed.), Th e Alkaloids. Chemistry and Physiology.
Vol. IX. Academic Press, New York, London, pp. 223–267.
Paje, F., Mossakowski, D., 1984. pH-preferences and habitat selection in
carabid beetles. Oecologia (Berlin) 64, 41–46.
Purtauf, T., Roschewitz, I., Dauber, J., Th ies, C., Tscharntke, T., Wolters,
V., 2005. Landscape context of organic and conventional farms: infl u-
ences on carabid beetle diversity. Agriculture, Ecosystems and Envi-
ronment 108, 165–174.
Reid, I.D., 1985. Biological delignifi cation of aspen wood by solid-state
fermentation with the white-rot fungus Merulius tremellosus. Applied
and Environmental Microbiology 50, 133–139.
Rüth, E., 1976. Elektrophysiologie der Sensilla Chaetica auf den Anten-
nen von Periplaneta americana. Journal of Comparative Physiology
105, 55–64.
Ryan, M.F., 2002. Insect Chemoreception. Fundamental and Applied.
Kluwer Academic Publishers, New York, Boston, Dordrecht, London,
Moscow, P. 330.

44
Sandoval, M., Albert, P.J., 2007. Chemoreception of sucrose and amino
acids in second and fourth instars of the spruce budworm Choristo-
neura fumiferana (Clem.) (Lepidoptera: Tortricidae). Journal of Insect
Physiology 53, 84–92.
Schoonhoven, L.M., van Loon, J.J.A., 2002. An inventory of taste in
caterpillars: each species its own key. Acta Zoologica Academiae Sci-
entiarum Hungaricae 48, 215–263.
Schoonhoven, L.M., van Loon, J.J.A., Dicke, M., 2005. Insect-Plant
Biology. 2nd edn.. Oxford University Press, Oxford, P. 421
Shields, V.D.C., Mitchell, B.K., 1995. Responses of maxillary stylocon-
ic receptors to stimulation by sinigrin, sucrose and inositol in two
crucifer-feeding, polyphagous lepidopterous species. Philosophical
Transactions of the Royal Society London 347, 447–457.
Silfverberg, H., 2004. Enumeratio nova Coleopterorum Fennoscandiae,
Daniae et Baltiae. Sahlbergia 9, 1–111.
Simmonds, M.S.J., Blaney, W.M., 1983. Some neurophysiological eff ects
of azadirachtin on lepidopterous larvae and their feeding responses.
In: Schmutterer, H., Ascher, K.R.S. (Eds.), Proceedings of the Second
International Neem Conference, G.T.Z., Eschborn, pp. 163–180
Zabel, R.A., Morrell, J.J., 1992. Wood Microbiology Decay and Its Pre-
vention. Academic Press, San Diego, pp. 195–224.
Th iele, H.-U., 1977. Carabid Beetles in Th eir Environment. Zoophysi-
ological Ecology 10.Springer, Berlin, P 369
Toft, S., Bilde, T. 2002. Carabid diets and food value. In: Holland, J.M.
(Ed.), Th e Agroecology of Carabid Beetles. Intercept, Andover, UK,
pp. 81–110.
Tooley, J., Brust, G. 2002. Weed seed predation by carabid beetles. In:
Holland, J.M. (Ed.), Th e Agroecology of Carabid Beetles. Intercept,
Andover, UK, pp. 215–229.
Valaškova, V., Baldrian, P., 2006. Degradation of cellulose and hemicel-
luloses by the brown rot fungus Piptoporus betulinus—production,
of extracellular enzymes and characterization of the major cellulases.
Microbiology 152, 3613–3622.
Van Loon, J.J.A., 1990. Chemoreception of phenolic acids and fl avonoids
in larvae of two species of Pieris. Journal of Comparative Physiology
A 166, 889–899.
Wang, Z., Singhvi, A., Kong, P. and Scott, K. 2004. Taste representations
in the Drosophila brain. Cell 117, 981–991.

45
Varela, E., Tien, M., 2003. Eff ect of pH and oxalate on hydroquinone-
derived hydroxyl radical formation during brown rot wood degrada-
tion. Applied and Environmental Microbiology 69, 6025–6031.
Wheater, C.P., 1989. Prey detection by some predatory Coleoptera (Car-
abidae and Staphylinidae). Journal of Zoology A 218, 171–185.

46
SUMMARY IN ESTONIAN
JOOKSIKLASTE (COLEOPTERA: CARABIDAE)
ANTENNAALNE KONTAKTNE KEMORETSEPTSIOON
Rööv- ja segatoidulised jooksiklased oma suure liigirikkuse ja arvukuse tõttu omavad arvestatavat majanduslikku tähtsust kui põllu- ja aiakahjurite arvukuse olulised looduslikud reguleerijad.
Mitme- ja segatoiduliste jooksiklaste jaoks leidub põldudel alati piisavalt toitu. Jooksiklaste peamisteks toiduobjektideks on lehetäid, hooghänna-lised, liblikate ja mardikate munad, vastsed ning valmikud, kahetiivaliste munad ja vastsed ning vaablaste ebaröövikud. Suuremate liikide toiduks sobivad ka vihmaussid ja teod. Suure osa sega- ja taimtoiduliste liikide toi-dumenüüsse kuuluvad taimede, sealhulgas umbrohtude seemned. Just selle tõttu võivad jooksiklased erinevalt kitsalt spetsialiseerunud röövtoidulistest putukatest ja parasitoididest migreeruda põldudele varakevadel, enne kui sinna ilmuvad kultuurtaimede kahjurid. Hävitades kahjureid arvukuse tõusu algfaasis hoitakse ära nende hilisem massesinemine ja majanduslik kahju. Seepärast on mahetootmise ja integreeritud kahjuritõrje tingimustes manipulatsioonid jooksiklaste liigirikkuse ja arvukusega tähtsal kohal.
Jooksiklaste seotus teatud elupaigatüüpide ning varje- ja talvituskohtadega on määratud mitmesuguste biootiliste ja abiootiliste teguritega nagu toitu-mistingimused, taimkattetüüp, konkurentide olemasolu ja levik, aastaaeg, maastiku iseärasused, maaharimise mõju, temperatuuri, valgus- ja niiskus-tingimused jne. Pinnase sooladesisaldus ja pH võivad olla samuti tähtsad tegurid jooksiklaste levikul. Käitumiskatsetega on näidatud, et vastavad kontaktsed kemoretseptoosed sensillid ehk maitseharjased paiknevad nende mardikate tundlatel. Paljude jooksiklaste elupaigavalik on nii spetsiifi line, et sageli kasutatakse neid elupaikade iseloomustamiseks. Välisstiimulid ja otsingulise käitumise mehhanismid, mille abil jooksiklased oma toidu asukoha kindlaks teevad ja saagirikastele aladele peatuma jäävad, pole täpselt teada. Samal ajal, kui paljud liigid leiavad oma toidu nähtavasti juhusliku otsingu teel, kütivad mitmed teised, päevase aktiivsusega liigid, saaki nägemise abil. Paljud liigid kasutavad aga toiduga seotud lõhna-signaale. Keemilise informatsiooni kasutamine on jooksiklastel ilmselt tunduvalt rohkem levinud kui väheste avaldatud andmete põhjal võiks arvata. Jooksiklaste käitumist reguleerivaid keemilisi välisstiimuleid ning neid vastuvõtvate kemoretseptorite talitlust on seni üllatavalt vähe uuritud.

47
Elektronmikroskoopilised uuringud on näidanud, et jooksiklaste tundlatel
esineb tuhandeid erinevaid kemoretseptoorseid sensille. Lisaks arvukatele
pisikestele pulkjatele haistesensillidele, paikneb nende fl agellumil 56 kuni
70 suurt 35–200 μm pikkust harjasjat kontaktset kemosensilli. Kivijook-
sikul Nebria brevicollis on need sensillid varustatud nelja kemoretseptoorse
ja ühe mehhanoretseptoorse neuroniga. See on tüüpiline neuronite komp-
lekt enamiku putukarühmade kontaktsetes kemosensillides. Nendest
on jooksiklastel seni elektrofüsioloogiliselt kindlaks tehtud ainult ühe,
soolaneuroni, funktsioon. Ülejäänud kolme kemoretseptoorse neuroni
täpsem funktsioon pole teada. Kontaktset kemoretseptsiooni on põhjali-
kumalt uuritud paljudel taimtoidulistel putukatel, kellel need neuronid
võivad reageerida väga mitmesugustele taimsetele keemilistele stiimulitele,
enamasti aga toitumise stimulantidele (taimsed suhkrud, aminohapped,
suhkuralkoholid jt.) ja deterrentsetele ühenditele (alkaloidid, glükosiidid
jt.). Kuna stimulantsetel ja deterrentsetel ühenditel on toidu- ja peremees-
taime äratundmisel keskne roll, on nad viimastel aastakümnetel pälvinud
üha suuremat tähelepanu.
Käesoleva töö eesmärgiks on elektrofüsioloogilise meetodi abil selgitada
antennaalseid kontaktseid kemoretseptoreid (maitseharjaseid) innerveeri-
vate neuronite funktsiooni ümarselg-süsijooksikul (Pterostichus aethiops) ja
metsa-süsijooksikul (P. oblongopunctatus). Erinevate meetoditega (puhver-
lahuste seeriad, leelistatud ja leelistamata soolad) testiti ärrituslahuste pH
mõju nende neuronite reaktsioonile ning spetsiifi lise pH-tundliku neuroni
võimalikku esinemist nende liikide antennaalsetes maitseharjastes. Kuna
mõlemad uuritavad liigid on osaliselt taimtoidulised (granivoorid), testiti
ka antennaalsete maitseneuronite reaktsiooni mitmesugustele taimsetele
suhkrutele ja aminohapetele, et selgitada nende ainete vastuvõtmisele
spetsialiseerunud neuronite olemasolu ja talitluse iseärasusi jooksiklaste
tundlatel. Olulise osa tööst moodustavad elektrofüsioloogilised ja etoloogi-
lised eksperimendid, mille käigus selgitati taimeseemnetes leiduvate toksi-
liste ja deterrentsete ainete (alkaloidid ja glükosiidid) stimuleerivat toimet
nimetatud jooksiklaste antennaalsete maitseharjaste kemosensoorsetele
neuronitele. Katsete eesmärgiks oli selgitada spetsiifi lise deterrendineuroni
olemasolu ja nende ainete võimalikku inhibeerivat toimet maitseharjase
teistele neuronitele. Nii ümarselg-süsijooksik kui ka metsa-süsijooksik
on videvikuaktiivsusega omnivoorsed metsaliigid, kelle toidumenüüs on
tähtsal kohal okaspuude seemned. Katsemardikad koguti Lõuna-Eesti
metsadest.

48
Elektrofüsioloogilistes katsetes mõõdeti spetsiaalse aparatuuri abil mardi-
kate antennaalseid maitseharjaseid innerveerivate neuronite reaktsioone
ärrituslahustele, kasutades sensilli tipust neuronite impulss-aktiivsuse
rakuvälise registreerimise meetodit. Selleks, et suurendada ärrituslahuste
elektrijuhtivust, lahustati suhkrud, aminohapped, alkaloidid ja glükosiidid
vajalikus kontsentratsioonis 10 mmol l-1 elektrolüüdi (NaCl, koliinklo-
riid) lahuses. Puhtad ained hangiti fi rmadelt AppliChem (Saksamaa) ja
Sigma-Aldrich. Lahuste pH taset mõõdeti portatiivse pH-meetri E6115
(Evikon, Estonia) abil. Neuronite reaktsiooni mõõdeti toimepotentsiaa-
lide (närviimpulsside) arvuna sekundis. Et testida elektrofüsioloogiliselt
aktiivsete alkaloidide deterrentset toimet mardikate toitumise aktiivsusele,
viidi läbi vajalikud käitumuslikud valikukatsed alkaloidiga töödeldud
ja töötlemata männiseemnetega. Katseandmete analüüsimisel kasutati
erinevaid statistilisi teste, kasutades arvutitarkvara Statistica (StatSoft,
Inc., USA).
Elektrofüsioloogilised katsed atsetaat- ja fosfaatpuhvrite seeriatega ning
leelistatud ja leelistamata sooladega pH vahemikus 3 kuni 11 näitasid, et
ümarselg-süsijooksiku ja metsa-süsijooksiku antennaalsetes maitseharjastes
paikneb kaks neuronit, soola- ja pH-tundlik neuron, mis reageerivad nende
elektrolüütide lahustele. pH-tundliku neuroni keskmine impulss-aktiiv-
sus tõusis alati koos ärrituslahuse pH-taseme tõusuga ulatudes aluselise
reaktsiooniga ärrituslahuste puhul (100 mmol l-1 NaCl + 10 mmol l-1
NaOH; pH 9,6) maksimaalselt kuni 30 imp/s, samal ajal kui soola-tund-
liku neuroni aktiivsus ei muutunud või muutus ainult vähesel määral.
Samas täheldati, et ka ärrituslahustes olevate soolade iooniline koostis
mõjutas märgatavalt pH-tundliku neuroni närviimpulsside sagedust.
Saadud tulemus on huvipakkuv, kuna varem pole putukate kontaktsetes
kemosensoorsetes sensillides spetsiifi lise pH-tundliku neuroni olemasolu
täheldatud. Uuritud jooksiklaste antennaalne pH-tundlik neuron näib
olevat seotud mardikate elupaiga ja talvituskohtade valikuga. pH-tundliku
neuroni impulssaktiivsus puudub või on madalseisus (alla 5 imp/s) pH
väärtustel 3 kuni 6. See happelisuse piirkond vastab pinnase pH tase-
mele uuritud jooksiklaste metsabiotoopides ja eelistatud talvituskohtades
pruunmädaniku poolt lagundatud puidus, kus mardikad veedavad 7–8
kuud, septembrist aprillini.
Käesoleva uurimuse raames identifi tseeriti esmakordselt granivoorsetel
lülijalgsetel (ümarselg-süsijooksikul ja metsa-süsijooksikul) elektrofüsio-

49
loogiliselt suhkrutundliku neuroni olemasolu. Katsed näitasid, et 12-st
testitud taimsest suhkrust 3 (maltoos, sahharoos ja glükoos) osutusid
füsioloogiliselt aktiivseteks, stimuleerides antennaalsete maitseharjaste
suhkrutundliku neuroni impulssaktiivsust. Tüübilt oli neuroni reakt-
sioon 1–1000 mmol l-1 suhkrulahusele faasilis-tooniline, sõltudes lahuse
kontsentratsioonist. Metsa-süsijooksikul osutusid 1,4-glükosiidsidemega
disahhariidid maltoos ja sahharoos füsioloogiliselt aktiivseimateks, tõstes
1000 mmol l-1 kontsentratsioonil suhkrutundliku neuroni impulss-aktiiv-
suse tasemele 70 imp/s. Glükoosi stimuleeriv toime oli ligi kaks korda
madalam. Kuna maltoosi leidub looduses peamiselt idanevates seemnetes
ja noortes idantites, võib oletada, et uuritud jooksiklaste suhkrutundlikkus
on seotud nende osalise taimtoidulisusega (mardikad toituvad okaspuude
seemnetest ja noortest idanditest). Ükski puidu ensümaatilise lagunemise
käigus tekkivast vees lahustuvast suhkrust (tsellobioos, arabinoos, ksüloos,
mannoos, ramnoos ja galaktoos) ei osutunud füsioloogiliselt aktiivseks,
mistõttu nad ei osale nähtavasti metsa-süsijooksiku otsingulises käitu-
mises. Mõned testitud seitsmest 100 mmol l-1 aminohappest (metioniin,
seriin, alaniin, glutamiin) avaldasid nõrka stimuleerivat toimet (alla 3
imp/s) metsa-süsijooksiku maitseharjase kemosensoorsetele neuronitele,
mida võib aga iseloomustada kui maitseharjase neuronite reaktsiooni
mittespetsiifi list modulatsiooni.
Antennaalsete maitseharjaste kemosensoorsete neuronite reaktsiooni taim-
setele alkaloididele ja glükosiididele testiti metsa-süsijooksikul. Katsete
tulemusena identifi tseeriti granivoorsetel lülijalgsetel esmakordselt alka-
loiditundlik neuron. Ilmnes, et testitud viiest alkaloidist avaldasid sellele
neuronile tugevat stimuleerivat toimet vaid kiniin ja selle sool kiniinhüd-
rokloriid. Alkaloiditundliku neuroni faasilis-toonilised reaktsioonid 0,001
kuni 50 mmol l-1 olid võrdelises sõltuvuses lahuse kontsentratsioonist.
Neuroni erutusläveks kiniinhüdrokloriidi suhtes osutus kontsentratsioon
0,01 mmol l-1, maksimaalseid reaktsioone (67 imp/s) täheldati kontsentrat-
sioonil 50 mmol l-1. Käitumiskatsetes osutus kiniinhüdrokloriid tugevaks
toitumise deterrendiks. Kofeiini stimuleeriv efekt sellele neuronile oli väga
nõrk (alla 2 imp/s), nikotiin aktiivsust ei mõjutanud, strühniin aga pärssis
nõrgalt selle neuroni impulssaktiivsust.
Lisaks kiniini ja kiniinhüdrokloriidi alkaloidineuronit stimuleerivale
toimele võivad nad tugevalt pärssida metsa-süsijooksiku maitseharja-
se soola- ja pH-tundliku neuroni impulssaktiivsust. Samuti ilmnes, et

50
glükosiidid sinigriin ja salitsiin ning alkaloidid kofeiin ja nikotiin, mille
mõju alkaloiditundlikule neuronile oli nõrk või puudus üldse, võivad
samuti tugevalt pärssida soola- ja pH-tundliku neuroni impulssaktiivsust.
Saadud tulemused lubavad oletada, et jooksiklase antennaalsed maitse-
harjased võivad taimseid toksilisi ja deterrentse toimega aineid avastada
nii alkaloiditundliku neuroni stimuleerimise kui ka maitseharjase teiste
kemosensoorsete neuronite periferaalse inhibitsiooni teel.

51
ACKNOWLEDGEMENTS
Th is study was carried out at the Institute of Agricultural and Environ-
mental Sciences of the Estonian University of Life Sciences. I would like
to express my gratitude to my supervisors Enno Merivee and Prof. Anne
Luik who has been supporting and helping me completing in this thesis.
I am grateful to Kaljo Voolma and Heino Õunap for their review of my
thesis and good comments on how to improve it.
I sincerely thank my colleagues in the Institute of Agricultural and En-
vironmental Sciences for their support.
Th e Estonian Science foundation (Grants No 5423 and 6958) fi nancially
supported this study.
I
PUBLICATIONS

Merivee, E., Ploomi, A., Milius, M., Luik, A., Heidemaa, M., 2005.
ELECTROPHYSIOLOGICAL IDENTIFICATION OF
ANTENNAL PH RECEPTORS IN THE GROUND BEETLE
PTEROSTICHUS OBLONGOPUNCTATUS.
Physiological Entomology 30(2), 122–33.

55
Electrophysiological identification of antennal pHreceptors in the ground beetle Pterostichusoblongopunctatus
ENNO MER I V E E 1 , A NGE LA P LOOM I 1 , MAR I T M I L I U S 1 , ANNE
LU IK 1 and M IKK HE I D EMAA 2
1Estonian Agricultural University, Institute of Plant Protection, Tartu and 2University of Tartu, Institute of Zoology and
Hydrobiology, Tartu, Estonia
Abstract. Electrophysiological responses of antennal taste bristles to 100mM
acetate and phosphate buffers were tested at pH 3–11 in the ground beetlePterostichus oblongopunctatus (F.) (Coleoptera, Carabidae). Additionally,responses of these sensilla to 10 and 100mM phosphate buffers were comparedwith each other. Generally, in response to these stimulating solutions, two sensorycells, classified as a salt cell (cation cell) and a pH cell, respectively, showed actionpotentials distinguished by differences in their amplitudes and polarity of spikes.The firing rate of the cation cell increased with increasing buffer concentration,and was influenced by buffer pH in a complicated way. The best stimulus for thesecond cell (pH cell) was pH of the stimulating buffer solution. As the pH of thestimulus solution increased, higher rates of firing were produced by the pH cell.For example, the number of action potentials elicited by 100mM phosphate bufferat pH11.1 was approximately 16-fold higher compared with that at pH 8.1, andfiring rates during the first second of the response were 27.9 and 1.7 imp/s,respectively. The pH cell did not fire or fired at very low frequency (first secondresponse below 5 imp/s) at pH3–6. This level of acidity probably represents thepH preferences of this ground beetle in its forest habitat and hibernating sites.By contrast to the cation cell, the pH cell responded to increases in bufferconcentration by decreasing its firing rate.
Key words. Action potentials, firing rate, taste sensilla, tip recording.
Introduction
The life of carabids from egg to death of the adult proceeds
in direct physical contact with the ground (from which the
name of the beetles is derived in English: ground beetles).
Due to scarce data, we can only assume that, in searching
and selection of their habitat, refugia and hibernating sites,
besides other stimuli, ground beetles perceive and analyse
external chemical information, such as soil pH, salt content
and concentration. However, some data obtained from
behavioural experiments concerning the ground beetles’
ability to perceive salt content and pH of the ground
are contradictory (Thiele, 1977). Nevertheless, Paje &
Mossakowski (1984) demonstrated convincingly that
behaviourally ground beetles have certain pH preferences
concerning their habitat; ablation experiments conducted
by these authors suggested that pH receptors are located
on the beetles’ antennae.
Scanning electron microscope studies demonstrate that
the most probable candidates for pH receptors (i.e. large
blunt-tipped sensilla) can be found abundantly on the
antennae of all ground beetles. These antennae consist of
a scapus, pedicel and nine flagellomeres. The taste receptors
stand in a whorl of six to seven bristles, in distal parts of the
flagellomeres. In addition, six bristles are located in pairs
Correspondence: Dr Enno Merivee, Estonian Agricultural
University, Institute of Plant Protection, 64 Kreutzwaldi Street,
51014 Tartu, Estonia. E-mail: [email protected]
Physiological Entomology (2005) 30, 122–133
122 # 2005 The Royal Entomological Society

symmetrically at the antennal tip (Kim & Yamasaki, 1996;
Merivee et al., 2000, 2001, 2002). According to Daly &
Ryan (1979), these taste bristles in the ground beetle
Nebria brevicollis are innerved with five neurones: four
chemoreceptor cells and one mechanoreceptor cell. In
electrophysiological experiments with acid, neutral and
alkaline salts, it was demonstrated that two chemoreceptor
cells of the antennal taste bristles respond to these salt
solutions in ground beetles. One of the cells, the salt cell,
was sensitive to ionic (cationic) content and concentration
of the stimulating solution. Firing rates of the second cell
were proportional to the pH level of the stimulating
salt solutions tested (Merivee et al., unpublished data).
Unfortunately, the tested salts were dissolved in distilled
water and not in buffer solutions. It is likely that the pH
level of the tested salt solutions was unstable and changed
during experiments. Thus, the function of the second salt
cell (pH cell) requires further testing using buffer-controlled
pH solutions. To date, electrophysiological evidence con-
cerning the occurrence of pH receptors in insects is lacking,
but it has been shown repeatedly that the pH of stimulating
solutions may affect neural responses of insect taste sensilla
(Shiraishi & Morita, 1969; Bernays et al., 1998).
The aim of the present study is to test responses of
antennal taste bristles to acetate and phosphate buffers in
a wide range of pH values (3–11) in the ground beetle
Pterostichus oblongopunctatus F. (Coleoptera, Carabidae)
to investigate the functioning of pH receptors in ground
beetles.
Materials and methods
Test beetles
Test beetles were collected in April 2003 from their
hibernation sites in forest margins in southern Estonia.
The beetles were kept in plastic boxes filled with moistened
moss in a refrigerator at þ5 �C. Three or four days before
the electrophysiological tests, the beetles were kept in
similar boxes at room temperature (þ 22 �C). The beetles
were given clean water to drink and fed with moistened cat
food (Kitekat, Master Foods, Poland) every day.
Test beetles were immobilized by placing them tightly
into a special conical tube made of thin sheet-aluminium
of a size that allowed their head and antennae to protrude
only a little from the narrower end. The wider rear end of
the conical tube was blocked with a piece of plasticine to
prevent the beetle from retreating out of the tube. The
antennae of immobilized intact beetles were fastened hori-
zontally on the edge of a special aluminium stand with tiny
amounts of beeswax, such that some horizontally located
antennal taste bristles were visible from above under a light
microscope, and easily accessible for micromanipulations
from the side. Contamination of tested taste bristles and
their contact with preparation instruments and beeswax was
avoided.
Chemicals and preparation of buffer solutions
Before experiments, 100mM acetate buffers in the pH
range 3–6 were prepared by dissolving glacial acetic acetate
and anhydrous sodium acetate in distilled water. Stimulat-
ing 100mM buffers with higher levels of pH (5.8–8.1) were
made with combinations of Na2HPO4 and NaH2PO4. For
some experiments, the pH of phosphate buffer was raised to
11.1 with NaOH. The pH of the 100mM borax used in some
test series with acetate buffers was 9.3. All chemicals
were obtained from AppliChem (Germany). To produce
stimulating buffers of approximately the same pH but of
different concentrations of ions, the 100mM phosphate
buffer (pH7.6) was diluted in distilled water to 10mM
solution (pH7.8). The final pH of the stimulating solutions
was measured with a pH meter E6115 (Evikon, Estonia).
Stimulation and recording of action potentials
Electrophysiological recordings were made from large
chaetoid taste sensilla on the antennal flagellum of the
beetle with a tip-recording technique (Hodgson et al.,
1955). To achieve a good signal-to-noise ratio and avoid
registration of muscular activities from two basal antenno-
meres, the scape and pedicel and from other parts of insect’s
body, an indifferent tungsten microelectrode was inserted
into the base of the flagellum. The recording microelec-
trode, a glass micropipette (diameter at the tip approxi-
mately 20 mm) filled with stimulating buffer solution was
placed over the sensillum tip by means of a micromanipu-
lator under visual control with a light microscope at
magnification � 300. Immediately before each stimulation,
two to three drops of the stimulating solution were squeezed
out of the micropipette tip, and absorbed onto a piece of
filter paper attached to an additional micromanipulator to
reduce concentration increases due to evaporation. A
copper wire made contact with high-impedance preampli-
fier input. The signal picked up by the recording electrode
was amplified and filtered with a band width set at
100–2000Hz and monitored on an oscilloscope. Recordings
of the first 5 s of the response were transferred directly to a
computer via an analogue-to-digital input board DAS-1401
(Keithley, Taunton, Massachusetts) for data acquisition,
storage and further analysis using TestPoint (Capital
Equipment Corp., Billerica, Massachusetts) software. The
sampling data rate was 10 kHz.
Due to variations in responsiveness of different taste
bristles to the same stimuli, on the antennae of one and
the same as well as different beetles, stimulating buffers
were tested in pairs to allow a more precise comparison of
the responses to the tested stimuli. Thus, each taste sensil-
lum was tested twice. Up to 15 sensilla on one antenna of a
test beetle were in a suitable position for stimulations and
recordings. Primarily, these sensilla were tested with the first
stimulus. Then, after approximately 30min, the stimulat-
ing/recording micropipette was rinsed with distilled water
several times and filled with the second stimulus to test the
pH receptors in P. oblongopunctatus 123
# 2005 The Royal Entomological Society, Physiological Entomology, 30, 122–133

57
same sensilla again. The space of time between the two
successive stimulations ensured complete disadaptation of
the receptor cells. After the test series, test beetles remained
in good condition, and no differences were observed in the
responses of their taste bristles recorded at the beginning
and the end of the test series. Antennal sensilla from four to
six beetles were tested in each pair of stimuli.
Data management and analysis
Action potentials from several sensory cells of taste sen-
silla were distinguished by differences in their amplitude,
duration, shape and polarity of spikes. In some cases, the
regularity of firing as the principal parameter for separating
and identifying action potentials was used. Automatic
classifying and counting of action potentials from taste
sensilla was clearly not an appropriate method for data
analysis because spike amplitude of a certain cell often
changed in time or with concentration, and two action
potentials frequently interacted to produce an irregular
waveform. Instead, the analysis used was visual, employing
TestPoint software (CEC Capital Equipment, Bedford,
New Hampshire). Firing rates of receptor cells were
expressed as a number of spikes per second. All data
were analysed using Microsoft Excel (Microsoft Corp.,
Redmond, Washington) and Statistica (StatSoft, Inc.,
Tulsa, Oklohoma) software.
Results
Taste sensilla on the antennal flagellum in
P. oblongopunctatus
Large blunt-tipped 100–200-mm long flagellar taste
bristles of the ground beetle P. oblongopunctatus can be
quantified at low magnification under the light microscope.
The largest bristles occur on the first flagellomere counting
from the beetle’s head. The length of the bristles towards the
tip of the antenna decreases, and the shortest of the bristles
can be found at the tip of the terminal flagellomere. Bristles
are located in whorls in the distal part of flagellomeres.
There are six on the first flagellomere and seven on the
following flagellomeres. In addition to those, there are six
bristles symmetrically at the tip of the terminal flagellomere.
Hence, the total number of taste bristles on each antennal
flagellum is 68. All these bristles seemed to respond in a
similar manner to the stimuli tested.
Characterization of action potentials recorded from antennal
taste sensilla
Two sensory cells innervating flagellar taste bristles in
P. oblongopunctatus responded to stimulation with acetate
buffers, phosphate buffers and borax: A- and B-cell. Action
potentials with positive spikes generated by the A-cell are
extremely large, approximately 8mV peak-to-peak. By con-
trast, action potentials with negative spikes originating
from the B-cell are three- to four-fold smaller in amplitude
compared with the action potentials generated by the A-cell
(Fig. 1A). The responses to tested stimuli were phasic–tonic
in type, with a more pronounced phasic component in
B-cells. On the recordings, action potentials from two cells
frequently interacted to produce an irregular waveform. In
these cases, the regularity of firing as the principal para-
meter for visual separating and identifying action potentials
was used (Fig. 1A). In some cases, responses of the A-cells
were of a bursting nature (Figs 1C,D). Usually action
potentials generated by the same cell had similar amplitude.
However, in a number of cases, especially at the beginning
of the B-cell’s response, a few action potentials could be
observed whose amplitude was remarkably greater than the
others (Fig. 1B).
Responses to acetate buffers at pH 3 and 4
The A-cell did not respond to 100mM acetate buffer at
pH 3 and only a very weak response was observed at pH4:
the average number of impulses during the first and follow-
ing seconds of the response was less than 1. At the same
time, the A-cell of the same bristles strongly responded to
borax: on average, 23 impulses per first second of the
response were recorded (Fig. 2A, A-cell), demonstrating
that the tested cells were in good functional condition.
The B-cell responded to 100mM acetate buffers at pH 3
and 4 also relatively weakly: 5.7 and 9.5 impulses per first
second of the response, respectively (P< 0.05). The
response of the same B-cells to borax was 24 imp/s. Dur-
ing the following seconds of the response (in the tonic
part), the firing rate of the B-cell stabilized at the level of
1–3 imp/s; compared with the first second of the response,
the opposite trend was noted. The firing rate of the cell at
pH 4 appeared to be lower than that at pH3, although
the difference was not statistically significant (P> 0.05;
Fig. 2A, B-cell). Some typical 5-s responses of antennal
taste sensilla to stimulating solutions at pH 3 and 4 are
shown in Fig. 3.
Responses to acetate buffers at pH 4 and 5
In response to stimulation with acetate buffer at pH5, the
A-cells generated significantly more action potentials (first
second mean response 3.6 impulses) compared with the
response at pH 4 (0.9 impulses per first second of the
response) (P< 0.05; Fig. 2B, A-cell).
During the first two seconds of the response, B-cells
fired at a significantly higher frequency at pH 5 than at
pH 4 (P< 0.05; first second responses 15.9 and 9.7 imp/s,
respectively). The third second response was approxi-
mately equal (P> 0.05). As observed in the test series
with stimulating solutions at pH 3 and 4, during the last
124 E. Merivee et al.
# 2005 The Royal Entomological Society, Physiological Entomology, 30, 122–133
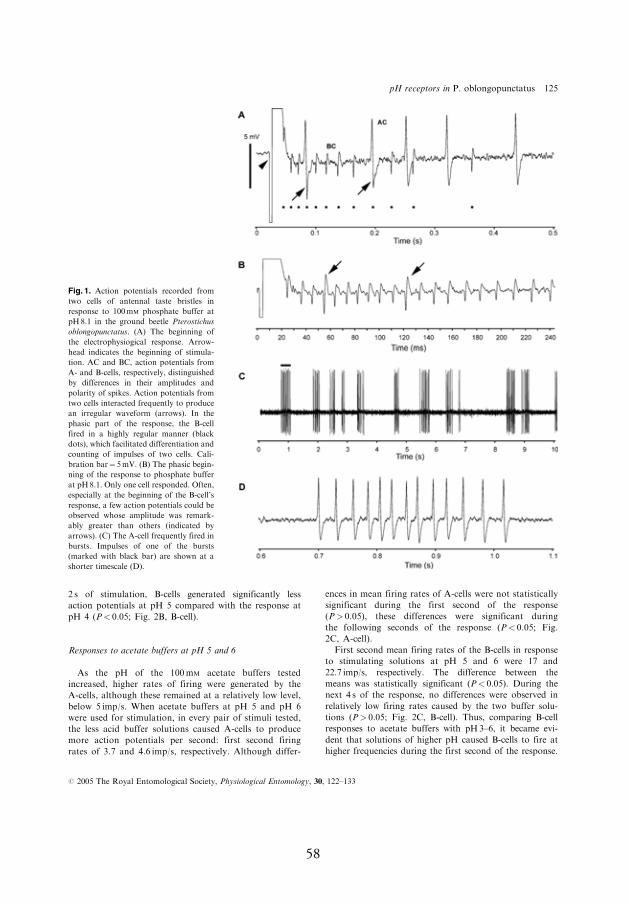
58
2 s of stimulation, B-cells generated significantly less
action potentials at pH 5 compared with the response at
pH 4 (P< 0.05; Fig. 2B, B-cell).
Responses to acetate buffers at pH 5 and 6
As the pH of the 100mM acetate buffers tested
increased, higher rates of firing were generated by the
A-cells, although these remained at a relatively low level,
below 5 imp/s. When acetate buffers at pH 5 and pH 6
were used for stimulation, in every pair of stimuli tested,
the less acid buffer solutions caused A-cells to produce
more action potentials per second: first second firing
rates of 3.7 and 4.6 imp/s, respectively. Although differ-
ences in mean firing rates of A-cells were not statistically
significant during the first second of the response
(P> 0.05), these differences were significant during
the following seconds of the response (P< 0.05; Fig.
2C, A-cell).
First second mean firing rates of the B-cells in response
to stimulating solutions at pH 5 and 6 were 17 and
22.7 imp/s, respectively. The difference between the
means was statistically significant (P< 0.05). During the
next 4 s of the response, no differences were observed in
relatively low firing rates caused by the two buffer solu-
tions (P> 0.05; Fig. 2C, B-cell). Thus, comparing B-cell
responses to acetate buffers with pH3–6, it became evi-
dent that solutions of higher pH caused B-cells to fire at
higher frequencies during the first second of the response.
Fig. 1. Action potentials recorded from
two cells of antennal taste bristles in
response to 100mM phosphate buffer at
pH8.1 in the ground beetle Pterostichus
oblongopunctatus. (A) The beginning of
the electrophysiogical response. Arrow-
head indicates the beginning of stimula-
tion. AC and BC, action potentials from
A- and B-cells, respectively, distinguished
by differences in their amplitudes and
polarity of spikes. Action potentials from
two cells interacted frequently to produce
an irregular waveform (arrows). In the
phasic part of the response, the B-cell
fired in a highly regular manner (black
dots), which facilitated differentiation and
counting of impulses of two cells. Cali-
bration bar¼ 5mV. (B) The phasic begin-
ning of the response to phosphate buffer
at pH8.1. Only one cell responded. Often,
especially at the beginning of the B-cell’s
response, a few action potentials could be
observed whose amplitude was remark-
ably greater than others (indicated by
arrows). (C) The A-cell frequently fired in
bursts. Impulses of one of the bursts
(marked with black bar) are shown at a
shorter timescale (D).
pH receptors in P. oblongopunctatus 125
# 2005 The Royal Entomological Society, Physiological Entomology, 30, 122–133

