
Caterpillar Labial Saliva Alters Tomato Plant Gene Expression
Upload
independentCategory
view
2download
0
Dev Genes Evol (2006) 216: 431–442DOI 10.1007/s00427-006-0086-3
ORIGINAL ARTICLE
Laurent Joulia . Jean Deutsch . Henri-Marc Bourbon .David L. Cribbs
The specification of a highly derived arthropod appendage,
the Drosophila labial palps, requires the joint action of selectors
and signaling pathways
Received: 5 October 2005 / Accepted: 1 May 2006 / Published online: 14 June 2006# Springer-Verlag 2006
Abstract The remarkable diversity of form in arthropodsreflects flexible genetic programs deploying many con-served genes. In the insect modelDrosophila melanogaster,diversity of form can be observed between serially homol-ogous appendages or when a single appendage is trans-formed by homeotic mutations, such as the adult labialmouthparts that can present alternative antennal, protho-racic, or maxillary identities. We have examined the roles ofthe Hox selector genes proboscipedia (pb) and Sex combsreduced (Scr), and the antennal selectors homothorax (hth)and spineless (ss) in labial specification, by tissue-directedmitotic recombination. Whereas loss of pb function trans-forms labium to prothoracic leg, loss of Scr, hth, or ssfunctions results in little or no change in labial specifica-tion. Results of analysis of single and multiple mutantcombinations support a genetic hierarchy in which thehomeotic pb gene possesses a primary role. It is surprisingto note that while loss of ss activity alone had no detectedeffect, all mutant combinations lacking both pb and ssyielded the most severe phenotype observed: stunted,apparently tripartite legs that may correspond to a defaultstate. The roles of the four selector genes are functionallylinked to a cell nonautonomous mechanism involving thecoupled activities of the decapentaplegic (dpp)/TGF-β andwingless (wg)/Wnt signaling pathways. Accordingly,several mutant combinations impaired in dpp signaling
were seen to reorient labial-to-leg transformations towardantennal aristae. A crucial aspect of selector function indevelopment and evolution may be in regulating diffusiblesignals, including those emitted by dpp and wg.
Keywords Selectors . Morphogens . Ground state .Proboscipedia . Spineless
Introduction
The arthropod body plan is composed of a series ofrepeated segments that are units for development. In theinsect model Drosophila, segmental imaginal discs inlarvae give rise to serially homologous pairs of append-ages, including antennas, legs, genitalia, and mouthparts(Cohen 1993). The Engrailed (En) transcription factor inposterior cells activates the expression of hedgehog (hh)(DiNardo et al. 1985; Lee et al. 1992; Tabata et al. 1992).Short-range diffusion of Hh protein induces localizedexpression of the Wnt and TGF-β growth factors encodedby wingless (wg) and decapentaplegic (dpp) in adjacentanterior cells (Basler and Struhl 1994; Heemskerk andDiNardo 1994; Tabata and Kornberg 1994; Zecca et al.1995). Wg and Dpp gradients in turn regulate proximo-distal (PD) positional information through the expressionof the PD genes Distal-less (Dll) distally, dachshund (dac)medially, and homothorax (hth) proximally (Abu-Shaarand Mann 1998; Casares and Mann 2001; Lecuit andCohen 1997; Yao et al. 1999). Controlled expression of thePD appendage genes Dll and hth also contributes tospecifying identity, and the Hth/Dll couple is viewed as anantennal selector that acts by regulatingmultiple target genes(Abu-Shaar and Mann 1998; Casares andMann 1998; Donget al. 2000). These target genes include spineless (ss), whosemisexpression can induce ectopic antennal development(Duncan et al. 1998), and the pipsqueak-motif genes distalantenna (dan) and dan-related (Dong et al. 2002; Emerald etal. 2003; Suzanne et al. 2003).
Another class of selectors comprises the eight homeoticHox genes that confer segmental identity along the antero-
Communicated by guest editors Jean Deutsch and Gerhard Scholtz
L. Joulia . H.-M. Bourbon . D. L. Cribbs (*)Centre de Biologie du Développement, CNRS,118 route de Narbonne,31062 Toulouse, Cedex 04, Francee-mail: [email protected].: +33-561-558287Fax: +33-561-556507e-mail: [email protected]
J. DeutschEquipe Développement et Evolution, UMR 7622,Biologie du Développement,CNRS and Université Pierre et Marie Curie,9 quai St-Bernard, case 24,75252 Paris, Cedex 05, France

posterior (AP) body axis. Hox genes code for home-odomain transcription factors thought to regulate panels ofdownstream genes that in turn distinguish segmentmorphologies (Kaufman et al. 1990; Casares and Mann2001; McGinnis and Krumlauf 1992). Drosophila anten-nas, legs, genitalia, and maxillary and labial palps areconsidered serially homologous. Homeotic mutations cantransform genitalia, maxillary palps, and labial palps intolegs and/or antennas (Bridges and Dobzhansky 1933;Duncan et al. 1998; Estrada and Sanchez-Herrero 2001).The differences among these appendages reflect the inputof Hox selector genes. Antennas are thought to correspondto the absence of Hox gene input and leg identitiesreflecting the action of a single Hox gene: Sex combsreduced (Scr) for prothoracic (T1), Antennapedia (Antp) formesothoracic (T2), or Ultrabithorax (Ubx) for metathor-acic (T3) legs (Struhl 1982). By contrast, labial imaginaldisc development yielding the adult proboscis involves thejoint action of two Hox genes, proboscipedia (pb) and Scr(Percival-Smith et al. 1997). Insect adult labial structuresare morphologically diverse appendages specialized forfeeding and taste (see, for example, Angelini and Kaufman2005; Hughes and Kaufman 2000, 2002). In Drosophila, thelabial palps are derived structures with distinctively striatedpseudotracheal rows used for drinking fruit juice. The labialdiscs that normally give rise to adult labial palps can betransformed to antennal aristae by partial loss of pb functionactivity, while complete loss-of-function mutants transformlabial mouthparts to distal T1 legs (Kaufman 1978).
Here we examine the genetic program of the highlyderived labial organ and its relationship to other ventralappendages. To this end, we have analyzed the functionalties among four selector genes, pb, Scr, hth, and ss. Usingloss-of-function mutations of these genes, singly or incombination, coupled with tissue-directed mitotic recom-bination, we removed their functions from developinglabial palps. Various mutant combinations transformedlabium alternately to antenna- or leg-related structures. It issurprising to note that though ss mutations are withouteffect in labial development, mutant combinations remov-ing both pb and ss functions yielded the most severe labialphenotype observed, namely, stunted tripartite legs. Wepresent evidence that tissue identity depends not only onselector composition but also to an unexpected extent oncell nonautonomous processes influenced by the wg anddpp signaling pathways. We discuss the roles of selectortranscription factors in the fine regulation of signalingpathways for tissue identity and evolution. We argue thatgenotypes giving rise to these appendages constitute agenetic default state, and that they approximate theprimitive or “evolutionary ground state” form that gaverise to ventral arthropod appendages.
Materials and methods
Drosophila strains and culture Drosophila stocks weremaintained at 22°C on standard cornmeal/agar mediumunless otherwise noted. The multiple mutant chromosomes
were generated by meiotic recombination. Mutant chro-mosomes were maintained as heterozygotes with theTM6B, HuTb balancer chromosome harboring the larval/pupal marker Tb. The transgenic pbGAL4(10) line wasused to generate mutant clones in labial imaginal discs asdescribed in Joulia et al. (2005).
Adult cuticle analysis Flies of interest were stored inethanol until dissection. Detailed examinations of dis-sected heads and legs were performed by light microscopy(Zeiss Axiophot) after mounting in Hoyer’s medium.
