
Electrophysiological Evidence of Semantic Interference in Visual Search
Upload
independentCategory
view
0download
0
Neuroscience Letters 374 (2005) 212–217
Electrophysiological evidence of enhanced distractibilityin ADHD children
V. Gumenyuka,∗, O. Korzyukova,b, C. Escerac, M. Hamalainend, M. Huotilainena,e,f ,T. Hayrinend, H. Oksanend, R. Naatanena,e, L. von Wendtd, K. Alhog
a Cognitive Brain Research Unit, Department of Psychology, University of Helsinki, Helsinki, Finlandb Department of Psychiatry, Yale University and VA-Connecticut Healthcare System, West Haven, CT, USA
c Cognitive Neuroscience Research Group, Department of Psychiatry and Clinical Psychology, Faculty of Psychology,University of Barcelona, Catalonia, Spain
d Department of Child Neurology, Hospital for Children and Adolescents, Helsinki University Central Hospital, Helsinki, Finlande Helsinki Brain Research Centre, Helsinki, Finland
f Collegium of Advanced Studies, University of Helsinki, Finlandg Department of Psychology, University of Helsinki, Helsinki, Finland
Received 28 June 2004; received in revised form 14 October 2004; accepted 21 October 2004
A
childrenw ted brainp visual taskw esponsesf frontal LateN recordings ative ERPc ietal scalpa e ADHDg involuntarya©
K
Adis
tap
taskvent-
im-eltyalp,imal
muli.orient-onon
0d
bstract
Abnormal involuntary attention leading to enhanced distractibility may account for different behavioral and cognitive problems inith attention deficit hyperactivity disorder (ADHD). This was investigated in the present experiment by recording event-relaotentials (ERPs) to distracting novel sounds during performance of a visual discrimination task. The overall performance in theas less accurate in the ADHD children than in the control children, and the ADHD children had a higher number of omitted r
ollowing novel sounds. In both groups, the distracting novel sounds elicited a biphasic P3a ERP component and a subsequentegativity (LN). The early phase of P3a (180–240 ms) had significantly smaller amplitudes over the fronto-central left-hemisphereites in the ADHD children than in the control group presumably due to an overlapping enhanced left-hemisphere dominant negomponent elicited in the ADHD group. Moreover, the late phase of P3a (300–350 ms) was significantly larger over the left parreas in the ADHD children than in the controls. The LN had a smaller amplitude and shorter latency over the frontal scalp in throup than in the controls. In conclusion, the ERP and behavioral effects caused by the novel sounds reveal deficient control ofttention in ADHD children that may underlie their abnormal distractibility.2004 Elsevier Ireland Ltd. All rights reserved.
eywords: Attention deficit hyperactivity disorder; Distraction; Children; Orienting; P3a; Reorienting negativity (LN/RON)
ttention deficit hyperactivity disorder (ADHD) is a multi-imensional disorder that has its onset in childhood and that
s characterized by persistent problems of inattention, impul-ivity, and hyperactivity[2,15].
In ADHD patients, difficulties with sustained attentionasks[11] can be caused by different alternatives, one beingbnormal distractibility. Therefore, the present study com-ared involuntary attention and distractibility in ADHD and
∗ Corresponding author. Tel.: +358 9 191 29460; fax: +358 9 191 29450.E-mail address:[email protected] (V. Gumenyuk).
healthy children, as indicated by distraction of visualperformance by task-irrelevant novel sounds and by erelated brain potentials (ERPs) to these sounds.
Highly deviant, task-irrelevant auditory and visual stuli elicit a P3 component of ERP called the P3a or novP3[5,21]. The P3a is maximal over the fronto-central scand therefore, it can be separated from the parietally maxP300 or P3b response to auditory and visual target stiThe P3a has been suggested to be associated with theing response[5,6,13,20]and with detection and evaluatiof novelty[7]. Association of P3a with involuntary attenti
304-3940/$ – see front matter © 2004 Elsevier Ireland Ltd. All rights reserved.oi:10.1016/j.neulet.2004.10.081

V. Gumenyuk et al. / Neuroscience Letters 374 (2005) 212–217 213
switching is evidenced by the fact that P3a-eliciting novelsounds also distract visual task performance in adults[6] andchildren[9].