59
Fig. 2. Responses of antennal taste bristles to 100mM acetate buffers at pH 3–6. Because the test insects and their taste sensilla varied
greatly in their responses to the same stimulus, stimuli were tested in pairs with one and the same bristle. (A) The A-cell did not fire,
or fired at very low frequency, when stimulated by acetate buffers at pH 3 and 4. However, strong responses to borax showed that the
same cells were in a good functional condition. (B and C) According to how the pH level of stimulus solution increased, higher rates
of firing were observed in A-cells. The increase in pH caused B-cells to fire more frequently, especially during the first second of the
response, but to a remarkably lesser extent than in A-cells. Vertical bars indicate SE of the means; asterisk (*) and n.s. indicate
whether the difference between the mean firing rates is significant or not, respectively (paired t-test, P< 0.05). n, Number of taste
bristles tested.
126 E. Merivee et al.
# 2005 The Royal Entomological Society, Physiological Entomology, 30, 122–133
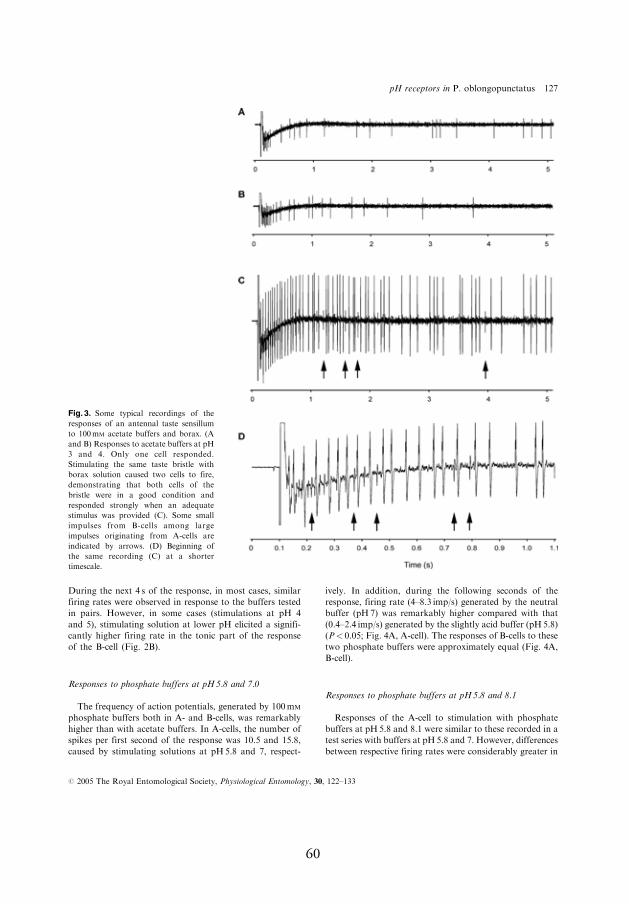
60
During the next 4 s of the response, in most cases, similar
firing rates were observed in response to the buffers tested
in pairs. However, in some cases (stimulations at pH 4
and 5), stimulating solution at lower pH elicited a signifi-
cantly higher firing rate in the tonic part of the response
of the B-cell (Fig. 2B).
Responses to phosphate buffers at pH 5.8 and 7.0
The frequency of action potentials, generated by 100mM
phosphate buffers both in A- and B-cells, was remarkably
higher than with acetate buffers. In A-cells, the number of
spikes per first second of the response was 10.5 and 15.8,
caused by stimulating solutions at pH5.8 and 7, respect-
ively. In addition, during the following seconds of the
response, firing rate (4–8.3 imp/s) generated by the neutral
buffer (pH7) was remarkably higher compared with that
(0.4–2.4 imp/s) generated by the slightly acid buffer (pH5.8)
(P< 0.05; Fig. 4A, A-cell). The responses of B-cells to these
two phosphate buffers were approximately equal (Fig. 4A,
B-cell).
Responses to phosphate buffers at pH 5.8 and 8.1
Responses of the A-cell to stimulation with phosphate
buffers at pH5.8 and 8.1 were similar to these recorded in a
test series with buffers at pH5.8 and 7. However, differences
between respective firing rates were considerably greater in
Fig. 3. Some typical recordings of the
responses of an antennal taste sensillum
to 100mM acetate buffers and borax. (A
and B) Responses to acetate buffers at pH
3 and 4. Only one cell responded.
Stimulating the same taste bristle with
borax solution caused two cells to fire,
demonstrating that both cells of the
bristle were in a good condition and
responded strongly when an adequate
stimulus was provided (C). Some small
impulses from B-cells among large
impulses originating from A-cells are
indicated by arrows. (D) Beginning of
the same recording (C) at a shorter
timescale.
pH receptors in P. oblongopunctatus 127
# 2005 The Royal Entomological Society, Physiological Entomology, 30, 122–133
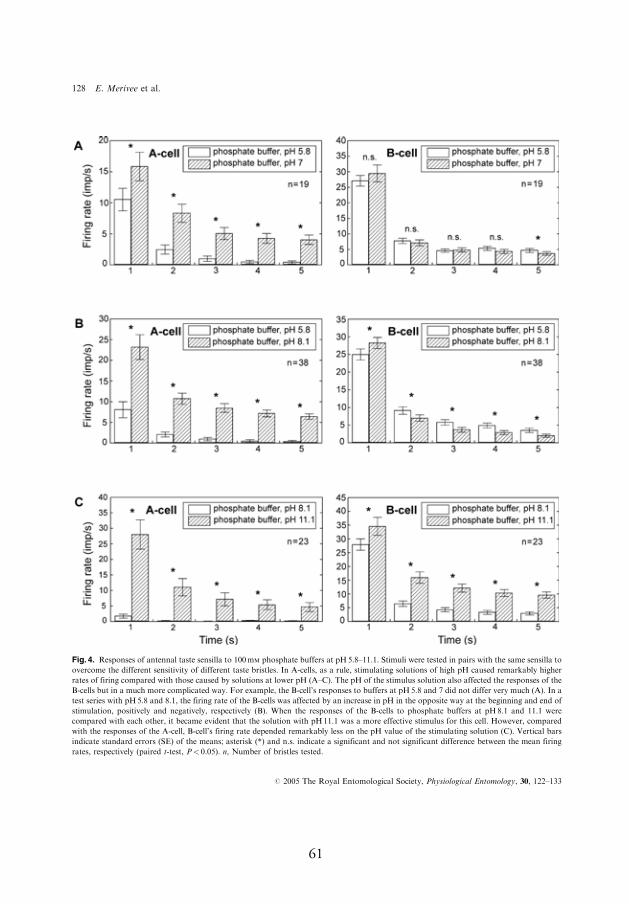
61
Fig. 4. Responses of antennal taste sensilla to 100mM phosphate buffers at pH5.8–11.1. Stimuli were tested in pairs with the same sensilla to
overcome the different sensitivity of different taste bristles. In A-cells, as a rule, stimulating solutions of high pH caused remarkably higher
rates of firing compared with those caused by solutions at lower pH (A–C). The pH of the stimulus solution also affected the responses of the
B-cells but in a much more complicated way. For example, the B-cell’s responses to buffers at pH5.8 and 7 did not differ very much (A). In a
test series with pH5.8 and 8.1, the firing rate of the B-cells was affected by an increase in pH in the opposite way at the beginning and end of
stimulation, positively and negatively, respectively (B). When the responses of the B-cells to phosphate buffers at pH8.1 and 11.1 were
compared with each other, it became evident that the solution with pH11.1 was a more effective stimulus for this cell. However, compared
with the responses of the A-cell, B-cell’s firing rate depended remarkably less on the pH value of the stimulating solution (C). Vertical bars
indicate standard errors (SE) of the means; asterisk (*) and n.s. indicate a significant and not significant difference between the mean firing
rates, respectively (paired t-test, P< 0.05). n, Number of bristles tested.
128 E. Merivee et al.
# 2005 The Royal Entomological Society, Physiological Entomology, 30, 122–133
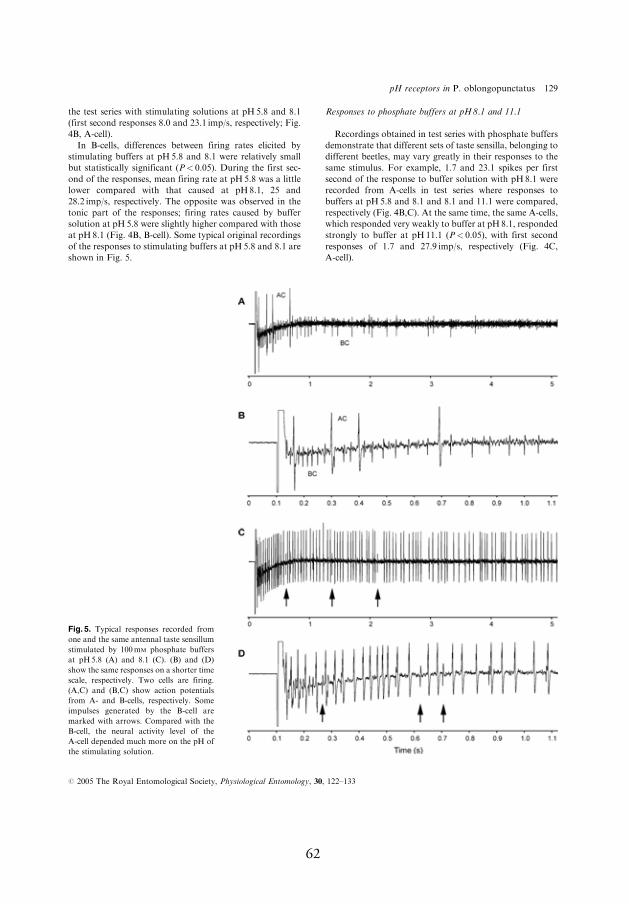
62
the test series with stimulating solutions at pH5.8 and 8.1
(first second responses 8.0 and 23.1 imp/s, respectively; Fig.
4B, A-cell).
In B-cells, differences between firing rates elicited by
stimulating buffers at pH5.8 and 8.1 were relatively small
but statistically significant (P< 0.05). During the first sec-
ond of the responses, mean firing rate at pH5.8 was a little
lower compared with that caused at pH8.1, 25 and
28.2 imp/s, respectively. The opposite was observed in the
tonic part of the responses; firing rates caused by buffer
solution at pH5.8 were slightly higher compared with those
at pH8.1 (Fig. 4B, B-cell). Some typical original recordings
of the responses to stimulating buffers at pH5.8 and 8.1 are
shown in Fig. 5.
Responses to phosphate buffers at pH 8.1 and 11.1
Recordings obtained in test series with phosphate buffers
demonstrate that different sets of taste sensilla, belonging to
different beetles, may vary greatly in their responses to the
same stimulus. For example, 1.7 and 23.1 spikes per first
second of the response to buffer solution with pH8.1 were
recorded from A-cells in test series where responses to
buffers at pH5.8 and 8.1 and 8.1 and 11.1 were compared,
respectively (Fig. 4B,C). At the same time, the same A-cells,
which responded very weakly to buffer at pH8.1, responded
strongly to buffer at pH11.1 (P< 0.05), with first second
responses of 1.7 and 27.9 imp/s, respectively (Fig. 4C,
A-cell).
Fig. 5. Typical responses recorded from
one and the same antennal taste sensillum
stimulated by 100mM phosphate buffers
at pH5.8 (A) and 8.1 (C). (B) and (D)
show the same responses on a shorter time
scale, respectively. Two cells are firing.
(A,C) and (B,C) show action potentials
from A- and B-cells, respectively. Some
impulses generated by the B-cell are
marked with arrows. Compared with the
B-cell, the neural activity level of the
A-cell depended much more on the pH of
the stimulating solution.
pH receptors in P. oblongopunctatus 129
# 2005 The Royal Entomological Society, Physiological Entomology, 30, 122–133
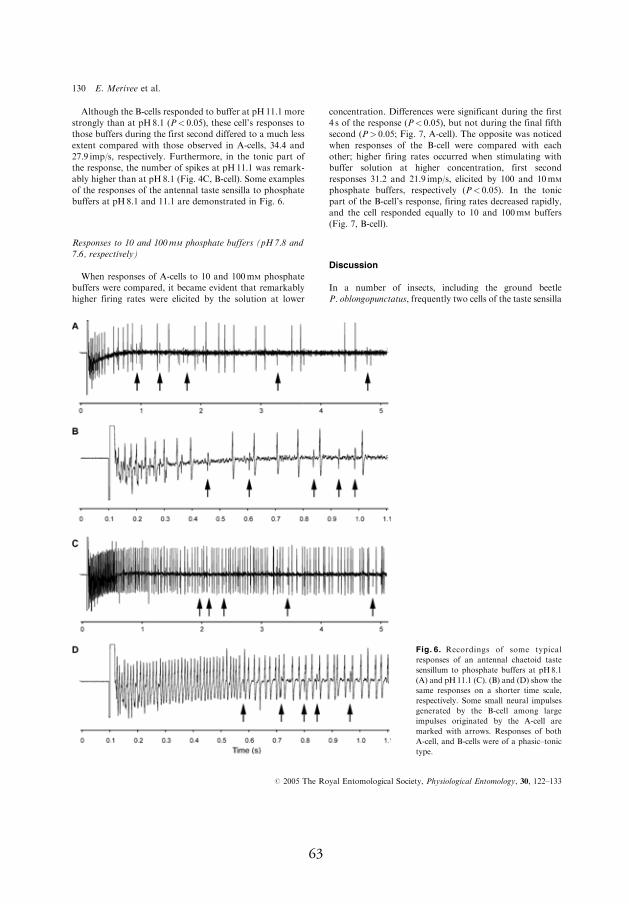
63
Although the B-cells responded to buffer at pH11.1 more
strongly than at pH8.1 (P< 0.05), these cell’s responses to
those buffers during the first second differed to a much less
extent compared with those observed in A-cells, 34.4 and
27.9 imp/s, respectively. Furthermore, in the tonic part of
the response, the number of spikes at pH11.1 was remark-
ably higher than at pH8.1 (Fig. 4C, B-cell). Some examples
of the responses of the antennal taste sensilla to phosphate
buffers at pH8.1 and 11.1 are demonstrated in Fig. 6.
Responses to 10 and 100mM phosphate buffers (pH7.8 and
7.6, respectively)
When responses of A-cells to 10 and 100mM phosphate
buffers were compared, it became evident that remarkably
higher firing rates were elicited by the solution at lower
concentration. Differences were significant during the first
4 s of the response (P< 0.05), but not during the final fifth
second (P> 0.05; Fig. 7, A-cell). The opposite was noticed
when responses of the B-cell were compared with each
other; higher firing rates occurred when stimulating with
buffer solution at higher concentration, first second
responses 31.2 and 21.9 imp/s, elicited by 100 and 10mM
phosphate buffers, respectively (P< 0.05). In the tonic
part of the B-cell’s response, firing rates decreased rapidly,
and the cell responded equally to 10 and 100mM buffers
(Fig. 7, B-cell).
Discussion
In a number of insects, including the ground beetle
P. oblongopunctatus, frequently two cells of the taste sensilla
Fig. 6. Recordings of some typical
responses of an antennal chaetoid taste
sensillum to phosphate buffers at pH8.1
(A) and pH11.1 (C). (B) and (D) show the
same responses on a shorter time scale,
respectively. Some small neural impulses
generated by the B-cell among large
impulses originated by the A-cell are
marked with arrows. Responses of both
A-cell, and B-cells were of a phasic–tonic
type.
130 E. Merivee et al.
# 2005 The Royal Entomological Society, Physiological Entomology, 30, 122–133
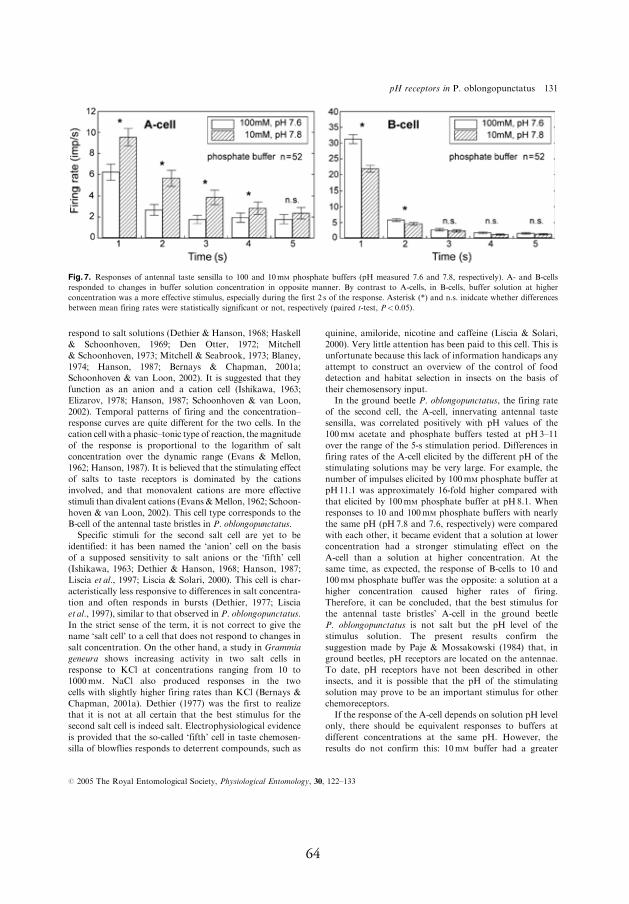
64
respond to salt solutions (Dethier & Hanson, 1968; Haskell
& Schoonhoven, 1969; Den Otter, 1972; Mitchell
& Schoonhoven, 1973; Mitchell & Seabrook, 1973; Blaney,
1974; Hanson, 1987; Bernays & Chapman, 2001a;
Schoonhoven & van Loon, 2002). It is suggested that they
function as an anion and a cation cell (Ishikawa, 1963;
Elizarov, 1978; Hanson, 1987; Schoonhoven & van Loon,
2002). Temporal patterns of firing and the concentration–
response curves are quite different for the two cells. In the
cation cell with a phasic–tonic type of reaction, themagnitude
of the response is proportional to the logarithm of salt
concentration over the dynamic range (Evans & Mellon,
1962; Hanson, 1987). It is believed that the stimulating effect
of salts to taste receptors is dominated by the cations
involved, and that monovalent cations are more effective
stimuli than divalent cations (Evans &Mellon, 1962; Schoon-
hoven & van Loon, 2002). This cell type corresponds to the
B-cell of the antennal taste bristles in P. oblongopunctatus.
Specific stimuli for the second salt cell are yet to be
identified: it has been named the ‘anion’ cell on the basis
of a supposed sensitivity to salt anions or the ‘fifth’ cell
(Ishikawa, 1963; Dethier & Hanson, 1968; Hanson, 1987;
Liscia et al., 1997; Liscia & Solari, 2000). This cell is char-
acteristically less responsive to differences in salt concentra-
tion and often responds in bursts (Dethier, 1977; Liscia
et al., 1997), similar to that observed in P. oblongopunctatus.
In the strict sense of the term, it is not correct to give the
name ‘salt cell’ to a cell that does not respond to changes in
salt concentration. On the other hand, a study in Grammia
geneura shows increasing activity in two salt cells in
response to KCl at concentrations ranging from 10 to
1000mM. NaCl also produced responses in the two
cells with slightly higher firing rates than KCl (Bernays &
Chapman, 2001a). Dethier (1977) was the first to realize
that it is not at all certain that the best stimulus for the
second salt cell is indeed salt. Electrophysiological evidence
is provided that the so-called ‘fifth’ cell in taste chemosen-
silla of blowflies responds to deterrent compounds, such as
quinine, amiloride, nicotine and caffeine (Liscia & Solari,
2000). Very little attention has been paid to this cell. This is
unfortunate because this lack of information handicaps any
attempt to construct an overview of the control of food
detection and habitat selection in insects on the basis of
their chemosensory input.
In the ground beetle P. oblongopunctatus, the firing rate
of the second cell, the A-cell, innervating antennal taste
sensilla, was correlated positively with pH values of the
100mM acetate and phosphate buffers tested at pH3–11
over the range of the 5-s stimulation period. Differences in
firing rates of the A-cell elicited by the different pH of the
stimulating solutions may be very large. For example, the
number of impulses elicited by 100mM phosphate buffer at
pH11.1 was approximately 16-fold higher compared with
that elicited by 100mM phosphate buffer at pH8.1. When
responses to 10 and 100mM phosphate buffers with nearly
the same pH (pH7.8 and 7.6, respectively) were compared
with each other, it became evident that a solution at lower
concentration had a stronger stimulating effect on the
A-cell than a solution at higher concentration. At the
same time, as expected, the response of B-cells to 10 and
100mM phosphate buffer was the opposite: a solution at a
higher concentration caused higher rates of firing.
Therefore, it can be concluded, that the best stimulus for
the antennal taste bristles’ A-cell in the ground beetle
P. oblongopunctatus is not salt but the pH level of the
stimulus solution. The present results confirm the
suggestion made by Paje & Mossakowski (1984) that, in
ground beetles, pH receptors are located on the antennae.
To date, pH receptors have not been described in other
insects, and it is possible that the pH of the stimulating
solution may prove to be an important stimulus for other
chemoreceptors.
If the response of the A-cell depends on solution pH level
only, there should be equivalent responses to buffers at
different concentrations at the same pH. However, the
results do not confirm this: 10mM buffer had a greater
Fig. 7. Responses of antennal taste sensilla to 100 and 10mM phosphate buffers (pH measured 7.6 and 7.8, respectively). A- and B-cells
responded to changes in buffer solution concentration in opposite manner. By contrast to A-cells, in B-cells, buffer solution at higher
concentration was a more effective stimulus, especially during the first 2 s of the response. Asterisk (*) and n.s. inidcate whether differences
between mean firing rates were statistically significant or not, respectively (paired t-test, P< 0.05).
pH receptors in P. oblongopunctatus 131
# 2005 The Royal Entomological Society, Physiological Entomology, 30, 122–133

65
stimulating effect on the A-cell than 100mM buffer. This
phenomenon can be explained by interaction of different
chemical stimuli to chemosensory cells. Other chemicals, as
well as the pH of stimulating solution, may influence the
responses of insect taste receptors to one or another stimu-
lating compound (Hanson, 1987; Liscia et al., 1997; Bernays
et al., 1998; Liscia & Solari, 2000; Bernays & Chapman,
2001b; Schoonhoven & van Loon, 2002). Presumably,
salts of stimulating buffer solutions at high concentration
suppressed the neural activity of pH cell when stimulated
with 100mM phosphate buffer in 10 and 100mM phosphate
buffer experiments in P. oblongopunctatus.
The pH cell of antennal taste sensilla in P. oblongopunc-
tatus does not fire, or fires at very low frequency (first
second response below 5 imp/s) at pH3–6. This acid range
of pH values most likely represents the pH values in the
forest habitat and hibernating sites of this ground beetle
(Thiele, 1977; Paje & Mossakowski, 1984).
It is noteworthy that the action potentials registered by
antennal taste bristles’ cation and pH cells in the ground
beetle P. oblongopuctatus are negative and positive in their
polarity, respectively. Furthermore, other electrophysiolo-
gists studying insects’ chemoreceptors have noticed that
spikes in sensillum tip-recording technique (Hodgson
et al., 1955) may have positive as well as negative polarity
(Hanson & Wolbarsht, 1962; Dethier & Hanson,
1965; Elizarov & Sinitzina, 1974; Elizarov, 1978; Hansen-
Delkeskamp, 2001; Albert, 2003). Because the sensillum is
an integral part of the recording circuit, any cellular
properties and changes may contribute to uncontrolled
variability in the shape of the recorded action potentials
(Frazier & Hanson, 1986). At present, there appears to be
no satisfactory explanation for this phenomenon.
Acknowledgements
Contract grant sponsor: Estonian Science Foundation;
Contract grant numbers: 5423 and 4105.
References
Albert, P.J. (2003) Electrophysiological responses to sucrose from a
gustatory sensillum on the larval maxillary palp of the spruce
budworm, Choristoneura fumiferana (Clem.) (Lepidoptera:
Tortricidae). Journal of Insect Physiology, 49, 733–738.
Bernays, E.A. & Chapman, R.F. (2001a) Taste cell responses in the
polyphagous arctiid,Grammia geneura: towards a general pattern
for caterpillars. Journal of Insect Physiology, 47, 1029–1043.
Bernays, E.A. & Chapman, R.F. (2001b) Electrophysiological
responses of taste cells to nutrient mixtures in the polyphagous
caterpillar of Grammia geneura. Journal of Comparative Physiol-
ogy A, 187, 205–213.
Bernays, E.A., Glendinning, J.I. & Chapman, R.F. (1998) Plant
acids modulate chemosensory responses in Manduca sexta
larvae. Physiological Entomology, 23, 193–201.
Blaney, W.M. (1974) Electrophysiological responses of the terminal
sensilla on the maxillary palps of Locusta migratoria (L.) to some
electrolytes and non-electrolytes. Journal of Experimental
Biology, 60, 275–293.
Daly, P.J. & Ryan, M.F. (1979) Ultrastructure of antennal sensilla
of Nebria brevicollis (Fab.) (Coleoptera: Carabidae). Interna-
tional Journal of Insect Morphology and Embryology, 8, 169–181.
den Otter, C.J. (1972) Differential sensitivity of insect chemo-
receptors to alkali cations. Journal of Insect Physiology, 18,
109–131.
Dethier, V.G. (1977) The taste of salts. American Scientist, 65,
744–751.
Dethier, V.G. & Hanson, F.E. (1965) Taste papillae of the blowfly.
Journal of Cellular and Comparative Physiology, 65, 93–100.
Dethier, V.G. & Hanson, F.E. (1968) Electrophysiological
responses of the blowfly to sodium salts of fatty acids.
Proceedings of the National Academy of Sciences, U.S.A., 60,
1269–1303.
Elizarov, Y.A. (1978) Insect Chemoreception. Moscow University
Press, Russia.
Elizarov, Y.A. & Sinitzina, E.E. (1974) Contact chemoreceptors in
Aedes aegypti L. Zoological Journal, 53, 577–584 (in Russian).
Evans, D.R. & Mellon, D. (1962) Stimulation of a primary taste
receptor by salts. Journal of General Physiology, 45, 651–661.
Frazier, J.L. & Hanson, F.E. (1986) Electrophysiological recording
and analysis of insect chemosensory responses. Insect–Plant
Interactions (ed. by T. A. Miller and J. Miller), pp. 285–330.
Springer, Germany.
Hansen-Delkeskamp, E. (2001) Responsiveness of antennal taste
hairs of the apterygotan insect, Thermobia domestica (Zygento-
ma); an electrophysiological investigation. Journal of Insect
Physiology, 47, 689–697.
Hanson, F.E. (1987) Chemoreception in the fly: the search for
the liverwurst receptor. Perspectives in Chemoreception
and Behaviour (ed. by R. F. Chapman, E. A. Bernays and
J. G. Stoffolano), pp. 99–122. Springer, Germany.
Hanson, F.E. & Wolbarsht, K.L. (1962) Dendritic action potentials
in insect chemoreceptors. American Zoologist, 2, 528–534.
Haskell, P.T. & Schoonhoven, L.M. (1969) The function of certain
mouth part receptors in relation to feeding in Schistocerca
gregaria and Locusta migratoria migratorioides. Entomologia
Experimentalis et Applicata, 12, 429–440.
Hodgson, E.S., Lettvin, J.Y. & Roeder, K.D. (1955) Physiology of
a primary chemoreceptor unit. Science, 122, 417–418.
Ishikawa, S. (1963) Responses of maxillary chemoreceptors in the
larvae of the silkworm, Bombyx mori, to stimulation by
carbohydrates. Journal of Cellular and Comparative Physiology,
61, 99–107.
Kim, J.L. & Yamasaki, T. (1996) Sensilla of Carabus (Isiocarabus)
fiduciarius saishutoicus Csiki (Coleoptera: Carabidae). Interna-
tional Journal of Insect Morphology and Embryology, 25,
153–172.
Liscia, A. & Solari, P. (2000) Bitter taste recognition in the blowfly:
Electrophysiological and behavioral evidence. Physiology and
Behaviour, 70, 61–65.
Liscia, A., Solari, P., Majone, R., Tomassini Barbarossa, I. &
Crnjar, R. (1997) Taste receptor mechanisms in the blowfly:
evidence of amiloride-sensitive and insensitive receptor sites.
Physiology and Behavior, 62, 875–879.7.
Merivee, E., Ploomi, A., Rahi, M., Luik, A. & Sammelselg, V.
(2000) Antennal sensilla of the ground beetle Bembidion lampros
Hbst (Coleoptera, Carabidae). Acta Zoologica (Stockholm), 81,
339–350.
132 E. Merivee et al.
# 2005 The Royal Entomological Society, Physiological Entomology, 30, 122–133

66
Merivee, E., Ploomi, A., Luik, A., Rahi, M. & Sammelselg, V.
(2001) Antennal sensilla of the ground beetle Platynus dorsalis
(Pontoppidan, 1763) (Coleoptera, Carabidae). Microscopy
Research and Technique, 55, 339–349.
Merivee, E., Ploomi, A., Rahi, M., Bresciani, J., Ravn, H.P., Luik, A. &
Sammelselg,V. (2002)Antennal sensilla of the groundbeetleBembidion
properans Steph. (Coleoptera, Carabidae).Micron, 33, 429–440.
Mitchell, B.K. & Schoonhoven, L.M. (1973) Taste receptors
in Colorado beetle larvae. Journal of Insect Physiology, 20,
1787–1793.
Mitchell, B.K. & Seabrook, W.D. (1973) Electrophysiological
investigations on tarsal chemoreceptors of the spruce budworm,
Choristoneura fumiferana (Lepidoptera). Journal of Insect
Physiology, 20, 1209–1218.
Paje, F. & Mossakowski, D. (1984) pH-preferences and habitat
selection in carabid beetles. Oecologia (Berlin), 64, 41–46.
Schoonhoven, L.M. & van Loon, J.J.A. (2002) An inventory of
taste in caterpillars: each species its own key. Acta Zoologica
Academiae Scientiarum Hungarica, 48, 215–263.
Shiraishi, A. & Morita, H. (1969) The effects of pH on the labellar
sugar receptor of the fleshfly. Journal of General Physiology, 53,
450–470.
Thiele, H.-U. (1977) Carabid beetles in their environment.
Zoophysiology and Ecology 10. Springer, Germany.
Accepted 20 October 2004
pH receptors in P. oblongopunctatus 133
# 2005 The Royal Entomological Society, Physiological Entomology, 30, 122–133
II

Milius, M., Merivee, E., Williams, I., Luik, A., Mänd, M., Must, A.,
2006.
A NEW METHOD FOR ELECTROPHYSIOLOGICAL
IDENTIFICATION OF ANTENNAL PH RECEPTOR CELLS
IN GROUND BEETLES: THE EXAMPLE OF PTEROSTICHUS
AETHIOPS (PANZER, 1796) (COLEOPTERA, CARABIDAE).
Journal of Insect Physiology 52, 960–967.