Immunohistochemistry and antibodies Antibody stainingswere carried out essentially as described (Pattatucci andKaufman 1991). Imaginal discs dissected and mounted onmicroscope slides in 0.5× phosphate-buffered saline/50%glycerol were viewed by confocal microscopy (Zeiss LSM410 or Leica TCS SPII). Images were processed usingAdobe Photoshop. Monoclonal antibodies directed againstWg and Dac proteins were obtained from the Develop-mental Studies Hybridoma Bank. Monoclonal mouse anti-Dll was provided by I. Duncan, polyclonal rabbit anti-Hthby A. Salzberg, rat anti-Dan by S. Cohen, and rabbit anti-En by M. Crozatier and A. Vincent. The dilutions ofantibodies were: mouse anti-Dac, 1:100; rabbit anti-Hth,1:500; mouse anti-Dll, 1:500; rat anti-Dan, 1:1,000; mouseanti-Wg, 1:100; and rabbit anti-En, 1:100. The secondaryantibodies employed, directed against rat, mouse, or rabbitIgGs, were coupled to rhodamine or fluorescein isothio-cyanate fluorochromes.
Clonal analysis Mitotic recombination events giving riseto clones were produced by the flippase recombinasetarget (Flp/FRT) system. Non tissue-directed clones of pb−,Scr−, and pb− Scr− cells as well as Scr− cells in pb−
context were obtained in larvae of genotype y w hs-Flp;FRT82BKi pb5/FRT82B P[Ub-GFP], y w hs-Flp; FRT82B
Scr−/FRT82B P[Ub-GFP], y w hs-Flp; FRT82Bpb−Scr−/FRT82B P[Ub-GFP], y w hs-Flp; and FRT82Bpb−Scr−/FRT82BKi pb5 P[Ub-GFP]. Clones were induced byexpression of Flp recombinase from the hsp70 promoter(hs-Flp) via heat shocks during the first or second larvalinstars (2 h at 37.5°C). Alternatively, localized recombi-nase expression from coupled pbGAL4>UAS:Flp trans-genes led to mitotic clones in the small labial imaginaldiscs at high frequency (≥1 clone/disc). Mutant clone sizewas enhanced by the Minute technique (Morata and Ripoll1975) in larvae derived from crosses of pbGAL4(10)UAS-Flp; FRT82BM(3)rps3 P[Ub-GFP]/T(2;3)CyO-TM6B, Tb flies with stocks harboring the followingchromosomes in combination with the TM6B, HuTbbalancer: FRT82BScr1 (from A. Percival-Smith); FRT82B-hth5E (from A. Salzberg); FRT82Bpygo130; FRT82BssD115-7
(from I. Duncan); and FRT82Bpb27Scr2 (from A. Percival-Smith).
Chromosomes constructed for this work and maintainedwith TM6B, Hu Tb were: FRT82BKi pb5pp; FRT82BKi pb5
hth5E; FRT82Bputp1; FRT82BKi pb5putp1; FRT82BKi pb5
432

pygo130; FRT82BKi pb5putp1pygo130; FRT82BScr1hth5E;FRT82BKi pb5ssa; FRT82Bpb27Scr2hth5E; FRT82Bpb27Scr2
ssa; FRT82Bpb27Scr2putp1; FRT82Bpb27Scr2hth5Eputp1;FRT82Bpb27Scr2hth5Essa; and FRT82Bpb27Scr2hth5Eputp1
ssa.
Results
The Hox selectors Pb and Scr differentially control hthand Dll
Mutations of selector genes can lead the labial palps ofD. melanogaster to adopt multiple alternative identities(Kumar et al. 1979). The labial palps are partiallytransformed into antennal aristae on reducing pb function,or to T1 legs in the pb− (i.e., null) condition (Kaufman1978). Labial palps of Scr hypomorphs are modifiedtoward structures considered to be maxillary (Abzhanov etal. 2001), while the loss of both pb and Scr results in theappearance of antennal structures (Percival-Smith et al.1997). Two putative “antennal” selector genes, hth and ss,are also normally expressed in labial discs. To comparedevelopmental programs yielding labium, leg, or antenna(1a1-c1), and to better understand the molecular basis oflabial transformations resulting from selector mutations,we examined the patterns of a number of position markersin normal labial discs and in mutant tissue destined towardalternative identities.
Mutant labial discs were generated in mosaic animals bymitotic recombination, using a tissue-targeted FLP/FRTsite-specific recombination strategy (Newsome et al. 2000)that efficiently couples the pbGAL4 driver with a UAS:FLP element expressing the recombinase (pb>FLP; ≥1clone per labial disc). On enhancing the relative growth ofmutant clones by the Minute technique (“Materials andmethods”), the resulting adult mouthparts were composedmostly of mutant cells. To determine the developmentaleffect of removing one or more Hox selector functions,pb>FLP was used to generate labial palps harboring clonesof mutant cells. Targeted Minute-enhanced clones of pb−
cells gave rise to T1 legs comparable to pb− homozygotes(Fig. 1d1). By contrast, though loss of Scr or hth functionsresulted in a reduction of pseudotracheal rows, thesedefects were unaccompanied by transformation (data notshown). Thus, both Scr and hth are necessary for normallabial development, but their loss-of-function does notreveal explicit specification roles in this appendage. Lossof ss function removed sensory hairs but had no otherdetectable effect on labial specification (data not shown).pb− Scr− labial discs devoid of Hox gene expression weretransformed to yield adult antennas harboring the dis-tinctive a3 segment and terminal arista (Fig. 1e1).
The expression patterns of molecular markers includingdac, dan, Dll, and hth were compared among imaginaldiscs giving rise to wild-type leg, antenna, or labium(Fig. 1, columns a, b, and c), or mutant labial discsharboring clones of pb− or pb− Scr− cells (Fig. 1, columnsd and e). Dac and Dan markers are differentially expressed
among the labial, leg, and antennal discs (Abzhanov et al.2001; Emerald et al. 2003). Localized nuclear Dacaccumulation is observed as a full ring in leg discs or acrescent in antennal discs, respectively (Fig. 1a2,b2), but isabsent from the wild-type labial disc (Fig. 1c2). Novel Dacprotein expression was detected within labial clones of pb−
and pb− Scr− cells that give rise to distal leg and antenna,respectively (Fig. 1d2,e2). Dan expression is normallyspecific to the antennal disc (Fig. 1b3), and absent from theleg and labial discs (Fig. 1a3,c3). Consistently, Dan wasnot expressed in clones of pb− cells giving rise to legstructures (Fig. 1d3), but was detected for pb− Scr− clonesthat yield antennas (Fig. 1e3). The reprogramming of adultlabium to antenna by simultaneous removal of both Hoxselectors is reflected in larval development by the Dac andDan identity markers. The appearance of adult antennalaristae is reliably presaged by Dan expression, whether inwild-type antennal discs that do not express Hox selectorproteins or in labial discs transformed by mutationsremoving pb and Scr functions.
In leg discs, normal expression of the hth and Dll genesleads to accumulation of their homeodomain proteins indistinct cell populations (Fig. 1a4,a5), while in antennaldiscs they overlap in a wide band (Fig. 1b4,b5). In wild-type labial disc pouches, Dll accumulates at the distal endopposite the site of attachment to the cuticular larvalmouthparts (Fig. 1c4), while Hth accumulates in a band ofproximal cells adjacent to the attachment site (Fig. 1c5). Inlabial discs harboring large pb− clones that yield a labium-to-leg transformation, the normal pouch opens to yield anepithelial monolayer where Dll expands (compare Fig. 1d4with Fig. 1c4), while Hth remains proximally restricted(Fig. 1d5) as in the leg disc (Fig. 1a2,a3). In pb− Scr− labialdiscs yielding antennas, expanded Dll expression encom-passes most of the tissue (Fig. 1e2) while Hth expressionextends centrally compared to pb− clones (compareFig. 1d5,e5). These results, validated by double labelingexperiments in wild-type or pb− labial discs (Fig. 1f,g), orin discs harboring large pb− Scr− clones (Fig. 1h,h′),confirm that Dll and Hth accumulate separately in leg-destined labial discs but overlap broadly in transformeddiscs yielding antennas. Labial discs with large pb− Scr−
clones give rise to a 100% penetrant antennal transforma-tion (Fig. 1e1; 48/48 hemi-labia), express Dan (Fig. 1e3),and show strongly overlapping Dll/Hth accumulation(Fig. 1h,h′). These observations support an instructiverole for overlapping Hth and Dll expression in mutantlabial discs comparable to that in normal antennal discs(Dong et al. 2000).
