
CHOXC-8 andCHOXD-13 expression in embryonic chick skin and cutaneous appendage specification
14
CHOXC-8 and CHOXD-13 Expression in Embryonic Chick Skin and Cutaneous Appendage Specification BENOI ˆ T KANZLER, FABRICE PRIN, JACQUES THELU, AND DANIELLE DHOUAILLY* Biologie de la Diffe ´renciation Epithe ´liale-UMR CNRS 5538, Etude de la Diffe ´renciation et de l’Adhe ´rence Cellulaires, Institut Albert Bonniot, Universite ´ Joseph Fourier, Grenoble, France ABSTRACT We studied the expression of two distantly clustered Hox genes which could, respectively, be involved in specification of dor- sal feather- and foot scale-forming skin in the chick embryo: cHoxc-8, a median paralog, and cHoxd-13, located at the 58 extremity of the HoxD cluster. The cHoxc-8 transcripts are present at embryonic day 3.5 (E3.5)in the somitic cells, which give rise to the dorsal dermis by E5, and at E6.5–8.5 in the dorsal dermal and epidermal cells during the first stages of feather morphogenesis. The cHoxd-13 transcripts are present at E4.5–9.5 in the autopodial mesenchyme and at E10.5–12.5 in the plantar dermis during the initiation of reticulate scale morphogenesis. Both the cHoxc-8 and cHoxd-13 transcripts are no longer detect- able after the anlagen stage of cutaneous append- age morphogenesis. Furthermore, heterotopic dermal–epidermal recombinations of dorsal, plan- tar, and apteric tissues revealed that the epider- mal ability or inability to form feathers is already established by the time of skin formation. Reti- noic acid (RA) treatment at E11 induces after 12 hr an inhibition of cHoxd-13 expression in the plantar dermis, followed by the formation of feather filaments on the reticulate scales. When E7.5 dorsal explants are treated with RA for 6 days, they form scale-like structures where the Hox transcripts are no more detectable. Protein analysis revealed that the plantar filaments, made up of feather b-keratins, corresponded to a homeo- tic transformation, whereas the scale-like struc- tures, composed also of feather b-keratins, were teratoid. These results strengthen the hypothesis that different homeobox genes play a significant role in specifying the regional identity of the differ- ent epidermal territories. Dev. Dyn. 1997;210: 274–287. r 1997 Wiley-Liss, Inc. Key words: chick; epidermis; feather; homeobox genes; keratin; retinoic acid; scale INTRODUCTION The vertebrate skin provides a powerful model sys- tem to study cellular interactions which govern organ morphogenesis because of its distinct developmental pathways and the heterogeneity of its appendages. Furthermore, during integument development, epider- mal keratinocytes synthetize specific keratin polypep- tides according to their regional origin, thus allowing the molecular identification of the protein content inherent to a given morphology. The a-keratins range from 40 to 73 kD in molecular weight and are expressed in all epithelial cells from fish to human (Moll et al., 1982). The expression of b-keratin polypeptides, which range from 10 to 25 kD, is restricted to reptiles and birds and characterizes such terminally differentiated tissues as scales, claws, beak, and feathers (Gregg et al., 1984). Feathers, which characterize avian skin, present some differences in their morphology and distri- bution pattern according to the body regions. Further- more, in the chick, the type of appendage changes from zeugopodial feathers to autopodial scales in the hind- limb. The scales themselves display two main subtypes: the scuta, which are large overlapping scales from the anterior tarsometatarsal region, and the reticula, which are small and roundish scales covering the plantar surface of the foot. Their structural protein content characterizes these three types of appendages: Feath- ers express lighter b-keratin polypeptides than scutate scales, while the reticulate epidermis is composed mostly of a-keratins (Kemp and Rogers, 1972; Dhou- ailly et al., 1978; Haake et al., 1984; Sawyer, 1983). These cutaneous appendages develop as a result of successive interactions between the two tissues compris- ing the skin, an epithelium, the epidermis, and an underlying mesenchyme, the dermis; their regional diversity depends both on the inductive properties of the dermis and on the competence of the epidermis (Sengel et al., 1969; Sengel, 1976; Dhouailly and Sen- gel, 1983, Cadi et al.; 1983; Dhouailly, 1977, 1984; Sawyer, 1983; Viallet and Dhouailly, 1994). The skin regionalization specification could involve transcription factors belonging to the homeoprotein family. Vertebrate homeobox genes, which are homolo- gous to the first identified genes of the Drosophila antennapedia and bithorax complexes are organized into four clusters, HoxA, B, C, and D, located on different chromosomes (for reviews, see MacGinnis and Grant sponsor: ARC; Grant number: 6233; Grant sponsor: Fonda- tion de la Recherche Me ´dacale. *Correspondence to: D. Dhouailly, Institut Albert Bonniot, LEDAC UMR CNRS 5538-Biologie de la Diffe ´renciation Epithe ´liale, Domaine de la Merci, 38706 La Tronche Cedex, France. E-mail: danielle. [email protected] Received 18 February 1997; Accepted 1 August 1997 DEVELOPMENTAL DYNAMICS 210:274–287 (1997) r 1997 WILEY-LISS, INC.
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of CHOXC-8 andCHOXD-13 expression in embryonic chick skin and cutaneous appendage specification

CHOXC-8 and CHOXD-13 Expression in Embryonic ChickSkin and Cutaneous Appendage SpecificationBENOIT KANZLER, FABRICE PRIN, JACQUES THELU, AND DANIELLE DHOUAILLY*Biologie de la Differenciation Epitheliale-UMR CNRS 5538, Etude de la Differenciation et de l’Adherence Cellulaires,Institut Albert Bonniot, Universite Joseph Fourier, Grenoble, France
ABSTRACT We studied the expression oftwo distantly clustered Hox genes which could,respectively, be involved in specification of dor-sal feather- and foot scale-forming skin in thechick embryo: cHoxc-8, a median paralog, andcHoxd-13, located at the 58 extremity of the HoxDcluster. The cHoxc-8 transcripts are present atembryonic day 3.5 (E3.5)in the somitic cells, whichgive rise to the dorsal dermis by E5, and atE6.5–8.5 in the dorsal dermal and epidermal cellsduring the first stages of feather morphogenesis.The cHoxd-13 transcripts are present at E4.5–9.5in the autopodial mesenchyme and at E10.5–12.5in the plantar dermis during the initiation ofreticulate scale morphogenesis. Both the cHoxc-8and cHoxd-13 transcripts are no longer detect-able after the anlagen stage of cutaneous append-age morphogenesis. Furthermore, heterotopicdermal–epidermal recombinations of dorsal, plan-tar, and apteric tissues revealed that the epider-mal ability or inability to form feathers is alreadyestablished by the time of skin formation. Reti-noic acid (RA) treatment at E11 induces after 12hr an inhibition of cHoxd-13 expression in theplantar dermis, followed by the formation offeather filaments on the reticulate scales. WhenE7.5 dorsal explants are treated with RA for 6days, they form scale-like structures where theHox transcripts are no more detectable. Proteinanalysis revealed that the plantar filaments, madeup of feather b-keratins, corresponded to a homeo-tic transformation, whereas the scale-like struc-tures, composed also of feather b-keratins, wereteratoid. These results strengthen the hypothesisthat different homeobox genes play a significantrole in specifying the regional identity of the differ-ent epidermal territories. Dev. Dyn. 1997;210:274–287. r 1997 Wiley-Liss, Inc.
Key words: chick; epidermis; feather; homeoboxgenes; keratin; retinoic acid; scale
INTRODUCTION
The vertebrate skin provides a powerful model sys-tem to study cellular interactions which govern organmorphogenesis because of its distinct developmentalpathways and the heterogeneity of its appendages.Furthermore, during integument development, epider-
mal keratinocytes synthetize specific keratin polypep-tides according to their regional origin, thus allowingthe molecular identification of the protein contentinherent to a given morphology. The a-keratins rangefrom 40 to 73 kD in molecular weight and are expressedin all epithelial cells from fish to human (Moll et al.,1982). The expression of b-keratin polypeptides, whichrange from 10 to 25 kD, is restricted to reptiles andbirds and characterizes such terminally differentiatedtissues as scales, claws, beak, and feathers (Gregg etal., 1984). Feathers, which characterize avian skin,present some differences in their morphology and distri-bution pattern according to the body regions. Further-more, in the chick, the type of appendage changes fromzeugopodial feathers to autopodial scales in the hind-limb. The scales themselves display two main subtypes:the scuta, which are large overlapping scales from theanterior tarsometatarsal region, and the reticula, whichare small and roundish scales covering the plantarsurface of the foot. Their structural protein contentcharacterizes these three types of appendages: Feath-ers express lighter b-keratin polypeptides than scutatescales, while the reticulate epidermis is composedmostly of a-keratins (Kemp and Rogers, 1972; Dhou-ailly et al., 1978; Haake et al., 1984; Sawyer, 1983).These cutaneous appendages develop as a result ofsuccessive interactions between the two tissues compris-ing the skin, an epithelium, the epidermis, and anunderlying mesenchyme, the dermis; their regionaldiversity depends both on the inductive properties ofthe dermis and on the competence of the epidermis(Sengel et al., 1969; Sengel, 1976; Dhouailly and Sen-gel, 1983, Cadi et al.; 1983; Dhouailly, 1977, 1984;Sawyer, 1983; Viallet and Dhouailly, 1994).
The skin regionalization specification could involvetranscription factors belonging to the homeoproteinfamily. Vertebrate homeobox genes, which are homolo-gous to the first identified genes of the Drosophilaantennapedia and bithorax complexes are organizedinto four clusters, HoxA, B, C, and D, located ondifferent chromosomes (for reviews, see MacGinnis and
Grant sponsor: ARC; Grant number: 6233; Grant sponsor: Fonda-tion de la Recherche Medacale.