69
Journal of Insect Physiology 52 (2006) 960–967
A new method for electrophysiological identification of antennal pHreceptor cells in ground beetles: The example of Pterostichus aethiops
(Panzer, 1796) (Coleoptera, Carabidae)
Marit Miliusa, Enno Meriveea,�, Ingrid Williamsb, Anne Luika, Marika Manda, Anne Musta
aEstonian University of Life Sciences, Institute of Agricultural and Environmental Sciences, 64 Kreutzwaldi Street, 51014 Tartu, EstoniabRothamsted Research, Harpenden, Hertfordshire, AL5 2JQ, UK
Received 20 March 2006; received in revised form 2 June 2006; accepted 7 June 2006
Abstract
The responses of antennal taste sensilla of the ground beetle Pterostichus aethiops to 100mM Na+-salts and their mixtures with 1 and
10mM NaOH were compared. An increase in pH by 0.3–0.6 units in 100mM Na+-salt solutions, caused by the content of 1mM NaOH,
was too small, except for alkaline Na2HPO4, to influence the firing rate of the cation cell and pH cell significantly. However, different
sensitivity of the two cells to increased pH was clearly demonstrated when the concentration of NaOH in 100mM stimulating salt
solutions was increased to 10mM. Increasing pH by 1.2–2 units caused the 1st s firing rate to increase by 140–1050% and 0–26% in the
pH cell and cation cell, respectively. Compared to the buffer series method used for identification of the pH receptors in ground beetles
earlier, considerably stronger responses of the pH cell to a similar increase in pH were observed when the NaOH method was used for
testing. At the same time, undesirable changes in salt ions concentration that occur when stimulating solutions differing by 1–2 pH units
are prepared were much smaller using the latter method. Behavioural and ecological relevance of the results is discussed.
r 2006 Elsevier Ltd. All rights reserved.
Keywords: Taste sensilla; Tip recording; Action potential rate; Predatory insects; Habitat preferences
1. Introduction
The ground beetles (Coleoptera, Carabidae) with 40,000species belong to the most numerous families of beetles,their distribution being cosmopolitan. Approximately 430species have been found in Fennoscandia, Denmark andBaltic countries, 530 species occur in Czech Republic(Lovei and Sunderland, 1996; Kula and Purchart, 2004;Silferberg, 2004). Their bio-indication importance isderived from their dependence on specific biotic andabiotic conditions of the site: food availability, competi-tors, vegetation type, humidity, light and temperatureconditions, soil pH and salt content etc. Habitat choice isso specific that carabids are often used to characterizehabitats (Thiele, 1977; Lovei and Sunderland, 1996).
External stimuli crucial in habitat choice of ground beetleshave been purely studied on the receptor level, however.Preferences for particular soil pH, varying between 3 and
9, in ground beetles were discovered using laboratorychoice experiments some decades ago (Krogerus, 1960;Paje and Mossakowski, 1984). More recently, in ecologicalfield experiments, a strong correlation between theabundance of some ground beetles and soil pH has beenfound (Irmler, 2001, 2003). Electrophysiological studieswith alkaline, neutral and acid salts have indicated that inaddition to the salt (cation) cell, the pH cell also probablyinnervates the antennal taste bristles in the ground beetlePterostichus aethiops (Panzer, 1796) (Merivee et al., 2004).Experiments with acetate and phosphate buffer series in thepH range of 3–11 confirmed this assumption in the groundbeetle Pterostichus oblongopunctatus (Fabricius, 1787)(Merivee et al., 2005). The pH cell discovered seems to bespecific to ground beetles, as it is not found in other insects.In order to explain the role of this cell in the beetles’
ARTICLE IN PRESS
www.elsevier.com/locate/jinsphys
0022-1910/$ - see front matter r 2006 Elsevier Ltd. All rights reserved.
doi:10.1016/j.jinsphys.2006.06.003
�Corresponding author.
E-mail address: [email protected] (E. Merivee).

70
behaviour more precisely, further electrophysiologicalstudies on the distribution and functioning of the pH-receptive cell in ground beetles are justified.
P. aethiops is a European species found from the BritishIsles east to Perm in Russia, south to Serbia and north intosouthern Fennoscandia and Finland. In mountains ofCentral and Eastern Europe their abundance considerablyincreases with increasing altitude up to subalpine zones.This spring-breeding, wingless, eurytopic ground beetlepopulates wet coniferous, deciduous as well as mixedforests with acid soil pH. Also, its preferred overwinteringsites in brown-rotted wood laying on the ground where, inautumn, the beetles concentrate in great numbers arestrongly acid (pH 3–5; Merivee, unpublished data).Frequently, hibernating beetles have been found in mossand litter. In its preferred habitats, it is one of the mostnumerous ground beetles (Haberman, 1968; Thiele, 1977;Lindroth, 1985, 1986; Wachmann et al., 1995; Ermakov,2004; Kula and Purchart, 2004; Silferberg, 2004; Sk"o-dowski, 2005).
In P. oblongopunctatus, it was demonstrated that the pHcell did not fire or fired at very low frequency at pH 3–6,first s response below 5 imp/s declining during the next fours of the stimulation close to zero. This level of acidityprobably represents the pH preferences of this groundbeetle in its forest habitat and hibernating sites (Meriveeet al., 2005). In this study we show that similar correlationsbetween electrophysiological responses of the pH cell andpH of the preferred habitats may occur also in the groundbeetle P. aethiops.
The limitation of the buffer series method used fortesting the responses of pH receptor cells to pH in groundbeetles earlier (Merivee et al., 2005) is that buffers withdifferent pH vary largely in both pH and electrolyteconcentration. The NaOH method reported here allows toprepare stimulating salt solutions over a wide range of pHwhereas changes in electrolyte concentrations are minimal.Over the dynamic range of the stimulus/response curve, theaction potential rate of a taste cell is a logarithmic functionof the stimulus concentration. The maximum firing rate ofa phasic-tonically responding taste cell evoked by strongstimuli does usually not exceed 40–50 imp/s (Evans andMellon, 1962; Hanson, 1987; Merivee et al., 2004).Consequently, a 1–10% increase in concentration ofthe stimulating salt solution causes a change of less than1 imp/s in the firing rate of the taste cell, which is hardlydetectable in electrophysiological experiments. These cal-culations were taken into account when designing thecurrent experiments.
To test for the presence of the pH cell in the antennaltaste bristles of the ground beetle P. aethiops moreprecisely, three solutions of 100mM Na+-salts, Na2HPO4,NaCl and NaH2PO4 were prepared, acid, neutral andalkaline, respectively; these served as control stimuli. Teststimuli were prepared by adding small amounts of NaOHto the 100mM salts resulting in three electrolyte mixtureswith 1mM NaOH and three mixtures with 10mM NaOH.
Addition of NaOH to these salt solutions does not changetheir ionic composition and the Na+ concentrationincreases only to a small extent (1% and 10%, respec-tively). At the same time, the pH level of these solutionsrises substantially. The stimulating effect of these sodiumsalt solutions with and without NaOH was expected to beapproximately similar for the salt cell but not for the pHcell if present. Results of these experiments are reported inthis paper. The size, number and location of antennal tastebristles in the ground beetle P. aethiops have beendescribed earlier (Merivee et al., 2004).
2. Material and methods
2.1. Test beetles
The specimens of P. aethiops used in the experimentswere collected in May 2005 in southern Estonia. They werestored in plastic boxes filled with moistened moss and sandin a refrigerator at +5 1C. Two or three days beforeelectrophysiological tests the beetles were transferred tosimilar boxes at room temperature (+22 1C). They weregiven clean water to drink and fed with moistened cat food(Kitekat, Master Foods, Poland) every day.For the electrophysiological tests, each test beetle was
restrained by placing it into a conical tube made of thinsheet of aluminium of a size that allowed its head andantennae to protrude a little from the narrower end. Thewider rear end of the conical tube was closed with a piece ofplasticine to prevent the beetle from retreating out of thetube. The antennae of the restrained beetle were fastenedhorizontally on the edge of a special aluminium stand withtiny amounts of beeswax, so that horizontally locatedantennal taste bristles were visible from above under a lightmicroscope, and easily accessible for micromanipulationsfrom the side. Contamination of tested taste bristles andtheir contact with preparation instruments and beeswaxwas carefully avoided.
2.2. Chemicals and preparation of stimulating solutions
Chemicals of analytical grade of purity used in theexperiments were obtained from AppliChem (Germany).Stimulating test solutions prepared are shown in Table 1.pH of the solutions ranging from 4.6 to 10.6 was measuredwith the pH meter E6115 (Evikon, Estonia).
2.3. Stimulation and recording of action potentials
Electrophysiological recordings were made from thelarge chaetoid taste sensilla on the antennal flagellum of therestrained beetle using a tip-recording technique (Hodgsonet al., 1955). To achieve a good signal-to-noise ratio andavoid registration of muscular activities from two basalantennomeres, the scape and pedicel and from other partsof insect’s body, an indifferent tungsten microelectrode wasinserted into the base of the flagellum. The recording
ARTICLE IN PRESSM. Milius et al. / Journal of Insect Physiology 52 (2006) 960–967 961
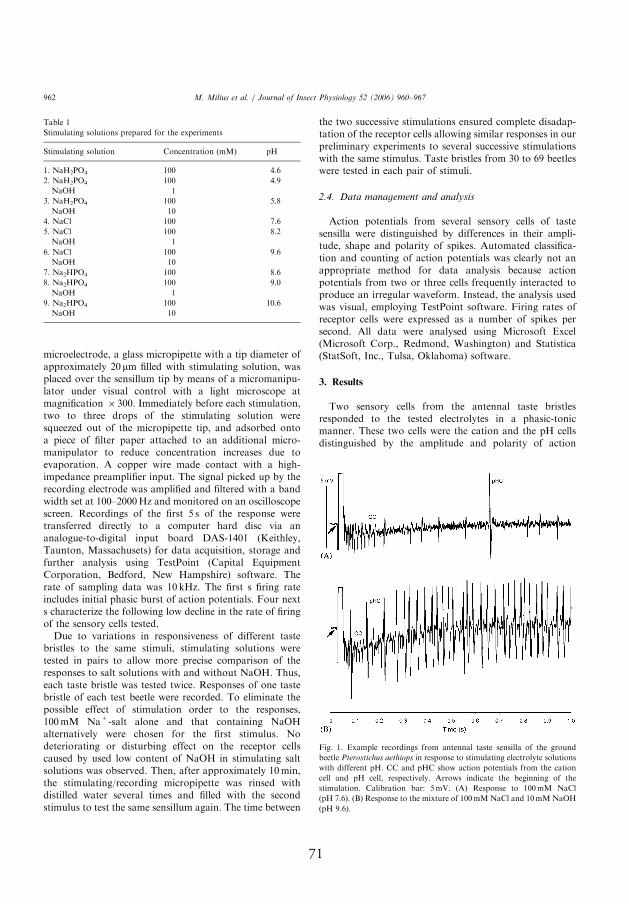
71
microelectrode, a glass micropipette with a tip diameter ofapproximately 20 mm filled with stimulating solution, wasplaced over the sensillum tip by means of a micromanipu-lator under visual control with a light microscope atmagnification � 300. Immediately before each stimulation,two to three drops of the stimulating solution weresqueezed out of the micropipette tip, and adsorbed ontoa piece of filter paper attached to an additional micro-manipulator to reduce concentration increases due toevaporation. A copper wire made contact with a high-impedance preamplifier input. The signal picked up by therecording electrode was amplified and filtered with a bandwidth set at 100–2000Hz and monitored on an oscilloscopescreen. Recordings of the first 5 s of the response weretransferred directly to a computer hard disc via ananalogue-to-digital input board DAS-1401 (Keithley,Taunton, Massachusets) for data acquisition, storage andfurther analysis using TestPoint (Capital EquipmentCorporation, Bedford, New Hampshire) software. Therate of sampling data was 10 kHz. The first s firing rateincludes initial phasic burst of action potentials. Four nexts characterize the following low decline in the rate of firingof the sensory cells tested.
Due to variations in responsiveness of different tastebristles to the same stimuli, stimulating solutions weretested in pairs to allow more precise comparison of theresponses to salt solutions with and without NaOH. Thus,each taste bristle was tested twice. Responses of one tastebristle of each test beetle were recorded. To eliminate thepossible effect of stimulation order to the responses,100mM Na+-salt alone and that containing NaOHalternatively were chosen for the first stimulus. Nodeteriorating or disturbing effect on the receptor cellscaused by used low content of NaOH in stimulating saltsolutions was observed. Then, after approximately 10min,the stimulating/recording micropipette was rinsed withdistilled water several times and filled with the secondstimulus to test the same sensillum again. The time between
the two successive stimulations ensured complete disadap-tation of the receptor cells allowing similar responses in ourpreliminary experiments to several successive stimulationswith the same stimulus. Taste bristles from 30 to 69 beetleswere tested in each pair of stimuli.
2.4. Data management and analysis
Action potentials from several sensory cells of tastesensilla were distinguished by differences in their ampli-tude, shape and polarity of spikes. Automated classifica-tion and counting of action potentials was clearly not anappropriate method for data analysis because actionpotentials from two or three cells frequently interacted toproduce an irregular waveform. Instead, the analysis usedwas visual, employing TestPoint software. Firing rates ofreceptor cells were expressed as a number of spikes persecond. All data were analysed using Microsoft Excel(Microsoft Corp., Redmond, Washington) and Statistica(StatSoft, Inc., Tulsa, Oklahoma) software.
3. Results
Two sensory cells from the antennal taste bristlesresponded to the tested electrolytes in a phasic-tonicmanner. These two cells were the cation and the pH cellsdistinguished by the amplitude and polarity of action
ARTICLE IN PRESS
Table 1
Stimulating solutions prepared for the experiments
Stimulating solution Concentration (mM) pH
1. NaH2PO4 100 4.6
2. NaH2PO4 100 4.9
NaOH 1
3. NaH2PO4 100 5.8
NaOH 10
4. NaCl 100 7.6
5. NaCl 100 8.2
NaOH 1
6. NaCl 100 9.6
NaOH 10
7. Na2HPO4 100 8.6
8. Na2HPO4 100 9.0
NaOH 1
9. Na2HPO4 100 10.6
NaOH 10
Fig. 1. Example recordings from antennal taste sensilla of the ground
beetle Pterostichus aethiops in response to stimulating electrolyte solutions
with different pH. CC and pHC show action potentials from the cation
cell and pH cell, respectively. Arrows indicate the beginning of the
stimulation. Calibration bar: 5mV. (A) Response to 100mM NaCl
(pH 7.6). (B) Response to the mixture of 100mMNaCl and 10mMNaOH
(pH 9.6).
M. Milius et al. / Journal of Insect Physiology 52 (2006) 960–967962
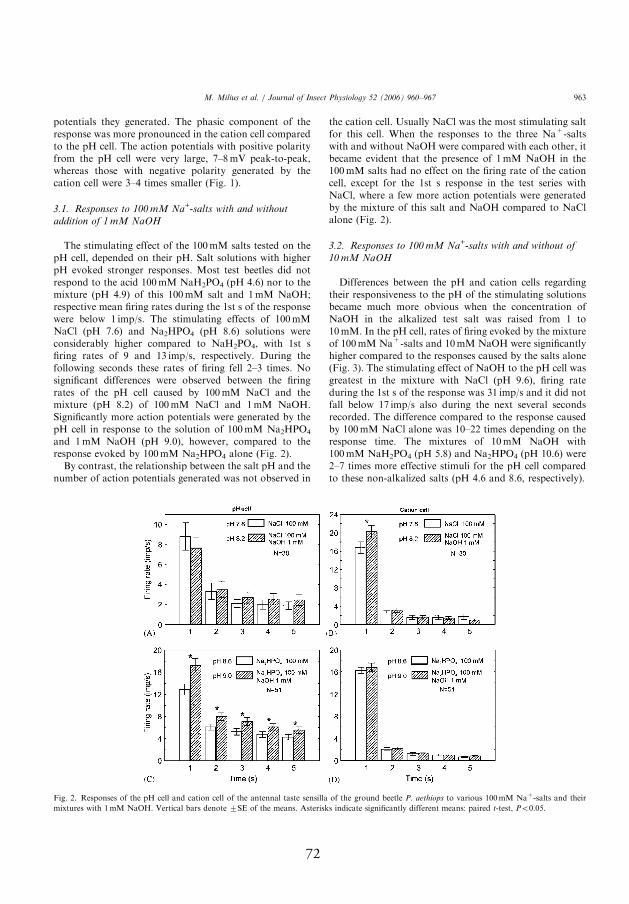
72
potentials they generated. The phasic component of theresponse was more pronounced in the cation cell comparedto the pH cell. The action potentials with positive polarityfrom the pH cell were very large, 7–8mV peak-to-peak,whereas those with negative polarity generated by thecation cell were 3–4 times smaller (Fig. 1).
3.1. Responses to 100mM Na+-salts with and without
addition of 1mM NaOH
The stimulating effect of the 100mM salts tested on thepH cell, depended on their pH. Salt solutions with higherpH evoked stronger responses. Most test beetles did notrespond to the acid 100mM NaH2PO4 (pH 4.6) nor to themixture (pH 4.9) of this 100mM salt and 1mM NaOH;respective mean firing rates during the 1st s of the responsewere below 1 imp/s. The stimulating effects of 100mMNaCl (pH 7.6) and Na2HPO4 (pH 8.6) solutions wereconsiderably higher compared to NaH2PO4, with 1st sfiring rates of 9 and 13 imp/s, respectively. During thefollowing seconds these rates of firing fell 2–3 times. Nosignificant differences were observed between the firingrates of the pH cell caused by 100mM NaCl and themixture (pH 8.2) of 100mM NaCl and 1mM NaOH.Significantly more action potentials were generated by thepH cell in response to the solution of 100mM Na2HPO4
and 1mM NaOH (pH 9.0), however, compared to theresponse evoked by 100mM Na2HPO4 alone (Fig. 2).
By contrast, the relationship between the salt pH and thenumber of action potentials generated was not observed in
the cation cell. Usually NaCl was the most stimulating saltfor this cell. When the responses to the three Na+-saltswith and without NaOH were compared with each other, itbecame evident that the presence of 1mM NaOH in the100mM salts had no effect on the firing rate of the cationcell, except for the 1st s response in the test series withNaCl, where a few more action potentials were generatedby the mixture of this salt and NaOH compared to NaClalone (Fig. 2).
3.2. Responses to 100mM Na+-salts with and without of
10mM NaOH
Differences between the pH and cation cells regardingtheir responsiveness to the pH of the stimulating solutionsbecame much more obvious when the concentration ofNaOH in the alkalized test salt was raised from 1 to10mM. In the pH cell, rates of firing evoked by the mixtureof 100mM Na+-salts and 10mM NaOH were significantlyhigher compared to the responses caused by the salts alone(Fig. 3). The stimulating effect of NaOH to the pH cell wasgreatest in the mixture with NaCl (pH 9.6), firing rateduring the 1st s of the response was 31 imp/s and it did notfall below 17 imp/s also during the next several secondsrecorded. The difference compared to the response causedby 100mM NaCl alone was 10–22 times depending on theresponse time. The mixtures of 10mM NaOH with100mM NaH2PO4 (pH 5.8) and Na2HPO4 (pH 10.6) were2–7 times more effective stimuli for the pH cell comparedto these non-alkalized salts (pH 4.6 and 8.6, respectively).
ARTICLE IN PRESS
Fig. 2. Responses of the pH cell and cation cell of the antennal taste sensilla of the ground beetle P. aethiops to various 100mM Na+-salts and their
mixtures with 1mM NaOH. Vertical bars denote 7SE of the means. Asterisks indicate significantly different means: paired t-test, Po0.05.
M. Milius et al. / Journal of Insect Physiology 52 (2006) 960–967 963
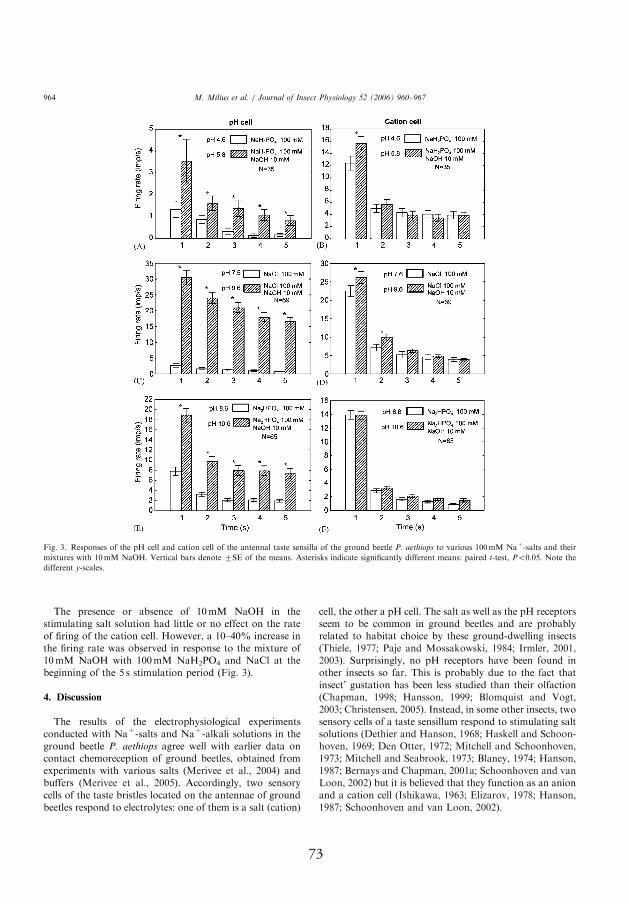
73
The presence or absence of 10mM NaOH in thestimulating salt solution had little or no effect on the rateof firing of the cation cell. However, a 10–40% increase inthe firing rate was observed in response to the mixture of10mM NaOH with 100mM NaH2PO4 and NaCl at thebeginning of the 5 s stimulation period (Fig. 3).
4. Discussion
The results of the electrophysiological experimentsconducted with Na+-salts and Na+-alkali solutions in theground beetle P. aethiops agree well with earlier data oncontact chemoreception of ground beetles, obtained fromexperiments with various salts (Merivee et al., 2004) andbuffers (Merivee et al., 2005). Accordingly, two sensorycells of the taste bristles located on the antennae of groundbeetles respond to electrolytes: one of them is a salt (cation)
cell, the other a pH cell. The salt as well as the pH receptorsseem to be common in ground beetles and are probablyrelated to habitat choice by these ground-dwelling insects(Thiele, 1977; Paje and Mossakowski, 1984; Irmler, 2001,2003). Surprisingly, no pH receptors have been found inother insects so far. This is probably due to the fact thatinsect’ gustation has been less studied than their olfaction(Chapman, 1998; Hansson, 1999; Blomquist and Vogt,2003; Christensen, 2005). Instead, in some other insects, twosensory cells of a taste sensillum respond to stimulating saltsolutions (Dethier and Hanson, 1968; Haskell and Schoon-hoven, 1969; Den Otter, 1972; Mitchell and Schoonhoven,1973; Mitchell and Seabrook, 1973; Blaney, 1974; Hanson,1987; Bernays and Chapman, 2001a; Schoonhoven and vanLoon, 2002) but it is believed that they function as an anionand a cation cell (Ishikawa, 1963; Elizarov, 1978; Hanson,1987; Schoonhoven and van Loon, 2002).
ARTICLE IN PRESS
Fig. 3. Responses of the pH cell and cation cell of the antennal taste sensilla of the ground beetle P. aethiops to various 100mM Na+-salts and their
mixtures with 10mM NaOH. Vertical bars denote 7SE of the means. Asterisks indicate significantly different means: paired t-test, Po0.05. Note the
different y-scales.
M. Milius et al. / Journal of Insect Physiology 52 (2006) 960–967964

74
In addition to the specific stimulus, also frequentlynamed the best stimulus, other chemical factors such asionic composition and concentration of electrolytes, thepresence of non-electrolytes, and pH of the stimulatingsolution, may all influence the response of different kindsof insect contact chemoreceptive cells to some extent(Hanson, 1987; Liscia et al., 1997; Bernays et al., 1998;Liscia and Solari, 2000; Bernays and Chapman, 2001b;Schoonhoven and van Loon, 2002). In this context, it is notsurprising that the response of the antennal cation cell ofP. aethiops, in some cases, was also increased by 17–40%by pH increase up to two pH units of the stimulatingsolution, which is some thirty times smaller increase infiring rate than was observed in the pH cell responses underthe same stimulating conditions. Similar stimulatory effectof pH to the cation cell responses was observed inelectrophysiological experiments with acetate and phos-phate buffers also in the ground beetle P. oblongopunctatus(Merivee et al., 2005).
Our tests demonstrated that an increase in pH by 0.3–0.6units in 100mM Na+-salt solutions, caused by the 1mMNaOH content, was too small, except for Na2HPO4, toinfluence the firing rate of the pH cell or cation cellsignificantly. However, different sensitivity of the two cellsto increased pH was clearly demonstrated when theconcentration of NaOH in 100mM stimulating saltsolutions was increased to 10mM resulting in pH increaseby 1.2–2 units depending on the salt. Compared to theacetate and phosphate buffers method (Merivee et al.,2005), a comparable increase in pH by 1–2 units of thestimulatory salt solution using the method of adding smallamounts of NaOH to 100mM Na+-salts evoked aconsiderably larger increase in the rate of firing of the pHcell: respective rates of firing during the 1st s of theresponse increased by 30–400% and 140–1050%, respec-tively, depending on buffers and salts used.
In the stimulating solutions differing by 1.2–2 pH units,and using the NaOH method, the difference in Na+
concentrations is only 10% and the salt anion concentra-tion does not change. By contrast, using the method ofbuffers, respective concentrations of content ions maydiffer up to 30 times, depending on the pH range and saltsinvolved. Taking into consideration that both salt cationsand anions of the stimulating solution may affect the pHcell firing rate, large differences in the concentration of saltions are undesirable when testing pH cell responses to pH.Thus, it seems that the NaOH method is, besides thebuffers method, a reliable tool for studying pH receptorfunctioning in insects.
Our results suggest that in P. aethiops, in its preferredacid forest habitats and overwintering sites in brown-rottedwood at pH 3–5, the antennal pH sensitive cell does notdischarge or discharges at very low frequency with the firsts firing rate close to 1 imp/s or lower. Areas with a higherpH seem to be unfavourable to this insect and whencontacted the pH cell signals with a stronger response. Thismay occur, for example, in places with decaying plant
material on the soil surface. Many of the materialscomposted contain significant amounts of protein, whichis converted to ammonia toxic to many organisms andserving as the primary substrate in the nitrificationprocesses. Nitrification processes have been observed atpH levels ranging from 6.6 to 9.7 (Odell et al., 1996).Obviously, the ground beetle P. aethiops avoids theseplaces with a high pH.Six to seven months, from September to April, adults of
P. aethiops may spend in its preferred overwintering sites inbrown-rotted wood in Estonia. Fungi often lower the pHof the extracellular medium through secretion of organicacids and thus establish a pH gradient around themycelium. The organic acid oxalate is produced by most,if not all, wood-degrading fungi. Usually, pH 3–6 isobserved in both white- and brown-rotted wood. Stablemoisture content is quaranteed as a result of metabolicwater production during the wood-degrading process. Forcultures of white-rot fungus Merulius tremellosus incubatedin air, the optimum water content was 2 g/1 g of wood.Moister conditions were less favourable to delignification,possibly because the water impeded aeration (Reid, 1985;Varela and Tien, 2003). Similar prolonged stability ofwater content and aeration is probably also quaranteed inbrown-rotting wood which seems to be a favourablemicroenvironment to ground beetles for their prolongedhibernation.Brown-rotted wood may offer good protection for
hibernating ground beetles against entomopathogenicfungi and parasites which play an important role in theirpopulation dynamic. Up to 41% parasitism by nematodesand ectoparasitic fungi was found on 14 species ofBembidion in Norway depending on habitat selectionof the host (Andersen and Skorping, 1991). Eggs ofP. oblongopunctatus suffered 83% mortality in fresh litterbut only 7% in sterilized soil (Heessen, 1981). Althoughpredators, parasites, and pathogens affect all ground beetledevelopmental stages, quantitative data remain scarce. So,antennal pH receptors of the ground beetles P. aethiops
seem to be a powerful tool in their selection for favourableacid habitats and microhabitats crucial to survive.Chemically mediated habitat selection has been shown to
occur in several species of ground beetles by Evans (1982,1983, 1984, 1988). He demonstrated that olfactory cellsreceptive to methyl esters of palmitic and oleic acid emittedby the mat-forming blue-green algae Oscillatoria animalis
Agardh and Oscillatoria subbrevis Schmidle are present insensillae on the antennae of those Bembidiini associatedwith the shores of saline lakes. Low concentrations offormic acid, a defence secretion in the ground beetleHarpalus rufipes, was shown by Luff (1986) to causeaggregation in this species. Formic acid is also produced bysome wood-decaying fungi as a result of enzymaticdecarboxylation of oxalic acid accumulating in largequantities in the cell sap of various plants and as a majorproduct of carbohydrate metabolism by many molds(Shimazono and Hayaishi, 1957). Probably, formic acid
ARTICLE IN PRESSM. Milius et al. / Journal of Insect Physiology 52 (2006) 960–967 965