Multiple Hox inputs regulate hth and Dll expression
Drosophila Hox functions are known to spatially limit theexpression of Dll and/or Hth. For example, Dll wasidentified as a transcriptional target repressed by the Ubxand Abd-A proteins in the embryonic abdomen (Vachon etal. 1992), while a role in limiting imaginal hth expressionwas imputed to Antp, abd-A, Scr, andUbx (Yao et al. 1999).
433
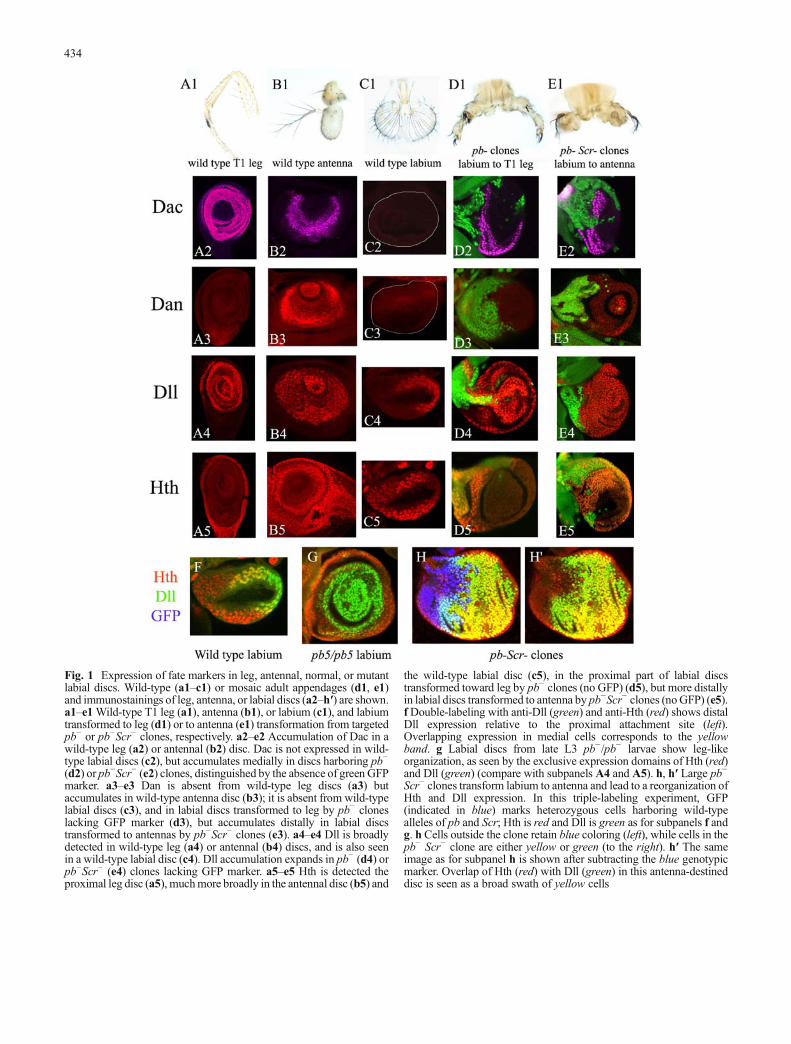
Fig. 1 Expression of fate markers in leg, antennal, normal, or mutantlabial discs. Wild-type (a1–c1) or mosaic adult appendages (d1, e1)and immunostainings of leg, antenna, or labial discs (a2–h′) are shown.a1–e1Wild-type T1 leg (a1), antenna (b1), or labium (c1), and labiumtransformed to leg (d1) or to antenna (e1) transformation from targetedpb− or pb−Scr− clones, respectively. a2–e2 Accumulation of Dac in awild-type leg (a2) or antennal (b2) disc. Dac is not expressed in wild-type labial discs (c2), but accumulates medially in discs harboring pb−
(d2) or pb−Scr− (e2) clones, distinguished by the absence of green GFPmarker. a3–e3 Dan is absent from wild-type leg discs (a3) butaccumulates in wild-type antenna disc (b3); it is absent from wild-typelabial discs (c3), and in labial discs transformed to leg by pb− cloneslacking GFP marker (d3), but accumulates distally in labial discstransformed to antennas by pb−Scr− clones (e3). a4–e4 Dll is broadlydetected in wild-type leg (a4) or antennal (b4) discs, and is also seenin a wild-type labial disc (c4). Dll accumulation expands in pb− (d4) orpb−Scr− (e4) clones lacking GFP marker. a5–e5 Hth is detected theproximal leg disc (a5), muchmore broadly in the antennal disc (b5) and
the wild-type labial disc (c5), in the proximal part of labial discstransformed toward leg by pb− clones (no GFP) (d5), but more distallyin labial discs transformed to antenna by pb−Scr− clones (noGFP) (e5).fDouble-labeling with anti-Dll (green) and anti-Hth (red) shows distalDll expression relative to the proximal attachment site (left).Overlapping expression in medial cells corresponds to the yellowband. g Labial discs from late L3 pb−/pb− larvae show leg-likeorganization, as seen by the exclusive expression domains of Hth (red)and Dll (green) (compare with subpanels A4 and A5). h, h′ Large pb−
Scr− clones transform labium to antenna and lead to a reorganization ofHth and Dll expression. In this triple-labeling experiment, GFP(indicated in blue) marks heterozygous cells harboring wild-typealleles of pb and Scr; Hth is red and Dll is green as for subpanels f andg. h Cells outside the clone retain blue coloring (left), while cells in thepb− Scr− clone are either yellow or green (to the right). h′ The sameimage as for subpanel h is shown after subtracting the blue genotypicmarker. Overlap of Hth (red) with Dll (green) in this antenna-destineddisc is seen as a broad swath of yellow cells
434

In the labial disc where the homeotic Pb and Scr selectorproteins accumulate ubiquitously, homozygotes for pb-nullmutations show a marked proximal expansion of Dllexpression to encompass most of the disc, showing thatnormal pb function serves to negatively regulate Dll(Abzhanov et al. 2001). Large clones of pb− cells yield thesame expansion of Dll (compare Fig. 1c4 and d4; Joulia etal. 2005). However, because Dll accumulates both inmutant and in pb+ cells in the case of large clones (yellowcells, Fig. 1d4), this effect is not cell-autonomous.Gradients of diffusible signals from localized sources ofdpp and wg expression lead to activation of Dll (Abu-Shaarand Mann 1998; Lecuit and Cohen 1997). Normal pbfunction is required in labial discs of third instar larvae (L3)where it strongly downregulates dpp and wg (Joulia et al.2005). Thus, pb might restrict labial Dll expression at leastin part indirectly by restricting cell-signaling. In agreementwith this possibility, small clones of pb− cells inducedbefore the L3 stage had no effect on Dll accumulation.Proximal clones did not express Dll, while mutant distalcells expressed Dll normally. The effects of pb on Dllexpression are therefore mostly or solely indirect.
By contrast, hth expression in leg discs is restricted to aproximal zone where wg and dpp concentrations arerelatively low (Abu-Shaar and Mann 1998; Lecuit andCohen 1997). Accordingly, labial discs harboring largeclones of pb− or pb−Scr− cells open toward a leg form withHth expression retracted to the disc circumference. Whilesmall distally situated clones of pb− or pb−Scr− mutantcells (induced by hsFlp, no Minute mutation) were withouteffect, some pb− cells in proximal clones showed enhancedHth protein accumulation (Fig. 2a,a′). Thus, pb normallyacts to limit Hth accumulation in expressing cells. In Scr−
clones Hth accumulation was unaltered (Fig. 2b,b′),suggesting that Scr does not regulate hth in this tissue.However, this is not the case because a pronouncedenhancement of Hth was seen in pb−Scr− cells (Fig. 2c,c′).Similarly, Scr− clones induced in a homozygous pb−
context (i.e., pb−Scr− cells) showed enhanced hth expres-sion compared with their pb− neighbors. We conclude thatPb and Scr contribute jointly and cell-autonomously tolimiting Hth accumulation, and that modifications of Dlland Hth spatial patterns in pb− and pb−Scr− mutant tissuesreflect Hox inputs at both the cellular and tissue levels oforganization.
