
Dynamics of Neutrophil Migration in Lymph Nodes during Infection

ARTICLE IN PRESS
Available at www.sciencedirect.com
Developmental and Comparative Immunology (2008) 32, 773–783
0145-305X/$ - see frodoi:10.1016/j.dci.20
�Corresponding au
E-mail address: p
journal homepage: www.elsevier.com/locate/dci
Natural killer cells in lymph nodes of healthy calvesexpress CD16 and show both cytotoxic andcytokine-producing properties
Preben Boysena,�, Gjermund Gunnesb, Daniela Pendec,Mette Valheimd, Anne K. Storseta
aDepartment of Food Safety and Infection Biology, Norwegian School of Veterinary Science,P.O. Box 8146 Dep., N-0033 Oslo, NorwaybDepartment of Basic Sciences and Aquatic Medicine, Norwegian School of Veterinary Science,P.O. Box 8146 Dep., N-0033 Oslo, NorwaycIstituto Nazionale per la Ricerca sul Cancro, Genova, ItalydDepartment for Animal Health, Section for Pathology, National Veterinary Institute,P.O. Box 8156 Dep., N-0033 Oslo, Norway
Received 23 October 2006; received in revised form 30 October 2007; accepted 17 November 2007Available online 18 December 2007
KEYWORDSNatural killer cells(NK cells);Lymph nodes/lymphoid organs;Animal models;Cattle/bovine;Phenotype/cellmarkers;Cytotoxicity;Cytokines;Interferon-gamma
nt matter & 200707.11.006
thor. Tel.: +47 229
reben.boysen@ve
SummaryNatural killer (NK) cells were recently shown to play an important immunomodulatory rolein lymph nodes. We here report the presence, phenotype and function of NK cells residentin lymph nodes of several anatomical sites of healthy calves. NKp46+/CD3�lymphocytes,recently demonstrated to precisely identify NK cells in all tested species, were present inthe paracortex and the medulla of bovine lymph nodes. Most lymph node-derived NK cellsexpressed CD16 and perforin, and a lytic capacity was demonstrated, while a well-developed interferon-gamma response to interleukin-2 and interleukin-12 stimulation wasalso seen. Lymph node-derived NK cells differed from those in blood by a higher expressionof the activation markers CD44 and CD25, as well as CD8. L-selectin (CD62L) was expressedby the majority of lymph node-derived NK cells, consistent with a dependency of thismolecule for migration to lymph nodes. Unlike in blood, the majority of lymph node NKcells had little or no CD2 expression. Compared to available literature, calf lymph nodescontained NK cells in numbers equal to or higher than reported in humans, and clearlyhigher than in mice. These findings suggest a cytotoxic role of lymph node residing NKcells, beyond the predominantly cytokine-producing role previously inferred from studieson human NK cells.& 2007 Elsevier Ltd. All rights reserved.
Elsevier Ltd. All rights reserved.
64817; fax: +47 22964818.
ths.no (P. Boysen).

ARTICLE IN PRESS
P. Boysen et al.774
from herds reported free from disease problems, and routinely
IntroductionNatural killer (NK) cells were until recently thought not to besignificantly present in normal lymph nodes (LNs) [1], perhapsbecause of limitations of suitable monoclonal antibodies forimmunohistochemical analysis [2,3]. However, this view wasrecently challenged by the demonstration of significantnumbers of NK cells in LNs and tonsils from healthy humans[4,5]. Despite low relative numbers, it was estimated thatthese organs pooled together contain around 10 times asmany NK cells as peripheral blood. In contrast, LNs of healthymice appear to harbour only few NK cells [6–8], but uponinflammatory stimuli they are recruited to lymph nodes,where they provide IFN-g for T-cell priming and a subsequentTh1 polarization [8]. Similar NK cell priming of T cells wasrecently shown also in man [9], and subsequent demonstra-tions of an intimate co-localization between NK cells,dendritic cells and T cells in LNs have provided evidencethat such interactions are pivotal in shaping both the innateand adaptive immune responses [6,10–12].
In sharp contrast to blood, the majority of NK cells inhuman LNs were found to be CD56bright/CD16� [4,5]. Previousstudies had suggested that NK cells with this phenotyperepresented a cytokine-producing NK cell subset distinct fromthe primarily cytolytic CD56dim/CD16+ NK cells [13–16], so itwas hypothesized that these subsets might have differentialtrafficking patterns to different organs. However, recentinvestigations have provided evidence that the formerphenotype is likely to be an immature stage that is terminallydifferentiated into the latter in lymphiod organs [17,18].
Like in the human [19], rat [20] and mouse [10], NK cellsin cattle can be identified as NKp46+/CD3� lymphocytes[21], comprising 1–10% of the lymphocyte population inperipheral blood [22]. Interestingly, NK cells were present inmesenterial LNs of healthy cattle, the majority being CD2�,unlike in blood where most are CD2+ [21]. We found thatthe CD2� NK cells may represent a distinct bovine NK cellsubset or differentiation stage [23]. Unfortunately, a directcomparison to the human subsets was not feasible, since likein rodents, CD56 appears not to be present on bovine NKcells [24], and CD16 expression has not been previouslydemonstrated on bovine lymphocytes. Nevertheless, it isnoteworthy that the CD2� subset in bovine blood displayed astrong proliferative and cytokine-producing capacity [23],characteristics shared with the NK cell subset that domi-nates in human LNs [5,18].
In the present study, we describe the presence andcharacteristics of NK cells found in a selection of LNs fromhealthy calves. We demonstrate the expression of CD16 bybovine NK cells, and show that the majority of NK cells incalf LNs are CD16+, contain perforin and are cytotoxic.Furthermore, these LN-derived NK cells display a distinctphenotype, including elevated expression of activationmarkers and little or no expression of CD2.
Materials and methods
Animals and cell preparations
Four unrelated Norwegian Red calves, two males and twofemales, were used in this study. The calves were purchased
examined for the absence of disease while housed isolatedfrom other animals. At 9–11 weeks of age (observing well-developed ruminant function), blood samples were collectedand the animals were euthanized by penetrating captive boltgun and thorough bleeding of the animal, in accordance withNorwegian animal protection laws. Tissue samples wereharvested aseptically and used immediately for most applica-tions or freezed for immunohistochemical analysis as pre-viously described [25]. Cell homogenates from fresh tissueswere prepared by slicing thin cross-sections representative ofthe organ, followed by mechanical disaggregation usingMedimachine (Beckton Dickinson), in the presence of EDTA,as described [25]. Mononuclear cells from these homogenatesas well as from blood collected in EDTA-containing tubes wereseparated as previously described [21]. Briefly, mononuclearcells were separated by gradient centrifugation (Lymphoprep;Axis-shield, Norway) and washed twice. For some CD16expression analyses, purified NK cell isolates were positivelyselected using Dynabeads (Dynal, Invitrogen, Oslo, Norway)and monoclonal antibodies (mAbs) against bovine NKp46(AKS1;IgG1, Serotec, Oxford, UK) and cultured in recombinantbovine interleukin-2 (rbIL-2) containing cell culture medium(RPMI; GIBCO, Invitrogen) for 12 days [21,23]. Analyses ofperforin and CD62L expression were performed on cryopre-served mononuclear cell suspensions as described [26]. Toverify the quality of these preparations, some markersanalysed in fresh cells were reassessed in the cryopreservedcells. Analysis of the CD3 marker in relation to NKp46 wasperformed on fresh slaughter house material of adult cattle.