75
could be perceived and used for long-distance orientationand aggregation in brown-rotted wood by P. aethiops aswell.
Acknowledgements
Contract grant sponsor: Estonian Science Foundation;Contract Grant No. 5423 and 5736.
References
Andersen, J., Skorping, A., 1991. Parasites of carabid beetles: prevalence
depends on habitat selection of the host. Canadian Journal of Zoology
69, 1216–1220.
Bernays, E.A., Chapman, R.F., 2001a. Taste cell responses in the
polyphagous arctiid, Grammia geneura: towards a general pattern for
caterpillars. Journal Insect Physiology 47, 1029–1043.
Bernays, E.A., Chapman, R.F., 2001b. Electrophysiological responses
of taste cells to nutrient mixtures in the polyphagous caterpillar
of Grammia geneura. Journal of Comparative Physiology A 187,
205–213.
Bernays, E.A., Glendinning, J.I., Chapman, R.F., 1998. Plant acids
modulate chemosensory responses in Manduca sexta larvae. Physio-
logical Entomology 23, 193–201.
Blaney, W.M., 1974. Electrophysiological responses of the terminal
sensilla on the maxillary palps of Locusta migratoria (L.) to some
electrolytes and non-electrolytes. Journal of Experimental Biology 60,
275–293.
Blomquist, G., Vogt, R. (Eds.), 2003. Insect Pheromone Biochemistry and
Molecular Biology. The Synthesis and Detection of Pheromones and
Plant Volatiles. Elsevier, Academic Press, Amsterdam, Boston,
Heidelberg, London, New York, Oxford, Paris, San Diego, San
Francisco, Singapore, Sidney, Tokyo, p. 745.
Chapman, R.F., 1998. The Insects Structure and Function. Cambridge
University Press, Cambridge, p. 770.
Christensen, T.A. (Ed.), 2005. Methods in Insect Sensory Neuroscience.
CRC Press, Boca Raton, London, New York, Washington, p. 435.
Den Otter, C.J., 1972. Differential sensitivity of insect chemoreceptors to
alkali cations. Journal of Insect Physiology 18, 109–131.
Dethier, V.G., Hanson, F.E., 1968. Electrophysiological responses of the
blowfly to sodium salts of fatty acids. Proceedings of the National
Academy of Sciences, USA 60, 1269–1303.
Elizarov, Yu.A., 1978. Insect Chemoreception. Moscow University Press,
Moscow, p. 232. (in Russian).
Ermakov, A.I., 2004. Structural changes in the carabid fauna of forest
ecosystems under a toxic impact. Russian Journal of Ecology 35,
403–408.
Evans, D.R., Mellon, D., 1962. Stimulation of a primary taste receptor by
salts. Journal of General Physiology 45, 651–661.
Evans, W.G., 1982. Oscillatoria sp. (Cyanophyta) mat metabolites
implicated in habitat selection in Bembidion obtusidens (Coleopt.:
Carabidae). Journal of Chemical Ecology 8, 671–678.
Evans, W.G., 1983. Habitat selection in the Carabidae. Coleopterists
Bulletin 37, 164–167.
Evans, W.G., 1984. Odour-mediated responses of Bembidion obtusidens
(Coleoptera: Carabidae) in a wind tunnel. Canadian Journal of
Entomology 116, 1653–1658.
Evans, W.G., 1988. Chemically mediated habitat recognition in shore
insects (Coleoptera: Carabidae, Hemiptera: Saldidae). Journal of
Chemical Ecology 14, 1441–1454.
Haberman, H., 1968. Ground Beetles of Estonia. Valgus, Tallinn. p. 598
(in Estonian).
Hanson, F.E., 1987. Chemoreception in the fly: the search for the
liverwurst receptor. In: Chapman, R.F., Bernays, E.A., Stoffolano,
J.G. (Eds.), Perspectives in Chemoreception and Behaviour. Springer,
Berlin, Heidelberg, New York, pp. 99–122.
Hansson, B.S. (Ed.), 1999. Insect Olfaction. Springer, Berlin, Heidelberg,
New York, Barcelona, Hong Kong, London, Milan, Paris, Singapore,
Tokyo, p. 457.
Haskell, P.T., Schoonhoven, L.M., 1969. The function of certain mouth
part receptors in relation to feeding in Schistocerca gregaria and
Locusta migratoria migratorioides. Entomologia Experimentalis et
Applicata 12, 429–440.
Heessen, H.J.L., 1981. Egg mortality in P. oblongopunctatus (Coleoptera,
Carabidae). Oecologia 50, 233–235.
Hodgson, E.S., Lettvin, J.Y., Roeder, K.D., 1955. Physiology of a primary
chemoreceptor unit. Science 122, 417–418.
Irmler, U., 2001. Characterisierung der Laufkafergemeinschaften schles-
wig-holsteinischer Walder und Moglichkeiten ihrer okologischen
Bewertung. Angewandte Carabidologie. Laufkafer im Wald
(Suppl. 2), 21–32.
Irmler, U., 2003. The spatial and temporal pattern of carabid beetles on
arable fields in northern Germany (Schleswig–Holstein) and their value
as ecological indicators. Agriculture, Ecosystems & Environment 98,
141–151.
Ishikawa, S., 1963. Responses of maxillary chemoreceptors in the
larvae of the silkworm, Bombyx mori, to stimulation by carbohy-
drates. Journal of Cellular and Comparative Physiology 61,
99–107.
Krogerus, R., 1960. Okologische Studien uber nordische Moorarthropo-
den. Societas Scientiarum. Fennica, Commentationes Biologicae 21
(3), 1–238.
Kula, E., Purchart, L., 2004. The ground beetles (Coleoptera: Carabidae)
of forest altitudinal zones of the eastern part of the Krusne hory Mts.
Journal of Forest Science 50, 456–463.
Lindroth, C.H., 1985. The Carabidae (Coleoptera) of Fennoscandia and
Denmark. Fauna Entomologica Scandinavica 15, 1–225.
Lindroth, C.H., 1986. The Carabidae (Coleoptera) of Fennoscandia and
Denmark. Fauna Entomologica Scandinavica 15, 233–497.
Liscia, A., Solari, P., 2000. Bitter taste recognition in the blowfly:
electrophysiological and behavioral evidence. Physiology & Behavior
70, 61–65.
Liscia, A., Solari, P., Majone, R., Tomassini Barbarossa, I., Crnjar, R.,
1997. Taste receptor mechanisms in the blowfly: evidence of amiloride-
sensitive and insensitive receptor sites. Physiology & Behavior 62,
875–879.
Lovei, G.L., Sunderland, K.D., 1996. Ecology and behavior of ground
beetles (Coleoptera:Carabidae). Annual Review of Entomology 41,
231–256.
Luff, M.L., 1986. Aggregation of some Carabidae in pitfall traps. In: den
Boer, P.J. (Ed.), Carabid Beetles. Gustav Fisher, Stuttgart,
pp. 385–397.
Merivee, E., Renou, M., Mand, M., Luik, A., Heidemaa, M., Ploomi, A.,
2004. Electrophysiological responses to salts from antennal chaetoid
taste sensilla of the ground beetle Pterostichus aethiops. Journal of
Insect Physiology 50, 1001–1013.
Merivee, E., Ploomi, A., Milius, M., Luik, A., Heidemaa, M., 2005.
Electrophysiological identification of antennal pH-receptors in the
ground beetle Pterostichus oblongopunctatus (Coleoptera, Carabidae).
Physiological Entomology 30, 122–133.
Mitchell, B.K., Schoonhoven, L.M., 1973. Taste receptors in Colorado
beetle larvae. Journal of Insect Physiology 20, 1787–1793.
Mitchell, B.K., Seabrook, W.D., 1973. Electrophysiological investiga-
tions on tarsal chemoreceptors of the spruce budworm,
Choristoneura fumiferana (Lepidoptera). Journal of Insect Physiology
20, 1209–1218.
Odell, L.H., Kirmeyer, G.J., Wilczak, A., Jasangelo, J.G., Marcinko, J.P.,
Wolfe, R.L., 1996. Controlling nitrification in chloraminated systems.
Journal AWWA 88, 86–98.
Paje, F., Mossakowski, D., 1984. pH-preferences and habitat selection in
carabid beetles. Oecologia (Berlin) 64, 41–46.
Reid, I.D., 1985. Biological delignification of aspen wood by solid-state
fermentation with the white-rot fungus Merulius tremellosus. Applied
and Environmental Microbiology 50, 133–139.
ARTICLE IN PRESSM. Milius et al. / Journal of Insect Physiology 52 (2006) 960–967966

76
Schoonhoven, L.M., van Loon, J.J.A., 2002. An inventory of taste in
caterpillars: each species its own key. Acta Zoologica Academiae
Scientiarum Hungarica 48, 215–263.
Shimazono, H., Hayaishi, O., 1957. Enzymatic decarboxylation of oxalic
acid. Journal of Biological Chemistry 227, 151–159.
Silferberg, H., 2004. Enumeratio nova Coleopterorum Fennoscandiae,
Daniae et Baltiae. Sahlbergia 9, 1–111.
Sk"odowski, J.J.W., 2005. European Carabidology 2003. Proceedings of the
11th European Carabidologist Meeting. DIAS Report 114, 291–303.
Thiele, H.-U., 1977. Carabid Beetles in Their Environment. Zoophysiol-
ogy and Ecology, vol. 10. Springer, Berlin, p. 369.
Varela, E., Tien, M., 2003. Effect of pH and oxalate on hy-
droquinone-derived hydroxyl radical formation during brown rot
wood degradation. Applied and Environmental Microbiology 69,
6025–6031.
Wachmann, E., Platen, R., Barndt, D., 1995. Laufkafer. Beobachtung,
Lebensweise. Naturbuch Verlag, Leipzig, p. 295.
ARTICLE IN PRESSM. Milius et al. / Journal of Insect Physiology 52 (2006) 960–967 967
III

Merivee, E., Must, A., Milius, M., Luik, A., 2007.
ELECTROPHYSIOLOGICAL IDENTIFICATION OF THE
SUGAR CELL IN ANTENNAL TASTE SENSILLA OF THE
PREDATORY GROUND BEETLE PTEROSTICHUS AETHIOPS.
Journal of Insect Physiology 53(4), 377–384.

79
Journal of Insect Physiology 53 (2007) 377–384
Electrophysiological identification of the sugar cell in antennal tastesensilla of the predatory ground beetle Pterostichus aethiops
Enno Merivee�, Anne Must, Marit Milius, Anne Luik
Estonian University of Life Sciences, Institute of Agricultural and Environmental Sciences, 64 Kreutzwaldi Street, 51014 Tartu, Estonia
Received 6 October 2006; received in revised form 14 December 2006; accepted 21 December 2006
Abstract
By single sensillum tip recording technique, in addition to the salt and pH cells found in antennal taste sensilla of some ground beetles
earlier, the third chemosensory cell of four innervating these large sensilla was electrophysiologically identified as a sugar cell in the
ground beetle Pterostichus aethiops. This cell generated action potentials of considerably smaller amplitude than those of the salt and pH
cells, and phasic-tonically responded to sucrose and glucose over the range of 1–1000mM tested. Responses were concentration
dependent, with sucrose generating more spikes than glucose. During the first second of the response, maximum rates of firing of the
sugar cell reached up to 19 and 37 imp/s when stimulated with 1000mM glucose and sucrose, respectively. Three to four seconds later,
the responses decreased close to zero. Both sugars are important in plant carbohydrate metabolism. These ground dwelling insects may
come into contact with live and decayed plant material everywhere in their habitat including their preferred overwintering sites in brown-
rot decayed wood. In conclusion, we hypothesize that high content of soluble sugars in their overwintering sites and refugia is
unfavourable for these ground beetles, most probably to avoid contact with dangerous fungi.
r 2007 Elsevier Ltd. All rights reserved.
Keywords: Ground beetles; Antennal taste sensilla; Sugar cell; Tip recording technique; Habitat selection; Brown-rotted wood; Hibernation
1. Introduction
Most electrophysiologists studying insect contact che-moreception have focused on taste sensilla of herbivores(Chapman, 1998,2003; Schoonhoven and van Loon, 2002)and some other insect groups (van der Starre, 1972; Ruth,1976; Hanson, 1987; Hansen-Delkeskamp, 1992; Murataet al., 2002), related to their feeding and oviposition.Usually, these sensilla are located on mouthparts, anten-nae, ovipositor and tarsi. Very little is known aboutfunctioning of taste sensilla in predatory insects such asground beetles (Carabidae). In the tribes Platynini,Pterostichini and Bembidiini, approximately 70 tastebristles are located on the antennae of adults (Meriveeet al., 2000, 2001, 2002). Four chemoreceptive and onemechanoreceptive cell innervate these sensilla in the groundbeetle Nebria brevicollis (Daly and Ryan, 1979). Similar setof sensory cells is widespread in contact chemoreceptors of
insects suggesting that it is characteristic for other groundbeetles too. In the ground beetles Pterostichus aethiops andPterostichus oblongopunctatus, two chemoreceptive cellshave been electrophysiologically identified: they are the salt(cation) cell and pH cell probably involved in choice ofhabitat and overwintering sites of these insects (Meriveeet al., 2004, 2005; Milius et al., 2006). The occurrence of thepH receptive cell in the sensillar apparatus seems to beunique for ground beetles, as it is not found in otherinsects. Stimulating chemical compounds, specific for othertwo chemoreceptive cells innervating antennal taste sensillaof ground beetles, are not known.Besides other chemosensory cells, phagostimulatory
sugar cells are present in all phytophagous insects that havebeen studied and are almost certainly present universally(Schoonhoven and van Loon, 2002; Chapman, 2003). Sugarcells have been also found in various flies (Shiraishi andMorita, 1969; van der Starre, 1972; Hanson, 1987; Liscia etal., 1998, 2002; Furuyama et al., 1999; Amakawa, 2001;Sadakata et al., 2002; Murata et al., 2004), cockroaches(Ruth, 1976; Becker and Peters, 1989; Hansen-Delkeskamp
ARTICLE IN PRESS
www.elsevier.com/locate/jinsphys
0022-1910/$ - see front matter r 2007 Elsevier Ltd. All rights reserved.
doi:10.1016/j.jinsphys.2006.12.012
�Corresponding author.
E-mail address: [email protected] (E. Merivee).

and Hansen, 1995; Hansen-Delkeskamp, 1998) andapterygotan insects (Zygentoma) (Hansen-Delkeskamp,2001). These results suggest that also in ground beetles,taste sensilla may house the sugar cell. The aim of this studyis to test stimulatory effect of glucose and sucrose, importantin plant carbohydrate metabolism (Kruger, 1993), toantennal taste bristles in the ground beetle P. aethiops
(Panzer, 1796). This ground dwelling forest insect(Thiele, 1977) may come into contact with live and decayedplant material everywhere in its habitat including itspreferred overwintering sites in brown-rot decayed wood.Results of these experiments are reported in this paper.
2. Material and methods
2.1. Test beetles
Test beetles were collected in their preferred overwinteringsites in brown-rot decayed wood at forest margins in southernEstonia in September and October 2005. Beetles were kept inplastic boxes filled with moistened moss and pieces of brown-rot decayed wood in refrigerator at 5–6 1C for a couple ofweeks. Three to four days before experiments, the beetleswere exposed to room temperature (20 1C), fed withmoistened cat food (Kitecat, Master Foods, Poland) andgiven clean water to drink every day.
Intact test beetles were restrained by placing them tightlyinto a special conical tube made of thin sheet aluminium ofa size that allowed their head and antennae to protrudeonly a little from the narrower end. The wider rear end ofthe conical tube was blocked with a piece of plasticine toprevent the beetle from retreating out of the tube.Thereafter, antennae of the beetles were fixed horizontallyon the edge of an aluminium stand with special clamps andtiny amounts of beeswax, so that horizontally located largechaetoid taste sensilla were well visible from above under alight microscope, and easily accessible for micromanipula-tions from the side. Contamination of tested taste bristlesand their contact with preparation instruments andbeeswax was carefully avoided.
2.2. Stimulation and recording of action potentials
Chemicals of analytical grade of purity used in theexperiments were obtained from AppliChem (Germany).Ten millimolar KCl was used as an electrolyte in the mixtureswith sugars, and as a control solution. pH of stimulatingsolutions varying between 6.1 and 6.2 was measured with thepH meter E6115 (Evikon, Estonia). To minimize the possiblemicrobial contamination, stimulating sugar solutions wereprepared no longer than 24h before experiments.
Action potentials from taste bristles were recorded byconventional single sensillum tip-recording technique. Toreach good signal-to-noise ratio, a grounded tungstenmicroelectrode was inserted into the base of the flagellum.The recording glass micropipette with tip diameter of 20mmand filled with stimulating solution was placed over the bristle
tip by means of a micromanipulator under visual controlthrough a light microscope at a magnification of 300� .Immediately before each stimulation, 2–3 drops of solutionwere squeezed out of the micropipette tip, and adsorbed ontoa piece of filter paper attached to an additional micromani-pulator, in order to avoid concentration changes in thecapillary tip due to evaporation. Due to response, variabilityof different taste bristles to the same stimulus stimulatingsolutions were tested in pairs to allow a more precisecomparison of the responses to the tested stimuli. Thus, eachtaste sensillum was tested twice. Primarily, three to five sensillaof each test beetle were tested with the control stimulus. Then,approximately after 30min, the stimulating/recording micro-pipette was rinsed with distilled water several times and filledwith the second stimulating solution containing sugar to testthe same sensilla again. In each next test beetle the order ofpresentation of stimulating solutions was alternated and thenfiltered with a band width set at 100–2000Hz and amplified(input impedance 10GO) signals were monitored on anoscilloscope screen, and relayed to a computer via ananalogue-to-digital input board DAS-1401 (Keithley, Taun-ton, MA, USA) for data acquisition, storage, and analysisusing TestPoint software (Capital Equipment Corporation,Billerica, MA, USA) at a sampling rate of 10kHz. Recordingswere made during 5-sec stimulation period in order tocharacterize both rapid phasic and relatively slow stabilizationperiod of the responses of the chemosensory cells involved.
2.3. Data analysis
Automatic classifying and counting of action potentialsfrom taste sensilla was clearly not an appropriate methodfor data analysis because two action potentials frequentlyinteracted to produce an irregular waveform. Instead, spikesfrom several chemosensory cells innervating antennal tastesensilla tested were visually distinguished by their amplitudeand polarity of spikes, and counted, using TestPointsoftware. In some cases, the regularity of firing as theprincipal parameter for separating and identifying actionpotentials was used, whereas contact artefact lasting up to25ms seen at onset of stimulus was not corrected by ourrecording amplifier, 3–4 spikes may have lost. So as thecontact artefact of the same duration occurs in both cases,when stimulated with control as well as with test solutioncontaining sugar, it does not essentially affect main resultsand conclusions of the work. Firing rate (imp/s) was used tomeasure the response. t-test for paired samples was used todetermine different means significantly. Responses fromthree to five taste sensilla of each test beetle were analysed.The number of beetles (N) in each test series was 7–16.
3. Results
3.1. Responses of antennal taste sensilla to 10mM KCl
Stimulation of antennal taste bristles of the groundbeetle P. aethiops by 10mM KCl only, serving as an
ARTICLE IN PRESSE. Merivee et al. / Journal of Insect Physiology 53 (2007) 377–384378

81
electrolyte and control solution in the experiments, causedtwo chemosensory cells to respond phasic-tonically. Thesecells were the salt (cation) cell and the pH cell. Comparedto the action potentials with negative polarity generated bythe salt cell those of the pH cell were positive andconsiderably larger in amplitude (Fig. 1A). In varioussamples of test insects, the first second response of the pHcell to this electrolyte varied to a great degree, from 4to15 imp/s. During the next 4 s of stimulation, however, therate of firing of the pH cell fell to the level of 0–2 imp/s(Figs. 2C and 3C). In contrast to the pH cell, firing rate ofthe salt cell was highly stable in different samples of testbeetles with first second responses varying between 11 and13 imp/s (Figs. 2B and 3B). In some recordings, a few smallaction potentials with negative polarity of the third cellwere observed. Their mean rate of firing usually did notexceed 1 imp/s at the beginning of stimulation. Thereafter,this low rate of firing fell to the level close to zero.
3.2. Responses of antennal taste sensilla to the mixtures of
10mM KCl and glucose
In response to the mixtures of 10mM KCl and sugars, inaddition to the action potentials from the salt and pH cells,
numerous spikes of the third cell were observed in therecordings (Fig. 1B). The responses of the third cell tostimulation solutions containing glucose were significantlystronger compared to control solution (po0.05), except forthe solutions with 1 and 1000mM glucose at the end of the5-s stimulation period, when the responses fell much below1 imp/s and they did not differ from control solution(p40.05). During the first second of stimulation, over therange of 1–1000mM glucose tested, firing rate of the thirdcell drastically increased with concentration increase from2 to 19 imp/s (po0.05). During the next 4 s of stimulation,firing rate of the third cell fell below 1–2 imp/s, and nodependence of the response on glucose concentration wasobserved any longer. This cell was classified as a phasic-tonic sugar cell (Fig. 2A).In contrast, the responses of the salt cell were not
significantly affected by the glucose content of thestimulating solutions (p40.05). Its first second responsesvaried between 11 and 14 imp/s. The responses fell below1 imp/s during the next s of stimulation, and close to zerothereafter (Fig. 2B). The responses of the pH cell were alsoaffected by the content of glucose in the stimulatingsolutions, but in a more complicated manner. Firing rate ofthis cell increased to some degree at low concentrations of
ARTICLE IN PRESS
Fig. 1. Typical responses from antennal taste bristles of the ground beetle Pterostichus aethiops: (A) 10mM KCl (control solution), (B) the mixture of 10
mMKCl and 100mM sucrose. Arrows indicate the beginning of stimulation. Action potentials of the sugar cell (SC), salt cell (CC) and pH cell (pHC) were
distinguished.
E. Merivee et al. / Journal of Insect Physiology 53 (2007) 377–384 379
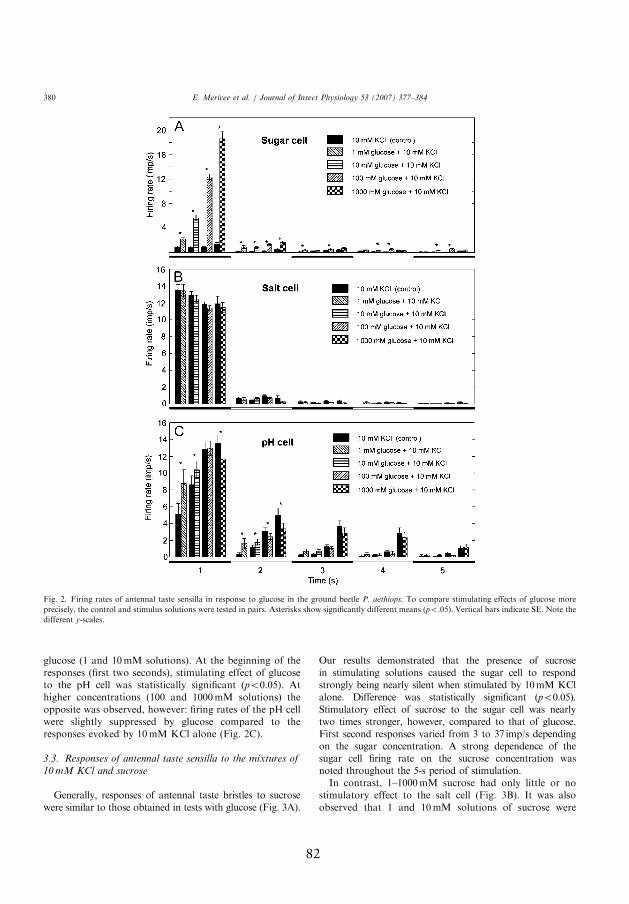
82
glucose (1 and 10mM solutions). At the beginning of theresponses (first two seconds), stimulating effect of glucoseto the pH cell was statistically significant (po0.05). Athigher concentrations (100 and 1000mM solutions) theopposite was observed, however: firing rates of the pH cellwere slightly suppressed by glucose compared to theresponses evoked by 10mM KCl alone (Fig. 2C).
3.3. Responses of antennal taste sensilla to the mixtures of
10mM KCl and sucrose
Generally, responses of antennal taste bristles to sucrosewere similar to those obtained in tests with glucose (Fig. 3A).
Our results demonstrated that the presence of sucrosein stimulating solutions caused the sugar cell to respondstrongly being nearly silent when stimulated by 10mM KClalone. Difference was statistically significant (po0.05).Stimulatory effect of sucrose to the sugar cell was nearlytwo times stronger, however, compared to that of glucose.First second responses varied from 3 to 37 imp/s dependingon the sugar concentration. A strong dependence of thesugar cell firing rate on the sucrose concentration wasnoted throughout the 5-s period of stimulation.In contrast, 1–1000mM sucrose had only little or no
stimulatory effect to the salt cell (Fig. 3B). It was alsoobserved that 1 and 10mM solutions of sucrose were
ARTICLE IN PRESS
Fig. 2. Firing rates of antennal taste sensilla in response to glucose in the ground beetle P. aethiops. To compare stimulating effects of glucose more
precisely, the control and stimulus solutions were tested in pairs. Asterisks show significantly different means (po.05). Vertical bars indicate SE. Note the
different y-scales.
E. Merivee et al. / Journal of Insect Physiology 53 (2007) 377–384380
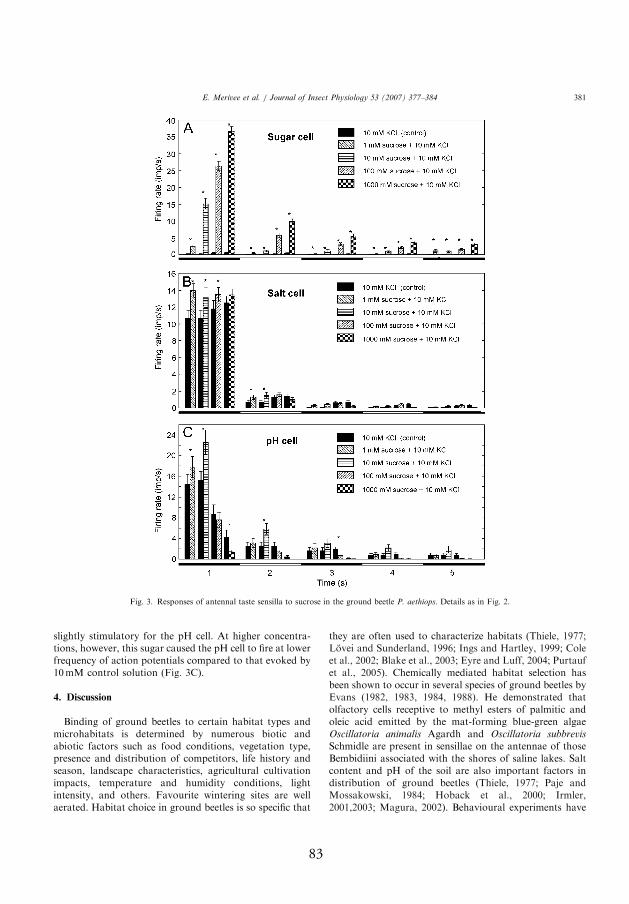
83
slightly stimulatory for the pH cell. At higher concentra-tions, however, this sugar caused the pH cell to fire at lowerfrequency of action potentials compared to that evoked by10mM control solution (Fig. 3C).
4. Discussion
Binding of ground beetles to certain habitat types andmicrohabitats is determined by numerous biotic andabiotic factors such as food conditions, vegetation type,presence and distribution of competitors, life history andseason, landscape characteristics, agricultural cultivationimpacts, temperature and humidity conditions, lightintensity, and others. Favourite wintering sites are wellaerated. Habitat choice in ground beetles is so specific that
they are often used to characterize habitats (Thiele, 1977;Lovei and Sunderland, 1996; Ings and Hartley, 1999; Coleet al., 2002; Blake et al., 2003; Eyre and Luff, 2004; Purtaufet al., 2005). Chemically mediated habitat selection hasbeen shown to occur in several species of ground beetles byEvans (1982, 1983, 1984, 1988). He demonstrated thatolfactory cells receptive to methyl esters of palmitic andoleic acid emitted by the mat-forming blue-green algaeOscillatoria animalis Agardh and Oscillatoria subbrevis
Schmidle are present in sensillae on the antennae of thoseBembidiini associated with the shores of saline lakes. Saltcontent and pH of the soil are also important factors indistribution of ground beetles (Thiele, 1977; Paje andMossakowski, 1984; Hoback et al., 2000; Irmler,2001,2003; Magura, 2002). Behavioural experiments have
ARTICLE IN PRESS
Fig. 3. Responses of antennal taste sensilla to sucrose in the ground beetle P. aethiops. Details as in Fig. 2.
E. Merivee et al. / Journal of Insect Physiology 53 (2007) 377–384 381