The labium-to-antenna transformation can bypass hthbut requires ss
The preceding results support the view that Hox genesrepress antennal fate at least in part by downregulating Hthaccumulation (Casares and Mann 1998; Yao et al. 1999).To test the requirement for hth in making these ectopicantennas, we removed labial hth activity via targeted triplemutant pb− Scr− hth− clones. The predominant outcome inresulting adults was compact leg appendages identified byclaws and bracted bristles (Fig. 3a; 53/66 hemi-labia, 81%).Accordingly, Dll (Fig. 3b) and dac (Fig. 3c) expression
patterns were unchanged compared with pb− and pb− Scr−
clones (Fig. 1d2,e2), while the Dan antennal fate markerwas generally absent from triple mutant labial discs(Fig. 3e). This confirms that normal hth function is neededfor pb− Scr− labial cells to form antennas. Unexpectedly,contrary to the fully penetrant antennal transformationobtained on generating pb− Scr− clones (48/48; e.g.,Fig. 1e1), the remaining 19% (13/66 hemi-labia) showed amix of leg and antennal tissue (Fig. 3f). This resultsuggested that antennal structures can be specified evenwithout hth. Accordingly, Dan accumulation prefiguringantenna was detected in a minority of imaginal discscontaining pb− Scr− hth− clones (Fig. 3g). Such expressionwas not limited to mutant cells but extended to pb+ Scr+
hth+ cells identified by GFP expression (Fig. 4h,h′; note theGFP+ cells accumulating Dan). This observation, consis-tent with a previous report (Emerald et al. 2003), indicatesthat dan is activated by a nonautonomous signal receivedby cells both within and outside the clone. Mixed leg/
Fig. 2 Hth accumulation is regulated by pb/Scr. pb−, Scr−, orpb−Scr− clones were produced using Flp/FRT-directed site-specificrecombination. a–c Immunostainings with anti-Hth. Wild-type cellsexpress GFP (green); wild-type cells expressing Dll or Hth areyellow. Mutant cells lack GFP (black), and mutant cells expressingDll or Hth are red. a′–c′ Immunostainings alone are shown onadjoining right panels with clones of mutant cells encircled by lightlines. Hth accumulation is enhanced in some but not all pb− mutantcells (a–a′, arrow vs asterisk). No enhancement is observed for Scr−
clones (b–b′). The effect of pb− cells is further enhanced in allmutant cells of pb−Scr− clones (c–c′)
435

antennal transformations likewise resulted from inducingScr− clones in a pb-mutant background; in these condi-tions, activation of dan in wild-type cells was also detected(Fig. 3i,i′,i″).
The unexpected detection of Dan in and around pb− Scr−
hth− clones suggested a diffusible signal acting betweenhth and dan. We therefore tested the function of anotherknown regulator of dan, the bHLH transcription factorgene ss (Duncan et al. 1998; Emerald and Cohen 2004;Emerald et al. 2003). Null mutants of ss are semiviable andtransform adult antennas into stunted legs that weresuggested to represent the ventral “ground state” (Duncanet al. 1998). ss is activated by de novo coexpression of hthand Dll, placing it downstream of hth (Dong et al. 2002).To ask whether residual dan activation depends on ssactivity, we looked at the role of ss in labial transforma-
tions. Though ss is normally expressed in wild-type labialdiscs (data not shown), loss-of-function mutants have nodiscernible effect on labial specification (Fig. 4a). Howev-er, clones for all mutant combinations removing both pband ss functions (including pb− ss−, pb− Scr− ss−, and pb−
Scr− hth− ss−) yielded a 100% penetrant transformation oflabium to stunted legs identified by their distal claws(Fig. 4b–d; ≥38 hemi-labia examined for each genotype).These legs appear to be composed of three segments, butlack all vestiges of the tarsae still detectable in pb− Scr−
hth− mouthparts (Fig. 3a). The full penetrance indicatesthat removing ss in these combinations stabilizes the newmutant state. We attribute the residual antenna-formingcapacity of pb− Scr− hth− triple mutants to ongoing ssactivity, and infer that ss acts downstream of the presump-tive antennal signal.
Fig. 3 The effect of removing the known selector from labial tissue,and visualizing a nonautonomous mechanism implicated in antennalfate specification. Transformed labial palps of mosaic adults (a andf) and immunostainings of corresponding labial discs (b–i″) areshown. All the clones were generated with the pb>Flp targetingsystem. Wild-type cells express GFP (green), mutant cells do not.a pb−Scr−hth− clones adopt a leg identity in 80% of cases, whileb pb−Scr−hth− labial discs broadly express Dll; c Dac accumulatesin mutant cells; d Wg is detected in the ventral side along the A/Pboundary; and e Dan is usually not detected. f In the remaining 20%of mosaic adults, pb−Scr−hth− clones exhibit a mixed transforma-
tion with both antennal aristae (arrow) and leg claws (asterisk).g Similarly, about 20% of discs harboring pb−Scr−hth− mutant cellsexpress Dan (violet). h–h′ Dan is detected here both in the mutantclone (no GFP) and in adjacent wild-type (circled) cells. Scr− clonesgenerated in a pb null context also yield mixed leg/antennaltransformations (data not shown). In this context, nonautonomousDan (red, subpanel i) expression is detected both in mutant pb−Scr−
cells (no GFP, subpanel i′) and in adjacent wild-type, GFP-expressing cells. This is best seen in subpanel i″ (yellow cells arenonmutant but Dan-expressing)
436

Wg and Dpp signaling pathways are implicatedin tissue identity
The preceding results, supporting the existence of adiffusible signal used in normal or mutant labial specifica-tion, led us to test known signaling pathways for a possiblerole. Recent observations link Hox functions to Sonichedgehog signaling in the mouse (Chen et al. 2004; Zakanyet al. 2004), while mutants for wg can lead to homeotictransformations (Emerald and Roy 1997). Similarly, wehave identified a crucial role for pb at the anteroposteriorfrontier of the labial disc where Pb antagonizes Cubitusinterruptus to repress expression of the diffusible signalingfactors encoded by wg and dpp in response to hh signaling(Joulia et al. 2005). Whereas normal labial expressionlevels of wg and dpp are low compared with leg or antennaldiscs, in pb− discs Hh efficiently activates wg and dpp withstriking consequences (Joulia et al. 2005). To test theeffects of Wg and/or Dpp signaling in specifying “labial”identity, we therefore modified their activities in a pb−
context.To inactivate the wg pathway in a pb− context, we
removed the function of the pygopus (pygo) gene thatencodes an obligatory component of the wg pathway(Belenkaya et al. 2002; Kramps et al. 2002; Parker et al.2002; Thompson et al. 2002; Thompson 2004; Townsley etal. 2004), then induced targeted clones of pb−pygo− cells.Similarly, to inactivate the dpp pathway we made use of thepunt (put) gene that encodes a requisite TGF-β type IIreceptor (Burke and Basler 1996; Letsou et al. 1995;Ruberte et al. 1995; Simin et al. 1998). Labial discsharboring wg-deficient pb− pygo− clones developed asdiminished stubs (compare pb− in Fig. 1d1 with pb− pygo−
in Fig. 5a) with residual leg identity as shown by the
presence of bracted bristles (n=48; see Fig. 5a). Bycontrast, when dpp signaling was abolished via pb− put−
double mutants, tissue identity was often redirected.Whereas targeted pb− clones led to prothoracic legs in100% of cases, labial discs with pb− put− clones gave risein equal proportions to T1 legs (25/50 hemi-labia) or toantennal aristae (25/50). This is illustrated by Fig. 5b whereone labial disc gave rise to T1 leg structures while the otheryielded an antennal arista. Mixed identity leg/antennalappendages arising from a single disc were not observed.Double mutant pb− pygo− discs retain the capacity toreceive and transduce Dpp but not Wg signals, while pb−
put− discs can receive extracellular information transmittedby Wg but not Dpp. These results thus implicate Wg signalin distinguishing leg from antennal identity in a pb mutantlabial disc. While Dll and dac PD patterning markers wereequivalently expressed in labial imaginal discs with pb−
(Fig. 1) or pb− put− clones (Fig. 5c,d), accumulation of theDan antennal marker was detected in some labial discsharboring pb− put− clones (Fig. 5f). The revived danexpression and antennal phenotype could result from adirect role of dpp signaling, or it might reflect an indirectperturbation of wg signaling due to the unbalancing of themutual inhibition of these two pathways (Brook and Cohen1996; Jiang and Struhl 1996; Penton and Hoffmann 1996).We favor the latter explanation for two reasons. First,overexpressing wg can alter labial identity (Johnston andSchubiger 1996; Joulia et al. 2005; Maves and Schubiger1995). Second, in labial discs harboring pb− put− clones,the Wg protein is no longer restricted to the ventral sectorbut accumulates along the full length of the AP boundary(Fig. 5e). To test whether the expanded wg signal is indeedessential for the antennal transformation, pb− put− pygo−
triple mutants were employed. In this case, only leg
Fig. 4 Joint action of pb and ssin transformations of adult labialpalps. a Labium of an adulthomozygous mutant for ss. Themutant proboscis appears wild-type except for the absence ofbristles. b–d Labial palps har-boring pb−ss− (b), pb−Scr−ss−
(c), or pb−Scr−hth−ss−
(d) clones show the same 100%penetrant labium-to-leg trans-formation. This shortened com-pact appendage shows no signof tarsal articulation, but regu-larly assumes the tripartite formindicated in subpanel b.A similar form can be notedin subpanel c
437
structures resembling pb−pygo− double mutants wereobserved (data not shown). We favor the interpretationthat wg signaling is inhibited by dpp in mutant pb− labialdiscs directed toward legs. This wg signal is apparentlyinstructive and can redirect labial cells toward antennalidentity. More generally, these results raise the possibilitythat the wg/dpp signaling balance, coupled with Hoxselector function, contributes to tissue identity.