Flow cytometric analysis
Two- or three-coloured flow cytometric analysis of surfacereceptors was performed on mononuclear cells, as previouslydescribed [21], using antibodies against the followingmolecules: CD2 (MUC2A;IgG2a), CD3 (MM1A;IgG1), CD8(BAQ111A;IgM or CACT80C;IgG1), CD25 (CACT108A;IgG2a),CD44 (BAG40A;IgG3), d-TCR chain (GB21;IgG2b), B-lympho-cytes (LCT30A;IgG1) (all from VMRD), CD14 (CAM36A;IgG1,VMRD or TUK4;IgG2a, Dako, Glostrup, Denmark), CD62L(FMC46;IgG2b, Serotec, Oxford, UK) and NKp46 (AKS1). Twoanti-human CD16 mAbs were screened for cross-reactivity(KD1;IgG2a [27] or GRM1;IgG2a, Southern Biotech, Birming-ham, AL, USA). Subtype-specific secondary antibodies wereconjugated with FITC or PE (Southern Biotech, Birmingham,AL, USA), or with Alexa Fluor (AF) 488 or AF633 (MolecularProbes, Invitrogen). In some cases, directly conjugated mAbswere used: NKp46 (AKS1)*AF488 and CD3 (MM1A)*AF647(the latter kindly provided by Will L. Goff, Animal DiseaseResearch Unit, USDA-ARS, Pullman, Washington, USA), alongwith appropriate isotype controls (MOPC-21, BD Pharmingen).The samples were analysed on a FACS CALIBUR flow cytometer(Becton Dickinson), equipped with Cell-Quest Pro software.Gating was set for viable mononuclear cells in the forward/side scatter plot and 5 to 10� 105 cells were analysed.
Transfection of bovine CD16 in 293T cells
An expression construct containing the entire codingregion of bovine FcgIIIA/CD16 (GeneBank X99695) [28] was

ARTICLE IN PRESS
Natural killer cells in lymph nodes of healthy calves 775
constructed by amplifying the gene from cDNA generatedfrom bovine NK cells as previously described [23] with genespecific primers with BamH1 and XhoI restriction sites:50-GGATCCACTCTAACACACAGCATGTG-30 and 50-CTCGAGTG-CCTCCATTTGTCCTAAGG-30. The product was cloned intopCRs 2.1-TOPO Vector (Invitrogen), released by BamHI andXhoI digestion, subcloned into pcDNA I (Invitrogen) and usedto transfect 293T cells as previously described [23]. 293Tcells were released by Cell dissociation solution (Sigma) 24 hafter transfection and assayed by flow cytometric analysis.
Immunohistochemical analysis
Immunohistochemical preparation of frozen histologicalsections was performed as previously described [25]. Briefly,10 mm frozen tissue samples were sectioned and collected onslides precoated with 10% poly-L-lysine (Sigma, St. Louis,MO, USA), and stored at �70 1C. Following thawing anddrying at room temperature for 1 h, the sections werefixed in acetone for 10min, then blocked for 10min with 20%BSA/TBS. Primary antibodies (against NKp46 and CD2 asabove and CD21 (DU2-74-25;IgG2b) [29]) diluted in 1%BSA/TBS were incubated for 1 h on a rotating table atroom temperature. The sections were rinsed 3� in PBSand incubated for 30min with AF-conjugated secondaryantibodies as above (AF488*IgG1, AF594*IgG2a andAF594*IgG2b). The rinsed sections were mounted in poly-vinyl alcohol at pH 8 and stored at 4 1C until examination.
A Zeiss Axiovert 200M fluorescence inverted microscopeequipped with an LSM 510 laser confocal unit (Carl Zeiss,Jena, Germany) was used to detect specific confocalfluorescent signal in the sections. A Plan-Neofluar 25/0.80oil objective was used and the tissue was scanned witha 488 nm argon laser, and a 546 nm helium/neon lasercorresponding to the absorption maxima of each of thefluorochromes used. The 546 nm helium/neon laser excitedthe AF594 fluorochrome. Digital Images were captured usingthe Zeiss LSM 510 3.0 software.
Some of the tissue sections were analysed by conventionalmicroscopy with a Leica DM RXA fluorescence microscope(Leica Microsystems AG, Wetzlar, Germany). These imageswere captured through an HC PL Fluotar 20� /0.50 dryobjective with a Spot RT Slider digital camera (DiagnosticInstruments Inc., St. Sterling Heights, MI, USA).
Detection of intracellular and secreted IFN-c
Intracellular IFN-g assay was performed as describedpreviously [23]. Briefly, mononuclear cells were stimulatedfor 16 h with 50U/ml rbIL-2 and/or 5U/ml rbIL-12 (kindlyprovided by Jayne Hope, Institute of Animal Health,Compton, UK), including 10 mg/ml Brefeldin A (Sigma) duringthe last 4 h. Cells incubated with mAbs against the surfaceantigens CD2, CD3 and/or NKp46 (as listed above) werepermeabilized using Cytofix/Cytoperm and Perm/Washsolutions (both Pharmingen, Beckton Dickinson) as describedpreviously [23], and further incubated with anti-bovineIFN-g mAb (clone 6.6;IgG2b, kindly provided by GregersJungersen, Danish Institute for Food and Veterinary Re-search, Copenhagen), and analysed by flow cytometry.
For assessment of secreted IFN-g, supernatants of 2� 105
mononuclear cells in 200 ml medium stimulated as abovewere assessed using an ELISA (Bovine IFN-g EASIA, Biosource,Nivelles, Belgium), according to the manufacturer’s instruc-tions and previous description [23].
Cytotoxicity assays and perforin expression
The cytotoxic capacity of bovine NK cells was tested using51Cr release assay, using the murine FcgR+ target line P815,to obtain a redirected lysis effect as previously described[21], with modifications: For CD16 functional studies, NKcells cultured in the presence of IL-2 for 9–11 days wereused as effector cells. For comparison of mononuclear cellsfrom different organs, freshly prepared mononuclear cellswere used as effector cells, without any cytokine addition.In order to make comparison of NKp46-mediated lysis acrossmononuclear cell samples with variable content of NK cells,NK per target cell (NK/T) ratios were calculated based onnumbers of NKp46+ cells in phenotypic analyses performedin parallel. Specific 51Cr release was calculated on the basisof the ratio [(sample release�spontaneous release)/(totalrelease�spontaneous release)].
The examination of perforin expression was performed oncells permeabilized as above, using anti-perforin mAb (clonedG9;IgG2b, Pharmingen, Beckton Dickinson), and analysedby flow cytometry.
Results
Presence of CD16 on bovine NK cells
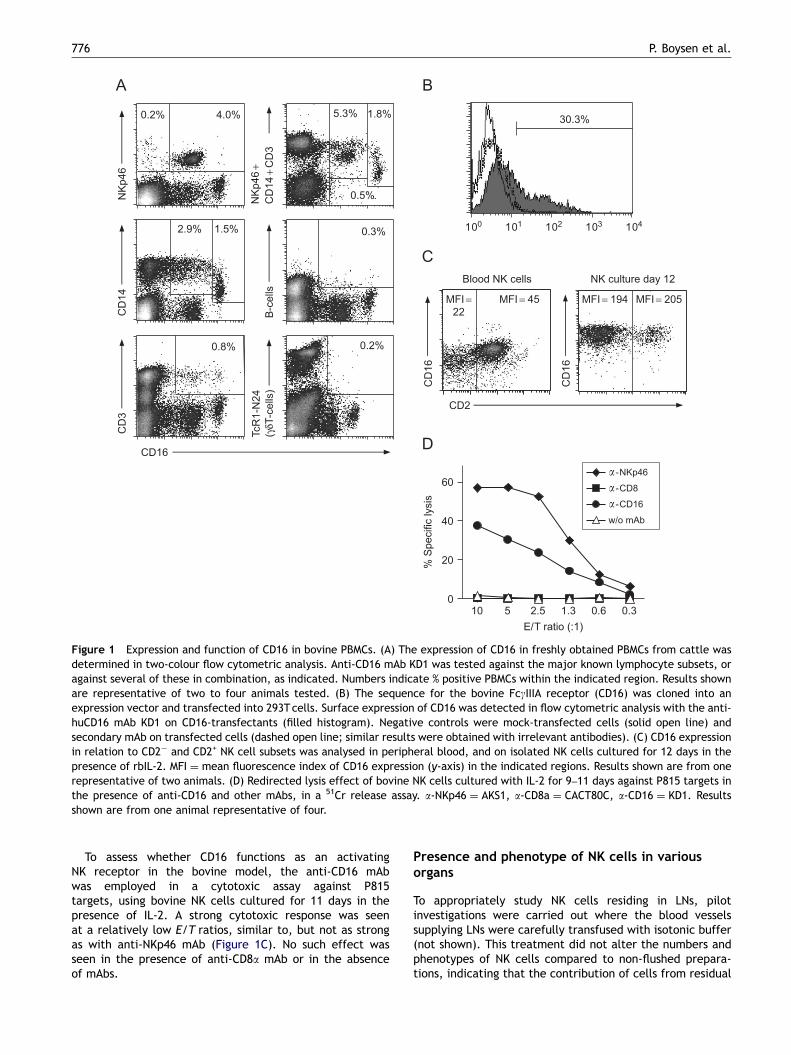
Two anti-human CD16 mAbs were screened for cross-reactivity with bovine peripheral blood mononuclear cells(PBMCs). One mAb (KD1) showed marked cross-reactivity(Figure 1A), while no binding was detected with the other(GRM1) (not shown). To confirm that KD1 recognized bovineCD16, bovine CD16 was transfected into 293T cells, and byflow cytometry the specific binding of KD1 was detected in30.3% of transfected cells (Figure 1B). In peripheral blood,�95% of NKp46+ cells expressed CD16 (Figure 1A), consistentwith the pattern in human NK cells [30]. The majority of theremaining CD16+ cells were CD14+, divided into distinctCD14bright/CD16dim and CD14dim/CD16bright populations, thelatter cells consistent with maturing monocytes [31]. CD16expression was found on a small fraction of CD3+ cells(T or NK–T cells), similar to findings in man [32], while all Bcells and gdT cells were CD16�. Simultaneous co-stainingwith a cocktail of NKp46, CD3 and CD14 mAbs indicatedthat CD16 is only expressed on NK cells, monocytes and T(or NK–T) cells.