*Correspondence to: D. Dhouailly, Institut Albert Bonniot, LEDACUMR CNRS 5538-Biologie de la Differenciation Epitheliale, Domainede la Merci, 38706 La Tronche Cedex, France. E-mail: [email protected]
Received 18 February 1997; Accepted 1 August 1997
DEVELOPMENTAL DYNAMICS 210:274–287 (1997)
r 1997 WILEY-LISS, INC.

Krumlauf, 1992; Krumlauf, 1994). Within each cluster,genes most similar in sequence to a particular Dro-sophila gene, or paralogous genes, occupy the samerelative positions within their respective cluster. Inaddition, the rostro-caudal boundary of one given Hoxexpression domain correlates with the position of thegene within its cluster, a property termed colinearity(Duboule and Dolle, 1989; Duboule and Morata, 1994).The same Hox sets have been shown to regulate thedevelopmental processes within antero-posterior andproximo-distal specification in patterning of the verte-brae and limb segments (among others: Kessel andGruss, 1991; Duboule, 1992). Previous results haveshown that the cHoxc-6 and cHoxd-4 homeoproteins aredifferentially expressed during dorsal chick skin forma-tion (Chuong et al., 1990, 1993). Likewise, in mouse,two different homeobox gene families, namely Otx andHox, identify the facial and body skin territories,respectively, (Kanzler et al., 1994).
In order to investigate the putative involvement ofHox genes in providing positional information for skincells, and consequently in specification of chick feather-versus scale-forming skin, we chose to study the expres-sion of a median paralog, of order 8, which may beexpressed at its highest level in the middorsal skin andthat of the most distal one, of order 13, which similarlycould be expressed in foot skin. A chick Hoxd-13 probewas provided to us by D. Duboule (Izpisua-Belmonte etal., 1991). In order to obtain a chick Hox gene belongingto the 8th paralog, we screened a E8.5 dorsal skin cDNAlibrary (Michaille et al., 1994) using a mouse Hoxc-8probe obtained from H. Le Mouellic (Le Mouellic et al.,1992) and consequently isolated and characterized thechick Hoxc-8 cDNA. The developmental expressionpattern of these two genes, analyzed by in situ hybrid-ization, suggests that the cHoxc-8 and cHoxd-13 pro-teins play a role in the specification of dorsal andplantar skin morphogenesis. To further define theinvolvement of both genes, we carried out two comple-mentary types of experiments. First, dermal–epider-mal recombinants were performed between chick dor-sal and plantar skin, at stages during which Hoxc-8 andHoxd-13 are expressed in skin and which correspond tothe initiation of appendage morphogenesis. To furthertest the morphogenetic abilities of the plantar dermis,some of the recombinants involved apteric midventralepidermis. Second, as it is well known that retinoic acid(RA) is both able to modulate homeobox gene expres-sion (Simeone et al., 1991; Boncinelli et al., 1991) and topromote the ectopic formation of feathers on chick feet(Dhouailly et al., 1980) and scale-like structures indorsal skin (Chuong et al., 1992), we repeated these twotypes of RA treatment in order to analyze the possiblecorrelative changes in cHoxc-8 and cHoxd-13 expression.
RESULTSOverview of Chick Tegument Morphogenesis
In the chick, while most of the body is covered withfeathers, the feet bear scales, and some areas, referred
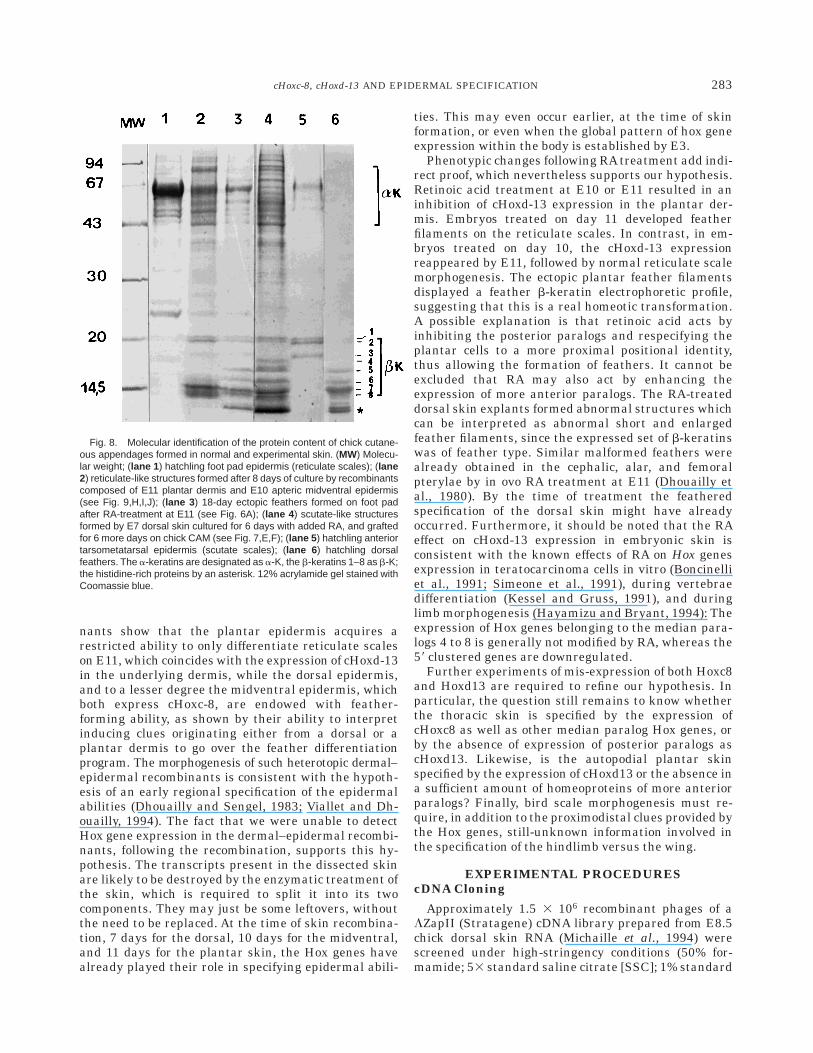
to as apteria, remain bare. All the epidermis, with theexception of the median facial epidermis (Couly and LeDouarin, 1988), originates from the ectoderm, whereasthe origin of the dermis varies according to the differentregions. The back dermis is formed by E5 by themigration of somitic dermatomal cells (Mauger, 1972),whereas the limb dermis is formed at E8.5 by mesoder-mal cells which originate from the somatopleure. Thefirst feather primordia (Fig. 1; stage f1 as referred byMichaille et al., 1994) appear at E7 along the midline ofthe lumbar region and consist of a circular epidermalthickening, or placode, which then becomes associatedwith an underlying dermal condensation (Sengel, 1976;Dhouailly, 1984). The epidermal and dermal cells thenproliferate to form the feather bud (stage f3) by E8.5,which slants backward. The feather bud then elongatesand invaginates at its base to form the feather filament(stage f7) by E14.5. Feather keratinization (a-keratinsin the outer epidermal sheath and b-keratins in thefeather itself) is first detectable at E12.5 at the apex ofthe filament and then proceeds downward (Haake etal., 1984). For a better comparison with the correspond-ing stages of feather morphogenesis, the three mainstages of scuta and reticula formation were calledstages 1, 3, and 7, respectively (Fig. 1). The firstindication of scutate scale formation in the anteriortarsometatarsal region is the appearance of oval plac-odes (stage s1) during E9. The scuta rudiments arecomposed of an epidermal thickening covering a lightlymarked dermal condensation. By E12, the placodes giverise to the scale buds (stage s3), then to the scutatescales (stage s7), which express a- and b-keratins byE16. Scutate scale b-keratins (Fig. 8, lane 5) involvethree major polypeptides (21 to 18 kD), numberedb1–b3. The polypeptides 1 to 3 are also present butbarely detectable in feather, whereas feather specificb-keratins (Fig. 8, lane 6) include a minor (b4) and fourmajor (b5–8) 16- to 10-kD polypeptides. The first step ofreticula formation (stage r1) occurs in the center of theplantar region by E11.5 and is characterized by anepidermal dome without any associated dermal conden-sation. Reticulate scale differentiation reaches stage r7by E16. These structures express a-keratins (Fig. 8,lane 1) with the exception of the embryonic peridermaland subperidermal shedding layers which express a25-kD b-keratin (Sawyer and Borg, 1979; Knapp et al.,1993).
cHoxc-8 cDNA Cloning
Three distinct positive clones were obtained by screen-ing the chick skin cDNA library under high-stringencyhybridization conditions with the mouse Hoxc-8 cDNAprobe, and their sequences were determined. The long-est cDNA clone (Fig. 2A) is 2,800 bp in length andencompasses the complete open reading frame ofcHoxc-8, as revealed by sequence comparison with theknown Hox genes belonging to the 8th paralog. Thepartial sequence of this cDNA (Fig. 2B) has beensubmitted to the EMBL/GenBank Data Libraries and
275cHoxc-8, cHoxd-13 AND EPIDERMAL SPECIFICATION

received the accession number X94179. The predictedcorresponding protein contains 242 amino acids andexhibits a Mr of 28,200 kD. Sequence comparison withthe human (Boncinelli et al., 1989) and mouse (LeMouellic et al., 1988) Hoxc-8 amino acid sequencesreveal a complete interspecies conservation of the ho-meodomain; the chick sequence (Fig. 2B) is 100%identical with its human and mouse homologues. Twoother regions of the predicted cHoxc-8 protein arestrongly conserved when compared with the consensussequence, namely a hexapeptide (MYPWMK) locatedfive amino acids upstream of the homeodomain and apeptide located at the N-terminal end. The YPWMmotif has been shown to be essential for establishingcooperative interactions between a subset of Hox pro-teins and Pbx proteins (Chang et al., 1995). The N-terminal region contains a high number of hydrophilicresidues, particularly serine and threonine. As reportedfor the mouse amino-acid sequence, the carboxy-terminal region comprises an acidic region that isparticularly rich in glutamate residues (15 out of 21amino acids). Such a region has been previously identi-fied in the Hoxa-7 gene, where 15 glutamate codonspreceed a stop codon (Colberg-Poley et al., 1985).