The P3a to novel sounds has two phases: an early phase(eP3a) peaking at 200–250 ms and a late phase (lP3a) peakingat 300–350 ms from sound onset[6]. In adults, the eP3a has itsmaximum amplitude over the central scalp, whereas the lP3ais distributed widely and has its maximum over the frontalscalp. According to scalp current density analysis[25], mag-netoencephalography[1], and studies in patients with localbrain lesions[14], the eP3a is generated in the auditory andtemporo-parietal cortices, while the lP3a gets a major con-tribution from the prefrontal cortex[13,25]. Thus, multiplebrain areas are involved in involuntary attention and auditorynovelty detection. This appears to be true also in children.The scalp distributions of the eP3a and lP3a to novel soundsin children aged over 11 years are similar to those in adults[9]. However, in 7–10-year-olds, the lP3a has its maximumover the centro-parietal scalp suggesting that the auditory at-tention networks are not yet fully developed at this age[9].
The P3a to distracting sounds is followed by a reorientingnegativity (RON) component in adult ERPs[19] and the LateNegativity (LN) in children[9]. RON and LN have maximumamplitudes over fronto-central scalp areas at 400–700 ms af-ter onset of a distracting sound and may be associated with thep tionb
HDa muli[ erP novels ught rast,K ns( isonw rgett tudy,w s tor andh g vi-s
lthyc duet isuald oupa eana oup).
ento s-p g-nf oseA wnf
asa l-
dren in the ADHD group had their verbal, performance, andfull-scale intelligence quotients (IQ) within the normal range90–126 (means: 108, 100, and 104, respectively). The ADHDchildren were rewarded with a ticket to a self-chosen movie(according to the protocol).
The control children were recruited from ordinary primaryschools (none of the control children were in gifted programsat school) and were rewarded for their participation with 6D .None of the control children had any diagnosed psycholog-ical or neurological problems. All of them were in the age-appropriate grade at school. All children had a normal orcorrected-to-normal vision and normal hearing.
Written informed consents were obtained from all parentsof children participating in the study. The Ethics Committeeof the Hospital for Children and Adolescents approved theprotocol of the present study for the ADHD children andThe Ethical Committee of the Department of Psychology,University of Helsinki, approved the protocol of the presentstudy for the control group.
The children were presented with auditory-visual stimuluspairs at a constant rate of one pair in every 1.7 s. Each soundhad a duration of 200 ms and was delivered binaurally throughheadphones at 50 dB SPL. The sound was either a sinusoidaltone of 600 Hz (P= 0.80; or 560 tones) or one of the complexenvironmental “novel” sounds presented randomly (P= 0.20)w n anyt rawnf h ast novels
gs( tallya 0 msa wered uters eyes.
ocust -o ond asa witht reena im-a in-d d tot
plingr F3,F T5,T sitesa ec-t linksw Fp1,F rightc he tipo s andn s pe-
refrontal brain mechanisms involved in reorienting attenack to task performance after a distracting event[9,19].
Previous studies found no differences between ADnd control children in P3a responses to novel visual sti
10,11]. However, Kemner et al.’s[11] data suggest small3a responses over the parietal scalp to task-irrelevantounds in ADHD children than in healthy controls, althohis effect did not reach statistical significance. In contilpelainen et al.[12] found enhanced frontal P3 deflectio
presumably the P3a) in distractible children in comparith non-distractible children in response to infrequent ta
ones occurring among frequent tones. In the present se investigated these issues further by recording ERP
epeating tones and novel sounds from ADHD childrenealthy controls while they concentrated on a demandinual task.
Eleven 8–10-year-old ADHD and 10 age-matched heahildren were studied. One ADHD child was excludedo a very poor performance (less than 50% hits) in the viscrimination task (see below). The remaining ADHD grnd the control group consisted of 10 subjects each (mges 8.7 and 8.6 years., respectively; one girl in each gr
The ADHD children were recruited from the Departmf Child Neurology of the Helsinki University Central Hoital. All of them had a thorough multidisciplinary diaostic assessment and all fulfilled the DSM-IV[2] criteria
or ADHD and had no other comorbidity disorders. ThDHD children who were on medication were withdra
rom it at least a day before the experiment.The general intellectual ability of the clinical group w
ssessed with Wechsler’s[22,23]intelligence scales. All chi
ith the exception that at least 3 tones occurred betweewo successive novel sounds. The novel sounds were drom a pool of 140 different environmental sounds, suchose produced by a hammer, rain, and car horn. Eachound was presented only once during the experiment.