The ventral appendage ground state and cellsignaling pathways
Multiple genetic conditions exist for executing an antennalor a “leg” program. Relevant parameters include theactivities of transcriptional regulators but also of cell-signaling pathways. As stated above, 19% of triple mutantpb− Scr− hth− labial discs adopt mixed leg/antennal fate.To ask whether cell-signaling input can condition identityindependently of pb, Scr, and hth selector functions, wetested a quadruple mutant pb− Scr− hth− put−. Labial palpsof these quadruple mutant animals lacking normal dppsignaling capacity were transformed to antennal aristae in76% of examined samples (47/62 hemi-labia), and mixedtransformations were not observed (see Fig. 6a). In discswith large pb− Scr− hth− put− clones, PD patterning(visualized here by Dac, Fig. 6b) appears to be allocated asfor pb−, pb− Scr−, or pb− Scr− hth− clones. However, Wgaccumulation was expanded along the AP boundary(Fig. 6c) as for pb− put− clones, and Dan was detected insome samples (Fig. 6d). We infer that the identity adoptedby a group of cells can be altered by modifying availablecell-signaling information and the selector activities of thePb, Scr, and Hth homeodomain proteins.
To test whether the residual antennal capacity revealed inpb− Scr− hth− put− discs reflects ongoing ss+ function, aquintuple mutant pb− Scr− hth− put− ss− chromosome wasconstructed. Progeny harboring targeted labial clonesshowed a fully penetrant transformation to stunted legs
(Fig. 6e; 36/36 hemi-labia). Dac and Wg markers wereunchanged in these conditions relative to pb− Scr− hth−
put− quadruple mutants (Fig. 6f and g, respectively), butthe Dan antennal marker was no longer detected (Fig. 6h)in agreement with the fully penetrant adult leg phenotype.We conclude that the antennal identity observed for pb−
Scr− hth− put− clones with modified wg/dpp signals(Fig. 6a,c) requires normal ss function.
Discussion
From labial palp transformations toward theevolutionary ground state
The Drosophila labial imaginal disc is organized similarlyto other ventral discs, making use of hh-mediated signalingto give rise to a PD organization and appendage structure(Joulia et al. 2005). Cells of the labial disc can clearlyassume three distinct morphologies—labium, antenna, orleg. These gene-dependent homeotic transformationsreveal that serially homologous genetic pathways specifyserially homologous appendages.
The remarkable developmental plasticity of the labialappendage makes it an intriguing model for examining howa genetic program can be modified in evolution. Thepremise that serially homologous structures reflect accu-mulated selector functions was tested by geneticallyablating combinations of known selector gene functionsin labial cells. We have identified four different geneticconstitutions that transform labium to antennas withterminal aristae, interspersed with genotypes that result informs of leg tissue terminating in distal claws, summarizedin Fig. 7.
It is important to note that while some outcomes are fullypenetrant and reliable, others are variable. We posit thatstable developmental outcomes should be favored duringevolution. Consequently, genetic changes concordant withstability may help us understand important developmental
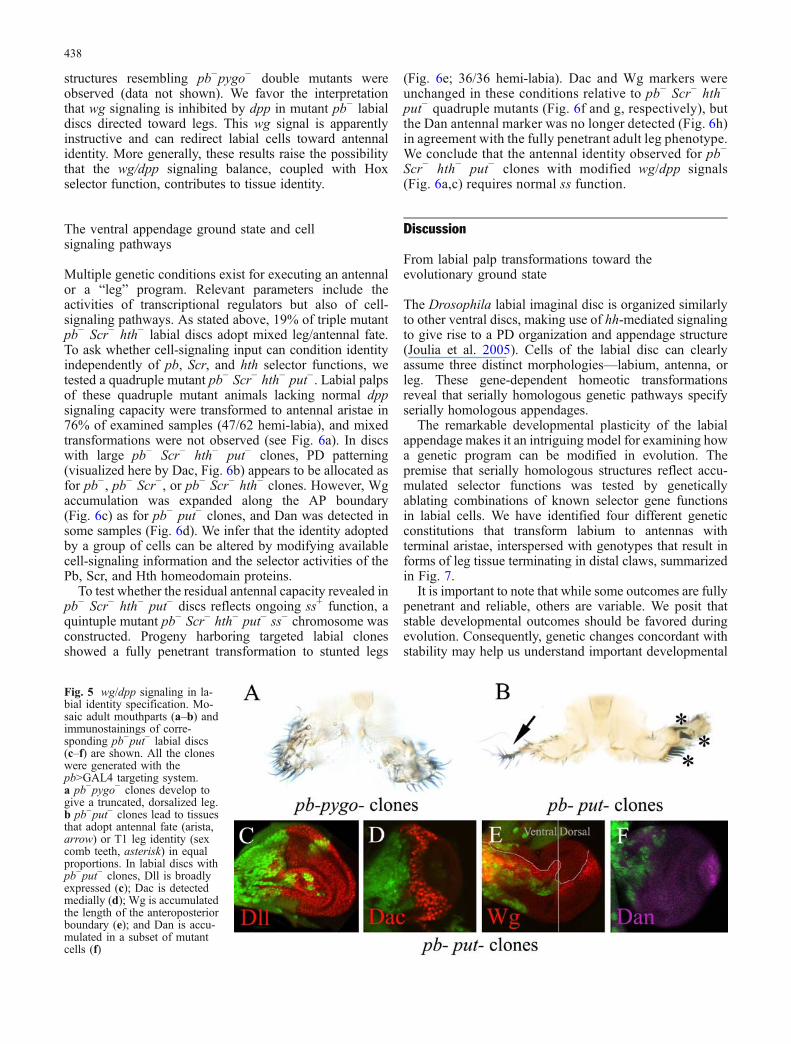
Fig. 5 wg/dpp signaling in la-bial identity specification. Mo-saic adult mouthparts (a–b) andimmunostainings of corre-sponding pb−put− labial discs(c–f) are shown. All the cloneswere generated with thepb>GAL4 targeting system.a pb−pygo− clones develop togive a truncated, dorsalized leg.b pb−put− clones lead to tissuesthat adopt antennal fate (arista,arrow) or T1 leg identity (sexcomb teeth, asterisk) in equalproportions. In labial discs withpb−put− clones, Dll is broadlyexpressed (c); Dac is detectedmedially (d); Wg is accumulatedthe length of the anteroposteriorboundary (e); and Dan is accu-mulated in a subset of mutantcells (f)
438
constraints on evolution as postulated by Gilbert et al.(1996) and Raff (1987). One such stable outcome forDrosophila labial cells is a stunted leg-like structurebearing distal claws that we interpret as a primitive leg. Thecommon element of the multiple genetic combinationsyielding such legs was loss of the AP identity functionprovided by the Hox gene pb and the antennal selectorfunction of ss. Indeed, in all tested pb−ss− mutantcombinations, the formation of stunted legs was fully
penetrant and their form was remarkably constant. Wepostulate that this form is consistent with prior use andrefinement in evolution.