Expression of the cHoxc-8 and cHoxd-13 GenesDuring Chick Skin Morphogenesis
Using in situ hybridization, we studied the spatiotem-poral distribution of cHoxc-8 and cHoxd-13 transcripts
in 3.5-, 4.5-, and 6.5-day whole embryos, as well as in8.5- to 10.5-day whole hindlimbs. The tissue distribu-tion of the cHoc-8 and cHoxd-13 transcripts werestudied on serial sagittal sections of 6.5- to 14.5-dayembryos and of 9.5- to 16-day hindlimb, that is duringdorsal and plantar skin morphogenesis. Antisense RNAprobes, labeled either with digoxygenin-UTP or 35S-CTP, were synthesized from the 38 cDNA flankingregion of the cHoxc-8 homeobox, as well as from thesimilar region of the cHoxd-13 cDNA.
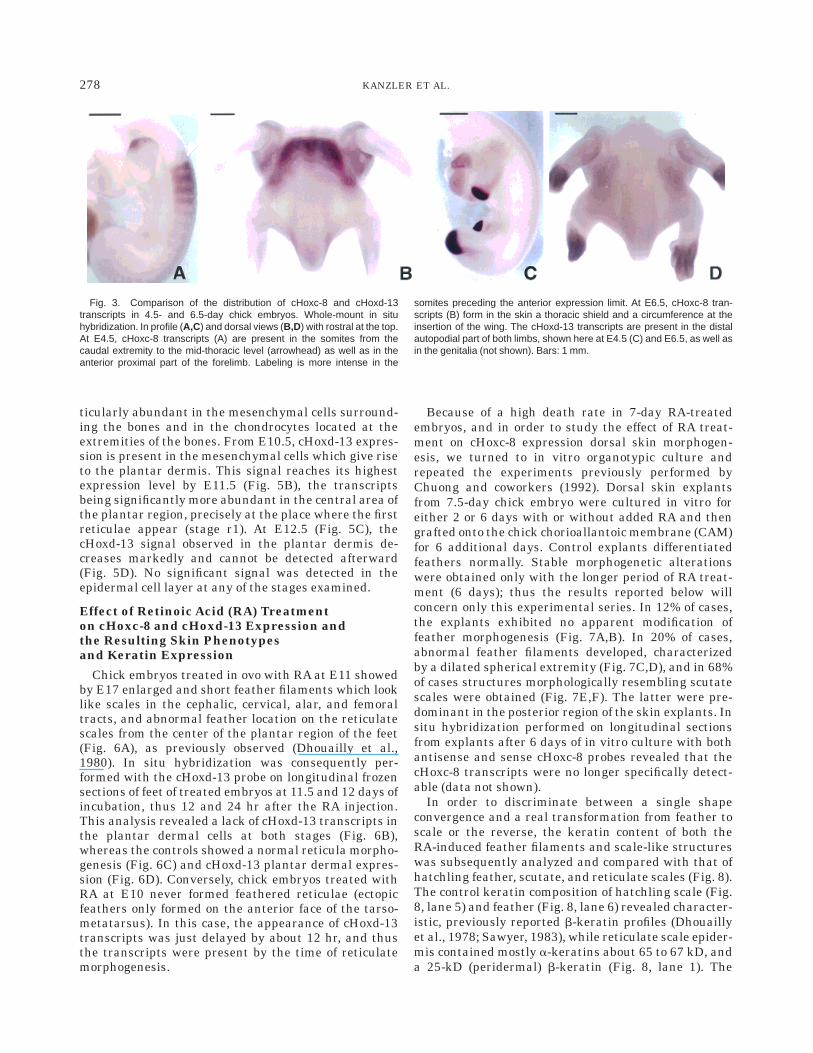
Whole-mount in situ hybridizations performed atE3.5 and at E4.5 (Fig. 3A) show that cHoxc-8 tran-scripts are present in the neural tube, in the mesoder-maly derived cells of somitic origin from the midtho-racic level to the caudal extremity, as well as in theantero-proximal region of the wing bud, and morelightly of that of the hindlimb bud. This gene displaysan anterior expression limit in the somites 23 and 24, alevel which is posterior to that observed in the neuraltube. These two somites later give rise to the fifththoracic vertebra and to the overlying thoracic dermis.By 6.5 days of incubation (Figs. 3B, 4A), the chick skin(stage f0) has formed in the dorsal region and thecHoxc-8 transcripts are detectable at the same level inthe vertebrae and the overlying mesenchymal, dermal,and epidermal cells. This expression reaches anteriorlyto the fifth thoracic vertebra, thus appearing colinearwith the limit observed earlier in the somites. ThecHoxc-8 signal is significantly more intense in thedorsal skin region corresponding to its anterior expres-sion boundary and decreases toward the caudal region.This dorsal antero-posterior pattern is conserved atE7.5 (stage f1), where transcripts are present in thedorsal feather primordia (Fig. 4B,C), both in the dermalcondensation and the epidermal placodes. By E8.5, theepidermal and dermal signal remain uniformly distrib-uted in the feather bud (stage f3) (Fig. 4E). This signalis also present in the mid-thoracic and at a less degreein the abdominal ventral skin, comprising the midven-tral apterium. At stage f7 (E14.5), the cHoxc-8 tran-scripts are no longer present in the interfollicular skinor in the feather follicle (Fig. 3G). No specific signal wasdetected with the corresponding sense probes on con-secutive sections (Fig. 4D,F,H). No expression of cHoxc-8was found with the antisense probe in the head andanterior thoracic skin regions at any of the studiedstages.
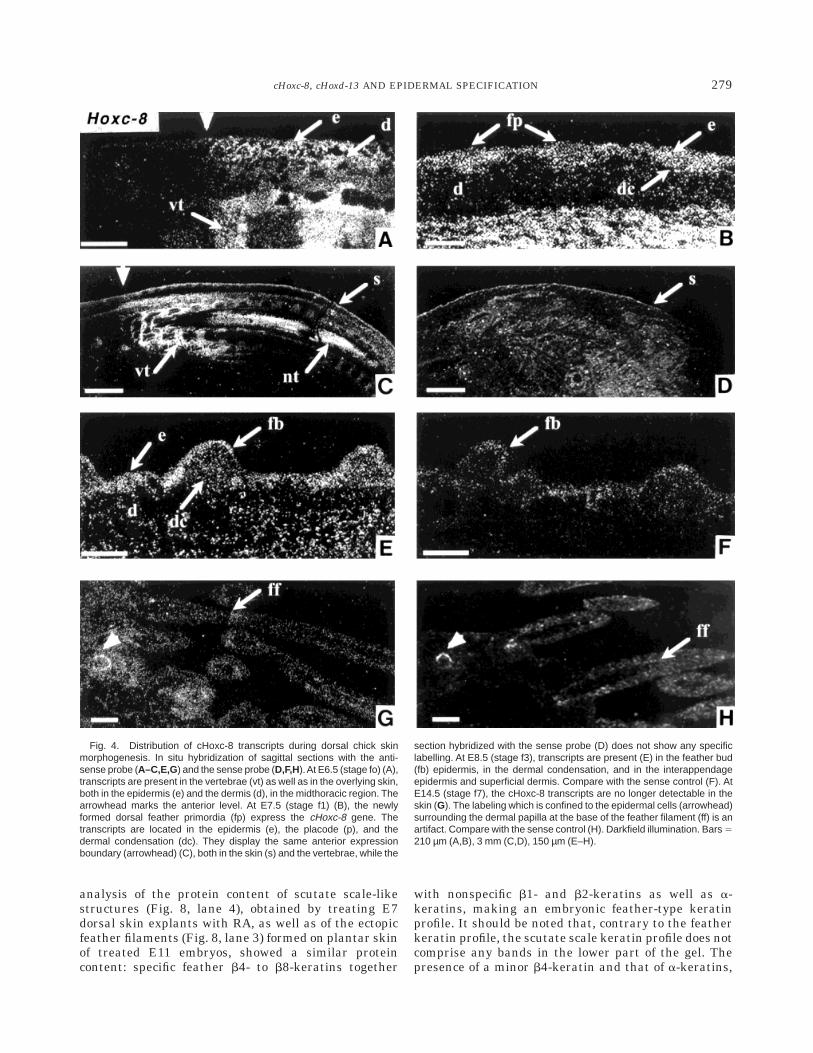
At E3.5, the cHoxd-13 transcripts were present insomites 39–40 at the distal end of the trunk (data notshown). They were never detected in either the dermisor the epidermis during trunk skin morphogenesis, aresult which is confirmed by their presence in thelombar spinal cord and caudal vertebrae in the samesections. At E4.5 and E6.5 (Fig. 3C,D) the cHoxd-13transcripts were detected in both the wing and hind-limb autopodes and then at E9.5 (Fig. 5A) in the distalregion of the feet, including the digits. On sagittalsections, it appears that these transcripts display anantero-posterior to proximo-distal gradient, being par-
Fig. 1. Main stages of chick feather (A), anterior tarsometatarsal scale(scuta) (B), and plantar scale (reticula) morphogenesis (C) (see text). br,barb ridge; d, dermis; dc, dermal condensation; dp, dermal pulp; e,epidermis; ec epidermal collar; ep, epidermal placode; fb, feather bud; ff,feather filament; ies, inner epithelial sheath; oes, outer epithelial sheath;p, papilla; r, reticula; rb, reticulate scale bud; s, scuta; sb, scutate scalebud.