The visual stimuli were 32 different white-line drawin16 animals and 16 non-animals) extending 9 cm horizonnd vertically, presented for 300 ms, and starting at 30fter the onset of the preceding sound. The drawingsisplayed in a random order in the center of a black compcreen located approximately 130 cm from the subject’s
The children were instructed to ignore the sounds, to fheir gaze on a white fixation cross (1 cm× 1 cm) continuusly presented at the center of the screen, and to respccurately as possible by pressing one response button
heir right thumb to an animal image appearing on the scnd another button with their left thumb to a non-animalge. The children’s motivation to perform the task wasicated by their genuine interest to their hit rate reporte
hem after each block.The electroencephalogram (EEG; 0.1–100 Hz, sam
ate 250 Hz) was recorded from frontal (Fp1, Fp2, F7,z, F4, and F8), central (C3, Cz, C4), temporal (T3, T4,6), parietal (P3, Pz, P4), and occipital (O1, O2) scalpnd from the left and right mastoids (LM and RM, resp
ively). Voltage changes caused by eye movements and bere monitored with recordings from the forehead sites (p2) and from additional electrodes placed at the left andanthi. The common reference electrode was placed at tf the nose. ERPs were obtained separately for the toneovel sounds by averaging EEG epochs over a 900 m

214 V. Gumenyuk et al. / Neuroscience Letters 374 (2005) 212–217
riod starting 100 ms before each sound onset. These EEGepochs were digitally band-pass filtered at 1–30 Hz. Epochswith extracerebral artifacts exceeding±100�V at any elec-trode were excluded from averaging. Also, the epochs for thefirst four stimuli of each block and the epochs for any toneoccurring right after a novel sound were excluded. For eachchild, averaged ERPs to tones and novel sounds consisted ofat least 380 and 90 acceptable EEG epochs, respectively.
Because the children were instructed to perform the visualdiscrimination task with an “accuracy instruction” (stress onperformance accuracy, not on speed, cf.[17]), changes in therates of correct responses (“hits”) and incorrect responses(wrong button presses and response omissions) caused by thetask-irrelevant novel sounds were used as major behavioralindexes distractibility.
A correct button press given 200–1500 ms after visualstimulus onset was classified as a hit. Within-group differenceof means rates of hit, wrong and missed responses, and hit re-action times (RTs) were analyzed witht-tests, while between-group difference of those were analyzed with Mann–WhitneyU-test (depended variables: tone versus novel).
The P3a and LN amplitudes were measured from ERPdifference waves obtained by subtracting ERPs to tones fromthose to novel sounds[6,9]. Their amplitudes were measuredas mean voltages over fixed latency windows determineda encew udeso and3 n tot 0 msp ea-s ms.B desw osth for theP P3a,i ver-s odes)a lineF es).
rongr or theA tione gnifi-c mbero om-p theAP aftern gnifi-c
ig-n ac-c one( vels l-
Table 1Mean (standard deviations in parentheses) RTs, hit rates, wrong-responseand miss rates of the ADHD and control children in performing the visualdiscrimination task after an occurrence of a task-irrelevant tone vs. novelsound
Performance Stimulus ADHD mean(S.D.)
Controlsmean (S.D.)
RT (ms) Tone 476 (78) 498 (122)Novel 488 (86) 553 (142)
Hit rate (%) Tone 65 (13) 81 (8)Novel 62 (12) 78 (10)
Wrong-response (%) Tone 13 (5) 6 (3)Novel 12 (6) 9 (3)
Miss-response (%) Tone 22 (9) 12 (10)Novel 26 (9) 13 (10)
dren had an overall smaller percentage of correct responsesthan the controls.
A Mann–WhitneyU-test for the rate of missed responsesindicated that an occurrence of a novel sound (in comparisonwith an occurrence of a tone) enhanced significantly more thenumber of response omissions in the ADHD children than inthe controls (U(17) = 2.49;P< 0.01), this effect of the pre-ceding sound on the number of omissions being 4% in theADHD group but only 1% in the control group (seeTable 1).No significant group difference or effect of preceding stimu-lus was found for the number of wrong responses.