Several related concepts dealing with comparativeanatomy are used in the literature. One, the bauplan relatedto the preevolutionary archetype concept of Owen is stillused to describe the deduced ancestral, primitive archi-tecture common to large groups of organisms such asclasses or phyla (Brusca and Brusca 2002). Another is thegenetic “default state,” which can in principle be created—and observed experimentally—by removing selector func-tions via loss-of-function mutations. Insofar as the defaultstate removes all evolutionarily acquired selector functions,it is equivalent to the primitive or evolutionary groundstate. Comparative morphology has suggested a bipartiteancestral limb consisting of a proximal coxopodite and asegmented distal telopodite. Accordingly, in animalsharboring large clones of hth or of Antp hth mutant cells,the stunted appendages observed were interpreted torepresent a ground state ventral appendage composed ofa coxopodite and a telopodite (Casares and Mann 2001).However, this direct interpretation is associated withseveral complicating factors. One is the intrinsic stochasticnature of clonal analyses. Another concerns the divergentroles played by hth, not only as an antennal “selector” butalso as a crucial element of PD patterning that affects cellmingling along the PD axis (Emerald and Cohen 2001). Inlight of our present results, yet another potentially importantvariable is the synergizing role of signaling-mediatedfeedback loops to modify developmental outcome.
pb−Scr−hth−ss− quadruple mutants correspond to aputative labial default state. This mutant tissue is devoidof known selector functions, and its developmentaloutcome should approach the evolutionary ground statefor ventral arthropod appendages. The resulting leg-likelabial appendage lacks tarsal articulations; indeed, it ismore stunted than for hth− legs, interpreted as a groundstate condition (cf. Casares and Mann 2001). These labialstructures closely resemble the transformed antennas of ssmutants, a condition likewise suggested to correspond to anevolutionary ground state (Duncan et al. 1998). Further,antennas harboring pb−Scr−hth−ss− mutant clones arestrongly similar to the quadruple mutant labial palps, or toss null antennas (Duncan et al. 1998). These similaroutcomes indicate a central role for ss in both antennal andlabial development.
The pb−Scr−hth−ss− “labial” discs express both Dll anddac, presumably in direct response to wg and dpp organizersignals. This observation is in agreement with recentmolecular findings from several groups showing expres-sion of the medial marker dac not only in other insects butalso in a spider (Chelicerata) and a millipede (Myriapoda)(Abzhanov and Kaufman 2000; Inoue et al. 2002; Prpicand Tautz 2003; Prpic et al. 2001). Thus, the subdivision ofthe leg into three domains distinguished by expression ofDll, dac, and hth may have occurred before thediversification of arthropods into chelicerates, myriapods,crustaceans, and insects (Kojima 2004). In agreement withthis assessment, quintuple mutant pb−Scr−hth−put−ss−
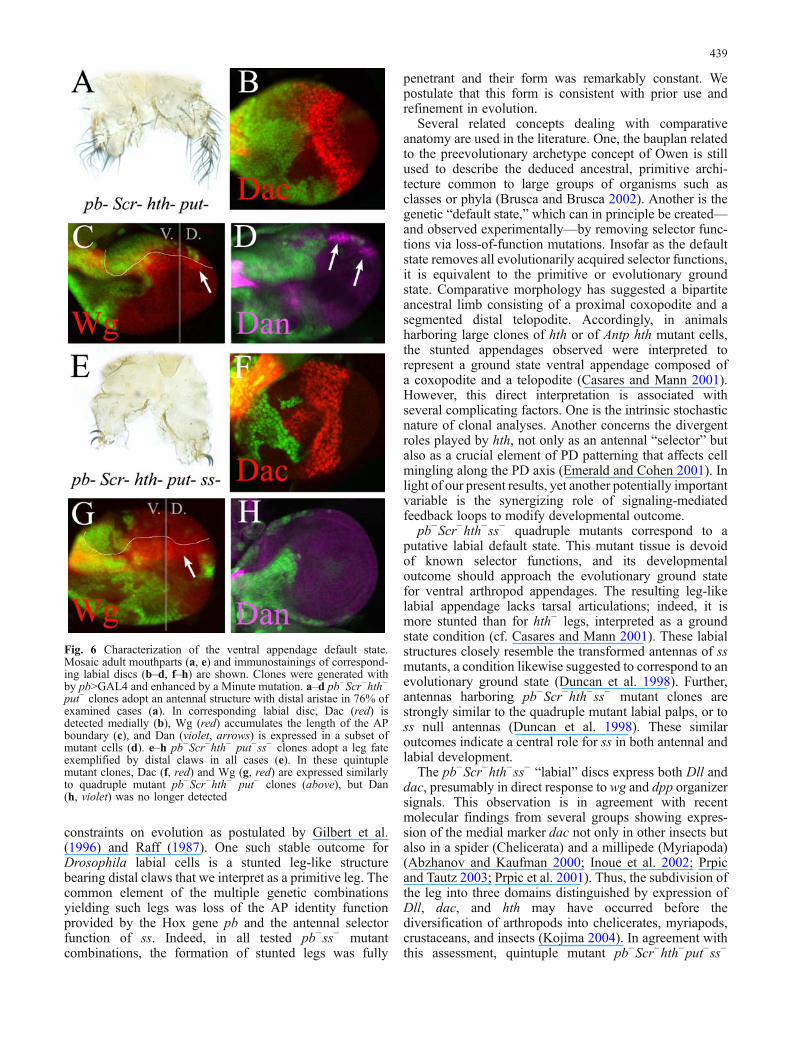
Fig. 6 Characterization of the ventral appendage default state.Mosaic adult mouthparts (a, e) and immunostainings of correspond-ing labial discs (b–d, f–h) are shown. Clones were generated withby pb>GAL4 and enhanced by a Minute mutation. a–d pb−Scr−hth−
put− clones adopt an antennal structure with distal aristae in 76% ofexamined cases (a). In corresponding labial disc, Dac (red) isdetected medially (b), Wg (red) accumulates the length of the APboundary (c), and Dan (violet, arrows) is expressed in a subset ofmutant cells (d). e–h pb−Scr−hth− put−ss− clones adopt a leg fateexemplified by distal claws in all cases (e). In these quintuplemutant clones, Dac (f, red) and Wg (g, red) are expressed similarlyto quadruple mutant pb−Scr−hth− put− clones (above), but Dan(h, violet) was no longer detected
439

discs yield a rudimentary appendage via imaginal discs thatexpress the medial dac marker, providing consistentgenetic evidence for a leg subdivided into three regionsin a common arthropod ancestor. We favor the interpreta-tion that the putative labial default state described here is agood approximation of the primitive state for an ancestralarthropod leg that was already structured into proximal,intermediate, and distal domains. We infer that theexplicitly segmented tripartite legs of present day insectsare derived from a simpler tripartite version lacking explicitsubdivisions. From a direct phylogenetic analysis it can bederived that the common ancestor of Onychophora,Tardigrada, and Arthropoda, i.e., the “Panarthropod”ancestor (Nielsen 1998), possessed a nonsegmented,claw-bearing appendage. Thus, the labial legs of pb ssmutants likely resemble the legs of the common ancestor ofArthropods and potentially even of Panarthropods.