Since bovine NK cells can be subdivided into twostable subsets depending on the CD2 expression [23], therelationship between CD2 and CD16 expression wasinvestigated. No such correlation was observed, either infreshly prepared PBMCs, or in NK cultures grown in thepresence of IL-2 (Figure 1C), or in cultures of separated CD2+
or CD2� NK cell subsets (not shown). A moderate upregula-tion of CD16 on NK cells was observed during IL-2 activation(Figure 1C).
ARTICLE IN PRESS
CD2
CD
16
CD
16
Blood NK cells NK culture day 12
MFI =22
MFI = 45 MFI = 194 MFI = 205
0
20
40
60
10 5 2.5 1.3 0.6 0.3
E/T ratio (:1)
% S
pecific
lysis
CD
3
0.8%
TcR
1-N
24
(γδT
-cells
)
0.2%
B-c
ells
0.3%
CD16
4.0%N
Kp46
0.2%C
D14
2.9% 1.5%
NK
p46
+C
D14
+C
D3
5.3% 1.8%
0.5%
100 101 102 103 104
30.3%
� -NKp46
� -CD8
� -CD16
w/o mAb
Figure 1 Expression and function of CD16 in bovine PBMCs. (A) The expression of CD16 in freshly obtained PBMCs from cattle wasdetermined in two-colour flow cytometric analysis. Anti-CD16 mAb KD1 was tested against the major known lymphocyte subsets, oragainst several of these in combination, as indicated. Numbers indicate % positive PBMCs within the indicated region. Results shownare representative of two to four animals tested. (B) The sequence for the bovine FcgIIIA receptor (CD16) was cloned into anexpression vector and transfected into 293Tcells. Surface expression of CD16 was detected in flow cytometric analysis with the anti-huCD16 mAb KD1 on CD16-transfectants (filled histogram). Negative controls were mock-transfected cells (solid open line) andsecondary mAb on transfected cells (dashed open line; similar results were obtained with irrelevant antibodies). (C) CD16 expressionin relation to CD2� and CD2+ NK cell subsets was analysed in peripheral blood, and on isolated NK cells cultured for 12 days in thepresence of rbIL-2. MFI ¼ mean fluorescence index of CD16 expression (y-axis) in the indicated regions. Results shown are from onerepresentative of two animals. (D) Redirected lysis effect of bovine NK cells cultured with IL-2 for 9–11 days against P815 targets inthe presence of anti-CD16 and other mAbs, in a 51Cr release assay. a-NKp46 ¼ AKS1, a-CD8a ¼ CACT80C, a-CD16 ¼ KD1. Resultsshown are from one animal representative of four.
P. Boysen et al.776
To assess whether CD16 functions as an activatingNK receptor in the bovine model, the anti-CD16 mAbwas employed in a cytotoxic assay against P815targets, using bovine NK cells cultured for 11 days in thepresence of IL-2. A strong cytotoxic response was seenat a relatively low E/T ratios, similar to, but not as strongas with anti-NKp46 mAb (Figure 1C). No such effect wasseen in the presence of anti-CD8a mAb or in the absenceof mAbs.
Presence and phenotype of NK cells in variousorgans
To appropriately study NK cells residing in LNs, pilotinvestigations were carried out where the blood vesselssupplying LNs were carefully transfused with isotonic buffer(not shown). This treatment did not alter the numbers andphenotypes of NK cells compared to non-flushed prepara-tions, indicating that the contribution of cells from residual

ARTICLE IN PRESS
Natural killer cells in lymph nodes of healthy calves 777
blood was minimal even in the absence of transfusion.Proper general bleeding of the animal during slaughterwas therefore considered sufficient, verified by visuallyinspecting the absence of blood in LNs. These pilotstudies also showed that all LNs from several parts of thejejunum comprised cells with a similar phenotype. Never-theless, specific attention was given to the distal jejunal LN(DJLN), which drains the ileocaecal valve, since previousstudies have shown a specific role for this area in pathogenuptake [33].
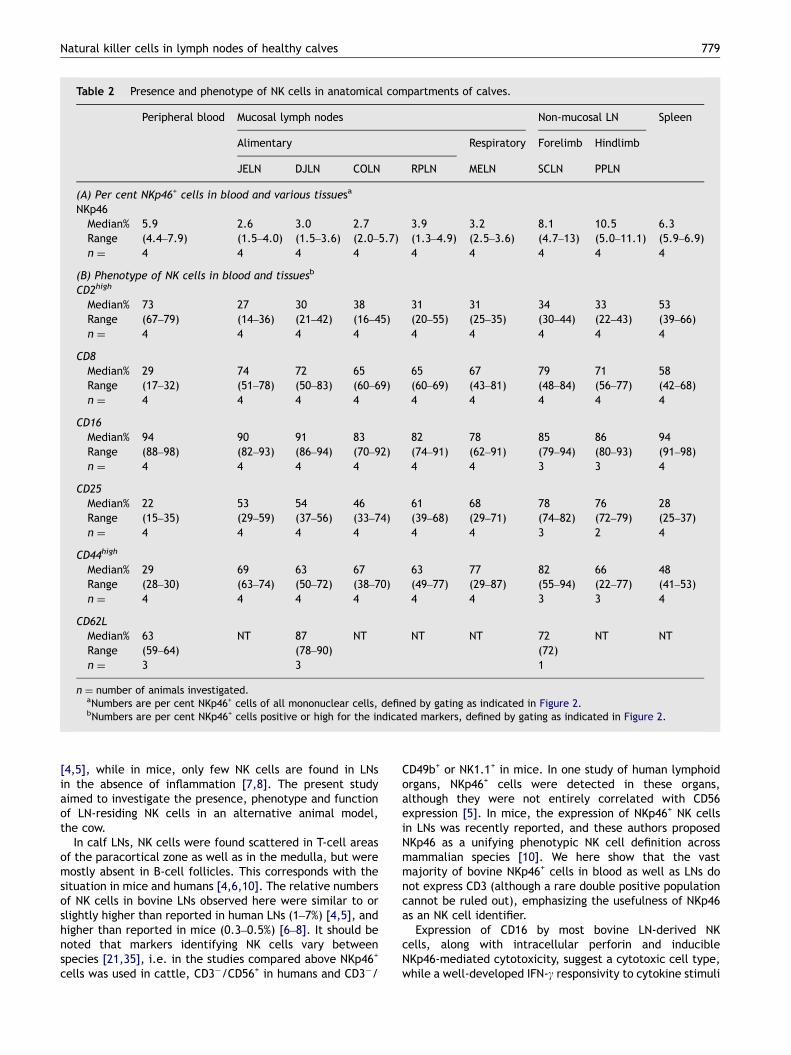
Mononuclear cells extracted from blood, a selection ofLNs (Table 1) [34] and some other tissues, were analysed forthe NK cell marker NKp46 together with other surfacemarkers (Figure 2A; Table 2). Bovine PBMCs contained 4–8%NK cells, LNs draining mucosal surfaces contained 1–6% NKcells, while non-mucosal LNs contained 5–13% NK cells(Table 2A). Non-mucosal LNs had a firm texture, yieldingrelatively few lymphocytes, compared to a soft texture ofmucosal LNs from which an abundance of cells washarvested. Analysis of two animals showed that also theproportion of CD3+ cells were higher in the non-mucosal LNs(�56% in SCLN and �33% in DJLN), while in one testedanimal B cells were considerably lower in such LNs (�20% inSCLN and �68% in DJLN) (not shown). The proportion of NKcells in mononuclear cells from the spleen was comparableto blood (6–7%). The vast majority of NKp46+ cells werenegative for the CD3 marker (in blood: median ¼ 97.7%,range ¼ 95.2–99.0%; in JLN: median ¼ 94.0%, range ¼87.7–96.9%) (Figure 2B), and also absent of the gdT-cellreceptor, CD4 and CD14 (not shown). The intensity of NKp46expression did not vary substantially between the organs(Figure 2A and B).