84
shown that presumptive contact chemoreceptive sensillaeare located on the ground beetles’ antennae (Paje andMossakowski, 1984; Hoback et al., 2000).
By electrophysiological experiments it is convincinglydemonstrated that the salt and pH cells innervatenumerous antennal taste bristles in ground beetles indeed.The used conventional single sensillum tip-recordingtechnique (Hodgson et al., 1955), and the location of thegrounded microelectrode at the base of antennal flagellumfree of muscle tissues guaranteed that action potentialsrecorded originated from the sensory cells of the tastesensillum contacted and stimulated by the recordingmicroelectrode. The unusual positive polarity and extre-mely long duration of the potentials generated by the pHcell have been repeatedly pointed out and discussed earlierin various ground beetles. Unfortunately, we have nosatisfactory explanation for this phenomenon today(Merivee et al., 2004, 2005). Detailed transmission electronmicroscope studies are needed to get new detailedinformation on the inner structure of these sensilla.Intracellular recordings of action potentials from the pHcell may probably finally solve the problem of the realpolarity of their unusual positive potentials recorded byextracellular single sensillum tip-recording technique. Dueto the technical complicacy of intracellular recording ofaction potentials from insect antennal taste sensilla coveredwith a thick and hard cuticular wall, no such attempts havebeen made so far, however. With regard to the pHreceptive cell, it was suggested that in P. aethiops andP. oblongopunctatus, in their preferred acid forest habitatsand overwintering sites in brown-rotted wood at pH 3–5the antennal pH sensitive cell does not discharge ordischarges at very low frequency with the first secondfiring rate close to 1 imp/s or lower. Areas with higher pHseems to be unfavourable for these insects and whencontacted the pH cell signals with a stronger response(Merivee et al., 2004, 2005; Milius et al., 2006). Therefore,we hypothesize that the function of the third chemorecep-tive cell of the taste bristles identified as a sugar cell in theforest ground beetle P. aethiops, is also related to habitatand/or microhabitat choice. This cell generated actionpotentials of considerably smaller amplitude than those ofthe salt and pH cells, and responded phasic-tonically tosucrose and glucose over the all range of 1–1000mMstimulating solutions tested, a span that covers the range ofsoluble sugar levels generally present in green plants, i.e.1–50mM/l (Schoonhoven and van Loon, 2002; Chapman,2003). Significantly, higher rates of firing were recorded inresponse to sucrose compared to that evoked by glucose.During the first second of the response, maximum rates offiring of the sugar cell reached up to 19 and 37 imp/s whenstimulated with glucose and sucrose, respectively. Bothsugars are important in plant carbohydrate metabolism.These results are in a good consistency with literature dataobtained by similar experiments with phytophagousinsects. Various mono-, di-, and trisaccharides maystimulate the sugar cell with different effectiveness in some
insects, whereas in other species this receptor respondsexclusively to sucrose or glucose or fructose or othercarbohydrate. These compounds function as strong pha-gostimulants and important stimuli for host recognition tomost herbivorous insects (Mitchell and Gregory, 1979;Schoonhoven, 1987; Bernays and Chapman, 2001; Bernayset al., 2002; Schoonhoven and van Loon, 2002; Chapman,2003) suggesting that the sugar cell found in antennal tastebristles of the predatory ground beetle P. aethiops may alsobe related to detection for food.On the other hand, gustatory apparatuses are usually
located in the labella, legs, wings, palpi and less frequently inthe antennae. Further, there exist strong evidences that thepH cell at least, and probably also the salt cell innervatingthese antennal taste bristles are responsible for habitatselection in some ground beetles (Thiele, 1977; Paje andMossakowski, 1984; Hoback et al., 2000). So, participationof the sugar cell in searching for suitable habitat is alsopossible in ground beetles. The predatory ground beetleP. aethiops may come into contact with live and decayedplant material everywhere in its habitat including itspreferred overwintering sites in brown-rot decayed woodwhich does not contain soluble sugars and cellulose, andwhich is composed of pure lignin. Stable moisture contentand aeration are quaranteed as a result of metabolic waterproduction during the wood-degrading process. The contentof sugars and pH depend on the assembly of wood-decayingfungi involved, and change during this process (Reid, 1985;Enoki et al., 1988; Zabel and Morrell, 1992; Varela and Tien,2003) reflecting the quality of decaying wood as a favourablemicroenvironment to ground beetles for their prolongedhibernation. In addition, brown-rotted wood may offer goodprotection for hibernating ground beetles against entomo-pathogenic fungi and parasites which play an important rolein their population dynamic. Up to 41% parasitism bynematodes and ectoparasitic fungi was found on 14 speciesof Bembidion in Norway depending on habitat selectionof the host (Andersen and Skorping, 1991). Eggs ofP. oblongopunctatus suffered 83% mortality in fresh litterbut only 7% in sterilized soil (Heessen, 1981). Althoughpredators, parasites, and pathogens affect all developmentalstages of ground beetles, quantitative data remain scarce.Hence, antennal pH, salt and sugar receptors of the groundbeetle P. aethiops seem to be a powerful tool in their selectionof suitable habitats and microhabitats crucial to survive.
Acknowledgements
Contract grant sponsor: Estonian Science Foundation;Contract grant numbers 5423 and 5736.
References
Amakawa, T., 2001. Effects of age and blood sugar levels on the proboscis
extension of the blow fly Phormia regina. Journal of Insect Physiology
47, 195–203.
ARTICLE IN PRESSE. Merivee et al. / Journal of Insect Physiology 53 (2007) 377–384382

85
Andersen, J., Skorping, A., 1991. Parasites of carabid beetles: prevalence
depends on habitat selection of the host. Canadian Journal of Zoology
69, 1216–1220.
Becker, A., Peters, W., 1989. Localization of sugar-binding sites in contact
chemosensilla of Periplaneta americana. Journal of Insect Physiology
35, 239–250.
Bernays, E.A., Chapman, R.F., 2001. Taste cell responses in the
polyphagous arctiid, Grammia geneura: towards a general pattern
for caterpillars. Journal of Insect Physiology 47, 1029–1043.
Bernays, E.A., Chapman, R.F., Hartmann, T., 2002. A highly sensitive
taste receptor for pyrrolizidine alkaloids in the lateral galeal sensillum
of a polyphagous caterpillar, Estigmene acraea. Journal of Compara-
tive Physiology A 188, 715–723.
Blake, S., McCracken, D.I., Eyre, M.D., Garside, A., Foster, G.N., 2003.
The relationship between the classification of Scottish ground
beetle assemblages (Coleoptera, Carabidae) and the national vegeta-
tion classification of British plant communities. Ecography 26,
602–616.
Chapman, R.F., 1998. The Insects. Structure and Function. Cambridge
University Press, Cambridge, p. 770.
Chapman, R.F., 2003. Contact chemoreception in feeding by phytopha-
gous insects. Annual Review of Entomology 48, 455–484.
Cole, L.J., McCracken, D.I., Dennis, P., Downie, I.S., Griffin, A.L.,
Foster, G.N., Murphy, K.J., Waterhouse, T., 2002. Relationships
between agricultural management and ecological groups of ground
beetles (Coleoptera: Carabidae) on Scotish farmland. Agriculture,
Ecosystems and Environment 93, 323–336.
Daly, P.J., Ryan, M.F., 1979. Ultrastructure of antennal sensilla of Nebria
brevicollis (Fab.) (Coleoptera: Carabidae). International Journal of
Insect Morphology and Embryology 8, 169–181.
Enoki, A., Tanaka, H., Fuse, G., 1988. Degradation of lignin-related
compounds, pure cellulose, and wood components by white-rot and
brown-rot fungi. Holzforschung 42, 85–93.
Evans, W.G., 1982. Oscillatoria sp. (Cyanophyta) mat metabolites
implicated in habitat selection in Bembidion obtusidens (Coleopt.:
Carabidae). Journal of Chemical Ecology 8, 671–678.
Evans, W.G., 1983. Habitat selection in the Carabidae. The Coleopterists’
Bulletin 37, 164–167.
Evans, W.G., 1984. Odour-mediated responses of Bembidion obtusidens
(Coleoptera: Carabidae) in a wind tunnel. Canadian Journal of
Entomology 116, 1653–1658.
Evans, W.G., 1988. Chemically mediated habitat recognition in shore
insects (Coleoptera: Carabidae, Hemiptera: Saldidae). Journal of
Chemical Ecology 14, 1441–1454.
Eyre, M.D., Luff, M.L., 2004. Ground beetle species (Coleoptera,
Carabidae) associations with land cover variables in northern England
and southern Scotland. Ecography 27, 417–426.
Furuyama, A., Koganezawa, M., Shimada, I., 1999. Multiple receptor
sites for nucleotide reception in the labellar taste receptor cells of the
fleshfly Boettcherisca peregrina. Journal of Insect Physiology 45,
249–255.
Hansen-Delkeskamp, E., 1992. Functional characterization of antennal
contact chemoreceptors in the cockroach Periplaneta americana.
Journal of Insect Physiology 38, 813–822.
Hansen-Delkeskamp, E., 1998. Development of specific responses in
antennal taste hairs after ecdysis. An electrophysiological investigation
of the cockroach, Periplaneta brunnea. Journal of Insect Physiology 44,
659–666.
Hansen-Delkeskamp, E., 2001. Responsiveness of antennal taste hairs of
the apterygotan insect, Thermobia domestica (Zygentoma); an electro-
physiological investigation. Journal of Insect Physiology 47, 689–697.
Hansen-Delkeskamp, E., Hansen, K., 1995. Responses and spike
generation in the largest antennal taste hairs of Periplaneta brunnea
Burm. Journal of Insect Physiology 41, 773–781.
Hanson, F.E., 1987. Chemoreception in the fly: the search for the
liverwurst receptor. In: Chapman, R.F., Bernays, E.A., Stoffolano,
J.G. (Eds.), Perspectives in Chemoreception and Behaviour. Springer,
Germany, pp. 99–122.
Heessen, H.J.L., 1981. Egg mortality in P. oblongopunctatus (Coleoptera,
Carabidae). Oecologia 50, 233–235.
Hoback, W.W., Golick, D.A., Svatos, T.M., Spomer, S.M., Higley, L.G.,
2000. Salinity and shade preferences result in ovipositional differences
between sympatric tiger beetle species. Ecological Entomology 25,
180–187.
Hodgson, E.S., Lettvin, J.Y., Roeder, K.D., 1955. Physiology of a primary
chemoreceptor unit. Science 122, 417–418.
Ings, T.C., Hartley, S.E., 1999. The effect of habitat structure on carabid
communities during the regeneration of a native Scotish forest. Forest
Ecology and Management 119, 123–136.
Irmler, U., 2001. Charakterisierung der Laufkafergemeinschaften schles-
wig-holsteinischer Walder und Moglichkeiten ihrer okologischen
Bewertung. Angewandte Carabidologie, Supplement 2. Laufkafer im
Wald, 21–32.
Irmler, U., 2003. The spatial and temporal pattern of carabid beetles on
arable fields in northern Germany (Schleswig–Holstein) and their value
as ecological indicators. Agriculture, Ecosystems and Environment 98,
141–151.
Kruger, N.J., 1993. Carbohydrate synthesis and degradation. In: Dennis,
D.T., Turpin, D.H. (Eds.), Plant Physiology, Biochemistry and
Molecular Biology. Longman Scientific and Technical, Singapore,
pp. 59–76.
Liscia, A., Majone, R., Solari, P., Tomassini Barbarossa, I., Crnjar, R.,
1998. Sugar response differences related to sensillum type and location
on the labella of Protophormia terraenovae: a contribution to spatial
representation of the stimulus. Journal of Insect Physiology 44, 471–481.
Liscia, A., Crnjar, R., Masala, C., Sollai, G., Solari, P., 2002. Sugar
reception in the blowfly: a possible Ca++ involvement. Journal of
Insect Physiology 48, 693–699.
Lovei, G.L., Sunderland, K.D., 1996. Ecology and behavior of ground
beetles (Coleoptera: Carabidae). Annual Review of Entomology 41,
231–256.
Magura, T., 2002. Carabids and forest edge: spatial pattern and edge
effect. Forest Ecology and Management 157, 23–37.
Merivee, E., Ploomi, A., Rahi, M., Luik, A., Sammelselg, V., 2000.
Antennal sensilla of the ground beetle Bembidion lampros Hbst
(Coleoptera, Carabidae). Acta Zoologica (Stockholm) 81, 339–350.
Merivee, E., Ploomi, A., Luik, A., Rahi, M., Sammelselg, V., 2001.
Antennal sensilla of the ground beetle Platynus dorsalis (Pontoppidan,
1763) (Coleoptera, Carabidae). Microscopy Research and Techique 55,
339–349.
Merivee, E., Ploomi, A., Rahi, M., Bresciani, J., Ravn, H.P., Luik, A.,
Sammelselg, V., 2002. Antennal sensilla of the ground beetle
Bembidion properans Steph. (Coleoptera, Carabidae). Micron 33,
429–440.
Merivee, E., Renou, M., Mand, M., Luik, A., Heidemaa, M., Ploomi, A.,
2004. Electrophysiological responses to salts from antennal chaetoid
taste sensilla of the ground beetle Pterostichus aethiops. Journal of
Insect Physiology 50, 1001–1013.
Merivee, E., Ploomi, A., Milius, M., Luik, A., Heidemaa, M., 2005.
Electrophysiological identification of antennal pH-receptors in the
ground beetle Pterostichus oblongopunctatus (Coleoptera, Carabidae).
Physiological Entomology 30, 122–133.
Milius, M., Merivee, E., Williams, I., Luik, A., Mand, M., Must, A., 2006.
A new method for Electrophysiological identification of antennal pH
receptor cells in ground beetles: the example of Pterostichus aethiops
(Panzer, 1796) (Coleoptera, Carabidae). Journal of Insect Physiology
52, 960–967.
Mitchell, B.K., Gregory, P., 1979. Physiology of the maxillary sugar
sensitive cell in the red turnip beetle, Entomoscelis americana. Journal
of Comparative Physiology 132, 167–178.
Murata, Y., Kataoka–Shirasugi, N., Amakawa, T., 2002. Electrophysio-
logical studies of salty taste modification by organic acids in the
labellar taste cell of the blowfly. Chemical Senses 27, 57–65.
Murata, Y., Mashiko, M., Ozaki, M., Amakawa, T., Nakamura, T., 2004.
Intrinsic nitric oxide regulates the taste response of the sugar receptor
cell in the blowfly, Phormia regina. Chemical Senses 29, 75–81.
ARTICLE IN PRESSE. Merivee et al. / Journal of Insect Physiology 53 (2007) 377–384 383

86
Paje, F., Mossakowski, D., 1984. pH-preferences and habitat selection in
carabid beetles. Oecologia (Berlin) 64, 41–46.
Purtauf, T., Roschewitz, I., Dauber, J., Thies, C., Tscharntke, T., Wolters,
V., 2005. Landscape context of organic and conventional farms:
influences on carabid beetle diversity. Agriculture, Ecosystems and
Environment 108, 165–174.
Reid, I.D., 1985. Biological delignification of aspen wood by solid-state
fermentation with the white-rot fungus Merulius tremellosus. Applied
and Environmental Microbiology 50, 133–139.
Ruth, E., 1976. Elektrophysiologie der Sensilla Chaetica auf den Antennen
von Periplaneta americana. Journal of comparative Physiology 105,
55–64.
Sadakata, T., Hatano, H., Koseki, T., Koganezawa, M., Shimada, I.,
2002. The effect of amiloride on the labellar taste receptor cells of the
fleshfly Boettcherisca peregrina. Journal of Insect Physiology 48,
565–570.
Schoonhoven, L.M., 1987. What makes a caterpillar eat? The sensory
code underlying feeding behavior. In: Chapman, R.F.et al. (Ed.),
Perspectives in Chemoreception and Behavior. Springer, New York,
pp. 69–97.
Schoonhoven, L.M., van Loon, J.J.A., 2002. An inventory of taste in
caterpillars: each species its own key. Acta Zoologica Academiae
Scientiarum Hungaricae 48, 215–263.
Shiraishi, A., Morita, H., 1969. The effects of pH on the labellar sugar
receptor of the fleshfly. The Journal of General Physiology 53, 450–470.
Thiele, H.-U., 1977. Carabid Beetles in Their Environment. Zoophysiology
and Ecology, vol.10. Springer, Berlin, p. 369.
Van der Starre, H., 1972. Tarsal taste discrimination in the blowfly,
Calliphora vicina Robineau-Desvoidy. Netherlands Journal of Zool-
ogy 22, 227–282.
Varela, E., Tien, M., 2003. Effect of pH and oxalate on hydroquinone-
derived hydroxyl radical formation during brown rot wood degrada-
tion. Applied and Environmental Microbiology 69, 6025–6031.
Zabel, R.A., Morrell, J.J., 1992. Chemical changes in wood caused by
decay fungi. In: Wood Microbiology Decay and its Prevention.
Academic, San Diego, pp. 195–224.
ARTICLE IN PRESSE. Merivee et al. / Journal of Insect Physiology 53 (2007) 377–384384
IV

Merivee, E., Märtmann, H., Must, A., Milius, M., Williams, I.,
Mänd, M., 2008.
ELECTROPHYSIOLOGICAL RESPONSES FROM NEURONS
OF ANTENNAL TASTE SENSILLA IN THE POLYPHAGOUS
PREDATORY GROUND BEETLE PTEROSTICHUS
OBLONGOPUNCTATUS (fABRICIUS 1787) TO PLANT SUGARS
AND AMINO ACIDS.
Journal of Insect Physiology 54, 1213–1219.

89
Electrophysiological responses from neurons of antennal taste sensilla inthe polyphagous predatory ground beetle Pterostichus oblongopunctatus(Fabricius 1787) to plant sugars and amino acids
Enno Merivee *, Helina Martmann, Anne Must, Marit Milius, Ingrid Williams, Marika Mand
Estonian University of Life Sciences, 1 Kreutzwaldi Street, 51014 Tartu, Estonia
1. Introduction
Many species of ground beetles (Coleoptera, Carabidae) areimportant as natural enemies of phytophagous pest insects(Kromp, 1999; Symondson et al., 2002), but the chemical cuesthat they use in habitat selection and food recognition have beenlittle studied. More knowledge of these cues and their sensorydetection may aid the development of strategies to manipulatecarabid assemblages for plant protection in the future. Thecommon Euro-Siberian ground beetle, Pterostichus oblongopuncta-tus, the subject of this study, is a eurytopic woodland species,occuring in both deciduous and coniferous forests. It reproduces
in the spring; newly emerged adults appear in the autumn andhibernate in decayingwood, under the bark of tree trunks, in forestlitter and under stones (Haberman, 1968; Lindroth, 1986).
Scanning electron microscope studies show that the antennaeof ground beetles are well equipped with various chemoreceptors.In addition to numerous small basiconic olfactory sensilla, largerbristle-like contact chemosensilla have been found on the antennalflagellum of adults in several genera: approximately 70 suchsensilla in Bembidion, Platynus and Pterostichus (Merivee et al.,2000, 2001, 2002, 2005) and 56 in Carabus (Kim and Yamasaki,1996). In Nebria brevicollis these large sensilla are each innervatedby one mechanoreceptive and four chemoreceptive neurons (Dalyand Ryan, 1979). In Pterostichus aethiops and P. oblongopunctatus,specific chemical stimuli have been identified by electrophysiologyfor three of the four chemosensory neurons. One responds tovarious salts and another to the pH of the stimulating solution(Merivee et al., 2004, 2005; Milius et al., 2006). In P. aethiops, thethird neuron is sensitive to sucrose and glucose, important in plant
Journal of Insect Physiology 54 (2008) 1213–1219
A R T I C L E I N F O
Article history:
Received 10 March 2008
Received in revised form 13 May 2008
Accepted 19 May 2008
Keywords:
Contact chemoreceptors
Electrophysiology
Firing rate
Feeding
Phytophagy
Polyphagy
A B S T R A C T
The responses of antennal contact chemoreceptors, in the polyphagous predatory ground beetle
Pterostichus oblongopunctatus, to twelve 1–1000 mmol l�1 plant sugars and seven 10–100 mmol l�1
amino acids were tested. The disaccharides with an a-1.4-glycoside linkage, sucrose and maltose, were
the twomost stimulatory sugars for the sugar-sensitive neuron innervating these contact chemosensilla.
The firing rates they evoked were concentration dependent and reached up to 70 impulses/s at
1000 mmol l�1. The stimulatory effect of glucose on this neuronwas approximately two times lower. This
can be partly explained by the fact that glucose exists in at least two anomeric forms, a and b. These two
forms interconvert over a timescale of hours in aqueous solution, to a final stable ratio of a:b 36:64, in a
process called mutarotation. So the physiologically active a-anomere forms only 36% of the glucose
solution which was reflected in its relatively low dose/response curve. Due to the partial herbivory of P.
oblongopunctatus these plant sugars are probably involved in its search for food, for example, for conifer
seeds. Several carbohydrates, in addition to glucose, such as cellobiose, arabinose, xylose, mannose,
rhamnose and galactose are known as components of cellulose and hemicelluloses. They are released by
brown-rot fungi during enzymatic wood decay. None of them stimulated the antennal sugar-sensitive
neuron. They are therefore not implicated in the search for hibernation sites, which include rottingwood,
by this beetle. The weak stimulating effect (below 3 impulses/s) of some 100 mmol l�1 amino acids
(methionine, serine, alanine, glutamine) to the 4th chemosensory neuron of these sensilla was
characterized as non-specific, or modulating the responses of non-target chemosensory neurons.
� 2008 Elsevier Ltd. All rights reserved.
* Corresponding author at: Estonian University of Life Sciences, Institute of
Agricultural and Environmental Sciences, 1 Kreutzwaldi Street, 51014 Tartu,
Estonia.
E-mail address: [email protected] (E. Merivee).
Contents lists available at ScienceDirect
Journal of Insect Physiology
journa l homepage: www.e lsev ier .com/ locate / j insphys
0022-1910/$ – see front matter � 2008 Elsevier Ltd. All rights reserved.
doi:10.1016/j.jinsphys.2008.05.002