Combinatorial action of transcription factors and cellsignaling functions in identity and evolution
It has long been known that mutations altering the functionof selector transcription factors can redirect developmentalidentity. More surprising, however, is the extent to whichmodulating cell signaling pathways common to allsegments can distinctively impact the identity acquiredby the cells giving rise to a specific appendage. Theobservations united in this work lead us to entertain theviewpoint that signaling information may be an integralpart of selector function. Our observations suggest that a“community effect” (Gurdon 1988) unites the cell popu-
lation in a coherent choice, and that the dpp and wg cellsignaling pathways inform identity specification. We arguethat this is indeed an inherent aspect of homeotic segmentidentity function because it seems reasonable that mod-ulating the communication between the compartmentscomposing a segment could instruct or modify the resultingidentity. Several recent papers indicate surprisingly directlinks between the segment polarity cascade and Hoxtranscriptional regulation of target genes (Chen et al. 2004;Gebelein et al. 2004; Hersh and Carroll 2005; Joulia et al.2005). Thus, it appears that segmental actions of Hoxproteins can be decomposed into compartmental actionslinked by actions of signaling pathways, again suggestingthat specifying segmental identity involves the integrationof the signals emitted from its compartments.
Cells that acquire antennal identity communicate thisinformation to their neighbors, as indicated by thenonautonomous induction of dan (Fig. 3g–j″). Theunidentified antennal signal appears to act between Dll/hth and ss function, as shown by the epistatic effect of ss inthe quintuple mutant pb−Scr−hth−put−ss− (leg) comparedwith the quadruple version pb−Scr−hth−put− (antenna/leg).Conversely, the existence of a leg signal can be deducedfrom clones of the same quintuple mutant where groups ofheterozygous labial cells expected to behave as wild typeassume leg identity (data not shown). The most parsimo-nious explanation is that the nonautonomous signal(s)directly reflects modulated wg/dpp output (variation insignal intensity, duration, et cetera). Accordingly, theobserved nonautonomous activation of dan outside mutantclones might reflect effects both of amplitude and ratios ofWg/Dpp signals emitted along the AP boundary that could
Fig. 7 The genetic “labial”program and the “ground state.”This figure summarizes theresults of the genetic dissection.Transformation of labial to an-tennal structures can be obtainedin several conditions: in pbhypomorphs, in pb Scr doublemutants, and in pb put doublemutants (Dpp-signaling defi-cient), or in pb Scr hth putquadruple mutants. Legs withdiffering degrees of articulationare obtained on removing pb, pbScr hth, or pb ss, regardless ofthe status of Scr, hth, and put.The genetic default state can beattained in pb ss double mutants,and corresponds to a rudimen-tary claw-bearing “ground state”leg
440

potentially be relayed and diversified by other diffusiblesignal(s).
Our data support a fundamental role for the joint actionof selector transcription factors and cell signaling pathwaysin orienting and coordinating the differentiation of a groupof cells. In contrast to Drosophila where imaginal discsprefiguring the appendages are already separate entities,cell signaling pathways that create a community effectmaintaining the unity of a group of cells may be of primaryimportance in a nonholometabolous insect such as thegrasshopper Schistocerca where appendages develop fromgroups of cells that are not spatially delimited. Thus, wepropose that a crucial developmental role of the gene classcalled selectors—all encoding transcription factors—is tochannel, integrate, and finely regulate available signalstoward stable, usable outcomes.
Acknowledgements We thank our colleagues in Toulouse formany helpful discussions and Fernando Roch and CorinneBenassayag for their critical readings of versions of the manuscript.The monoclonal antibodies directed against Wg (4D4, developed byS. Cohen) and Dac proteins (developed by G. M. Rubin) used in thiswork were obtained from the Developmental Studies HybridomaBank developed under the auspices of the National Institute of ChildHealth and Human Development and maintained by The Universityof Iowa, Department of Biological Sciences, Iowa City, IA 52242,USA. Mouse anti-Dll, the plasmid sscA6 harboring a ss cDNA, andthe ssD115-7 allele were provided by Dr. I. Duncan; rabbit anti-Hthand the hth5E chromosome by A. Salzberg; rat anti-Dan by S. Cohen,FRT Scr1 and FRT pb27Scr2 by A. Percival-Smith. We thank AgnèsLepage for her expert technical assistance and Bruno Savelli andBrice Ronsin for their help with the confocal microscope. This workbenefited from the ongoing support of the Centre National deRecherche Scientifique (CNRS) and grants from the Associationpour la Recherche sur le Cancer (ARC). L. J. was supported bygraduate fellowships from the French Ministère de l’EducationNationale et de la Recherche and the Ligue Nationale Contre leCancer.
References
Abu-Shaar M, Mann RS (1998) Generation of multiple antagonisticdomains along the proximodistal axis during Drosophila legdevelopment. Development 125:3821–3830
Abzhanov A, Kaufman TC (2000) Homologs of Drosophilaappendage genes in the patterning of arthropod limbs. DevBiol 227:673–689
Abzhanov A, Holtzman S, Kaufman TC (2001) The Drosophilaproboscis is specified by two Hox genes, proboscipedia andSex combs reduced, via repression of leg and antennalappendage genes. Development 128:2803–2814
Angelini DR, Kaufman, TC (2005) Insect appendages andcomparative ontogenetics. Dev Biol 286:57–77
Basler K, Struhl G (1994) Compartment boundaries and the controlof Drosophila limb pattern by hedgehog protein. Nature368:208–214
Belenkaya TY, Han C, Standley HJ, Lin X, Houston DW, HeasmanJ, Lin X (2002) Pygopus encodes a nuclear protein essential forwingless/Wnt signaling. Development 129:4089–4101
Bridges CB, Dobzhansky T (1933) The mutant “proboscipedia” inDrosophila melanogaster—a case of heredity homoösis.Wilhelm Roux Arch Dev Biol 127:575–590
Brook WJ, Cohen SM (1996) Antagonistic interactions betweenwingless and decapentaplegic responsible for dorsal-ventralpattern in the Drosophila leg. Science 273:1373–1377
Brusca RC, Brusca GJ (2002) Invertebrates. Sinauer, Sunderland,Massachusetts
Burke R, Basler K (1996) Dpp receptors are autonomously requiredfor cell proliferation in the entire developing Drosophila wing.Development 122:2261–2269
Casares F, Mann RS (1998) Control of antennal versus legdevelopment in Drosophila. Nature 392:723–726
Casares F, Mann RS (2001) The ground state of the ventralappendage in Drosophila. Science 293:1477–1480
Chen Y, Knezevic V, Ervin V, Hutson R, Ward Y, Mackem S (2004)Direct interaction with Hoxd proteins reverses Gli3-repressorfunction to promote digit formation downstream of Shh.Development 131:2339–2347
Cohen S (1993) Imaginal disc development. In: Bate M, MartinezArias A (eds) The development of Drosophila melanogaster.Cold Spring Harbor Laboratory, Cold Spring Harbor, NewYork, p 1564
DiNardo S, Kuner JM, Theis J, O’Farrell PH (1985) Development ofembryonic pattern in D. melanogaster as revealed byaccumulation of the nuclear Engrailed protein. Cell 43:59–69
Dong PD, Chu J, Panganiban G (2000) Coexpression of thehomeobox genes Distal-less and homothorax determines Dro-sophila antennal identity. Development 127:209–216
Dong PD, Dicks JS, Panganiban G (2002) Distal-less andhomothorax regulate multiple targets to pattern the Drosophilaantenna. Development 129:1967–1974
Duncan DM, Burgess EA, Duncan I (1998) Control of distalantennal identity and tarsal development in Drosophila byspineless-aristapedia, a homolog of the mammalian dioxinreceptor. Genes Dev 12:1290–1303