276 KANZLER ET AL.

Fig. 2. A: Organization of the cHoxc-8 cDNA clone. The isolatedcDNA contains the complete cHoxc-8 coding region (hatched box; nt439–1168). The black box demarcates the position of the homeodomain(HD; nt 883–1063). Nucleotide positions of several restriction sites areindicated. E, EcoRI; P, PstI; K, KpnI. B: Partial nucleotide sequence of thechick cHoxc-8 cDNA and predicted amino acid sequence of the codingregion. The arrow points to the 38 end of the cDNA clone. Amino acid andnucleotide positions are indicated on the left and right sides, respectively.The sequence of the putative cHoxc-8 protein is indicated below the open
reading frame. Amino acid changes between the chick and mouse proteinsequences are underlined. Three consensus regions, i.e., the N-terminus,the conserved hexapeptide, as well as the homeodomain (HD) are boxed.The position of the intron as deduced from the mouse Hoxc-8 cDNAsequence is indicated by a black arrowhead. The splicing site (ACG/CT)fits well with the consensus sequence (Cech, 1983). These sequencedata are available from EMBL/GenBank/DDBJ sequence database underthe accession number X94179.
ticularly abundant in the mesenchymal cells surround-ing the bones and in the chondrocytes located at theextremities of the bones. From E10.5, cHoxd-13 expres-sion is present in the mesenchymal cells which give riseto the plantar dermis. This signal reaches its highestexpression level by E11.5 (Fig. 5B), the transcriptsbeing significantly more abundant in the central area ofthe plantar region, precisely at the place where the firstreticulae appear (stage r1). At E12.5 (Fig. 5C), thecHoxd-13 signal observed in the plantar dermis de-creases markedly and cannot be detected afterward(Fig. 5D). No significant signal was detected in theepidermal cell layer at any of the stages examined.
Effect of Retinoic Acid (RA) Treatmenton cHoxc-8 and cHoxd-13 Expression andthe Resulting Skin Phenotypesand Keratin Expression
Chick embryos treated in ovo with RA at E11 showedby E17 enlarged and short feather filaments which looklike scales in the cephalic, cervical, alar, and femoraltracts, and abnormal feather location on the reticulatescales from the center of the plantar region of the feet(Fig. 6A), as previously observed (Dhouailly et al.,1980). In situ hybridization was consequently per-formed with the cHoxd-13 probe on longitudinal frozensections of feet of treated embryos at 11.5 and 12 days ofincubation, thus 12 and 24 hr after the RA injection.This analysis revealed a lack of cHoxd-13 transcripts inthe plantar dermal cells at both stages (Fig. 6B),whereas the controls showed a normal reticula morpho-genesis (Fig. 6C) and cHoxd-13 plantar dermal expres-sion (Fig. 6D). Conversely, chick embryos treated withRA at E10 never formed feathered reticulae (ectopicfeathers only formed on the anterior face of the tarso-metatarsus). In this case, the appearance of cHoxd-13transcripts was just delayed by about 12 hr, and thusthe transcripts were present by the time of reticulatemorphogenesis.
Because of a high death rate in 7-day RA-treatedembryos, and in order to study the effect of RA treat-ment on cHoxc-8 expression dorsal skin morphogen-esis, we turned to in vitro organotypic culture andrepeated the experiments previously performed byChuong and coworkers (1992). Dorsal skin explantsfrom 7.5-day chick embryo were cultured in vitro foreither 2 or 6 days with or without added RA and thengrafted onto the chick chorioallantoic membrane (CAM)for 6 additional days. Control explants differentiatedfeathers normally. Stable morphogenetic alterationswere obtained only with the longer period of RA treat-ment (6 days); thus the results reported below willconcern only this experimental series. In 12% of cases,the explants exhibited no apparent modification offeather morphogenesis (Fig. 7A,B). In 20% of cases,abnormal feather filaments developed, characterizedby a dilated spherical extremity (Fig. 7C,D), and in 68%of cases structures morphologically resembling scutatescales were obtained (Fig. 7E,F). The latter were pre-dominant in the posterior region of the skin explants. Insitu hybridization performed on longitudinal sectionsfrom explants after 6 days of in vitro culture with bothantisense and sense cHoxc-8 probes revealed that thecHoxc-8 transcripts were no longer specifically detect-able (data not shown).
In order to discriminate between a single shapeconvergence and a real transformation from feather toscale or the reverse, the keratin content of both theRA-induced feather filaments and scale-like structureswas subsequently analyzed and compared with that ofhatchling feather, scutate, and reticulate scales (Fig. 8).The control keratin composition of hatchling scale (Fig.8, lane 5) and feather (Fig. 8, lane 6) revealed character-istic, previously reported b-keratin profiles (Dhouaillyet al., 1978; Sawyer, 1983), while reticulate scale epider-mis contained mostly a-keratins about 65 to 67 kD, anda 25-kD (peridermal) b-keratin (Fig. 8, lane 1). The
Fig. 3. Comparison of the distribution of cHoxc-8 and cHoxd-13transcripts in 4.5- and 6.5-day chick embryos. Whole-mount in situhybridization. In profile (A,C) and dorsal views (B,D) with rostral at the top.At E4.5, cHoxc-8 transcripts (A) are present in the somites from thecaudal extremity to the mid-thoracic level (arrowhead) as well as in theanterior proximal part of the forelimb. Labeling is more intense in the
somites preceding the anterior expression limit. At E6.5, cHoxc-8 tran-scripts (B) form in the skin a thoracic shield and a circumference at theinsertion of the wing. The cHoxd-13 transcripts are present in the distalautopodial part of both limbs, shown here at E4.5 (C) and E6.5, as well asin the genitalia (not shown). Bars: 1 mm.
278 KANZLER ET AL.
analysis of the protein content of scutate scale-likestructures (Fig. 8, lane 4), obtained by treating E7dorsal skin explants with RA, as well as of the ectopicfeather filaments (Fig. 8, lane 3) formed on plantar skinof treated E11 embryos, showed a similar proteincontent: specific feather b4- to b8-keratins together
with nonspecific b1- and b2-keratins as well as a-keratins, making an embryonic feather-type keratinprofile. It should be noted that, contrary to the featherkeratin profile, the scutate scale keratin profile does notcomprise any bands in the lower part of the gel. Thepresence of a minor b4-keratin and that of a-keratins,
Fig. 4. Distribution of cHoxc-8 transcripts during dorsal chick skinmorphogenesis. In situ hybridization of sagittal sections with the anti-sense probe (A–C,E,G) and the sense probe (D,F,H).At E6.5 (stage fo) (A),transcripts are present in the vertebrae (vt) as well as in the overlying skin,both in the epidermis (e) and the dermis (d), in the midthoracic region. Thearrowhead marks the anterior level. At E7.5 (stage f1) (B), the newlyformed dorsal feather primordia (fp) express the cHoxc-8 gene. Thetranscripts are located in the epidermis (e), the placode (p), and thedermal condensation (dc). They display the same anterior expressionboundary (arrowhead) (C), both in the skin (s) and the vertebrae, while the
section hybridized with the sense probe (D) does not show any specificlabelling. At E8.5 (stage f3), transcripts are present (E) in the feather bud(fb) epidermis, in the dermal condensation, and in the interappendageepidermis and superficial dermis. Compare with the sense control (F). AtE14.5 (stage f7), the cHoxc-8 transcripts are no longer detectable in theskin (G). The labeling which is confined to the epidermal cells (arrowhead)surrounding the dermal papilla at the base of the feather filament (ff) is anartifact. Compare with the sense control (H). Darkfield illumination. Bars 5210 µm (A,B), 3 mm (C,D), 150 µm (E–H).
279cHoxc-8, cHoxd-13 AND EPIDERMAL SPECIFICATION
the latter originating either from the interfollicularepidermis or the epidermal sheath of feather filaments,is usual when analyzing embryonic feathers or feath-ered skin explants.