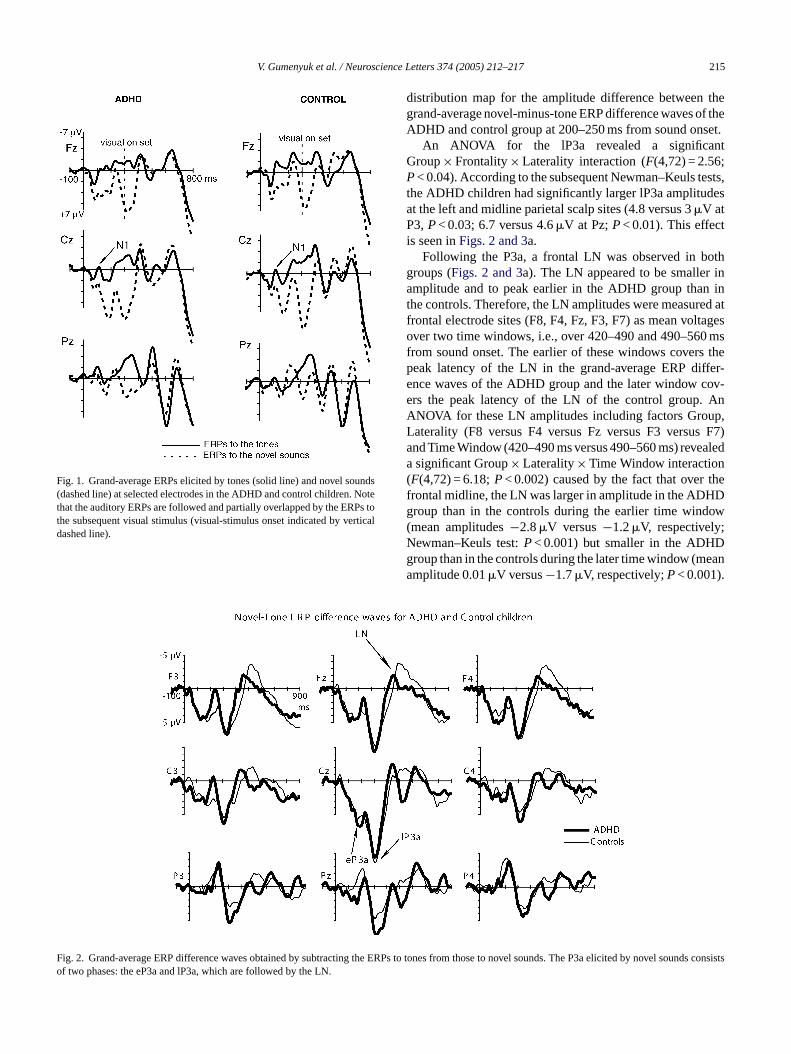
Fig. 1 displays grand-average ERPs elicited by theauditory-visual stimulus pairs. In the ADHD and controlgroups, tones elicited only a small auditory N1 component(peak around 100 ms from sound onset at the Cz electrode),presumably due to a relative immaturity of the auditory cor-tex[18]. A P3a response was elicited by novel sounds in eachage group (Fig. 1).
Fig. 2shows the grand-average ERP difference waves ofthe P3a and LN responses to the novel sounds for the ADHDand control groups. Consistent with our previous studies inhealthy children[9], the P3a had a biphasic structure. As seenin Fig. 2, the eP3a was largest over the fronto-central scalp inboth groups, whereas the lP3a had a wider distribution overthe midline scalp areas.
Cz,C upFS due toa nto-ct didt s: 0.9vs( y ano s inA r thel hisp
fter visual inspection of the grand-average ERP differaves at Fz and Cz electrodes. The mean amplitf the eP3a and lP3a were measured over 180–25000–350 ms from sound onset, respectively, in relatio
he mean difference-wave amplitude during the 10restimulus baseline. The LN amplitudes, in turn, were mured over two latency windows: 420–490 and 490–560etween-group differences in the P3a and LN amplituere examined with ANOVAs and Newman–Keuls poc tests. Greenhouse–Geisser corrections were used-values when appropriate. ANOVAs for the eP3a and l
ncluded electrode factors Frontality (frontal F3, Fz, F4us central C3, Cz, C4 versus parietal P3, Pz, P4 electrnd Laterality (left-hemisphere F3, C3, P3 versus midz, Cz, Pz versus right-hemisphere F4, C4, P4 electrod
Table 1shows mean RTs, and the mean rates of hits, wesponses, and response omissions in the visual task fDHD and control groups. In both groups, the distracffect in the visual task caused by novel sounds was siant: An occurrence of a novel sound decreased the nuf correct responses to the following visual stimulus, in carison with visual stimuli preceded by a tone both inDHD and control groups (t(9) = 2.54;P< 0.03;t(9) = 2.96;< 0.01, respectively). Both groups showed longer RTsovel sounds than after tones, but this effect reached siance only in the controls (t(9) =−3.79;P< 0.004).