90
carbohydrate metabolism (Merivee et al., 2007). As habitatselection in ground beetles appears to be influenced by soil pHand salinity (Krogerus, 1960; Thiele, 1977; Paje and Mossakowski,1984; Hoback et al., 2000; Irmler, 2001), the antennal pH- and salt-sensitive neuron may be involved in this process. Electrophysio-logical experiments with the pH-sensitive neuron in P. oblongo-
punctatus and P. aethiops demonstrated that this neuron did not fireor fired at very low frequency (first second response below5 impulses/s) when stimulated at pH 3–6. This level of acidityreflects the pH preferences of these species in their forest habitatand hibernation sites in brown-rotted wood. Firing rates ofthe neuron increased with increase in pH, up to 20–30 impulses/sat pH 8–11 (Merivee et al., 2005; Milius et al., 2006). Theseelectrophysiological results are in agreement with the acid soilpH preferences of P. oblongopunctatus obtained in behaviouraland ecological experiments (Paje and Mossakowski, 1984; Irmler,2001).
In addition to the salt- and pH-sensitive neuron, the sugar-sensitive neuron present in the same antennal taste sensillummayalso be involved in habitat/microhabitat selection by these species.Various sugars and amino acids function as strong phagostimu-lants in most phytophagous insects equipped with specializedreceptor neurons for these chemicals (Schoonhoven and van Loon,2002; Chapman, 2003). Adults of the genus Pterostichus appear tobe primarily carnivorous, feeding on small prey animals, but inmany species, plant matter also forms part of their diet, and, insome the proportion of animal to plant matter eaten varies withseason (Thiele, 1977; Hengeveld, 1980; Lovei and Sunderland,1996). Johnson and Cameron (1969) collected data on the extent ofherbivory in more than 150 species of ground beetles. Somespecies in the genera Pterostichus, including P. oblongopunctatus,Amara, Agonum and Harpalus consume conifer seeds and youngseedlings immediately following germination. In Sweden, P.
oblongopunctatus and Calathus micropterus are the most importantpredators of Pinus sylvestris seed. In the field, seed predationtypically resulted in >20% seed mortality, reaching even higherlevels (up to 60%) on some occasions andmost predation (81% of alldamaged seeds) was due to ground beetles (Heikkila, 1977;Nystrand and Granstrom, 2000). Therefore, as in phytophagousinsects, polyphagous predatory ground beetles, including P.
oblongopunctatus, may also have plant sugar- and amino acid-sensitive neurons associated with their feeding habits.
The aim of this study was to determine whether sugar- andamino acid-sensitive receptor neurons are present in the antennaltaste sensilla of adult P. oblongopunctatus by testing for stimulatoryeffects of various sugars, mostly of plant origin, and amino acids onthese sensilla by electrophysiology. As the composition and contentof water-soluble sugars differ substantially in live and decayingplant matter (Eriksson et al., 1990; Zabel and Morrell, 1992;Valaskova and Baldrian, 2006) the reaction spectrum of any sugar-sensitive antennal neurons, if present, should give some informationon the functioning of antennal contact chemoreceptors and howthey might be involved in habitat selection and food recognition bythis species.
2. Materials and methods
2.1. Insects
Adult P. oblongopunctatus were collected from a local popula-tion in southern Estonia. Theywere kept in plastic boxes filledwithmoistened moss and pieces of brown-rotted wood in a refrigeratorat 5–6 8C. Three to four days prior to the experiments, the beetleswere transferred to room temperature (23 8C), fed with moistenedcat food (Friskies Vitality+, Nestle Purina, Hungary) and given clean
water to drink every day. Electrophysiological experiments withhibernating beetles were conducted in February and March 2006and with active beetles from June to August 2007.
2.2. Electrophysiology
Each beetle to be testedwas restrained securely in a conical tubemade of thin sheet-aluminium of a size that allowed the head andantennae to protrude from the narrower end. The wider rear end ofthe tube was blocked with plasticine to prevent the beetle fromretreating outof the tube. Its antennaewerefixedhorizontally to theedge of an aluminium stand with special clamps and beeswax, sothat the horizontally located large chaetoid taste sensilla werevisible fromaboveundera lightmicroscope, andeasily accessible formicromanipulation from the side. Care was taken not to contam-inate the taste sensilla by contact with instruments or beeswax.Each prepared test beetle was placed in a Faraday cage (15 cm�10 cm� 5 cm) for electrophysiology.
Action potentials from taste neurons of the sensilla wererecorded by the conventional single sensillum tip-recordingtechnique. To achieve a good signal-to-noise ratio, the indifferenttungsten microelectrode was inserted into the base of the flage-llum free of muscular tissue. It was connected to an AC amplifiervia a calibration source, and earthed. The recording glass micro-pipette filled with stimulating solution was brought into contactwith the tip of the sensillum by means of a micromanipulatorunder visual control through a lightmicroscope at amagnificationof 300�. Immediately before each stimulation, 2–3 drops ofsolution were squeezed out of the micropipette tip, and adsorbedonto a piece of filter paper attached to anothermicromanipulator,in order to avoid concentration changes in the capillary tip dueto evaporation. Action potentials picked up by the recordingelectrode were filtered with a bandwidth set at 100–2000 Hz,amplified (input impedance 10 GV), monitored on an oscillo-scope screen, and relayed to a computer via an analogue-to-digitalinput board DAS-1401 (Keithley, Taunton, MA, USA) for dataacquisition, storage, and analysis using TestPoint software(Capital Equipment Corp., Billerica, MA, USA) at a sampling rateof 10 kHz. Responses were recorded from neurons of three to fiverandomly selected taste sensilla on the flagellomeres 1–9 of eachtest beetle. Each sensillum was stimulated once only with onesolution. Action potentials generated during the first second afterthe beginning of stimulus applicationwere counted and analysed.Responses of the sensilla to mechanical stimulation were alsotested, especially in those rare cases when it was not clearwhether a certain class of recorded action potentials originatedfrom the chemosensory ormechanosensory neuron. The tip of thesensillumwas rapidly moved from its resting position toward theantennal shaft by 15–208, held in this position for 2–3 s, and thenrapidly moved back to its initial position using the recordingelectrode.
2.3. Chemical stimuli
Themain tests were carried out using twelve 1–1000 mmol l�1
sugars and seven 10–100 mmol l�1 amino acids (Table 1) dis-solved in 10 mmol l�1 choline chloride to give good conductivity.Compared to KCl and NaCl, which are frequently used aselectrolytes in solutions of stimulating non-electrolytes, cholinechloride elicits only a few action potentials of large amplitudegenerated by the salt- and pH-sensitive neuron innervating theantennal taste sensilla in ground beetles. As a result, discrimina-tion and counting of small spikes generated by the sugar-sensitiveneuron is easier, especially at the beginning of the response.Chemicals of analytical grade purity were obtained from
E. Merivee et al. / Journal of Insect Physiology 54 (2008) 1213–12191214
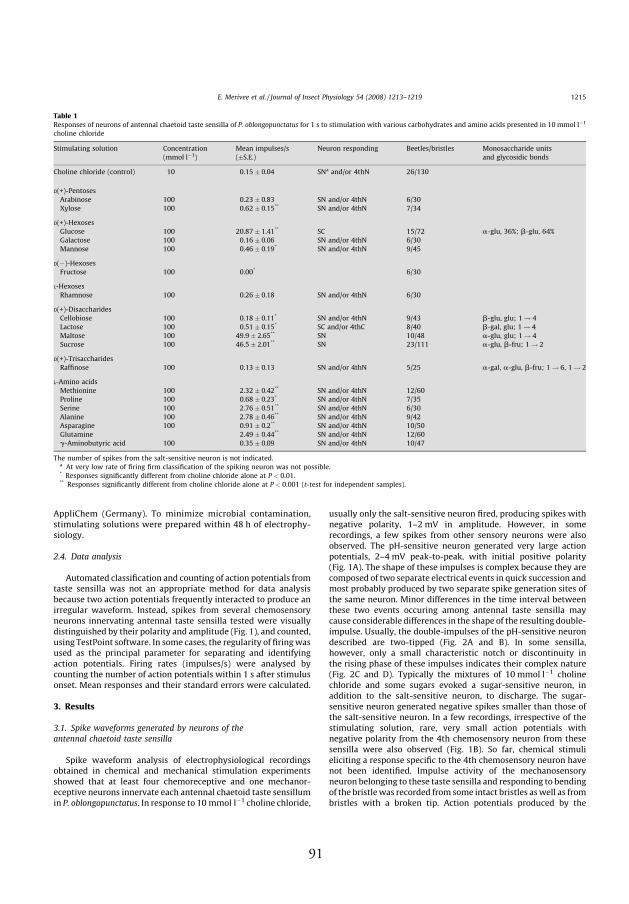
91
AppliChem (Germany). To minimize microbial contamination,stimulating solutions were prepared within 48 h of electrophy-siology.
2.4. Data analysis
Automated classification and counting of action potentials fromtaste sensilla was not an appropriate method for data analysisbecause two action potentials frequently interacted to produce anirregular waveform. Instead, spikes from several chemosensoryneurons innervating antennal taste sensilla tested were visuallydistinguished by their polarity and amplitude (Fig. 1), and counted,using TestPoint software. In some cases, the regularity of firingwasused as the principal parameter for separating and identifyingaction potentials. Firing rates (impulses/s) were analysed bycounting the number of action potentials within 1 s after stimulusonset. Mean responses and their standard errors were calculated.
3. Results
3.1. Spike waveforms generated by neurons of the
antennal chaetoid taste sensilla
Spike waveform analysis of electrophysiological recordingsobtained in chemical and mechanical stimulation experimentsshowed that at least four chemoreceptive and one mechanor-eceptive neurons innervate each antennal chaetoid taste sensillumin P. oblongopunctatus. In response to 10 mmol l�1 choline chloride,
usually only the salt-sensitive neuron fired, producing spikes withnegative polarity, 1–2 mV in amplitude. However, in somerecordings, a few spikes from other sensory neurons were alsoobserved. The pH-sensitive neuron generated very large actionpotentials, 2–4 mV peak-to-peak, with initial positive polarity(Fig. 1A). The shape of these impulses is complex because they arecomposed of two separate electrical events in quick succession andmost probably produced by two separate spike generation sites ofthe same neuron. Minor differences in the time interval betweenthese two events occuring among antennal taste sensilla maycause considerable differences in the shape of the resulting double-impulse. Usually, the double-impulses of the pH-sensitive neurondescribed are two-tipped (Fig. 2A and B). In some sensilla,however, only a small characteristic notch or discontinuity inthe rising phase of these impulses indicates their complex nature(Fig. 2C and D). Typically the mixtures of 10 mmol l�1 cholinechloride and some sugars evoked a sugar-sensitive neuron, inaddition to the salt-sensitive neuron, to discharge. The sugar-sensitive neuron generated negative spikes smaller than those ofthe salt-sensitive neuron. In a few recordings, irrespective of thestimulating solution, rare, very small action potentials withnegative polarity from the 4th chemosensory neuron from thesesensilla were also observed (Fig. 1B). So far, chemical stimulieliciting a response specific to the 4th chemosensory neuron havenot been identified. Impulse activity of the mechanosensoryneuron belonging to these taste sensilla and responding to bendingof the bristlewas recorded from some intact bristles aswell as frombristles with a broken tip. Action potentials produced by the
Table 1Responses of neurons of antennal chaetoid taste sensilla of P. oblongopunctatus for 1 s to stimulation with various carbohydrates and amino acids presented in 10 mmol l�1
choline chloride
Stimulating solution Concentration
(mmol l�1)
Mean impulses/s
(�S.E.)
Neuron responding Beetles/bristles Monosaccharide units
and glycosidic bonds
Choline chloride (control) 10 0.15 � 0.04 SNa and/or 4thN 26/130
D(+)-Pentoses
Arabinose 100 0.23 � 0.83 SN and/or 4thN 6/30
Xylose 100 0.62 � 0.15** SN and/or 4thN 7/34
D(+)-Hexoses
Glucose 100 20.87 � 1.41** SC 15/72 a-glu, 36%; b-glu, 64%Galactose 100 0.16 � 0.06 SN and/or 4thN 6/30
Mannose 100 0.46 � 0.19* SN and/or 4thN 9/45
D(�)-Hexoses
Fructose 100 0.00* 6/30
L-Hexoses
Rhamnose 100 0.26 � 0.18 SN and/or 4thN 6/30
D(+)-Disaccharides
Cellobiose 100 0.18 � 0.11* SN and/or 4thN 9/43 b-glu, glu; 1! 4
Lactose 100 0.51 � 0.15* SC and/or 4thC 8/40 b-gal, glu; 1! 4
Maltose 100 49.9 � 2.65** SN 10/48 a-glu, glu; 1! 4
Sucrose 100 46.5 � 2.01** SN 23/111 a-glu, b-fru; 1! 2
D(+)-Trisaccharides
Raffinose 100 0.13 � 0.13 SN and/or 4thN 5/25 a-gal, a-glu, b-fru; 1! 6, 1! 2
L-Amino acids
Methionine 100 2.32 � 0.42** SN and/or 4thN 12/60
Proline 100 0.68 � 0.23* SN and/or 4thN 7/35
Serine 100 2.76 � 0.51** SN and/or 4thN 6/30
Alanine 100 2.78 � 0.46** SN and/or 4thN 9/42
Asparagine 100 0.91 � 0.2** SN and/or 4thN 10/50
Glutamine 2.49 � 0.44** SN and/or 4thN 12/60
g-Aminobutyric acid 100 0.35 � 0.09 SN and/or 4thN 10/47
The number of spikes from the salt-sensitive neuron is not indicated.a At very low rate of firing firm classification of the spiking neuron was not possible.* Responses significantly different from choline chloride alone at P < 0.01.** Responses significantly different from choline chloride alone at P < 0.001 (t-test for independent samples).
E. Merivee et al. / Journal of Insect Physiology 54 (2008) 1213–1219 1215
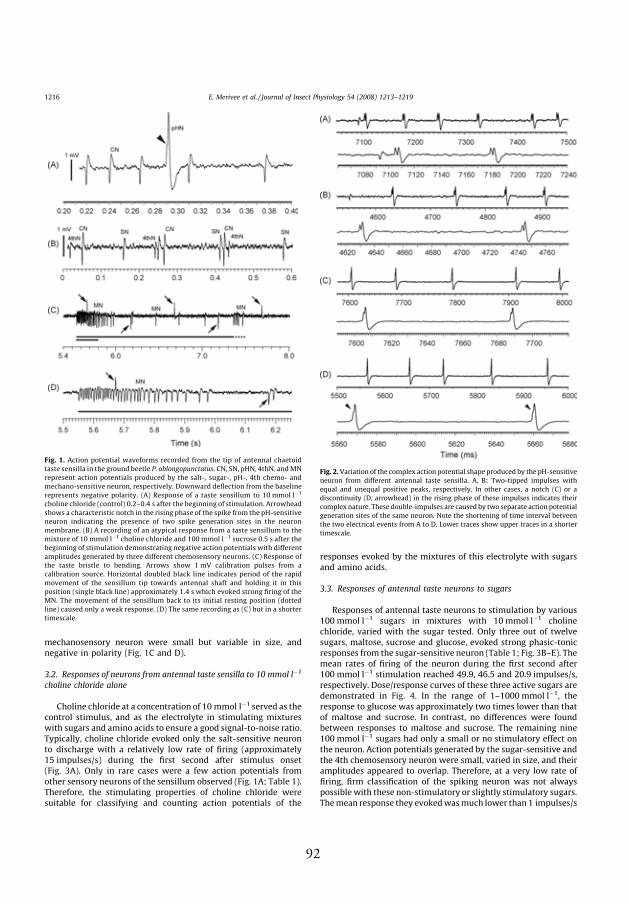
92
mechanosensory neuron were small but variable in size, andnegative in polarity (Fig. 1C and D).
3.2. Responses of neurons from antennal taste sensilla to 10 mmol l�1
choline chloride alone
Choline chloride at a concentration of 10 mmol l�1 served as thecontrol stimulus, and as the electrolyte in stimulating mixtureswith sugars and amino acids to ensure a good signal-to-noise ratio.Typically, choline chloride evoked only the salt-sensitive neuronto discharge with a relatively low rate of firing (approximately15 impulses/s) during the first second after stimulus onset(Fig. 3A). Only in rare cases were a few action potentials fromother sensory neurons of the sensillum observed (Fig. 1A; Table 1).Therefore, the stimulating properties of choline chloride weresuitable for classifying and counting action potentials of the
responses evoked by the mixtures of this electrolyte with sugarsand amino acids.
3.3. Responses of antennal taste neurons to sugars
Responses of antennal taste neurons to stimulation by various100 mmol l�1 sugars in mixtures with 10 mmol l�1 cholinechloride, varied with the sugar tested. Only three out of twelvesugars, maltose, sucrose and glucose, evoked strong phasic-tonicresponses from the sugar-sensitive neuron (Table 1; Fig. 3B–E). Themean rates of firing of the neuron during the first second after100 mmol l�1 stimulation reached 49.9, 46.5 and 20.9 impulses/s,respectively. Dose/response curves of these three active sugars aredemonstrated in Fig. 4. In the range of 1–1000 mmol l�1, theresponse to glucose was approximately two times lower than thatof maltose and sucrose. In contrast, no differences were foundbetween responses to maltose and sucrose. The remaining nine100 mmol l�1 sugars had only a small or no stimulatory effect onthe neuron. Action potentials generated by the sugar-sensitive andthe 4th chemosensory neuron were small, varied in size, and theiramplitudes appeared to overlap. Therefore, at a very low rate offiring, firm classification of the spiking neuron was not alwayspossible with these non-stimulatory or slightly stimulatory sugars.Themean response they evokedwasmuch lower than 1 impulses/s
Fig. 1. Action potential waveforms recorded from the tip of antennal chaetoid
taste sensilla in the ground beetle P. oblongopunctatus. CN, SN, pHN, 4thN, andMN
represent action potentials produced by the salt-, sugar-, pH-, 4th chemo- and
mechano-sensitive neuron, respectively. Downward deflection from the baseline
represents negative polarity. (A) Response of a taste sensillum to 10 mmol l�1
choline chloride (control) 0.2–0.4 s after the beginning of stimulation. Arrowhead
shows a characteristic notch in the rising phase of the spike from the pH-sensitive
neuron indicating the presence of two spike generation sites in the neuron
membrane. (B) A recording of an atypical response from a taste sensillum to the
mixture of 10 mmol l�1 choline chloride and 100 mmol l�1 sucrose 0.5 s after the
beginning of stimulation demonstrating negative action potentials with different
amplitudes generated by three different chemosensory neurons. (C) Response of
the taste bristle to bending. Arrows show 1 mV calibration pulses from a
calibration source. Horizontal doubled black line indicates period of the rapid
movement of the sensillum tip towards antennal shaft and holding it in this
position (single black line) approximately 1.4 s which evoked strong firing of the
MN. The movement of the sensillum back to its initial resting position (dotted
line) caused only a weak response. (D) The same recording as (C) but in a shorter
timescale.
Fig. 2.Variation of the complex action potential shape produced by the pH-sensitive
neuron from different antennal taste sensilla. A, B: Two-tipped impulses with
equal and unequal positive peaks, respectively. In other cases, a notch (C) or a
discontinuity (D, arrowhead) in the rising phase of these impulses indicates their
complex nature. These double-impulses are caused by two separate action potential
generation sites of the same neuron. Note the shortening of time interval betveen
the two electrical events from A to D. Lower traces show upper traces in a shorter
timescale.
E. Merivee et al. / Journal of Insect Physiology 54 (2008) 1213–12191216
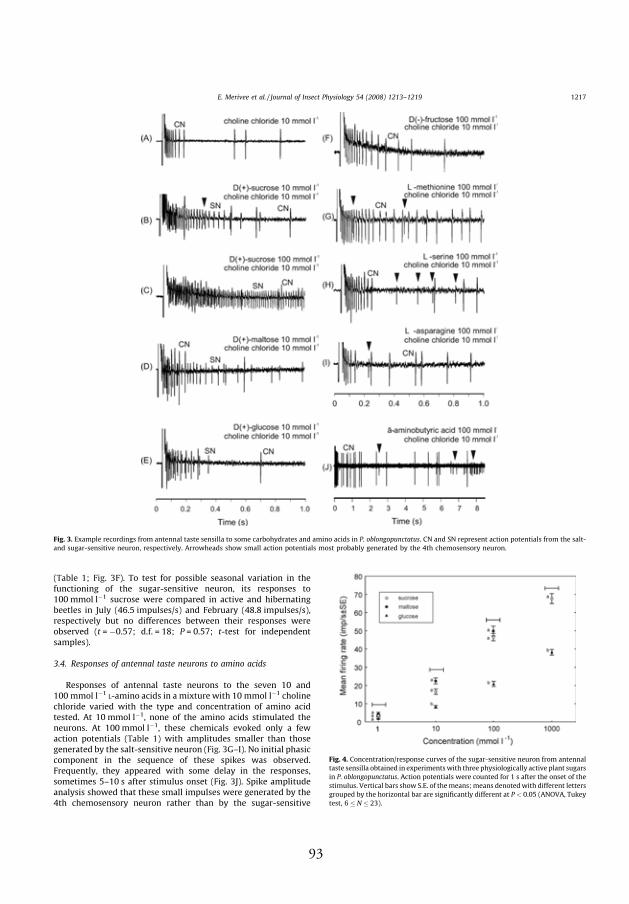
93
(Table 1; Fig. 3F). To test for possible seasonal variation in thefunctioning of the sugar-sensitive neuron, its responses to100 mmol l�1 sucrose were compared in active and hibernatingbeetles in July (46.5 impulses/s) and February (48.8 impulses/s),respectively but no differences between their responses wereobserved (t = �0.57; d.f. = 18; P = 0.57; t-test for independentsamples).
3.4. Responses of antennal taste neurons to amino acids
Responses of antennal taste neurons to the seven 10 and100 mmol l�1
L-amino acids in a mixture with 10 mmol l�1 cholinechloride varied with the type and concentration of amino acidtested. At 10 mmol l�1, none of the amino acids stimulated theneurons. At 100 mmol l�1, these chemicals evoked only a fewaction potentials (Table 1) with amplitudes smaller than thosegenerated by the salt-sensitive neuron (Fig. 3G–I). No initial phasiccomponent in the sequence of these spikes was observed.Frequently, they appeared with some delay in the responses,sometimes 5–10 s after stimulus onset (Fig. 3J). Spike amplitudeanalysis showed that these small impulses were generated by the4th chemosensory neuron rather than by the sugar-sensitive
Fig. 3. Example recordings from antennal taste sensilla to some carbohydrates and amino acids in P. oblongopunctatus. CN and SN represent action potentials from the salt-
and sugar-sensitive neuron, respectively. Arrowheads show small action potentials most probably generated by the 4th chemosensory neuron.
Fig. 4. Concentration/response curves of the sugar-sensitive neuron from antennal
taste sensilla obtained in experimentswith three physiologically active plant sugars
in P. oblongopunctatus. Action potentials were counted for 1 s after the onset of the
stimulus. Vertical bars show S.E. of themeans; means denotedwith different letters
grouped by the horizontal bar are significantly different at P < 0.05 (ANOVA, Tukey
test, 6 � N � 23).
E. Merivee et al. / Journal of Insect Physiology 54 (2008) 1213–1219 1217
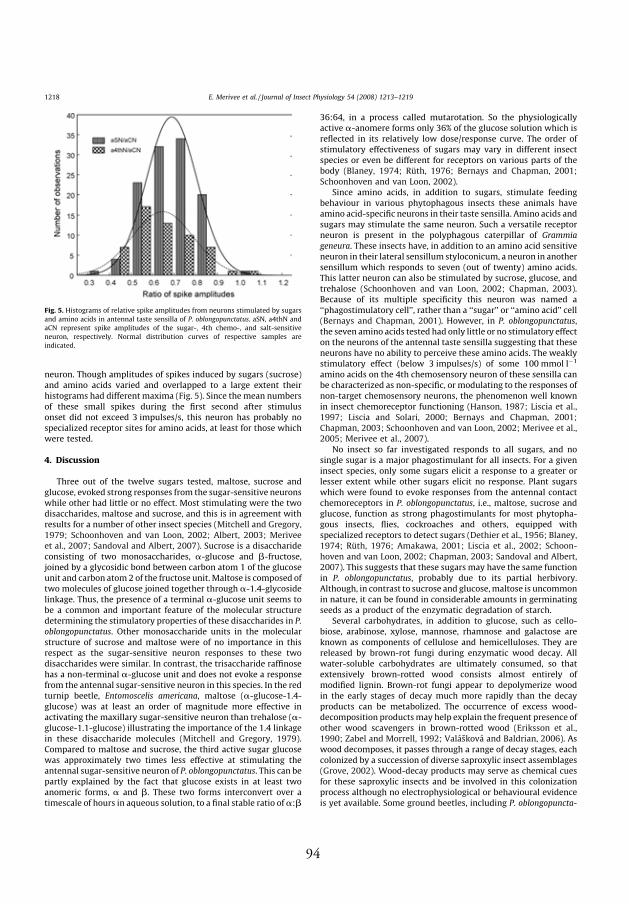
94
neuron. Though amplitudes of spikes induced by sugars (sucrose)and amino acids varied and overlapped to a large extent theirhistograms had different maxima (Fig. 5). Since the mean numbersof these small spikes during the first second after stimulusonset did not exceed 3 impulses/s, this neuron has probably nospecialized receptor sites for amino acids, at least for those whichwere tested.
4. Discussion
Three out of the twelve sugars tested, maltose, sucrose andglucose, evoked strong responses from the sugar-sensitive neuronswhile other had little or no effect. Most stimulating were the twodisaccharides, maltose and sucrose, and this is in agreement withresults for a number of other insect species (Mitchell and Gregory,1979; Schoonhoven and van Loon, 2002; Albert, 2003; Meriveeet al., 2007; Sandoval and Albert, 2007). Sucrose is a disaccharideconsisting of two monosaccharides, a-glucose and b-fructose,joined by a glycosidic bond between carbon atom 1 of the glucoseunit and carbon atom2 of the fructose unit. Maltose is composed oftwo molecules of glucose joined together through a-1.4-glycosidelinkage. Thus, the presence of a terminal a-glucose unit seems tobe a common and important feature of the molecular structuredetermining the stimulatory properties of these disaccharides in P.
oblongopunctatus. Other monosaccharide units in the molecularstructure of sucrose and maltose were of no importance in thisrespect as the sugar-sensitive neuron responses to these twodisaccharides were similar. In contrast, the trisaccharide raffinosehas a non-terminal a-glucose unit and does not evoke a responsefrom the antennal sugar-sensitive neuron in this species. In the redturnip beetle, Entomoscelis americana, maltose (a-glucose-1.4-glucose) was at least an order of magnitude more effective inactivating the maxillary sugar-sensitive neuron than trehalose (a-glucose-1.1-glucose) illustrating the importance of the 1.4 linkagein these disaccharide molecules (Mitchell and Gregory, 1979).Compared to maltose and sucrose, the third active sugar glucosewas approximately two times less effective at stimulating theantennal sugar-sensitive neuron of P. oblongopunctatus. This can bepartly explained by the fact that glucose exists in at least twoanomeric forms, a and b. These two forms interconvert over atimescale of hours in aqueous solution, to a final stable ratio ofa:b
36:64, in a process called mutarotation. So the physiologicallyactive a-anomere forms only 36% of the glucose solution which isreflected in its relatively low dose/response curve. The order ofstimulatory effectiveness of sugars may vary in different insectspecies or even be different for receptors on various parts of thebody (Blaney, 1974; Ruth, 1976; Bernays and Chapman, 2001;Schoonhoven and van Loon, 2002).
Since amino acids, in addition to sugars, stimulate feedingbehaviour in various phytophagous insects these animals haveamino acid-specific neurons in their taste sensilla. Amino acids andsugars may stimulate the same neuron. Such a versatile receptorneuron is present in the polyphagous caterpillar of Grammia
geneura. These insects have, in addition to an amino acid sensitiveneuron in their lateral sensillum styloconicum, a neuron in anothersensillum which responds to seven (out of twenty) amino acids.This latter neuron can also be stimulated by sucrose, glucose, andtrehalose (Schoonhoven and van Loon, 2002; Chapman, 2003).Because of its multiple specificity this neuron was named a‘‘phagostimulatory cell’’, rather than a ‘‘sugar’’ or ‘‘amino acid’’ cell(Bernays and Chapman, 2001). However, in P. oblongopunctatus,the seven amino acids tested had only little or no stimulatory effecton the neurons of the antennal taste sensilla suggesting that theseneurons have no ability to perceive these amino acids. The weaklystimulatory effect (below 3 impulses/s) of some 100 mmol l�1
amino acids on the 4th chemosensory neuron of these sensilla canbe characterized as non-specific, or modulating to the responses ofnon-target chemosensory neurons, the phenomenon well knownin insect chemoreceptor functioning (Hanson, 1987; Liscia et al.,1997; Liscia and Solari, 2000; Bernays and Chapman, 2001;Chapman, 2003; Schoonhoven and van Loon, 2002; Merivee et al.,2005; Merivee et al., 2007).
No insect so far investigated responds to all sugars, and nosingle sugar is a major phagostimulant for all insects. For a giveninsect species, only some sugars elicit a response to a greater orlesser extent while other sugars elicit no response. Plant sugarswhich were found to evoke responses from the antennal contactchemoreceptors in P. oblongopunctatus, i.e., maltose, sucrose andglucose, function as strong phagostimulants for most phytopha-gous insects, flies, cockroaches and others, equipped withspecialized receptors to detect sugars (Dethier et al., 1956; Blaney,1974; Ruth, 1976; Amakawa, 2001; Liscia et al., 2002; Schoon-hoven and van Loon, 2002; Chapman, 2003; Sandoval and Albert,2007). This suggests that these sugars may have the same functionin P. oblongopunctatus, probably due to its partial herbivory.Although, in contrast to sucrose and glucose,maltose is uncommonin nature, it can be found in considerable amounts in germinatingseeds as a product of the enzymatic degradation of starch.
Several carbohydrates, in addition to glucose, such as cello-biose, arabinose, xylose, mannose, rhamnose and galactose areknown as components of cellulose and hemicelluloses. They arereleased by brown-rot fungi during enzymatic wood decay. Allwater-soluble carbohydrates are ultimately consumed, so thatextensively brown-rotted wood consists almost entirely ofmodified lignin. Brown-rot fungi appear to depolymerize woodin the early stages of decay much more rapidly than the decayproducts can be metabolized. The occurrence of excess wood-decomposition productsmay help explain the frequent presence ofother wood scavengers in brown-rotted wood (Eriksson et al.,1990; Zabel and Morrell, 1992; Valaskova and Baldrian, 2006). Aswood decomposes, it passes through a range of decay stages, eachcolonized by a succession of diverse saproxylic insect assemblages(Grove, 2002). Wood-decay products may serve as chemical cuesfor these saproxylic insects and be involved in this colonizationprocess although no electrophysiological or behavioural evidenceis yet available. Some ground beetles, including P. oblongopuncta-
Fig. 5. Histograms of relative spike amplitudes from neurons stimulated by sugars
and amino acids in antennal taste sensilla of P. oblongopunctatus. aSN, a4thN and
aCN represent spike amplitudes of the sugar-, 4th chemo-, and salt-sensitive
neuron, respectively. Normal distribution curves of respective samples are
indicated.
E. Merivee et al. / Journal of Insect Physiology 54 (2008) 1213–12191218

95
tus, prefer brown-rotted wood for their prolonged hibernation(Haberman, 1968; Lindroth, 1986). Our results showed, however,that P. oblongopunctatuswas not able to sense water-soluble woodsugars released by brown-rot fungi, except for glucose, indicatingthat these chemicals are not involved in the search for hibernationsites by this species, although others may be.
Acknowledgements
This work was supported by contract grant sponsor EstonianScience Foundation (contract grant numbers: 6958 and 7391).
References
Albert, P.J., 2003. Electrophysiological responses to sucrose from a gustatorysensillum on the larval maxillary palp of the spruce budworm, Choristoneurafumiferana (Clem.) (Lepidoptera: Tortricidae). J. Insect Physiol. 49, 733–738.
Amakawa, T., 2001. Effects of age and blood sugar levels on the proboscis extensionof the blow fly Phormia regina. J. Insect Physiol. 47, 195–203.
Bernays, E.A., Chapman, R.F., 2001. Gustatory neuron responses in the polyphagousarctiid, Grammia geneura: towards a general pattern for caterpillars. J. InsectPhysiol. 47, 1029–1043.
Blaney, W.M., 1974. Electrophysiological responses of the terminal sensilla on themaxillary palps of Locusta migratoria (L.) to some electrolytes and non-electro-lytes. J. Exp. Biol. 60, 275–293.
Chapman, R.F., 2003. Contact chemoreception in feeding by phytophagous insects.Ann. Rev. Entomol. 48, 455–484.
Daly, P.J., Ryan, M.F., 1979. Ultrastructure of antennal sensilla of Nebria brevicollis(Fab) (Coleoptera: Carabidae). Int. J. Insect Morphol. Embryol. 8, 169–181.
Dethier, V.G., Evans, D.R., Rhoades, M.V., 1956. Some factors controlling the inges-tion of carbohydrates by the blowfly. Biol. Bull. 111, 204–222.
Eriksson, K.-E.L., Blanchette, R.A., Ander, P., 1990. Microbial and Enzymatic Degra-dation of Wood Components. Springer-Verlag, New York.
Grove, S.J., 2002. Saproxylic insect ecology and the sustainable management offorests. Ann. Rev. Ecol. Syst. 33, 1–23.
Haberman, H., 1968. Ground Beetles of Estonia. Valgus, Tallinn, p. 598 [in Estonian]Hanson, F.E., 1987. In: Chapman, R.F., Bernays, E.A., Stoffolano, J.G. (Eds.), Chem-
oreception in the Fly: The Search for the Liverwurst Receptor. Perspectives inChemoreception and Behaviour. Springer, Germany, pp. 99–122.
Heikkila, R., 1977. Elaimet kylvetynmannyn ja kuusen siemenen tuhoojina Pohjois-Suomessa [Summary: Destruction caused by animals to sown pine and spruceseed in northern Finland]. Metsantutkimuslaitoksen Julkaisuja 89, 1–35.
Hengeveld, R., 1980. Polyphagy, oligophagy and food specialization in groundbeetles (Coleoptera Carabidae). Netherlands J. Zool. 30, 564–584.
Hoback, W.W., Golick, D.A., Svatos, T.M., Spomer, S.M., Higley, L.G., 2000. Salinityand shade preferences result in ovipositional differences between sympatrictiger beetle species. Ecol. Entomol. 25, 180–187.
Irmler, U., 2001. Charakterisierung der Laufkafergemeinschaften schleswig-holstei-nischer Walder undMoglichkeiten ihrer okologischen Bewertung. AngewandteCarabidologie, Suppl 2. Laufkafer im Wald, pp. 21–32.
Johnson, U.E., Cameron, R.S., 1969. Phytophagous ground beetles. Ann. Entomol.Soc. Am. 62, 909–914.
Kim, J.L., Yamasaki, T., 1996. Sensilla of Carabus (Isiocarabus) fiduciarius saishutoicusCsiki (Coleoptera: Carabidae). Int. J. Insect Morphol. Embryol. 25, 153–172.
Krogerus, R., 1960. Okologische Studien uber nordische Moorarthropoden. SocietasScientiarum. Fennica Commentationes Biologicae 21 (3), 1–238.
Kromp, B., 1999. Carabid beetles in sustainable agriculture: a review on pest controlefficacy, cultivation impacts and enhancement. Agric. Ecosyst. Environ. 74,187–228.
Lindroth, C.H., 1986. Fauna Entomologica Scandinavica, Vol. 15: The Carabidae(Coleoptera) of Fennoscandia and Denmark, II. E.J. Brill, Leiden, pp. 230–497.With an appendix on the family Rhysodidae.
Liscia, A., Solari, P., 2000. Bitter taste recognition in the blowfly: electrophysiolo-gical and behavioral evidence. Physiol. Behav. 70, 61–65.
Liscia, A., Solari, P., Majone, R., Tomassini Barbarossa, I., Crnjar, R., 1997. Tastereceptor mechanisms in the blowfly: evidence of amiloride-sensitive andinsensitive receptor sites. Physiol. Behav. 62, 875–879.
Liscia, A., Crnjar, R., Masala, C., Sollai, G., Solari, P., 2002. Sugar reception in theblowfly: a possible Ca2+ involvement. J. Insect Physiol. 48, 693–699.
Lovei, G.L., Sunderland, K.D., 1996. Ecology and behavior of ground beetles (Coleop-tera: Carabidae). Ann. Rev. Entomol. 41, 231–256.
Merivee, E., Ploomi, A., Rahi, M., Luik, A., Sammelselg, V., 2000. Antennal sensilla ofthe ground beetle Bembidion lampros Hbst (Coleoptera Carabidae). Acta Zool.(Stockholm) 81, 339–350.
Merivee, E., Ploomi, A., Luik, A., Rahi, M., Sammelselg, V., 2001. Antennal sensilla ofthe ground beetle Platynus dorsalis (Pontoppidan, 1763) (Coleoptera Carabidae).Microsc. Res. Tech. 55, 339–349.
Merivee, E., Ploomi, A., Rahi, M., Bresciani, J., Ravn, H.P., Luik, A., Sammelselg, V.,2002. Antennal sensilla of the ground beetle Bembidion properans Steph.(Coleoptera Carabidae). Micron 33, 429–440.
Merivee, E., Renou, M., Mand, M., Luik, A., Heidemaa, M., Ploomi, A., 2004. Electro-physiological responses to salts from antennal chaetoid taste sensilla of theground beetle Pterostichus aethiops. J. Insect Physiol. 50, 1001–1013.
Merivee, E., Ploomi, A., Milius, M., Luik, A., Heidemaa, M., 2005. Electrophysiologicalidentification of antennal pH-receptors in the ground beetle Pterostichus oblon-gopunctatus (Coleoptera Carabidae). Physiol. Entomol. 30, 122–133.
Merivee, E., Must, A., Milius, M., Luik, A., 2007. Electrophysiological identification ofthe sugar cell in antennal taste sensilla of the predatory ground beetle Pter-ostichus aethiops. J. Insect Physiol. 53, 377–384.
Milius, M., Merivee, E., Williams, I., Luik, A., Mand, M., Must, A., 2006. A newmethodfor electrophysiological identification of antennal pH receptor cells in groundbeetles: the example of Pterostichus aethiops (Panzer, 1796) (Coleoptera, Car-abidae). J. Insect Physiol. 52, 960–967.
Mitchell, B.K., Gregory, P., 1979. Physiology of the maxillary sugar sensitive cell inthe red turnip beetle, Entomoscelis americana. J. Comp. Physiol. 132, 167–178.
Nystrand, O., Granstrom, A., 2000. Predation on Pinus sylvestris seeds and juvenileseedlings in Swedish boreal forests in relation to stand disturbance by logging. J.Appl. Ecol. 37, 449–463.
Paje, F., Mossakowski, D., 1984. pH-preferences and habitat selection in carabidbeetles. Oecologia (Berlin) 64, 41–46.
Ruth, E., 1976. Elektrophysiologie der Sensilla Chaetica auf den Antennen vonPeriplaneta americana. J. Comp. Physiol. 105, 55–64.
Sandoval, M., Albert, P.J., 2007. Chemoreception of sucrose and amino acids insecond and fourth instars of the spruce budworm Choristoneura fumiferana(Clem.) (Lepidoptera: Tortricidae). J. Insect Physiol. 53, 84–92.
Schoonhoven, L.M., van Loon, J.J.A., 2002. An inventory of taste in caterpillars: eachspecies its own key. Acta Zoologica Academiae Scientiarum Hungaricae 48,215–263.
Symondson, W.O.C., Sunderland, K.D., Greenstone, M.H., 2002. Can generalistpredators be effective biocontrol agents. Ann. Rev. Entomol. 47, 561–594.
Thiele, H.-U., 1977. Carabid Beetles in Their Environment. Zoophysiology andEcology 10. Springer, Berlin, p. 369.
Valaskova, V., Baldrian, P., 2006. Degradation of cellulose and hemicelluloses by thebrown rot fungus Piptoporus betulinus—production, of extracellular enzymesand characterization of the major cellulases. Microbiology 152, 3613–3622.
Zabel, R.A., Morrell, J.J., 1992. Wood Microbiology Decay and Its Prevention. Aca-demic Press, San Diego, pp. 195–224.
E. Merivee et al. / Journal of Insect Physiology 54 (2008) 1213–1219 1219
V

Komendant, M., Merivee, E., Must, A., Tooming, E., Williams, I.,
Luik, A.
ELECTROPHYSIOLOGICAL RESPONSE OF THE CHEMO-
RECEPTOR NEURONS IN THE ANTENNAL TASTE SEN-
SILLA TO PLANT ALKALOIDS AND GLUCOSIDES IN THE
GRANIVOROUS GROUND BEETLE PTEROSTICHUS OBLON-GOPUNCTATUS.
Journal of Chemical Ecology (in review)

ELECTROPHYSIOLOGICAL RESPONSE OF
THE CHEMORECEPTOR NEURONS IN THE
ANTENNAL TASTE SENSILLA TO PLANT
ALKALOIDS AND GLUCOSIDES
IN THE GRANIVOROUS GROUND BEETLE
Pterostichus oblongopunctatus
MARIT KOMENDANT, ENNO MERIVEE*, ANNE MUST, ENE
TOOMING, INGRID WILLIAMS, ANNE LUIK
Estonian University of Life Sciences
1 Kreutzwaldi Street, 51014 Tartu, Estonia

Abstract Electrophysiological responses of chemoreceptor neurons from
the antennal chaetoid taste sensilla to several plant alkaloids and glucosides
were tested in the ground beetle Pterostichus oblongopunctatus. For the
fi rst time, a specifi c alkaloid-sensitive neuron phasic-tonically responding
to quinine and quinine hydrochloride was found in these insects, most
probably related to their granivorous feeding habit. Th e response to qui-
nine hydrochloride was concentration-dependent at 0.001 to 50 mmol
l-1 with the response threshold at 0.01 mmol l-1, and a maximum rate of
fi ring of 67 spikes/sec at 50 mmol l-1. Th e stimulatory eff ect of caff eine
was very weak. In addition, both quinine and quinine hydrochloride
strongly inhibited spike production by the salt- and pH-sensitive neurons
when presented in mixtures with 10 mmol l-1 NaCl. It was also observed
that several tested plant secondary compounds, salicin, sinigrin, caff eine
and nicotine, which had only little or no eff ect on the fi ring rate of the
alkaloid-sensitive neuron, would greatly reduce the responses of the salt-
and pH-sensitive neurons, suggesting that the antennal taste sensilla of P. oblongopunctatus may detect plant defensive compounds both through the
activation of a specifi c alkaloid-sensitive neuron and through peripheral
inhibition of other chemoreceptor neurons of the taste sensillum.
Key Words Granivory, Contact chemoreception, Plant secondary com-
pounds, Alkaloid-sensitive neuron, Peripheral inhibition, Feeding deterrent