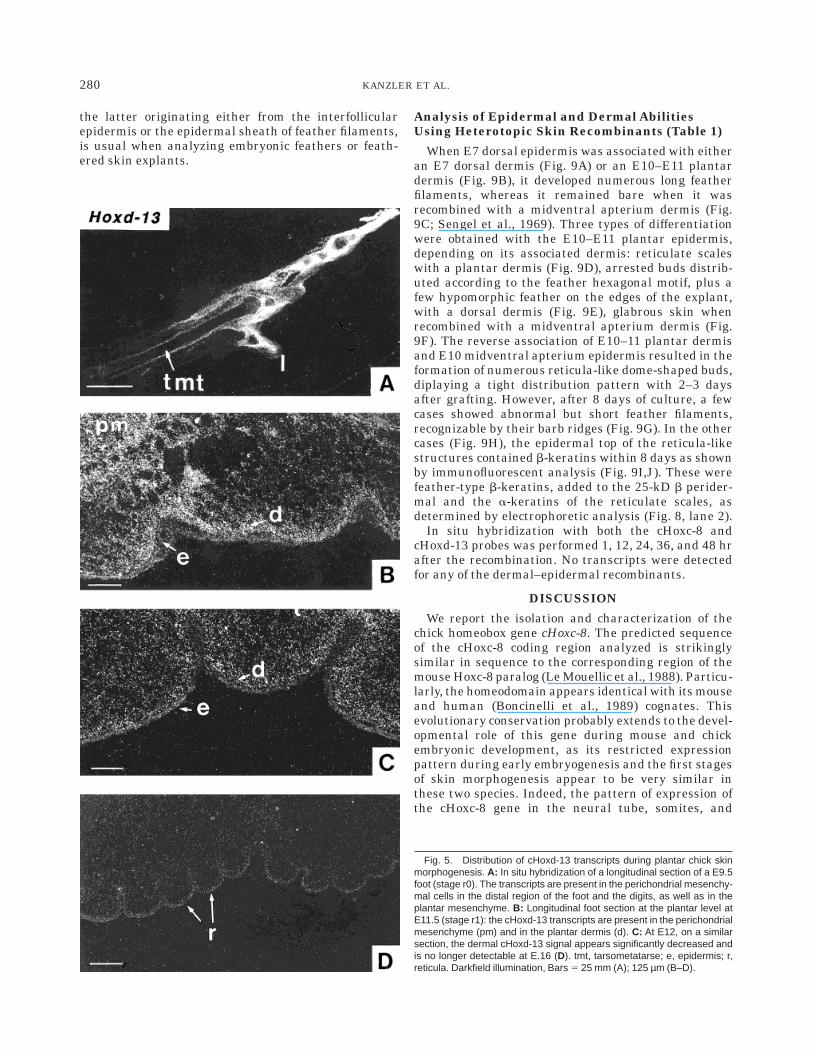
Analysis of Epidermal and Dermal AbilitiesUsing Heterotopic Skin Recombinants (Table 1)
When E7 dorsal epidermis was associated with eitheran E7 dorsal dermis (Fig. 9A) or an E10–E11 plantardermis (Fig. 9B), it developed numerous long featherfilaments, whereas it remained bare when it wasrecombined with a midventral apterium dermis (Fig.9C; Sengel et al., 1969). Three types of differentiationwere obtained with the E10–E11 plantar epidermis,depending on its associated dermis: reticulate scaleswith a plantar dermis (Fig. 9D), arrested buds distrib-uted according to the feather hexagonal motif, plus afew hypomorphic feather on the edges of the explant,with a dorsal dermis (Fig. 9E), glabrous skin whenrecombined with a midventral apterium dermis (Fig.9F). The reverse association of E10–11 plantar dermisand E10 midventral apterium epidermis resulted in theformation of numerous reticula-like dome-shaped buds,diplaying a tight distribution pattern with 2–3 daysafter grafting. However, after 8 days of culture, a fewcases showed abnormal but short feather filaments,recognizable by their barb ridges (Fig. 9G). In the othercases (Fig. 9H), the epidermal top of the reticula-likestructures contained b-keratins within 8 days as shownby immunofluorescent analysis (Fig. 9I,J). These werefeather-type b-keratins, added to the 25-kD b perider-mal and the a-keratins of the reticulate scales, asdetermined by electrophoretic analysis (Fig. 8, lane 2).
In situ hybridization with both the cHoxc-8 andcHoxd-13 probes was performed 1, 12, 24, 36, and 48 hrafter the recombination. No transcripts were detectedfor any of the dermal–epidermal recombinants.
DISCUSSION
We report the isolation and characterization of thechick homeobox gene cHoxc-8. The predicted sequenceof the cHoxc-8 coding region analyzed is strikinglysimilar in sequence to the corresponding region of themouse Hoxc-8 paralog (Le Mouellic et al., 1988). Particu-larly, the homeodomain appears identical with its mouseand human (Boncinelli et al., 1989) cognates. Thisevolutionary conservation probably extends to the devel-opmental role of this gene during mouse and chickembryonic development, as its restricted expressionpattern during early embryogenesis and the first stagesof skin morphogenesis appear to be very similar inthese two species. Indeed, the pattern of expression ofthe cHoxc-8 gene in the neural tube, somites, and
Fig. 5. Distribution of cHoxd-13 transcripts during plantar chick skinmorphogenesis. A: In situ hybridization of a longitudinal section of a E9.5foot (stage r0). The transcripts are present in the perichondrial mesenchy-mal cells in the distal region of the foot and the digits, as well as in theplantar mesenchyme. B: Longitudinal foot section at the plantar level atE11.5 (stage r1): the cHoxd-13 transcripts are present in the perichondrialmesenchyme (pm) and in the plantar dermis (d). C: At E12, on a similarsection, the dermal cHoxd-13 signal appears significantly decreased andis no longer detectable at E.16 (D). tmt, tarsometatarse; e, epidermis; r,reticula. Darkfield illumination, Bars 5 25 mm (A); 125 µm (B–D).
280 KANZLER ET AL.
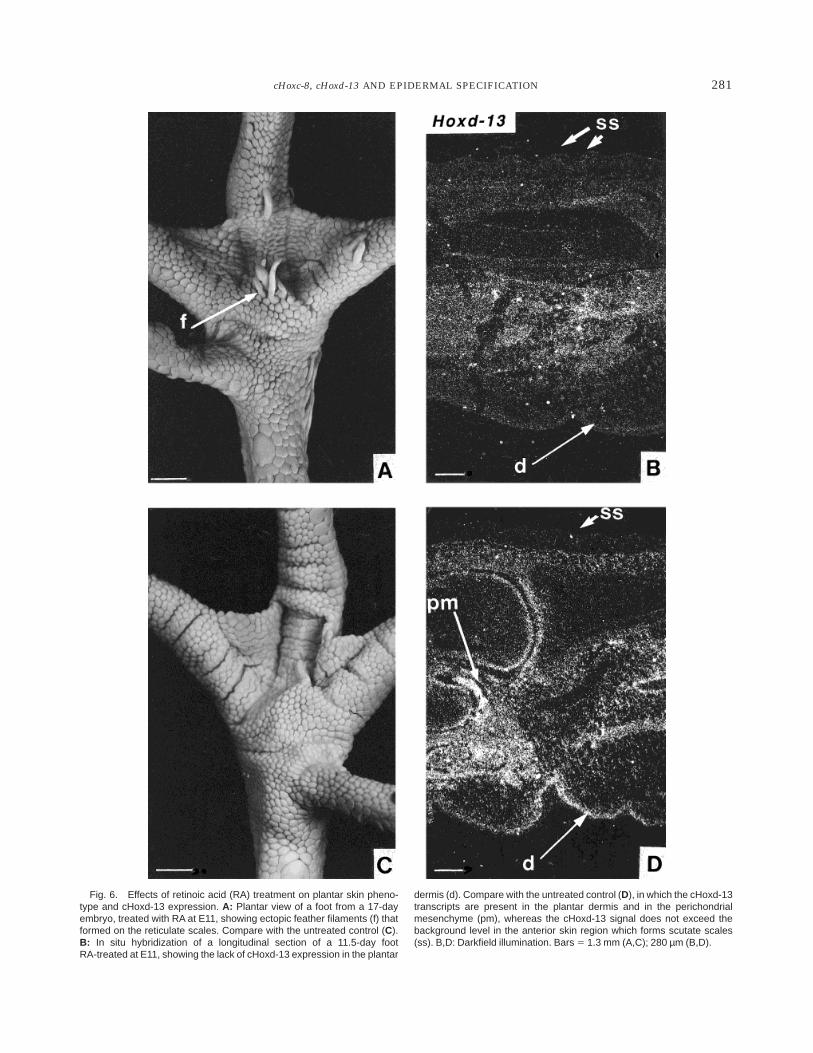
Fig. 6. Effects of retinoic acid (RA) treatment on plantar skin pheno-type and cHoxd-13 expression. A: Plantar view of a foot from a 17-dayembryo, treated with RA at E11, showing ectopic feather filaments (f) thatformed on the reticulate scales. Compare with the untreated control (C).B: In situ hybridization of a longitudinal section of a 11.5-day footRA-treated at E11, showing the lack of cHoxd-13 expression in the plantar
dermis (d). Compare with the untreated control (D), in which the cHoxd-13transcripts are present in the plantar dermis and in the perichondrialmesenchyme (pm), whereas the cHoxd-13 signal does not exceed thebackground level in the anterior skin region which forms scutate scales(ss). B,D: Darkfield illumination. Bars 5 1.3 mm (A,C); 280 µm (B,D).
281cHoxc-8, cHoxd-13 AND EPIDERMAL SPECIFICATION
proximodistal region of the limb buds of the 3-day chickembryo is similar to the pattern previously reported forthe mouse at a comparable stage of development (E10.5) (Le Mouellic et al., 1988, 1992). Likewise, thepattern of expression of cHoxc8 in the skin at E7.5corresponds with the overall expression of this genealong the cephalo-caudal and dorso-ventral axes previ-ously reported in embryonic murine skin at a similarstage (Kanzler et al., 1994). The cHoxc-8 transcripts arepresent in both the epidermis and the dermis of thefeather primordia and feather buds and disappearafterward. These results suggest that cHoxc-8 expres-sion plays a role in the thoracic region during theinitiation and first stages of feather embryonic morpho-genesis. In contrast, no cHoxd-13 expression was foundin the trunk skin over the period of morphogenesis inthe different feather pterylae, including the caudaltract. The cHoxd-13 expression characterizes the ven-tral skin of both wing and leg autopode, and particu-larly the plantar dermis during skin morphogenesis.