The majority of NK cells from all organs tested wereCD16+, with relatively little variation between organs(Figure 2; Table 2B). In blood, 67–79% of NK cells wereCD2high, the remaining population displaying a distinct CD2�
phenotype. In LNs around one-third of the NK cells wereCD2high, the majority were CD2low, while entirely CD2� NKcells were almost absent. In the spleen half of the NK cellsfell into either CD2low or CD2high subsets. Remarkabledifferences in CD44 expression were observed, as no orweak expression of this marker was seen in blood NK cells,while in all LNs a high expression of CD44 was seen in thegreat majority of NK cells, with no NK cells being completelynegative. In the spleen, NK cells were divided into equalproportions of CD44low and CD44high populations. More NKcells expressed CD25 and CD8 in LNs compared to blood and
Table 1 Description of lymph nodes investigated
Abbreviation English name Area of drenati
RPLN Retropharyngeal LN Upper alimentaJJLN Jejunal LN Small intestineDJLN Distal jejunal LN Small intestineCOLN Colonic LN Large intestineMELN Mediastinal LN Trachea, heart,SCLN Superficial cervical LN Skin and musclePPLN Popliteal LN Distal part of th
aRef. [34].
other tissues. NK cells in blood expressed CD62L as distinctnegative and positive populations, the latter in relativelyhigh density. In DJLN a more homogeneous CD62L expressionwas seen, few cells being entirely negative while thepositive cells had expression of lower density than in blood.A similar pattern was observed in SCLN from one animal.CD62L was downregulated on blood-derived NK cellscultured for 7 days in the presence of IL-2 (not shown).
Additional analysis of fresh slaughterhouse material of adultcattle showed moderately lower relative NK cell numbers inJLN (median ¼ 1.2%, range ¼ 0.6–1.8%, n ¼ 6; Figure 2B), butno apparent differences in phenotypic profile (not shown).
Localization of NK cells inside lymph nodes
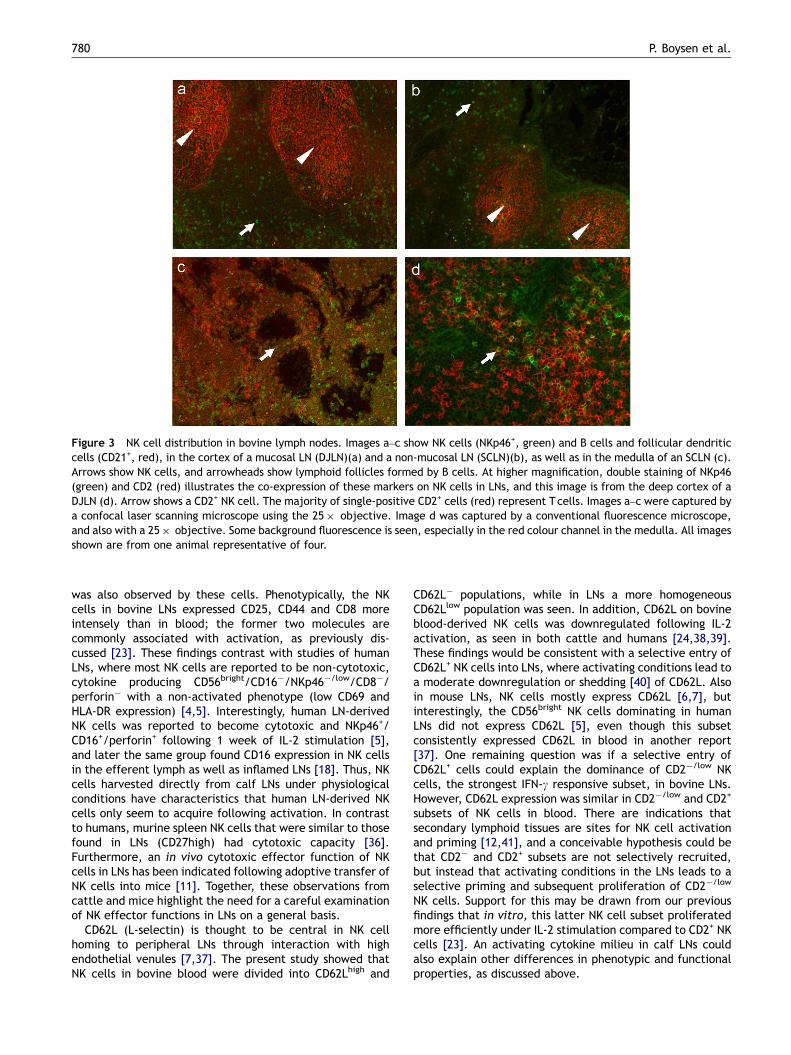
Immunohistochemical analysis showed that NKp46, whichis an appropriate identifier of bovine NK cells in blood [21],is a well-suited marker also for NK cell detection in tissues(Figure 3). NK cells were present in the paracortical (Figure 3aand b) as well as in the medullary (Figure 3c) areas of bovineLNs. In paracortical areas, NK cells appeared to be incomparable densities in both types of LNs, evenly distributedexcept in follicular areas where only few NK cells were seen. Inmucosal LNs, lymphoid follicles were typically numerous andlarge (Figure 3a), while non-mucosal LNs had smaller andfewer follicles (Figure 3b). Co-staining of LNs with mAbsagainst NKp46 and CD2 showed that a minor part of NK cells inthe LNs was double positive for these markers (Figure 3d),confirming findings made in flow cytometry (Table 2B).