101
INTRODUCTION
Ground beetles (Coleoptera, Carabidae) are the dominant ground-living
invertebrate predators in many ecosystems. Because of their potential role
in foodweb dynamics and pest control, the food and feeding habits of
ground beetles have been investigated in more detail than of any other
group of predatory arthropods. Data on food items are available for ap-
proximately 1100 species and subspecies of ground beetles of the world. A
total of 73% are carnivorous, 19% are omnivorous and 8% are exclusively
herbivorous (Th iele 1977; Larochelle 1990; Toft and Bilde 2002). Of these,
many species predate on seeds (Honek et al. 2003). Ground beetles use a
variety of external stimuli in order to locate food. Behavioural tests and
observations have indicated that visual, tactile, olfactory and gustatory
cues may be involved (Wheater 1989; Kielty et al. 1996; Toft and Bilde
2002). Scanning electron microscopic studies have demonstrated that
thousands of various olfactory and gustatory sensilla are located on the
ground beetles’ antennae (Daly and Ryan 1979; Merivee et al. 2000, 2001,
2002). Very little is known, however, about the chemical compounds to
which they respond.
Electrophysiological investigations of the chemoreception capabilities of
ground beetles started only recently with experiments on antennal chaetoid
contact chemoreceptors. Approximately 70 of these large sensilla (35-
200 μm in length) have been found on the antennal fl agellum of various
ground beetles including the ground beetle Pterostichus oblongopunctatus (Merivee et al. 2000, 2001, 2002, 2005). Typically, four chemoreceptive
and one mechanoreceptive neuron innervate these sensilla in various in-
sect groups (Morita and Shiraishi 1985; Chapman 1998; Jørgensen et al.
2006). A similar set of sensory neurons has been found for the antennal
chaetoid contact chemoreceptors of the ground beetle Nebria brevicollis (Daly and Ryan 1979) suggesting that it may be characteristic for these
sensilla in P. oblongopunctatus, too, though, histological data for this
species are not available. In the ground beetles Pterostichus aethiops and P.
oblongopunctatus, specifi c chemical stimuli have been identifi ed by elect-
rophysiology for three of the four chemosensory neurons. Two respond
to various electrolyte solutions. By their specifi c response characteristics
these neurons were classifi ed as salt- (cation-) and pH-sensitive neurons
(Merivee et al. 2004, 2005; Milius et al. 2006). Th e third chemoreceptor
neuron is sensitive to sucrose, maltose and glucose, important in plant

102
carbohydrate metabolism (Merivee et al. 2007, 2008), most probably
related to granivory of these two forest species (Heikkilä 1977; Nystrand
and Granström 2000). Th e action potentials (spikes) produced by the
salt-, pH-, and sugar-sensitive neurons can be distinguished by their wave-
form, polarity and amplitude. Th e pH-sensitive neuron generates large
two-tipped spikes with positive polarity. In contrast, the polarity of the
one-tipped spikes from the salt- and sugar-sensitive neurons is negative.
It has been also observed that the spikes generated by the sugar-sensitive
neuron are shorter in amplitude compared to those generated by the
salt-sensitive neuron (Merivee et al. 2004, 2005, 2007). Carbohydrates
function as strong phagostimulants to most herbivorous insects, equipped
with specialized receptors to detect these compounds (Chapman 1998;
Schoonhoven and van Loon 2002; Chapman 2003) suggesting that these
chemicals may have phagostimulatory role in herbivorous and omnivorous
ground beetles, too. Unfortunately, no data exist on the function of the
4th chemoreceptor neuron probably also located in these antennal sensilla
of ground beetles, including P. oblongopunctatus (Daly and Ryan 1979;
Merivee et al. 2008).
In addition to detecting phagostimulants, most animals, including her-
bivorous insects, possess chemoreceptor neurons responding to a diverse
range of feeding deterrent compounds. When stimulated, deterrent neurons
reduce or fully stop feeding activity (Dethier 1980; Schoonhoven and van
Loon 2002; Chapman 2003; Wang et al. 2004). Natural deterrent stimuli
recognized by chemoreceptor neurons of herbivorous insects constitute
the largest and most structurally diverse class of gustatory stimuli. It has
been shown that plant glucosides, for example, sinigrin and salicin, and
various alkaloids like nicotine (pyrrolidine group), strychnine and quinine
(quinoline group), caff eine (purine group) and several others stimulate the
deterrent neuron in many herbivorous insects. Th reshold concentrations
for compounds that stimulate a deterrent neuron vary from 10-7 to 10-2
mol l-1, but the majority of these are in the range of 10-4 to 10-2 mol l-1.
Th erefore, deterrent neurons fulfi l a central role in host-plant recognition
and have, since their discovery (Ishikawa 1966), attracted much inter-
est (Glendinning 2002; Ryan 2002; Schoonhoven and van Loon 2002;
Chapman 2003; Jørgensen et al. 2007).
Herbivory, including pre- and post-dispersal plant seed predation, is ex-
tremely common in virtually all ecosystems. Seeds with their very high

103
nutrient values per unit volume are extremely attractive to granivores as
food (Janzen 1971). To counterbalance the eff ects of herbivory, plants
have developed a broad spectrum of secondary metabolites involved in
plant defense, which are collectively known as antiherbivory compounds
and can be classifi ed into three main sub-groups: nitrogen compounds
(including alkaloids, cyanogenic glycosides and glucosinolates), terpenoids,
and phenolics. Th ese compounds can act as feeding deterrents or toxins
with various modes of action to herbivores, or reduce plant digestibility
(Janzen 1971; Jolivet 1998; Ryan 2002; Chapman 2003; Hulme and
Benkman 2004; Davis et al. 2008). Seeds are sources of some of the most
toxic natural products known to humans and the secondary chemicals in
seeds present formidable challenges to granivores. However, seed chemical
defenses can only be assessed with reference to specifi c target organisms
since a secondary chemical may not be equally toxic to all granivores.
Some seeds are so toxic that all seed-predators avoid them (Harborne 1993;
Hulme and Benkman 2004). Most toxins are usually in small concentra-
tions (<5%) in the seed and some alkaloids, for example, can be lethal
at concentrations as low as 0.1% (Hatzold et al. 1983; Harborne 1993).
Unfortunately, there is no evidence that plant secondary compounds cause
feeding deterrency in ground beetles.
However, due to selective seed predation by ground beetles (Toft and
Bilde, 2002; Tooley and Brust, 2002), the presence of a deterrent neuron
in their antennal contact chemoreceptors should be expected. We tested
the stimulatory eff ect of some plant alkaloids and glucosides on the che-
moreceptor neurons innervating the antennal chaetoid taste sensilla in
the omnivorous P. oblongopunctatus, known as an important predator
on conifer seeds (Heikkilä 1977; Nystrand and Granström 2000). Th ese
experiments are reported in this paper.

104
METHODS AND MATERIALS
Insects Adult test beetles of P. oblongopunctatus were collected from a local
population in southern Estonia in September and October 2008. Th ey
were kept in 30×20×10 cm plastic boxes fi lled with moistened moss and
pieces of brown-rotted wood in a refrigerator at 5-6 ºC. A week prior
to the experiments, the beetles were transferred to room temperature
(22 ºC), fed with moistened cat food (Friskies Vitality+, Nestlé Purina,
Hungary) and given clean water to drink every day. Electrophysiological
experiments were conducted from November 2009 to February 2010.
Electrophysiology Th e beetle to be tested was intact and restrained securely
in a conical tube made of thin sheet-aluminium of a size that allowed the
head and antennae to protrude from the narrower end. Th e wider rear
end of the tube was blocked with plasticine to prevent the beetle from
retreating out of the tube. Its antennae were fi xed horizontally to the
edge of an aluminium stand with special clamps and beeswax, so that the
horizontally-located large chaetoid taste sensilla were visible from above
under a light microscope, and easily accessible for micromanipulation from
the side. Care was taken not to contaminate the taste sensilla by contact
with instruments or beeswax. Each prepared test beetle was placed in a
Faraday cage (15x10x5 cm) for electrophysiology.
Spikes from chemoreceptor neurons of the sensilla were recorded by the
conventional single sensillum tip-recording technique (Hodgson et al.
1955). To achieve a good signal to noise ratio, the indiff erent tungsten
microelectrode was inserted into the base of the fl agellum free of muscular
tissue. It was connected to an AC amplifi er via a calibration source, and
earthed. Th e recording glass micropipette fi lled with stimulating solu-
tion was brought into contact with the tip of the sensillum by means of
a micromanipulator under visual control through a light microscope at
a magnifi cation of 300X. Approximately, two sec before each stimula-
tion, 2-3 drops of solution were squeezed out of the micropipette tip by
a syringe connected to the rear end of the micropipette by means of a
silicone tube, and adsorbed onto a piece of fi lter paper attached to another
micromanipulator, in order to avoid concentration changes in the capil-
lary tip due to evaporation. During each 5 sec stimulation period, spikes
picked up by the recording electrode were fi ltered with a bandwidth set at
100–2000 Hz, amplifi ed (input impedance 10 GΩ; amplifi cation factor

105
2000X), monitored on an oscilloscope screen, and relayed to a computer
via an analogue-to-digital input board DAS-1401 (Keithley, Taunton,
MA, USA) for data acquisition, storage, and analysis using TestPoint
software (Capital Equipment Corp., Billerica, MA, USA) at a sampling
rate of 10 kHz. Responses were recorded from neurons of two to three
randomly-selected taste sensilla on the fl agellomeres 1 to 9 of each test
beetle. Each sensillum was stimulated twice, with control (10 mmol l-1
NaCl) and test stimulus solution, respectively. In each subsequent test
beetle, the order of stimulation was reversed.
Chemicals Sodium chloride, sucrose, sinigrin, salicin and caff eine were
purchased from AppliChem (Germany). Quinine, quinine hydrochloride
dihydrate, nicotine and L-strychnine were obtained from Sigma-Aldrich
company. Except for quinine and strychnine, stimulating chemicals, at
concentration of 10 mmol l-1, and were diluted in 10 mmol l-1 sodium
chloride. Due to their poor solubility in water, 1 mmol l-1 quinine and
0.1 mmol l-1 strychnine in 10 mmol l-1 sodium chloride were used in
the experiments. Additionally, to assess the threshold concentration and
concentration/response curve for one of the most active compounds, qui-
nine hydrochloride dihydrate, a water soluble salt of quinine, was tested
at concentrations ranging from 0.001 to 50 mmol l-1. All solutions were
kept at 5°C for less than one week.
Feeding bioassays Two-choice feeding experiments were carried out in
110-mm diameter glass Petri dishes. To ensure high air humidity inside
the dishes, each was lined with a moistened Whatman fi lter paper circle
(Schleicher & Schuell, England). Two pine seeds (Pinus sibirica), treated
for 20 min with distilled water (control) and 0.01 to 50 mmol l-1 quinine
hydrochloride, respectively, were placed in the centre of each Petri dish,
50 mm from each other. Only seeds with seed coats removed were used
in the experiments. Test beetles, one in each Petri dish, were allowed to
choose between, and feed on the seeds for 24 h (20 °C, 14:10 light:dark).
After 24 h, the beetles were removed and the percentage of predation on
control seeds and seeds treated with alkaloid, was visually determined.
At each concentration of quinine hydrochloride, the feeding preference
of the beetles was tested in fi ve replications (10 beetles in each).
Data analysis Automated classifi cation and counting of action potentials
from taste sensilla was not an appropriate method for data analysis be-

106
cause two action potentials frequently coincided to produce an irregular
waveform. Instead, spikes from several chemosensory neurons innervating
antennal taste sensilla tested were visually distinguished by their polarity
and amplitude (Fig. 1), and counted, using TestPoint software. Firing rates
(spikes/sec) were analysed by counting the number of action potentials
within 5 sec after stimulus onset. Mean fi ring rates and their standard
errors were calculated. Paired sample Wilcoxon signed rank was used to
determine the signifi cance of diff erences between means. Spike amplitude
analysis was performed in order to explain which specifi c neuron was
stimulated by the chemical classes tested.

107
RESULTS
Spike shapes and amplitudes evoked by tested chemicals Spike waveform
analysis showed that spikes from four diff erent chemoreceptor neurons
innervating antennal taste sensilla of the ground beetle Pterostichus oblon-gopunctatus could be distinguished in the recordings obtained in response
to tested chemicals. Earlier, three of the neurons, the salt- (Merivee et al.
2004), pH- (Merivee et al. 2005; Milius et al. 2006) and sugar-sensitive
neurons (Merivee et al. 2007, 2008), were identifi ed in this species. Spikes
with negative polarity, 1–2 mV in amplitude, arose from the salt-sensi-
tive neuron. Th is neuron generated spikes in a phasic-tonic manner and
had an extremely short response latency lasting less than 10 msec (Fig.
1A–J). Two-tipped spikes with positive polarity, 2–4 mV in amplitude,
were generated by the pH-sensitive neuron (Merivee et al. 2005; Milius
et al. 2006). Compared to the spikes from the salt-sensitive neuron, these
positive spikes appeared with a much longer delay (Fig. 1A,F). Th e sugar-
sensitive neuron also produced spikes with negative polarity, but smaller
in amplitude than those of the salt-sensitive neuron (Fig. 1J). Some tested
alkaloids (quinine, caff eine) and an alkaloid salt (quinine hydrochloride)
induced a neuron of the taste sensilla to produce spikes with negative
polarity, but with a considerably longer period of latency compared to
the spikes of the salt- and sugar-sensitive neurons (Fig. 1B–E). Spike
amplitude histograms show two maxima, indicating that spikes evoked
by salt (NaCl), sugar (sucrose) and quinine hydrochloride were generated
by three diff erent neurons, the salt-, sugar- and alkaloid-sensitive neuron,
respectively (Fig. 2). Spike amplitudes of the chemoreceptor neurons from
diff erent taste sensilla varied to a some extent, from 0.7 to 2 mV (Fig.
1C,E). As a result, in most cases, spike amplitudes generated by the alka-
loid-sensitive neuron were shorter than those of the salt-sensitive neuron
(Fig. 1B,C). In some cases, however, opposite spike amplitude ratios of
the neurons were observed (Fig. 1D).
Eff ect of plant alkaloids and glucosides on the responses of the antennal che-
moreceptor neurons Only two of the tested chemicals (Table 1), 1 mmol l-1
quinine and 10 mmol l-1quinine hydrochloride, were highly stimulating for
the antennal alkaloid-sensitive neuron, evoking 8.9 and 22.8 spikes/sec,
respectively. Th e stimulatory eff ect of 10 mmol l-1 caff eine was very weak,
2.4 spikes/sec. In contrast, some inhibiting eff ect of strychnine and sinigrin
to the activity of the neuron was observed. Respective fi ring rates were

108
very low, however (Table 1). Other plant chemicals did not considerably
aff ect the spike production of the alkaloid-sensitive neuron. Phasic-tonic
response of the alkaloid-sensitive neuron to quinine hydrochloride was
concentration dependent over the range of 0.001 to 50 mmol l-1 with the
response threshold at 0.01 mmol l-1, and maximum rate of fi ring equal
to 67 spikes/sec at 50 mmol l-1 (Figs. 3 and 4). However, compared to
the spike trains of the salt-sensitive neuron (Fig. 1; Fig. 5A–F), the phasic
component of the response produced by the alkaloid-sensitive neuron was
less pronounced (Fig. 1B–E; Fig. 4; Fig. 5C–F).
In addition to stimulation of the alkaloid-sensitive neuron, both quinine
and quinine hydrochloride strongly inhibited spike generation by the salt-
and pH-sensitive neurons when presented in mixtures with 10 mmol l-1
NaCl (Table 1). Increasing the quinine hydrochloride concentration while
keeping the concentration of NaCl constant at 10 mmol l-1 caused a dose-
dependent decrease in spike generation by the salt-sensitive neuron (Figs.
3–5). In most tested sensilla, at concentrations above 10 mmol l-1 quinine
hydrochloride, only the alkaloid-sensitive neuron fi red while the fi ring
rate of the salt-sensitive neuron of the taste sensillum decreased close to
zero during the 5 sec adaptation period (Fig. 4). In response to 10 mmol
l-1 NaCl alone, the mean number of spikes produced by the pH-sensitive
neuron was relatively low compared to that of the salt-sensitive neuron,
and varied to some extent in diff erent sensilla (Table 1). At quinine hy-
drochloride concentrations above 0.01 mmol l-1, the mean fi ring rate of
the pH-sensitive neuron dropped to zero (Fig. 3). It was also observed that
nicotine and salicin, which did not aff ect the fi ring rate of the alkaloid-
sensitive neuron, greatly reduced the response of the pH-sensitive neuron
(Table 1). Sinigrin and caff eine suppressed neural activity of the salt- and
pH-sensitive neuron, respectively. Th is suggested that the antennal taste
sensilla of P. oblongopunctatus may detect plant defensive compounds not
only through the activation of specifi c alkaloid-sensitive neuron but also
through inhibition of the salt- and pH-sensitive neurons.
Eff ect of quinine hydrochloride on the feeding response of P. oblongopunctatus
Feeding choice experiments showed that one of the most stimulating
chemicals for the alkaloid-sensitive neuron, quinine hydrochloride, had a
concentration dependent deterrent eff ect on seed predation in the ground
beetle P. oblongopunctatus, with the threshold concentration at 1 mmol l-1.
Th us, the threshold concentration of this alkaloid to evoke a behavioural

109
response was a hundred times higher than that for an electrophysiological
response of the alkaloid-sensitive neuron. At higher concentrations, the
deterrent eff ect of quinine hydrochloride was very strong. It was observed
that pine seeds treated with 50 mmol l-1 quinine hydrochloride were never
predated by the beetles (Fig. 6).

110
DISCUSSION
Th e experimental data presented show that the “fourth” chemoreceptor
neuron in the antennal taste sensilla of the ground beetle Pterostichus oblongopunctatus may respond to some plant alkaloids such as quinine.
Th is fi nding is in agreement with literature data on the feeding habits of
this species. P. oblongopunctatus is one of the most common and wide-
spread ground beetles in the forests of northern Europe (Lindroth 1985;
Niemelä et al. 1994). In most stands of Pinus sylvestris, seed predation
results in 20% seed mortality, although occasionally it reaches 60%.
Post-dispersal pine seed predation in the Swedish boreal forests studied
was due mainly to the two ground beetle species P. oblongopunctatus and
Calathus micropterus. Both are able to consume conifer seeds. Th e larger
species, P. oblongopunctatus, is considered to be the more important pine
seed predator of the two (Heikkilä 1977; Nystrand and Granström 2000).
Like many herbivores, numerous insect seed-predators have biochemical
adaptations to deal with toxic plant secondary compounds, including vari-
ous detoxifi cation and sequestration mechanisms (Jolivet 1998; Hulme and
Benkman 2004; Schoonhoven et al. 2005; Després et al. 2007). However,
the diffi culties of dealing with more than a few diff erent kinds of defen-
sive compounds has favoured specialization in seed-predators and helps
to explain why a large proportion of them are specialist feeders (Hulme
and Benkman 2004). Th e omnivorous ground beetle P. oblongopunctatus predates on many small animals, most frequently on Coleoptera, Diptera,
Collembola, Acari etc. (Larochelle 1990), as well as on conifer seeds
(Heikkilä 1977; Nystrand and Granström 2000). Probably, the species
feeds on seeds of various plants but respective data are not available. Se-
lective seed predation has been observed in many other ground beetles,
however (Toft and Bilde, 2002; Tooley and Brust, 2002).
To discriminate between toxic and nontoxic seeds, P. oblongopunctatus
possesses a specifi c feeding deterrent neuron innervating its antennal chae-
toid taste sensilla. A strong stimulatory eff ect of quinine (quinoline group)
and quinine hydrochloride to this neuron was observed. Th e threshold
concentration for the latter at 0.01 mmol l-1 was close to the upper limit
of the threshold concentrations for the plant secondary compounds that
stimulate a deterrent neuron in other herbivorous insects (Schoonhoven
and van Loon 2002; Chapman 2003; Schoonhoven et al. 2005), probably

111
due to the fact that toxin concentration in seeds is usually much higher
than in other parts of the plant (Janzen 1971; Hulme and Benkman 2004;
Davis et al. 2008). Th e response of the neuron to quinine hydrochloride
was concentration dependent with maximum rates of fi ring reaching up
to 70 spikes/secat 50 mmol l-1. In two-choice behavioural experiments,
quinine hydrochloride had a strong concentration dependent deterrent
eff ect on feeding of P. oblongopunctatus beetles with the response thresh-
old at 1 mmol l-1. Th us, the threshold concentration of this alkaloid to
evoke a behavioural response was a hundred times higher than that for an
electrophysiological response of the alkaloid-sensitive neuron. In contrast
to quinine hydrochloride, the response of the alkaloid-sensitive neuron
to caff eine (purine group) was very weak, only 2.4 spikes/sec at 10 mmol
l-1 concentration. Th e eff ect of another quinoline alkaloid, strychnine,
and sinigrin, to the neuron was rather inhibitory. No response of the
neuron to tested pyrrolidine alkaloid, nicotine, and salicin (glucoside)
was observed suggesting that the antennal alkaloid-sensitive neuron of P. oblongopunctatus is not equally sensitive to all plant defensive chemicals.
Nevertheless, the list of toxic compounds capable of stimulating the an-
tennal alkaloid-sensitive neuron of P. oblongopunctatus is likely to grow
longer as more plant defence chemicals are tested.
Quinoline alkaloids are among the most potent insect feeding deterrents
occurring mainly in Rutaceous and Rubiaceous plants. Th ey can be pres-
ent in virtually every part of the plant or just a specifi c part like the bark,
rhizome, leaf or seed (Openshaw 1967; Michael 2003; Aniszewski 2007).
Th ese alkaloids represent a large and structurally diverse group of plant
defensive compounds. When contacted, some of them may be detected by
stimulation of a specifi c antennal alkaloid-sensitive neuron of the ground
beetles predating on seeds, similarly to quinoline alkaloid-sensitivity of
P. oblongopunctatus. Th e ability of granivores to perceive plant defensive
compounds allows them to avoid toxic seeds, and survive. High sensitiv-
ity of P. oblongopunctatus to plant sugars, commonly known as strong
phagostimulants for most phytophagous insects, seems to be also related
to its partially granivorous feeding habits. Th e disaccharide maltose was
one of the most stimulatory sugars for the sugar-sensitive neuron also
innervating antennal taste sensilla of P. oblongopunctatus (Merivee et al.
2008). Maltose is uncommon in nature, it can be found in considerable
amounts in germinating seeds as a product of the enzymatic degradation
of starch.

112
While stimulating the alkaloid-sensitive neuron, quinine hydrochloride
strongly reduced spike production by the salt- and pH-sensitive neurons
which also innervate these sensilla in Pterostichus spp. (Merivee et al.
2004, 2005; Milius et al. 2006). Increasing the quinine hydrochloride
concentration, while keeping the concentration of NaCl constant, caused
a dose-dependent decrease in spike generation by the salt-sensitive neu-
ron. At 10 mmol l-1 and higher concentrations of quinine hydrochloride,
predominant fi ring of the alkaloid-sensitive neuron was observed while
the fi ring rate of other neurons dropped almost to zero. It was also ob-
served that several tested plant secondary compounds, salicin, sinigrin,
caff eine and nicotine, which had only little or no eff ect on the fi ring rate
of the alkaloid-sensitive neuron, would greatly reduce the responses of
the salt- and pH-sensitive neurons of the antennal taste sensillum of P. oblongopunctatus. Th ese results suggest that the antennal taste sensilla in
P. oblongopunctatus may detect toxic and deterrent plant secondary com-
pounds not only through activation of the specifi c alkaloid-sensitive neuron
but also through inhibition of taste neurons activated by salts and stimulus
pH; this is similar to deterrent inhibition of taste neurons activated by
sugars and water in Drosophila (Meunier et al. 2003). Since foodplant
acceptability to herbivores depends on the balance between stimulation
and deterrency, similar interactions between stimulating chemicals at
peripheral taste neurons of herbivorous insects are of widespread occur-
rence. Plant toxins and deterrents may inhibit sugar neurons (Frazier 1986;
van Loon 1990; Messchendorp et al. 1996; Bernays and Chapman 2000;
Schoonhoven and van Loon 2002; Meunier et al. 2003; Schoonhoven
et al. 2005), water neurons (Meunier et al. 2003), and sugars and salts
may inhibit deterrent neurons (Simmonds and Blaney 1983; Shields and
Mitchell 1995; Glendinning et al. 2000; Schoonhoven and van Loon
2002; Schoonhoven et al. 2005). Deterrent compounds that on their
own do not stimulate any neuron within a sensillum may also decrease
the responsiveness of a neuron responding to a nutrient, as exemplifi ed
by sinigrin inhibiting the inositol neuron in Heliothis virescens and H. subfl exa (Bernays and Chapman 2000; Schoonhoven and van Loon 2002).
Survival of most terrestrial animals requires ability to detect environmental
NaCl, a nutrient essential for fl uid and electrolyte homeostasis and for a
multitude of other physiological processes (Lindemann 1996; Contreras
and Lundy 2000). Th e gustatory system allows insects to detect and ingest
salt, to discriminate between diff erent salts, and to avoid high salt concen-

113
trations. However, there are only few data that relate input from inorganic
salts to feeding behaviour in phytophagous insects. In some insects, salts
have a deterrent eff ect on feeding (Chapman 2002; Schoonhoven and
van Loon 2003). In contrast, larvae of Drosophila show dose-dependent
responses to NaCl: the appetitive responses to low concentration gradually
turn into aversion as concentration is increased up to 200 mmol l-1 and
higher (Miyakawa 1982; Liu et al. 2003; Niewalda et al. 2008). Some other
insects also respond positively to dilute salts. Cockroaches and housefl ies
prefer dilute sodium and potassium chlorides and refuse distilled water.
In the two-choice situation, many animals give a bimodal response to
sodium chloride. Acceptance at low concentrations switches to rejection
at high concentrations (Dethier 1977). In these cases, inorganic salt at
low concentration may act as a phagostimulant. Th erefore, NaCl at 10
mmol l-1 may also have a phagostimulatory eff ect in P. oblongopunctatus although no behavioural data is available. Th is assumption could explain
the inhibition of spike production of the salt-sensitive neuron by some
toxic compounds in P. oblongopunctatus as being similar to how plant
deterrents inhibit activity of other phagostimulatory neurons in other
phytophagous insects.
In conclusion we state that
• the occurrence of an alkaloid-sensitive neuron in the antennal taste
sensilla of the ground beetle P. oblongopunctatus was documented;
• the alkaloid-sensitive neuron found is most probably related to the
granivorous feeding habit of the species;
• some tested compounds (quinine and quinine hydrochloride) that
stimulate the alkaloid-sensitive neuron may inhibit activity of other
neurons (salt- and pH-sensitive neuron) of the sensillum;
• several tested plant secondary compounds, salicin, sinigrin, caff eine
and nicotine, which had only little or no eff ect on the fi ring rate of
the alkaloid-sensitive neuron, through peripheral inhibition, would
greatly reduce the responses of other chemoreceptor neurons of the
sensillum.

114
Acknowledgements Th e study was supported by Estonian target fi nancing
project SF0170057s09 and Estonian Science Foundation grant no. 6958.

115
REFERENCES
ANISZEWSKI, T. 2007. Alkaloids – secrets of life. Alkaloid chemistry,
biological signifi cance, applications and ecological role. P 334. Else-
vier, Amsterdam, Boston, Heidelberg, London, New York, Oxford,
Paris, San Diego, San Francisco, Singapore, Sydney, Tokyo.
BERNAYS, E. A. and CHAPMAN, R. F. 2000. A neurophysiological
study of sensitivity to a feeding deterrent in two sister species of
Heliothis with diff erent diet breadths. J. Insect Physiol. 46:905-912.
CHAPMAN, R. F. 1998. Th e Insects. Structure and Function. 4th edn.
P. 770. Cambridge University Press, Cambridge.
CHAPMAN, R. F., 2003. Contact chemoreception in feeding by phy-
tophagous insects. Annu. Rev. Entomol. 48:455-484.
CONTRERAS, R. J. and LUNDY, R. F. 2000. Gustatory neuron types
in the periphery: a functional perspective. Physiol. Behav. 69:41-52.
DALY, P. J. and RYAN, M.F. 1979. Ultrastructure of antennal sensilla of
Nebria brevicollis (Fab.) (Coleoptera: Carabidae). Int. J. Insect Morphol.
Embryol. 8:169-181.
DAVIS, A. S., SCHUTTE, B. J., IANNUZZI, J. and RENNER, K.
A. 2008. Chemical and physical defense of weed seeds in relation to
soil seedbank persistence. Weed Sci. 56: 676-684.
DESPRÉS, L., DAVID, J. P. and GALLET, C. 2007. Th e evolution-
ary ecology of insect resistance to plant chemicals. Trends Ecol. Evol. 22:298-307.
DETHIER, V. G. 1977. Th e taste of salt. Amer. Sci. 65, 744-751.
DETHIER, V. G. 1980. Evolution of receptor sensitivity to second-
ary plant substances with special reference to deterrents. Am. Nat. 115:45–66.
FRAZIER, J. L. 1986. Th e perception of plant allelochemicals that in-
hibit feeding. Pp. 1-42 in L. B. BRATTSTEN and S. AHMAD
(eds.). Molecular aspects of insect-plant associations. Plenum Press,
New York.
GLENDINNING, J. I. 2002. How do herbivorous insects cope with
noxious secondary plant compounds in their diet? Entomol. Exp. Appl. 104:15-25.
GLENDINNING, J. I., NELSON, N. and BERNAYS, E. A. 2000.
How do inositol and glucose modulate feeding in Manduca sexta
caterpillars? J. Exp. Biol. 203: 1299-1315.

116
HARBORNE, J. B. 1993. Introduction to Ecological Biochemistry. 4th
edn. P. 318. Academic Press, London, San Diego.
HATZOLD, T., ELMADFA, I., GROSS, R., WINK, M., HART
MANN, T. and WITTE, L. 1983. Quinolizidine alkaloids in seeds
of Lupinus mutabilis. J. Agric. Food Chem. 31:934-930.
HEIKKILÄ, R. 1977. Eläimet kylvetyn männyn ja kuusen siemenen
tuhoojina Pohjois-Suomessa (Summary: Destruction caused by ani-
mals to sown pine and spruce seed in northern Finland). Comm. Inst. Forest. Fenniae 89:1-35.
HODGSON, E. S., LETTVIN, J. Y. and ROEDER, K. D. 1955. Physi-
ology of a primary chemoreceptor unit. Science 122:417-418.
HONEK, A., MARTINKOVA, Z. and JAROSIK, V. 2003. Ground
beetles (Carabidae) as seed predators. Eur. J. Entomol. 100:531-544.
HULME, P. E. and BENKMAN C. W. 2004. Granivory. Pp. 132-
154 in C. M. HERRERA and O. PELLMYR (eds.). Plant-animal
interactions. An evolutionary approach. Blackwell Science, Oxford.
ISHIKAWA, S. 1966. Electrical response and function of bitter substance
receptor associated with the maxillary sensilla of the larva of the
silkworm, Bombyx mori L. J. Cell Comp. Physiol. 67:1-12.
JANZEN, D. H. 1971. Seed predation by animals. Annu. Rev. Ecol.
Syst. 2:465-492.
JOLIVET, P. 1998. Interrelationships between insects and plants. P. 309.
CRC Press, Boca Raton, Florida.
JØRGENSEN, K., ALMAAS, T. J., MARIONPOLL, F. and MU
STAPARTA, H. 2007. Electrophysiological characterization of re-
sponses from gustatory receptor neurons of sensilla chaetica in the
moth Heliothis virescens. Chem. Senses 32:863-879.
JØRGENSEN, K., KVELLO, P., ALMAAS, T. J. and MUSTAPARTA,
H. 2006. Two closely located areas in the suboesophageal ganglion
and the tritocerebrum receive projections of gustatory receptor neu-
rones located on the antennae and the proboscis in the moth Heliothis
virescens. J. Comp. Neurol. 496:121-134.
KIELTY, J. P., ALLENWILLIAMS, L. J., UNDERWOOD, N. and
EASTWOOD, E. A. 1996. Behavioural responses of three species
of ground beetle (Coleoptera: Carabidae) to olfactory cues associated
with prey and habitat. J. Insect Physiol. 9:237-250.
LAROCHELLE, A. 1990. Th e food of the carabid beetles (Coleoptera:
Carabidae, including Cicindelinae). Fabreries, Supplément 5:1-132.
LINDEMANN, B. 1996. Taste reception. Physiol. Rev.76:719-766.