Emerald BS, Roy JK (1997) Homeotic transformation in Drosoph-ila. Nature 389:684
Emerald BS, Cohen SM (2001) Limb development: getting down tothe ground state. Curr Biol 11:R1025–R1027
Emerald BS, Cohen SM (2004) Spatial and temporal regulation ofthe homeotic selector gene Antennapedia is required for theestablishment of leg identity in Drosophila. Dev Biol 267:462–472
Emerald BS, Curtiss J, Mlodzik M, Cohen SM (2003) Distalantenna and distal antenna related encode nuclear proteinscontaining pipsqueak motifs involved in antenna developmentin Drosophila. Development 130:1171–1180
Estrada B, Sanchez-Herrero E (2001) The Hox gene Abdominal-Bantagonizes appendage development in the genital disc ofDrosophila. Development 128:331–339
Gebelein B, McKay DJ, Mann RS (2004) Direct integration of Hoxand segmentation gene inputs during Drosophila development.Nature 431:653–659
Gilbert SF, Opitz JM, Raff RA (1996) Resynthesizing evolutionaryand developmental biology. Dev Biol 173:357–372
Gurdon JB (1988) A community effect in animal development.Nature 336:772–774
Heemskerk J, DiNardo S (1994) Drosophila hedgehog acts as amorphogen in cellular patterning. Cell 76:449–460
Hersh BM, Carroll SB (2005) Direct regulation of knot geneexpression by Ultrabithorax and the evolution of cis-regulatoryelements in Drosophila. Development 132:1567–1577
Hughes CL, Kaufman TC (2000) RNAi analysis of Deformed,proboscipedia and Sex combs reduced in the milkweed bugOncopeltus fasciatus: novel roles for Hox genes in thehemipteran head. Development 127:3683–3694
Hughes CL, Kaufman TC (2002) Hox genes and the evolution of thearthropod body plan. Evol Dev 4:459–499
Inoue Y, Mito T, Miyawaki K, Matsushima K, Shinmyo Y, HeanueTA, Mardon G, Ohuchi H, Noji S (2002) Correlation ofexpression patterns of homothorax, dachshund and Distal-lesswith the proximodistal segmentation of the cricket leg bud.Mech Dev 113:141–148
Jiang J, Struhl G (1996) Complementary and mutually exclusiveactivities of decapentaplegic and wingless organize axialpatterning duringDrosophila leg development. Cell 86:401–409
441

Johnston LA, Schubiger G (1996) Ectopic expression of wingless inimaginal discs interferes with decapentaplegic expression andalters cell determination. Development 122:3519–3529
Joulia L, Bourbon HM, Cribbs DL (2005) Homeotic proboscipediafunction modulates hedgehog-mediated organizer activity topattern adult Drosophila mouthparts. Dev Biol 278:496–510
Kaufman TC (1978) Cytogenetic analysis of chromosome 3 inDrosophila melanogaster: isolation and characterization of fournew alleles of the proboscipedia (pb) locus. Genetics 90:579–596
Kaufman TC, Seeger MA, Olsen G (1990) Molecular and geneticorganization of the Antennapedia gene complex of Drosophilamelanogaster. Adv Genet 27:309–362
Kojima T (2004) The mechanism of Drosophila leg developmentalong the proximodistal axis. Dev Growth Differ 46:115–129
Kramps T, Peter O, Brunner E, Nellen D, Froesch B, Chatterjee S,Murone M, Zullig S, Basler K (2002) Wnt/wingless signalingrequires BCL9/legless-mediated recruitment of pygopus to thenuclear beta-catenin-TCF complex. Cell 109:47–60
Kumar K, Ouweneel WJ, Faber J (1979) Differentiation capacitiesof the labial imaginal disc of Drosophila melanogaster.Wilhelm Roux Arch Dev Biol 186:51–64
Lecuit T, Cohen SM (1997) Proximal-distal axis formation in theDrosophila leg. Nature 388:139–145
Lee JJ, von Kessler DP, Parks S, Beachy PA (1992) Secretion andlocalized transcription suggest a role in positional signaling forproducts of the segmentation gene hedgehog. Cell 71:33–50
Letsou A, Arora K, Wrana JL, Simin K, Twombly V, Jamal J,Staehling-Hampton K, Hoffmann FM, Gelbart WM, MassagueJ et al (1995) Drosophila Dpp signaling is mediated by the puntgene product: a dual ligand-binding type II receptor of the TGFbeta receptor family. Cell 80:899–908
Maves L, Schubiger G (1995) Wingless induces transdeterminationin developing Drosophila imaginal discs. Development121:1263–1272
McGinnis W, Krumlauf R (1992) Homeobox genes and axialpatterning. Cell 68:283–302
Morata G, Ripoll P (1975) Minutes: mutants of Drosophilaautonomously affecting cell division rate. Dev Biol 42:211–221
Newsome TP, Asling B, Dickson BJ (2000) Analysis of Drosophilaphotoreceptor axon guidance in eye-specific mosaics. Devel-opment 127:851–860
Nielsen C (1998) The phylogenetic position of the Arthropoda. In:Fortey RA, Thomas RH (eds) Arthropod relationships. Chap-man & Hall, London, pp 11–22
Parker DS, Jemison J, Cadigan KM (2002) Pygopus, a nuclearPHD-finger protein required for Wingless signaling in Dro-sophila. Development 129:2565–2576
Pattatucci AM, Kaufman TC (1991) The homeotic gene Sex combsreduced of Drosophila melanogaster is differentially regulatedin the embryonic and imaginal stages of development. Genetics129:443–461
Penton A, Hoffmann FM (1996) Decapentaplegic restricts thedomain of wingless during Drosophila limb patterning. Nature382:162–164
Percival-Smith A, Weber J, Gilfoyle E, Wilson P (1997) Geneticcharacterization of the role of the two HOX proteins,Proboscipedia and Sex Combs Reduced, in determination ofadult antennal, tarsal, maxillary palp and proboscis identities inDrosophila melanogaster. Development 124:5049–5062
Prpic NM, Tautz D (2003) The expression of the proximodistal axispatterning genes Distal-less and dachshund in the appendagesof Glomeris marginata (Myriapoda: Diplopoda) suggests aspecial role of these genes in patterning the head appendages.Dev Biol 260:97–112
Prpic NM, Wigand B, Damen WG, Klingler M (2001) Expression ofdachshund in wild-type and Distal-less mutant Triboliumcorroborates serial homologies in insect appendages. DevGenes Evol 211:467–477
Raff RA (1987) Constraint, flexibility, and phylogenetic history inthe evolution of direct development in sea urchins. Dev Biol119:6–19
Ruberte E, Marty T, Nellen D, Affolter M, Basler K (1995) Anabsolute requirement for both the type II and type I receptors,punt and thick veins, for dpp signaling in vivo. Cell 80:889–897
Simin K, Bates EA, Horner MA, Letsou A (1998) Genetic analysisof punt, a type II Dpp receptor that functions throughout theDrosophila melanogaster life cycle. Genetics 148:801–813
Struhl G (1982) Genes controlling segmental specification in theDrosophila thorax. Proc Natl Acad Sci USA 79:7380–7384
Suzanne M, Estella C, Calleja M, Sanchez-Herrero E (2003) Thehernandez and fernandez genes of Drosophila specify eye andantenna. Dev Biol 260:465–483
Tabata T, Kornberg TB (1994) Hedgehog is a signaling protein witha key role in patterning Drosophila imaginal discs. Cell 76:89–102
Tabata T, Eaton S, Kornberg TB (1992) The Drosophila hedgehoggene is expressed specifically in posterior compartment cellsand is a target of engrailed regulation. Genes Dev 6:2635–2645
Thompson BJ (2004) A complex of Armadillo, Legless, andPygopus coactivates dTCF to activate wingless target genes.Curr Biol 14:458–466
Thompson B, Townsley F, Rosin-Arbesfeld R, Musisi H, Bienz M(2002) A new nuclear component of the Wnt signallingpathway. Nat Cell Biol 4:367–373
Townsley FM, Thompson B, Bienz M (2004) Pygopus residuesrequired for its binding to Legless are critical for transcriptionand development. J Biol Chem 279:5177–5183
Vachon G, Cohen B, Pfeifle C, McGuffin ME, Botas J, Cohen SM(1992) Homeotic genes of the Bithorax—complex repress limbdevelopment in the abdomen of the Drosophila embryo throughthe target gene Distal-less. Cell 71:437–450
Yao LC, Liaw GJ, Pai CY, Sun YH (1999) A common mechanismfor antenna-to-leg transformation in Drosophila: suppression ofhomothorax transcription by four HOM-C genes. Dev Biol211:268–276
Zakany J, Kmita M, Duboule D (2004) A dual role for Hox genes inlimb anterior-posterior asymmetry. Science 304:1669–1672
Zecca M, Basler K, Struhl G (1995) Sequential organizing activitiesof engrailed, hedgehog and decapentaplegic in the Drosophilawing. Development 121:2265–2278
442
Copyright © 2022 FDOKUMEN


