A Mann–WhitneyU-test for the hit rates yielded a sificant between-group differences in the visual-taskuracy both for the visual stimuli preceded by a tU(11) =−2.9;P< 0.002) and for those preceded by a noound (U(14) =−2.7;P< 0.005), indicating that ADHD chi
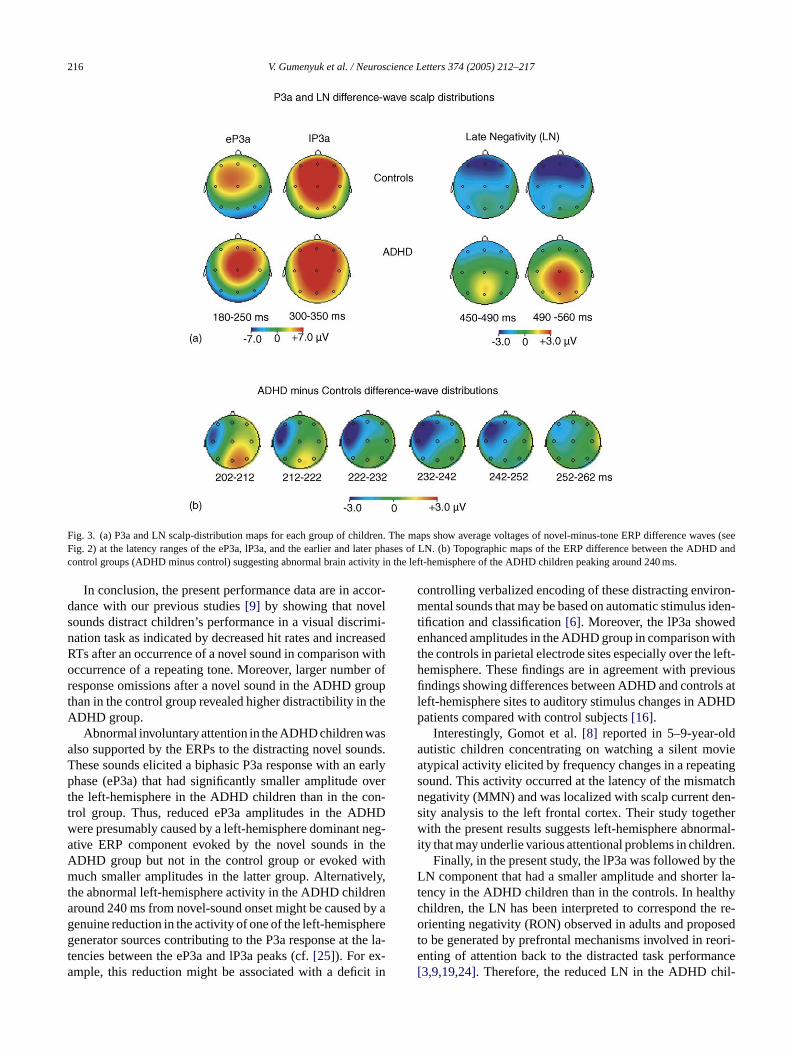
An ANOVA for the eP3a amplitudes at F3, FZ, F4, C3,4, P3, Pz, and P4 electrodes indicated a significant Gro×rontality× Laterality interaction (F(4,72) = 4.64;P< 0.01).ubsequent Newman–Keuls tests showed that this wasn amplitude difference between the groups over the froentral left-hemisphere for the eP3a: as seen inFigs. 2 and 3a,he ADHD children had smaller eP3a amplitudes thanhe controls (mean amplitude measured at 180–250 mersus 3.35�V at F3, and−0.9 versus 3.27�V at C3, re-pectively;P< 0.04 for both comparisons). As seen inFig. 2e.g., the F3 electrode), this effect might be caused bverlapping negative component peaking around 240 mDHD children and reducing their eP3a amplitudes ove
eft frontal scalp. The left-frontal scalp distribution of tossible negative component is seen inFig. 3b showing a
V. Gumenyuk et al. / Neuroscience Letters 374 (2005) 212–217 215
Fig. 1. Grand-average ERPs elicited by tones (solid line) and novel sounds(dashed line) at selected electrodes in the ADHD and control children. Notethat the auditory ERPs are followed and partially overlapped by the ERPs tothe subsequent visual stimulus (visual-stimulus onset indicated by verticaldashed line).