117
LINDROTH, C. H. 1985. Th e Carabidae (Coleoptera) of Fennoscandia
and Denmark. Fauna Entomol. Scand. 15:1-225.
LIU, L., LEONARD, A. S., MOTTO, D. G., FELLER, M. A., PRICE, M. P., JOHNSON, W. A. and WELSH, M. J. 2003. Contribution
of Drosophila DEG/ENaC Genes to Salt Taste. Neuron 39:133-146.
MERIVEE, E., MUST, A., MILIUS, M. and LUIK, A. 2007. Electro-
physiological identifi cation of the sugar cell in antennal taste sensilla
of the predatory ground beetle Pterostichus aethiops. J. Insect Physiol. 53:377-384.
MERIVEE, E., MÄRTMANN, H., MUST, A., MILIUS, M., WIL
LIAMS, I. and MÄND, M. 2008. Electrophysiological responses
from neurons of antennal taste sensilla in the polyphagous predatory
ground beetle Pterostichus oblongopunctatus (Fabritius 1787) to plant
sugars and amino acids. J. Insect Physiol. 54:1213-1219.
MERIVEE, E., PLOOMI, A., LUIK, A., RAHI, M. and SAM
MELSELG, V. 2001. Antennal sensilla of the ground beetle Platynus dorsalis (Pontoppidan, 1763) (Coleoptera, Carabidae). Microsc. Res.
Tech. 55:339-349.
MERIVEE, E., PLOOMI, A., MILIUS, M., LUIK, A. and HEIDE
MAA, M. 2005. Electrophysiological identifi cation of antennal pH-
receptors in the ground beetle Pterostichus oblongopunctatus (Coleop-
tera, Carabidae). Physiol. Entomol. 30: 122-133.
MERIVEE, E., PLOOMI, A., RAHI, M., BRESCIANI, J., RAVN, H.
P., LUIK, A. and SAMMELSELG, V. 2002. Antennal sensilla of the
ground beetle Bembidion properans Steph. (Coleoptera, Carabidae).
Micron 33:429-440.
MERIVEE, E., PLOOMI, A., RAHI, M., LUIK, A. and SAMMEL
SELG, V. 2000. Antennal sensilla of the ground beetle Bembidion lam-
pros Hbst (Coleoptera, Carabidae). Acta Zool. (Stockholm) 81:339-350.
MERIVEE, E., RENOU, M., MÄND, M., LUIK, A., HEIDEMAA,
M. and PLOOMI A., 2004. Electrophysiological responses to salts
from antennal chaetoid taste sensilla of the ground beetle Pterostichus aethiops. J. Insect Physiol. 50:1001-1013.
MESSCHENDORP, L., VAN LOON, J. J. A. and GOLS, G. J. Z.,
1996. Behavioural and sensory responses to drimane antifeedants in
Pieris brassicae larvae. Entomol. Exp. Appl. 79:195-202.
MEUNIER, N., MARIONPOLL, F., ROSPARS, J. P. and TANI
MURA, T. 2003. Peripheral coding of bitter taste in Drosophila. J.
Neurobiol. 56:139-152.

118
MICHAEL, J. P. 2003. Quinoline, quinazoline and acridone alkaloids.
Nat. Prod. Rep., 20:476-493.
MILIUS, M., MERIVEE, E., WILLIAMS, I., LUIK, A., MÄND, M.
and MUST, A. 2006. A new method for electrophysiological identifi -
cation of antennal pH receptor cells in ground beetles: the example of
Pterostichus aethiops (Panzer, 1796) (Coleoptera, Carabidae). J. Insect
Physiol. 52:960-967.
MIYAKAWA, Y. 1982. Behavioural evidence for the existence of sugar,
salt and amino acid taste receptor cells and some of their properties
in Drosophila larvae. J. Insect Physiol. 28:405-410.
MORITA, H. and SHIRAISHI, A. 1985. Chemoreception physiol-
ogy. Pp. 133-170 in G. A. KERKUT and L. I. GILBERT (eds.).
Comprehensive Insect Physiology, Biochemistry and Pharmacology.
Pergamon Press, Oxford,.
NIEMELÄ, J., SPENCE, J. R., LANGOR, D., HAILA, Y. and TUKIA,
H. 1994. Logging and boreal ground-beetle assemblages on two con-
tinents: implications for conservation. Pp. 29-50 in K. GASTON, M.
SAMWAYS and T. NEW (eds.). Perspectives in insect conservation.
Intercept Publications, Andover.
NIEWALDA, T., SINGHAL, N., FIALA, A., SAUMWEBER, T.,
WEGENER S. and GERBER, B. 2008. Salt Processing in Larval
Drosophila: Choice, Feeding, and Learning Shift from Appetitive to
Aversive in a Concentration-Dependent Way. Chem. Senses 33:685-
692.
NYSTRAND, O. and GRANSTRÖM, A. 2000. Predation on Pinus sylvestris seeds and juvenile seedlings in Swedish boreal forests in
relation to stand disturbance by logging. J. Appl. Ecol. 37:449-463.
OPENSHAW, H. T. 1967. Quinoline alkaloids other than those of Cinc-
hona. Pp. 223-267 in R. H. F. MANSKE (ed.). Th e Alkaloids. Che-
mistry and Physiology. Vol. IX. Academic Press, New York, London.
RYAN, M. F. 2002. Insect Chemoreception. Fundamental and Applied.
P. 330. Kluwer Academic Publishers, New York, Boston, Dordrecht,
London, Moscow.
SCHOONHOVEN, L. M. and VAN LOON, J. J. A. 2002. An inven-
tory of taste in caterpillars: each species its own key. Acta Zool. Hung. 48:215-263.
SCHOONHOVEN, L. M., VAN LOON, J. J. A. and DICKE, M.,
2005. Insect-Plant Biology. 2nd edn. P. 421. Oxford University Press,
Oxford.

119
SHIELDS, V. D. C. and MITCHELL, B. K. 1995. Responses of max-
illary styloconic receptors to stimulation by sinigrin, sucrose and
inositol in two crucifer-feeding, polyphagous lepidopterous species.
Philos. Trans. R. Soc. London 347:447-457.
SIMMONDS, M. S. J. and BLANEY, W. M. 1983. Some neurophysi-
ological eff ects of azadirachtin on lepidopterous larvae and their feed-
ing responses. Pp. 163-180 in H. SCHMUTTERER and K. R.
S. ASCHER (eds.). Proceedings of the Second International Neem
Conference, G.T.Z., Eschborn.
THIELE, H. U. 1977. Carabid Beetles in Th eir Environment. Zoo-
physiol. Ecol. 10. P 369. Springer, Berlin.
TOFT, S. and BILDE, T. 2002. Carabid diets and food value. Pp. 81-
110 in J. M. Holland (ed.). Th e Agroecology of Carabid Beetles.
Intercept, Andover, UK.
TOOLEY, J. and BRUST, G. 2002. Weed seed predation by carabid
beetles. Pp. 215-229 in J. M. Holland (ed.). Th e Agroecology of Ca-
rabid Beetles. Intercept, Andover, UK.
VAN LOON, J. J. A. 1990. Chemoreception of phenolic acids and fl avo-
noids in larvae of two species of Pieris. J. Comp. Physiol., A 166:889-
899.
WANG, Z., SINGHVI, A., KONG, P. and SCOTT, K. 2004. Taste
representations in the Drosophila brain. Cell 117:981-991.
WHEATER, C. P. 1989. Prey detection by some predatory Coleoptera
(Carabidae and Staphylinidae). J. Zool., A 218:171-185.

120
Legends for fi gures
Figure 1. Spike waveforms and response types of the chemoreceptor
neurons from the antennal chaetoid taste sensilla in P. oblongopunctatus. CN, AN, pHN and SN represent spikes produced by the salt-, alkaloid-,
pH- and sugar-sensitive neuron, respectively. Downward defl ection from
the baseline represents negative polarity. Recordings A–C, D, and E were
obtained from three diff erent sensilla from the same animal, respectively.
F–J, recordings were made from the antennal taste sensilla of diff erent
beetles. B, C, E, F, I, note the long response latency of the AN; fi rst spike
in the spike train of the AN is shown by an arrow. Usually, spike ampli-
tude of the AN was smaller compared to that of CN (see B and C), but in
some sensilla they were larger than CN spikes (demonstrated in D). Note
the pronounced phasic component of the response from CN (see A–J)
and SN (see J), and response with no clear phasic component of the AN
(see B, C, E) in these 0.5 sec fragments of the original 5 sec recordings.
Figure 2. (A) Histogram of relative spike amplitudes from neurons stimu-
lated by sucrose and quinine hydrochloride in antennal taste sensillum
of P. oblongopunctatus. aAN, aCN and aSN represent spike amplitudes
of the alkaloid-, salt- and sugar-sensitive neuron, respectively. Normal
distribution curves of respective samples are indicated. Two successive
stimulations of the same sensillum with 10 mmol l-1 sucrose and 10 mmol
l-1 quinine hydrochloride in the mixture with 10 mmol l-1 NaCl, respec-
tively, were performed. Spikes amplitudes from the two 30 sec record-
ings were measured and analysed. (B,C) Sample 1 sec fragments of the
respective original recordings.
Figure 3. Spike frequencies of the salt- and alkaloid-sensitive neurons in
the antennal chaetoid taste sensilla of P. oblongopunctatus upon stimulation
with 10 mmol l-1 NaCl mixed with diff erent concentrations of quinine
hydrochloride (QHC). CN, AN, pHN, salt-, alkaloid and pH-sensitive
neuron, respectively. N, the number of tested beetles. Responses from
two to three taste sensilla were tested from each test beetle. Vertical bars
denote ± SE of the means.
Figure 4. Responses of the salt- and alkaloid-sensitive neurons from
antennal taste sensilla of P. oblongopunctatus during 5 sec stimulation
with 10 mmol l-1 NaCl mixed with diff erent concentrations of quinine

121
hydrochloride. CN and AN show spike numbers produced by the salt- and
alkaloid-sensitive neuron, respectively; N, the number of tested beetles.
Responses from two to three taste sensilla were tested from each test
beetle. Vertical bars denote ± SE of the means.
Figure 5. Sample responses of an antennal taste sensillum of P. oblon-gopunctatus to diff erent concentrations of quinine hydrochloride. Note
that spike production of the alkaloid-sensitive neuron increased and that
of the salt-sensitive neuron decreased with quinine hydrochloride (QHC)
concentration increase. CN and AN show spikes from the salt- and al-
kaloid-sensitive neurons, respectively.
Figure 6. Deterrent eff ect of quinine hydrochloride (QHC) to the feeding
response of P. oblongopunctatus in two-choice experiments. At each variant
of seed treatment the feeding preference of the beetles between treated
(QHC) and control (distilled water) seeds was tested in fi ve replications
(10 beetles in each).
Table 1. Eff ect of tested plant alkaloids and glucosides to the spike pro-
duction of the chemoreceptor neurons from antennal chaetoid taste sen-
silla of P. oblongopunctatus. All test chemicals were diluted in 10 mmol
l-1 sodium chloride. Due to low solubility of quinine and strychnine in
water these chemicals were tested at a lower concentration than others.
pHN, CN, AN indicate spikes from pH-, salt- and alkaloid-sensitive
neurons, respectively. Asterisks indicate signifi cantly diff erent means,
P<0.05 (paired t-test).

122
A
F
B
I
C
H
D
G
0 0.1 0.2 0.3 0.4 0.5
CN
CN
CN
CN
pHN
pHN
1 mV
CN
CN
CN
CN
CN
CN
AN
AN
AN
SN
quinine hydrochloride dihydrate 1 mmol -1l
caffeine 10 mmol -1l
NaCl 10 mmol -1l
NaCl 10 mmol -1l
NaCl 10 mmol -1l
NaCl 10 mmol -1l
NaCl 10 mmol -1l
NaCl 10 mmol -1l
NaCl 10 mmol -1l
NaCl 10 mmol -1l
NaCl 10 mmol -1l
NaCl 10 mmol -1l
nicotine 10 mmol -1l
strychnine 0.1 mmol -1l
salicin 10 mmol -1l
sinigrin 10 mmol -1l
sucrose 10 mmol -1l
quinine hydrochloride dihydrate 50 mmol -1l
quinine hydrochloride dihydrate 1 mmol -1l
AN
AN
AN
AN
AN
AN
Time (sec)
E
J
Figure 1.
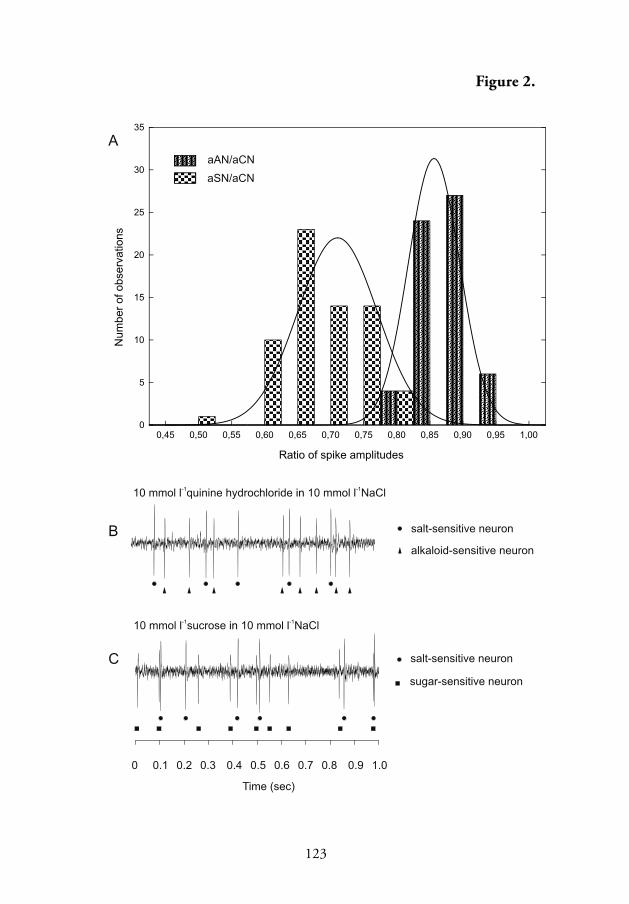
123
0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1.0
Time (sec)
10 mmol quinine hydrochloride 10 mmol NaCl -1 -1l in l
10 mmol sucrose 10 mmol NaCl -1 -1l in l
salt-sensitive neuron
salt-sensitive neuron
alkaloid-sensitive neuron
sugar-sensitive neuron
B
A
C
0,45 0,50 0,55 0,60 0,65 0,70 0,75 0,80 0,85 0,90 0,95 1,000
5
10
15
20
25
30
35N
um
be
r o
f o
bse
rva
tio
ns
Ratio of spike amplitudes
aAN/aCN
aSN/aCN
Figure 2.

124
10
0
0 0.010.001
20
30
40
50
60
70
80
0.1 1 10 50-1
Concentration of QHC (mmol l )
First se
co
nd
re
sp
on
se
(sp
ike
s/s
ec)
ANpHN
CN
N=7
Figure 3.
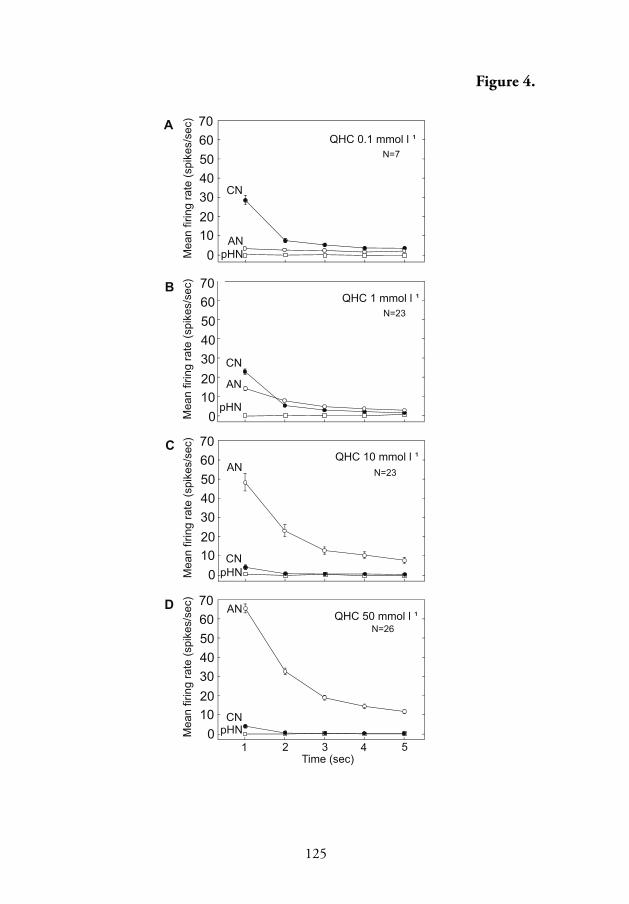
125
0
0
0
0
10
10
10
10
20
20
20
20
30
30
30
30
40
40
40
40
50
50
50
50
60
60
60
60
70
70
70
70
Time (sec)1 2 3 4 5
Me
an
firin
g r
ate
(sp
ike
s/s
ec)
Me
an
firin
g r
ate
(sp
ike
s/s
ec)
Me
an
firin
g r
ate
(sp
ike
s/s
ec)
Me
an
firin
g r
ate
(sp
ike
s/s
ec)
AN
AN
AN
AN
CN
pHN
pHN
pHN
pHN
CN
CN
CN
A
B
C
D
N=7
N=23
N=23
N=26
QHC 1 mmol l ¹
QHC 10 mmol l ¹
QHC 50 mmol l ¹
QHC 0.1 mmol l ¹
Figure 4.
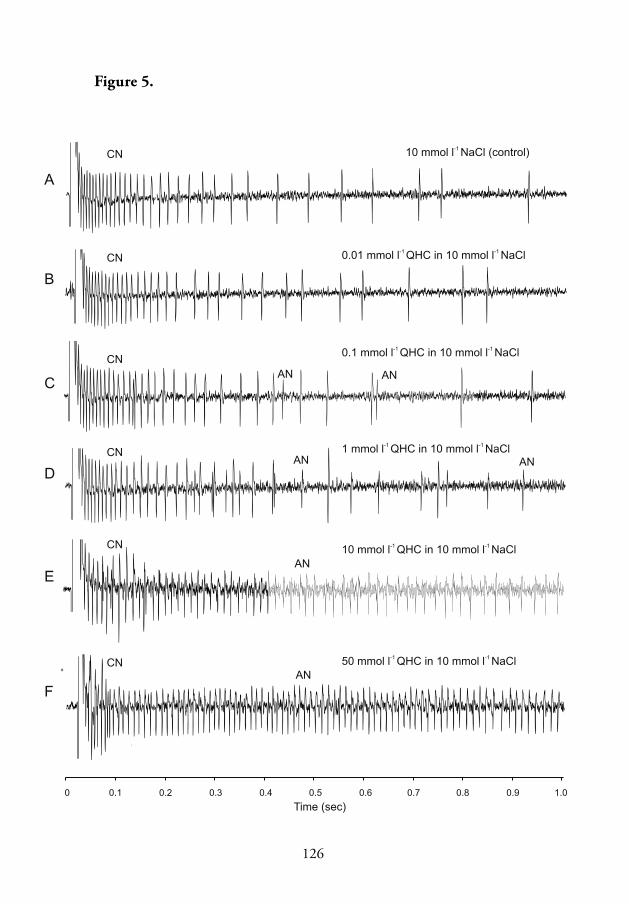
126
A
B
C
D
E
CN
CN
CN
CN
CN
AN
AN
AN
8
F
CNAN
AN
AN
-1 -1 0.01 mmol l QHC in 10 mmol l NaCl
-1 -1 1 mmol l QHC in 10 mmol l NaCl
-1 -1 10 mmol l QHC in 10 mmol l NaCl
-1 -1 50 mmol l QHC in 10 mmol l NaCl
-1 -1 0.1 mmol l QHC in 10 mmol l NaCl
-1 10 mmol l NaCl (control)
0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1.00
Time (sec)
Figure 5.
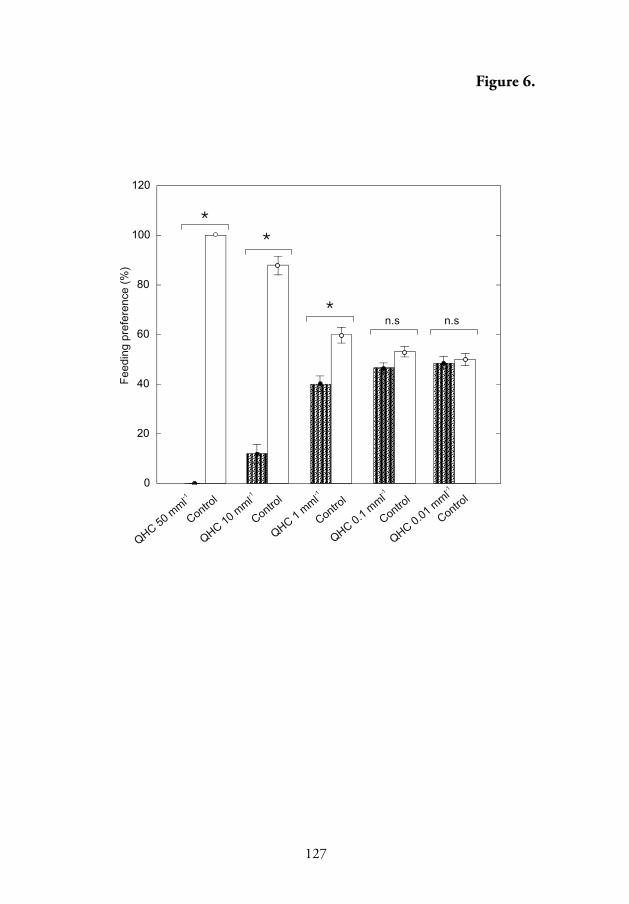
127
0
20
40
60
80
100
120
Fe
ed
ing
pre
fere
nce
(%
)
1-
QHC
1m
0
ml-1
QHC
50 m
ml
-1
HC1
m
Q
m
l-1
0
QHC
.1 m
ml
1-
HC0
Q
.0
1 mm
l
C
l
ontro
C
rl
ont o
Contrlo
Contrlo
C
l
ontro
*
*
*
n.s n.s
Figure 6.
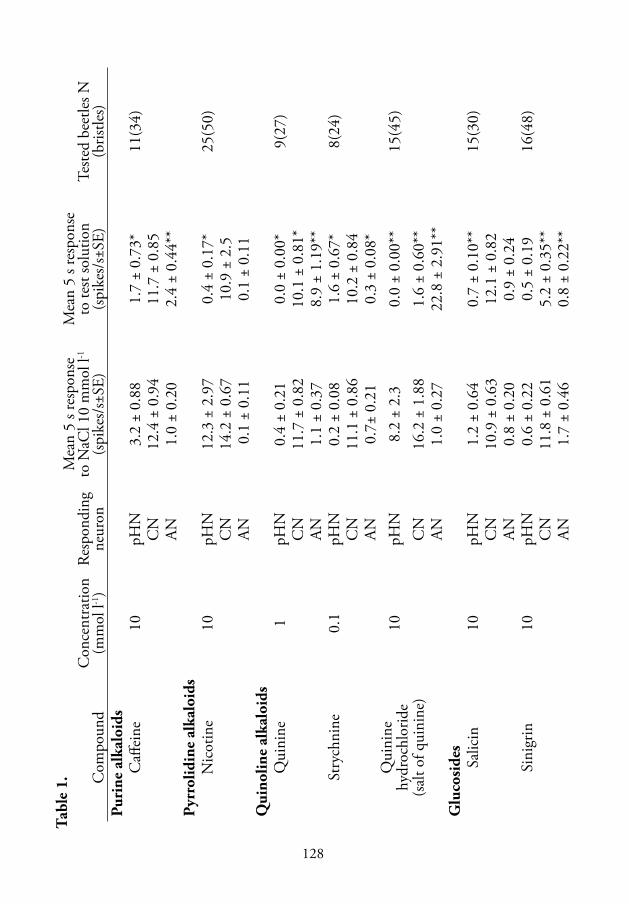
128
Tab
le 1
. Com
pou
nd
Con
cen
trat
ion
(mm
ol l-1
)R
esp
on
din
gn
euro
n
Mea
n 5
s r
esp
on
se
to N
aCl 10 m
mol l-1
(s
pik
es/s
±SE
)
Mea
n 5
s r
esp
on
se
to t
est
solu
tion
(s
pik
es/s
±SE
)T
este
d b
eetl
es N
(bri
stle
s)P
uri
ne
alk
alo
ids
Caff
ein
e10
pH
N3.2
± 0
.88
1.7
± 0
.73*
11(3
4)
CN
12.4
± 0
.94
11.7
± 0
.85
AN
1.0
± 0
.20
2.4
± 0
.44**
Pyr
roli
din
e al
kal
oid
sN
icoti
ne
10
pH
N12.3
± 2
.97
0.4
± 0
.17*
25(5
0)
CN
14.2
± 0
.67
10.9
± 2
.5A
N 0
.1 ±
0.1
10.1
± 0
.11
Qu
ino
lin
e al
kal
oid
sQ
uin
ine
1p
HN
0.4
± 0
.21
0.0
± 0
.00*
9(2
7)
CN
11.7
± 0
.82
10.1
± 0
.81*
AN
1.1
± 0
.37
8.9
± 1
.19**
Stry
chn
ine
0.1
pH
N0.2
± 0
.08
1.6
± 0
.67*
8(2
4)
CN
11.1
± 0
.86
10.2
± 0
.84
AN
0.7
± 0
.21
0.3
± 0
.08*
Qu
inin
e h
ydro
chlo
rid
e10
pH
N8.2
± 2
.30.0
± 0
.00**
15(4
5)
(sal
t of
qu
inin
e)C
N16.2
± 1
.88
1.6
± 0
.60**
AN
1.0
± 0
.27
22.8
± 2
.91**
Glu
cosi
des
Sali
cin
10
pH
N1.2
± 0
.64
0.7
± 0
.10**
15(3
0)
CN
10.9
± 0
.63
12.1
± 0
.82
AN
0.8
± 0
.20
0.9
± 0
.24
Sin
igri
n10
pH
N0.6
± 0
.22
0.5
± 0
.19
16(4
8)
CN
11.8
± 0
.61
5.2
± 0
.35**
AN
1.7
± 0
.46
0.8
± 0
.22**

129
CURRICULUM VITAE
First name: Marit
Last name: Komendant
Date birth: May 10th 1980
Address: Institute of Agricultural and Environmental
Sciences, Estonian University of Life Sciences,
Kreutzwaldi 1, Tartu 51014, Estonia
E-mail: [email protected]
Institution and position held: 2006–... Selteret OÜ offi ce manager
Education:2006–2010 PhD studies in Entomology, Estonian University
of Life Sciences
2004–2006 MSc studies in Plant Protection, Estonian
University of Life Sciences
2000–2004 BSc studies in Horticulture, Estonian Agricultural
University
1998–2000 Räpina Higher Gardening School
1986–1998 Gymnasium of Forseliuse
Academic degree: Master’s Degree (2006), Th esis: Antennal pH
receptors in the ground beetles Pterostichus aethiops and P. oblongopunctatus (Coleoptera,
Carabidae), Estonian University of Life Sciences.
Research interests: Biosciences and Environment, Ecology,
Biosystematics and -physiology (Insect behaviour
and sensor physiology)
Languages spoken:Estonian, English, German, Russian

130
ELULOOKIRJELDUS
Eesnimi: Marit
Perekonnanimi: Milius
Sünniaeg: 10. mai 1980
Aadress: Põllumajandus- ja keskkonnainstituut, Eesti
Maaülikool, Kreutzwaldi 1, Tartu 51014
E-mail: [email protected]
Töökoht ja amet: 2006–... Selteret OÜ büroojuht
Haridus:2006–2010 doktoriõpe entomoloogia erialal, Eesti
Maaülikool
2004–2006 magistriõpe taimekaitse erialal, Eesti Maaülikool
2000–2004 bakalaureuseõpe aianduse erialal, Eesti
Põllumajandusülikool
1998–2000 Räpina Kõrgem Aianduskool
1986–1998 Forseliuse Gümnaasium
Teaduskraad: Teadusmagister 2006, väitekiri: “Antennaalsed
pH-retseptorid süsijooksikutel Pterostichus aethiops ja P. oblongopunctatus (Coleoptera,
Carabidae)”, Eesti Maaülikool
Teadustöö põhisuunad: Bio- ja keskkonnateadused, Ökoloogia,
biosüstemaatika ja -füsioloogia (Putukate
käitumine ja sensoorne füsioloogia)

131
LIST OF PUBLICATIONS
1.1. Publications indexed in the ISI Web of Science database:
Merivee, E., Märtmann, H., Must, A., Milius, M., Williams, I., Mänd,
M., 2008. Electrophysiological responses from neurons of antennal
taste sensilla in the polyphagous predatory ground beetle Pterostichus
oblongopunctatus (Fabricius 1787) to plant sugars and amino acids.
Journal of Insect Physiology 54, 1213–1219.
Merivee, E., Must, A., Milius, M., Luik, A., 2007. Electrophysiological
identifi cation of the sugar cell in antennal taste sensilla of the predatory
ground beetle Pterostichus aethiops. Journal of Insect Physiology 53,
377–384.
Milius, M., Merivee, E., Williams, I., Luik, A., Mänd, M., Must, A.,
2006. A new method for Electrophysiological identifi cation of antennal
pH receptor cells in ground beetles: the example of Pterostichus aethiops (Panzer, 1796) (Coleoptera, Carabidae). Journal of Insect Physiology 52,
960–967.
Merivee, E., Ploomi, A., Milius, M., Luik, A., Heidemaa, M., 2005.
Electrophysiological identifi cation of antennal pH receptors in the
ground beetle Pterostichus oblongopunctatus. Physiological Entomology
30(2), 122–33.
1.2. Papers published in other peer-reviewed international journals with a registered code:
Merivee, E., Must, A., Milius, M., Luik, A., 2006. External stimuli
in searching for favourable habitat, overwintering sites and refugia of
ground beetles: a short review. Agronomy Research 4, 299–302.
3.5. Papers in Estonian and in other peer-reviewed research journals with a local editorial board:
Milius, M., Merivee, E., Mänd, M., Ploomi, A., 2005. Metsa-
süsijooksiku antennaalsete maitseharjaste reaktsioonid 10 ja 100 mM
fosfaatpuhvritele. EPMÜ Teadustööde kogumik 220, 228–230.

ISBN 978-9949-484-00-3
Trükitud taastoodetud paberile looduslike trükivärvidega © Ecoprint
VIIS VIIMAST KAITSMIST
THEA KULL
REPRODUCTION ECOLOGY AND GENETIC DIVERSITY OF
DECLINING SEDGE (CAREX) SPECIES
Prof. Tiiu Kull Dr. Tatjana Oja
November 23, 2011
MARGIT HEINLAAN
ECOTOXICOLOGICAL EVALUATION OF SYNTHETIC NANOPARTICLES AND
PARTICULATE ENVIRONMENTAL SAMPLES.
Prof. Henri-Charles Dubourguier
Prof. Kalev Sepp
Dr. Anne Kahru
December 14, 2011
REIN DRENKHAN
EPIDEMIOLOGICAL INVESTIGATION OF PINE FOLIAGE DISEASES
BY THE USE OF THE NEEDLE TRACE METHOD
Dr. Märt Hanso
January 13, 2011
MIHKEL KIVISTE
CONDITION AND RESIDUAL BEARING CAPACITY OF
EXISTING REINFORCED CONCRETE STRUCTURES
Prof. Jaan Miljan
January 21, 2011
TATJANA KUZNETSOVA
PLANTATIONS OF NATIVE AND INTRODUCED TREE SPECIES IN THE
RECLAMATION OF OIL SHALE POST-MINING AREAS
Dr. Malle Mandre
Dr. Henn Pärn January 25, 2011
Copyright © 2022 FDOKUMEN




