IFN-c production by NK cells from various organs
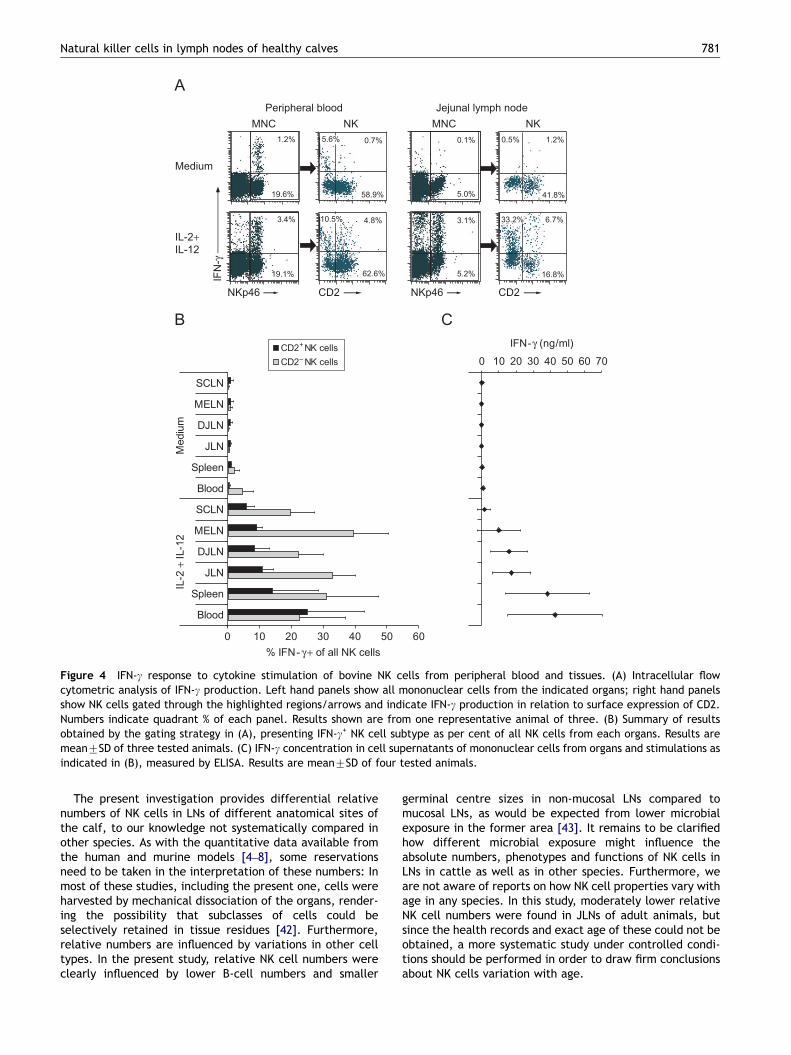
The IFN-g-producing capacity of NK cells and subsets fromvarious organs was analysed in an intracellular IFN-g assay(Figure 4A and B). In the absence of added cytokines, almostnone of the LN-derived NK cells, and a variable number(1–8%) of blood and spleen NK cells, mostly the CD2�/low
cells, contained IFN-g. Stimulation by rbIL-2 and rbIL-12induced IFN-g production in 18–61% of NK cells from LNs, andin 15–77% of NK cells from blood and spleen, hence with anindividual variation that did not reveal any differencesbetween organs. More CD2�/low NK cells produced IFN-g thanCD2high NK cells, except in blood where equal numbersof IFN-g+ NK cells from the two subsets were found. Asnoted above, the distribution of these subsets variedbetween organs (Table 2B), and when adjusting for this,
on (tributary territory)a Category
ry and respiratory tract Mucosal, jejunal area Mucosal, near ileocaecal valve Mucosal, ascending colon Mucosaloesophagus, thymus, lung, liver, spleen Mucosals of the neck and forelimb Non-mucosale hindlimb Non-mucosal

ARTICLE IN PRESS
Peripheral
blood
Jejunal
lymph
node
CD2 CD44CD25CD8 CD16
NK
p46
CD62L
30%70%79%21%
30%70% 78%22%
94%6%
84%16%
35%65%
59%41%
29%71%
74%26% 87%13%
63%37%
Peripheral
blood
Jejunal
lymph
node
NK
p46
CD3
3.1% 0.2%
45.7%
1.7% 0.1%
49.8%
Figure 2 Phenotype of NK cells in peripheral blood and jejunal LNs. Mononuclear cells from peripheral blood or LNs were analysedin two-colour flow cytometry, applying mAbs against NKp46 and the indicated markers. (A) Numbers indicate the population ofNKp46+ cells above or below an expression threshold (dashed line) for the shown representative animal. Summarized results for alltested animals are listed in Table 2. (B) The relation between CD3 and NKp46 on mononuclear cells is shown by quadrant percentagesfor each shown individual, which are representative of four (PBMC) or six (JLN) individuals.
P. Boysen et al.778
the CD2�/low NK cell subset of all organs consistently hadmore IFN-g responding cells compared to the CD2high subset(not shown). Stimulation by either rbIL-2 or rbIL-12separately resulted in similar patterns of IFN-g production,but at lower levels in all samples (not shown).
To complement these findings, IFN-g secretion to mono-nuclear cell culture supernatants was analysed in an ELISAfollowing stimulation with the same cytokines. No IFN-g wasdetected in cultures from any of the organs in the absenceof added cytokines (Figure 4C). Stimulation by rbIL-2 andrbIL-12 resulted in a high IFN-g secretion to supernatantsfrom blood and spleen cells, intermediate amounts frommucosal LN cells, and only low amounts from non-mucosal(SCLN) LN cells. Stimulation by each cytokine alone resultedin a similar pattern, but at lower levels in all samples(not shown). When comparing the ELISA and intracellularIFN-g detection methods for each animal individually, thesecreted IFN-g production to a great extent reflected theproportion of IFN-g+ NK cells (not shown).
Cytotoxicity and perforin expression of lymph nodeNK cells
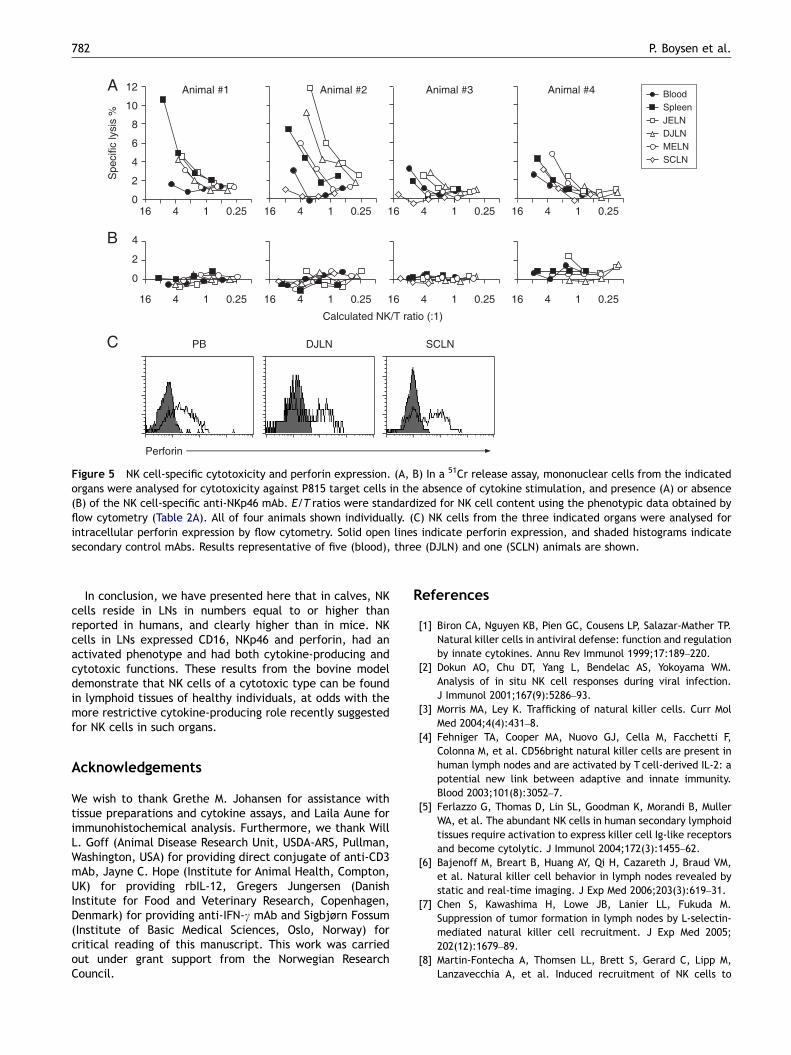
Mononuclear cells from blood and LNs were assessed forspontaneous cytotoxic activity in a 51Cr-release assay, usingE/T ratios from 100:1 to 6:1, without addition of cytokines.To assess NK-specific cytotoxicity, anti-NKp46 mAb and
FcR-bearing P815 target cells were used to obtain a redirectedlysis effect, and the results were made comparable byadjusting the effector cell numbers by NKp46+ expression. Inthe presence of mAb, low but detectable cytotoxic activitywas present in cells from all organs tested (Figure 5A). Thematerial was too small to allow a statistical comparisonbetween organs, but in three animals (#2–4) NK cells from non-mucosal LNs (SCLN) appeared to show a weaker cytotoxicitythan from mucosal LNs, while NK cells from blood appearedless cytotoxic than from mucosal LNs in three animals (#1, 2and 4). In the absence of mAbs no detectable cytotoxicactivity was detected (Figure 5B). When anti-CD16 (asdescribed above) was present, a slight tendency of redirectedlysis response was observed (not shown, n ¼ 2), but less thanwith IL-2-activated NK cells (Figure 1D), and not sufficientlyabove the background level to make any comparisons.