Fig. 2. Grand-average ERP difference waves obtained by subtracting the ER unds consiof two phases: the eP3a and lP3a, which are followed by the LN.
distribution map for the amplitude difference between thegrand-average novel-minus-tone ERP difference waves of theADHD and control group at 200–250 ms from sound onset.
An ANOVA for the lP3a revealed a significantGroup× Frontality× Laterality interaction (F(4,72) = 2.56;P< 0.04). According to the subsequent Newman–Keuls tests,the ADHD children had significantly larger lP3a amplitudesat the left and midline parietal scalp sites (4.8 versus 3�V atP3,P< 0.03; 6.7 versus 4.6�V at Pz;P< 0.01). This effectis seen inFigs. 2 and 3a.
Following the P3a, a frontal LN was observed in bothgroups (Figs. 2 and 3a). The LN appeared to be smaller inamplitude and to peak earlier in the ADHD group than inthe controls. Therefore, the LN amplitudes were measured atfrontal electrode sites (F8, F4, Fz, F3, F7) as mean voltagesover two time windows, i.e., over 420–490 and 490–560 msfrom sound onset. The earlier of these windows covers thepeak latency of the LN in the grand-average ERP differ-ence waves of the ADHD group and the later window cov-ers the peak latency of the LN of the control group. AnANOVA for these LN amplitudes including factors Group,Laterality (F8 versus F4 versus Fz versus F3 versus F7)and Time Window (420–490 ms versus 490–560 ms) revealeda significant Group× Laterality× Time Window interaction(F(4,72) = 6.18;P< 0.002) caused by the fact that over thef Dg ow( ;N Dg eana
Ps to tones from those to novel sounds. The P3a elicited by novel sosts
rontal midline, the LN was larger in amplitude in the ADHroup than in the controls during the earlier time windmean amplitudes−2.8�V versus−1.2�V, respectivelyewman–Keuls test:P< 0.001) but smaller in the ADHroup than in the controls during the later time window (mmplitude 0.01�V versus−1.7�V, respectively;P< 0.001).
216 V. Gumenyuk et al. / Neuroscience Letters 374 (2005) 212–217
Fig. 3. (a) P3a and LN scalp-distribution maps for each group of children. The maps show average voltages of novel-minus-tone ERP difference waves (seeFig. 2) at the latency ranges of the eP3a, lP3a, and the earlier and later phases of LN. (b) Topographic maps of the ERP difference between the ADHD andcontrol groups (ADHD minus control) suggesting abnormal brain activity in the left-hemisphere of the ADHD children peaking around 240 ms.
In conclusion, the present performance data are in accor-dance with our previous studies[9] by showing that novelsounds distract children’s performance in a visual discrimi-nation task as indicated by decreased hit rates and increasedRTs after an occurrence of a novel sound in comparison withoccurrence of a repeating tone. Moreover, larger number ofresponse omissions after a novel sound in the ADHD groupthan in the control group revealed higher distractibility in theADHD group.
Abnormal involuntary attention in the ADHD children wasalso supported by the ERPs to the distracting novel sounds.These sounds elicited a biphasic P3a response with an earlyphase (eP3a) that had significantly smaller amplitude overthe left-hemisphere in the ADHD children than in the con-trol group. Thus, reduced eP3a amplitudes in the ADHDwere presumably caused by a left-hemisphere dominant neg-ative ERP component evoked by the novel sounds in theADHD group but not in the control group or evoked withmuch smaller amplitudes in the latter group. Alternatively,the abnormal left-hemisphere activity in the ADHD childrenaround 240 ms from novel-sound onset might be caused by agenuine reduction in the activity of one of the left-hemispheregenerator sources contributing to the P3a response at the la-tencies between the eP3a and lP3a peaks (cf.[25]). For ex-ample, this reduction might be associated with a deficit in
controlling verbalized encoding of these distracting environ-mental sounds that may be based on automatic stimulus iden-tification and classification[6]. Moreover, the lP3a showedenhanced amplitudes in the ADHD group in comparison withthe controls in parietal electrode sites especially over the left-hemisphere. These findings are in agreement with previousfindings showing differences between ADHD and controls atleft-hemisphere sites to auditory stimulus changes in ADHDpatients compared with control subjects[16].