This is in concordance with the restricted pattern ofthis gene in the distal part of the limb, already shownby D. Duboule and coworkers (Dolle et al., 1991, 1993).In the mouse, the Hoxd-13 gene is expressed in aslightly more distal skin region, namely in the anteriorand plantar dermis from the digits when skin differen-tiation takes place (unpublished results from Dr. J.P.Viallet in our group). It should be noted that in bothspecies, the cHoxd-13 transcripts were only found inthe dermis. Given their distinct expression pattern inskin, the cHoxc-8 and cHoxd-13 could be part of differ-ent homeoproteins sets, providing positional informa-tion specifying the dorsal thorax and autopodial ventralskin territories, respectively. Nevertheless, the differ-ence between the ventral skin of the wing, which isfeathered, and the plantar skin implies that the forma-tion of reticulate scales must involve a further set ofinstructions which prevents the plantar epidermis fromdeveloping into feathers and which remains to beidentified. Indeed, the results of heterotopic recombi-
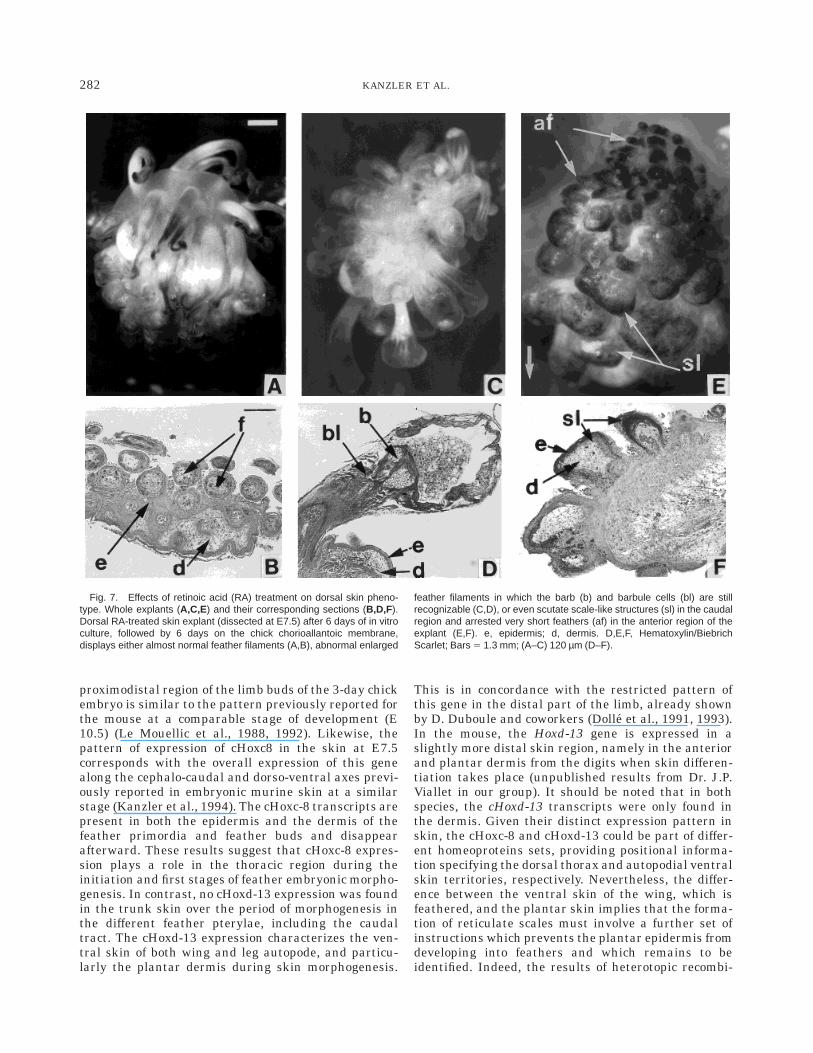
Fig. 7. Effects of retinoic acid (RA) treatment on dorsal skin pheno-type. Whole explants (A,C,E) and their corresponding sections (B,D,F).Dorsal RA-treated skin explant (dissected at E7.5) after 6 days of in vitroculture, followed by 6 days on the chick chorioallantoic membrane,displays either almost normal feather filaments (A,B), abnormal enlarged
feather filaments in which the barb (b) and barbule cells (bl) are stillrecognizable (C,D), or even scutate scale-like structures (sl) in the caudalregion and arrested very short feathers (af) in the anterior region of theexplant (E,F). e, epidermis; d, dermis. D,E,F, Hematoxylin/BiebrichScarlet; Bars 5 1.3 mm; (A–C) 120 µm (D–F).
282 KANZLER ET AL.
nants show that the plantar epidermis acquires arestricted ability to only differentiate reticulate scaleson E11, which coincides with the expression of cHoxd-13in the underlying dermis, while the dorsal epidermis,and to a lesser degree the midventral epidermis, whichboth express cHoxc-8, are endowed with feather-forming ability, as shown by their ability to interpretinducing clues originating either from a dorsal or aplantar dermis to go over the feather differentiationprogram. The morphogenesis of such heterotopic dermal–epidermal recombinants is consistent with the hypoth-esis of an early regional specification of the epidermalabilities (Dhouailly and Sengel, 1983; Viallet and Dh-ouailly, 1994). The fact that we were unable to detectHox gene expression in the dermal–epidermal recombi-nants, following the recombination, supports this hy-pothesis. The transcripts present in the dissected skinare likely to be destroyed by the enzymatic treatment ofthe skin, which is required to split it into its twocomponents. They may just be some leftovers, withoutthe need to be replaced. At the time of skin recombina-tion, 7 days for the dorsal, 10 days for the midventral,and 11 days for the plantar skin, the Hox genes havealready played their role in specifying epidermal abili-
ties. This may even occur earlier, at the time of skinformation, or even when the global pattern of hox geneexpression within the body is established by E3.
Phenotypic changes following RA treatment add indi-rect proof, which nevertheless supports our hypothesis.Retinoic acid treatment at E10 or E11 resulted in aninhibition of cHoxd-13 expression in the plantar der-mis. Embryos treated on day 11 developed featherfilaments on the reticulate scales. In contrast, in em-bryos treated on day 10, the cHoxd-13 expressionreappeared by E11, followed by normal reticulate scalemorphogenesis. The ectopic plantar feather filamentsdisplayed a feather b-keratin electrophoretic profile,suggesting that this is a real homeotic transformation.A possible explanation is that retinoic acid acts byinhibiting the posterior paralogs and respecifying theplantar cells to a more proximal positional identity,thus allowing the formation of feathers. It cannot beexcluded that RA may also act by enhancing theexpression of more anterior paralogs. The RA-treateddorsal skin explants formed abnormal structures whichcan be interpreted as abnormal short and enlargedfeather filaments, since the expressed set of b-keratinswas of feather type. Similar malformed feathers werealready obtained in the cephalic, alar, and femoralpterylae by in ovo RA treatment at E11 (Dhouailly etal., 1980). By the time of treatment the featheredspecification of the dorsal skin might have alreadyoccurred. Furthermore, it should be noted that the RAeffect on cHoxd-13 expression in embryonic skin isconsistent with the known effects of RA on Hox genesexpression in teratocarcinoma cells in vitro (Boncinelliet al., 1991; Simeone et al., 1991), during vertebraedifferentiation (Kessel and Gruss, 1991), and duringlimb morphogenesis (Hayamizu and Bryant, 1994): Theexpression of Hox genes belonging to the median para-logs 4 to 8 is generally not modified by RA, whereas the58 clustered genes are downregulated.
Further experiments of mis-expression of both Hoxc8and Hoxd13 are required to refine our hypothesis. Inparticular, the question still remains to know whetherthe thoracic skin is specified by the expression ofcHoxc8 as well as other median paralog Hox genes, orby the absence of expression of posterior paralogs ascHoxd13. Likewise, is the autopodial plantar skinspecified by the expression of cHoxd13 or the absence ina sufficient amount of homeoproteins of more anteriorparalogs? Finally, bird scale morphogenesis must re-quire, in addition to the proximodistal clues provided bythe Hox genes, still-unknown information involved inthe specification of the hindlimb versus the wing.
EXPERIMENTAL PROCEDUREScDNA Cloning
Approximately 1.5 3 106 recombinant phages of aLZapII (Stratagene) cDNA library prepared from E8.5chick dorsal skin RNA (Michaille et al., 1994) werescreened under high-stringency conditions (50% for-mamide; 53 standard saline citrate [SSC]; 1% standard
Fig. 8. Molecular identification of the protein content of chick cutane-ous appendages formed in normal and experimental skin. (MW) Molecu-lar weight; (lane 1 ) hatchling foot pad epidermis (reticulate scales); (lane2) reticulate-like structures formed after 8 days of culture by recombinantscomposed of E11 plantar dermis and E10 apteric midventral epidermis(see Fig. 9,H,I,J); (lane 3 ) 18-day ectopic feathers formed on foot padafter RA-treatment at E11 (see Fig. 6A); (lane 4 ) scutate-like structuresformed by E7 dorsal skin cultured for 6 days with added RA, and graftedfor 6 more days on chick CAM (see Fig. 7,E,F); (lane 5 ) hatchling anteriortarsometatarsal epidermis (scutate scales); (lane 6 ) hatchling dorsalfeathers. The a-keratins are designated as a-K, the b-keratins 1–8 as b-K;the histidine-rich proteins by an asterisk. 12% acrylamide gel stained withCoomassie blue.
283cHoxc-8, cHoxd-13 AND EPIDERMAL SPECIFICATION

saline sulfate [SDS]; 50 mM Tris-HCl pH 7.5; 0.1mg · ml21 denatured salmon sperm DNA at 44°C). Theprobe, a gift from Dr. H. Le Mouellic, was a 840-bpSalI-EcoRI probe comprising the main part of themouse Hoxc-8 homeodomain sequence and that ex-tended to the 38 end (Le Mouellic et al., 1988). Wefollowed the automatic excision protocol with helperphage and recircularization to generate the subclonescontaining the different chick cDNA inserts inpBluescripttII SK phagemid vector, as stated in theSTRATAGENE Kit instructions.
DNA Sequencing
The nucleotide sequences were determined with thedideoxy chain-termination method, using (35S)-dATPand the Pharmacia T7-sequencingy Kit, according tothe manufacturer’s instructions. The sequence of thecDNAs were read on both DNA strands.
Embryos
Warren breed fertile chick eggs were obtained fromthe ‘‘Centre d’aviculture de Cerveloup’’ (Moirans,France). Some Bresse breed eggs were obtained fromthe ‘‘Centre de Selection de la Race Bressane’’(Bechanne, France) as we have previously shown (un-published data) that this scaled-feet breed is moreprone than the Warren breed to develop feathers ontheir feet through RA treatment (70% instead of 10% oftreated embryos), which facilitates the correspondingin situ hybridization analysis. The eggs were incubatedat 38°C. For in situ hybridization, embryonic andpostnatal samples were embedded in Tissue Tecktmedium and frozen.