The perforin content of NK cells from blood (n ¼ 5), DJLN(n ¼ 3) and SCLN (n ¼ 1) was analysed by intracellular flowcytometric assay. A major proportion of NK cells from allthese tested organs contained perforin (Figure 5C). Theperforin expression did not differ between CD2� and CD2+
NK cell subsets from blood (not shown).
Discussion
Recent studies have shown that contrary to prior under-standing, NK cells reside in non-reactive LNs of humans
ARTICLE IN PRESS
Table 2 Presence and phenotype of NK cells in anatomical compartments of calves.
Peripheral blood Mucosal lymph nodes Non-mucosal LN Spleen
Alimentary Respiratory Forelimb Hindlimb
JELN DJLN COLN RPLN MELN SCLN PPLN
(A) Per cent NKp46+ cells in blood and various tissuesa
NKp46Median% 5.9 2.6 3.0 2.7 3.9 3.2 8.1 10.5 6.3Range (4.4–7.9) (1.5–4.0) (1.5–3.6) (2.0–5.7) (1.3–4.9) (2.5–3.6) (4.7–13) (5.0–11.1) (5.9–6.9)n ¼ 4 4 4 4 4 4 4 4 4
(B) Phenotype of NK cells in blood and tissuesb
CD2high
Median% 73 27 30 38 31 31 34 33 53Range (67–79) (14–36) (21–42) (16–45) (20–55) (25–35) (30–44) (22–43) (39–66)n ¼ 4 4 4 4 4 4 4 4 4
CD8Median% 29 74 72 65 65 67 79 71 58Range (17–32) (51–78) (50–83) (60–69) (60–69) (43–81) (48–84) (56–77) (42–68)n ¼ 4 4 4 4 4 4 4 4 4
CD16Median% 94 90 91 83 82 78 85 86 94Range (88–98) (82–93) (86–94) (70–92) (74–91) (62–91) (79–94) (80–93) (91–98)n ¼ 4 4 4 4 4 4 3 3 4
CD25Median% 22 53 54 46 61 68 78 76 28Range (15–35) (29–59) (37–56) (33–74) (39–68) (29–71) (74–82) (72–79) (25–37)n ¼ 4 4 4 4 4 4 3 2 4
CD44high
Median% 29 69 63 67 63 77 82 66 48Range (28–30) (63–74) (50–72) (38–70) (49–77) (29–87) (55–94) (22–77) (41–53)n ¼ 4 4 4 4 4 4 3 3 4
CD62LMedian% 63 NT 87 NT NT NT 72 NT NTRange (59–64) (78–90) (72)n ¼ 3 3 1
n ¼ number of animals investigated.aNumbers are per cent NKp46+ cells of all mononuclear cells, defined by gating as indicated in Figure 2.bNumbers are per cent NKp46+ cells positive or high for the indicated markers, defined by gating as indicated in Figure 2.
Natural killer cells in lymph nodes of healthy calves 779
[4,5], while in mice, only few NK cells are found in LNsin the absence of inflammation [7,8]. The present studyaimed to investigate the presence, phenotype and functionof LN-residing NK cells in an alternative animal model,the cow.
In calf LNs, NK cells were found scattered in T-cell areasof the paracortical zone as well as in the medulla, but weremostly absent in B-cell follicles. This corresponds with thesituation in mice and humans [4,6,10]. The relative numbersof NK cells in bovine LNs observed here were similar to orslightly higher than reported in human LNs (1–7%) [4,5], andhigher than reported in mice (0.3–0.5%) [6–8]. It should benoted that markers identifying NK cells vary betweenspecies [21,35], i.e. in the studies compared above NKp46+
cells was used in cattle, CD3�/CD56+ in humans and CD3�/
CD49b+ or NK1.1+ in mice. In one study of human lymphoidorgans, NKp46+ cells were detected in these organs,although they were not entirely correlated with CD56expression [5]. In mice, the expression of NKp46+ NK cellsin LNs was recently reported, and these authors proposedNKp46 as a unifying phenotypic NK cell definition acrossmammalian species [10]. We here show that the vastmajority of bovine NKp46+ cells in blood as well as LNs donot express CD3 (although a rare double positive populationcannot be ruled out), emphasizing the usefulness of NKp46as an NK cell identifier.
Expression of CD16 by most bovine LN-derived NKcells, along with intracellular perforin and inducibleNKp46-mediated cytotoxicity, suggest a cytotoxic cell type,while a well-developed IFN-g responsivity to cytokine stimuli
ARTICLE IN PRESS
Figure 3 NK cell distribution in bovine lymph nodes. Images a–c show NK cells (NKp46+, green) and B cells and follicular dendriticcells (CD21+, red), in the cortex of a mucosal LN (DJLN)(a) and a non-mucosal LN (SCLN)(b), as well as in the medulla of an SCLN (c).Arrows show NK cells, and arrowheads show lymphoid follicles formed by B cells. At higher magnification, double staining of NKp46(green) and CD2 (red) illustrates the co-expression of these markers on NK cells in LNs, and this image is from the deep cortex of aDJLN (d). Arrow shows a CD2+ NK cell. The majority of single-positive CD2+ cells (red) represent T cells. Images a–c were captured bya confocal laser scanning microscope using the 25� objective. Image d was captured by a conventional fluorescence microscope,and also with a 25� objective. Some background fluorescence is seen, especially in the red colour channel in the medulla. All imagesshown are from one animal representative of four.
P. Boysen et al.780
was also observed by these cells. Phenotypically, the NKcells in bovine LNs expressed CD25, CD44 and CD8 moreintensely than in blood; the former two molecules arecommonly associated with activation, as previously dis-cussed [23]. These findings contrast with studies of humanLNs, where most NK cells are reported to be non-cytotoxic,cytokine producing CD56bright/CD16�/NKp46�/low/CD8�/perforin� with a non-activated phenotype (low CD69 andHLA-DR expression) [4,5]. Interestingly, human LN-derivedNK cells was reported to become cytotoxic and NKp46+/CD16+/perforin+ following 1 week of IL-2 stimulation [5],and later the same group found CD16 expression in NK cellsin the efferent lymph as well as inflamed LNs [18]. Thus, NKcells harvested directly from calf LNs under physiologicalconditions have characteristics that human LN-derived NKcells only seem to acquire following activation. In contrastto humans, murine spleen NK cells that were similar to thosefound in LNs (CD27high) had cytotoxic capacity [36].Furthermore, an in vivo cytotoxic effector function of NKcells in LNs has been indicated following adoptive transfer ofNK cells into mice [11]. Together, these observations fromcattle and mice highlight the need for a careful examinationof NK effector functions in LNs on a general basis.
CD62L (L-selectin) is thought to be central in NK cellhoming to peripheral LNs through interaction with highendothelial venules [7,37]. The present study showed thatNK cells in bovine blood were divided into CD62Lhigh and
CD62L� populations, while in LNs a more homogeneousCD62Llow population was seen. In addition, CD62L on bovineblood-derived NK cells was downregulated following IL-2activation, as seen in both cattle and humans [24,38,39].These findings would be consistent with a selective entry ofCD62L+ NK cells into LNs, where activating conditions lead toa moderate downregulation or shedding [40] of CD62L. Alsoin mouse LNs, NK cells mostly express CD62L [6,7], butinterestingly, the CD56bright NK cells dominating in humanLNs did not express CD62L [5], even though this subsetconsistently expressed CD62L in blood in another report[37]. One remaining question was if a selective entry ofCD62L+ cells could explain the dominance of CD2�/low NKcells, the strongest IFN-g responsive subset, in bovine LNs.However, CD62L expression was similar in CD2�/low and CD2+
subsets of NK cells in blood. There are indications thatsecondary lymphoid tissues are sites for NK cell activationand priming [12,41], and a conceivable hypothesis could bethat CD2� and CD2+ subsets are not selectively recruited,but instead that activating conditions in the LNs leads to aselective priming and subsequent proliferation of CD2�/low
NK cells. Support for this may be drawn from our previousfindings that in vitro, this latter NK cell subset proliferatedmore efficiently under IL-2 stimulation compared to CD2+ NKcells [23]. An activating cytokine milieu in calf LNs couldalso explain other differences in phenotypic and functionalproperties, as discussed above.