Interestingly, Gomot et al.[8] reported in 5–9-year-oldautistic children concentrating on watching a silent movieatypical activity elicited by frequency changes in a repeatingsound. This activity occurred at the latency of the mismatchnegativity (MMN) and was localized with scalp current den-sity analysis to the left frontal cortex. Their study togetherwith the present results suggests left-hemisphere abnormal-ity that may underlie various attentional problems in children.
Finally, in the present study, the lP3a was followed by theLN component that had a smaller amplitude and shorter la-tency in the ADHD children than in the controls. In healthychildren, the LN has been interpreted to correspond the re-orienting negativity (RON) observed in adults and proposedto be generated by prefrontal mechanisms involved in reori-enting of attention back to the distracted task performance[3,9,19,24]. Therefore, the reduced LN in the ADHD chil-

V. Gumenyuk et al. / Neuroscience Letters 374 (2005) 212–217 217
dren might be associated with a prefrontal cortical dysfunc-tion [4] leading to problems in reorienting attention back tothe distracted task performance. This problem in reorient-ing attention is indicated in the present study by enhancednumber of omitted responses in the visual task after a novelsound. The decrement in the LN peak latency observed inthe ADHD children in relation to the controls might, in turn,be associated with a higher degree of impulsivity, which is asymptom of ADHD[2,15].
In summary, the present study suggests that ERPs recordedto the task-irrelevant novel stimuli distracting the children’sperformance provide us with evidence supporting the ob-servations of attentional and behavioral deficits in ADHD.Although, the present study should be considered as prelim-inary, given the relatively small size of the two groups, thepresent results suggest that a multimethodological approachcombining neuropsychological, behavioral and electrophysi-ological measures of distractibility and involuntary orientingof attention improve our understanding of the brain–behaviorrelationship in ADHD.
Acknowledgements
The authors thank Ms. Kirsi Jarvinen for her helpi naW thee rtedbO andg ted byt ncea
R
.P.ovelortex:1998)
tis-tric
tions. 10
M.A.func-and
taskysiol.
ntials,
[7] D. Friedman, Y.M. Cycowicz, H. Gaeta, The novelty P3: an event-related potential (ERP) sign of the brain’s evaluation of novelty,Neurosci. Biobehav. Rev. 25 (2001) 355–373.
[8] M. Gomot, M.H. Giard, J.L. Adrien, C. Barthelemy, N. Bruneau,Hypersensitivity to acoustic change in children with autism: elec-trophysiological evidence of left frontal cortex dysfunction, Psy-chophysiology 39 (2002) 577–584.
[9] V. Gumenyuk, O. Korzyukov, K. Alho, C. Escera, R. Naatanen,Effects of auditory distraction on electrophysiological brain activityand performance in children aged 8–13 years, Psychophysiology 41(2004) 30–36.
[10] L.M. Jonkman, C. Kemner, H.S. Koelega, G. Camfferman, R.J. vdGaag, J.K. Buitelaar, H. van Engeland, M.N. Verbaten, Event-relatedpotentials and performance of attention-deficit hyperactivity disorder:children and normal controls in auditory and visual selective attentiontasks, Biol. Psychiatry 41 (1997) 595–611.
[11] C. Kemner, M.N. Verbaten, H.S. Koelega, J.K. Buitelaar, R.J.van der Gaag, G. Camfferman, H. van Engeland, Event-relatedbrain potentials in children with attention-deficit and hyperactiv-ity disorder: effects of stimulus deviancy and task relevance inthe visual and auditory modality, Biol. Psychiatry 40 (1996) 522–534.
[12] R. Kilpelainen, L. Luoma, E. Herrgard, H. Ypparila, J. Partanen, J.Karhu, Persistent frontal P300 brain potential suggests abnormal pro-cessing of auditory information in distractible children, NeuroReport10 (1999) 3405–3410.
[13] R.T. Knight, Decreased response to novel stimuli after prefrontallesions in man, Clin. Neurophysiol. 59 (1984) 9–20.
[14] R.T. Knight, D. Scabini, D.L. Woods, C.C. Clayworth, Contributionsof temporal–parietal junction to the human auditory P3, Brain Res.