In Situ Hybridization
A 600-bp XhoI-PstI fragment of the cHoxd-13 cDNA(a gift from Dr. D. Duboule) containing the homeodo-main region and the following 38 sequences and a 1,500bp KpnI-EcoRI fragment of the isolated chick Hoxc-8cDNA clone were used as templates for synthesis ofeither (35S)-CTP or digoxygenin-11-UTP–labeled ribo-probes for in situ hybridization on frozen sections orwhole-mount embryos, respectively. In vitro transcrip-tion reactions were performed using either T7 or T3
RNApolymerase as directed by the manufacturer (Boeh-ringer Mannheim Biochemicals).
Whole-mount in situ hybridization protocol was basedon the Conlon and Rossant (1992) procedure with minormodifications. Following fixation, bleaching, and pro-teinase K treatment, either 3.5-, 4.5-, or 6.5-day em-bryos as well as 9.5-day feet were hybridized overnightat 70°C with 1 µg · ml21 of probe. After several washesunder stringent conditions and RNase A and T1 treat-ment, embryos and feet were incubated with anti-digoxigenin Fab conjugated to alkaline phosphatase(Boehringer Mannheim). Staining was allowed to pro-ceed at room temperature by the addition of alkalinephosphatase substrates NBT/BCIP. In situ hybridiza-tion on frozen sections was carried out essentially asfollows. Serial cryostat sections of 6–10 µm thicknesswere treated successively with acetone, 4% formalde-hyde at 4°C, 0.1 M triethanolamine/0.25% acetic anhy-dride, 50% formamide/13 SSC at 60°C, and two ethanolwashes. The hybridization buffer included 50% for-mamide, 10 mM DTT, 500 µg/ml tRNA, 13 salts-Denhardts, 10% dextran sulfate, and heat-denatured,(35S)-labeled antisense riboprobe (specific activity of5 3 108 c.p.m.21 · mg21; final concentration: 2 · 104
c.p.m./µl) or the corresponding opposite control probe.After overnight hybridization at 54°C, slides werewashed under stringent conditions and treated withRNase A and T1. After ethanol dehydration, slides weredipped in Kodak NTB-2 nuclear track emulsion andexposed for about 3 weeks before developing. Sectionswere stained with propidium iodide, mounted in Sur-gipatht, and then analyzed with an AX70 Olympusmicroscope using both darkfield and fluorescence illumi-nations.
In Vivo and In Vitro Retinoic Acid Treatment
Retinoic acid (125 µg of all-trans retinoic acid, a giftfrom Hoffmann-La Roche, Basel, Switzerland), previ-ously dissolved in absolute ethanol (50 µl) was injectedinto the amniotic cavity of 10- or 11-day chick embryo(Bresse breed). Preliminary experiments (Dhouailly etal., 1980) showed that 125 µg was the most suitabledose to obtain a high percentage of both surviving
TABLE 1. Skin Morphogenesis in Chick Heterotopic Epidermal–Dermal RecombinantsCultured 6–8 Days on Chick Chorioallantoic Membrane
Dermis
EpidermisDorsal E7
(number of cases)Plantar E10–11
(number of cases)Midventral apteriumE10 (number of cases)
Dorsal E7 Feather filaments (5) Arrested buds (18)Arrested buds plus
hypomorphicfeathers (18)
Feather filaments(Sengel et al., 1969)
Plantar E11 Feather filaments (10) Reticulate scales (5) Abnormal reticulae (12)Abnormal feathers and
reticulae (11)Midventral apterium
E10Glabrous skin (2) (and
Sengel et al., 1969)Glabrous skin (4) Glabrous skin (2)
284 KANZLER ET AL.
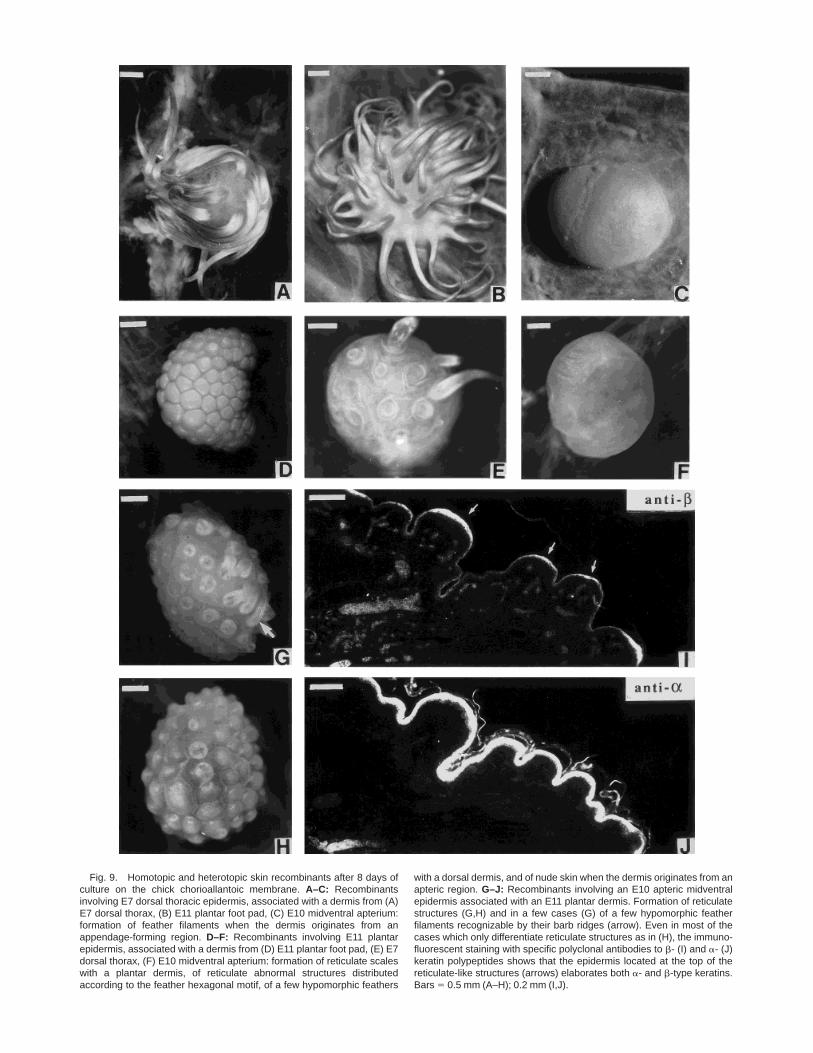
Fig. 9. Homotopic and heterotopic skin recombinants after 8 days ofculture on the chick chorioallantoic membrane. A–C: Recombinantsinvolving E7 dorsal thoracic epidermis, associated with a dermis from (A)E7 dorsal thorax, (B) E11 plantar foot pad, (C) E10 midventral apterium:formation of feather filaments when the dermis originates from anappendage-forming region. D–F: Recombinants involving E11 plantarepidermis, associated with a dermis from (D) E11 plantar foot pad, (E) E7dorsal thorax, (F) E10 midventral apterium: formation of reticulate scaleswith a plantar dermis, of reticulate abnormal structures distributedaccording to the feather hexagonal motif, of a few hypomorphic feathers
with a dorsal dermis, and of nude skin when the dermis originates from anapteric region. G–J: Recombinants involving an E10 apteric midventralepidermis associated with an E11 plantar dermis. Formation of reticulatestructures (G,H) and in a few cases (G) of a few hypomorphic featherfilaments recognizable by their barb ridges (arrow). Even in most of thecases which only differentiate reticulate structures as in (H), the immuno-fluorescent staining with specific polyclonal antibodies to b- (I) and a- (J)keratin polypeptides shows that the epidermis located at the top of thereticulate-like structures (arrows) elaborates both a- and b-type keratins.Bars 5 0.5 mm (A–H); 0.2 mm (I,J).

embryos and morphogenetic alterations. Feet fromcontrol and treated embryos were collected 12 and 24 hrafter injection for in situ hybridization, as well as 6days later to study the resulting skin phenotype and todetermine the keratin composition of the appendagesby SDS-polyacrylamide gel electrophoresis (PAGE).
Dorsal skin tissues from 7.5-day chick embryo (War-ren breed) were microdissected from the level of thewing to the caudal extremity. Cultures were then set upby placing the explants onto a grid in a Falcon dish withDMEM (Dulbecco’s modified Eagle medium) supple-mented with 20% fetal calf serum and either all-transretinoic acid previously dissolved in ethanol (finalconcentration: 5 µg/ml) or an equivalent volume ofethanol alone. The medium was changed every 2 days.After 2 or 6 days of in vitro culture, the explants weregrafted onto the chick chorioallantoic membrane (CAM)for 6 more days. Some of the control and RA-treatedgrafts were conserved at 220°C for further proteincontent analysis, and some of them were embedded inTissue Teckt medium for cryostat sectioning and in situhybridization analysis.
Heterotopic Dermal–Epidermal Recombinations
Midthoracic dorsal, ventral, and plantar skin frag-ments were dissected from 7-, 10-, and 11-day Warrenchick embryos, respectively, in Ca21- and Mg21-freeEarle’s saline. Dermis and epidermis were separated inTyrode’s solution after incubation in 2% trypsin and 1%pancreatin in Earle’s saline for 10 min at 4°C; proteasedigestion was stopped in 50% fetal calf serum. Tissueswere reassociated in heterotopic recombinations (re-ferred in Table 1) on a semisolid nutritive agar andplaced for 30 min at 37°C to obtain sufficient mutualadhesion between dermis and epidermis. Recombi-nants were transferred to the CAM of 10-day chickembryos, cultured for 6 to 8 days, photographed, andthen some of them were processed for electrophoretickeratin analysis or immunofluorescent staining with apolyclonal rabbit antibody which recognizes all b-keratins (Dhouailly and Sawyer, 1984).