ARTICLE IN PRESS
1.2%
19.6%
3.4%
19.1%
0.1%
5.0%
3.1%
5.2%
5.6%
10.5%
0.7%
4.8%
0.5%
33.2%
1.2%
6.7%
NKp46 NKp46CD2 CD2
IFN
-γ
Peripheral blood Jejunal lymph node
Medium
IL-2+IL-12
MNC MNCNK NK
58.9%
62.6%
41.8%
16.8%
Blood
Spleen
JLN
DJLN
MELN
SCLN
Blood
Spleen
JLN
DJLN
MELN
SCLN
IL-2
+ IL
-12
Mediu
m
% IFN- γ+ of all NK cells
0 10 20 30 40 50 60 70
IFN-γ (ng/ml)
0 10 20 30 40 50 60
CD2+ NK cells
CD2– NK cells
Figure 4 IFN-g response to cytokine stimulation of bovine NK cells from peripheral blood and tissues. (A) Intracellular flowcytometric analysis of IFN-g production. Left hand panels show all mononuclear cells from the indicated organs; right hand panelsshow NK cells gated through the highlighted regions/arrows and indicate IFN-g production in relation to surface expression of CD2.Numbers indicate quadrant % of each panel. Results shown are from one representative animal of three. (B) Summary of resultsobtained by the gating strategy in (A), presenting IFN-g+ NK cell subtype as per cent of all NK cells from each organs. Results aremean7SD of three tested animals. (C) IFN-g concentration in cell supernatants of mononuclear cells from organs and stimulations asindicated in (B), measured by ELISA. Results are mean7SD of four tested animals.
Natural killer cells in lymph nodes of healthy calves 781
The present investigation provides differential relativenumbers of NK cells in LNs of different anatomical sites ofthe calf, to our knowledge not systematically compared inother species. As with the quantitative data available fromthe human and murine models [4–8], some reservationsneed to be taken in the interpretation of these numbers: Inmost of these studies, including the present one, cells wereharvested by mechanical dissociation of the organs, render-ing the possibility that subclasses of cells could beselectively retained in tissue residues [42]. Furthermore,relative numbers are influenced by variations in other celltypes. In the present study, relative NK cell numbers wereclearly influenced by lower B-cell numbers and smaller
germinal centre sizes in non-mucosal LNs compared tomucosal LNs, as would be expected from lower microbialexposure in the former area [43]. It remains to be clarifiedhow different microbial exposure might influence theabsolute numbers, phenotypes and functions of NK cells inLNs in cattle as well as in other species. Furthermore, weare not aware of reports on how NK cell properties vary withage in any species. In this study, moderately lower relativeNK cell numbers were found in JLNs of adult animals, butsince the health records and exact age of these could not beobtained, a more systematic study under controlled condi-tions should be performed in order to draw firm conclusionsabout NK cells variation with age.
ARTICLE IN PRESS
4 1
Calculated NK/T ratio (:1)
0
4
8
6
2
10
12
Specific
lysis
%
Animal #1 Animal #2 Animal #3 Animal #4
16 0.25 4 116 0.25 4 116 0.25 4 116 0.25
0
4
2
4 116 0.25 4 116 0.25 4 116 0.25 4 116 0.25
PB DJLN SCLN
Perforin
JELN
DJLN
SCLN
MELN
Blood
Spleen
Figure 5 NK cell-specific cytotoxicity and perforin expression. (A, B) In a 51Cr release assay, mononuclear cells from the indicatedorgans were analysed for cytotoxicity against P815 target cells in the absence of cytokine stimulation, and presence (A) or absence(B) of the NK cell-specific anti-NKp46 mAb. E/T ratios were standardized for NK cell content using the phenotypic data obtained byflow cytometry (Table 2A). All of four animals shown individually. (C) NK cells from the three indicated organs were analysed forintracellular perforin expression by flow cytometry. Solid open lines indicate perforin expression, and shaded histograms indicatesecondary control mAbs. Results representative of five (blood), three (DJLN) and one (SCLN) animals are shown.
P. Boysen et al.782
In conclusion, we have presented here that in calves, NKcells reside in LNs in numbers equal to or higher thanreported in humans, and clearly higher than in mice. NKcells in LNs expressed CD16, NKp46 and perforin, had anactivated phenotype and had both cytokine-producing andcytotoxic functions. These results from the bovine modeldemonstrate that NK cells of a cytotoxic type can be foundin lymphoid tissues of healthy individuals, at odds with themore restrictive cytokine-producing role recently suggestedfor NK cells in such organs.
Acknowledgements
We wish to thank Grethe M. Johansen for assistance withtissue preparations and cytokine assays, and Laila Aune forimmunohistochemical analysis. Furthermore, we thank WillL. Goff (Animal Disease Research Unit, USDA-ARS, Pullman,Washington, USA) for providing direct conjugate of anti-CD3mAb, Jayne C. Hope (Institute for Animal Health, Compton,UK) for providing rbIL-12, Gregers Jungersen (DanishInstitute for Food and Veterinary Research, Copenhagen,Denmark) for providing anti-IFN-g mAb and Sigbjørn Fossum(Institute of Basic Medical Sciences, Oslo, Norway) forcritical reading of this manuscript. This work was carriedout under grant support from the Norwegian ResearchCouncil.
References
[1] Biron CA, Nguyen KB, Pien GC, Cousens LP, Salazar-Mather TP.Natural killer cells in antiviral defense: function and regulationby innate cytokines. Annu Rev Immunol 1999;17:189–220.
[2] Dokun AO, Chu DT, Yang L, Bendelac AS, Yokoyama WM.Analysis of in situ NK cell responses during viral infection.J Immunol 2001;167(9):5286–93.
[3] Morris MA, Ley K. Trafficking of natural killer cells. Curr MolMed 2004;4(4):431–8.
[4] Fehniger TA, Cooper MA, Nuovo GJ, Cella M, Facchetti F,Colonna M, et al. CD56bright natural killer cells are present inhuman lymph nodes and are activated by T cell-derived IL-2: apotential new link between adaptive and innate immunity.Blood 2003;101(8):3052–7.
[5] Ferlazzo G, Thomas D, Lin SL, Goodman K, Morandi B, MullerWA, et al. The abundant NK cells in human secondary lymphoidtissues require activation to express killer cell Ig-like receptorsand become cytolytic. J Immunol 2004;172(3):1455–62.
[6] Bajenoff M, Breart B, Huang AY, Qi H, Cazareth J, Braud VM,et al. Natural killer cell behavior in lymph nodes revealed bystatic and real-time imaging. J Exp Med 2006;203(3):619–31.
[7] Chen S, Kawashima H, Lowe JB, Lanier LL, Fukuda M.Suppression of tumor formation in lymph nodes by L-selectin-mediated natural killer cell recruitment. J Exp Med 2005;202(12):1679–89.
[8] Martin-Fontecha A, Thomsen LL, Brett S, Gerard C, Lipp M,Lanzavecchia A, et al. Induced recruitment of NK cells to

ARTICLE IN PRESS
Natural killer cells in lymph nodes of healthy calves 783
lymph nodes provides IFN-gamma for T(H)1 priming. NatImmunol 2004;5(12):1260–5.
[9] Morandi B, Bougras G, Muller WA, Ferlazzo G, Munz C. NK cellsof human secondary lymphoid tissues enhance T cell polariza-tion via IFN-gamma secretion. Eur J Immunol 2006;36(9):2394–400.