[ VshipChild
[ chil-sio-hav.
[ pell,Clin.
[ tionulti-20–
[ di-1998)
[ giates,
[ ng-uli,
[ sed,
[ ed.,
[ re-rt 15
[ osa,lated
n recruiting the control children, as well as Ms. Leeallendahr and Mr. Timo Saarinen for assistance with
lectrophysiological recording. V. Gumenyuk was suppoy the Academy of Finland grant no. 55606. R. Naatanen and. Korzyukov were supported by the Academy of Finlrants nos. 77322 and 102316. C. Escera was suppor
he grant no. PM99-0167 of the Spanish Ministry of Sciend Technology.
eferences
[1] K. Alho, I. Winkler, C. Escera, M. Huotilainen, J. Virtanen, IJaaskelainen, E. Pekkonen, R.J. Ilmoniemi, Processing of nsounds and frequency changes in the human auditory cmagnetoencephalographic recordings, Psychophysiology 35 (211–224.
[2] American Psychiatric Association, DSM-IV, Diagnostic and Statical Manual of Mental Disorders, fourth ed., American PsychiaAssociation, Washington, DC, 1994.
[3] S. Berti, E. Schroger, A comparison of auditory and visual distraceffects: behavioral and event-related indices, Cogn. Brain Re(2001) 265–273.
[4] G. Bush, J.A. Frazier, S.L. Rauch, L.J. Seidman, P.J. Whalen,Jenike, B.R. Rosen, J. Biederman, Anterior cingulate cortex dystion in attention-deficit/hyperactivity disorder revealed by fMRIthe Counting Stroop, Biol. Psychiatry 45 (1999) 1542–1552.
[5] E. Courchesne, S.A. Hillyard, R. Galambos, Stimulus novelty,relevance and the visual evoked potential in man, Clin. Neuroph39 (1975) 131–143.
[6] C. Escera, K. Alho, E. Schroger, I. Winkler, Involuntary attentioand distractibility as evaluated with Event-related brain potenAudiol. Neurootol. 5 (2000) 151–166.
502 (1989) 109–116.15] A.E. Morgan, G.W. Hynd, C.A. Riccio, J. Hall, Validity of DSM-I
ADHD predominantly inattentive and combined types: relationto previous DSM diagnoses/subtype differences, J. Am. Acad.Adolesc. Psychiatry 35 (1996) 325–333.
16] R.D. Oades, Frontal, temporal and lateralized brain function indren with attention-deficit hyperactivity disorder: a psychophylogical and neuropsychological viewpoint on development, BeBrain Res. 94 (1998) 83–95.
17] A. Pfefferbaum, J. Ford, R. Johnson Jr., B. Wenegrat, B.S. KoManipulation of P3 latency: speed vs. accuracy instructions,Neurophysiol. 55 (1983) 188–197.
18] C.W. Ponton, J.J. Eggermont, B. Kwong, M. Don, Maturaof human central auditory system activity: evidence from mchannel evoked potentials, Clin. Neurophysiol. 111 (2000) 2236.
19] E. Schroger, C. Wolff, Attentional orienting and reorienting is incated by human event-related brain potentials, NeuroReport 9 (3355–3358.
20] E.N. Sokolov, J.A. Sprinks, R. Naatanen, H. Lyytinen, The OrientinResponse in Information Processing, Lawrence Erlbaum AssocLondon, NJ, 2002.
21] N.K. Squires, K.C. Squires, S.A. Hillyard, Two varieties of lolatency positive waves evoked by unpredictable auditory stimClin. Neurophysiol. 38 (1975) 387–401.
22] D. Wechsler, Wechsler Intelligence Scale for Children, reviPsykologien kustannus, Helsinki, 1984 (in Finnish).
23] D. Wechsler, Wechsler Intelligence Scale for Children, thirdPsykologien kustannus, Helsinki, 1999 (in Finnish).
24] N. Wetzel, S. Berti, A. Widmann, E. Schroger, Distraction andorientation in children: a behavioral and ERP study, NeuroRepo(2004) 1355–1358.
25] E. Yago, C. Escera, K. Alho, M.-H. Giard, J.M. Serra-GrabulSpatiotemporal dynamics of the auditory novelty-P3 event-rebrain potential, Cogn. Brain Res. 16 (2003) 383–390.
Copyright © 2022 FDOKUMEN




