Keratin Analysis
Control skin fragments were dissected from thetarsometatarsus, central foot pad, and feather pluckedfrom the back of a 21-day hatchling chicken. Skinrecombinants and RA-treated dorsal explants wereretrieved from the CAM 7 or 8 days after grafting.Ectopic feather filaments formed after RA-treatment onthe reticula were plucked at E19. For gel electrophore-sis, keratin polypeptides from control and experimentalspecimens were isolated and S-carboxymethylated ac-cording to the procedure of Dhouailly et al. (1978). Thenone-dimensional SDS-PAGE (12% acrylamide) was per-formed according to Laemmli (1970).
ACKNOWLEDGMENTS
We are grateful to Dr. D. Duboule for the gift of thechick Hoxd-13 probe and to Dr. H. Le Mouellic for the
mouse Hoxc-8 probe, as well as for their advice, and toDr. R.H. Sawyer, in the laboratory of whom D. Dhou-ailly became interested in chick reticulate morphogen-esis. We thank Dr. J.P. Viallet for his advice in perform-ing the whole mounts, Mr. Ray Dunn and Mrs. G.Chevalier for their help in keratin analysis, and Mrs. B.Peyrusse for the illustrations. We are indebted to Dr.Bernigaud for providing the Bresse breed eggs and toDr. Jennifer Seed for critical reading of the manuscript.This work was supported by ARC grant 6233 and a‘‘Fondation de la Recherche Medicale’’ grant to D.Dhouailly.
REFERENCESBoncinelli E, Acampora D, Pannese M, D’Esposito M, Somma R,
Gaudino G, Stornaiuolo A, Cafiero M, Faiella A, Simeone A. Organi-zation of human class I homeobox genes. Genome 1989;31:745–756.
Boncinelli E, Simeone A, Acampora D, Mavilio F. HOX gene activationby retinoic acid. TIG 1991;7:329–334.
Cadi R, Dhouailly D, Sengel P. Use of retinoic acid for the analysis ofdermal-epidermal interactions in the tarsometatarsal skin of thechick embryo. Dev Biol 1983;100:489–495.
Cech TR. RNAsplicing: three themes with variations. Cell 1983;34:713–716.
Chang C-P, Shen W-F, Rozenfeld S, Lawrence HJ, Largman C, ClearyML. Pbx proteins display hexapeptide-dependent cooperative DNAbinding with a subset of Hox proteins. Genes Dev 1995;9:663–674.
Chuong CM, Olivier G, Ting SA, Jegalian BG, Chen HM, De RobertisEM. Gradients of homeoproteins in developing feather buds. Devel-opment 1990;110:1021–1030.
Chuong CM, Ting SA, Widelitz RB, Lee YS. Mechanism of skinmorphogenesis. II. Retinoic acid modulates axis orientation andphenotypes of skin appendages. Development 1992;115:839–852.
Chuong CM, Randall B, Widelitz B, Ting-Xin J. Adhesion moleculesand homeoproteins in the phenotypic determination of skin append-ages. J Invest Dermatol 1993;101:10S–14S.
Colberg-Poley AM, Voss SD, Chowdhury K, Gruss P. Structuralanalysis of murine genes containing homeobox sequences and theirexpression in embryonal carcinoma cells. Nature 1985;314:713–718.
Conlon RA, Rossant J. Exogenous retinoic acid rapidly inducesanterior ectopic expression of murine Hox-2 genes in vivo. Develop-ment 1992;116:357–368.
Couly G, Le Douarin N. The fat map of the cephalic neural primordiumat the presomitic to the 3-somite stage in the avian embryo.Development 1988;103s:101–113.
Dhouailly D. Dermo-epidermal interactions during morphogenesis ofcutaneous appendages in amniotes. Front Matrix Biol 1977;4:86–121.
Dhouailly D. Specification of feather and scale patterns. In: Malincin-ski GM, Bryant SV, eds. Pattern Formation. New York: MacmillanPub. C., 1984:581–601.
Dhouailly D, Sengel P. Feather forming properties of the foot integu-ment in avian embryos. In: Sawyer RH, Fallon JF, eds. Epithelial-Mesenchymal Interactions in Development. New York: PraegerPress, 1983:147–161.
Dhouailly D, Sawyer RH. Avian scale development: XI. Initial appear-ance of the dermal defect in Scaleless skin. Dev Biol 1984;105:343–350.
Dhouailly D, Rogers GE, Sengel P. The specification of feather andscale protein synthesis in dermal-epidermal recombinations. DevBiol 1978;65:58–68.
Dhouailly D, Hardy MH, Sengel P. Formation of feathers on chick footscales: a stage-dependant morphogenetic response to retinoic acid. JEmbryol Exp Morphol 1980;58:63–78.
Dolle P, Izpisua-Belmonte JC, Boncinelli E, Duboule D. The Hox4–8gene is localised at the extremity of the Hox-4 complex and isexpressed in the most posterior parts of the body during develop-ment. Mech Dev 1991;36:3–14.
286 KANZLER ET AL.

Dolle P, Dierich A, LeMeur M, Schimmang T, Schuhbaur B, ChambonP, Duboule D. Disruption of the Hoxd-13 gene induces localizedheterochrony leading to mice with neotenic limbs. Cell 1993;75:431–444.
Duboule D. The vertebrate limb: a model system to study theHox/HOM gene network during development and evolution. Bioas-says 1992;14:375–383.
Duboule D, Dolle P. The structural and functional organization of themurine HOX gene family resembles that of Drosophila homeoticgenes. EMBO J 1989;8:1497–1505.
Duboule D, Morata G. Colinearity and functional hierarchy amonggenes of the homeotic complexes. TIG 1994;10:358–364.
Gregg K, Wilton SD, Parry DAD, Rogers GE. A comparison of genomiccoding sequences for feather and scale keratins: structural andevolutionary implications. EMBO J 1984;3:175–178.
Haake AR, Konig G, Sawyer RH. Avian feather development: relation-ships between morphogenesis and keratinization. Dev Biol 1984;106:406–413.
Hayamizu TF, Bryant SV. Reciprocal changes in Hoxd-13 and RAR-b2expression in response to retinoic acid in chick limb buds. Dev Biol1994;166:123–132.
Izpisua-Belmonte J-C, Tickle C, Dolle P, Wolpert L, Duboule D.Expression of the homeobox Hox-4 genes and the specification ofposition in chick wing development. Nature 1991;350:585–589.
Kanzler B, Viallet JP, Le Mouellic H, Boncinelli E, Duboule D,Dhouailly D. Differential expression of two different homeobox genefamilies during mouse tegument morphogenesis. Int J Dev Biol1994;38:633–640.
Kemp DJ, Rogers GE. Differentiation of avian keratinocytes. Charac-terization and relationship of the keratin proteins of the adult andembryonic feathers and scales. Biochemestry 1972;11:969–975.
Kessel M, Gruss P. Homeotic transformations of murine vertebrae andconcomitant alteration of Hox codes induced by retinoic acid. Cell1991;67:89–104.
Knapp LW, Shames RB, Barnes GL, Sawyer RH. Region-specificpatterns of beta keratin expression during avian skin development.Dev Dyn 1993;196:283–290.
Krumlauf R. Hox genes in vertebrate development. Cell 1994;78:191–201.
Laemmli UK. Cleavage of structural proteins during the assembly ofthe head of bacteriophage T4. Nature 1970;264:377–379.
Le Mouellic H, Condamine H, Brulet P. Pattern of transcription of thehomeogene Hox-3.1 in the mouse embryo. Genes Dev 1988;2:125–135.
Le Mouellic H, Lallemand Y, Brulet P. Homeosis in the mouse inducedby a null mutation in the Hox-3.1 gene. Cell 1992;69:251–264.
MacGinnis W, Krumlauf R. Homeobox genes and axial patterning.Cell 1992;68:283–302.
Mauger A. Role du mesoderme somitique dans le developpement duplumage dorsal chez l’embryon de poulet. II Regionalisation dumesoderme plumigene. J Embryol Exp Morphol 1972;28:343–366.
Michaille JJ, Blanchet S, Kanzler B, Garnier JM, Dhouailly D.Characterization of cDNAs encoding the chick retinoic acid receptorg2 and preferential distribution of retinoic acid receptor g tran-scripts during chick skin development. Dev Dyn 1994;201:334–343.
Moll R, Franke WW, Shiller DL. The catalog of human cytokeratins:patterns of expression in normal epithelia, tumors and culturedcells. Cell 1982;31:11–24.
Sawyer RH. The role of epithelial-mesenchymal interactions in regu-lating gene expression during avian scale morphogenesis. In: Saw-yer RH, Fallon JF, eds. Epithelial-Mesenchymal Interactions inDevelopment. New York: Praeger Press, 1983:115–146.
Sawyer RH, Borg TK. Avian scale development. VI. Ultrastructure ofthe keratinizing cells of reticulate scales. J Morphol 1979;161:111–122.
Sengel P. Morphogenesis of skin. In Abercrombie M, Newth DR, TorreyJG, eds. Developmental and Cell Biology Series. Cambridge: Cam-bridge University Press, 1976:1–277.
Sengel P, Dhouailly D, Kieny M. Aptitude des constituants cutanes del’apterie medio-ventrale du poulet a former des plumes. Dev Biol1969;19:436–446.
Simeone A, Acampora D, Nigro V, Faiella A, D’Esposito M, StornaiuoloA, Mavilio F, Boncinelli E. Differential regulation by retinoic acid ofthe homeobox genes of the four HOX loci in human embryonalcarcinoma cells. Mech Dev 1991;33:215–227.
Viallet JP, Dhouailly D. Retinoic acid and mouse skin morphogenesis:II. Role of epidermal competence in hair glandular metaplasia. DevBiol 1994;166:277–288.
287cHoxc-8, cHoxd-13 AND EPIDERMAL SPECIFICATION




