[10] Walzer T, Blery M, Chaix J, Fuseri N, Chasson L, Robbins SH, etal. Identification, activation, and selective in vivo ablation ofmouse NK cells via NKp46. Proc Natl Acad Sci USA 2007;104(9):3384–9.
[11] Garrod KR, Wei SH, Parker I, Cahalan MD. Natural killer cellsactively patrol peripheral lymph nodes forming stable con-jugates to eliminate MHC-mismatched targets. Proc Natl AcadSci USA 2007;104(29):12081–6.
[12] Lucas M, Schachterle W, Oberle K, Aichele P, Diefenbach A.Dendritic cells prime natural killer cells by trans-presentinginterleukin 15. Immunity 2007;26(4):503–17.
[13] Lanier LL, Le AM, Civin CI, Loken MR, Phillips JH. Therelationship of CD16 (Leu-11) and Leu-19 (NKH-1) antigenexpression on human peripheral blood NK cells and cytotoxic Tlymphocytes. J Immunol 1986;136(12):4480–6.
[14] Nagler A, Lanier LL, Cwirla S, Phillips JH. Comparative studiesof human FcRIII-positive and negative natural killer cells.J Immunol 1989;143(10):3183–91.
[15] Cooper MA, Fehniger TA, Caligiuri MA. The biology of humannatural killer-cell subsets. Trends Immunol 2001;22(11):633–40.
[16] Jacobs R, Hintzen G, Kemper A, Beul K, Kempf S, Behrens G,et al. CD56bright cells differ in their KIR repertoire andcytotoxic features from CD56dim NK cells. Eur J Immunol2001;31(10):3121–7.
[17] Freud AG, Yokohama A, Becknell B, Lee MT, Mao HC, FerketichAK, et al. Evidence for discrete stages of human natural killercell differentiation in vivo. J Exp Med 2006;203(4):1033–43.
[18] Romagnani C, Juelke K, Falco M, Morandi B, D’Agostino A,Costa R, et al. CD56brightCD16- killer Ig-like receptor-NK cellsdisplay longer telomeres and acquire features of CD56dim NKcells upon activation. J Immunol 2007;178(8):4947–55.
[19] Moretta L, Bottino C, Pende D, Vitale M, Mingari MC, Moretta A.Human natural killer cells: molecular mechanisms controllingNK cell activation and tumor cell lysis. Immunol Lett 2005;100(1):7–13.
[20] Westgaard IH, Berg SF, Vaage JT, Wang LL, Yokoyama WM,Dissen E, et al. Rat NKp46 activates natural killer cellcytotoxicity and is associated with FcepsilonRIgamma andCD3zeta. J Leukoc Biol 2004;76(6):1200–6.
[21] Storset AK, Kulberg S, Berg I, Boysen P, Hope JC, Dissen E.NKp46 defines a subset of bovine leukocytes with natural killercell characteristics. Eur J Immunol 2004;34(3):669–76.
[22] Kulberg S, Boysen P, Storset AK. Reference values for relativenumbers of natural killer cells in cattle blood. Dev CompImmunol 2004;28(9):941–8.
[23] Boysen P, Olsen I, Berg I, Kulberg S, Johansen GM, Storset AK.Bovine CD2-/NKp46+ cells are fully functional natural killercells with a high activation status. BMC Immunol 2006;7(1):10.
[24] Endsley JJ, Endsley MA, Estes DM. Bovine natural killer cellsacquire cytotoxic/effector activity following activation withIL-12/15 and reduce Mycobacterium bovis BCG in infectedmacrophages. J Leukoc Biol 2006;79(1):71–9.
[25] Gunnes G, Valheim M, Press CM, Tverdal A, Storset A.Comparison of flow cytometry and image morphometry in thequantitative analysis of cell population markers in the lymphnode of sheep. Vet Immunol Immunopathol 2003;94(3-4):177–83.
[26] Maecker HT, Moon J, Bhatia S, Ghanekar SA, Maino VC, PayneJK, et al. Impact of cryopreservation on tetramer, cytokineflow cytometry, and ELISPOT. BMC Immunol 2005;6:17.
[27] Moretta A, Tambussi G, Ciccone E, Pende D, Melioli G, MorettaL. CD16 surface molecules regulate the cytolytic function ofCD3CD16+ human natural killer cells. Int J Cancer 1989;44(4):727–30.
[28] Collins RA, Gelder KI, Howard CJ. Nucleotide sequence ofcattle FcGRIII: its identification in gammadelta T cells.Immunogenetics 1997;45(6):440–3.
[29] Hein WR, Dudler L, Marston WL, Landsverk T, Young AJ, Avila D.Ubiquitination and dimerization of complement receptor type2 on sheep B cells. J Immunol 1998;161(1):458–66.
[30] Sivori S, Vitale M, Morelli L, Sanseverino L, Augugliaro R,Bottino C, et al. p46, a novel natural killer cell-specific surfacemolecule that mediates cell activation. J Exp Med 1997;186(7):1129–36.
[31] Ziegler-Heitbrock HW, Fingerle G, Strobel M, Schraut W, StelterF, Schutt C, et al. The novel subset of CD14+/CD16+ bloodmonocytes exhibits features of tissue macrophages. EurJ Immunol 1993;23(9):2053–8.
[32] Lanier LL, Kipps TJ, Phillips JH. Functional properties of aunique subset of cytotoxic CD3+ T lymphocytes that express Fcreceptors for IgG (CD16/Leu-11 antigen). J Exp Med 1985;162(6):2089–106.
[33] Valheim M, Storset AK, Aleksersen M, Brun-Hansen H. Lesions insubclinical paratuberculosis of goats are associated withpersistent gut-associated lymphoid tissue. J Comp Pathol 2002;127(2-3):194–202.
[34] Schummer A, Nickel R, Schummer A, Seiferle E, editors. Theanatomy of domestic animals. Volume 3: the circulatorysystem, the skin, and the cutaneous organs of the domesticmammals. Berlin: Verlag Paul Parey; 1981.
[35] Hercend T, Schmidt RE. Characteristics and uses of naturalkiller cells. Immunol Today 1988;9(10):291–3.
[36] Hayakawa Y, Smyth MJ. CD27 dissects mature NK cells into twosubsets with distinct responsiveness and migratory capacity.J Immunol 2006;176(3):1517–24.
[37] Campbell JJ, Qin S, Unutmaz D, Soler D, Murphy KE, Hodge MR,et al. Unique subpopulations of CD56+ NK and NK–T peripheralblood lymphocytes identified by chemokine receptor expres-sion repertoire. J Immunol 2001;166(11):6477–82.
[38] Frey M, Packianathan NB, Fehniger TA, Ross ME, Wang WC,Stewart CC, et al. Differential expression and function ofL-selectin on CD56bright and CD56dim natural killer cellsubsets. J Immunol 1998;161(1):400–8.
[39] Uksila J, Salmi M, Butcher EC, Tarkkanen J, Jalkanen S.Function of lymphocyte homing-associated adhesion moleculeson human natural killer and lymphokine-activated killer cells.J Immunol 1997;158(4):1610–7.
[40] Tedder TF, Steeber DA, Chen A, Engel P. The selectins: vascularadhesion molecules. FASEB J 1995;9(10):866–73.
[41] Ferlazzo G, Pack M, Thomas D, Paludan C, Schmid D, Strowig T,et al. Distinct roles of IL-12 and IL-15 in human natural killercell activation by dendritic cells from secondary lymphoidorgans. Proc Natl Acad Sci USA 2004;101(47):16606–11.
[42] Fossum S, Rolstad B, Tjernshaugen H. Selective loss of S-phasecells when making cell suspensions from lymphoid tissue. CellImmunol 1979;48(1):149–54.
[43] Gunnes G, Press CM, Tverdal A, Landsverk T. Compartmentswithin the lymph node cortex of calves and adult cattlediffer in the distribution of leukocyte populations: animmunohistochemical study using computer-assisted morpho-metric analysis. Dev Comp Immunol 1998;22(1):111–23.
Copyright © 2022 FDOKUMEN




















